
Physiological tools for irrigation scheduling in grapevine (Vitis vinifera L.)
12
Physiological tools for irrigation scheduling in grapevine (Vitis vinifera L.) An open gate to improve water-use efficiency? J. Cifre, J. Bota, J.M. Escalona, H. Medrano, J. Flexas * Laboratori de Fisiologia Vegetal, Departament de Biologia, Universitat de les Illes Balears, Carretera de Valldemossa Km 7.5, 07122 Palma de Mallorca, Balearic Islands, Spain Abstract Grapevine is a traditionally non-irrigated crop that occupies quite an extensive agricultural area in dry lands and semi-arid regions. Recently, irrigation was introduced to increase the low land yield, but a good compromise between grape quality and yield is of major importance for the achievement of high-quality products as wine. Therefore, water-use-efficient irrigation and regulated deficit irrigation programs need to be developed to improve water-use efficiency, crop productivity and quality in semi- arid crops. In the present review, current knowledge on grapevine responses to water stress is summarised. Based on this knowledge, the usefulness of different physiological parameters is discussed, and current knowledge on their applicability for water stress detection and irrigation management in grapevines is reviewed. Partial root drying (PRD) and regulated deficit irrigation (RDI) programs are proposed as the most promising tools for such purpose. Among RDI programs, several parameters are proposed as putative indicators of irrigation convenience and dosage. These are sap flow (measured by sap flow meters), trunk growth variations (measured by linear transducers of displacement, LTDs), canopy temperature (assessed by infrared thermometry), reflectance indices (measured by spectroradiometers or specifically designed instruments) and chlorophyll fluorescence (measured by either active or passive fluorometers). Advantages and disadvantages of all these tools are discussed. # 2004 Elsevier B.V. All rights reserved. Keywords: Grapevines; Water stress; Water-use efficiency; Irrigation scheduling; Physiological indicators; Sap flow; Remote sensing; Reflectance indices; Chlorophyll fluorescence; Infrared thermometry; Linear transducers of displacement 1. Introduction Water is the most limiting resource in the Mediterranean region. Rainfall is scarce and irregu- larly distributed along the year, and climate change models predict even more arid conditions for the near future (Ragab and Prudhomme, 2002). Therefore, increasing water-use efficiency (WUE) should be a key issue for research (Al-Kaisi and Yin, 2003) and it is currently a priority for the United Nations policy, what is called the ‘Blue Revolution’ and summarised as ‘more crop per drop’(Annan, 2000). www.elsevier.com/locate/agee Agriculture, Ecosystems and Environment 106 (2005) 159–170 * Corresponding author. Tel.: +34 971 172365; fax: +34 971 173184. E-mail address: jaume.fl[email protected] (J. Flexas). 0167-8809/$ – see front matter # 2004 Elsevier B.V. All rights reserved. doi:10.1016/j.agee.2004.10.005
Transcript of Physiological tools for irrigation scheduling in grapevine (Vitis vinifera L.)

www.elsevier.com/locate/agee
Agriculture, Ecosystems and Environment 106 (2005) 159–170
Physiological tools for irrigation scheduling in
grapevine (Vitis vinifera L.)
An open gate to improve water-use efficiency?
J. Cifre, J. Bota, J.M. Escalona, H. Medrano, J. Flexas*
Laboratori de Fisiologia Vegetal, Departament de Biologia, Universitat de les Illes Balears,
Carretera de Valldemossa Km 7.5, 07122 Palma de Mallorca, Balearic Islands, Spain
Abstract
Grapevine is a traditionally non-irrigated crop that occupies quite an extensive agricultural area in dry lands and semi-arid
regions. Recently, irrigation was introduced to increase the low land yield, but a good compromise between grape quality and
yield is of major importance for the achievement of high-quality products as wine. Therefore, water-use-efficient irrigation and
regulated deficit irrigation programs need to be developed to improve water-use efficiency, crop productivity and quality in semi-
arid crops.
In the present review, current knowledge on grapevine responses to water stress is summarised. Based on this knowledge, the
usefulness of different physiological parameters is discussed, and current knowledge on their applicability for water stress
detection and irrigation management in grapevines is reviewed. Partial root drying (PRD) and regulated deficit irrigation (RDI)
programs are proposed as the most promising tools for such purpose. Among RDI programs, several parameters are proposed as
putative indicators of irrigation convenience and dosage. These are sap flow (measured by sap flow meters), trunk growth
variations (measured by linear transducers of displacement, LTDs), canopy temperature (assessed by infrared thermometry),
reflectance indices (measured by spectroradiometers or specifically designed instruments) and chlorophyll fluorescence
(measured by either active or passive fluorometers). Advantages and disadvantages of all these tools are discussed.
# 2004 Elsevier B.V. All rights reserved.
Keywords: Grapevines; Water stress; Water-use efficiency; Irrigation scheduling; Physiological indicators; Sap flow; Remote sensing;
Reflectance indices; Chlorophyll fluorescence; Infrared thermometry; Linear transducers of displacement
1. Introduction
Water is the most limiting resource in the
Mediterranean region. Rainfall is scarce and irregu-
* Corresponding author. Tel.: +34 971 172365;
fax: +34 971 173184.
E-mail address: [email protected] (J. Flexas).
0167-8809/$ – see front matter # 2004 Elsevier B.V. All rights reserved
doi:10.1016/j.agee.2004.10.005
larly distributed along the year, and climate change
models predict even more arid conditions for the near
future (Ragab and Prudhomme, 2002). Therefore,
increasing water-use efficiency (WUE) should be a
key issue for research (Al-Kaisi and Yin, 2003) and it
is currently a priority for the United Nations policy,
what is called the ‘Blue Revolution’ and summarised
as ‘more crop per drop’ (Annan, 2000).
.

J. Cifre et al. / Agriculture, Ecosystems and Environment 106 (2005) 159–170160
Grapevine (Vitis vinifera L.) is a traditionally non-
irrigated crop that occupies quite an extensive
agricultural area in dry lands and semi-arid regions.
Recently, irrigation was introduced to increase the low
land yield. Grapevine is cultivated to obtain table
grape, raisins or wine. In the latter case at least, a good
compromise between grape quality and yield is of
major importance for the achievement of high-quality
premium wines. In the case of producing grapevines,
even when a certain variability in the water-use
efficiency is present (Bota et al., 2001), the wine-
market-oriented production is restricted for the
necessity of the highest grape quality standards as
well as for certain regional authorised varieties, which
commonly offer the tipicity of wine characteristics
(‘Denominaciones de Origen’, ‘Appellations d’Ori-
gine Controlees’, etc.). Therefore, for grapevine as for
other extensive crops irrigation remains up to now the
most likely way to reduce drought impacts on yield.
Hence, water-use-efficient irrigation or regulated
deficit irrigation (RDI) programs would still be one
of the most desirable tools to improve water-use
efficiency and crop productivity in semi-arid areas
(Boyer, 1996).
