
Grapevine fleck virus-like viruses in Vitis
13
Arch Virol (2000) 145: 553–565 Grapevine fleck virus-like viruses in Vitis S. Sabanadzovic 1 , N. Abou-Ghanem 1 , M. A. Castellano 2 , M. Digiaro 1 , and G. P. Martelli 2 1 Istituto Agronomico Mediterraneo, Valenzano, Bari, Italy 2 Dipartimento di Protezione delle Piante, Università degli Studi and Centro di Studio del CNR sui Virus e le Virosi delle Colture Mediterranee, Bari, Italy Accepted September 27, 1999 Summary. Two sets of degenerate primers for the specific amplification of 572– 575 nt and 386 nt segments of the methyltransferase and RNA- dependent RNA polymerase cistrons of members of the genera Tymovirus and Marafivirus and of the unassigned virus Grapevine fleck virus (GFkV) were designed on the basis of available sequences. These primers were used for amplifying and subsequent cloning and sequencing part of the open reading frame 1 of the genome of GFkV, Grapevine asteroid mosaic-associated virus (GAMaV) and of another previously unreported virus, for which the name Grapevine red globe virus (GRGV) is pro- posed. Computer-assisted analysis of the amplified genome portions showed that the three grapevine viruses are phylogenetically related with one another and with sequenced tymoviruses and marafiviruses. The relationships with tymoviruses was confirmed by the type of ultrastructural modifications induced in the host cells. RdRp-specific degenerate primers were successfully used for the aspecific detection of the three viruses in crude grapevine sap extracts. Specific virus iden- tification was obtained with RT-PCR using antisense virus-specific primers. Introduction Grapevine fleck virus (GFkV) is a phloem-limited non mechanically transmis- sible virus with isometric particles c. 30 nm in diameter, rounded contour, and surface structure like that of tymoviruses [12] and marafiviruses [13], suggestive of clustering of coat protein subunits in pentamers and hexamers [1, 4]. The vi- ral genome is a positive sense single-stranded RNA c. 7,500 nucleotides in size, which was almost totally sequenced ([30], unpubl. res). GFkV is latent in Vitis vinifera but induces specific foliar symptoms in the indicator Vitis rupestris [18], in whose phloem cells it elicits highly characteristics cythopatic structures [5].
Transcript of Grapevine fleck virus-like viruses in Vitis

Arch Virol (2000) 145: 553–565
Grapevine fleck virus-like viruses inVitis
S. Sabanadzovic1, N. Abou-Ghanem1, M. A. Castellano2,M. Digiaro 1, andG. P. Martelli 2
1Istituto Agronomico Mediterraneo, Valenzano, Bari, Italy2Dipartimento di Protezione delle Piante, Università degli Studiand Centro di Studio del CNR sui Virus e le Virosi delle Colture
Mediterranee, Bari, Italy
Accepted September 27, 1999
Summary. Two sets of degenerate primers for the specific amplification of 572–575 nt and 386 nt segments of the methyltransferase and RNA- dependent RNApolymerase cistrons of members of the generaTymovirusandMarafivirusand ofthe unassigned virus Grapevine fleck virus (GFkV) were designed on the basisof available sequences. These primers were used for amplifying and subsequentcloning and sequencing part of the open reading frame 1 of the genome of GFkV,Grapevine asteroid mosaic-associated virus (GAMaV) and of another previouslyunreported virus, for which the name Grapevine red globe virus (GRGV) is pro-posed. Computer-assisted analysis of the amplified genome portions showed thatthe three grapevine viruses are phylogenetically related with one another and withsequenced tymoviruses and marafiviruses. The relationships with tymoviruseswas confirmed by the type of ultrastructural modifications induced in the hostcells. RdRp-specific degenerate primers were successfully used for the aspecificdetection of the three viruses in crude grapevine sap extracts. Specific virus iden-tification was obtained with RT-PCR using antisense virus-specific primers.
Introduction
Grapevine fleck virus (GFkV) is a phloem-limited non mechanically transmis-sible virus with isometric particles c. 30 nm in diameter, rounded contour, andsurface structure like that of tymoviruses [12] and marafiviruses [13], suggestiveof clustering of coat protein subunits in pentamers and hexamers [1, 4]. The vi-ral genome is a positive sense single-stranded RNA c. 7,500 nucleotides in size,which was almost totally sequenced ([30], unpubl. res). GFkV is latent inVitisviniferabut induces specific foliar symptoms in the indicatorVitis rupestris[18],in whose phloem cells it elicits highly characteristics cythopatic structures [5].

