

Occurrence of low molecular weight phenolics in Vitis vinifera red grape cultivars and their...
14
Occurrence of low molecular weight phenolics in Vitis vinifera red grape cultivars and their winemaking by-products from São Paulo (Brazil) Milene Teixeira Barcia a , Paula Becker Pertuzatti a , Daniele Rodrigues a , Sergio Gómez-Alonso b,c , Isidro Hermosín-Gutiérrez c,d, ⁎, Helena Teixeira Godoy a a Faculdade de Engenharia de Alimentos, Universidade Estadual de Campinas, Cidade Universitária “Zeferino Vaz”, s/n, 13083862, Campinas, São Paulo, Brazil b Fundación Parque Científico y Tecnológico de Albacete, Paseo de la Innovación, 1, 02006 Albacete, Spain c Instituto Regional de Investigación Científica Aplicada, Universidad de Castilla-La Mancha, Campus Universitario s/n, 13071 Ciudad Real, Spain d Escuela de Ingenieros Agrónomos, Universidad de Castilla-La Mancha, Ronda de Calatrava, 7, 13071 Ciudad Real, Spain abstract article info Article history: Received 15 January 2014 Accepted 29 March 2014 Available online 5 April 2014 Keywords: Vitis vinifera Grape marc Lees Anthocyanins Flavonols Hydroxycinnamic acids Stilbenes Drying process The content of low molecular weight phenolic compounds present in winemaking by-products (skins from grape marc and wine lees) of red wine made from the Vitis vinifera grape cultivars Cabernet Sauvignon and Cabernet Franc in the state of São Paulo (Brazil) has been examined. The study was extended to two consecutive years and showed that winemaking by-products still contained high concentrations of interesting functional and bioactive phenolic compounds. In addition to original grape phenolics, the winemaking by-products contained new phenolics originated by the action of enzymatic (yeast mediated) and chemical reactions during winemaking, including formation of anthocyanin-derived pigments and hydrolysis products from flavonol glycosides and tartaric esters of hydroxycinnamic acids. A total of 19 anthocyanins, 9 pyranoanthocyanins, 18 flavonols, 7 hydroxycinnamic acid derivatives, and 3 resveratrol-based stilbenes were identified. Wine lees have been revealed as an interesting source of flavonol aglycones and pyranoanthocyanins. The best sample preparation technique prior to the extraction of phenolic compounds, with regard to the maintenance of phenolic compound profiles and concentrations, was freeze-drying. However, in the case of lees, spray-drying showed to be also effective in the preservation of phenolic compounds and is a less expensive technique. Oven-drying at 50 °C should be avoided if there is interest in using winemaking by-products for further purposes, since phenolic compound concentrations were strongly reduced by thermal degradation. © 2014 Elsevier Ltd. All rights reserved. Introduction São Paulo is a southeastern Brazilian State where wine production activity is currently increasing. The wine industry produces a large amount of by-products and residues. All these residues contain biodegradable organic matter, the disposal of which creates serious environmental problems (Mendes, Prozil, Evtuguin, & Lopes, 2013; Rebello et al., 2013). Grape marc (a set of skins and seeds) is the most abundant residue of wine making, which is generated after concomitant fermentation and maceration of the grapes during red wine production. After fermen- tation of the must, a decanting wine process occurs, where the supernatant is separated from the lees. The lees are composed of fine particles of grape residue and yeasts (Cortés, Rodríguez, Salgado, & Domínguez, 2011; Maragkoudakis et al., 2013; Paradelo, Moldes, & Barral, 2010). Over the past few years, the by-products of wine have attracted attention as possible sources of phenolic compounds since these com- pounds in the grape are only partially transferred to the wine. However, the majority of the phenolic compounds still remain in the residue as they are present in the solid parts of the grape. Thus, the recovery of phenols from industrial residues is gaining considerable attention, especially because of the antioxidant and biological activities attributed to these compounds (Cheng, Bekhit, McConnell, Mros, & Zhao, 2012; Rubilar, Pinelo, Shene, Sineiro, & Nuñez, 2007; Shrikhande, 2000). The use of bioactive compounds obtained from vegetable residue for various purposes in the food, cosmetic and pharmaceutical industries is good for the environment and also helps to reduce cost and to increase efficiency. However, in many cases, there is a significant lack of research about the viability of such residue exploitation and, as a result, its use is still under investigation (Babbar, Oberoi, Uppal, & Patil, 2011). The objective of this study was to perform a characterization of the different classes of low molecular weight phenolic compounds present in winemaking by-products (skin from grape marc and lees) obtained during the preparation of red wine from grapes Cabernet Sauvignon and Cabernet Franc, both Vitis vinifera, generated by a winery in the Food Research International 62 (2014) 500–513 ⁎ Corresponding author at: Instituto Regional de Investigación Científica Aplicada, Universidad de Castilla-La Mancha, Campus Universitario s/n, 13071 Ciudad Real, Spain. E-mail address: [email protected] (I. Hermosín-Gutiérrez). http://dx.doi.org/10.1016/j.foodres.2014.03.051 0963-9969/© 2014 Elsevier Ltd. All rights reserved. Contents lists available at ScienceDirect Food Research International journal homepage: www.elsevier.com/locate/foodres
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Occurrence of low molecular weight phenolics in Vitis vinifera red grape cultivars and their...

Food Research International 62 (2014) 500–513
Contents lists available at ScienceDirect
Food Research International
j ourna l homepage: www.e lsev ie r .com/ locate / foodres
Occurrence of low molecular weight phenolics in Vitis vinifera red grapecultivars and their winemaking by-products from São Paulo (Brazil)
Milene Teixeira Barcia a, Paula Becker Pertuzatti a, Daniele Rodrigues a, Sergio Gómez-Alonso b,c,Isidro Hermosín-Gutiérrez c,d,⁎, Helena Teixeira Godoy a
a Faculdade de Engenharia de Alimentos, Universidade Estadual de Campinas, Cidade Universitária “Zeferino Vaz”, s/n, 13083862, Campinas, São Paulo, Brazilb Fundación Parque Científico y Tecnológico de Albacete, Paseo de la Innovación, 1, 02006 Albacete, Spainc Instituto Regional de Investigación Científica Aplicada, Universidad de Castilla-La Mancha, Campus Universitario s/n, 13071 Ciudad Real, Spaind Escuela de Ingenieros Agrónomos, Universidad de Castilla-La Mancha, Ronda de Calatrava, 7, 13071 Ciudad Real, Spain
⁎ Corresponding author at: Instituto Regional de InvUniversidad de Castilla-La Mancha, Campus Universitario
E-mail address: [email protected] (I. Hermosín
http://dx.doi.org/10.1016/j.foodres.2014.03.0510963-9969/© 2014 Elsevier Ltd. All rights reserved.