The recent generalisation of vine irrigation in
countries with dry summer has lead to some
controversy, due to the not fully understood relation-
ships between grapevine photosynthesis and fruit
yield and quality. Irrigation substantially increases
photosynthesis, and grape yield is increased by 1.5–4-
fold, depending on the irrigation timing, the amount of
water applied, the cultivar, the environmental condi-
tions and other cultural practices (Bravdo et al., 1985;
Hepner et al., 1985; Matthews et al., 1987; Schultz,
1996; Williams, 1996; Escalona et al., 2003). On the
other hand, it seems that, up to a certain amount of
added water, no effects are observed on grape and
wine quality, even when grape yield is increased
(Bravdo et al., 1985; Hepner et al., 1985; Medrano et
al., 2003; Escalona and Medrano, personal commu-
nication). However, larger amounts of water, although
further increasing grape yield, have a negative effect
on quality, mainly due to colour losses, low sugar
content and acidity imbalances (Bravdo et al., 1985;
Hepner et al., 1985; Matthews et al., 1990; Cacho et
al., 1992; Esteban et al., 1999, 2001). Therefore, a
deep knowledge of the mechanisms that regulate plant
carbon assimilation and partitioning under different
water regimes is of great interest in the frame of
precision agriculture, since these mechanisms play an
important role in the regulation of the fragile balance
between grape yield and quality. Eventually, this will
lead to the description of physiologically based
criteria for irrigation scheduling.
The aim of the present work is to review recent
contributions on the effects of soil water availability
on grapevines and to show the potential interest of
some physiological indicators of grapevine water
status which could better assess the irrigation schedule
and dosage for a sustainable, water-use-efficient
production of high-quality grapevines.
2. Stomatal responses of grapevines
to soil water deficit
It has been known for many years that soil water
deficit reduces photosynthesis (Kriedemann and
Smart, 1971; Liu et al., 1978), vegetative growth
(Matthews et al., 1987; Schultz and Matthews, 1988a),
and reproductive growth and yield in grapevines
(Kliewer et al., 1983; Bravdo et al., 1985; Hepner et
al., 1985; Matthews and Anderson, 1988, 1989).
Stomatal closure is among the first processes
occurring in the leaves in response to drought. A good
relationship between stomatal conductance (gs) and
leaf water potential and/or water content has been
observed in some grapevine genotypes and conditions
(Liu et al., 1978; Gamon and Pearcy, 1990; Rodrigues
et al., 1993; Naor et al., 1994), but not in many others
(Quick et al., 1992; Naor and Wample, 1994; Poni et
al., 1994; Bota et al., 2001; Flexas et al., 2002a;
Medrano et al., 2002; Schultz, 2003). The latter
represent a near-isohydric behaviour, which implies
that the same minimum leaf water potential can be
achieved at midday irrespective of soil water status.
This fact makes midday leaf water potential a poor
indicator of water stress, in contrast with pre-dawn
leaf water potential or the stem water potential that
may serve as good indicators (Schultz, 1998; Escalona
et al., 1999; Chone et al., 2001). Differences between
near-isohydric and anisohydric V. vinifera L. cultivars
seem to be related to differences in hydraulic
architecture (Schultz, 2003).
Whatever the regulation mechanisms of plant water
relations, tight stomatal closure occur at relatively low

J. Cifre et al. / Agriculture, Ecosystems and Environment 106 (2005) 159–170 161
Fig. 1. The relationship between daily maximum stomatal conduc-
tance in grapevines (V. vinifera L.) growing outdoors in Mallorca,
and submitted to different water supply. Triangles are values for 22
different varieties grown in pots without rootstock, as described by
Bota et al. (2001); open circles are values for potted plants of the
variety Tempranillo, grafted on rootstock R-110 and grown as
described by Flexas et al. (1999); closed circles are values from
a 10-year study in two varieties, Tempranillo and Manto Negro,
grafted on rootstock R-110 and growing in a commercial vineyard,
as described by Medrano et al. (2003). All data are average values of
6–8 replicates.
soil water deficit in many grapevine cultivars, leading
to a rapid reduction of gs as pre-dawn leaf water
potential decreases (Fig. 1, r2 = 0.50, see also Schultz,
2003). This fine regulation of stomatal conductance
depends on some hydraulic conductance variations
and on synthesis of abscisic acid (ABA) in the roots,
which is transported to leaves (Lovisolo et al., 2002).
Grapevines were among the first plant species in
which a direct role of ABA in stomatal closure was
demonstrated (Loveys and Kriedemann, 1974; Liu et
al., 1978; Loveys, 1984; Loveys and During, 1984).
More recently, Correia et al. (1995) were able to
demonstrate that the maximum daily stomatal con-
ductance (but not its diurnal fluctuation) was
determined by the xylem ABA concentration in
field-grown grapevines. In addition, it has been shown
that partial root drying in grapevines induces stomatal
closure and shoot growth reduction without modifying
leaf water status, a process in which a five-fold
increase in leaf ABA content is involved (Dry and
Loveys, 1999; Dry et al., 2000a, 2000b; Stoll et al.,
2000). On the basis of these observations, it is likely
that root ABA synthesis in response to drought
controls to some extent the stomatal responses in
grapevines, although this could also be modulated by
osmotic adjustment, xylem hydraulic conductivity,
and environmental factors such as air humidity
(During, 1987; Naor et al., 1994; Lovisolo et al.,
2002).
In summary, the tight regulation of stomatal closure
in grapevines in response to very mild soil water
deficits makes stomatal conductance itself a more
precise indicator of water stress than common water
relations’ parameters.
3. Downregulation of photosynthesis under
drought in grapevines
A close, curvilinear correlation between stomatal
conductance (gs) and net photosynthesis (AN) has been
described in grapevines, as in other species (Fig. 2A,
r2 = 0.89, see also Chaves et al., 1987; During, 1987;
Naor and Wample, 1994; Escalona et al., 1999; Flexas
et al., 2002a). Due to this correlation and to the fact
that stomatal closure is among the first events to occur
under drought, it was assumed that the drought-
induced decrease of photosynthesis was mediated by
stomatal closure (Kriedemann and Smart, 1971).
Under mild water stress, it is likely that grapevine
photosynthesis is depressed almost exclusively by
stomatal closure, as indicated by increased water-use
efficiency (i.e., the ratio of photosynthesis to
transpiration or stomatal conductance to water vapour,
see Fig. 2B). This is presumably a general feature for
most species. However, under more severe water stress
situations, non-stomatal inhibition of photosynthesis
has been described (Chaves, 1991; Cornic, 2000;
Flexas et al., 2004).
Flexas et al. (2002a) have recently shown that
drought-induced changes in many photosynthetic
parameters are more related to variations in maximum
daily gs than to variations in the most commonly used
water status parameters, like leaf water potential or
relative water content. In fact, using gs as an
integrative parameter reflecting the severity of water
stress, different grapevine cultivars (Flexas et al.,
2002a) or even different C3 species (Flexas and
Medrano, 2002; Medrano et al., 2002) followed a very
similar pattern of downregulation of different photo-
synthetic parameters in response to progressive soil

J. Cifre et al. / Agriculture, Ecosystems and Environment 106 (2005) 159–170162
Fig. 2. The relationship between (A) net CO2 assimilation (AN) and
stomatal conductance and (B) intrinsic water-use efficiency (AN/gs)
and stomatal conductance in grapevines (V. vinifera L.) growing
outdoors in Mallorca, and submitted to different water supply.
Symbols as in Fig. 1. All data are single values, taken during the
central hours of the day (i.e., at saturating light).
water depletion. In the case of grapevines, this
observation is consistent with the above-described
near-isohydric behaviour of many drought-adapted
cultivars and with the suggestion that ABA-induced
stomatal closure (and not decreased leaf water status)
is the most immediate leaf response to drought.