554 S. Sabanadzovic et al.
Asteroid mosaic, a semilatent disease of European grapes, is also indexed byV. rupestris, in which it elicits foliar symptoms different from those induced byfleck [17]. Vines affected by asteroid mosaic contain a largely uncharacterized iso-metric virus morphologically similar but serologically distinct from GFkV, whichwas provisionally called Grapevine asteroid mosaic-associated virus (GAMaV)[2].
In the course of a recent study of a severe case of leafroll disease inV. viniferacv. Red globe [8], partially purified preparations from leaves of naturally infectedvines revealed the presence of isometric particles with the same size and outwardaspect as GFkV and GAMaV virions (Fig. 7 D). Comparable particles were alsoseen in partially purified preparations from Albanian accessions of unidentifiedcultivars ofV. viniferainfected by grapevine leafroll-associated virus 7 (GLRaV-7) [7]. As reported in this paper, a comparative study was conducted to determinethe nature of the isometric particles present in the grapevine accessions of differentgeographical origin and their relationship with GFkV.
Materials and methods
Virus sources
The virus sources used in this investigation were LN33 MT48 infected by GFkV [30], aV.rupestrisplant (USA9) infected with an authentic Californian isolate of GAMaV [2] andV.viniferaaccessions from southern Italy (RG40/5) and Albania (AA41, AA42) [7, 8]. Maturecanes from field-grown vines were used for double stranded RNA (dsRNA) extraction andmolecular testing, whereas cuttings rooted in sand and forced to grow under glasshouseconditions served as donors of tissue for thin sectioning.
Double-stranded RNA extraction
Double-stranded RNAs (dsRNAs) were extracted from cortical scrapings of mature grapevinecanes by phenol extraction and purification through cellulose CF-11 columns as described byDodds [11]. Preparations were digested with RNase-free DNase (60mg/ml) and DNase-freepancreatic RNase (0.5mg/ml) [31] prior to denaturation and reverse transcrption.
Reverse transcription
dsRNA preparations were denatured with 20 mM methyl mercuric hydroxide, primed with1mg of random DNA hexanucleotides (Roche Biochemicals) and reverse transcribed with200 units of Moloney murine leukemia virus reverse transcriptase (Gibco-BRL) in a 30mlreaction for 1 h at 39◦C.
Primers
Two sets of degenerate oligonucleotide primers were designed for RT-PCR, based on pub-lished nucleotide sequences ofTurnip yellow mosaic virus(TYMV) [19], Eggplant mosaicvirus (EMV) [26], Kennedia yellow mosaic virus(KYMV) [10], Scrophularia mottle virus(ScrMV, = Ononis yellow mosaic virus) [9] andPhysalis mottle virus(PhyMV) [27], Oatblue dwarf virus(OBDV) [13], and GFkV ([29] and unpubl.)
The nucleotide sequence, amino acid sequence, degeneracy level of these primers andthe target genomic regions are reported in Table 1 and Fig. 1, respectively. In particular, thefirst set of primers (MTR1 and MTR2) was selected for a portion of the polymerase cistron,including Rozanov et al. [28] motifs III and IV of the methyltransferase domain and extending

Grapevine fleck virus-like viruses 555
Table 1. PCR primers for the amplification of the methyltransferase and polymerase codingsequences of grapevine fleck virus-like viruses
Primer Nucleotide sequencea Degeneracy Amplified product (bp)
MTR1 5′TTC ATG CAY GAY GCY MTS ATG T3′ 48 575–689MTR2 5′TCC CAV GCN BHB GVR GTG ACC CA3′ 1944RD1 5′CYC ARC AYA ARG TVA ACG A3′ 48 386RD2 5′GCG CAT GCA BGT SAG RGG G3′ 12
aB = T + C+ G; H= A + T + C; M = A + C; N= A + C+ G+ T; R= A + G; S= C+ G;V = A + G+ C; Y = T + C
Fig. 1. Diagramatic representation of the replicase cistron comprised in ORF1 of Grapevinefleck virus and in members of the generaTymovirusandMarafivirus. The target sequencesfor PCR amplification (MTR-PCR and RD-PCR) are indicated.MTRMethyltransferase;Hel
helicase;RdRpRNA-dependent RNA polymerase
towards the helicase domain (Fig. 1, MTR-PCR). These primers were designed to amplifyDNA fragments of various size, ranging from 575 nts in OBDV to 689 nt in EMV.
The second set of primers was designed on the RdRp domain of virus replicases (RD set)comprising most of the sequence between Koonin’s [20] motifs I-V. This primer set consistedof a slightly degenerate sense primer RD1 and the complementary RD2 (Fig. 1, RD-PCR)which amplified a highly conserved region of 386 nt in all the above viruses.