a b s t r a c t
a r t i c l e i n f oArticle history:Received 15 January 2014Accepted 29 March 2014Available online 5 April 2014
Keywords:Vitis viniferaGrape marcLeesAnthocyaninsFlavonolsHydroxycinnamic acidsStilbenesDrying process
The content of lowmolecularweight phenolic compounds present inwinemaking by-products (skins fromgrapemarc and wine lees) of red wine made from the Vitis vinifera grape cultivars Cabernet Sauvignon and CabernetFranc in the state of São Paulo (Brazil) has been examined. The study was extended to two consecutive yearsand showed that winemaking by-products still contained high concentrations of interesting functional andbioactive phenolic compounds. In addition to original grape phenolics, the winemaking by-products containednew phenolics originated by the action of enzymatic (yeast mediated) and chemical reactions duringwinemaking, including formation of anthocyanin-derived pigments and hydrolysis products from flavonolglycosides and tartaric esters of hydroxycinnamic acids. A total of 19 anthocyanins, 9 pyranoanthocyanins, 18flavonols, 7 hydroxycinnamic acid derivatives, and 3 resveratrol-based stilbenes were identified. Wine leeshave been revealed as an interesting source of flavonol aglycones and pyranoanthocyanins. The best samplepreparation technique prior to the extraction of phenolic compounds,with regard to themaintenance of phenoliccompound profiles and concentrations, was freeze-drying. However, in the case of lees, spray-drying showed tobe also effective in the preservation of phenolic compounds and is a less expensive technique. Oven-drying at50 °C should be avoided if there is interest in usingwinemaking by-products for further purposes, since phenoliccompound concentrations were strongly reduced by thermal degradation.
© 2014 Elsevier Ltd. All rights reserved.
Introduction
São Paulo is a southeastern Brazilian State where wine productionactivity is currently increasing. The wine industry produces a largeamount of by-products and residues. All these residues containbiodegradable organic matter, the disposal of which creates seriousenvironmental problems (Mendes, Prozil, Evtuguin, & Lopes, 2013;Rebello et al., 2013).
Grapemarc (a set of skins and seeds) is themost abundant residue ofwine making, which is generated after concomitant fermentationandmaceration of the grapes during redwine production. After fermen-tation of the must, a decanting wine process occurs, where thesupernatant is separated from the lees. The lees are composed of fineparticles of grape residue and yeasts (Cortés, Rodríguez, Salgado, &Domínguez, 2011; Maragkoudakis et al., 2013; Paradelo, Moldes, &Barral, 2010).
estigación Científica Aplicada,s/n, 13071 Ciudad Real, Spain.-Gutiérrez).
Over the past few years, the by-products of wine have attractedattention as possible sources of phenolic compounds since these com-pounds in the grape are only partially transferred to thewine. However,the majority of the phenolic compounds still remain in the residue asthey are present in the solid parts of the grape. Thus, the recovery ofphenols from industrial residues is gaining considerable attention,especially because of the antioxidant and biological activities attributedto these compounds (Cheng, Bekhit, McConnell, Mros, & Zhao, 2012;Rubilar, Pinelo, Shene, Sineiro, & Nuñez, 2007; Shrikhande, 2000).
The use of bioactive compounds obtained from vegetable residue forvarious purposes in the food, cosmetic and pharmaceutical industries isgood for the environment and also helps to reduce cost and to increaseefficiency. However, inmany cases, there is a significant lack of researchabout the viability of such residue exploitation and, as a result, its use isstill under investigation (Babbar, Oberoi, Uppal, & Patil, 2011).
The objective of this study was to perform a characterization of thedifferent classes of low molecular weight phenolic compounds presentin winemaking by-products (skin from grape marc and lees) obtainedduring the preparation of red wine from grapes Cabernet Sauvignonand Cabernet Franc, both Vitis vinifera, generated by a winery in the

501M.T. Barcia et al. / Food Research International 62 (2014) 500–513
State of São Paulo (Brazil) in two consecutive years. In addition, theeffects of three different by-product drying techniques (drying-ovenat 50 °C, spray-drying, and freeze-drying) on the content of phenoliccompounds were evaluated.
Material and methods
Chemicals
All solvents were of LC quality and all chemicals were of analyticalgrade (N99%). Water was of Milli-Q quality. The following commercialstandards were used for identification: malvidin 3-glucoside, caffeicacid, p-coumaric acid, trans-caftaric acid, and trans-piceid (Phytolab,Vestenbergsgreuth, Germany); cyanidin 3-glucoside, kaempferol,quercetin, isorhamnetin, myricetin, syringetin, and the 3-glucosides ofkaempferol, quercetin, isorhamnetin, and syringetin (Extrasynthese,Genay, France); trans-resveratrol (Sigma, Madrid, Spain); myricetin 3-glucoside, quercetin 3-glucuronide, and laricitrin 3-glucoside werepreviously isolated from Petit Verdot grape skins (Castillo-Muñoz,Fernández-Gonzalez, Gómez-Alonso, García-Romero, & Hermosín-Gutiérrez, 2009). The trans isomers of resveratrol and its 3-glucoside(piceid) were transformed into their respective cis isomers by UV-irradiation (366 nm light for 5 min in quartz vials) of 25% MeOHsolutions of the trans isomers.
Samples
Samples of grape berries used for elaboration of wine and theirresulting winemaking by-products (skins and lees) of two V. viniferavarieties (Cabernet Sauvignon and Cabernet Franc) were collected intriplicate (3 × 500 g) during the harvest of 2011 and 2012 (with theexception of the skin of Cabernet Franc in the year 2011, which waslost in the industry) from a winery located at the municipal district ofSão Roque (São Paulo, Brazil), which lies at 23° 31′44″ S and 47° 08′06″ W, and 771 m above sea level (referred to datum WGS84, WorldGeodetic System1984) andwhich has a subtropical climate (maximum,23.1 °C; minimum, 15.5 °C). Grape berry samples were frozen at−20 °C for 24h, cut in twohalves, re-frozen at−20 °C for 24h, and fur-ther freeze-dried for 48 h. Samples of fermented grape marc and leesfrom both grape varieties were frozen at −20 °C for 24 h and thenfreeze-dried for 48 h. After that, the skins were manually separatedfrom dried grape marc samples. Dried samples of skins from grapemarc (Cabernet Sauvignon from year 2011) and lees (Cabernet Francand Cabernet Sauvignon, both from year 2011) were also obtained byoven-drying at 50 °C under forced air flow. Finally, a third drying treat-ment was applied to lees of the year 2011 from both grape cultivars;that of spray-drying. After drying, the samples of entire grapes andskins were crushed and homogenized. All dried samples were storedat−18 °C until their analysis.
Extraction of phenolic compounds
Dried samples in amounts of 2.5, 0.25, and 0.25 g (grapes, skins andlees, respectively) were extracted with 25mL of a mixture of methanol,water, and formic acid (50:48.5:1.5 v/v), with the help of an ultrasonicbath for 2 min and were then centrifuged at 5000 g at 5 °C for 5 min.Second (grape and lees) and third (skins) extractions of the resultingpellets yielded nearly 99% of the sample phenolic content, as confirmedby the HPLC analysis of successive extractions (up to five). Thecombined supernatants were stored at −18 °C until use. The sampleextracts (grape: 1 mL, skin and lees: 5 mL) were dried in a rotaryevaporator (35 °C) and re-dissolved in 0.5, 0.5 and 1 mL (grape, skinand lees, respectively) of 0.1 N HCl, filtered (0.20 μm, polyestermembrane, Chromafil PET 20/25, Macherey-Nagel, Düren, Germany)and directly injected into the HPLC system for anthocyanindetermination.
