
Hepatitis B virus promotes hepatocarcinogenesis in transgenic mice
Upload
independentCategory
view
1download
0
ORIGINAL PAPER
Effects of transgenic rootstocks on growth and developmentof non-transgenic scion cultivars in apple
Anders Smolka • Xue-Yuan Li • Catrin Heikelt •
Margareta Welander • Li-Hua Zhu
Received: 3 September 2009 / Accepted: 18 January 2010 / Published online: 5 February 2010
� Springer Science+Business Media B.V. 2010
Abstract Although cultivation of genetic modified
(GM) annual crops has been steadily increasing in the
recent 10 years, the commercial cultivation of GM
fruit tree is still very limited and reports of field trials
on GM fruit trees are rare. This is probably because
development and evaluation of GM fruit trees require
a long period of time due to long life cycles of trees. In
this study, we report results from a field trial on three
rolB transgenic dwarfing apple rootstocks of M26 and
M9 together with non-transgenic controls grafted with
five non-transgenic scion cultivars. We intended to
investigate the effects of transgenic rootstock on non-
transgenic scion cultivars under natural conditions as
well as to evaluate the potential value of using the
rolB gene to modify difficult-to-root rootstocks of
fruit trees. The results showed that all rolB transgenic
rootstocks significantly reduced vegetative growth
including tree height regardless of scion cultivar,
compared with the non-transgenic rootstocks. Flow-
ering and fruiting were also decreased for cultivars
grown on the transgenic rootstocks in most cases, but
the fruit quality was not clearly affected by the
transgenic rootstocks. Cutting experiment and RT-
PCR analysis showed that the rolB gene was stably
expressed under field conditions. PCR and RT-PCR
analyses displayed that the rolB gene or its mRNA
were not detectable in the scion cultivars, indicating
no translocation of the transgene or its mRNA from
rootstock to scion. Our results suggest that rolB
modified rootstocks should be used in combination
with vigorous scion cultivars in order to obtain
sufficient vegetative growth and good yield. Alterna-
tively, the rolB gene could be used to dwarf vigorous
rootstocks of fruit trees or produce bonzai plants as
it can significantly reduce the vegetative growth of
plants.
Keywords rolB � GM apple rootstock �Scion cultivar � Growth � Flower � Fruit quality
Introduction
Genetic engineering is becoming more and more
important in modern plant breeding as it can modify
specific traits that are not easily reached using
conventional breeding methods. Commercial cultiva-
tion of genetically modified plants (GMPs) has
steadily increased worldwide in the past 10 years.
By 2008, the cultivation of GMPs had reached over
100 million hectares in 25 countries. The cultivated
GMPs are predominantly annual crops with increased
insect and pesticide resistance or herbicide tolerance,
A. Smolka � X.-Y. Li � C. Heikelt � M. Welander (&) �L.-H. Zhu (&)
Department of Plant Breeding and Biotechnology,
Swedish University of Agricultural Sciences, Box 101,
230 53 Alnarp, Sweden
e-mail: [email protected]
L.-H. Zhu
e-mail: [email protected]
123
Transgenic Res (2010) 19:933–948
DOI 10.1007/s11248-010-9370-0

while the commercial cultivation of GM fruit tree is
very limited. So far, the only GM tree species that are
commercially cultivated are GM poplar with insect
resistance grown in China and GM papaya with virus
resistance grown in USA and China (James 2008).
Fruit trees are commonly propagated by grafting a
scion cultivar onto a rootstock. Rootstocks have a
special value for fruit production as they can
effectively control tree size, influence growth and
development of cultivars grafted onto them and
tolerate adverse soil conditions (Drake et al. 1988;
Fallahi et al. 1985; Ferree et al. 2001a, b; Hirst and
Ferree 1995, 1996; Lauri et al. 2006; Tubbs 1974;
Tworkoski and Miller 2007; Webster et al. 1985).
However, there are no ideal rootstocks available for
commercial production and genetic improvement of
existing rootstocks or breeding new rootstocks is thus
a continuous goal for fruit tree breeding. Genetic
modification has been proved to be efficient for
improving currently available rootstocks where only
one or a few traits need to be improved (Zhu et al.
2001, 2003). Studies on GM fruit trees under field
conditions are still very scarce with the only reports
on fire blight resistance by Norelli et al. (2003) and
Aldwinckle et al. (2003), fruit firmness by Hrazdina
et al. (2003) and fruit flavour by Dandekar et al.
(2004). However, there is little information about the
field trials and evaluation of field-grown trees
regarding growth and development in these reports.
The information about effects of transgenic root-
stocks on non-transgenic scions of fruit trees under
field conditions is not available since there is no field
trial on GM rootstocks of fruit trees. Clearly,
systematic evaluation of GM fruit trees concerning
growth and development is necessary to increase our
knowledge on GM trees under field conditions.
Another issue related to GM rootstocks is that,
unlike GM scion cultivars, the use of GM rootstocks
in combination with non-transgenic scion cultivars
may circumvent the food safety issue if transgenes or
their products are not present in scion fruits. The
results reported so far have been inconsistent in this
regard. Dutt et al. (2007) and Bortolotti et al. (2005)
have reported that transgene proteins were found in
sap and phloem of non-transgenic scions grafted onto
transgenic rootstock in grape and tobacco, respec-
tively, but Youk et al. (2009) reported that no
transgenic products were translocated from transgenic
rootstock to non-transgenic scion in watermelon.
Since reports on this issue are very limited, further
studies are needed to facilitate the future application
of GM rootstocks in fruit production.
In modern apple production, dwarfing rootstocks
are commonly used for achieving high production
efficiency. Dwarfing rootstocks are often propagated
vegetatively, but they are often difficult-to-root,
especially when they reach the adult phase. Improve-
ment of the rooting ability is thus necessary for these
rootstocks. The rolB (rooting locus B) gene, isolated
from the soil bacterium Agrobacterium rhizogenes
(Cardarelli et al. 1987; Vilaine and Cassedelbart
1987), is a well documented rooting related gene and
has been proved to stimulate rooting in different plant
species when overexpressed (Capone et al. 1989; Dai
et al. 2004; Feyissa et al. 2007; Geier et al. 2008;
Rugini et al. 1991; Spena et al. 1987; Tzfira et al.
1996; Welander et al. 1998). Except for its effect on
adventitious rooting, the rolB gene can also stimulate
flowering in transgenic tobacco plants (Altamura et al.
1994). The rolB gene has been used for modifying
difficult-to-root dwarfing apple and pear rootstocks
and showed a great increase in rooting ability both in
vitro and ex vitro as well as reduced plant size under
greenhouse conditions (Zhu et al. 2001, 2003). It has
also been reported that, under non-limiting nutrient
conditions, the relative growth rate of the transgenic
apple rootstock was not altered by the rolB gene
compared to the untransformed control (Zhu and
Welander 1999). It is however unknown how the rolB
transgenic rootstocks will affect growth and develop-
ment of non-transgenic scion cultivars under field
conditions. The answer to this question would facil-
itate the potential use of the rolB gene in breeding of
fruit tree rootstocks in the future.
In this study we evaluated the effects of rolB
transgenic apple rootstocks on growth, flowering and
fruit quality of non-transgenic scion cultivars grafted
onto these rootstocks, the stability of the rolB gene
expression under natural conditions, possibility of
translocation of the rolB gene or its mRNA from
rootstock to scion. Our results show that the rolB
transgenic rootstocks reduced growth and flowering
of the scion cultivars, while the fruit quality was not
apparently affected. These effects varied among
different cultivars with the strongest effect on the
cultivar Discovery. The rolB gene is stably expressed
under field conditions. No translocation of the rolB
gene or its mRNA was detected in the scion cultivars.
934 Transgenic Res (2010) 19:933–948
123

Materials and methods
Plant material
The field trial was located in the Southern part of
Sweden and initiated by planting in vitro rooted
rootstocks in a nursery in 2001. Rootstocks used in
the trial consisted of non-transgenic M26, transgenic
M26-rolB (clone F), non-transgenic M9, transgenic
M9-rolB1 (clone ARB3) and M9-rolB2 (clone
ARB9). These transgenic rootstocks have previously
been described by Welander et al. (1998) and Zhu
et al. (2001). The reasons for choosing these clones
are because they do not contain the gus gene and have
either single or two copies of the rolB gene in order to
maintain the stable gene expression under field
conditions. The clone F of M26 contains one copy
of the rolB gene and rooted better than clone C.
ARB3 and ARB9 contain two copies of the rolB gene
(all clones with one copy of the rolB gene were
unfortunately lost). Both clones had significantly
increased rooting compared to the control, while
clone ARB9 is dwarfer than ARB3. Five types of
apple cultivars (Aroma, Discovery, Elise, Elstar and
Jonagold) were budded onto the five rootstocks in
July 2002 in the nursery. The budded trees were
allowed to grow two seasons in the nursery before
being planted in the field trial. Well-developed
budded trees were then planted in the field trial in
May 2004. Every scion-rootstock combination con-
sisted of 10 trees which were randomly distributed in
the field to avoid potential soil effects. The trees were
planted in a distance of 2.5 9 3 m and managed with
a general maintenance including spraying with pes-
ticides, pruning and fertilization. The spaces between
the trees were covered with grass. The pruning was
carried out every spring in a way to obtain crown
trees and the degree of pruning may differ from year
to year as well as from cultivar to cultivar based on
their growth habits. In order to secure vegetative
growth of small trees, flowers were removed during
the first 2 years of the field trial. The non-grafted rolB
rootstocks and non-transgenic control rootstocks
were also planted in the field in 2004.
Registration of growth data, flowers and fruits
Tree height, trunk diameter at about 10 cm above the
ground and annual shoot growth were registered from
all trees after growth cessation from 2003 to 2007
(the registration was started when the trees were still
in the nursery). Flower buds and the number of
flowers from all trees were counted before full bloom
from 2005 to 2007 and the number of fruits from all
trees was recorded in autumn from 2007 to 2009.
Fruit quality analysis
Fruits were harvested when they were fully ripen and
stored in a paper bag covered with a plastic bag at
4�C before fruit quality analysis. Fruit quality
parameters analysed include fresh weight, diameter,
firmness, colour, total soluble solids (TSS), titratable
acidity (TA), total phenols and vitamin C content.
Depending on the available amount of fruits at
harvest time (some were damaged due to insects and
diseases), fruit quality analysis was carried out only
on three cultivars, namely Elise, Elstar and Jonagold
in 2008. Fruit fresh weight, diameter, firmness and
colour were measured on 10 randomly picked apples
from 5 trees for each combination. The measurements
were carried out after 1 month of storage for Elise
and Elstar, and after 6 months of storage for Jona-
gold. For analysing TSS, TA, total phenols and
vitamin C, 2 apples were made into one biological
sample, and 5 biological replicates were made for
each parameter with 3 measurements from each
biological replicate.