PCR conditions
Denaturation temperature (94◦C for 2 min) was the same for both set of primers. Denatura-tion was followed by 35 amplification cycles under slightly different conditions (annealingtemperature and MgCl2 concentration) according to the primer set: (i) 3 mM MgCl2 andannealing temperature of 55◦C for MTR primers; (ii) 2 mM MgCl2 and annealing at 58◦Cfor RD primers. Annealing was preceeded by denaturation at 94◦C for 30 sec and followedby extension at 72◦C for 45 sec. Final elongation was at 72◦C for 7 min. Tenml aliquots ofPCR products were electrophoresed in 1.5% TBE agarose or 5% acrylamide and stained withethidium bromide or silver. Controls consisted of reverse transcribed dsRNA extracts fromvines infected by viruses other than GFkV-like viruses, i.e. vitiviruses or closteroviruses.
cDNA cloning and sequencing
Threeml aliquots of PCR products were ligated to a pGEM-T Easy vector (Roche Biochemi-cals) immediately after electrophoresis, and cloned inEscherichia coliDH5a cells. Sequenc-ing was performed on both cDNA strands by the dideoxynucleotide chain termination method[32] using35S-ATP and the Thermo Sequenase cycle sequencing kit (Amersham).

556 S. Sabanadzovic et al.
Computer-assisted analysis of nucleotide and protein sequences
Nucleotide and protein sequences were analysed using the Strider 1.1 program [21]. Proteinsequences were aligned with CLUSTAL W [33]. Tentative phylogenetic trees were con-structed, and bootstrap analysis made with the NEIGHBOR, SEQBOOT, PROTDIST, andCONSENSE programs of the PHYLIP package [15].
Virus-specific PCR detection
Based on the sequenced portions of the three viral genomes (GFkV, GAMaV, and RG40/5)virus-specific antisense primers denoted AS-AP (5′GGCCAACTGGGTGGAGACA3′), RG-AP (5GGCCAAACTGGGTGCTCACG3′), and FK-AP (5′GGCCAAATTGGCATTG-CATG3′) were designed and used with the sense primer RD1 (Fig. 1) for PCR assays directlyfrom crude grapevine sap extracts. The samples were prepared and reverse transcribed aspreviously described [29]. cDNAs were denaturated at 94◦C for 2 min and the target regionwas amplified by 35 cycles of PCR (94◦C for 30 sec, 58◦C for 35 sec and 72◦C for 35 sec)followed by final extension at 72◦C for 5 min.
Serology
Leaf extracts fromV. rupestrisinfected with GAMaV and fromV. viniferaaccessions GR40/5,AA41 and AA42 were tested in ELISA with a polyclonal antiserum to GFkV [4] and mon-oclonal antibodies to the same virus [3]. Furthermore, particles from concentrated partiallypurified preparations from leaf tissues of accession RG50/5 were exposed to the polyclonalantiserum to GFkV, as described by Milne and Luisoni [24].
Thin sectioning
Tissue fragments from newly produced or mature leaves or young succulent roots were ex-cised and processed according to standard procedures [22], i.e. fixation in 4% glutaraldehydein 0.05 M phosphate buffer, post-fixation in 1% osmium tetroxide for 2 h at 4◦C and stainingovernight in 2% aqueous uranyl acetate in the cold (4◦C). Embedding was in Spurr’s resinafter dehydration in graded ethanol dilutions. Thin section were stained with lead citratebefore viewing with a Philips 201 C electron microscope. Comparable samples from healthyseedlings ofV. viniferaand rooted cuttings ofV. rupestriswere similarly processed to serveas controls.
Results
RT-PCR analysis
The MTR primer set amplified a single DNA fragment of about 600 nt fromreverse transcribed cDNA synthesized on dsRNAs extracted from all grapevinesources (Fig. 2A). Conversely, PCR products amplified by the RD primer set hadan approximate size of 400 nts (Fig. 2B). No amplification products were obtainedfrom the controls (Fig. 2A, B).
Sequence analysis
Nucleotide sequences reported in this paper are deposited in EMBL databaseunder the accession numbers listed in Table 2. Analysis of the cloned cDNAfragments corresponding to the methyltransferase coding sequence showed that

Grapevine fleck virus-like viruses 557
Table 2. EMBL database accession numbers of thesequences reported in the present paper
Virus (isolate) RdRp sequence MTR sequence
GRGV (40/5) AJ249355 AJ249356GRGV (AA41) AJ249359 AJ249360GRGV (AA42) AJ249361 AJ249362GAMaV AJ249357 AJ249358GFkV AJ249363 AJ249364
GRGVGrapevine red globe virus;GAMaVgrapevine as-teroid mosaic-associated virus;GFkVgrapevine fleck virus
Fig. 2. RT-PCR products amplified from reverse transcribed dsRNA using degenerate MTR(A) and RD (B) primer sets. The order of samples is the same in both panels: GFkV (2);GAMaV (3); GRGV (RG 40/5, AA41 and AA42,4, 5 and6, respectively). Control is in7
and molecular markers (SIGMA PCR MW ladder) in1
the sequences varied slightly in size. Amplified DNA products from GFkV andGAMaV samples were 575 nucleotides (nt) in size, while those from RG40/5,AA41 and AA42 were three nucleotide shorter (572 nts). All these amplifiedfragments had high C content ranging from 39% in RG40/5 to 47% in GFkV.The deduced protein products were polypeptides with a size of 191 amino acids(aa) for GFkV and GaMAV, and 190 aa for RG40/5, AA41, and AA42. Databankcomparisons confirmed that the amplified sequences belonged in the replicasegene of positive sense RNA virus genomes, specifically in the MTR domain,extending towards the 3′ end of the ORF, as shown in Fig. 1. Computer-assistedanalysis of the amplified sequences showed that no ORFs other than replicaseoccurred in that part of the genome. Motifs characterizing putative movementproteins were not detected, as would have been expected with tymoviruses [25].