Using gs as an integrative parameter for the degree
of drought, three phases of photosynthesis response
can be differentiated along a water stress gradient, that
are shared by different grapevine cultivars (Flexas et
al., 2002a; Medrano et al., 2002):
(1) A
phase of mild water stress is defined for adecreasing range of gs from 0.5–0.7 to 0.15 mol
H2O m�2 s�1. This is characterised by a relatively
small decline of AN, which results in a progressive
increase of intrinsic water-use efficiency (AN/gs)
and a decline of sub-stomatal CO2 concentration
(Ci, Flexas et al., 2002a). As a consequence of
decreased CO2 availability in the mesophyll, the
rate of photorespiration increases, which enables
the maintenance of the thylakoid electron trans-
port rate (ETR, Flexas et al., 1999, 2000, 2002a).
At this stage, stomatal closure is probably the only
limitation to photosynthesis.
(2) A
moderate water stress phase is characterisedby intermediate gs values (0.15 > gs > 0.05 mol
H2O m�2 s�1). During this phase, a further
reduction in AN occurs and water-use efficiency
usually increases, but it decreases in some
cultivars (e.g. Naor et al., 1994). Ci still decreases,
but ETR and the carboxylation efficiency (e; i.e.,
the initial slope of AN–Ci curve) characteristically
decline during this phase (Flexas et al., 1999,
2002a). The decline of e is dominated by
decreased mesophyll conductance at this stage
(Flexas et al., 2002a), since the activity of
photosynthetic enzymes, such as Rubisco, is
mostly unaffected (Bota et al., 2004; but see
Maroco et al., 2002). NPQ, a chlorophyll
fluorescence parameter indicative of thermal
dissipation in the antenna of photosystem II,
increases under these conditions, and steady-state
chlorophyll fluorescence (Fs) drops under high
light (Flexas et al., 2000, 2002b). Therefore, in
this phase, stomatal limitations seem dominant
and photosynthesis is rapidly reversed upon re-
watering (Flexas et al., 1999), but non-stomatal
limitations are already developing (Naor et al.,
1994; Flexas et al., 2002a, 2002b; Maroco et al.,
2002).
(3) A
phase of severe water stress takes place when gsis very low (<0.05 mol H2O m�2 s�1). In this
stress phase, steeper reductions of AN, AN/gs, ETR
and e occur. NPQ further increases, and the
excitation capture efficiency of photosystem II
(Fv/Fm) is eventually reduced, especially during
very hot days (Flexas et al., 2002a; Medrano et al.,
2002). During this phase AN/gs decreases (During,
1987) and Ci steeply increases (Flexas et al.,
2002a), indicating that non-stomatal limitations to
photosynthesis become dominant. The rate of
photorespiration is decreased, but the ratio of
photorespiration to photosynthesis still increases,

J. Cifre et al. / Agriculture, Ecosystems and Environment 106 (2005) 159–170 163
maintaining ETR high relative to AN (During,
1988; Flexas et al., 1999, 2002a). In this phase,
photosynthesis did not recover after irrigation
(During, 1988; Quick et al., 1992), indicating that
non-stomatal inhibition is dominant. Further
decreases of e during this phase are likely to be
due to impaired Rubisco activity (Bota et al.,
2002, 2004; Flexas et al., 2002a; Maroco et al.,
2002). A general decline in the activity and
amount of photosynthetic enzymes is observed in
this phase (Maroco et al., 2002; Flexas et al.,
2004).
4. Physiological tools for water stress detection
and management of grapevine irrigation
As mentioned in previous sections, for water stress
situations characterised by gs values above 0.05–
0.1 mol H2O m�2 s�1 photosynthesis is mainly lim-
ited by stomatal closure, and a complete recovery of
the maximum AN occurred just one night after
irrigation (Flexas et al., 1999). For more prolonged
and/or more pronounced water deficits leading to
further decreases of gs, non-stomatal limitations
(metabolic and/or restricted internal CO2 diffusion)
are dominant and photosynthesis did not recover
rapidly after irrigation (During, 1988; Quick et al.,
1992). Therefore, maximising WUE implies a certain
reduction in crop yield. Interestingly, in grapevines
(see above sections), highest crop load is linked to low
grape quality. Thus, in fact, limitations to grape yield
are a common practice (if not compulsory) for a
market standard wine production and premium wines.
Thus, being an extensive crop in drought-prone areas,
irrigation would give more benefits if schedules and
dosages were oriented to maximise water-use effi-
ciency (Fereres, 2003).
Based on this knowledge, physiologically based
irrigation tools may attempt to maintain the plants at a
limit between water stress and excess water con-
sumption, thus making a rational use of irrigation
water. This kind of irrigation may save water with
respect to empirical irrigation, improving yield as
compared with rainfed grapevines, and maintaining
the high fruit quality. Partial root drying (PRD) and
regulated deficit irrigation (RDI) programs are
currently envisaged as the most promising irrigation
tools based on physiological knowledge of grapevine
response to water stress.
Partial root drying (PRD) consists in drip irrigation
alternating from one side of the row to the other, thus
allowing that some roots of the plants are always in
contact with drying soil. The aim of this technique is to
control excessive vegetative vigour and save irrigation
water without influencing fruit yield and quality. This
treatment induces root ABA synthesis in the dried
zone (Dry and Loveys, 1999; Dry et al., 2000a,
2000b), leading to partial stomatal closure without
reducing leaf water status. Therefore, by adjusting the
proportion of roots that are irrigated, it is possible to
maintain the plants at the inflexion point of the AN–gs
relationship, thus saving a large amount of water with
only a small reduction of photosynthesis (Stoll et al.,
2000). Trials in Australia with this type of irrigation
have shown a significant reduction in vegetative
growth while maintaining fruit yield and quality,
saving in irrigation (Dry et al., 2001).
Regulated deficit irrigation (RDI) programs consist
in continuously monitoring some physiological para-
meters indicative of vine water stress. Irrigation
should be applied only when such a parameter drops
below a certain threshold value. Although leaf water
potential has been proposed as an indicative parameter
for RDI programs in grapevines (Girona et al., 2002)
and other extensive crops (Goldhamer et al., 1999), the
near-isohydric behaviour of many cultivars will
probably preclude its use, specially to detect mild
to moderate water stress situations, in which large
reductions of photosynthesis and yield occur without
much effects on canopy water relations. Presently, and
based on the knowledge of grapevine response to
progressive water stress described on the above
sections, it seems reasonable that continuously
monitoring of some direct or indirect indicators of
crop gs or AN may allow deciding when and how much
water must be applied for a rational use of irrigation
water. To give priority to high water-use efficiency
over maximum yield, irrigation should be applied only
when the indicator parameter drops below a certain
threshold value, preferably corresponding to some gs
value between 0.05 and 0.15 mmol H2O m�2 s�1.
According to current knowledge on water stress
effects on grapevine photosynthesis, yield and quality,
maintaining grapevines at such gs range may, in
principle, allow: (1) maximum water-use efficiency,

J. Cifre et al. / Agriculture, Ecosystems and Environment 106 (2005) 159–170164
(2) rapid reversibility of photosynthesis upon irriga-
tion, (3) relatively moderate yield loses as compared to
intensive irrigation and (4) maintenance of optimum
grape quality characters.