Fruit firmness and colour measurements
Firmness was measured with a penetrometer FT 327
with 11 mm plunger. The measurements were made
on two sides (sun-side/shade-side) of apples and the
means were presented as kg/cm2. Fruit colour was
measured on three positions of a fruit with a Minolta
Chroma Meter CR-200 according to manufacturer’s
instructions. The results were expressed as hue angle,
which is determined as H = tan - 1(b/a) where a
represents chromaticity on a green (-) to red (?) axis
and b represents chromaticity on a blue (-) to yellow
(?) axis. H ranges from 0 (red colour) to 90 (yellow
colour).
Analysis of TSS and titratable acidity
About equally large sectors from the sun-side and
shade-side of apples were used to prepare juice in a
Transgenic Res (2010) 19:933–948 935
123

food mixer. TSS was analysed using Precision
Instrument’s digital refractometer RFM 80. Titratable
acidity (TA) was titrated as malic acid with 0.05 M
NaOH to pH 8.
Analysis of total phenolics
Total phenolics from flesh and peel were analysed
separately using a modified Folin-Ciocalteu colorimet-
ric method according to Dewanto et al. (2002). In brief,
chopped apple flesh or peels were extracted in 50%
ethanol for 10 min. After centrifuging at 13,000 rpm for
15 min, 63 ll of the supernatant was added to a 1.0 ml
cuvette together with 250 ll water and 63 ll Folin–
Ciocalteau’s reagent. After reaction for 6 min, 625 ll
Na2CO3 7% was added into the cuvette to raise the pH
for phenols to be oxidized to phenolates (Dewanto et al.
2002). The samples were then allowed to stand for
75 min before being measured at 765 nm with a
Shimadzu Recording Spectrophotometer UV-240
Grapicord. A standard curve of gallic acid at the
concentrations of 0, 5, 10, 20 and 40% was made to
calculate the content of total phenols in the samples.
Total phenol content was expressed as mg gallic acid
equivalent per g fresh weight.
Vitamin C analysis
The content of vitamin C was analysed in form of
L-ascorbic acid using HPLC. Tissues of about 5 g
fresh weight were homogenised with 25 ml 1.5%
metaphosphoric acid with an Ultra-Turrax homoge-
nizer. The homogenised mixture was centrifuged at
13,000 rpm at 4�C. 500 ll of the supernatant was
added to an Eppendorf tube, followed by addition of
550 ll 7 mM DTT to reduce dehydroascorbic acid
(DHA) to ascorbic acid. After 30 min of reaction, the
sample was centrifuged for 5 min at 10,000 rpm at
4�C. The whole extraction process was executed
under green light conditions to minimize degradation
of vitamin C. Supernatant of 600 ll was transferred
to a HPLC vial for HPLC analysis (LaChrome Merck
Hitachi with the data program D-7000 HSM HPLC
and the column Phenomenex Synergi 4u polar RP).
The mobile phase was single, containing 20 mM
KH2PO4 buffer and 4% methanol with pH adjusted to
2.3 with H3PO4. Ten microliters were injected and
the detection wavelength was 248 nm with the flow
rate of 1 ml/min. The content of ascorbic acid was
calculated based on a standard curve of known
concentrations of ascorbic acid.
Cutting experiment
In order to confirm that the rolB gene is stably
expressed and can still stimulate adventitious rooting
under field conditions, a cutting experiment was
carried out in greenhouse. Cuttings were made of
annual shoots taken from non-grafted non-transgenic
and transgenic rootstocks grown in the field trial in
July of 2007. About 2–3 nodes were taken for each
cutting with two half leaves. If leaves were small, two
full leaves were kept. The prepared cuttings were
then planted into a mixture of soil and perlite (1:1) in
a plate covered with plastic film to maintain high
humility. About 30 cuttings were taken for each
rootstock. The temperature in the greenhouse was 23/
18�C (day/night) with natural light. The rooting result
was recorded after 2 months.
Expression of the rolB gene under field conditions
To investigate the stability of the transgene expression
under field conditions, growing shoot tips of the non-
grafted rolB rootstocks and non-transgenic control
rootstocks were taken in July 2007 (when shoots grew
vigorously) for RT-PCR analysis of the rolB gene.
Total RNA extraction and RT-PCR were carried out
according to Feyissa et al. (2007) and Zhu et al. (2008).
The primers were 50-ATGGATCCCAAATTGCTAT
TCCTTCCACGA-30 and 50-TTAGGCTTCTTTCTT
CAGGTTTACTGCAGC-30, yielding a 776 bp product.
Detection of the rolB gene in scion cultivar
Leaf and flower samples were taken from the scion
cultivars grafted onto the transgenic rootstocks.
Genomic DNA extraction and PCR analysis were
carried out according to Zhu et al. (2001, 2003).
Detection of rolB mRNA in scion cultivar
In order to verify if the rolB mRNA could be
translocated from rootstock to scion, growing shoot
tips were taken from the scion cultivars grafted onto
the different rootstocks. Total RNA was extracted
and RT-PCR was performed according to Feyissa
et al. (2007).
936 Transgenic Res (2010) 19:933–948
123

Statistic analysis
Data was subjected to analysis of variance (ANOVA)
with Duncan’s multiple range test using the Stat-
graphics program.
Results
Effect of transgenic rootstock on success
of budding
The budding union between rootstock and scion was
formed smoothly for all cultivars grown on the
transgenic and control rootstocks, observed by naked
eyes. The success of budding was almost 98% in
general for all combinations, indicating that the rolB
gene did not have any negative effect on budding
(data not shown).
Effect of transgenic rootstock on tree growth
The bud breaking time was not affected by the
transgenic rootstocks, as observed by naked eyes each
spring. However, the rolB transgenic rootstocks
significantly reduced tree growth in most cases, as
measured by tree height, stem diameter, annual shoot
length and annual shoot number as shown in Tables 1,
2, 3 and 4. From 2005 onward, this reduction is
significant for all combinations when compared with
Table 1 Plant height (cm) of five apple cultivars grafted onto transgenic and non-transgenic apple rootstocks
Combination
(cultivar/rootstock)
2003
(Mean ± SE)
2004
(Mean ± SE)
2005
(Mean ± SE)
2006
(Mean ± SE)
2007
(Mean ± SE)
Average of
5 years
(mean ± SE)
Aroma/M26 85 ± 2.7b 110 ± 3.2a 130 ± 2.9a 160 ± 4.8a 170 ± 2.9a 131 ± 1.5a
Aroma/M26-rolB 69 ± 2.2c 102 ± 3.3a 115 ± 2.9b 129 ± 4.8b 149 ± 2.9b 113 ± 1.5b
Aroma/M9 95 ± 2.2a 109 ± 3.3a 135 ± 2.9a 152 ± 5.1a 180 ± 2.8a 134 ± 1.5a
Aroma/M9-rolB1 73 ± 2.2c 111 ± 3.1a 117 ± 2.9b 121 ± 5.1b 153 ± 2.9b 115 ± 1.5b
Aroma/M9-rolB2 60 ± 2.2d 92 ± 3.2b 107 ± 2.9b 107 ± 4.8c 139 ± 2.9c 101 ± 1.5c
Discovery/M26 91 ± 2.2a 95 ± 3.7a 116 ± 2.9a 132 ± 3.9a 136 ± 2.9a 114 ± 1.6a
Discovery/M26-rolB 73 ± 2.2c 72 ± 3.7b 92 ± 2.9b 99 ± 4.1b 119 ± 2.9b 91 ± 1.5b
Discovery/M9 92 ± 2.2a 95 ± 3.3a 116 ± 2.9a 129 ± 3.9a 136 ± 2.6a 114 ± 1.5a
Discovery/M9-rolB1 80 ± 2.2b 88 ± 3.8a 87 ± 2.9b 88 ± 3.9b 105 ± 3.0b 90 ± 2.0b
Discovery/M9-rolB2 62 ± 2.3d 90 ± 3.1a 96 ± 2.9b 95 ± 3.9b 112 ± 2.9b 91 ± 1.6b
Elise/M26 96 ± 2.1a 111 ± 3.5a 130 ± 2.9a 151 ± 4.0a 149 ± 2.9a 127 ± 1.7a
Elise/M26-rolB 70 ± 2.1b 85 ± 3.0b 108 ± 2.9b 115 ± 4.0c 113 ± 2.9bc 98 ± 1.6c
Elise/M9 95 ± 2.2a 95 ± 3.2b 136 ± 2.9a 137 ± 4.0b 149 ± 2.7a 122 ± 1.6a
Elise/M9-rolB1 72 ± 2.2b 99 ± 3.2b 111 ± 2.9b 115 ± 4.3c 123 ± 2.9b 104 ± 1.7b
Elise/M9-rolB2 57 ± 2.3c 83 ± 3.2b 97 ± 2.9c 97 ± 4.0d 110 ± 2.9c 89 ± 1.7d
Elstar/M26 78 ± 2.2b 108 ± 3.2b 148 ± 2.9a 182 ± 4.1a 165 ± 2.9b 136 ± 1.6b
Elstar/M26-rolB 85 ± 2.2b 112 ± 3.1b 132 ± 2.9b 154 ± 4.3b 146 ± 2.9d 126 ± 1.7c
Elstar/M9 92 ± 2.2a 125 ± 3.1a 146 ± 2.9a 186 ± 4.4a 179 ± 2.6a 146 ± 1.6a
Elstar/M9-rolB1 83 ± 2.1b 116 ± 3.1b 125 ± 2.9b 132 ± 4.1c 156 ± 3.0c 122 ± 1.6d
Elstar/M9-rolB2 68 ± 2.2c 112 ± 3.2b 116 ± 2.9c 128 ± 4.4c 145 ± 2.9d 114 ± 1.5e
Jonagold/M26 78 ± 2.2b 119 ± 3.2ab 136 ± 2.9a 164 ± 5.5a 158 ± 2.9b 131 ± 1.5b
Jonagold/M26-rolB 77 ± 2.2bc 121 ± 3.1ab 120 ± 2.9b 130 ± 5.5c 145 ± 2.9c 119 ± 1.5c
Jonagold/M9 101 ± 2.2a 125 ± 3.6a 138 ± 2.9a 150 ± 5.5b 175 ± 2.9a 138 ± 1.5a
Jonagold/M9-rolB1 72 ± 2.2c 120 ± 3.3ab 120 ± 2.9b 126 ± 5.5cd 142 ± 2.7c 116 ± 1.5c
Jonagold/M9-rolB2 63 ± 2.3d 114 ± 3.2b 119 ± 2.9c 111 ± 5.5d 138 ± 2.9c 109 ± 1.5d
Data were collected from 10 trees for each combination. Duncan’s multiple test was carried out among five different rootstocks
within one cultivar. Different letters in each column within one cultivar indicate significant differences at P = 0.05
Transgenic Res (2010) 19:933–948 937
123
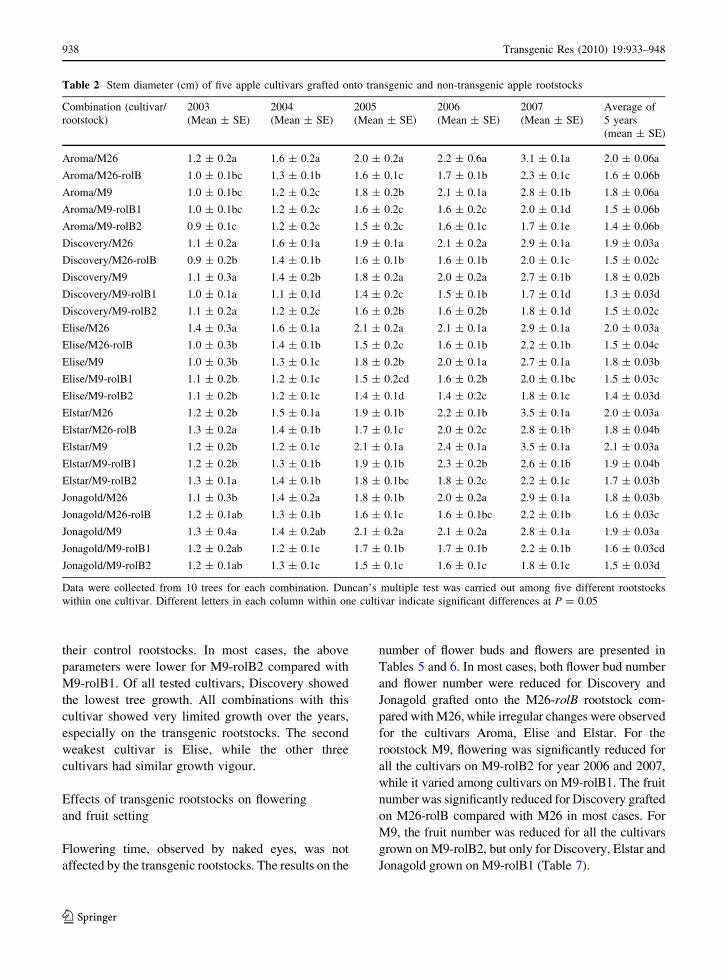
their control rootstocks. In most cases, the above
parameters were lower for M9-rolB2 compared with
M9-rolB1. Of all tested cultivars, Discovery showed
the lowest tree growth. All combinations with this
cultivar showed very limited growth over the years,
especially on the transgenic rootstocks. The second
weakest cultivar is Elise, while the other three
cultivars had similar growth vigour.