For analysis of non-anthocyanin phenolics, a clean-up step usingECX SPE cartridges (40 μm, 500 mg, 6 mL; Scharlab, Sentmenat,Barcelona, Spain) for removing anthocyanins was applied following apreviously described method (Rebello et al., 2013). The eluates weredried in a rotary evaporator (35 °C) and re-dissolved in 0.5 mL of 20%methanol in water and directly injected into the HPLC equipment foranalyzing flavonols, hydroxycinnamic acid derivatives and stilbenes.
Identification and quantification of phenolic compounds by HPLC–DAD–ESI–MSn
Analysis of anthocyanins and derived compoundsHPLC separation, identification, and quantification of anthocyanins
and pyranoanthocyanins were performed on an Agilent 1100 Seriessystem (Agilent, Germany), equipped with DAD (G1315B) and aLC/MSD Trap VL (G2445C VL) electrospray ionization mass spectrometry(ESI-MSn) system, and coupled to an Agilent ChemStation (versionB.01.03) data-processing station. The mass spectra data was processedwith the Agilent LC/MS Trap software (version 5.3). 10 μL of sampleextracts was injected into a reversed-phase column Zorbax EclipseXDB-C18 (2.1 × 150 mm; 3.5 μm particle; Agilent, Germany),thermostatized at 40 °C (Rebello et al., 2013). The solvents werebased on mixtures of water, acetonitrile, and formic acid (88.5:3:8.5,v/v/v, solvent A; 41.5:50:8.5, v/v/v, solvent B), and the flow rate was0.19 mL/min. The linear gradient for solvent B was as follows: 0 min,6%; 10 min, 30%; 30 min, 50%; 34 min, 100%; 36 min, 100%; 42 min,6%. For identification, ESI-MSn was used setting the following parame-ters: positive ionization mode; dry gas, N2, 11 mL/min; drying temper-ature, 350 °C; nebulizer, 65 psi; capillary,−2500 V; capillary exit offset,70 V; skimmer 1, 20 V; skimmer 2, 6 V; compound stability, 100%; andscan range, 50–1200 m/z. For quantification, DAD-chromatogramswere extracted at 520 nm and their total concentrations wereexpressed as equivalents of malvidin-3-diglucoside (g/kg of dry weightsample).
Analysis of non-anthocyanin phenolic compoundsHPLC separation, identification and quantification of non-anthocyanin
phenolic compounds were performed on the same chromatographicsystem previously described for anthocyanins. In this case, 20 μL ofanthocyanin-free extract fractions was injected into a reversed-phasecolumn Zorbax Eclipse XDB-C18 (2.1 × 150 mm; 3.5 μm particle;Agilent, Germany), thermostatized at 40 °C (Rebello et al., 2013).The solvents were as follows: solvent A (acetonitrile/water/formicacid, 3:88.5:8.5, v/v/v), solvent B (acetonitrile/water/formic acid,50:41.5:8.5, v/v/v), and solvent C (methanol/water/formic acid,90:1.5:8.5, v/v/v). The flow rate was 0.19 mL/min. The linear solventgradient was as follows: 0 min, 98% A, and 2% B; 8 min, 96% A, and 4%B; 37 min, 70% A, 17% B, and 13% C; 51 min, 50% A, 30% B, and 20% C;51.5 min, 30% A, 40% B, and 30% C; 56 min, 50% B, and 50% C; 57 min,50% B, and 50% C; 64 min, 98% A, and 2% B. For quantification, DADchromatograms were extracted at 360 nm (flavonols) and 320 nm(hydroxycinnamic acid derivatives and resveratrol forms), and theirconcentrations were expressed as in mg per kg of dried sampleequivalents of quercetin 3-glucoside (flavonols), trans-caftaric acid(hydroxycinnamic acid), and trans-resveratrol (resveratrol forms).
StatisticsThe ANOVA Tukey test (STATISTIC 7.0, p b 0.05) was used for
mean value comparison. In addition, Principal Component Analysis(Pirouette 3.11.) was applied to the data matrix in order to better visu-alize similarities and/or differences within samples in a determined setof data. Data was previously pre-escalated before submitting them tostatistics analysis.
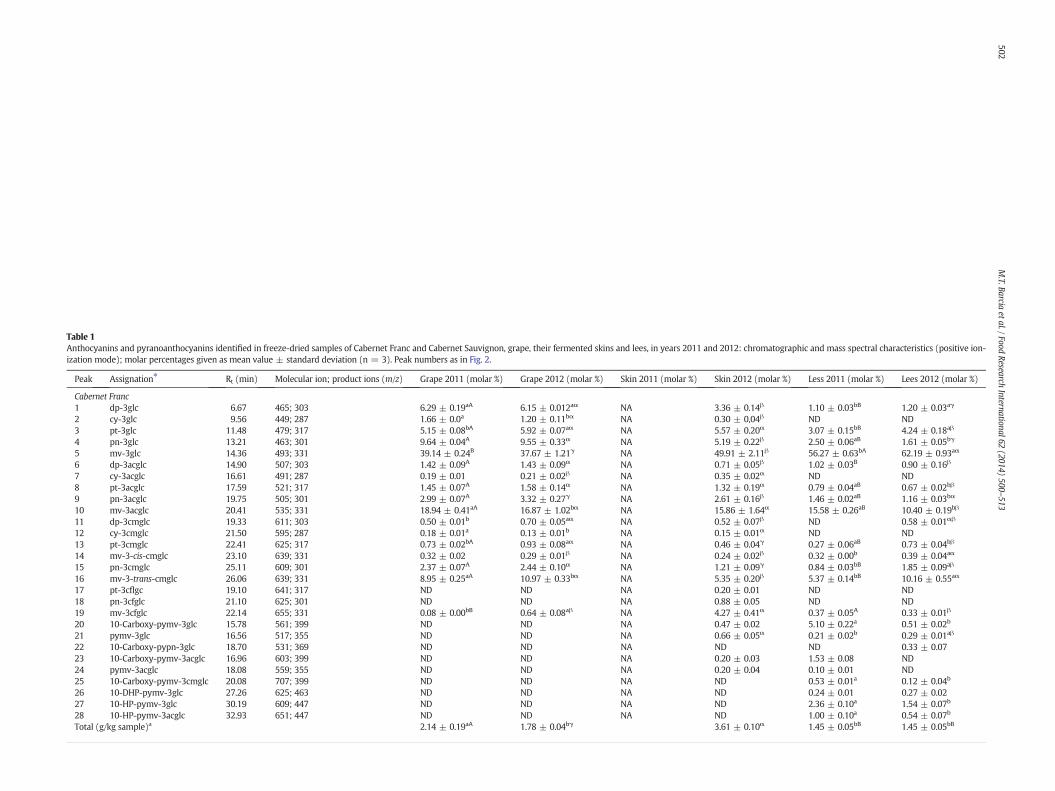
Table 1Anthocyanins and pyranoanthocyanins identified in freeze-dried samples of Cabernet Franc and Cabernet Sauvignon, grape, their fermented skins and lees, in years 2011 and 2012: chromatographic and mass spectral characteristics (positive ion-ization mode); molar percentages given as mean value ± standard deviation (n = 3). Peak numbers as in Fig. 2.
