
Rootstock control of scion response to water stress in grapevine
7
Environmental and Experimental Botany 93 (2013) 20–26 Contents lists available at SciVerse ScienceDirect Environmental and Experimental Botany journal h om epa ge : www.elsevier.com/locate/envexpbot Rootstock control of scion response to water stress in grapevine Sara Tramontini a,b,∗ , Marco Vitali b , Luna Centioni b , Andrea Schubert b , Claudio Lovisolo b,c a European Food Safety Authority (EFSA), Plant Health Unit (PLH), via Carlo Magno 1/a, 43126, Parma, Italy 1 b University of Turin, Department of Agricultural, Forest and Food Sciences, (DISAFA) via Leonardo da Vinci 44, 10095, Grugliasco, Italy c Plant Virology Institute, National Research Council (IVV-CNR), Grugliasco unit, Via Leonardo da Vinci 44, 10095, Grugliasco, TO, Italy a r t i c l e i n f o Article history: Received 26 February 2013 Received in revised form 18 April 2013 Accepted 22 April 2013 Keywords: Hydraulic conductance Mercuric chloride Stomatal conductance Transmembrane pathway Vitis genotypes Water potential a b s t r a c t Rootstocks play a major role in grapevine tolerance to water stress by controlling and adjusting the water supply to shoot transpiration demand. This study aimed to characterize the influence of rootstock genotypes in the adaptive response of scions to water limiting conditions. The effect of rootstock genotype (140Ru and SO4) was observed in the different availability of water provided to the scions (Cabernet Sauvignon, Grenache, Merlot, Syrah), while scions influenced stomatal control of water transpiration. Implication on the cell-to-cell component of plant water transport in both rootstock and scion impacted on embolisms formation in roots and on hydraulics of leaves. The main conclusion of the present study was that rootstock and scion genotypes are able to confer to the plant traits of drought adaptability influencing respectively the capacity of water extraction from the soil and the sensitivity of the stomatal control. © 2013 Elsevier B.V. All rights reserved. 1. Introduction The dehydration observed under drought conditions appears when an imbalance occurs between water extracted from the substrate and water lost by transpiration (Aroca et al., 2012). There- fore, in plants that are routinely grafted, such as grapevine (Vitis vinifera L.), rootstock effect on scion performance must be consid- ered in the study of adaptability to stress conditions. Rootstocks provide tolerance to exogenous limiting factors, biotic (e.g., soil- borne pests) and abiotic (e.g., salinity, water or oxygen deficit), while influencing the ecophysiological behaviour of the scion and its berry quality (Bavaresco and Lovisolo, 2000; Padgett-Johnson et al., 2000; Soar et al., 2006; Tramontini et al., 2012; de Souza et al., 2009; Ibacache and Sierra, 2009; Rizk-Alla et al., 2011; Marguerit et al., 2012). Stomata are considered direct responsible of opti- mizing the balance between carbon gain and water loss of the plant (Rogiers et al., 2012), and the patterns of their response, in terms of timing and intensity, are genetically determined (Chaves et al., 2010). Significant efforts have been done in the selection of the optimal rootstock/scion combinations to satisfy specific ∗ Corresponding author at: European Food Safety Authority (EFSA), Plant Health Unit (PLH), via Carlo Magno 1/a, 43126, Parma, Italy. Tel.: +39 0521 036 878; fax: +39 0521 036 0878. E-mail address: [email protected] (S. Tramontini). 1 The positions and opinions presented in this article are those of the author alone and are not intended to represent the views or scientific works of EFSA. grape growing needs (e.g., Koundouras et al., 2008; Hamdan and Basheer-Salimia, 2010; Komar et al., 2010). However, the effect of rootstock/scion interaction on plant adaptation to stress is still very much debated (Gambetta et al., 2012). Since drought condi- tions affect water transport from the soil through the plant into the atmosphere in a soil–plant–air continuum that is interconnected by a continuous film of water, measuring its influence on plant water transport requires considering both water extraction at the soil–root interface and water release at the leaf–air interface (Janott et al., 2011). Although the key role played by both rootstock and scion is acknowledged their relative contribution and differences obtained by specific combination of genotypes require some further analysis. The aim of this study was therefore to clarify the relative role of rootstock and scion in the hydraulic response to limitations in water availability. In order to do so, we evaluated the differences observed between two grapevine rootstock genotypes (considered represen- tative of two of the most widespread hybrids: Vitis berlandieri × Vitis rupestris, and V. berlandieri × Vitis riparia) and among four scion genotypes (cultivars), separately and in combination (i.e. on grafted plants). The relationship between stomatal conductance (g s ) and leaf water potential ( leaf ) was used to rank the scion genotypes for their ability to withstand water stress (Rogiers et al., 2012). Root hydraulic conductivity, in terms of short and long distance trans- port (over membranes and xylem, respectively) was considered in order to characterize the responses of the rootstocks. Aquaporins play a major role in transcellular water movement by facilitating the transport of water through cell membranes (Kaldenhoff et al., 0098-8472/$ – see front matter © 2013 Elsevier B.V. All rights reserved. http://dx.doi.org/10.1016/j.envexpbot.2013.04.001
Transcript of Rootstock control of scion response to water stress in grapevine

R
SCa
b
c
ARRA
KHMSTVW
1
wsfvepbwie2empteo
Uf
a
0h
Environmental and Experimental Botany 93 (2013) 20– 26
Contents lists available at SciVerse ScienceDirect
Environmental and Experimental Botany
journa l h om epa ge : www.elsev ier .com/ locate /envexpbot
ootstock control of scion response to water stress in grapevine
ara Tramontinia,b,∗, Marco Vitalib, Luna Centionib, Andrea Schubertb,laudio Lovisolob,c
European Food Safety Authority (EFSA), Plant Health Unit (PLH), via Carlo Magno 1/a, 43126, Parma, Italy1
University of Turin, Department of Agricultural, Forest and Food Sciences, (DISAFA) via Leonardo da Vinci 44, 10095, Grugliasco, ItalyPlant Virology Institute, National Research Council (IVV-CNR), Grugliasco unit, Via Leonardo da Vinci 44, 10095, Grugliasco, TO, Italy
a r t i c l e i n f o
rticle history:eceived 26 February 2013eceived in revised form 18 April 2013ccepted 22 April 2013
a b s t r a c t
Rootstocks play a major role in grapevine tolerance to water stress by controlling and adjusting thewater supply to shoot transpiration demand. This study aimed to characterize the influence of rootstockgenotypes in the adaptive response of scions to water limiting conditions. The effect of rootstock genotype(140Ru and SO4) was observed in the different availability of water provided to the scions (CabernetSauvignon, Grenache, Merlot, Syrah), while scions influenced stomatal control of water transpiration.