Effects of transgenic rootstocks on flowering
and fruit setting
Flowering time, observed by naked eyes, was not
affected by the transgenic rootstocks. The results on the
number of flower buds and flowers are presented in
Tables 5 and 6. In most cases, both flower bud number
and flower number were reduced for Discovery and
Jonagold grafted onto the M26-rolB rootstock com-
pared with M26, while irregular changes were observed
for the cultivars Aroma, Elise and Elstar. For the
rootstock M9, flowering was significantly reduced for
all the cultivars on M9-rolB2 for year 2006 and 2007,
while it varied among cultivars on M9-rolB1. The fruit
number was significantly reduced for Discovery grafted
on M26-rolB compared with M26 in most cases. For
M9, the fruit number was reduced for all the cultivars
grown on M9-rolB2, but only for Discovery, Elstar and
Jonagold grown on M9-rolB1 (Table 7).
Table 2 Stem diameter (cm) of five apple cultivars grafted onto transgenic and non-transgenic apple rootstocks
Combination (cultivar/
rootstock)
2003
(Mean ± SE)
2004
(Mean ± SE)
2005
(Mean ± SE)
2006
(Mean ± SE)
2007
(Mean ± SE)
Average of
5 years
(mean ± SE)
Aroma/M26 1.2 ± 0.2a 1.6 ± 0.2a 2.0 ± 0.2a 2.2 ± 0.6a 3.1 ± 0.1a 2.0 ± 0.06a
Aroma/M26-rolB 1.0 ± 0.1bc 1.3 ± 0.1b 1.6 ± 0.1c 1.7 ± 0.1b 2.3 ± 0.1c 1.6 ± 0.06b
Aroma/M9 1.0 ± 0.1bc 1.2 ± 0.2c 1.8 ± 0.2b 2.1 ± 0.1a 2.8 ± 0.1b 1.8 ± 0.06a
Aroma/M9-rolB1 1.0 ± 0.1bc 1.2 ± 0.2c 1.6 ± 0.2c 1.6 ± 0.2c 2.0 ± 0.1d 1.5 ± 0.06b
Aroma/M9-rolB2 0.9 ± 0.1c 1.2 ± 0.2c 1.5 ± 0.2c 1.6 ± 0.1c 1.7 ± 0.1e 1.4 ± 0.06b
Discovery/M26 1.1 ± 0.2a 1.6 ± 0.1a 1.9 ± 0.1a 2.1 ± 0.2a 2.9 ± 0.1a 1.9 ± 0.03a
Discovery/M26-rolB 0.9 ± 0.2b 1.4 ± 0.1b 1.6 ± 0.1b 1.6 ± 0.1b 2.0 ± 0.1c 1.5 ± 0.02c
Discovery/M9 1.1 ± 0.3a 1.4 ± 0.2b 1.8 ± 0.2a 2.0 ± 0.2a 2.7 ± 0.1b 1.8 ± 0.02b
Discovery/M9-rolB1 1.0 ± 0.1a 1.1 ± 0.1d 1.4 ± 0.2c 1.5 ± 0.1b 1.7 ± 0.1d 1.3 ± 0.03d
Discovery/M9-rolB2 1.1 ± 0.2a 1.2 ± 0.2c 1.6 ± 0.2b 1.6 ± 0.2b 1.8 ± 0.1d 1.5 ± 0.02c
Elise/M26 1.4 ± 0.3a 1.6 ± 0.1a 2.1 ± 0.2a 2.1 ± 0.1a 2.9 ± 0.1a 2.0 ± 0.03a
Elise/M26-rolB 1.0 ± 0.3b 1.4 ± 0.1b 1.5 ± 0.2c 1.6 ± 0.1b 2.2 ± 0.1b 1.5 ± 0.04c
Elise/M9 1.0 ± 0.3b 1.3 ± 0.1c 1.8 ± 0.2b 2.0 ± 0.1a 2.7 ± 0.1a 1.8 ± 0.03b
Elise/M9-rolB1 1.1 ± 0.2b 1.2 ± 0.1c 1.5 ± 0.2cd 1.6 ± 0.2b 2.0 ± 0.1bc 1.5 ± 0.03c
Elise/M9-rolB2 1.1 ± 0.2b 1.2 ± 0.1c 1.4 ± 0.1d 1.4 ± 0.2c 1.8 ± 0.1c 1.4 ± 0.03d
Elstar/M26 1.2 ± 0.2b 1.5 ± 0.1a 1.9 ± 0.1b 2.2 ± 0.1b 3.5 ± 0.1a 2.0 ± 0.03a
Elstar/M26-rolB 1.3 ± 0.2a 1.4 ± 0.1b 1.7 ± 0.1c 2.0 ± 0.2c 2.8 ± 0.1b 1.8 ± 0.04b
Elstar/M9 1.2 ± 0.2b 1.2 ± 0.1c 2.1 ± 0.1a 2.4 ± 0.1a 3.5 ± 0.1a 2.1 ± 0.03a
Elstar/M9-rolB1 1.2 ± 0.2b 1.3 ± 0.1b 1.9 ± 0.1b 2.3 ± 0.2b 2.6 ± 0.1b 1.9 ± 0.04b
Elstar/M9-rolB2 1.3 ± 0.1a 1.4 ± 0.1b 1.8 ± 0.1bc 1.8 ± 0.2c 2.2 ± 0.1c 1.7 ± 0.03b
Jonagold/M26 1.1 ± 0.3b 1.4 ± 0.2a 1.8 ± 0.1b 2.0 ± 0.2a 2.9 ± 0.1a 1.8 ± 0.03b
Jonagold/M26-rolB 1.2 ± 0.1ab 1.3 ± 0.1b 1.6 ± 0.1c 1.6 ± 0.1bc 2.2 ± 0.1b 1.6 ± 0.03c
Jonagold/M9 1.3 ± 0.4a 1.4 ± 0.2ab 2.1 ± 0.2a 2.1 ± 0.2a 2.8 ± 0.1a 1.9 ± 0.03a
Jonagold/M9-rolB1 1.2 ± 0.2ab 1.2 ± 0.1c 1.7 ± 0.1b 1.7 ± 0.1b 2.2 ± 0.1b 1.6 ± 0.03cd
Jonagold/M9-rolB2 1.2 ± 0.1ab 1.3 ± 0.1c 1.5 ± 0.1c 1.6 ± 0.1c 1.8 ± 0.1c 1.5 ± 0.03d
Data were collected from 10 trees for each combination. Duncan’s multiple test was carried out among five different rootstocks
within one cultivar. Different letters in each column within one cultivar indicate significant differences at P = 0.05
938 Transgenic Res (2010) 19:933–948
123
Weight, diameter, firmness and colour
Results for fruit weight, diameter, firmness and
colour are presented in Table 8. In general, there
were no consistent changes in fruit weight and
diameter when comparing the transgenic rootstocks
with their control counterparts. In some cases, the
fruit weight and diameter did not differ or were
inconsistent between the transgenic and control
rootstock. This is also true for firmness. For M26,
the fruit colour tended to be more reddish for trees on
the transgenic rootstock for all cultivars, but no
significant difference was found for Elise. For M9,
there was almost no difference in fruit colour in most
cases. Compared with the other cultivars, Jonagold
fruits had much lower firmness and this is apparently
due to the much longer storage period for this cultivar
before analysis was carried out.