Comparisons of these sequences among themselves and with the correspond-ing sequences of tymoviruses and OBDV revealed extensive homologies. In par-ticular, the sequenced MTR region of GFkV (Fig. 3, Table 3) showed an averageaa identity slightly higher than 50% with the equivalent region of tymoviruses,and aa identity of 57% with OBDV. The MTR of GAMaV had aa identity of 78%with the corresponding region of OBDV, which was higher than that (56–61% aaidentity) found with tymoviruses. The MTR of viruses from accessions RG40/5,AA41, and AA42 had the highest aa identity among themselves (92–95%, Table 3in bold). The aa identity with OBDV (62%) was higher than with tymoviruses (52–

558 S. Sabanadzovic et al.
Fig. 3. Multiple amino acid sequence alignment of products amplified from MTR motifs IIIand IV of GFkV, GAMaV, GRGV (RG40/5, AA41, AA42) and the corresponding regions of
tymoviruses (EMV, PhMV, ScrMV, KYMV, TYMV) and marafiviruses (OBDV)
Table 3. Percent amino acid identity between methyltransferase (above the diagonal) and polymerase(below the diagonal) domains of grapevine fleck virus-like viruses and representatives of tymoviruses
and marafiviruses
GFkV RG40/5 AA41 AA42 GAMaV OBDV TYMV EMV KYMV ScrMV PhMV
GFkV 57 57 56 57 55 56 54 50 51 50RG40/5 63 95 92 60 62 52 55 54 54 55AA41 62 96 92 61 62 52 55 54 54 55AA42 65 95 97 58 60 45 48 47 50 47GAMaV 70 75 77 78 78 56 55 56 57 58OBDV 77 76 75 77 85 56 56 58 57 56TYMV 73 72 71 72 73 73 55 64 66 61EMV 73 66 65 67 68 73 78 69 72 72KYMV 71 66 65 66 71 76 81 75 68 67ScrMV 62 68 69 69 67 64 68 69 72 71PhMV 69 67 66 68 69 70 73 75 75 68

Grapevine fleck virus-like viruses 559
Fig. 4. Multiple amino acid sequence alignment of RD-amplified products from GFkV,GAMaV, GRGV (RG40/5, AA41, AA42) and the corresponding regions of tymoviruses(EMV, PhMV, ScrMV, KYMV, TYMV) and marafiviruses (OBDV). Highly conserved
regions are shaded
55%). When the comparison was among MTRs of GFkV, GAMaV and virusesfrom accessions RG40/5, AA41, and AA42 the identity level was 56% to 61%.(Fig. 3, Table 3).
A comparable homology was detected in the polymerase domain (Fig. 4,Table 3). However, the highest homology values (95–97% aa identitiy) werefound among viruses from accessions RG40/5, AA41, and AA42 (Table 3, inbold).
Phylogenetic analysis of the MTR domains of grapevine fleck-like viruseswith comparable sequences of members of the generaTymovirusandMarafivirus,showed the presence of two distinct clusters in the tree, comprising, respectively,all tymoviruses and all GFkV-like viruses plus OBDV. Three subgroups wereevident in this latter grouping, separating GFkV from RG40/5, AA41, and AA42,which were in turn distinct from OBDV and GAMaV (Fig. 5A). The phylogenetictree constructed with sequences from the RdRp domain resembled the MTR treein that one cluster contained tymoviruses and the other was again subdividedin three subgroups comprising: (i) GFkV; (ii) RG40/5, AA41, and AA42; (iii)GAMaV and OBDV (Fig. 5B).