Peak Assignation⁎ Rt (min) Molecular ion; product ions (m/z) Grape 2011 (molar %) Grape 2012 (molar %) Skin 2011 (molar %) Skin 2012 (molar %) Less 2011 (molar %) Lees 2012 (molar %)
Cabernet Franc1 dp-3glc 6.67 465; 303 6.29 ± 0.19aA 6.15 ± 0.012aα NA 3.36 ± 0.14β 1.10 ± 0.03bB 1.20 ± 0.03aγ
2 cy-3glc 9.56 449; 287 1.66 ± 0.0a 1.20 ± 0.11bα NA 0.30 ± 0,04β ND ND3 pt-3glc 11.48 479; 317 5.15 ± 0.08bA 5.92 ± 0.07aα NA 5.57 ± 0.20α 3.07 ± 0.15bB 4.24 ± 0.18aβ
4 pn-3glc 13.21 463; 301 9.64 ± 0.04A 9.55 ± 0.33α NA 5.19 ± 0.22β 2.50 ± 0.06aB 1.61 ± 0.05bγ
5 mv-3glc 14.36 493; 331 39.14 ± 0.24B 37.67 ± 1.21γ NA 49.91 ± 2.11β 56.27 ± 0.63bA 62.19 ± 0.93aα
6 dp-3acglc 14.90 507; 303 1.42 ± 0.09A 1.43 ± 0.09α NA 0.71 ± 0.05β 1.02 ± 0.03B 0.90 ± 0.16β
7 cy-3acglc 16.61 491; 287 0.19 ± 0.01 0.21 ± 0.02β NA 0.35 ± 0.02α ND ND8 pt-3acglc 17.59 521; 317 1.45 ± 0.07A 1.58 ± 0.14α NA 1.32 ± 0.19α 0.79 ± 0.04aB 0.67 ± 0.02bβ
9 pn-3acglc 19.75 505; 301 2.99 ± 0.07A 3.32 ± 0.27γ NA 2.61 ± 0.16β 1.46 ± 0.02aB 1.16 ± 0.03bα
10 mv-3acglc 20.41 535; 331 18.94 ± 0.41aA 16.87 ± 1.02bα NA 15.86 ± 1.64α 15.58 ± 0.26aB 10.40 ± 0.19bβ
11 dp-3cmglc 19.33 611; 303 0.50 ± 0.01b 0.70 ± 0.05aα NA 0.52 ± 0.07β ND 0.58 ± 0.01αβ
12 cy-3cmglc 21.50 595; 287 0.18 ± 0.01a 0.13 ± 0.01b NA 0.15 ± 0.01α ND ND13 pt-3cmglc 22.41 625; 317 0.73 ± 0.02bA 0.93 ± 0.08aα NA 0.46 ± 0.04γ 0.27 ± 0.06aB 0.73 ± 0.04bβ
14 mv-3-cis-cmglc 23.10 639; 331 0.32 ± 0.02 0.29 ± 0.01β NA 0.24 ± 0.02β 0.32 ± 0.00b 0.39 ± 0.04aα
15 pn-3cmglc 25.11 609; 301 2.37 ± 0.07A 2.44 ± 0.10α NA 1.21 ± 0.09γ 0.84 ± 0.03bB 1.85 ± 0.09aβ
16 mv-3-trans-cmglc 26.06 639; 331 8.95 ± 0.25aA 10.97 ± 0.33bα NA 5.35 ± 0.20β 5.37 ± 0.14bB 10.16 ± 0.55aα
17 pt-3cflgc 19.10 641; 317 ND ND NA 0.20 ± 0.01 ND ND18 pn-3cfglc 21.10 625; 301 ND ND NA 0.88 ± 0.05 ND ND19 mv-3cfglc 22.14 655; 331 0.08 ± 0.00bB 0.64 ± 0.08aβ NA 4.27 ± 0.41α 0.37 ± 0.05A 0.33 ± 0.01β
20 10-Carboxy-pymv-3glc 15.78 561; 399 ND ND NA 0.47 ± 0.02 5.10 ± 0.22a 0.51 ± 0.02b
21 pymv-3glc 16.56 517; 355 ND ND NA 0.66 ± 0.05α 0.21 ± 0.02b 0.29 ± 0.01aβ
22 10-Carboxy-pypn-3glc 18.70 531; 369 ND ND NA ND ND 0.33 ± 0.0723 10-Carboxy-pymv-3acglc 16.96 603; 399 ND ND NA 0.20 ± 0.03 1.53 ± 0.08 ND24 pymv-3acglc 18.08 559; 355 ND ND NA 0.20 ± 0.04 0.10 ± 0.01 ND25 10-Carboxy-pymv-3cmglc 20.08 707; 399 ND ND NA ND 0.53 ± 0.01a 0.12 ± 0.04b
26 10-DHP-pymv-3glc 27.26 625; 463 ND ND NA ND 0.24 ± 0.01 0.27 ± 0.0227 10-HP-pymv-3glc 30.19 609; 447 ND ND NA ND 2.36 ± 0.10a 1.54 ± 0.07b
28 10-HP-pymv-3acglc 32.93 651; 447 ND ND NA ND 1.00 ± 0.10a 0.54 ± 0.07b
Total (g/kg sample)a 2.14 ± 0.19aA 1.78 ± 0.04bγ 3.61 ± 0.10α 1.45 ± 0.05bB 1.45 ± 0.05bB
502M.T.Barcia
etal./FoodResearch
International62(2014)
500–513

Peak
Assignation⁎ Rt (min) Molecular ion; product ions (m/z) Grape 2011 (molar %) Grape 2012 (molar %) Skin 2011 (molar %) Skin 2012 (molar %) Less 2011 (molar %) Lees 2012 (molar %)
Cabernet Sauvignon1 dp-3glc 6.67 465; 303 6.11 ± 0.26aA 2.40 ± 0.32bα 2.92 ± 0.29B 3.22 ± 0.68α 1.56 ± 0.16aC 1.08 ± 0.05bβ
2 cy-3glc 9.56 449; 287 2.05 ± 0.12aA 0.41 ± 0.12bα 0.56 ± 0.12aB 0.35 ± 0.03bα 0.15 ± 0.02C 0.13 ± 0.01β
3 pt-3glc 11.48 479; 317 4.06 ± 0.17A 3.91 ± 0.56α 3.16 ± 0.33B 3.85 ± 0.76α 3.28 ± 0.37aB 2.28 ± 0.26bβ
4 pn-3glc 13.21 463; 301 9.79 ± 0.39aA 5.26 ± 0.17bα 4.06 ± 0.12aB 2.37 ± 0.22bβ 1.86 ± 0.20aC 0.69 ± 0.01bγ
5 mv-3glc 14.36 493; 331 39.22 ± 1.74bC 53.86 ± 0.24aγ 57.19 ± 0.53bB 62.54 ± 2.52aβ 63.54 ± 3.33A 66.23 ± 0.13α
6 dp-3acglc 14.90 507; 303 1.26 ± 0.11aA 0.63 ± 0.06bβ 0.63 ± 0.08B 0.46 ± 0.12β 0.64 ± 0.07bB 1.22 ± 0.08aα
7 cy-3acglc 16.61 491; 287 0.24 ± 0.05b 0.44 ± 0.06a ND ND ND ND8 pt-3acglc 17.59 521; 317 1.16 ± 0.12aA 0.76 ± 0.05bα 0.69 ± 0.08aB 0.42 ± 0.13bβ 0.87 ± 0.11aB 0.54 ± 0.02bβ
9 pn-3acglc 19.75 505; 301 2.93 ± 0.18aA 1.29 ± 0.07bα 1.08 ± 0.07aB 0.50 ± 0.15bβ 0.87 ± 0.12bB 1.39 ± 0.11aα