Several contact and remote sensing approaches
have been proposed as such indicators of gs. Among
contact approaches, the most promising are (1) sap
flow meters and (2) linear transducers of displacement
(LTDs). Among the remote sensing approaches, (3)
thermal imaging, (4) reflectance indices and (5)
chlorophyll fluorescence have been evaluated with
some success.
4.1. Sap flow meters
Sap flow measurements give reliable, direct
estimates of plant or shoot water loss without
disturbing the conditions of the leaf environment
(Shulze et al., 1985; Ansley et al., 1994; Fernandez et
al., 2001). In irrigated grapevines, sap flow measure-
ments were shown as a good tool to estimate canopy
transpiration (Yunusa et al., 2000). More recently,
Escalona et al. (2002) have demonstrated a very high
correlation between single leaf gs and the instanta-
neous sap flow in water-stressed, potted grapevines
(see also Table 1). For the total daily water
consumption, the correlation was even higher
(r2 = 0.98) and close to a 1:1 relationship. Net
photosynthesis rate was correlated as well with sap
flow values (r2 = 0.78), and this correlation increased
(r2 = 0.91) when the daily balances of both parameters
were compared. Ginestar et al. (1998) have already
started to apply sap flow measurements to establish
different irrigation coefficients to Shiraz grapevines,
obtaining a gradient of grape yield and qualities. The
main advantages of sap flow measurements may be:
(a) the fact that they directly reflect canopy sap flow
and transpiration, therefore being directly related to
Table 1
Threshold values of sap flow, net CO2 assimilation (AN), and stomatal co
50 mm) null and minimum (around �50 mm)
Trunk
growth (mm)
Maximum daily
Sap flow (g m�2 h�1)
Daily Sap flow
(g m�2 day�1)
50 150 1174
0 83 630
�50 36 320
Values inferred from data in Escalona et al. (2002).
whole-canopy gs, and (b) the fact that they correlate
with gs at any given range of the latter, therefore
allowing a continuous monitoring of gs from full water
availability to severe water depletion. The principal
disadvantages are: (a) that the sensors are quite
expensive, thus not allowing establishing a high
number of them per field; and (b) the fact that this is a
contact or intrusive technique, which may imply some
interference with plant performance.
4.2. Linear transducers of displacement
LTDs were mainly introduced by Garnier and
Berger (1986), and envisaged as a powerful tool for
irrigation scheduling (Fereres et al., 1999; Moriana et
al., 2000). They permit the continuous measurements
of the stem diameter, which can be related to plant
growth, water-use and status. The shrinking and
swelling of the extensible plant tissues provide an
indirect measure of the transpiration streams during
daylight periods, and is related to changes in water
content and turgor potential of their cells. In spite of its
potential usefulness, little attention has been paid to
this technique in grapevine studies, although Escalona
et al. (2002) have recently demonstrated that diurnal
stem diameter variations closely track sap flow (see
also Table 1). The main advantage of this technique is
that the sensors are very inexpensive, thus allowing the
establishment of a high number of them per field. As
remarkable disadvantages, there is the fact that this is a
contact technique, which may imply some interfer-
ence with plant performance, and that any possible
correlation with gs would be somewhat indirect.
4.3. Infrared thermometry
It has been long recognised that leaf or canopy
temperature is highly dependent on the rate of
nductance (gs), at which trunk growth is either maximum (around
Leaf-level AN
(mmol m�2 s�1)
Daily carbon gain
(g m�2 day�1)
Leaf-level gs
(mol m�2 s�1)
11.1 3.4 0.15
6.9 2.6 0.09
3.7 2.1 0.02

J. Cifre et al. / Agriculture, Ecosystems and Environment 106 (2005) 159–170 165
transpiration and can therefore be used as an indicator
of stomatal opening (Sinclair et al., 1984). Accord-
ingly, infrared thermometry has been developed as a
means for irrigation scheduling (Jones, 1999; Jones et
al., 2002). The main advantages of this tool are: (a) the
potential amplitude of its fields of view, allowing
measurements at different scales ranging from single
leaves to several hectares, and (b) the fact that, in
principle, thermometric signals should correlate with
gs at any given range of the latter, therefore allowing a
continuous monitoring of gs from full water avail-
ability to severe water depletion. Riou et al. (2000)
have recently described a thermometric parameter
sensible to water stress in grapevines, and Jones et al.
(2002) have shown reasonable estimations of whole
canopy gs based on thermometric determinations. The
principal disadvantages are: (a) the possibility of
misleading interpretations whenever the temperature
sensed is biased by, for example, bare soil in the
sensor’s field of view, although recent investigations
have shown that this problem can be avoided by the
use of modern thermal imagers with associated image
analysis software (Jones et al., 2002); and (b) the fact
that scaling-up to satellite level for regional-scale
purposes would be difficult due to the strong
absorption of infrared radiation by the atmosphere.
Despite its big potential interest, much knowledge has
still to be gained prior to this tool becomes easily used
for irrigation scheduling.
4.4. Canopy reflectance indices
Different parameters reflecting changes in canopy
reflectance at different wavelengths have been used
for monitoring plant performance. The general
advantage of all these parameters or indexes is that
they are not restricted by the dimension of their field of
view, allowing measurements from small to large
scales, including airborne and satellite detection
(Gamon et al., 1990; Sellers et al., 1992; Penuelas
and Filella, 1998).
Among them, the simplest indexes, as the simple
ratio (SR = NIR/R, i.e., the ratio between near-
infrared to visible red) and the so-called normalised
difference vegetation index (NDVI = NIR � R/
NIR + R), correlate with the amount of green biomass
over an area and with the absorbed PPFD, and they
have even been shown to reflect stomatal conductance
and photosynthesis in unstressed plants (Sellers et al.,
1992; Penuelas and Filella, 1998). The main
disadvantage, that precludes their use for irrigation
purposes, is that they are quite static, failing to follow
gs and photosynthesis changes with developing
drought (Penuelas et al., 1994; Runyon et al., 1994).
A water index has been defined (WI = R900/R970)
that closely follows leaf water content and gs during
drought (Penuelas et al., 1993, 1994; Penuelas and
Filella, 1998). This seems a very promising tool for
drought assessment, provided that it performs better at
the canopy than at the leaf scale (Penuelas et al.,
1993). Its main disadvantage is that, because it
involves structural changes like loss of cell wall
elasticity, its variations appear only when RWC drops
below 85% (Penuelas et al., 1993). This precludes its
use for mild water stress assessment or for assessment
of plants, like many near-isohydric grapevine culti-
vars, capable of maintaining RWC within narrow
ranges.
The photochemical reflectance index (PRI = R531
� R570/R531 + R570) is strongly correlated to the
de-epoxidation state of the xanthophyll cycle (Gamon
et al., 1990; Penuelas et al., 1994; Penuelas and
Filella, 1998). PRI has been shown to correlate with
photosynthesis and gs, thus been capable to reflect
drought at the leaf level in many species (Penuelas et
al., 1998; Stylinski et al., 2002; Winkel et al., 2002),
including grapevines (Evain et al., 2004). Its principal
disadvantages are that (a) the precise PRI values as
well as their relationship to AN and gs seem dependent
on the species and/or the instrument used (Stylinski et
al., 2002; Winkel et al., 2002) and (b) on occasions it
fails to follow drought at the canopy scale, which is
probably due to leaf changes in orientation as a
consequence of wilting (Gamon et al., 1990; Penuelas
et al., 1994).