560 S. Sabanadzovic et al.
Fig. 5. Comparative phylogenetic analysis of the methyltransferase motifs III and IV (A)and RdRp motifs II-V (B) of GFkV, GAMaV and GRGV (RG40/5, AA41, AA42) with ty-moviruses and marafiviruses. EMBL accession numbers of the sequences used are those listedin Table 2, plus: EMV (J04374); KYMV (D00637); ScrMV (J04375); TYMV (X16378);
PhMV S97776; OBDV (U87832). Potato X virus (PVX, M31541) was used as outgroup
Serology
RG40/5 particles were not decorated by the antiserum to GFkV and no positivereaction was observed in any of the tests in which GAMaV-infectedV. rupestrisandV. vinifera accessions GR40/5, AA41 and AA42 were assayed by ELISAwith a polyclonal antiserum and monoclonal antibodies to GFkV.
Virus-specific PCR detection
The RD degenerate primer set, based on a highly conserved region of the RdRpcoding sequence, amplified the expected fragment of 386 bp from cortical scrap-ing extracts from vines infected by GFkV (Fig. 6, lane 2), GAMaV (Fig. 6, lane6), and RG40/5 (Fig. 6, lane 10). However, when virus-specific antisense primerswere combined with the degenerate sense primer RD1 (Table 1) and used underthe conditions described in ‘Materials and methods’, only homologous virus se-quences were amplified, thus allowing a clear-cut discrimination between viruses(Fig. 6, lanes 3, 8, and 13). Using this primer combination GAMaV was identi-fied in an additional Californian accession (USA 11), known by indexing to beaffected by asteroid mosaic, and RG40/5 in six other Red Globe accessions fromdifferent Apulian (southern Italy) vineyards (not shown).
Cytology
The cytology of mesophyll and vascular bundle cells ofV. rupestrisleaves infectedwith GAMaV did not differ from that of the controls. Regardless of their nature,

Grapevine fleck virus-like viruses 561
Fig. 6. Identification of GFkV, GAMaV, and RG40/5 by RT-PCR in crude grapevine sapextracts. The degenerate primer set based on RdRp motifs (RD) aspecifically amplifies allthree viruses (2, 6, and10). Specific amplification is obtained with the virus-specific antisenseprimers FK-AP (GFkV,3), AS-AP (GAMaV,8), and RG-AP (GRGV,13). Molecular markers
in 1. Control (C, healthy grape) in14
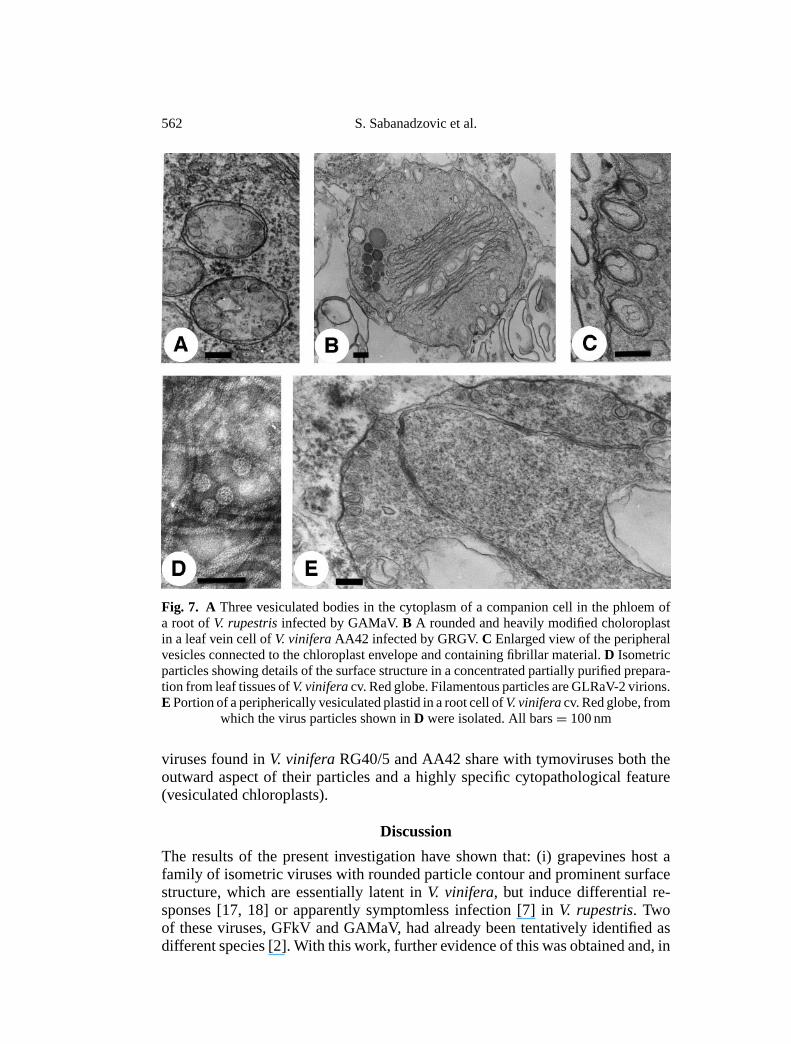
the cells were apparently normal in structural organization and outward appear-ance of the organelles and did not contain detectable virus particles. A comparablesituation was also encountered in most of the root specimens examined. However,in some GAMaV-infected samples, cytopathic structures were observed in differ-entiating sieve tubes, consisting of double-membraned oval-shaped bodies withperipheral vesiculation (Fig. 7A). The vesicles were also double-membraned, hada diameter not exceeding 80 nm, contained fine fibrils, and were connected to theouter envelope of the bodies, from the invagination of which they seemed to orig-inate (Fig. 7A). These cytopathic structures recalled very much the “vesiculatedbodies” derived from modified mitochondria, that characterize GFkV infections[6]. However, contrary to what was observed with GFkV infections, where in-tracellular particles are plentiful [5, 6], no virus particles were identified in thecells of GAMaV-infectedV. rupestris, regardless of whether or not they containedvesiculated mitochondria. GFkV and GAMaV had common features in the sizeand morphology of their particles, as seen in negatively stained preparations (Fig.7 D), and in the induction of seemingly identical cytopathic structures (vesiculatedbodies).