10 mv-3acglc 20.41 535; 331 22.87 ± 1.05A 24.02 ± 0.28α 16.52 ± 0.75aB 9.72 ± 0.10bγ 13.06 ± 1.02aC 11.01 ± 0.04bβ
11 dp-3cmglc 19.33 611; 303 0.24 ± 0.02 0.36 ± 0.12 0.18 ± 0.04b 0.29 ± 0.01a 0.25 ± 0.03b 0.49 ± 0.09a
12 cy-3cmglc 21.50 595; 287 0.13 ± 0.01 ND 0.17 ± 0.05 ND ND 0.07 ± 0.0113 pt-3cmglc 22.41 625; 317 0.40 ± 0.02aA 0.24 ± 0.03bβ 0.18 ± 0.04bB 0.33 ± 0.03aα 0.36 ± 0.01A 0.37 ± 0.03α
14 mv-3-cis-cmglc 23.10 639; 331 0.34 ± 0.03aA 0.16 ± 0.01bβ 0.25 ± 0.03B 0.25 ± 0.02β 0.34 ± 0.04bA 0.55 ± 0.07aα
15 pn-3cmglc 25.11 609; 301 1.83 ± 0.14aA 0.55 ± 0.05bγ 0.70 ± 0.06B 0.81 ± 0.08β 0.77 ± 0.13bB 1.08 ± 0.03aα
16 mv-3-trans-cmglc 26.06 639; 331 7.37 ± 0.14aA 4.90 ± 0.57bβ 5.57 ± 0.58aB 4.45 ± 0.26bβ 5.73 ± 0.50bB 9.09 ± 0.06aα
17 pt-3cflgc 19.10 641; 317 ND ND ND ND ND ND18 pn-3cfglc 21.10 625; 301 ND ND ND ND ND ND19 mv-3cfglc 22.14 655; 331 ND 0.77 ± 0.07β 1.39 ± 0.11bA 1.96 ± 0.04aα 0.34 ± 0.03B 0.39 ± 0.05γ
20 10-Carboxy-pymv-3glc 15.78 561; 399 ND ND 2.73 ± 0.12A 2.57 ± 0.62α 1.54 ± 0.16aB 0.45 ± 0.02bβ
21 pymv-3glc 16.56 517; 355 ND ND 0.73 ± 0.06bA 3.64 ± 0.25aα 0.27 ± 0.02B 0.27 ± 0.02β
22 10-Carboxy-pypn-3glc 18.70 531; 369 ND ND ND ND 0.10 ± 0.02b 0.81 ± 0.15a
23 10-Carboxy-pymv-3acglc 16.96 603; 399 ND ND 0.93 ± 0.18A 1.00 ± 0.13α 0.44 ± 0.05aB 0.14 ± 0.00bβ
24 pymv-3acglc 18.08 559; 355 ND ND 0.32 ± 0.03bA 1.09 ± 0.09a 0.10 ± 0.02B ND25 10-Carboxy-pymv-3cmglc 20.08 707; 399 ND ND 0.18 ± 0.03B 0.20 ± 0.01 0.24 ± 0.02A ND26 10-DHP-pymv-3glc 27.26 625; 463 ND ND ND ND 0.40 ± 0.05a 0.20 ± 0.01b
27 10-HP-pymv-3glc 30.19 609; 447 ND ND ND ND 2.46 ± 0.28a 1.23 ± 0.15b
28 10-HP-pymv-3acglc 32.93 651; 447 ND ND ND ND 0.84 ± 0.06a 0.30 ± 0.03b
Total (g/kg sample)a 1.78 ± 0.20aB 1.11 ± 0.20bβ 2.59 ± 0.36aA 0.58 ± 0.04bγ 1.47 ± 0.06bB 2.22 ± 0.02aα
⁎ Nomenclature abbreviations: dp, delphinidin; cy, cyanidin; pt, petunidin; pn, peonidin; mv, malvidin; glc, glucoside; acglc, 6″-acetyl-glucoside; cfglc, 6″-caffeoyl-glucoside; cmglc, 6″-p-coumaroyl-glucoside; pymv, pyranomalvidin; pypn,pyranopeonidin; 10-DHP, 10-(3″,4″-hydroxy)-phenyl; 10-HP, 10-(4″-hydroxy)-phenyl. 10-Carboxy-pymv-3glc and pymv-3-glc are also known as vitisins A and B, respectively.
a As malvidin 3-glucoside equivalents (mv-3glc). NA, not analyzed, ND, not detected. Different lower case letters for the samples of different vintages (2011 and 2012) of each kind of sample (grape, skin, or lees) mean significant differences(ANOVA, p b 0.05). Different capital or Greek letters for the three samples (grape, fermented skin, and lees) of each vintage (years 2011 or 2012, respectively) mean significant differences (ANOVA, p b 0.05).
503M.T.Barcia
etal./FoodResearch
International62(2014)
500–513

504 M.T. Barcia et al. / Food Research International 62 (2014) 500–513
Results and discussion
Anthocyanins and derived compounds in grapes and their winemakingby-products
A total of 19 anthocyanins and 9 pyranoanthocyanins weretentatively identified (Table 1), although not all of them were alwaysdetected in all samples. The identification of each anthocyanin wasmainly based on the comparison of spectroscopic data, especiallythose of MS/MS spectra, with those obtained from available standardsor previously reported (Lago-Vanzela, Da-Silva, Gomes, García-Romero,& Hermosín-Gutiérrez, 2011a; Nixdorf & Hermosín-Gutiérrez, 2010).
The expected structures of the anthocyanidins delphinidin (dp),cyanidin (cy), petunidin (pt), peonidin (pn) and malvidin (mv) wereidentified on the basis of their respective product ions at m/z 303, 287,317, 301 and 331, generated after fragmentation (MS/MS spectra) ofsuspected anthocyanins. In contrast, pelargonidin-based anthocyaninswere not found in the samples, as noted by the absence of any signalattributable to their product ions atm/z 271.