TSS, TA, Vitamin C and phenolics
For Jonagold, a significantly increased TSS was
detected in fruits grown on both transgenic M9
rootstocks compared with non-transgenic M9 root-
stock. For Elstar, TSS was significantly increased in
fruits grown on the transgenic rootstocks of both M26
and M9. There was no difference in TSS for Elise
between the transgenic rootstocks and non-transgenic
ones (Table 9). There was no significant difference in
TA in most cases for all cultivars when comparing
the transgenic rootstocks with the non-transgenic
control for both M26 and M9 (Table 9). There was no
significant difference in the content of ascorbic acid
for Jonagold when comparing the transgenic root-
stocks with the non-transgenic control. For Elstar, the
Vc content was lower in fruits on M26-rolB com-
pared to M26, but higher on M9-rolB1 compared
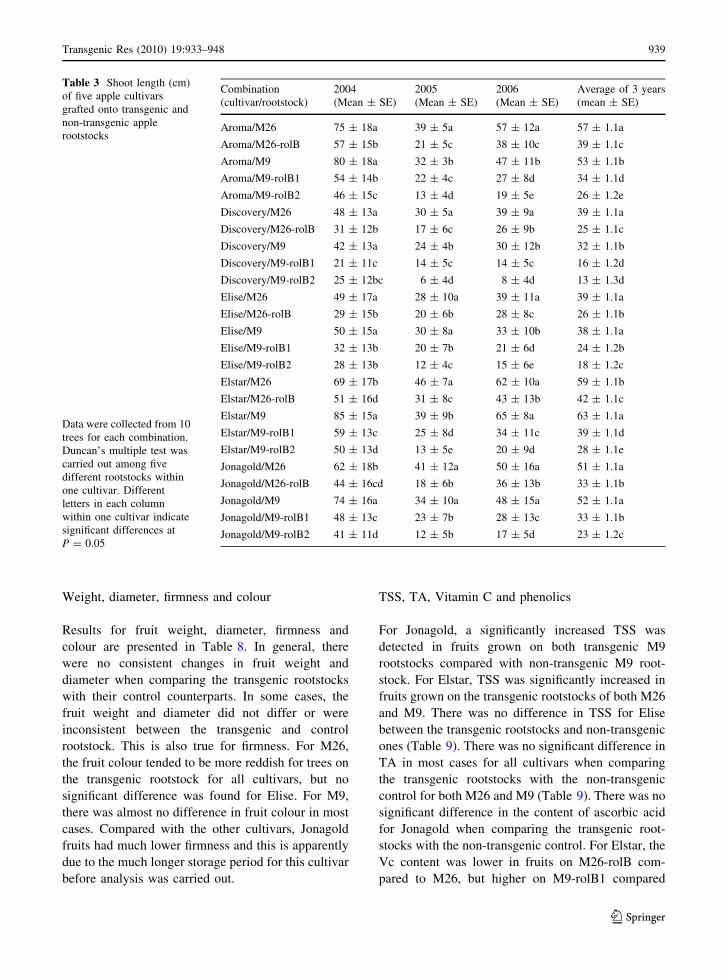
Table 3 Shoot length (cm)
of five apple cultivars
grafted onto transgenic and
non-transgenic apple
rootstocks
Data were collected from 10
trees for each combination.
Duncan’s multiple test was
carried out among five
different rootstocks within
one cultivar. Different
letters in each column
within one cultivar indicate
significant differences at
P = 0.05
Combination
(cultivar/rootstock)
2004
(Mean ± SE)
2005
(Mean ± SE)
2006
(Mean ± SE)
Average of 3 years
(mean ± SE)
Aroma/M26 75 ± 18a 39 ± 5a 57 ± 12a 57 ± 1.1a
Aroma/M26-rolB 57 ± 15b 21 ± 5c 38 ± 10c 39 ± 1.1c
Aroma/M9 80 ± 18a 32 ± 3b 47 ± 11b 53 ± 1.1b
Aroma/M9-rolB1 54 ± 14b 22 ± 4c 27 ± 8d 34 ± 1.1d
Aroma/M9-rolB2 46 ± 15c 13 ± 4d 19 ± 5e 26 ± 1.2e
Discovery/M26 48 ± 13a 30 ± 5a 39 ± 9a 39 ± 1.1a
Discovery/M26-rolB 31 ± 12b 17 ± 6c 26 ± 9b 25 ± 1.1c
Discovery/M9 42 ± 13a 24 ± 4b 30 ± 12b 32 ± 1.1b
Discovery/M9-rolB1 21 ± 11c 14 ± 5c 14 ± 5c 16 ± 1.2d
Discovery/M9-rolB2 25 ± 12bc 6 ± 4d 8 ± 4d 13 ± 1.3d
Elise/M26 49 ± 17a 28 ± 10a 39 ± 11a 39 ± 1.1a
Elise/M26-rolB 29 ± 15b 20 ± 6b 28 ± 8c 26 ± 1.1b
Elise/M9 50 ± 15a 30 ± 8a 33 ± 10b 38 ± 1.1a
Elise/M9-rolB1 32 ± 13b 20 ± 7b 21 ± 6d 24 ± 1.2b
Elise/M9-rolB2 28 ± 13b 12 ± 4c 15 ± 6e 18 ± 1.2c
Elstar/M26 69 ± 17b 46 ± 7a 62 ± 10a 59 ± 1.1b
Elstar/M26-rolB 51 ± 16d 31 ± 8c 43 ± 13b 42 ± 1.1c
Elstar/M9 85 ± 15a 39 ± 9b 65 ± 8a 63 ± 1.1a
Elstar/M9-rolB1 59 ± 13c 25 ± 8d 34 ± 11c 39 ± 1.1d
Elstar/M9-rolB2 50 ± 13d 13 ± 5e 20 ± 9d 28 ± 1.1e
Jonagold/M26 62 ± 18b 41 ± 12a 50 ± 16a 51 ± 1.1a
Jonagold/M26-rolB 44 ± 16cd 18 ± 6b 36 ± 13b 33 ± 1.1b
Jonagold/M9 74 ± 16a 34 ± 10a 48 ± 15a 52 ± 1.1a
Jonagold/M9-rolB1 48 ± 13c 23 ± 7b 28 ± 13c 33 ± 1.1b
Jonagold/M9-rolB2 41 ± 11d 12 ± 5b 17 ± 5d 23 ± 1.2c
Transgenic Res (2010) 19:933–948 939
123

with M9. For Elise, there was no consistent difference
in ascorbic acid when comparing the transgenic
rootstocks with the non-transgenic rootstocks
(Table 9). Total phenolics did not differ significantly
in either flesh or peels of Jonagold when comparing
the transgenic and non-transgenic rootstocks, while
the results on Elise and Elstar did not give a
consistent trend in the content of total phenolics
(Table 9).
Rooting of cuttings taken from the non-grafted
rolB transgenic rootstocks
In order to verify if the rolB effect on rooting is
stable under natural conditions, we took cuttings
from non-grafted transgenic rootstocks that were
planted together with the other field trial material
(Fig. 1). The results showed that the rooting
percentage was 73% for M26-rolB compared with
0% for M26, while 67 and 43% for M9-rolB1 and
M9-rolB2, respectively, compared to 3% for M9.
The rooting percentage of transgenic rootstocks was
significantly higher than their control counterparts
(Table 10).
Stability of transgene expression under field
conditions
In order to verify stability of transgene expression
under field conditions, RT-PCR was performed to
detect mRNA of the rolB gene in non-grafted
transgenic rootstocks that were planted together with
the other field trial material. The result showed that
the gene was expressed in all the three transgenic
clones grown in the field for several years, indicating
that the rolB gene is stably expressed (Fig. 2).
Table 4 Shoot number of
five apple cultivars grafted
onto transgenic and non-
transgenic apple rootstocks
Data were collected from 10
trees for each combination.
Duncan’s multiple test was
carried out among five
different rootstocks within
one cultivar. Different
letters in each column
within one cultivar indicate
significant differences at
P = 0.05
Combination
(cultivar/rootstock)
2005
(Mean ± SE)
2006
(Mean ± SE)
Average of 2 years
(mean ± SE)
Aroma/M26 17 ± 0.7a 26 ± 1.5a 22 ± 0.8a
Aroma/M26-rolB 12 ± 0.7b 15 ± 1.5b 14 ± 0.8b
Aroma/M9 18 ± 0.7a 22 ± 1.6a 20 ± 0.9a
Aroma/M9-rolB1 11 ± 0.7b 9 ± 1.6c 10 ± 0.8c
Aroma/M9-rolB2 9 ± 0.7c 7 ± 1.5c 8 ± 0.8d
Discovery/M26 14 ± 0.7a 18 ± 0.8a 16 ± 0.8a
Discovery/M26-rolB 10 ± 0.7b 9.4 ± 0.9c 10 ± 0.9b
Discovery/M9 13 ± 0.7a 15 ± 0.8b 14 ± 0.8a
Discovery/M9-rolB1 8 ± 0.7c 4 ± 0.8d 6 ± 0.8c
Discovery/M9-rolB2 7 ± 0.7c 0.9 ± 0.8e 4 ± 0.8c
Elise/M26 10 ± 0.6a 16 ± 1.2a 13 ± 0.8a
Elise/M26-rolB 6 ± 0.6b 8 ± 1.2b 7 ± 0.8bc
Elise/M9 9 ± 0.6b 14 ± 1.2a 12 ± 0.8a
Elise/M9-rolB1 5 ± 0.6b 7 ± 1.2bc 6 ± 0.9c
Elise/M9-rolB2 4 ± 0.6b 4 ± 1.2c 4 ± 0.8c
Elstar/M26 17 ± 0.8b 27 ± 1.3a 22 ± 0.8a
Elstar/M26-rolB 13 ± 0.8d 17 ± 1.3b 15 ± 0.8b
Elstar/M9 20 ± 0.8a 28 ± 1.4a 24 ± 0.9a
Elstar/M9-rolB1 15 ± 0.8c 18 ± 1.3b 17 ± 0.8b
Elstar/M9-rolB2 11 ± 0.8d 10 ± 1.4c 11 ± 0.9c
Jonagold/M26 14 ± 0.9a 23 ± 1.1a 19 ± 0.8a
Jonagold/M26-rolB 10 ± 0.8b 12 ± 1.1b 11 ± 0.8b
Jonagold/M9 15 ± 0.8a 19 ± 1.1a 17 ± 0.8a
Jonagold/M9-rolB1 9 ± 0.8b 10 ± 1.1b 10 ± 0.8b
Jonagold/M9-rolB2 7 ± 0.8b 5 ± 1.1c 6 ± 0.8c
940 Transgenic Res (2010) 19:933–948
123

Detection of the rolB gene for its presence
in the scion cultivars
PCR analysis was performed to investigate if the
transgene could be translocated from rootstock to
scion cultivar. No rolB band was detected in leaves or
flowers of the scion cultivars, suggesting that the
transgene was not translocated from rootstock to
scion (data not shown).
Detection of rolB mRNA in the scion cultivars
RT-PCR was conducted to check if the rolB mRNA
could be detected in the scion cultivars. No rolB
mRNA was detected in growing shoot tips of the
scion cultivars, indicating no translocation of the rolB
mRNA from rootstock to scion (Fig. 3).
Discussion
The rolB transgenic rootstocks significantly affect
growth of scions, but not the fruit quality
The effect of dwarfing rootstocks on scions has
already been studied extensively. In general, dwarfing
rootstocks can effectively control scion vigour and
reduce plant height, thus resulting in smaller trees
(Lauri et al. 2006; Tworkoski and Miller 2007). It has
also been reported that the growth rate of scion
cultivar can be independent of rootstock (Bulley et al.
2005). In this study, all growth parameters of the
scions grafted onto the transgenic rootstocks were
lower than those on the non-transgenic rootstocks
regardless of scion cultivar, indicating that the rolB
gene could reduce the growth rate of scion cultivars.
Table 5 Number of flower
buds of five apple cultivars
grafted onto transgenic and
non-transgenic apple
rootstocks
Data were collected from 10
trees for each combination.