4.5. Chlorophyll fluorescence indices
Chlorophyll fluorescence indices have been pro-
posed as well as irrigation tools. With current
instrumentation, chlorophyll fluorescence can be
measured at short distances, of several meters, with
laser-based fluorometers (Flexas et al., 2000; Ounis
et al., 2001), and from airborne using sun-induced
chlorophyll fluorescence (Moya et al., 1998). How-
ever, satellite detection is theoretically possible if
J. Cifre et al. / Agriculture, Ecosystems and Environment 106 (2005) 159–170166
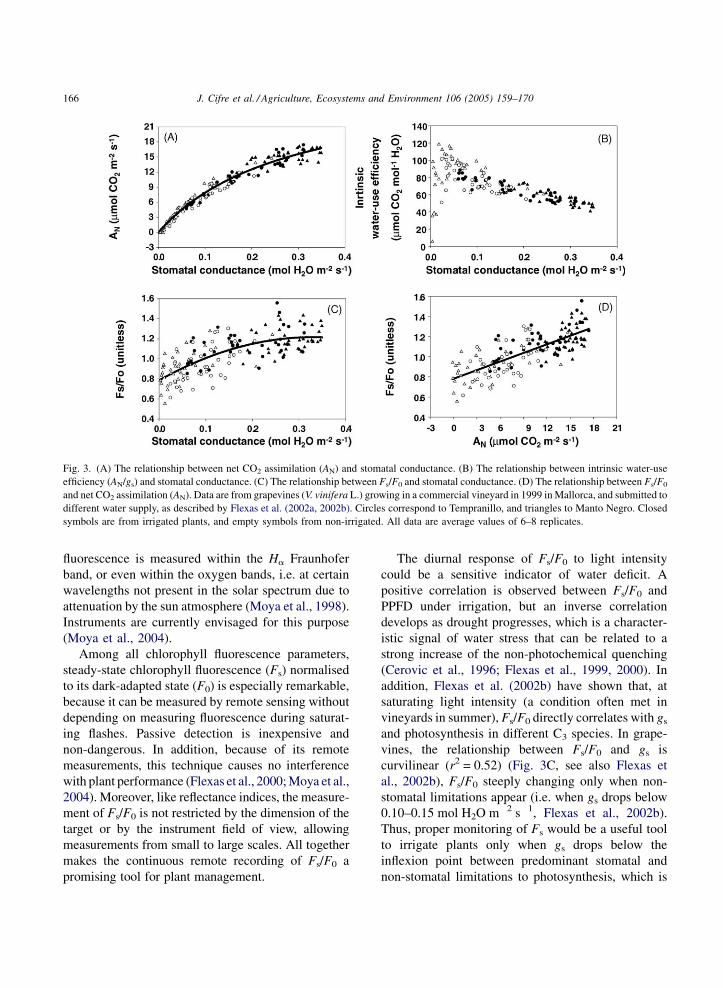
Fig. 3. (A) The relationship between net CO2 assimilation (AN) and stomatal conductance. (B) The relationship between intrinsic water-use
efficiency (AN/gs) and stomatal conductance. (C) The relationship between Fs/F0 and stomatal conductance. (D) The relationship between Fs/F0
and net CO2 assimilation (AN). Data are from grapevines (V. vinifera L.) growing in a commercial vineyard in 1999 in Mallorca, and submitted to
different water supply, as described by Flexas et al. (2002a, 2002b). Circles correspond to Tempranillo, and triangles to Manto Negro. Closed
symbols are from irrigated plants, and empty symbols from non-irrigated. All data are average values of 6–8 replicates.
fluorescence is measured within the Ha Fraunhofer
band, or even within the oxygen bands, i.e. at certain
wavelengths not present in the solar spectrum due to
attenuation by the sun atmosphere (Moya et al., 1998).
Instruments are currently envisaged for this purpose
(Moya et al., 2004).
Among all chlorophyll fluorescence parameters,
steady-state chlorophyll fluorescence (Fs) normalised
to its dark-adapted state (F0) is especially remarkable,
because it can be measured by remote sensing without
depending on measuring fluorescence during saturat-
ing flashes. Passive detection is inexpensive and
non-dangerous. In addition, because of its remote
measurements, this technique causes no interference
with plant performance (Flexas et al., 2000; Moya et al.,
2004). Moreover, like reflectance indices, the measure-
ment of Fs/F0 is not restricted by the dimension of the
target or by the instrument field of view, allowing
measurements from small to large scales. All together
makes the continuous remote recording of Fs/F0 a
promising tool for plant management.
The diurnal response of Fs/F0 to light intensity
could be a sensitive indicator of water deficit. A
positive correlation is observed between Fs/F0 and
PPFD under irrigation, but an inverse correlation
develops as drought progresses, which is a character-
istic signal of water stress that can be related to a
strong increase of the non-photochemical quenching
(Cerovic et al., 1996; Flexas et al., 1999, 2000). In
addition, Flexas et al. (2002b) have shown that, at
saturating light intensity (a condition often met in
vineyards in summer), Fs/F0 directly correlates with gs
and photosynthesis in different C3 species. In grape-
vines, the relationship between Fs/F0 and gs is
curvilinear (r2 = 0.52) (Fig. 3C, see also Flexas et
al., 2002b), Fs/F0 steeply changing only when non-
stomatal limitations appear (i.e. when gs drops below
0.10–0.15 mol H2O m�2 s�1, Flexas et al., 2002b).
Thus, proper monitoring of Fs would be a useful tool
to irrigate plants only when gs drops below the
inflexion point between predominant stomatal and
non-stomatal limitations to photosynthesis, which is

J. Cifre et al. / Agriculture, Ecosystems and Environment 106 (2005) 159–170 167
Fig. 4. Relationship between Fs/F0 and stomatal conductance in
grapevine plants (V. vinifera L.) growing outdoors conditions during
summer 1999 in Mallorca, as described by Flexas et al. (1999).
Closed circles correspond to data during progressive water stress
development, and open circles correspond to recovery upon irriga-
tion of the same plants.
close to the inflexion point of the AN–gs relationship
(Fig. 3A, r2 = 0.97). On the other hand, the relation-
ship between Fs/F0 and AN is linear (Fig. 3D,
r2 = 0.57), which may be an even more interesting
feature for irrigation purposes. The large scattering
present in Fig. 3 may not preclude the use of Fs/F0 as
an indicator parameter since, as shown by Ounis et al.
(2001), scattering disappear when Fs/F0 is measured at
a larger canopy spot, instead of at leaf level. Remote
sensing instruments are currently being designed for
measuring at large canopy spots (Moya et al., 2004).
Beyond the advantages already mentioned, this
parameter is of particular interest because its
correlation to gs seems to be identical during drought
and recovery (Fig. 4, r2 = 0.80). A main disadvantage
is that, presently, no commercial instrument available
is capable of passive remote sensing of chlorophyll
fluorescence. In addition, before this technique
became useful for irrigation purposes, more studies
are needed to establish its applicability in different
crops and under different conditions.