eywords:ydraulic conductanceercuric chloride
tomatal conductanceransmembrane pathwayitis genotypes
Implication on the cell-to-cell component of plant water transport in both rootstock and scion impactedon embolisms formation in roots and on hydraulics of leaves. The main conclusion of the present studywas that rootstock and scion genotypes are able to confer to the plant traits of drought adaptabilityinfluencing respectively the capacity of water extraction from the soil and the sensitivity of the stomatalcontrol.
ater potential
. Introduction
The dehydration observed under drought conditions appearshen an imbalance occurs between water extracted from the
ubstrate and water lost by transpiration (Aroca et al., 2012). There-ore, in plants that are routinely grafted, such as grapevine (Vitisinifera L.), rootstock effect on scion performance must be consid-red in the study of adaptability to stress conditions. Rootstocksrovide tolerance to exogenous limiting factors, biotic (e.g., soil-orne pests) and abiotic (e.g., salinity, water or oxygen deficit),hile influencing the ecophysiological behaviour of the scion and
ts berry quality (Bavaresco and Lovisolo, 2000; Padgett-Johnsont al., 2000; Soar et al., 2006; Tramontini et al., 2012; de Souza et al.,009; Ibacache and Sierra, 2009; Rizk-Alla et al., 2011; Margueritt al., 2012). Stomata are considered direct responsible of opti-izing the balance between carbon gain and water loss of the
lant (Rogiers et al., 2012), and the patterns of their response, in
erms of timing and intensity, are genetically determined (Chavest al., 2010). Significant efforts have been done in the selectionf the optimal rootstock/scion combinations to satisfy specific∗ Corresponding author at: European Food Safety Authority (EFSA), Plant Healthnit (PLH), via Carlo Magno 1/a, 43126, Parma, Italy. Tel.: +39 0521 036 878;
ax: +39 0521 036 0878.E-mail address: [email protected] (S. Tramontini).
1 The positions and opinions presented in this article are those of the author alonend are not intended to represent the views or scientific works of EFSA.
098-8472/$ – see front matter © 2013 Elsevier B.V. All rights reserved.ttp://dx.doi.org/10.1016/j.envexpbot.2013.04.001
© 2013 Elsevier B.V. All rights reserved.
grape growing needs (e.g., Koundouras et al., 2008; Hamdan andBasheer-Salimia, 2010; Komar et al., 2010). However, the effectof rootstock/scion interaction on plant adaptation to stress is stillvery much debated (Gambetta et al., 2012). Since drought condi-tions affect water transport from the soil through the plant into theatmosphere in a soil–plant–air continuum that is interconnectedby a continuous film of water, measuring its influence on plantwater transport requires considering both water extraction at thesoil–root interface and water release at the leaf–air interface (Janottet al., 2011). Although the key role played by both rootstock andscion is acknowledged their relative contribution and differencesobtained by specific combination of genotypes require some furtheranalysis.
The aim of this study was therefore to clarify the relative role ofrootstock and scion in the hydraulic response to limitations in wateravailability. In order to do so, we evaluated the differences observedbetween two grapevine rootstock genotypes (considered represen-tative of two of the most widespread hybrids: Vitis berlandieri × Vitisrupestris, and V. berlandieri × Vitis riparia) and among four sciongenotypes (cultivars), separately and in combination (i.e. on graftedplants). The relationship between stomatal conductance (gs) andleaf water potential (� leaf) was used to rank the scion genotypesfor their ability to withstand water stress (Rogiers et al., 2012). Roothydraulic conductivity, in terms of short and long distance trans-
port (over membranes and xylem, respectively) was considered inorder to characterize the responses of the rootstocks. Aquaporinsplay a major role in transcellular water movement by facilitatingthe transport of water through cell membranes (Kaldenhoff et al.,
and E
2n2amnph(tM2stf(etttgce
2
2
stt(strcy(anipS2Rg(wisoSftswoe
2
wm
S. Tramontini et al. / Environmental
008; Maurel et al., 2008; Wudick et al., 2009), therefore they sig-ificantly control root hydraulic conductivity (Javot and Maurel,002; Lee et al., 2005; Van As, 2007; Postaire et al., 2010). Theirctivity was measured with the application of an inhibitor of celletabolism, HgCl2, to root and leaf tissues. Mercurial reagents can-
ot affect all plant aquaporins (Daniels et al., 1994), and couldroduce side effects (Gaspar et al., 2001; Hachez et al., 2006),owever they remain general but nonspecific aquaporin inhibitorsWudick et al., 2009) useful for obtaining preliminary informa-ion on transmembrane water transport (Javot and Maurel, 2002;
oshelion et al., 2002; Terashima and Ono, 2002; Lovisolo et al.,008; Miyazawa et al., 2008; Knipfer et al., 2011). While the waterymplastic path in the root is mainly driven by osmotic forces,he apoplastic path, radial and axial, is maintained by hydrostaticorces, limited by the resistance produced by cavitation eventsAroca et al., 2012). For this reason, a measurement on the level ofmbolisms of the root system was conducted. Our results show thathe rootstock control (based on root hydraulic conductance) andhe scion control (based on stomatal conductance) follow an addi-ive pattern in cooperating to adaptation to drought, confirming therapevine as an optimal model plant for molecular and physiologi-al studies on both plant drought avoidance and tolerance (Lovisolot al., 2010).