Duncan’s multiple test was
carried out among five
different rootstocks within
one cultivar. Different
letters in each column
within one cultivar indicate
significant differences at
P = 0.05
Combination
(cultivar/rootstock)
2005
(Mean ± SE)
2006
(Mean ± SE)
2007
(Mean ± SE)
Average of 3 years
(mean ± SE)
Aroma/M26 3.3 ± 0.7a 3.3 ± 0.8c 113 ± 11a 40 ± 3.2a
Aroma/M26-rolB 2.2 ± 0.9a 3.8 ± 0.7b 81 ± 11bc 29 ± 3.9b
Aroma/M9 2.4 ± 0.8a 5.6 ± 0.8a 85 ± 12b 31 ± 3.8ab
Aroma/M9-rolB1 3.1 ± 0.7a 1.0 ± 0.8d 67 ± 11bc 24 ± 3.5bc
Aroma/M9-rolB2 2.2 ± 0.9a 0.7 ± 0.7d 48 ± 10c 17 ± 3.6c
Discovery/M26 5.1 ± 0.8a 6.3 ± 0.8a 91 ± 11a 34 ± 3.6a
Discovery/M26-rolB 2.8 ± 0.7b 3.1 ± 0.7c 63 ± 12b 23 ± 3.6b
Discovery/M9 5.8 ± 0.7a 3.6 ± 0.8b 72 ± 11b 27 ± 3.6ab
Discovery/M9-rolB1 2.0 ± 0.9b 0.6 ± 0.8d 45 ± 11c 16 ± 3.8c
Discovery/M9-rolB2 1.8 ± 0.7b 0.3 ± 0.7d 46 ± 11c 16 ± 3.4c
Elise/M26 5.7 ± 1.0a 4.2 ± 0.7a 77 ± 11a 29 ± 4.1a
Elise/M26-rolB 1.0 ± 1.2b 2.2 ± 0.8b 57 ± 11ab 20 ± 4.4b
Elise/M9 2.6 ± 0.9b 3.6 ± 0.8a 69 ± 11ab 25 ± 3.8ab
Elise/M9-rolB1 1.6 ± 0.9b 1.7 ± 0.7b 48 ± 12bc 17 ± 3.9bc
Elise/M9-rolB2 1.7 ± 0.8b 0.4 ± 0.7c 37 ± 10c 13 ± 3.5c
Elstar/M26 2.3 ± 1.0a 1.2 ± 0.7d 110 ± 11a 38 ± 3.5a
Elstar/M26-rolB 1.5 ± 1.2a 2.2 ± 0.8c 83 ± 11b 29 ± 3.5bc
Elstar/M9 3.0 ± 1.1a 2.7 ± 0.7b 108 ± 11a 38 ± 3.6a
Elstar/M9-rolB1 4.0 ± 1.0a 3.1 ± 0.8a 95 ± 11ab 34 ± 3.8ab
Elstar/M9-rolB2 2.5 ± 1.1a 1.0 ± 0.8e 57 ± 11c 20 ± 3.8c
Jonagold/M26 6.2 ± 1.0a 1.9 ± 0.7a 133 ± 11a 47 ± 3.5a
Jonagold/M26-rolB 2.5 ± 0.9ab 2.0 ± 0.8a 79 ± 11c 28 ± 3.8bc
Jonagold/M9 4.0 ± 0.9ab 2.7 ± 0.7a 107 ± 11b 38 ± 3.5ab
Jonagold/M9-rolB1 6.0 ± 0.8a 1.7 ± 0.7a 70 ± 11cd 26 ± 3.9c
Jonagold/M9-rolB2 0.1 ± 0.8b 0.3 ± 0.8b 30 ± 10d 10 ± 4.1d
Transgenic Res (2010) 19:933–948 941
123

This result is in line with the previous reports about
the rolB gene under greenhouse conditions (Zhu et al.
2001, 2003). A separate study carried out in a growth
unit with well-controlled climate conditions showed
that, under non-limiting nutrient conditions, the
relative growth rate of the rolB transformed apple
rootstock was not altered compared to the untrans-
formed control (Zhu and Welander 1999). The
explanation of this inconsistency could be that the
rolB transgenic rootstocks are likely to be more
sensitive to soil conditions than non-transgenic ones,
probably due to more hairy roots caused by the rolB
gene, which are mainly distributed on the soil
surface. Our current study clearly showed that the
flowering and fruiting were significantly reduced for
the cultivar Discovery grown on all the transgenic
rootstocks and the tendency is most dramatic for the
transgenic M9 rootstocks. This is likely due to the
significantly reduced tree growth. It has been reported
that yields are lower for scions grown on dwarfing
rootstocks compared with those on vigorous root-
stocks (Di Vaio et al. 2009).
Some studies have showed that dwarfing root-
stocks can affect the fruit quality (Autio et al. 1996;
Daugaard and Callesen 2002; Drake et al. 1988;
Fallahi and Kilby 1997; Fallahi et al. 1985), while
others reported that the fruit quality was not obvi-
ously affected by the rootstocks (Antognozzi et al.
1993; Riesen and Monney 1996). This inconsistency
may be attributed to differences in yield, choice of
cultivar and rootstock combinations, climate and
cultivation. In this study, the fruit quality was
generally not affected by the transgenic rootstocks,
indicating that the rolB transgenic rootstocks do not
clearly affect the fruit quality of non-transgenic scion
cultivars. Since our results on fruit quality analysis
Table 6 Number of
flowers of five apple
cultivars grafted onto
transgenic and non-
transgenic apple rootstocks
Data were collected from 10
trees for each combination.
Duncan’s multiple test was
carried out among five
different rootstocks within
one cultivar. Different
letters in each column
within one cultivar indicate
significant differences at
P = 0.05
Combination
(cultivar/rootstock)
2005
(Mean ± SE)
2006
(Mean ± SE)
2007
(Mean ± SE)
Average of
3 years
(mean ± SE)
Aroma/M26 7.1 ± 2.1ab 14 ± 2.3b 567 ± 56a 196 ± 16.4a
Aroma/M26-rolB 5.7 ± 3.1ab 17 ± 0.7ab 406 ± 56bc 143 ± 19.7ab
Aroma/M9 5.1 ± 2.7b 27 ± 0.7a 436 ± 60b 156 ± 18.8a
Aroma/M9-rolB1 9.4 ± 2.4a 7 ± 0.7bc 338 ± 56bc 118 ± 17.6bc
Aroma/M9-rolB2 5.7 ± 2.8ab 3 ± 0.7c 243 ± 50c 84 ± 18.5c
Discovery/M26 24.4 ± 2.5a 31 ± 0.7a 458 ± 56a 171 ± 18.0a
Discovery/M26-rolB 13.9 ± 2.3b 14 ± 0.7b 323 ± 60b 117 ± 17.8bc
Discovery/M9 32.6 ± 2.6a 17 ± 0.7b 359 ± 56b 136 ± 17.8ab
Discovery/M9-rolB1 11.0 ± 2.3b 2.9 ± 0.7c 232 ± 56c 82 ± 19.0c
Discovery/M9-rolB2 9.3 ± 0.7b 1.5 ± 0.7c 232 ± 53c 81 ± 17.0c
Elise/M26 18.7 ± 3.5a 18.8 ± 0.7a 385 ± 56a 141 ± 20.7a
Elise/M26-rolB 4.7 ± 4.2b 11 ± 0.7ab 287 ± 56bc 101 ± 21.9ab
Elise/M9 10.3 ± 2.9b 15.6 ± 0.7a 343 ± 56ab 123 ± 18.8a
Elise/M9-rolB1 7.9 ± 2.9b 6.5 ± 0.7ab 237 ± 60cb 84 ± 19.7b
Elise/M9-rolB2 6.5 ± 2.5b 0.8 ± 0.7b 182 ± 50c 63 ± 17.5b
Elstar/M26 6.4 ± 2.4a 6.4 ± 0.7b 554 ± 56a 189 ± 17.6a
Elstar/M26-rolB 4.0 ± 2.4a 10.2 ± 0.7ab 412 ± 56b 142 ± 17.6ab
Elstar/M9 7.3 ± 2.5a 14.1 ± 0.7ab 543 ± 56a 188 ± 18.0a
Elstar/M9-rolB1 7.9 ± 2.8a 15.7 ± 0.7a 477 ± 56ab 167 ± 19.1b
Elstar/M9-rolB2 5.0 ± 2.8a 5.2 ± 0.7b 278 ± 53c 96 ± 19.1b
Jonagold/M26 18.0 ± 2.4a 8 ± 0.7ab 661 ± 56a 229 ± 17.6a
Jonagold/M26-rolB 9.7 ± 2.8ab 9.3 ± 0.7ab 398 ± 56c 139 ± 19.1bc
Jonagold/M9 11.0 ± 2.4ab 11.7 ± 0.7a 526 ± 56b 183 ± 17.8ab
Jonagold/M9-rolB1 16.7 ± 2.8a 6.1 ± 0.7ab 352 ± 56cd 125 ± 19.4c
Jonagold/M9-rolB2 0.7 ± 4.1b 1.5 ± 0.7b 154 ± 50d 52 ± 20.6d
942 Transgenic Res (2010) 19:933–948
123

are based mainly on 1 year study, more studies may
be needed for a final conclusion. To our knowledge,
this study is the first report regarding the effect of
GM rootstocks on non-transgenic scion cultivars in
fruit trees under field conditions.
Expression of the rolB gene under field conditions
Stable expression of transgenes under field conditions
has been demonstrated in various species, such as
poplar (Li et al. 2008; Meilan et al. 2002), sunflower
(Rousselin et al. 2002), pineapple (Sripaoraya et al.
2006) and soybean (Padgette et al. 1995). However,
loss of transgene expression does occur, as has been
demonstrated in aspen (Kumar and Fladung 2001),
tall fescue (Bettany et al. 1998), oilseed rape (Metz
et al. 1997), oat (Pawlowski et al. 1998) and many
other species. The loss of transgene expression can be
due either to deletion of the transgene itself (Cherd-
shewasart et al. 1993; Heberlebors et al. 1988;
Tencate et al. 1990), or to epigenetic silencing of
the transgene. Silencing can occur at the transcrip-
tional level or at the posttranscriptional level (for
reviews, see Baulcombe 2004; Brodersen and Voin-
net 2009). Since apple trees have a long life cycle, the
stable expression of transgenes is particularly impor-
tant for potential commercialisation of GM root-
stocks. The results on cutting and RT-PCR in this
study showed that the rolB gene is stably expressed in
the transgenic rootstocks under field conditions over a
period of several years. Since this result was only
based on three transgenic clones, further studies with
more transgenic clones over even longer period are
required to draw final conclusion.
The rooting percentage of cuttings of the trans-
genic rolB rootstocks is lower than the rooting
Table 7 Number of fruits
of five apple cultivars
grafted onto transgenic and
non-transgenic apple
rootstocks
Data were collected from 10
trees for each combination.