5. Conclusions
In summary, current knowledge on physiological
responses of grapevines to water stress allows
envisaging physiologically based irrigation tools for
water-use-efficient irrigation programs. Partial root
drying (PRD) and regulated deficit irrigation (RDI)
programs are such tools. Among RDI programs,
several indicator parameters seem promising. These
are sap flow, trunk growth variations, canopy
temperature, photochemical reflectance index (PRI)
and steady-state chlorophyll fluorescence. All these
tools have yielded promising results at the ecophy-
siological level. A combination of PRD and RDI
techniques has not been tested, and could yield even
more promising results. In addition, it would be time
now to conduct a number of studies at the agronomical
level to confirm the usefulness of these approaches,
prior to their extension as common, practical tools, for
vineyard management and irrigation.
Acknowledgements
We thank Herederos de Ribas S.A. (Mallorca) for
the management of the experimental vineyard. We
gratefully acknowledge the work of many people in
field data collection and processing (especially Joana
Barcelo, Javier Gulıas, Elkadri Lefi, Bartolome
Sampol and Diego Riera). Suggestions on the manu-
script by Dr. M. Ribas-Carbo are indebted. Financial
support from CICYT-Projects AGF94-0687, AGF97-
1180 and AGL-2001-1285-CO3-01 of the Plan
Nacional (Spain) are greatly acknowledged. Finally,
we wish to express our gratitude to Dr. C. Dıez de
Bethancourt for her encouragement to work in this
field and for her advice on chemical analysis.
References
Al-Kaisi, M.M., Yin, X., 2003. Effects of nitrogen rate, irrigation
rate and plant population on corn yield and water use efficiency.
Agron. J. 95, 1475–1482.
Annan, K., 2000. UN Millennium Summit. http://www.un.org/mil-
lennium/summit.htm.
Ansley, S.J., Dugas, W.A., Heuer, M.L., Trevino, B.A., 1994. Stem
flow and porometer measurements of transpiration from honey
mesquite (Prosopis glandulosa). J. Exp. Bot. 45, 847–856.
Bota, J., Flexas, J., Medrano, H., 2001. Genetic variability of
photosynthesis and water use in Balearic grapevine cultivars.
Ann. Appl. Biol. 138, 353–361.
Bota, J., Flexas, J., Keys, A.J., Loveland, J., Parry, M.A.J., Medrano,
H., 2002. CO2/O2 specificity factor of ribulose-1,5-bisphosphate
carboxylase/oxygenase in grapevines (Vitis vinifera L.): first in

J. Cifre et al. / Agriculture, Ecosystems and Environment 106 (2005) 159–170168
vitro determination and comparison to in vivo estimations. Vitis
41 (4), 163–168.
Bota, J., Medrano, H., Flexas, J., 2004. Is photosynthesis limited by
decreased Rubisco activity and RuBP content under progressive
water stress? New Phytol. 162, 671–681.
Boyer, J.S., 1996. Advances in drought tolerance in plants. Adv.
Agron. 56, 187–218.
Bravdo, B., Hepner, Y., Loinger, C., Cohen, S., Tabacman, H., 1985.
Effect of irrigation and crop level on growth, yield and wine
quality of Cabernet Sauvignon. Am. J. Enol. Viticult. 36 (2),
132–139.
Cacho, J., Fernandez, P., Ferreira, V., y Castells, J.E., 1992. Evolu-
tion of five anthocyanidin-3-glucosides in the skin of the Tem-
pranillo Moristel and Garnacha grape varieties and influence of
climatological variables. Am. J. Enol. Vitic. 43, 244–248.
Cerovic, Z.G., Goulas, Y., Goburnov, M., Briantais, J.M., Camenen,
L., Moya, I., 1996. Fluorosensing of water stress in plants:
diurnal changes of the mean lifetime and yield of chlorophyll
fluorescence, measured simultaneously and at distance with t-
LIDAR and modified PAM fluorometer in maize, sugar beet and
kalanchoe. Remote Sens. Environ. 58, 311–321.
Chaves, M.M., 1991. Effects of water deficits on carbon assimila-
tion. J. Exp. Bot. 42, 1–16.
Chaves, M.M., Tenhunen, J.D., Harley, P., Lange, O.L., 1987. Gas
exchange studies in two Portuguese grapevine cultivars. Physiol.
Plant. 70 (4), 639–647.
Chone, X., van Leeuwen, C., Dubordieu, D., Gaudillere, J.P., 2001.
Stem water potential is a sensitive indicator of grapevine water
status. Ann. Bot. 87, 477–483.
Cornic, G., 2000. Drought stress inhibits photosynthesis by decreas-
ing stomatal aperture – not by affecting ATP synthesis. Trends
Plant Sci. 5, 187–188.
Correia, M.J., Pereira, J.S., Chaves, M.M., Rodrigues, M.L.,
Pacheco, C.A., 1995. ABA xylem concentrations determine
maximum daily leaf conductance of field-grown Vitis vinifera
L. plants. Plant Cell Environ. 18, 511–521.
Dry, P.R., Loveys, B.R., 1999. Grapevine shoot growth and stomatal
conductance are reduced when part of the root system is dried.
Vitis 38, 151–156.
Dry, P.R., Loveys, B.R., During, H., 2000a. Partial drying of the
rootzone of grape. I. Transient changes in shoot growth and gas
exchange. Vitis 39, 3–7.
Dry, P.R., Loveys, B.R., During, H., 2000b. Partial drying of the
rootzone of grape. II. Changes in the pattern of root develop-
ment. Vitis 39, 3–7.
Dry, P.R., Loveys, B.R., McCarthy, M.G., Stoll, M., 2001. Strategic
irrigation management in Australian vineyards. J. Int. Sci. Vigne
Vin. 35, 129–139.
During, H., 1987. Stomatal responses to alterations of soil and air
humidity in grapevines. Vitis 26, 9–18.
During, H., 1988. CO2 assimilation and photorespiration of
grapevine leaves: responses to light and drought. Vitis 27,
199–208.
Escalona, J.M., Flexas, J., Medrano, H., 1999. Stomatal and
non-stomatal limitations of photosynthesis under water stress
in field-grown grapevines. Aust. J. Plant Physiol. 26, 421–
433.
Escalona, J.M., Flexas, J., Medrano, H., 2002. Drought effects on
water flow, photosynthesis and growth of potted grapevines.
Vitis 41, 57–62.
Escalona, J.M., Flexas, J., Bota, J., Medrano, H., 2003. From leaf
photosynthesis to grape yield: influence of soil water availabil-
ity. Vitis 42 (2), 57–64.
Esteban, M.A., Villanueva, M.J., Lissarrague, J.R., 1999. Effect of
irrigation on changes in berry composition during maturation.
Sugars, organic acids, and mineral elements. Am. J. Enol. Vitic.
50, 418–434.
Esteban, M.A., Villanueva, M.J., Lissarrague, J.R., 2001. Effect of
irrigation on changes in the anthocyanin composition of the skin
of cv Tempranillo (Vitis vinifera L) grape berries during ripen-
ing. J. Sci. Food Agric. 81, 409–420.
Evain, S., Flexas, J., Moya, I., 2004. A new instrument for passive
remote sensing. 2. Measurement of leaf and canopy reflectance
changes at 531 nm and their relationship with photosynthesis
and chlorophyll fluorescence. Remote Sens. Environ. 91, 175–
185.
Fereres, E., 2003. Water productivity of crop plants in the
Mediterranean. In: Proceedings of the Workshop Optimisation
of Water Use by Plants in the Mediterranean, Cala Bona
(Mallorca).
Fereres, E., Goldhamer, D., Cohen, M., Girona, J., Marta, M., 1999.