. Materials and methods
.1. Plant material
For those experiments requiring the destruction of the rootystem, the investigation was conducted on 2-year-old plants ofwo different grapevine rootstocks, one derived from hybridiza-ion of V. berlandieri with the xerophylic species V. rupestris140Ru), and the other from V. berlandieri with the mesophylicpecies V. riparia (SO4), with three replicates for genotype. Thewo genotypes were selected as characterizing the extremes in theesults of Lovisolo et al. (2008). All the other experiments wereonducted during two consecutive years (2011 and 2012) on 2-ear-old (and 3-year-old the next year) plants of four cultivarsCabernet Sauvignon, Grenache, Merlot, Syrah) grafted on the twobove-mentioned rootstocks, with five replicates for each combi-ation rootstock/cultivar. The four cultivars were selected taking
nto account their ecophysiological characterization obtained byrevious authors: Grenache is near-isohydric (Schultz, 2003;antesteban et al., 2009), Merlot is anisohydric (Williams and Baeza,007; Shellie and Glenn, 2008), Syrah is anisohydric (Schultz, 2003;ogiers et al., 2009; Santesteban et al., 2009), and Cabernet Sauvi-non is classified as isohydric (Chalmers, 2007) or anisohydricWilliams and Baeza, 2007), depending from the authors. All plantsere grown in a greenhouse with no supplementary light or heat-
ng, in 4 L pots filled with a substrate composed of sandy-loamoil/expanded clay/peat mixture (4:2:1 in volume), with a final pHf 7.3. Additional leaves of Cabernet Sauvignon, Grenache, Merlot,yrah needed for the experiment on detached leaves were collectedrom the experimental vineyard located at the Faculty of Agricul-ure of the University of Turin (Grugliasco, Italy), from plants of theame clonal origin. To this aim, mature leaves (10th node onwards)ere collected in the month of July. These leaves were used in
rder to study stomatal behaviour in absence of rootstock and shootffects.
.2. Leaf, root and soil water potentials
Leaf water potential (� leaf) and root water potential (� root)ere estimated with a pressure chamber (Scholander et al., 1965;odel used: Soil Moisture Equipment Corp., Santa Barbara, CA,
xperimental Botany 93 (2013) 20– 26 21
USA). � leaf measurements were done between 10:00 and 12:00AM on fully expanded leaves from primary stems, immediatelyafter detaching the leaf from the stem, unless differently speci-fied (e.g., experiment on detached leaves). � root was measured onthe two most basal leaves early in the morning (around 8:00) afterkeeping them enclosed overnight in a reflective plastic envelope.Leaf bagging blocks transpiration and allows leaf water potential toreach equilibrium with root water potential. Soil water potential(� soil) was calculated from the moisture potential curves previ-ously obtained for that specific substrate (Lovisolo et al., 2008).
2.3. Root hydraulic conductance and putative transmembranepath
We measured hydraulic conductance, its transmembrane watermovement component, and the intensity of xylem embolization.All measurements were based on a destructive method with acontrolled tension–pressure apparatus (Lovisolo et al., 2002) andtaken in sequence on the same plant material as described byLovisolo et al. (2008). The root system was gently cleaned fromsoil under water and cut at the interface between root and shoot.After measurement of weight and volume, it was immersed into atension–pressure chamber filled with tap water and its stalk wasclamped with a rubber sleeve. An initial conductance was measuredby applying a negative pressure of −80 kPa for 5 min through thesleeve: this tension simulates the suction forces that cause watertranspirative transport in the potted roots. The same pressure wasthen imposed for 5 min after having treated the root with a solutionof 0.05 mM HgCl2 for 60 min (+15 min for stabilizing the system)in order to inhibit mercury-sensitive aquaporins. The differencebetween this conductance and the previous conductance definesthe portion of conductance sustained by transmembrane watermovement. The same pressure was again imposed for other 5 minafter a flushing of +100 kPa for 5 min to the whole root, in order tofree the system from embolisms. The difference between this valueand the previous defines the conductance achievable by the systemthrough the apoplastic pathway in absence of embolisms.
2.4. Stomatal conductance
In order to exclude the root effect and to concentrate on thecharacteristics induced by the grafted cultivar, this measurementwas conducted on detached leaves, treated as self-contained func-tional units, as described by Brodribb (2009). Stomatal conductance(gs) was measured on fully developed leaves with a portableGas Exchange Fluorescence System (GFS-3000, Heinz Walz GmbH,Effeltrich, Germany). Measurements were conducted by clampingthe leaves in the leaf chamber, where photosynthetic active radia-tion, (PAR: 1200 �mol m−2 s−1) and temperature (25 ◦C) were keptconstant. A zero-point for CO2 was set at the beginning of each dayof measurements. In the case of the experiment on attached leaves,after measuring gs, the same leaf was used for measuring � leaf. Thecollection of data was concentrated within two weeks (between31 May and 15 June 2012) in order to minimize any potential sea-sonal fluctuation (Alsina et al., 2011). In the case of the experimenton detached leaves, seven leaves per replicate were excised underwater and immediately placed in 50 mL tubes with their petioleimmersed in deionised water. Leaves were kept in the laboratoryunder constant artificial light (400 �mol m−2 s−1 PAR) and temper-ature (24–27 ◦C) and transpiration was continuously monitored onone of them. When the leaf reached a steady state in transpira-tion (usually after 20 min, with a coefficient of variation <3% for
3 min, Nardini and Salleo, 2005), all tubes were simultaneouslyemptied from water. Six measurements of � leaf were then takenwith a 4-min interval on the remaining leaves. Later on, in analy-sis of the correlation between gs and � leaf, it was decided not to
22 S. Tramontini et al. / Environmental and Experimental Botany 93 (2013) 20– 26
F tial (�a 40Rut h tren
c‘iF
2
wbdhobttwwPfwvt
A
cst
2
etCc
3
3
o�S
H2O2 aquaporin inhibition. Pressure flushing induced an increaseof conductance of 197% and 369% respectively (Fig. 2).
Fig. 2. Hydraulic conductance measured on excised root of 140Ru (filled boxes) and
ig. 1. Relationship between soil water potential (� soil , MPa) and leaf water potennd Syrah) potted on the same substrate and grafted on two different rootstocks: 1wo years: 2011 (a) and 2012 (b) between end of May and beginning of August. Eac
onsider the first two measurements of gs due to the observedIwanoff effect’ described by Düring (1993), which produced anncrease in gs immediately after removal of the leaf from water.our replicate measurements were done for each cultivar.