Duncan’s multiple test was
carried out among five
different rootstocks within
one cultivar. Different
letters in each column
within one cultivar indicate
significant differences at
P = 0.05
Combination
(cultivar/rootstock)
2007
(Mean ± SE)
2008
(Mean ± SE)
2009
(Mean ± SE)
Average of
3 years
(mean ± SE)
Aroma/M26 20 ± 5.4a 14 ± 5.8c 47 ± 7.8a 27 ± 4.9ab
Aroma/M26-rolB 5.8 ± 5.8a 16 ± 5.8c 32 ± 7.0ab 18 ± 5.1ab
Aroma/M9 13.7 ± 5.9a 56 ± 5.8a 23 ± 7.5ab 31 ± 5.2a
Aroma/M9-rolB1 7.3 ± 5.9a 48 ± 5.8ab 22 ± 7.0ab 26 ± 5.1ab
Aroma/M9-rolB2 5.6 ± 5.9a 31 ± 5.8bc 11 ± 7.5b 16 ± 5.1b
Discovery/M26 2.4 ± 5.9c 32 ± 5.8ab 85 ± 7.8a 40 ± 5.1a
Discovery/M26-rolB 1.6 ± 5.9c 15 ± 5.8c 56 ± 7.5bc 24 ± 5.4b
Discovery/M9 13 ± 5.9a 34 ± 5.8a 78 ± 7.5ab 42 ± 5.1a
Discovery/M9-rolB1 3.2 ± 5.9c 34 ± 5.8a 30 ± 7.6cd 22 ± 5.2bc
Discovery/M9-rolB2 6.9 ± 5.9b 20 ± 5.8bc 20 ± 5.9d 16 ± 5.1c
Elise/M26 26 ± 5.9ab 43 ± 5.8b 44 ± 6.7a 38 ± 5.1ab
Elise/M26-rolB 14 ± 5.9b 37 ± 5.8b 18 ± 6.7b 23 ± 5.1c
Elise/M9 35 ± 5.9a 56 ± 5.8ab 56 ± 6.7a 49 ± 5.1a
Elise/M9-rolB1 9 ± 6.2b 79 ± 6.1a 13 ± 6.7b 34 ± 5.4bc
Elise/M9-rolB2 8.5 ± 5.9b 44 ± 5.8b 11 ± 6.7b 21 ± 5.1c
Elstar/M26 66 ± 6.2a 22 ± 5.8ab 57 ± 6.5a 48 ± 5.4a
Elstar/M26-rolB 41 ± 5.9b 13 ± 5.8b 30 ± 6.5a 28 ± 5.1cd
Elstar/M9 65 ± 5.9a 34 ± 5.8a 42 ± 6.5a 47 ± 5.2ab
Elstar/M9-rolB1 26 ± 5.9bc 33 ± 5.8a 39 ± 6.5a 33 ± 5.1bc
Elstar/M9-rolB2 13 ± 6.2c 18 ± 6.1b 15 ± 6.5a 15 ± 5.3d
Jonagold/M26 27 ± 5.9ab 19 ± 5.8a 60 ± 7.8a 35 ± 5.1a
Jonagold/M26-rolB 17 ± 5.9abc 12 ± 5.8ab 41 ± 7.8ab 23 ± 4.9b
Jonagold/M9 36 ± 5.9a 19 ± 5.8a 64 ± 7.8a 40 ± 5.1a
Jonagold/M9-rolB1 14 ± 5.9bc 17 ± 5.8ab 27 ± 7.8bc 19 ± 5.2bc
Jonagold/M9-rolB2 7 ± 5.9c 10 ± 5.8b 9 ± 7.8c 9 ± 5.1c
Transgenic Res (2010) 19:933–948 943
123
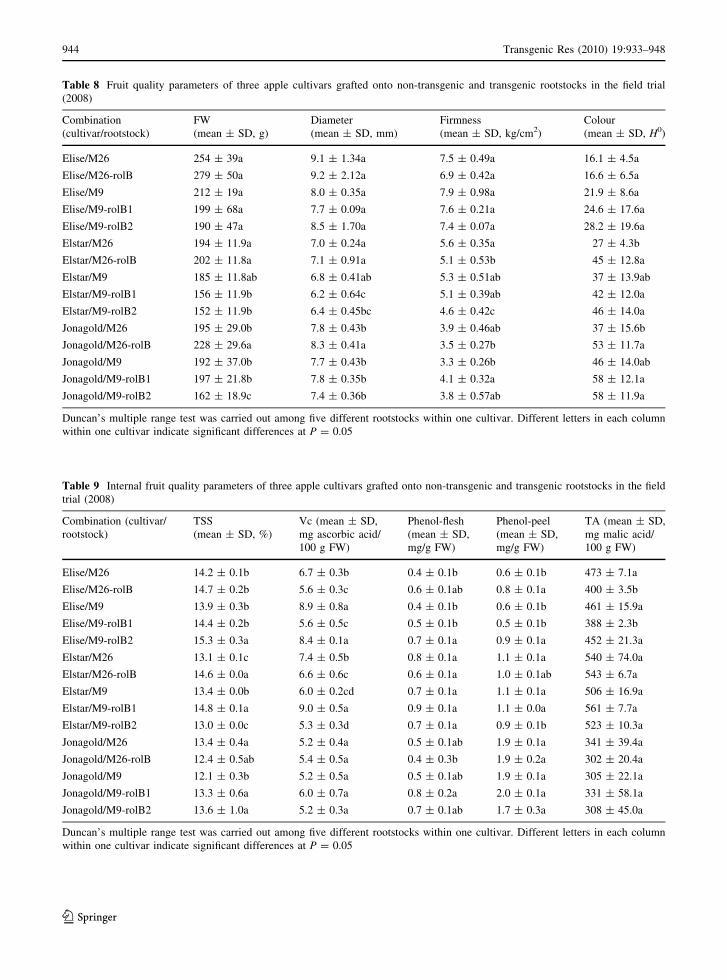
Table 8 Fruit quality parameters of three apple cultivars grafted onto non-transgenic and transgenic rootstocks in the field trial
(2008)
Combination
(cultivar/rootstock)
FW
(mean ± SD, g)
Diameter
(mean ± SD, mm)
Firmness
(mean ± SD, kg/cm2)
Colour
(mean ± SD, H0)
Elise/M26 254 ± 39a 9.1 ± 1.34a 7.5 ± 0.49a 16.1 ± 4.5a
Elise/M26-rolB 279 ± 50a 9.2 ± 2.12a 6.9 ± 0.42a 16.6 ± 6.5a
Elise/M9 212 ± 19a 8.0 ± 0.35a 7.9 ± 0.98a 21.9 ± 8.6a
Elise/M9-rolB1 199 ± 68a 7.7 ± 0.09a 7.6 ± 0.21a 24.6 ± 17.6a
Elise/M9-rolB2 190 ± 47a 8.5 ± 1.70a 7.4 ± 0.07a 28.2 ± 19.6a
Elstar/M26 194 ± 11.9a 7.0 ± 0.24a 5.6 ± 0.35a 27 ± 4.3b
Elstar/M26-rolB 202 ± 11.8a 7.1 ± 0.91a 5.1 ± 0.53b 45 ± 12.8a
Elstar/M9 185 ± 11.8ab 6.8 ± 0.41ab 5.3 ± 0.51ab 37 ± 13.9ab
Elstar/M9-rolB1 156 ± 11.9b 6.2 ± 0.64c 5.1 ± 0.39ab 42 ± 12.0a
Elstar/M9-rolB2 152 ± 11.9b 6.4 ± 0.45bc 4.6 ± 0.42c 46 ± 14.0a
Jonagold/M26 195 ± 29.0b 7.8 ± 0.43b 3.9 ± 0.46ab 37 ± 15.6b
Jonagold/M26-rolB 228 ± 29.6a 8.3 ± 0.41a 3.5 ± 0.27b 53 ± 11.7a
Jonagold/M9 192 ± 37.0b 7.7 ± 0.43b 3.3 ± 0.26b 46 ± 14.0ab
Jonagold/M9-rolB1 197 ± 21.8b 7.8 ± 0.35b 4.1 ± 0.32a 58 ± 12.1a
Jonagold/M9-rolB2 162 ± 18.9c 7.4 ± 0.36b 3.8 ± 0.57ab 58 ± 11.9a
Duncan’s multiple range test was carried out among five different rootstocks within one cultivar. Different letters in each column
within one cultivar indicate significant differences at P = 0.05
Table 9 Internal fruit quality parameters of three apple cultivars grafted onto non-transgenic and transgenic rootstocks in the field
trial (2008)
Combination (cultivar/
rootstock)
TSS
(mean ± SD, %)
Vc (mean ± SD,
mg ascorbic acid/
100 g FW)
Phenol-flesh
(mean ± SD,
mg/g FW)
Phenol-peel
(mean ± SD,
mg/g FW)
TA (mean ± SD,
mg malic acid/
100 g FW)
Elise/M26 14.2 ± 0.1b 6.7 ± 0.3b 0.4 ± 0.1b 0.6 ± 0.1b 473 ± 7.1a
Elise/M26-rolB 14.7 ± 0.2b 5.6 ± 0.3c 0.6 ± 0.1ab 0.8 ± 0.1a 400 ± 3.5b
Elise/M9 13.9 ± 0.3b 8.9 ± 0.8a 0.4 ± 0.1b 0.6 ± 0.1b 461 ± 15.9a
Elise/M9-rolB1 14.4 ± 0.2b 5.6 ± 0.5c 0.5 ± 0.1b 0.5 ± 0.1b 388 ± 2.3b
Elise/M9-rolB2 15.3 ± 0.3a 8.4 ± 0.1a 0.7 ± 0.1a 0.9 ± 0.1a 452 ± 21.3a
Elstar/M26 13.1 ± 0.1c 7.4 ± 0.5b 0.8 ± 0.1a 1.1 ± 0.1a 540 ± 74.0a
Elstar/M26-rolB 14.6 ± 0.0a 6.6 ± 0.6c 0.6 ± 0.1a 1.0 ± 0.1ab 543 ± 6.7a
Elstar/M9 13.4 ± 0.0b 6.0 ± 0.2cd 0.7 ± 0.1a 1.1 ± 0.1a 506 ± 16.9a
Elstar/M9-rolB1 14.8 ± 0.1a 9.0 ± 0.5a 0.9 ± 0.1a 1.1 ± 0.0a 561 ± 7.7a
Elstar/M9-rolB2 13.0 ± 0.0c 5.3 ± 0.3d 0.7 ± 0.1a 0.9 ± 0.1b 523 ± 10.3a
Jonagold/M26 13.4 ± 0.4a 5.2 ± 0.4a 0.5 ± 0.1ab 1.9 ± 0.1a 341 ± 39.4a
Jonagold/M26-rolB 12.4 ± 0.5ab 5.4 ± 0.5a 0.4 ± 0.3b 1.9 ± 0.2a 302 ± 20.4a
Jonagold/M9 12.1 ± 0.3b 5.2 ± 0.5a 0.5 ± 0.1ab 1.9 ± 0.1a 305 ± 22.1a
Jonagold/M9-rolB1 13.3 ± 0.6a 6.0 ± 0.7a 0.8 ± 0.2a 2.0 ± 0.1a 331 ± 58.1a
Jonagold/M9-rolB2 13.6 ± 1.0a 5.2 ± 0.3a 0.7 ± 0.1ab 1.7 ± 0.3a 308 ± 45.0a
Duncan’s multiple range test was carried out among five different rootstocks within one cultivar. Different letters in each column
within one cultivar indicate significant differences at P = 0.05
944 Transgenic Res (2010) 19:933–948
123

percentage of the same rootstocks reported earlier
from in vitro rooting (Welander and Zhu 2000; Zhu
et al. 2001). This is probably due to weak growth of
the transgenic rootstocks under field conditions.