Continuous trunk diameter recording is a sensitive indicator for
irrigation of peach. Calc. Agric. 53 (4), 21–25.
Fernandez, J.E., Palomo, M.J., Diaz-Espejo, A., Clothier, B.E.,
Green, S.R., Giron, I.F., Moreno, F., 2001. Heat-pulse measure-
ments of sap flow in olives for automating irrigation: tests, root
flow and diagnostics of water stress. Agric. Water Manage. 51
(2), 99–123.
Flexas, J., Medrano, H., 2002. Drought-inhibition of photosynthesis
in C3 plants: stomatal and non-stomatal limitation revisited.
Ann. Bot. 89, 183–189.
Flexas, J., Escalona, J.M., Medrano, H., 1999. Water stress induces
different levels of photosynthesis and electron transport rate
regulations in grapevines. Plant Cell Environ. 22, 39–48.
Flexas, J., Briantais, J.-M., Cerovic, Z., Medrano, H., Moya, I.,
2000. Steady-state and maximum chlorophyll fluorescence
responses to water stress in grapevine leaves: a new remote
sensing system. Remote Sens. Environ. 73, 283–297.
Flexas, J., Bota, J., Escalona, J.M., Sampol, B., Medrano, H., 2002a.
Effects of drought on photosynthesis in grapevines under field
conditions: an evaluation of stomatal and mesophyll limitations.
Funct. Plant Biol. 29, 461–471.
Flexas, J., Escalona, J.M., Evain, S., Gulıas, J., Moya, I., Osmond,
C.B., Medrano, H., 2002b. Steady-state chlorophyll fluorescence
(Fs) measurements as a tool to follow variations of net CO2
assimilation and stomatal conductance during water-stress in C3
plants. Physiol. Plant. 114, 231–240.
Flexas, J., Bota, J., Loreto, F., Cornic, G., Sharkey, T.D., 2004.
Diffusive and metabolic limitations to photosynthesis under
drought and salinity in C3 plants. Plant Biol. 6, 1–11.
Gamon, J.A., Field, C.B., Bilger, W., Bjorkman, O., Fredeen, A.L.,
Penuelas, J., 1990. Remote sensing of the xanthophyll cycle and
chlorophyll fluorescence in sunflower leaves and canopies.
Oecologia 85, 1–7.

J. Cifre et al. / Agriculture, Ecosystems and Environment 106 (2005) 159–170 169
Gamon, J.A., Pearcy, R.W., 1990. Photoinhibition in Vitis califor-
nica: interactive effects of sunlight, temperature and water
status. Plant Cell Environ. 13, 267–275.
Garnier, E., Berger, A., 1986. Effect of water stress on stem diameter
changes of peach trees growing in the field. J. Appl. Ecol. 23,
193–209.
Ginestar, C., Eastham, J., Gray, S., Iland, P., 1998. Use of sap-flow
sensors to schedule vineyard irrigation. I. Effects of post-ver-
aison water deficit on water relations, vine growth, and yield of
Shiraz grapevines. Am. J. Enol. Viticult. 49, 413–420.
Girona, J., Mata, M., Del Campo, J., Arbones, A., 2002. Programa-
cion de riego en vina en base al potencial hıdrico de la hoja.
Resultados del primer ano (2001). Actas del VI Symposium
Hispano-Portugues de Relaciones Hıdricas en Plantas, Navarra,
Spain.
Goldhamer, D., Fereres, E., Cohen, M., Mata, M., Girona, J., 1999.
Sensitivity of continuous and discrete plant and soil water status
monitoring in peach trees subjected to deficit irrigation. J. Am.
Soc. Hort. Sci. 124 (4), 437–444.
Hepner, Y., Bravdo, B., Loinger, C., Cohen, S., Tabacman, H., 1985.
Effect of drip irrigation schedules on growth, yield, must
composition and wine quality of Cabernet Sauvignon. Am. J.
Enol. Viticult. 36, 77–85.
Jones, H.G., 1999. Use of thermography for quantitative studies of
spatial and temporal variation of stomatal conductance over leaf
surfaces. Plant Cell Environ. 22, 1043–1055.
Jones, H.G., Stoll, M., Santos, T., de Sousa, C., Chaves, M.M.,
Grant, O.M., 2002. Use of infrared thermography for monitoring
stomatal closure in the field: application to grapevine. J. Exp.
Bot. 53, 2249–2260.
Kliewer, W.M., Freeman, B.M., Hosssom, C., 1983. Effect of
irrigation, crop level and potassium fertilization on Carignane
vines. I. Degree of water stress and effect on growth and yield.
Am. J. Plant Physiol. 34, 186–196.
Kriedemann, P.E., Smart, R.E., 1971. Effects of irradiance, tem-
perature, and leaf water potential on photosynthesis of vine
leaves. Photosynthetica 5, 15–19.
Liu, W.T., Pool, R., Wenkert, W., Kriedemann, P.E., 1978. Changes
in photosynthesis, stomatal resistance and abscisic acid of Vitis
labruscana through drought and irrigation cycles. Am. J. Enol.
Viticult. 29, 239–246.
Loveys, B.R., 1984. Diurnal changes in water relations and abscisic
acid in field-grown Vitis vinifera cultivars. III. The influence of
xylem-derived abscisic acid on leaf gas exchange. New Phytol.
98, 563–573.
Loveys, B.R., During, H., 1984. Diurnal changes in water relations
and abscisic acid in field-grown Vitis vinifera cultivars. II.
Abscisic acid changes under semiarid conditions. New Phytol.
97, 37–47.
Loveys, B.R., Kriedemann, P.E., 1974. Internal control of stomatal
physiology and photosynthesis. I. Stomatal regulation and asso-
ciated changes in endogenous levels of abscisic and phaseic
acids. Aust. J. Plant Physiol. 1, 407–415.
Lovisolo, C., Hartung, W., Schubert, A., 2002. Whole-plant hydrau-
lic conductance and root-to-shoot flow of abscisic acid inde-
pendently affected by water stress in grapevines. Funct. Plant
Biol. 29, 1349–1356.
Maroco, J.P., Rodrigues, M.L., Lopes, C., Chaves, M.M., 2002.
Limitations to leaf photosynthesis in field-grown grapevine
under drought – metabolic and modelling approaches. Funct.
Plant Biol. 29, 451–459.
Matthews, M.A., Anderson, M.M., 1988. Fruit ripening in Vitis
vinifera L.: responses to seasonal water deficits. Am. J. Enol.
Viticult. 39, 313–320.
Matthews, M.A., Anderson, M.M., 1989. Reproductive development
in grape (Vitis vinifera L.): responses to seasonal water deficits.
Am. J. Enol. Viticult. 40, 52–60.
Matthews, M.A., Anderson, M.M., Schultz, H.R., 1987. Phenologic
and growth-responses to early and late season water deficits in
Cabernet franc. Vitis 26 (3), 147–160.
Matthews, M.A., Ishii, R., Anderson, M.M., O’Mahony, M., 1990.
Dependence of wine sensory attributes on vine water status. J.
Sci. Food Agric. 51, 321–335.
Medrano, H., Escalona, J.M., Bota, J., Gulıas, J., Flexas, J., 2002.
Regulation of photosynthesis of C3 plants in response to pro-
gressive drought: the interest of stomatal conductance as a
reference parameter. Ann. Bot. 89, 895–905.