.5. Transmembrane water path in leaves
In order to compare the four cultivars for their transmembraneater movement on foliar tissues, the methodology proposed
y Terashima and Ono (2002) was followed and adapted. Eachetached leaf was divided in two parts along the median vein. Thealf without petiole was used as control and the half with peti-le was left absorbing a solution of 50 mM of HgCl2 for 60 minefore the measurement. During this period, the leaf was exposedo light (1000 �mol m−2 s−1) to help uptake of the solution throughhe petiole by transpiration. From each half-leaf 20 foliar disksith a diameter of 1.2 cm were obtained with a circular cutter andeighted. The disks were floated on 40 mL of deionized water in a
etri dish with the adaxial surface upwards and were shaken gentlyor 15 min. They were then dried on absorbing paper for 5 min andeighted again. Twelve replicate leaves were used for each culti-
ar. The increase in turgidity of the leaf discs was calculated withhe following equation:
verage weight increase =[
(final weight − initial weight)initial weight
]× 100
The obtained value was treated as a measure of the hydrauliconductivity of the plasma membrane of the leaf disc, such as a rea-onable approximation of the transmembrane component of waterransport, independent from water movement via vasculature.
.6. Statistical analysis
Data were expressed by means and corresponding standardrrors. Results were submitted to one-way ANOVA using the statis-ical software package SPSS (version 20, SPSS Inc, Cary; NC, USA).orrelated variables were interpolated in figures by regressionurves plotted by means of Microsoft Excel© software.
. Results
.1. Rootstock – soil and leaf water potentials
In order to characterize rootstock components of the controlf stress effects, we analysed the relationship between � soil andleaf, during two consecutive years, in plants grafted on 140Ru or
O4. At the same values of � leaf, plants grafted on 140Ru showed
leaf, MPa) on vine plants of four cultivars (Cabernet Sauvignon, Grenache, Merlot (filled squares) and SO4 (empty circles). The figure refers to data collected duringdline label is bordered with the same style of the line to which it refers.
more negative � soil than plants grafted on SO4. This difference wasincreasingly evident with lower � leaf and was not influenced by thescion cultivar (Fig. 1). Plant development, measured as leaf area,was not different between the two rootstocks (data not shown).
3.2. Rootstock – root hydraulic conductance and transmembranewater path
The diverse relationship between � soil and � leaf, observed in thetwo rootstocks suggested differences in the hydraulic conductanceof the root system. We analysed this parameter in excised roots ofnot grafted plants of 140Ru and SO4, at the same time evaluatingthe component of conductance controlled by xylem embolization.Root hydraulic conductance, measured at −80 kPa on the excisedroot before and after Hg treatment, was always higher in 140Ruthan in SO4. However, Hg treatment reduced the conductance by56% in 140Ru and 16% in SO4, a result not far from the rangerecently obtained on rootstocks by Gambetta et al. (2012) after
SO4 (empty boxes) during three sequential treatments: (i) depressurizing at the api-cal cut with a vacuum application of −80 kPa, (ii) treating with Hg and depressurizingat −80 kPa, and (iii) imposing a pressure flushing of +100 kPa on the Hg-treated roots.Means ± standard errors. Differences were tested within each treatment separately.Values with different letters differ significantly (P ≤ 0.05).

S. Tramontini et al. / Environmental and Experimental Botany 93 (2013) 20– 26 23
F af waM and S2 fers.
3
ldatgslco‘vbi
F(i
ig. 3. Relationship between stomatal conductance (gs, mmol H2O m−2 s−1) and leerlot (c) and Syrah (d) grafted on two different rootstocks: 140Ru (filled squares)
012. Each trendline label is bordered with the same style of the line to which it re
.3. Scion – stomatal conductance on attached leaves
While assessing the pattern of gs/� leaf in detached scion cultivareaves, we established in grafted plants whether the genotype-ependent patterns of gs adaptation to leaf water status would beffected by the rootstock. A situation consistent to all the four cul-ivars can be observed: at the same level of � leaf, in fact, plantsrafted on 140Ru tend to have higher gs values than plants of theame cultivar grafted on SO4 (Fig. 3). This was observed at � leafevels higher than −1 MPa; only Grenache (Fig. 3b) expressed thisondition for � leaf values more negative than −1.1 MPa. The resultf a two way ANOVA conducted on gs values for the two factors
cultivar’ and ‘rootstock’ confirmed that the difference in the meanalues among the different cultivars was greater than would haveeen expected by chance after allowing for effects of differences
n rootstocks (P = 0.031) and that the difference in the mean values
ig. 4. Relationship between stomatal conductance (gs, mmol H2O m−2 s−1) and leaf watempty diamonds), Merlot (filled squares) and Syrah (empty circles) grafted on two diffen Fig. 3. Each trendline label is bordered with the same style of the line to which it refers
ter potential (� leaf, MPa) on vine plants of Cabernet Sauvignon (a), Grenache (b),O4 (empty circles). The figures refer to data collected between 31 May and 15 June
among the different rootstocks was greater than would have beenexpected by chance after allowing for effects of differences in cul-tivars (P = 0.037), but without a statistically significant interactionbetween ‘cultivar’ and ‘rootstock’ (P = 0.559). When the data wereclustered per rootstock (Fig. 4), it was more evident that, on bothrootstocks, gs of Merlot was the less influenced by � leaf while theopposite was for Syrah, while Cabernet Sauvignon and Grenacheconfirmed the intermediate condition already observed at Fig. 3,the latter presenting strong variations of gs when grafted on 140Ruand minimal variations when on SO4 (Fig. 4).
3.4. Scion – stomatal conductance on detached leaves
The next step was to focus on the scion cultivar control of gs,which was measured on progressively drying leaves (Fig. 5). Forthe cultivars Cabernet Sauvignon and Grenache, the dynamics of gs
er potential (� leaf, MPa) on vine plants of Cabernet Sauvignon (crosses), Grenacherent rootstocks: 140Ru (a) and SO4 (b). The figures represent the same data shown.