When the cuttings were made, the annual growth
was limited, especially for the transgenic M9 root-
stocks. A weak growth would limit accumulation of
nitrogen and/or carbohydrates in stems, which in turn
might lead to a low rooting percentage as has been
demonstrated for many species (Hambrick et al.
1991; Rapaka et al. 2005; Zerche and Druege 2009).
No translocation of the rolB gene or its mRNA
from rootstock to scion
The results from investigations on translocation of
transgenes and their products from rootstock to scion
reported in the literature have been inconsistent. It
has been reported that graft-transmissible signals can
be translocated from stock to scion (Garcia-Perez
et al. 2004; Kudo and Harada 2007; Tournier et al.
2006). The influence between rootstock and scion is
in some cases mutual, for example a rootstock
transformed with a construct for gene silencing could
induce gene-specific silencing in an originally non-
silenced scion (Palauqui et al. 1997; Voinnet et al.
1998), while the silencing effect could also be
transmitted from scion to rootstock (Sonoda and
Nishiguchi 2000; Tournier et al. 2006). The trans-
ported products could be small RNAs (siRNAs;
Hewezi et al. 2005; Tournier et al. 2006; Yoo et al.
2004), mRNAs (Kim et al. 2001), and proteins
(Golecki et al. 1999; Gomez et al. 2005). However,
Ayre and Turgeon (2004) could not detect transmis-
sion of the CO (CONSTANS) gene or its transcripts
from stock to scion in Arabidopsis thaliana. In this
study, PCR and RT-PCR could not detect the rolB
gene or its mRNA in scion cultivar, suggesting no
Fig. 1 Rooting of cuttings of rolB transgenic apple rootstocks
and non-transgenic controls
Table 10 Rooting results of cuttings taken from the rootstocks
grown in field
Rootstock Total cuttings Rooted cuttings Rooting %
M26 30 0 0c
M26-rolB 30 22 73a
M9 30 1 3c
M9-rolB1 30 20 67a
M9-rolB2 30 13 43b
Cutting was carried out in greenhouse without auxin treatment.
Different letters in the root% column indicate significant
differences at P = 0.05
Fig. 2 RT-PCR of rolB mRNA in non-grafted rolB transgenic
rootstocks. M markers, P plasmid DNA, B blank, 1 M26-rolB,
2 M9-rolB1, 3 M9-rolB2
Fig. 3 RT-PCR of rolB mRNA in scion cultivars. M markers,
P plasmid DNA, B blank, 1–5 Jonagold and 6–10 Elise, grafted
onto M26, M26-rolB, M9, M9-rolB1, M9-rolB2 rootstocks,
respectively
Transgenic Res (2010) 19:933–948 945
123

translocation of the rolB gene and its mRNA from
rootstock to scion. However, it is unclear if small
RNAs could be translocated from rootstock and
sicon. It also remains uncertain if the ROLB protein
could be translocated from rootstock into scion. We
have made several attempts for producing ROLB
antibodies for Western blot analysis without success.
We have encountered great difficulties in purifying
the recombinant ROLB protein and ROLB antibod-
ies, probably because the ROLB protein most likely
is a membrane protein (Filippini et al. 1996). The
extremely low sensitivity of the ROLB antibodies
made detection of ROLB protein at nanogram levels
from plant material impossible (Smolka et al., unpub-
lished data). It has been reported that the phloem
provides a pathway for the transport of small and
macromolecules including soluble proteins (Bor-
tolotti et al. 2005). As mentioned above, the ROLB
protein is not water soluble and may not be possible
or easy to transport from rootstock to scion. However,
this needs to be proven.
The perspective of using the rolB gene
in modifying horticultural crops
Over the years, there have been many attempts to
utilise the rolB gene for improving the rooting ability
in a number of plant species and the results have been
satisfactory in most cases. However, there is so far no
report about how the rolB gene will affect growth and
development under field conditions over a long
period of time. It has been unclear if the rolB
transgenic plants can be of commercial interest for
fruit growers. Our current study shows that the rolB
gene indeed reduced growth and flowering as well as
fruiting of non-transgenic scion cultivars, especially
cultivars with weak growth vigour. This reduction
might be positive for vigorous cultivars in commer-
cial production where thinning is often required when
excessive flowering occurs. From our results, we
recommend using rolB modified rootstocks in com-
bination with vigorous scion cultivars or dwarfing
vigorous rootstocks with the rolB gene.
Another potential use of the rolB gene may be for
making bonzai plants of fruit trees and ornamentals
where extremely slow growth rates are desirable.
Acknowledgments We thank Annelie Ahlman for her
excellent technical support, Carin Emanuelsson and Miao
Zhou for collecting some data and Karl-Erik Gustavsson for
support with the HPLC analysis. FORMAS, The Royal
Physiographic Society in Lund, Nilsson’s fund are acknow-
ledged for financial support.
References
Aldwinckle HS, Borejsza-Wysocka EE, Malnoy M, Brown SK,
Norelli JL, Beer SV, Meng X, He SY, Jin Q-L (2003)
Development of fire blight resistant apple cultivars by
genetic engineering. Acta Hortic 622:105–111
Altamura MM, Capitani F, Gazza L, Capone I, Costantino P
(1994) The plant oncogene rolb stimulates the formation
of flower and root meristemoids in tobacco thin cell-lay-
ers. New Phytol 126(2):283–293
Antognozzi E, Proiett P, Famiani F (1993) Effect of rootstocks
and training systems on growth and yield of two apple
cultivars. Acta Hortic 349:187–190
Autio WR, Hayden RA, Micke WC, Brown GR (1996) Root-
stock affects ripening, color, and shape of ‘starkspur
supreme delicious’ apples in the 1984 NC-140 coopera-
tive planting. Fruit Varieties J 50(1):45–53
Ayre BG, Turgeon R (2004) Graft transmission of a floral
stimulant derived from CONSTANS. Plant Physiol
135(4):2271–2278
Baulcombe D (2004) RNA silencing in plants. Nature
431(7006):356–363
Bettany AJE, Dalton SJ, Timms E, Morris P (1998) Stability of
transgene expression during vegetative propagation of
protoplast-derived tall fescue (Festuca arundinacea
Schreb.) plants. J Exp Bot 49(328):1797–1804
Bortolotti C, Murillo I, Fontanet P, Coca M, Segundo BS
(2005) Long-distance transport of the maize pathogenesis-
related PRms protein through the phloem in transgenic
tobacco plants. Plant Sci 168(3):813–821
Brodersen P, Voinnet O (2009) Revisiting the principles of
microRNA target recognition and mode of action. Nat
Rev Mol Cell Biol 10(2):141–148
Bulley SM, Wilson FM, Hedden P, Phillips AL, Croker SJ,
James DJ (2005) Modification of gibberellin biosynthesis
in the grafted apple scion allows control of tree height
independent of the rootstock. Plant Biotechnol J 3(2):215–
223
Capone I, Spano L, Cardarelli M, Bellincampi D, Petit A,
Costantino P (1989) Induction and growth-properties of
carrot roots with different complements of Agrobacte-rium-rhizogenes T-DNA. Plant Mol Biol 13(1):43–52
Cardarelli M, Mariotti D, Pomponi M, Spano L, Capone I,
Costantino P (1987) Agrobacterium-rhizogenes T-DNA
genes capable of inducing hairy root phenotype. Mol Gen
Genet 209(3):475–480
Cherdshewasart W, Ghartichhetri GB, Saul MW, Jacobs M,
Negrutiu I (1993) Expression instability and genetic-dis-
orders in transgenic Nicotiana-plumbaginifolia l plants.
Transgenic Res 2(6):307–320
Dai WH, Cheng ZM, Sargent WA (2004) Expression of the
rolB gene enhances adventitious root formation in hard-
wood cuttings of aspen. In Vitro Cel Dev Biol Plant
40(4):366–370
946 Transgenic Res (2010) 19:933–948
123

Dandekar AM, Teo G, Defilippi BG, Uratsu SL, Passey AJ,
Kader AA, Stow JR, Colgan RJ, James DJ (2004) Effect
of down-regulation of ethylene biosynthesis on fruit flavor
complex in apple fruit. Transgenic Res 13(4):373–384
Daugaard H, Callesen O (2002) The effect of rootstock on
yield and quality of ‘Mutsu’ apples. J Hortic Sci Bio-
technol 77(2):248–251
Dewanto V, Wu XZ, Liu RH (2002) Processed sweet corn has
higher antioxidant activity. J Agric Food Chem
50(17):4959–4964
Di Vaio C, Cirillo C, Buccheri M, Limongelli F (2009) Effect
of interstock (M.9 and M.27) on vegetative growth and
yield of apple trees (cv ‘‘Annurca’’). Sci Hortic
119(3):270–274
Drake SR, Larsen FE, Fellman JK, Higgins SS (1988) Matu-
rity, storage quality, carbohydrate, and mineral-content of
Goldspur apples as influenced by rootstock. J Am Soc
Hortic Sci 113(6):949–952
Dutt M, Li ZT, Kelley KT, Dhekney SA, Van Aman M, Tattersall
J, Gray DJ (2007) Transgenic rootstock protein transmis-
sion in grapevines. Acta Hort (ISHS) 738:749–754
Fallahi E, Kilby M (1997) Rootstock and pruning influence on
yield and fruit quality of ‘Lisbon’ lemon. Fruit Varieties J
51(4):242–246
Fallahi E, Richardson DG, Westwood MN (1985) Quality of
apple fruit from a high-density orchard as influenced by
rootstocks, fertilizers, maturity, and storage. J Am Soc
Hortic Sci 110(1):71–74
Ferree DC, Erb WA, Morrison FA (2001a) Influence of four
apple cultivars on five dwarfing rootstocks on spur qual-
ity. J Am Pomol Soc 55(3):166–172
Ferree DC, Erb WA, Morrison FD (2001b) Influence of four
apple cultivars on five dwarfing rootstocks on morphology
of two-year-old limb sections. J Am Pomol Soc
55(3):159–165
Feyissa T, Zhu LH, Negash L, Welander M (2007) Regener-
ation and genetic transformation of Hagenia abyssinica(Bruce) JF Gmel. (Rosaceae) with rolB gene. Plant Cell
Tiss Organ Cult 88(3):277–288
Filippini F, Rossi V, Marin O, Trovato M, Costantino P,
Downey PM, LoSchiavo F, Terzi M (1996) A plant
oncogene as a phosphatase. Nature 379(6565):499–500
Garcia-Perez RD, Van Houdt H, Depicker A (2004) Spreading
of post-transcriptional gene silencing along the target
gene promotes systemic silencing. Plant J 38(4):594–602
Geier T, Eimert K, Scherer R, Nickel C (2008) Production and
rooting behaviour of rolB-transgenic plants of grape
rootstock ‘Richter 110’ (Vitis berlandieri X V-rupestris).