Medrano, H., Escalona, J.M., Cifre, J., Bota, J., Flexas, J., 2003. A
ten-year study on the physiology of two Spanish grapevine
cultivars under field conditions: effects of water availability
from leaf photosynthesis to grape yield and quality. Funct. Plant
Biol. 30, 607–619.
Moriana, A., Castro, J., Humanes, D., Orgaz, F., Pastor, M., Fereres,
E., 2000. The relations between trunk diameter fluctuations and
tree water status in olive trees (Olea europaea L.). Acta Hort.
537 (1), 293–297.
Moya, I., Camenen, L., Latouche, G., Mauxion, C., Evain, S.,
Cerovic, Z., 1998. An instrument for the measurement of sun-
light excited plant fluorescence. In: Garab, G. (Ed.), Photo-
synthesis: Mechanisms and Effects, vol. V. Kluwer Academic
Publishers, Dordrecht, pp. 4265–4270.
Moya, I., Camenen, L., Evain, S., Goulas, Y., Cerovic, Z.G.,
Latouche, G., Flexas, J., Ounis, A., 2004. A new instrument
for passive remote sensing. 1. Measurements of sunlight induced
chlorophyll fluorescence. Remote Sens. Environ. 91, 186–197.
Naor, A., Wample, R.L., 1994. Gas exchange and water relations of
field-grown Concord (Vitis labruscana Bailey) grapevines. Am.
J. Enol. Viticult. 45, 333–337.
Naor, A., Bravdo, B., Gelobter, J., 1994. Gas exchange and water
relations in field-grown Sauvignon blanc grapevines. Am. J.
Enol. Viticult. 45, 423–428.
Ounis, A., Evain, S., Flexas, J., Tosti, S., Moya, I., 2001. Adaptation
of a PAM-fluorometer for remote sensing of chlorophyll fluor-
escence. Photosynth. Res. 68, 113–120.
Penuelas, J., Filella, I., 1998. Visible and near-infrared reflectance
techniques for diagnosing plant physiological status. Trends
Plant Sci. 3, 151–156.
Penuelas, J., Filella, I., Biel, C., Serrano, L., Save, R., 1993. The
reflectance at the 950–970 nm region as an indicator of plant
water status. Int. J. Remote Sens. 14, 1887–1905.
Penuelas, J., Gamon, J.A., Fredeen, A.L., Merino, J., Field, C.B.,
1994. Reflectance indices associated with physiological changes
in nitrogen- and water-limited sunflower leaves. Remote Sens.
Environ. 48, 135–146.

J. Cifre et al. / Agriculture, Ecosystems and Environment 106 (2005) 159–170170
Penuelas, J., Filella, I., Llusia, J., Siscart, D., Pinol, J., 1998.
Comparative field study of spring and summer leaf gas exchange
and photobiology of the Mediterranean trees Quercus ilex and
Phillyrea latifolia. J. Exp. Bot. 49 (319), 229–238.
Poni, S., Lakso, A.N., Turner, J.R., Melious, R.E., 1994. Interactions
of crop level and late season water stress on growth and
physiology of field-grown Concord grapevines. Am. J. Enol.
Viticult. 45, 252–257.
Quick, W.P., Chaves, M.M., Wendler, R., David, M., Rodrigues,
M.L., Passaharinho, J.A., Pereira, J.S., Adcock, M.D., Leegood,
R.C., Stitt, M., 1992. The effect of water stress on photosynthetic
carbon metabolism in four species grown under field conditions.
Plant Cell Environ. 15, 25–35.
Ragab, R., Prudhomme, C., 2002. Climate change and water
resources management in arid and semi-arid regions: prospec-
tive and challenges for the 21st century. Biosyst. Eng. 81, 3–34.
Riou, C., Lebon, E., Lacroute, J., 2000. Validation of canopy
temperature as a water stress field indicator in vines. In:
Proceedings of the Sixth International Symposium on Grapevine
Physiology Biotechnology, Book of Abstracts, Heraklion,
Greece.
Rodrigues, M.L., Chaves, M.M., Wendler, R., David, M.M., Quick,
W.P., Leegood, R.C., Stitt, M., Pereira, J.S., 1993. Osmotic
adjustment in water stressed grapevine leaves in relation to
carbon assimilation. Aust. J. Plant Physiol. 20, 309–321.
Runyon, J., Waring, R.H., Goward, S.N., Welles, J.M., 1994.
Environmental limits on net primary production and light-use
efficiency across the Oregon transect. Ecol. Appl. 4, 226–237.
Schultz, H.R., 1996. Water relations and photosynthetic responses of
two grapevine cultivars of different geographical origin during
water stress. Acta Hort. 427, 251–266.
Schultz, H.R., 1998. Photosynthetic responses to drought of two
grapevine cultivars of contrasted origin. In: Garab, G. (Ed.),
Photosynthesis: Mechanisms and Effects, vol. V. Kluwer Aca-
demic Publishers, The Netherlands, pp. 3755–3760.
Schultz, H.R., 2003. Differences in hydraulic architecture account
for near-isohydric and anisohydric behaviour of two field-grown
Vitis vinifera L. cultivars during drought. Plant Cell Environ. 26
(8), 1393–1405.
Schultz, H.R., Matthews, M.A., 1988a. Vegetative growth distribu-
tion during water deficits in Vitis vinifera L. Aust. J. Plant
Physiol. 15, 641–656.
Sellers, P.J., Berry, J.A., Collatz, G.J., Field, C.B., Hall, F.G., 1992.
Canopy reflectance, photosynthesis, and transpiration. III. A
reanalysis using improved leaf models and a new canopy
integration scheme. Remote Sens. Environ. 42, 187–216.
Shulze, E.D., Cermark, J., Matyssek, R., Penka, M., Zimmermann,
R., Vasıcek, F., Gries, W., Kucera, J., 1985. Canopy transpiration
and water fluxes in the xylem of the trunk of Larix and Picea
trees – a comparison of xylem flow, porometer and cuvette
measurements. Oecologia 66, 475–483.
Sinclair, T.R., Tanner, C.B., Bennett, J.M., 1984. Water-use effi-
ciency in crop production. Bioscience 34, 36–40.
Stoll, M., Loveys, B., Dry, P., 2000. Hormonal changes induced by
partial rootzone drying of irrigated grapevine. J. Exp. Bot. 51,
1627–1634.
Stylinski, C.D., Gamon, J.A., Oechel, W.C., 2002. Seasonal patterns
of reflectance indices, carotenoid pigments and photosynthesis
of evergreen chaparral species. Oecologia 131, 366–374.
Williams, L.E., 1996. Grape. In: Zamski, E. (Ed.), Photoassimilate
Distribution in Plants and Crops: Source–Sink Relationships.
Marcel Dekker, New York, pp. 851–881.
Winkel, T., Methy, M., Thenot, F., 2002. Radiation use efficiency,
chlorophyll fluorescence, and reflectance indices associated with
ontogenic changes in water-limited Chenopodium quinoa
leaves. Photosynthetica 40, 227–232.
Yunusa, I.A.M., Walker, R.R., Loveys, B.R., Blackmore, D.H.,
2000. Determination of transpiration in irrigated grapevines:
comparison of the heat-pulse technique with gravimetric and
micrometeorological methods. Irrig. Sci. 20, 1–8.




