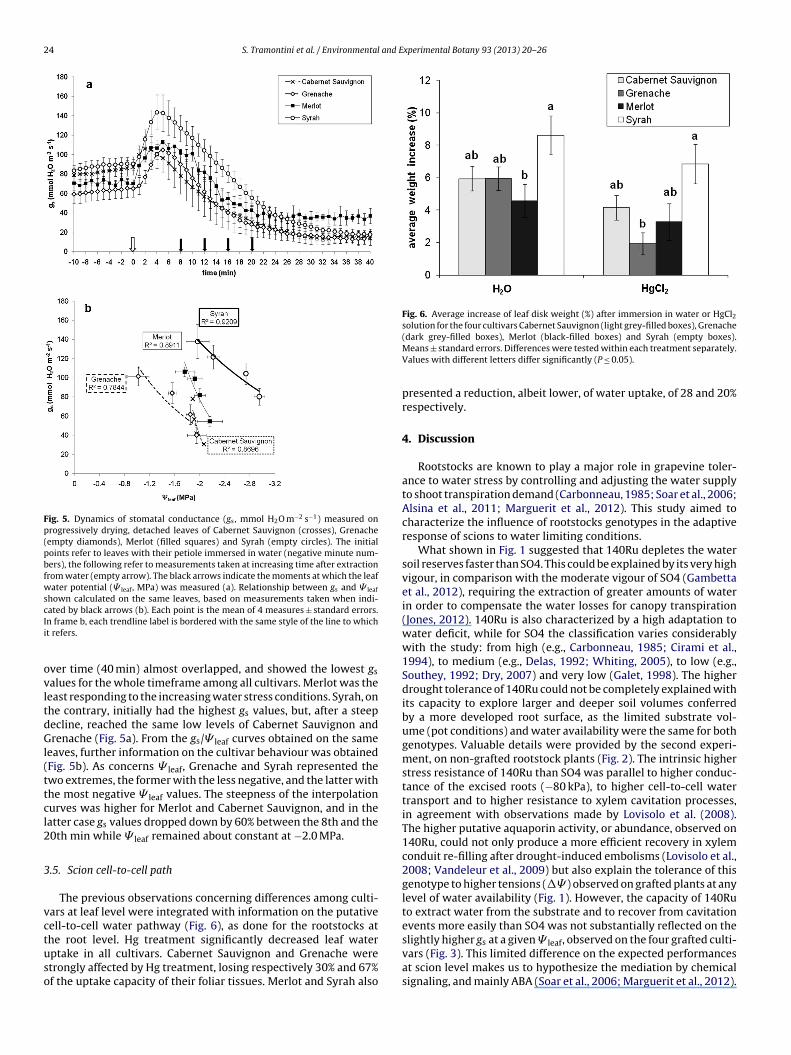
24 S. Tramontini et al. / Environmental and Experimental Botany 93 (2013) 20– 26
Fig. 5. Dynamics of stomatal conductance (gs, mmol H2O m−2 s−1) measured onprogressively drying, detached leaves of Cabernet Sauvignon (crosses), Grenache(empty diamonds), Merlot (filled squares) and Syrah (empty circles). The initialpoints refer to leaves with their petiole immersed in water (negative minute num-bers), the following refer to measurements taken at increasing time after extractionfrom water (empty arrow). The black arrows indicate the moments at which the leafwater potential (� leaf, MPa) was measured (a). Relationship between gs and � leaf
shown calculated on the same leaves, based on measurements taken when indi-cIi
ovltdGl(ttcl2
3
vctuso
Fig. 6. Average increase of leaf disk weight (%) after immersion in water or HgCl2solution for the four cultivars Cabernet Sauvignon (light grey-filled boxes), Grenache
slightly higher g at a given � , observed on the four grafted culti-
ated by black arrows (b). Each point is the mean of 4 measures ± standard errors.n frame b, each trendline label is bordered with the same style of the line to whicht refers.
ver time (40 min) almost overlapped, and showed the lowest gs
alues for the whole timeframe among all cultivars. Merlot was theeast responding to the increasing water stress conditions. Syrah, onhe contrary, initially had the highest gs values, but, after a steepecline, reached the same low levels of Cabernet Sauvignon andrenache (Fig. 5a). From the gs/� leaf curves obtained on the same
eaves, further information on the cultivar behaviour was obtainedFig. 5b). As concerns � leaf, Grenache and Syrah represented thewo extremes, the former with the less negative, and the latter withhe most negative � leaf values. The steepness of the interpolationurves was higher for Merlot and Cabernet Sauvignon, and in theatter case gs values dropped down by 60% between the 8th and the0th min while � leaf remained about constant at −2.0 MPa.
.5. Scion cell-to-cell path
The previous observations concerning differences among culti-ars at leaf level were integrated with information on the putativeell-to-cell water pathway (Fig. 6), as done for the rootstocks athe root level. Hg treatment significantly decreased leaf water
ptake in all cultivars. Cabernet Sauvignon and Grenache weretrongly affected by Hg treatment, losing respectively 30% and 67%f the uptake capacity of their foliar tissues. Merlot and Syrah also(dark grey-filled boxes), Merlot (black-filled boxes) and Syrah (empty boxes).Means ± standard errors. Differences were tested within each treatment separately.Values with different letters differ significantly (P ≤ 0.05).
presented a reduction, albeit lower, of water uptake, of 28 and 20%respectively.
4. Discussion
Rootstocks are known to play a major role in grapevine toler-ance to water stress by controlling and adjusting the water supplyto shoot transpiration demand (Carbonneau, 1985; Soar et al., 2006;Alsina et al., 2011; Marguerit et al., 2012). This study aimed tocharacterize the influence of rootstocks genotypes in the adaptiveresponse of scions to water limiting conditions.