Springer, New York, pp 269–280
Golecki B, Schulz A, Thompson GA (1999) Translocation of
structural P proteins in the phloem. Plant Cell 11(1):127–140
Gomez G, Torres H, Pallas V (2005) Identification of trans-
locatable RNA-binding phloem proteins from melon,
potential components of the long-distance RNA transport
system. Plant J 41(1):107–116
Hambrick CE, Davies FT, Pemberton HB (1991) Seasonal
changes in carbohydrate nitrogen levels during field
rooting of Rosa-multiflora Brooks 56 hardwood cuttings.
Sci Hortic 46(1–2):137–146
Heberlebors E, Charvat B, Thompson D, Schernthaner JP,
Barta A, Matzke AJM, Matzke MA (1988) Genetic-
analysis of T-DNA insertions into the tobacco genome.
Plant Cell Rep 7(7):571–574
Hewezi T, Alibert G, Kallerhoff J (2005) Local infiltration of
high- and low-molecular-weight RNA from silenced
sunflower (Helianthus annuus L.) plants triggers post-
transcriptional gene silencing in non-silenced plants. Plant
Biotechnol J 3(1):81–89
Hirst PM, Ferree DC (1995) Rootstock effects on the flowering
of Delicious apple. 1. Bud development. J Am Soc Hortic
Sci 120(6):1010–1017
Hirst PM, Ferree DC (1996) Effects of rootstock on bud
development and flower formation of ‘starkspur supreme
delicious’ apple. Fruit Varieties J 50(1):25–34
Hrazdina G, Kiss E, Galli Z, RosenField C, Norelli JL, Ald-
winckle HS (2003) Down regulation of ethylene produc-
tion in ‘royal gala’ apples. Acta Hortic 628:239–251
James C (2008) Global status of commercialized biotech/GM
crops: 2008. International Service for the Acquisition of
Agri-biotech Applications, Ithaca
Kim M, Canio W, Kessler S, Sinha N (2001) Developmental
changes due to long-distance movement of a homeobox
fusion transcript in tomato. Science 293(5528):287–289
Kudo H, Harada T (2007) A graft-transmissible RNA from
tomato rootstock changes leaf morphology of potato
scion. Hortscience 42(2):225–226
Kumar S, Fladung M (2001) Gene stability in transgenic aspen
(Populus). II. Molecular characterization of variable
expression of transgene in wild and hybrid aspen. Planta
213(5):731–740
Lauri PE, Maguylo K, Trottier C (2006) Architecture and size
relations: an essay on the apple (Malus x domestica,
Rosaceae) tree. Am J Bot 93(3):357–368
Li JL, Meilan R, Ma C, Barish M, Strauss SH (2008) Stability
of herbicide resistance over 8 years of coppice in field-
grown, genetically engineered poplars. West J Appl For
23(2):89–93
Meilan R, Auerbach DJ, Ma C, DiFazio SP, Strauss SH (2002)
Stability of herbicide resistance and GUS expression in
transgenic hybrid poplars (Populus sp.) during four years
of field trials and vegetative propagation. Hortscience
37(2):277–280
Metz PLJ, Jacobsen E, Stiekema WJ (1997) Occasional loss of
expression of phosphinothricin tolerance in sexual off-
spring of transgenic oilseed rape (Brassica napus L.).
Euphytica 98(3):189–196
Norelli JL, Boresza-Wysocka E, Reynoird JP, Aldwinckle HS
(2003) Transgenic ‘royal gala’ apple expressing attacin e
has increased field resistance to erwinia amylovora (fire
blight). Acta Hortic 538:631–633
Padgette SR, Kolacz KH, Delannay X, Re DB, Lavallee BJ, Tinius
CN, Rhodes WK, Otero YI, Barry GF, Eichholtz DA, Pes-
chke VM, Nida DL, Taylor NB, Kishore GM (1995)
Development, identification, and characterization of a gly-
phosate-tolerant soybean line. Crop Sci 35(5):1451–1461
Palauqui JC, Elmayan T, Pollien JM, Vaucheret H (1997) Sys-
temic acquired silencing: transgene-specific post-transcrip-
tional silencing is transmitted by grafting from silenced
stocks to non-silenced scions. EMBO J 16(15):4738–4745
Pawlowski WP, Torbert KA, Rines HW, Somers DA (1998)
Irregular patterns of transgene silencing in allohexaploid
oat. Plant Mol Biol 38(4):597–607
Transgenic Res (2010) 19:933–948 947
123

Rapaka VK, Bessler B, Schreiner M, Druege U (2005) Inter-
play between initial carbohydrate availability, current
photosynthesis and adventitious root formation in Pelar-
gonium cuttings. Plant Sci 168(6):1547–1560
Riesen W, Monney P (1996) Apfelunterlagen: Zwische-
nergebnisse deutsch- und westschweizerischer Versuche.
Obst- und Weinbau 132(21):548–552
Rousselin P, Molinier J, Himber C, Schontz D, Prieto-Dapena
P, Jordano J, Martini N, Weber S, Horn R, Ganssmann M,
Grison R, Pagniez M, Toppan A, Friedt W, Hahne G
(2002) Modification of sunflower oil quality by seed-
specific expression of a heterologous Delta 9-stearoyl-
(acyl carrier protein) desaturase gene. Plant Breed
121(2):108–116
Rugini E, Pellegrineschi A, Mencuccini M, Mariotti D (1991)
Increase of rooting ability in the woody species kiwi
(Actinidia-deliciosa a chev) by transformation with
Agrobacterium-rhizogenes-rol genes. Plant Cell Rep
10(6–7):291–295
Sonoda S, Nishiguchi M (2000) Graft transmission of post-
transcriptional gene silencing: target specificity for RNA
degradation is transmissible between silenced, non-
silenced plants, but not between silenced plants (vol 21,
pg 1, 2000). Plant J 22(3):275
Spena A, Schmulling T, Koncz C, Schell JS (1987) Indepen-
dent and synergistic activity of rol-a, rol-b and rol-c loci
in stimulating abnormal growth in plants. EMBO J
6(13):3891–3899
Sripaoraya S, Keawsompong S, Insupa P, Power JB, Davey MR,
Srinives P (2006) Genetically manipulated pineapple:
transgene stability, gene expression and herbicide tolerance
under field conditions. Plant Breed 125(4):411–413
Tencate CHH, Loonen A, Ottaviani MP, Ennik L, Vaneldik G,
Stiekema WJ (1990) Frequent spontaneous deletions of Ri
T-DNA in Agrobacterium-rhizogenes transformed potato
roots and regenerated plants. Plant Mol Biol 14(5):735–
741
Tournier B, Tabler M, Kalantidis K (2006) Phloem flow
strongly influences the systemic spread of silencing in
GFP Nicotiana benthamiana plants. Plant J 47(3):383–394
Tubbs FR (1974) Rootstock/scion relations in horticultural
crop physiology. Sci Hortic 2(3):221–230
Tworkoski T, Miller S (2007) Rootstock effect on growth of
apple scions with different growth habits. Sci Hortic
111(4):335–343
Tzfira T, Yarnitzky O, Vainstein A, Altman A (1996) Agro-
bacterium rhizogenes-mediated DNA transfer in Pinushalepensis Mill. Plant Cell Rep 16(1–2):26–31
Vilaine F, Cassedelbart F (1987) A new vector derived from
Agrobacterium-rhizogenes plasmids—a micro-Ri plasmid
and its use to construct a mini-Ri plasmid. Gene
55(1):105–114
Voinnet O, Vain P, Angell S, Baulcombe DC (1998) Systemic
spread of sequence-specific transgene RNA degradation in
plants is initiated by localized introduction of ectopic
promoterless DNA. Cell 95(2):177–187
Webster AD, Oehl VH, Jackson JE, Jones OP (1985) The
orchard establishment, growth and precocity of 4 micro-
propagated apple scion cultivars. J Hortic Sci 60(2):169–
180
Welander M, Zhu LH (2000) The rooting ability of rolb
transformed clones of the apple rootstock M26 and its
relation to gene expression. Acta Hort (ISHS) 521:133–
138
Welander M, Pawlicki N, Holefors A, Wilson F (1998) Genetic
transformation of the apple rootstock M26 with the RolB
gene and its influence on rooting. J Plant Physiol 153(3–4):
371–380
Yoo BC, Kragler F, Varkonyi-Gasic E, Haywood V, Archer-
Evans S, Lee YM, Lough TJ, Lucas WJ (2004) A sys-
temic small RNA signaling system in plants. Plant Cell
16(8):1979–2000
Youk ES, Pack IS, Kim YJ, Yoon WK, Kim CG, Ryu SB, Harn
CH, Jeong SC, Kim HM (2009) A framework for
molecular genetic assessment of a transgenic watermelon
rootstock line. Plant Sci 176(6):805–811
Zerche S, Druege U (2009) Nitrogen content determines
adventitious rooting in Euphorbia pulcherrima under
adequate light independently of pre-rooting carbohydrate
depletion of cuttings. Sci Hortic 121(3):340–347
Zhu LH, Welander M (1999) Growth characteristics of apple
cultivar Gravenstein plants grafted onto the transformed
rootstock M26 with rolA and rolB genes under non-lim-
iting nutrient conditions. Plant Sci 147(1):75–80
Zhu LH, Holefors A, Ahlman A, Xue ZT, Welander M (2001)
Transformation of the apple rootstock M.9/29 with the
rolB gene and its influence on rooting and growth. Plant
Sci 160(3):433–439
Zhu LH, Li XY, Ahlman A, Welander M (2003) The rooting
ability of the dwarfing pear rootstock BP10030 (Pyrus
communis) was significantly increased by introduction of
the rolB gene. Plant Sci 165(4):829–835
Zhu LH, Li XY, Welander M (2008) Overexpression of the
Arabidopsis gai gene in apple significantly reduces plant
size. Plant Cell Rep 27(2):289–296
948 Transgenic Res (2010) 19:933–948
123
Copyright © 2022 FDOKUMEN




