The cytology of mesophyll cells ofV. vinifera AA42 was also apparentlynormal. By contrast, differentiating sieve tubes and companion cells containedchloroplasts with a much altered shape and internal structure. Many of these or-ganelles were rounded, exhibited a profoundly modified internal membrane sys-tem, and a number of double-membraned vesicles connected to the organelle’slimiting membrane (Fig. 7B). These vesicles were rounded or flask-shaped, mea-sured up to 150 nm along the major axis, and contained fine fibrils (Fig. 7C).
Similarly modified plastids were also present in the phloem elements ofV.viniferaRG40/5 roots (Fig. 7E). In AA42 and RG40/5 samples, cells of leaf androot tissues with altered plastids were few, thus difficult to locate. Furthermore,none of the cells of the many specimens observed appeared to contain modifiedmitochondria.
Peripherally vesiculated chloroplasts are a hallmark of tymovirus infections,vesicles being the site of genome replication [12]. Thus, the GFkV-like isometric
562 S. Sabanadzovic et al.
Fig. 7. A Three vesiculated bodies in the cytoplasm of a companion cell in the phloem ofa root ofV. rupestrisinfected by GAMaV.B A rounded and heavily modified choloroplastin a leaf vein cell ofV. viniferaAA42 infected by GRGV.C Enlarged view of the peripheralvesicles connected to the chloroplast envelope and containing fibrillar material.D Isometricparticles showing details of the surface structure in a concentrated partially purified prepara-tion from leaf tissues ofV. viniferacv. Red globe. Filamentous particles are GLRaV-2 virions.E Portion of a peripherically vesiculated plastid in a root cell ofV. viniferacv. Red globe, from
which the virus particles shown inD were isolated. All bars= 100 nm
viruses found inV. viniferaRG40/5 and AA42 share with tymoviruses both theoutward aspect of their particles and a highly specific cytopathological feature(vesiculated chloroplasts).
Discussion
The results of the present investigation have shown that: (i) grapevines host afamily of isometric viruses with rounded particle contour and prominent surfacestructure, which are essentially latent inV. vinifera, but induce differential re-sponses [17, 18] or apparently symptomless infection [7] inV. rupestris. Twoof these viruses, GFkV and GAMaV, had already been tentatively identified asdifferent species [2]. With this work, further evidence of this was obtained and, in

Grapevine fleck virus-like viruses 563
addition, a seemingly new viral species was identified in cv Red Globe accessionRG50/5 and in Albanian grapevine accessions AA41 and AA42, for which theprovisional name Grapevine red globe virus (GRGV) is proposed; (ii) there areintriguing similarities in particle morphology and certain cytopathological fea-tures and a clear-cut phylogenetic relationship between the three above virusesand members of theTymovirusandMarafivirusgenera.