The profile of anthocyanins (molar percentage for each individualanthocyanin; Table 1; Fig. 2; Fig. SM-1 of Supplementary Materials)for both grape cultivars contained the complete series of non-acylatedand acylated 3-glucosides, which were accounted for in the followingdecreasing order: 3-glucosides (3-glc), 3-(6″-acetyl)-glucosides(3-acglc), and 3-(6″-p-coumaroyl)-glucosides (3-cmglc). Moreover,some 3-(6″-caffeoyl)-glucosides (3-cfglc) were also found as minorcompounds, usually only those derived from malvidin. The presenceof only 3-glucoside derivatives confirmed the nature of V. viniferagrape cultivars (Nixdorf & Hermosín-Gutiérrez, 2010). The assignmentof the 3-glucoside moiety was based on the fragmentation patterns ob-served in the spectra, because the entire glucoside restwas released andonly one product ion corresponding to the remaining anthocyanidinwas observed, regardless of whether glucose was acylated or not(Lago-Vanzela, Da-Silva, Gomes, García-Romero, &Hermosín-Gutiérrez,2011b; Nixdorf & Hermosín-Gutiérrez, 2010).
The major individual anthocyanins in grapes were, on average, mv-3-glc (49–57%), mv-3-acglc (around 16%), and mv-3-trans-cmglc(6–8%), whereas the caffeoyl derivative (mv-3-cfglc) was found inconcentrations below 2%. Most of the anthocyanins found in thegrape samples were also detected in both winemaking by-products(fermented skins and lees), with the exception of some minorcompounds (mainly cyanidin derivatives). Another peculiarity wasthat the fermented skin of Cabernet Franc (from the year 2012) showedhigh levels of mv-3-cfglc (4.27%), and other very minor caffeoyl deriva-tives that could be also detected were pt-3-cfglc (0.20%) and pn-3-cfglc(0.88%), which were not detected in any of the other samples of grape,fermented skin and lees. Finally, theminor cy-3-acglcwas detected onlyin both grape samples from the two years (average, 0.20–0.34%) andalso in the fermented skin of Cabernet Franc (0.35%) from the year 2012.
The anthocyanin profiles found in our studywere in accordancewithliterature reported data for Cabernet Sauvignon and similar grapecultivars like CabernetMitos, Lemberger, Spatburgunder, Schwarzriesling,and Trollinger (Ferrandino, Carra, Rolle, Schneider, & Schubert, 2012;Kammerer, Claus, Carle, & Schieber, 2004; Kammerer, Claus, Schieber, &Reinhold, 2005; Ryan & Revilla, 2003). It iswidely known that the profileof the anthocyanins of a grape cultivar is closely linked to their geneticinheritance, although environmental factors may have some influenceon this profile (Poudel, Mochioka, Beppu, & Kataoka, 2009). However,the differences in anthocyanin profiles between samples of grape andwinemaking by-products may be due to differences in solid/liquid parti-tion coefficients of individual compounds and their solubility in thewine that modulate the transfer of the anthocyanin from the grape tothe wine.
During winemaking, pyranoanthocyanins are formed fromanthocyanidin 3-glucosides, and can be easily identified by their charac-teristic UV–vis and MSn spectra (Nixdorf & Hermosín-Gutiérrez, 2010;
Blanco-Vega, López-Bellido, Alía-Robledo, & Hermosín-Gutiérrez, 2011).In red wine made from V. vinifera grape cultivars, vitisin-typepyranoanthocyanins are formed in the initial steps of fermentation,most-ly derived from the reaction of pyruvic acid and acetaldehydewith antho-cyanins, which are two yeast intermediate metabolites (Blanco-Vegaet al., 2011). These compounds can be partially retained by grape marcafter wine separation, and the presence of pyranoanthocyanins inwinemaking by-products of the grape Sicilian cultivar Nerello Mascalesehas been reported (Amico et al., 2004). The following vitisin-type pyrano-anthocyanins were found in the samples under study (Table 1; Figs. 1and 2): vitisin A (10-carboxy-pyranomalvidin-3-glucoside) and itsacetyl and p-coumaroyl derivatives, the A-type vitisin derived frompn-3-glc (10-carboxy-pyranopeonidin-3-glucoside), and vitisin B(pyranomalvidin-3-glucoside) and its acetyl derivative. The A-typevitisin from pn-3-glc was only found in lees samples. All the aforemen-tioned vitisin-type pyranoanthocyanins accounted for low molarpercentages and reflected themolar percentages found for their respec-tive anthocyanin precursors. However, in the case of Cabernet Francthey accounted for higher proportions in fermented skins whereas inthe case of Cabernet Sauvignon they did in the lees.
Hydroxyphenyl-pyranoanthocyanins are another type of pyrano-anthocyanins, which result from the reaction of anthocyanidin 3-glucosides with hydroxycinnamic acids or its decarboxylation products(Blanco-Vega et al., 2011). Maybe because of their low polarity, theywere only found in lees samples and the detected structures wereidentified as: 10-(4″-hydroxyphenyl)-pyranomalvidin-3-glucoside(10-HP-pymv-3-glc) and its acetyl derivative (10-HP-pymc-3-acglc),derived from p-coumaric acid, and 10-(3″,4″-dihydroxyphenyl)-pyranomalvidin-3-glcucoside (10-DHP-pymv-3-glc), derived fromcaffeic acid. According to Morata, Gómez-Cordovés, Colomo, and Suárez(2005) these compoundsmay be formed during thewinemaking pro-cess and they may be absorbed by the lees. These hydroxyphenyl-pyranoanthocyanins were formed in lower amounts than vitisin-typepyranoanthocyanins overall, but their presence only in lees meansthat they usually account for higher amounts in this type ofwinemakingby-product, with the only exception being the lees of Cabernet Francfrom 2011.
The total anthocyanin content in Cabernet Sauvignon and CabernetFranc grapes varied statistically between the studied years (1.78 and2.14 g/kg dry sample, for the year 2011; 1.11 and 1.78 g/kg dry sample,for the year 2012), but they were similar when compared to each other.However, the Cabernet Franc grape was statistically different (p b 0.05)from the Cabernet Sauvignon grape only in the year 2012, as was theanthocyanin content in fermented skins of Cabernet Franc which wasgreater than that of Cabernet Sauvignon. The lees of Cabernet Sauvignonand Cabernet Franc showed no significant difference (p b 0.05) in thetotal anthocyanin concentration (g/kg dry sample) in their analysis forone specific year, but they were different for years 2011 and 2012.Multiple factors such as climate, the degree of maturity, the berry'ssize, the grape variety and the applied technology for winemakingmay be the cause of variations in the profile and content of anthocya-nins in different years (Kammerer et al., 2005). Koundouras et al.(2009) investigated the importance of irrigation and root stocks onthe concentration of phenolic compounds in Cabernet Sauvignongrapes. According to that experiment, limited water could cause asubstantial increase in the concentration of anthocyanins in grapeskins andmalvidin 3-glucosidewas seen to be affected bywater supply.