What shown in Fig. 1 suggested that 140Ru depletes the watersoil reserves faster than SO4. This could be explained by its very highvigour, in comparison with the moderate vigour of SO4 (Gambettaet al., 2012), requiring the extraction of greater amounts of waterin order to compensate the water losses for canopy transpiration(Jones, 2012). 140Ru is also characterized by a high adaptation towater deficit, while for SO4 the classification varies considerablywith the study: from high (e.g., Carbonneau, 1985; Cirami et al.,1994), to medium (e.g., Delas, 1992; Whiting, 2005), to low (e.g.,Southey, 1992; Dry, 2007) and very low (Galet, 1998). The higherdrought tolerance of 140Ru could not be completely explained withits capacity to explore larger and deeper soil volumes conferredby a more developed root surface, as the limited substrate vol-ume (pot conditions) and water availability were the same for bothgenotypes. Valuable details were provided by the second experi-ment, on non-grafted rootstock plants (Fig. 2). The intrinsic higherstress resistance of 140Ru than SO4 was parallel to higher conduc-tance of the excised roots (−80 kPa), to higher cell-to-cell watertransport and to higher resistance to xylem cavitation processes,in agreement with observations made by Lovisolo et al. (2008).The higher putative aquaporin activity, or abundance, observed on140Ru, could not only produce a more efficient recovery in xylemconduit re-filling after drought-induced embolisms (Lovisolo et al.,2008; Vandeleur et al., 2009) but also explain the tolerance of thisgenotype to higher tensions (�� ) observed on grafted plants at anylevel of water availability (Fig. 1). However, the capacity of 140Ruto extract water from the substrate and to recover from cavitationevents more easily than SO4 was not substantially reflected on the
s leafvars (Fig. 3). This limited difference on the expected performancesat scion level makes us to hypothesize the mediation by chemicalsignaling, and mainly ABA (Soar et al., 2006; Marguerit et al., 2012).

and E
ctptcoegwusmaltctsbswlfCaaomufcsPgeHtmw
aoFrdrotttep
5
sas(rrtr
S. Tramontini et al. / Environmental
The component which conferred specific characteristics to theorrelation between stomatal conductance and leaf water poten-ial was the scion cultivar: the gs/� leaf curves measured in graftedlants (Fig. 4) have different slopes and shapes, correspondingo the ecophysiological behavior of each genotype. Slopes wereultivar-specific and not dependent on the rootstock. The curvef Cabernet Sauvignon, in spite of the most evident rootstockffect among the four cultivars (Fig. 3), maintained intermediates values on both rootstocks (Fig. 4). This intermediate rankingas consistent with the intermediate percentage loss in waterptake observed on foliar disks after Hg treatment, suggesting thattomatal conductance is linked with the transmembrane waterovement capacity of the leaf, probably dependent on aquaporin
ctivity (Pou et al., 2013). However, the gs/� leaf curve on excisedeaves, where stomatal closure delayed the � leaf drop below thehreshold of −2 MPa (Fig. 5b), could not be explained within theontext of the above-mentioned hydraulic forces and could implyhe involvement of complex hormonal signalling in whole plants,o that Cabernet Sauvignon assumes isohydric or anisohydricehaviour, depending on the experimental conditions. Grenache, inpite of its gs/� leaf curves resemblance to Cabernet Sauvignon onhole plants, presented the tightest stomatal control on excised
eaves (Fig. 5) and the highest effect of Hg-treatment among theour cultivars. This result is consistent with what was observed inabernet Sauvignon, with a more characterized isohydricity, prob-bly supported by the predominance of a hydraulic control. Syrah,s expected, displayed a behavior opposite to Grenache, but onlyn excised leaves, with the highest values of gs, and the most dra-atic � leaf drop (Fig. 5), and the lowest percentage loss in water
ptake on Hg-treated leaves. However, on whole plants, there-ore at less negative � leaf, the same cultivar expressed a behaviourloser to Grenache (near-isohydric) than to Merlot (anisohydric),upporting the results obtained under moderate water stress byou et al. (2012). Finally, Merlot showed the lowest variations ofs both in whole plants, and in excised leaves, confirming the gen-ral identifications as anisohydric cultivar. The scarce effect of theg treatment, similar as observed in Syrah, suggests that the low
ransmembrane water transport capacity negatively affected sto-atal control, limiting transpiration in the former and favoring soilater depletion in the latter.
The observations done on whole plants and on excised leavesllowed to obtain a more complete picture of the cultivar effectn the hydraulic dynamics involved at plant and at leaf level.urther studies will be needed in order to integrate the cur-ent results with additional details on the component of variationue to hydraulic and hormonal signaling between the scion andootstock (Marguerit et al., 2012). From the combination of thebtained results, we could interpret the rootstock effect as ‘quan-itative’: the ability of roots to supply water relative to shootranspiration demand displaced reciprocally the rootstock curveso higher/lower parallel positions. On the other hand, the scionffect was ‘qualitative’: the feedback loop between gs and � leafroduced at leaf level modified the slope of the curves.
. Conclusions
From the present study, it was concluded that rootstock andcion genotypes are able to confer to the plant traits of droughtdaptability influencing the capacity of water extraction from theoil and sensitivity of the stomatal control. On both components
roots and canopy) the transcellular pathway seems to have strongelationships with strategies related to water translocation. Furtheresearch will be needed in order to identify the potential use ofhis information in studies of stress tolerance and adaptability ofootstocks and cultivars to specific ecological conditions.xperimental Botany 93 (2013) 20– 26 25
Acknowledgements
We wish to thank Tiziano Strano and Elzbieta Ceglarska for theinvaluable support and Vivai Rauscedo for providing the plantingmaterial needed for this research. We acknowledge financial sup-port by the AGER Foundation, project SERRES, grant no. 2010-2015.
References
Alsina, M.M., Smart, D.R., Bauerle, T., de Herralde, F., Biel, C., Stockert, C., Negron,C., Save, R., 2011. Seasonal changes of whole root system conductance by adrought-tolerant grape root system. Journal of Experimental Botany 62, 99–109.
Aroca, R., Porcel, R., Ruiz-Lozano, J.M., 2012. Regulation of root water uptake underabiotic stress conditions. Journal of Experimental Botany 63, 43–57.
Bavaresco, L., Lovisolo, C., 2000. Effect of grafting on grapevine chlorosis andhydraulic conductivity. Vitis 39, 89–92.
Brodribb, T.J., 2009. Xylem hydraulic physiology: the functional backbone of terres-trial plant productivity. Plant Science 177, 245–251.
Carbonneau, A., 1985. The early selection of grapevine rootstocks for resistance todrought conditions. American Journal of Enology and Viticulture 36, 195–198.
Chalmers, Y.M., 2007. Influence of sustained deficit irrigation on physiologyand phenolic compounds in winegrapes and wine. Adelaide University, PhDthesis. http://digital.library.adelaide.edu.au/dspace/bitstream/2440/50101/1/02whole.pdf.