These conclusions are based on the following: (i) the GFkV antiserum did notdecorate GAMaV and GRGV particles, indicating that this virus is either serolog-ically unrelated to, or is so distant from the other two, that possible relationshipscould not be detected with the method used. (ii) GRGV elicits peripheral vesic-ulation of plastids, thus differing from GFkV and GAMaV, both of which causevesiculation of mitochondria. However, differences exist also between GAMaVand GFkV, the latter of which incites mitochondrial modifications more frequentand severe [5, 6] than those seen in GAMaV-infected cells; (iii) the amino acididentity level in the MTR and RdRp domains of GFkV, GAMaV and GRGVis in the range of 60–70%. Comparable identity levels were determined whenthese viruses were compared with five sequenced tymoviruses and OBDV, whichbelong in different genera. The likelihood that GFkV, GAMaV, and GRGV areseparate viral species is sustained by fact that homologies are much higher (92–97% identity) among RG40/5, AA41, and AA42 which, therefore, can be regardedas isolates of the same virus (GRGV). (iv) in both phylogenetic trees (Fig. 5A,B) GFkV, GAMaV, and GRGV appear to share the same replicase lineage amongthemselves and with a marafivirus (OBDV), but are positioned in distinct subclus-ters. GAMaV and OBDVseem to be phylogenetically closer to one another thanto GFkV or GRGV. However, unlike OBDV, GAMaV does not have a recognizedvector and OBDV, contrary to GAMaV, does not elicit mitochondrial vesiculation[13].
The PCR primers designed on the sequences obtained proved useful for thedifferential detection of each of these viruses in crude grapevine sap extracts. Thisfacilitates the identification of GAMaV, which until now was possible only byindexing onV. rupestris, and opens the way to reliable diagnosis of related viralspecies, such as GRGV.
GRGV is the 47th virus found in grapevines [34] and may not be the last. Avirus sharing high sequence homology with GRGV occurs in California (A.R.Rowhani, personal communication). Moreover, isometric viruses that incite pe-ripheral vesiculation of mitochondria or chloroplasts and have the same particlemorphology as GFkV, GAMaV, and GRGV, were recorded from Japan inV. ru-pestriswith a necrotic disease [23] and from Switzerland inV. viniferacv Gamayand Chasselas [14, 16]. The relationship of these latter viruses with the threeGFkV-like viruses discussed in this paper, remains to be ascertained.
Acknowledgements
Thanks are expressed to Dr. M.C. Edwards, USDA-ARS, ND State University, Fargo, USAfor supplying an isolate of oat blue dwarf virus.

564 S. Sabanadzovic et al.
References
1. Boscia D, Martelli GP, Savino V, Castellano MA (1991) Identification of the agent ofgrapevine fleck disease. Vitis 30: 97–105
2. Boscia D, Sabanadzovic S, Savino V, Kyriakopoulou PE, Martelli GP, Lafortezza R(1994) A non-mechanically transmissible isometric virus associated with asteroid mosaicof the grapevine. Vitis 33: 101–102
3. Boscia D, Elicio V, Savino V, Martelli GP (1995) Production of monoclonal antibodiesto grapevine fleck virus. Plant Pathol 44: 160–163
4. Boulila M, Boscia D, Di Terlizzi B, Castellano MA, Minafra A, Savino V, Martelli GP(1990) Some properties of a phloem-limited non mechanically-transmissibile grapevinevirus. J Phytopathol 129: 151–158
5. Castellano MA, Martelli GP (1984) Ultrastructure and nature of vesiculated bodiesassociated with isometric virus-like particles in diseased grapevines. J Ultrastruct Res89: 56–64
6. Castellano MA, Martelli GP, Savino V (1983) Virus-like particles and ultrastructuralmodifications in the phloem of leafroll-affected grapevines. Vitis 22: 23–39
7. Choueiri E, Boscia D, Digiaro M, Castellano MA, Martelli GP (1996) Some propertiesof a hitherto undescribed filamentous virus of the grapevine. Vitis 35: 91–93
8. Digiaro M, Boscia D, Simeone V, Savino V (1998) Gravi casi di accartocciamentofogliare su cultivar ad uva da tavola di recente introduzione in Puglia. Inf Fitopatol 48:76–79
9. Ding SW, Keese P, Gibbs A (1989) Nucleotide sequence of the ononis yellow mosaictymovirus genome. Virology 172: 555–563
10. Ding SW, Keese P, Gibbs AJ (1990) The nucleotide sequence of the genomic RNAof Kennedya yellow mosaic tymovirus-Jervis Bay isolate: relationship with potex- andcarlaviruses. J Gen Virol 71: 925–931
11. Dodds JA (1993) DsRNA in diagnosis. In: Matthews REF (ed) Diagnosis of plant virusdiseases. CRC Press, Boca Raton, pp 273–294