The transfer of anthocyanins from the grapes towine is very limited,usually below40% (Boulton, 2001). Thus,most of the anthocyanins fromgrape remain in the winemaking by-products, and even those trans-ferred to the wine can partially be adsorbed by the lees (Morata et al.,2005). The fermented skin separated from the grape marcs retainedlarge quantities of anthocyanins, despite the grapes initially showingless total anthocyanins than the fermented skin due to the sugarspresent in the dry matter; however, the content of anthocyanins waslower in the skins of Cabernet Sauvignon in the year 2012 (0.58 g/kg)
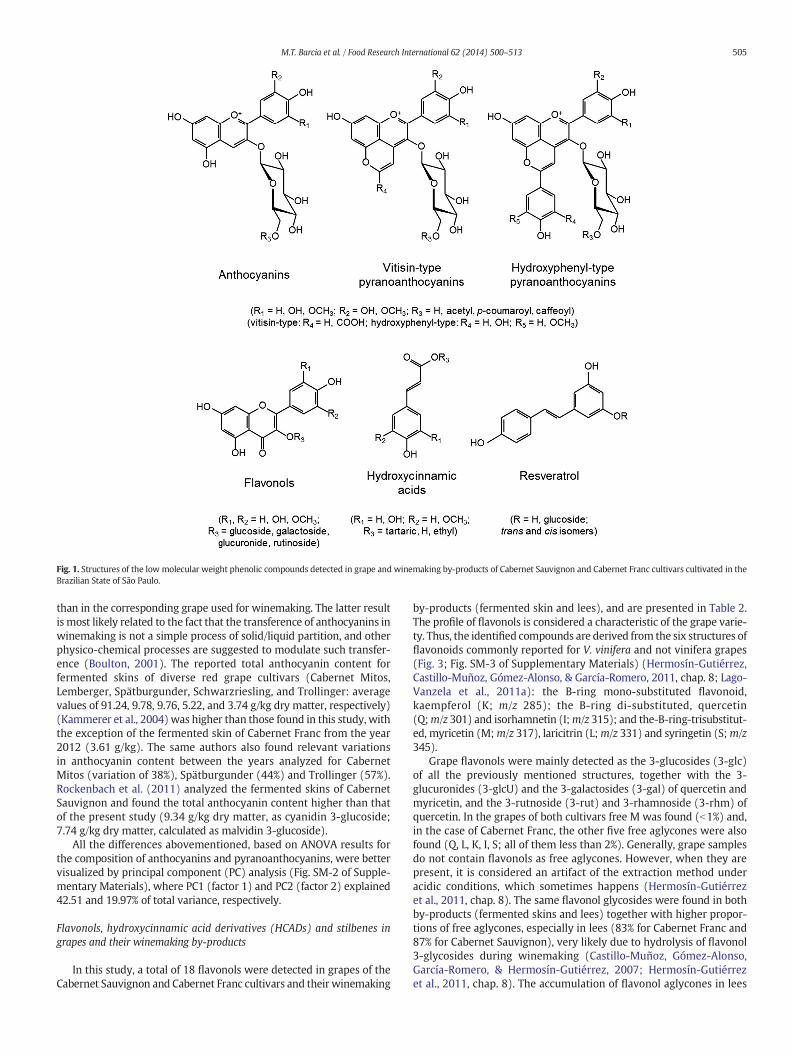
Fig. 1. Structures of the low molecular weight phenolic compounds detected in grape and winemaking by-products of Cabernet Sauvignon and Cabernet Franc cultivars cultivated in theBrazilian State of São Paulo.
505M.T. Barcia et al. / Food Research International 62 (2014) 500–513
than in the corresponding grape used for winemaking. The latter resultis most likely related to the fact that the transference of anthocyanins inwinemaking is not a simple process of solid/liquid partition, and otherphysico-chemical processes are suggested to modulate such transfer-ence (Boulton, 2001). The reported total anthocyanin content forfermented skins of diverse red grape cultivars (Cabernet Mitos,Lemberger, Spätburgunder, Schwarzriesling, and Trollinger: averagevalues of 91.24, 9.78, 9.76, 5.22, and 3.74 g/kg dry matter, respectively)(Kammerer et al., 2004) was higher than those found in this study, withthe exception of the fermented skin of Cabernet Franc from the year2012 (3.61 g/kg). The same authors also found relevant variationsin anthocyanin content between the years analyzed for CabernetMitos (variation of 38%), Spätburgunder (44%) and Trollinger (57%).Rockenbach et al. (2011) analyzed the fermented skins of CabernetSauvignon and found the total anthocyanin content higher than thatof the present study (9.34 g/kg dry matter, as cyanidin 3-glucoside;7.74 g/kg dry matter, calculated as malvidin 3-glucoside).
All the differences abovementioned, based on ANOVA results forthe composition of anthocyanins and pyranoanthocyanins, were bettervisualized by principal component (PC) analysis (Fig. SM-2 of Supple-mentary Materials), where PC1 (factor 1) and PC2 (factor 2) explained42.51 and 19.97% of total variance, respectively.
Flavonols, hydroxycinnamic acid derivatives (HCADs) and stilbenes ingrapes and their winemaking by-products
In this study, a total of 18 flavonols were detected in grapes of theCabernet Sauvignon and Cabernet Franc cultivars and their winemaking
by-products (fermented skin and lees), and are presented in Table 2.The profile of flavonols is considered a characteristic of the grape varie-ty. Thus, the identified compounds are derived from the six structures offlavonoids commonly reported for V. vinifera and not vinifera grapes(Fig. 3; Fig. SM-3 of Supplementary Materials) (Hermosín-Gutiérrez,Castillo-Muñoz, Gómez-Alonso, & García-Romero, 2011, chap. 8; Lago-Vanzela et al., 2011a): the B-ring mono-substituted flavonoid,kaempferol (K; m/z 285); the B-ring di-substituted, quercetin(Q;m/z 301) and isorhamnetin (I;m/z 315); and the-B-ring-trisubstitut-ed, myricetin (M;m/z 317), laricitrin (L;m/z 331) and syringetin (S;m/z345).
Grape flavonols were mainly detected as the 3-glucosides (3-glc)of all the previously mentioned structures, together with the 3-glucuronides (3-glcU) and the 3-galactosides (3-gal) of quercetin andmyricetin, and the 3-rutnoside (3-rut) and 3-rhamnoside (3-rhm) ofquercetin. In the grapes of both cultivars free M was found (b1%) and,in the case of Cabernet Franc, the other five free aglycones were alsofound (Q, L, K, I, S; all of them less than 2%). Generally, grape samplesdo not contain flavonols as free aglycones. However, when they arepresent, it is considered an artifact of the extraction method underacidic conditions, which sometimes happens (Hermosín-Gutiérrezet al., 2011, chap. 8). The same flavonol glycosides were found in bothby-products (fermented skins and lees) together with higher propor-tions of free aglycones, especially in lees (83% for Cabernet Franc and87% for Cabernet Sauvignon), very likely due to hydrolysis of flavonol3-glycosides during winemaking (Castillo-Muñoz, Gómez-Alonso,García-Romero, & Hermosín-Gutiérrez, 2007; Hermosín-Gutiérrezet al., 2011, chap. 8). The accumulation of flavonol aglycones in lees

Fig. 2.DAD-chromatograms corresponding to the anthocyaninprofiles (detection at 520 nm) of BRS Cabernet Franc samples: A) grapes; (B) fermented skins, (C)wine lees. Identification ofthe peaks referred to Table 1.