Chaves, M.M., Zarrouk, O., Francisco, R., Costa, J.M., Santos, T., Regalado, A.P.,Rodrigues, M.L., Lopes, C.M., 2010. Grapevine under deficit irrigation: hints fromphysiological and molecular data. Annals of Botany 105, 661–676.
Cirami, R., Furkaliev, J., Radford, R., 1994. Summer drought and vine rootstocks.Australian Grapegrower and Winemaker – Annual Technical Issue 336, 145.
Daniels, M.J., Mirkov, T.E., Chrispeels, M.J., 1994. The plasma membrane of Arabidop-sis thaliana contains a mercury-insensitive aquaporin that is a homolog of thetonoplast water channel protein TIP. Plant Physiology 106, 1325–1333.
Delas, J.J., 1992. Criteria used for rootstock selection in France. In: Wolpert, J.,Walker, M.A., Weber, E., American Society for Enology Viticulture: Davis (Eds.),Proceedings of Rootstock Seminar: A Worldwide Perspective. Reno, NV, USA, pp.83–89.
de Souza, C.R., Bassoi, L.H., Lima Filho, J.M.P., Silva, F.F.S.da, Viana, L.H., Dantas, B.F.,Pereira, M.S., Ribeiro, P.R., de, A., 2009. Water relations of field-grown grapevinesin the São Francisco Valley. Brazil, under different rootstocks and irrigationstrategies. Scientia Agricola 66, 436–446.
Dry, N., 2007. Grapevine Rootstocks: Selection and Management for South Aus-tralian Vineyards. Lythrum Press (Adelaide), Stepney, Australia, pp. 85.
Düring, H., 1993. Rapid stomatal and photosynthetic responses of Vitis berlandierileaves after petiole excision in water. Vitis 32, 63–68.
Galet, P., 1998. Grape Varieties and Rootstock Varieties. Oenoplurimedia, Chaintré,France, pp. 315.
Gambetta, G.A., Manuck, C.M., Drucker, S.T., Shaghasi, T., Fort, K., Matthews, M.A.,Walker, M.A., McElrone, A.J., 2012. The relationship between root hydraulics andscion vigour across Vitis rootstocks: what role do root aquaporins play? Journalof Experimental Botany 63, 6445–6455.
Gaspar, M., Sissoef, I., Bousser, A., Roche, O., Mahe, A., Hoarau, J., 2001. Transientvariations of water transfer induced by HgCl2 in excised roots of young maizeplants: new data on the inhibition process. Australian Journal of Plant Physiology28, 1175–1186.
Hachez, C., Zelazny, E., Chaumont, F., 2006. Modulating the expression of aquaporingenes in planta: a key to understand their physiological functions? Biochimicaet Biophysica Acta (BBA) – Biomembranes 1758, 1142–1156.
Hamdan, A., Basheer-Salimia, R., 2010. Preliminary compatibility between sometable-grapevine scions and phylloxera-resistant rootstock cultivars. Jordan Jour-nal of Agricultural Sciences 6, 1–10.
Ibacache, G.A., Sierra, C.B., 2009. Influence of rootstocks on nitrogen, phosphorusand potassium content in petioles of four table grape varieties. Chilean Journalof Agricultural Research 69, 503–508.
Janott, M., Gayler, S., Gessler, A., Javaux, M., Klier, C., Priesacket, E., 2011. Aone-dimensional model of water flow in soil–plant systems based on plantarchitecture. Plant and Soil 341, 233–256.
Javot, H., Maurel, C., 2002. The role of aquaporins in root water uptake. Annals ofBotany (Lond) 90, 301–313.
Jones, H.G., 2012. How do rootstocks control shoot water relations? New Phytologist194, 301–303.
Kaldenhoff, R., Ribas Carbo, M., Flexas Sans, J., Lovisolo, C., Heckwolf, M., Uehlein,N., 2008. Aquaporins and Plant Water Balance. Plant. Cell and Environment 31,658–666.
Knipfer, T., Besse, M., Verdeil, J.L., Fricke, W., 2011. Aquaporin-facilitated wateruptake in barley (Hordeum vulgare L.) roots. Journal of Experimental Botany 62,4115–4126.
Komar, V., Vigne, E., Demangeat, G., Lemaire, O., Fuchs, M., 2010. Comparative per-formance of virus-infected Vitis vinifera cv. Savagnin rose grafted onto three
rootstocks. American Society for Enology and Viticulture 61, 68–73.Koundouras, S., Tsialtas, I.T., Zioziou, E., Nikolaou, N., 2008. Rootstock effects on theadaptive strategies of grapevine (Vitis vinifera L. cv. Cabernet-Sauvignon) undercontrasting water status: leaf physiological and structural responses. Agricul-ture, Ecosystems & Environment 128, 86–96.

2 and E
L
L
L
L
M
M
M
M
N
P
P
P
P
R
6 S. Tramontini et al. / Environmental
ee, S.H., Chung, G.C., Steudle, E., 2005. Low temperature and mechanical stressesdifferently gate aquaporins of root cortical cells of chilling-sensitive cucumberand -resistant figleaf gourd. Plant, Cell & Environment 28, 1191–1202.
ovisolo, C., Hartung, W., Schubert, A., 2002. Whole-plant hydraulic conductanceand root-to-shoot flow of abscisic acid are independently affected by waterstress in grapevines. Functional Plant Biology 29, 1349–1356.
ovisolo, C., Perrone, I., Carra, A., Ferrandino, A., Flexas, J., Medrano, H., Schubert, A.,2010. Drought-induced changes in development and function of grapevine (Vitisspp.) organs and in their hydraulic and non-hydraulic interactions at the whole-plant level: a physiological and molecular update. Functional Plant Biology 37,98–116.
ovisolo, C., Tramontini, S., Flexas, J., Schubert, A., 2008. Mercurial inhibition of roothydraulic conductance in Vitis spp. rootstocks under water stress. Environmen-tal and Experimental Botany 63, 178–182.
arguerit, E., Brendel, O., Lebon, E., Van Leeuwen, C., Ollat, N., 2012. Rootstockcontrol of scion transpiration and its acclimation to water deficit are controlledby different genes. New Phytologist 194, 416–429.