12. Dreher TW, Haenni AL, Mayo MA (2000) GenusTymovirus. In: Van RegenmortelMHV, Fauquet C, Bishop DHL, Carstens E, Estes MK, Lemon S, McGeoch D, Wick-ner, RB, Mayo MA, Pringle CR, Maniloff J (eds) Virus Taxonomy. Seventh Reportof the International Committee on Taxonomy of Viruses. Academic Press, New York(in press)
13. Edwards MC (2000) GenusMarafivirus.In: Van Regenmortel MHV, Fauquet C, BishopDHL, Karstens E, Estes MK, Lemon S, McGeoch D, Wickner RB, Mayo MA, PringleCR, Maniloff J (eds) Virus Taxonomy. Seventh Report of the International Committeeon Taxonomy of Viruses Academic Press, New York (in press)
14. Faoro F, Gugerli P (1997) Cytological alterations associated with an unidentified isomer-tric grapevine virus (UIGV). Extended Abstracts 12th Meeting of IGVG, Lisbon 1997:31–32
15. Felsenstein J (1989) PHYLIP-Phylogeny inference package (version 3.2). Cladistics5: 164–166
16. Gugerli P, Brugger JJ, Ramel ME (1997). Immunochemical and biological distinc-tion of grapevine leafroll-associated viruses 2 and 6 in complex infections with otherknown and unidentified viruses. Extended Abstracts 12th Meeting of IGVG, Lisbon 1997pp: 33–34
17. Hewitt WB, Goheen AC, Raski DJ, Gooding GV (1962) Studies on virus diseases of thegrapevine in California. Vitis 3: 57–83
18. Hewitt WB, Goheen AC, Cory L, Luhn C (1972) Grapevine fleck disease, latent in manyvarieties , is transmitted by graft inoculation. Ann Phytopathol: 43–47

Grapevine fleck virus-like viruses 565
19. Keese P, Mackenzie A, Gibbs A (1989) Nucleotide sequence of the genome of an Aus-tralian isolate of turnip yellow mosaic tymovirus. Virology 172: 536–546
20. Koonin EV (1991) The phylogeny of RNA-dependant RNA polymerases of positive-strand RNA viruses. J Gen Virol 72: 2 197–2 206
21. Marck C (1988) DNA Strider: a “C” program for the fast analysis of DNA and proteinsequences on the Apple Macintosh family of computers. Nucleic Acids Res 16: 1 829–1 836
22. Martelli GP, Russo M (1984) Use of thin sectioning for visualization and identificationof plant viruses. Methods Virol 8: 143–224
23. Matsumoto T, Ohki ST (1998) A possible new necrotic disease of grapevine associatedwith small isometric particles and novel membrane bound large particles. Ann Phy-topathol Soc Japan 64: 560–564
24. Milne RG, Luisoni E (1977) Rapid immune electron microscopy of virus preparations.Methods Virol 6: 265–281
25. Morch MD, Boyer JD, Haenni AL (1988) Overlapping open reading frames revealed bycomplete nucleotide sequencing of turnip yellow mosaic virus genomic RNA. NucleicAcids Res 16: 6 157–6 173
26. Osorio-Keese ME, Keese P, Gibbs A (1989) Nucleotide sequence of the genome ofeggplant mosaic tymovirus. Virology 172: 547–554
27. Ranjith-Kumar CT, Gopinath K, Jacob ANK, Srividhya V, Elango P, Savithri H (1998)Genomic sequence of physalis mottle virus and its evolutionary relationship with othertymoviruses. Arch Virol 143: 1 489–1 500
28. Rozanov MN, Koonin EV, Gorbalenya AE (1992) Conservation of the putative methyl-transferase domain: a hallmark of the ‘Sindbis-like’ supergroup of positive-strand RNAviruses. J Gen Virol 73: 2 129–2 134
29. Sabanadzovic S, Saldarelli P, Savino V (1996) Molecular detection of grapevine fleckvirus. Vitis 35: 137–140
30. Sabanadzovic S, Abou-Ghanem N, Saldarelli P, Martelli GP (1997) Physico-chemicaland molecular characterization of grapevine fleck virus. Extended Abstracts 12th Meet-ing ICVG, Lisbon, 1997 pp: 25–26
31. Saldarelli P, Minafra A, Martelli GP, Walter B (1994) Detection of grapevine leafroll-associated closterovirus III by molecular hybridization. Plant Pathol 43: 91–96
32. Sanger F, Nicklen S, Coulson AR (1977) DNA sequencing with chain-terminating in-hibitors. Proc Natl Acad Sci USA 74: 5 463–5 467
33. Thompson JD, Higgins DG, Gibson TJ (1994) CLUSTAL W: improving the sensitivityof progressive multiple alignment through sequence weighting, position-specific gappenalties and weight matrix choice. Nucleic Acids Res 22: 4 673–4 680
34. Walter B, Martelli GP (1997) Clonal and sanitary selection of the grapevine. In: Walter B(ed) Sanitary selection of the grapevine. Protocols for detection of viruses and virus-likediseases. Les Colloques 86, 43–95. INRA Editions, Paris
Authors’ address: Dr. G. P. Martelli, Dipartimento di Protezione delle Piante, ViaAmendola 165/A I-70126 Bari, Italy.
Received July 15, 1999




