aurel, C., Verdoucq, L., Luu, D.-T., Santoni, V., 2008. Plant aquaporins: membranechannels with multiple integrated functions. Annual Review of Plant Biology 59,595–624.
oshelion, M., Becker, D., Biela, A., Uehlein, N., Hedrich, R., Otto, B., Levi, H., Moran,N., Kaldenhoff, R., 2002. Plasma membrane aquaporins in the motor cells ofSamanea saman: diurnal and circadian regulation. Plant Cell 3, 727–739.
iyazawa, S.-I., Yoshimura, S., Shinazaki, Y., Maeshima, M., Miyake, C., 2008.Deactivation of aquaporins decreases internal conductance to CO2 diffusionin tobacco leaves grown under longterm drought. Functional Plant Biology 35,553–564.
ardini, A., Salleo, S., 2005. Water stress-induced modifications of leaf hydraulicarchitecture in sunflower: co-ordination with gas exchange. Journal of Experi-mental Botany 56, 3093–3101.
adgett-Johnson, M., Williams, L.E., Walker, M.A., 2000. The influence of Vitis ripariarootstock on water relations and gas exchange of Vitis vinifera cv. Carignane scionunder non-irrigated conditions. American Journal of Enology and Viticulture 51,137–143.
ostaire, O., Tournaire-Roux, C., Grondin, A., Boursiac, Y., Morillon, R., Shäffner, A.R.,Maurel, C., 2010. A PIP1 aquaporin contributes to hydrostatic pressure-inducedwater transport in both the root and rosette of Arabidopsis. Plant Physiology 152,1418–1430.
ou, A., Medrano, H., Tomás, M., Martorell, S., Ribas-Carbó, M., Flexas, J., 2012. Aniso-hydric behaviour in grapevines results in better performance under moderatewater stress and recovery than isohydric behaviour. Plant and Soil 359, 335–349.
ou, A., Medrano, H., Flexas, J., Tyerman, S., 2013. A putative role for TIP and PIP aqua-
porins in dynamics of leaf hydraulic and stomatal conductances in grapevineunder water stress and re-watering. Plant, Cell & Environment 36, 828–843.izk-Alla, M.S., Sabry, G.H., Adb El-Wahab, M.A., 2011. Influence of some rootstockson the performance of red globe grape cultivar. The Journal of American Science7, 71–81.
xperimental Botany 93 (2013) 20– 26
Rogiers, S.Y., Greer, D.H., Hatfield, J.M., Hutton, R.J., Clarke, S.J., Hutchinson, P.A.,Somers, A., 2012. Stomatal response of an anisohydric grapevine cultivar toevaporative demand, available soil moisture and abscisic acid. Tree Physiology32, 249–261.
Rogiers, S.Y., Greer, D.H., Hutton, R.J., Landsberg, J.J., 2009. Does nigh-time transpi-ration contribute to anisohydric behaviour in a Vitis vinifera cultivar? Journal ofExperimental Botany 60, 3751–3763.
Santesteban, L.G., Miranda, C., Royo, J.B., 2009. Effect of water deficit and rewateringon leaf gas exchange and transpiration decline of excised leaves of four grapevine(Vitis vinifera L.) cultivars. Scientia Horticulturae 121, 434–439.
Scholander, P., Hammel, H., Edda, D., Bradstreet, E., Hemmingsen, E., 1965. Sappressure in vascular plants. Science 148, 339–346.
Schultz, H.R., 2003. Differences in hydraulic architecture account for near-isohydricand anisohydric behaviour of two field-grown Vitis vinifera L. cultivars duringdrought. Plant, Cell and Environment 26, 1393–1405.
Shellie, K., Glenn, D.M., 2008. Wine grape response to kaolin particle film underdeficit and well-watered conditions. Acta Horticulturae 792, 587–591.
Soar, C.J., Dry, P.R., Loveys, B.R., 2006. Scion photosynthesis and leaf gas exchangein Vitis vinifera L. cv. Shiraz: mediation of rootstock effects via xylem sap ABA.Australian Journal of Grape and Wine Research 12, 82–96.
Southey, J.M., 1992. Grapevine rootstock performance under diverse conditions inSouth Africa. In: Wolpert, J., Walker, M.A., Weber, E., American Society for Enol-ogy Viticulture: Davis (Eds.), Proceedings of Rootstock Seminar: A WorldwidePerspective. Reno, NV, USA, pp. 27–51.
Terashima, I., Ono, K., 2002. Effects of HgCl2 on CO2 dependence of leaf photosyn-thesis: evidence indicating involvement of aquaporins in CO2 diffusion acrossthe plasma membrane. Plant and Cell Physiology 43, 70–78.
Tramontini, S., Van Leeuwen, C., Domec, J.C., Destrac-Irvine, A., Basteau, C., Vitali, M.,Mosbach-Schulz, O., Lovisolo, C., 2012. Impact of soil texture and water avail-ability on the hydraulic control of plant and grape-berry development. Plant andSoil, http://dx.doi.org/10.1007/s11104-012-1507-x.
Van As, H., 2007. Intact plant MRI for the study of cell water relations, membrane per-meability, cell-to-cell and long distance water transport. Journal of ExperimentalBotany 58, 743–756.
Vandeleur, R.K., Mayo, G., Shelden, M.C., Gilliham, M., Kaiser, B.N., Tyerman, S.D.,2009. The role of plasma membrane intrinsic protein aquaporins in watertransport through roots: diurnal and drought stress responses reveal differ-ent strategies between isohydric and anisohydric cultivars of grapevine. PlantPhysiology 149, 445.
Whiting, J.R., 2005. Grapevine rootstocks. In: Dry, P.R., Coombe, B.G. (Eds.), Viticul-ture: vol. 1 – Resources. Winetitles Pty Ltd., Ashford, Australia, pp. 167–188.
Williams, L.E., Baeza, P., 2007. Relationships among ambient temperature and vapor
pressure deficit and leaf and stem water potentials of fully irrigated, field-growngrapevines. American Journal of Enology and Viticulture 58, 173–181.Wudick, M.M., Luu, D.-T., Maurel, C., 2009. A look inside: localization pat-terns and functions of intracellular plant aquaporins. New Phytologist 184,289–302.




