506 M.T. Barcia et al. / Food Research International 62 (2014) 500–513
could be explained by considering the low solubility of these com-pounds in water and hydroalcoholic mixtures of low ethanol contentsuch as wine. Q-3-rutinoside was missing in the lees of both grapecultivars while L-3-glucoside was not found in Cabernet Sauvignonlees and Q-3-rhamnoside was absent in Cabernet Franc lees samples.Flavonols based on quercetin were predominant in all samples mainlyas: an average of 51% for free Q in lees; averages of 16% for free Q, 22%for Q-3-glcU and 13% for Q-3-glc in fermented skins; and averages of27% for Q-3-glcU and 34% for Q-3-glc in grapes.
Ferrandino et al. (2012) examined the fresh skins of CabernetSauvignon cultivated in Italy and found 34.3% Q-3-glc, 25.8% M-3-glc,25.0% Q-3-glcU, and 14.5% K-3-glc. Slier et al. (2010) also identifiedM-3-glc and Q-3-glc in fresh skins of Cabernet Sauvignon from Chile,as well as Q-3-gal, K-3-gal, K-3-glc, and I-3-glc. A similar profile was
observed in the fresh skins of Spanish Cabernet Sauvignon (Castillo-Muñoz et al., 2007): 6.0% M-3-glcU, 20.9% M-3-glc, 23.3% Q-3-glcU,26.7% Q-3-glc, 6.5% L-3-glc, 3.8% K-3-glc, 4.5% I-3-glc, and 8.9% S-3-glc.Rockenbach et al. (2011) noted the presence of derivatives of quercetinand kaempferol in the fermented skin of Primitivo, Sangiovese, Pinotnoir, Negro amaro, Cabernet Sauvignon and Isabel cultivars fromBrazilian winery by-products. Spanish winery by-products from Caber-net Sauvignon showed a similar flavonol profile to the latter work,together with undefined glycosides of myricetin and isorhamnetin(Rubilar et al., 2007).
Variations of flavonol composition between harvests can beexplained by several factors (Kammerer et al., 2005; Poudel et al.,2009) as alreadymentioned elsewhere. In our study, themolar percent-ages of flavonols showed variations between the years 2011 and 2012,

507M.T. Barcia et al. / Food Research International 62 (2014) 500–513
with the exception of: Q-3-gal, M-3-gal, S-3-glc, free L, free K, and free Iin samples of Cabernet Sauvignon, andM-3-gal, L-3-glc, freeM, and freeK in samples of Cabernet Franc, which did not show statistical differ-ences (p b 0.05). Regarding the content of total flavonols, there werealso statistical variations between the years analyzed for both samples(grape, fermented skin and lees) of the two cultivars (Table 2). Thesame was observed when comparing grapes and fermented skinsof Cabernet Franc with Cabernet Sauvignon each year, with CabernetFranc showing predominantly higher concentrations. The lees ofthe two cultivars were not statistically different (at p b 0.05) whencompared in the same year. However, we cannot suggest the causes ofvariations. Besides the diverse factors which affect the biosynthesis offlavonols in grapes, their transfer to wine, their degree of hydrolysis,and their absorption by lees, they may be also affected by many otherfactors and may be different depending on the class of phenoliccompounds. The lees of both cultivars were found as important sourcesof flavonols showing higher amounts than the grapes and the fermentedskin: averages of 1274.2 mg/kg (Cabernet Franc) and 1347.1 mg/kg(Cabernet Sauvignon) in the lees, averages of 776.6 mg/kg (CabernetFranc) and 535.5 mg/kg (Cabernet Sauvignon) in the fermented skins,and averages of 281.5 mg/kg (Cabernet Franc) and 140.5 mg/kg(Cabernet Sauvignon) in the grapes.
Hydroxycinnamic acid derivatives (HCADs) were identified in thesamples based on the comparison of spectroscopic data, especiallyMS/MS spectral data, as shown in Table 2. The hydroxycinnamoyl-tartaric acids were detected in all samples. Two HCADs predominated,namely caftaric acid (from caffeic acid) with an average of 53% andcoutaric acid (from p-coumaric acid) with an average of 17%. Lowconcentrations of a third HCAD, the trans isomer of fertaric acid(from ferulic acid) with an average of 5%, were found in addition to itscis isomer that was also detected in Cabernet Franc grapes and inCabernet Sauvignon grape and fermented skin (only for the year2011). Hydroxycinnamoyl-tartaric acids might undergo hydrolysisduring the winemaking process thus releasing free hydroxycinnamicacids which can further react with the ethanol to form ethyl esters.Therefore, caffeic acid and p-coumaric acid were found in the lees ofboth samples. In the fermented skin, the caffeic acid was detected onlyfor Cabernet Sauvignon and ethyl caffeate was present in the lees ofCabernet Franc (year 2012). The HCAD profile in fresh skins of CabernetSauvignonwas previously reported by Ferrandino et al. (2012) showingthe presence of trans-caftaric acid, cis-coutaric acid, trans-coutaric acid,and trans-fertaric acid. In winemaking by-products of Cabernet Mitos,caffeic, p-coumaric, ferulic, caftaric, coutaric, and fertaric acids weredetected (Kammerer et al., 2005). The lees of both cultivars showedstatistically (p b 0.05) higher concentrations of HCADs compared withthe grapes and fermented skin, with average values of 884.8 mg/kg(Cabernet Franc) and 510.4 mg/kg (Cabernet Sauvignon) versus149.7 mg/kg (Cabernet Franc) and 107.0 mg/kg (Cabernet Sauvignon)in the grapes and 63.35 mg/kg (Cabernet Franc) and 44.55 mg/kg(Cabernet Sauvignon) in the fermented skins.
The presence of trans-resveratrol was expected. Several studieshave reported that fresh skins contain about 50–100 μg/g of resveratrol,while concentration in red wine varies between 1.5 and 3.0 mg/L.Several studies on the bioactive properties of resveratrol and itsderivatives have recently demonstrated its potential as an antimutagenic,antioxidant, anti-inflammatory and anti-proliferative agent (Rockenbachet al., 2011). The detection of stilbenes is difficult because they eluted inthe same area where flavonols and residual anthocyanins are eluted,making it difficult to separate and identify the appropriate stilbenesof interest. The latter may be one reason for the observed lack ofcis-resveratrol in grapes, fermented skin and lees of Cabernet Franc(Table 2). In the grapes and lees of Cabernet Sauvignon, trans-resveratrol was detected in addition to trans-piceid and cis-piceid(Table 2). It is suggested that some of the resveratrol may have beentransferred to the wine and some further absorbed by the lees, thusthe absence of resveratrol in the fermented skins could be explained.
Quantitatively, the grape of Cabernet Franc of year 2011 showed astilbene content of 29.88 mg/kg, whereas that of Cabernet Sauvignonshowed contents of 63.87 and 6.21 mg/kg for the years 2011 and2012 respectively. The lees of Cabernet Sauvignon (year 2011) showedno significant difference (p b 0.05) compared with the grape from thesame year. In contrast, in the year 2012, the lees exhibited a concentra-tion greater than grape (184.41 vs. 6.21 mg/kg).
All the differences abovementioned, based on ANOVA results for thecomposition of flavonols, HCAD and stilbenes, were better visualized byprincipal component (PC) analysis (Fig. SM-4 of SupplementaryMaterials), where PC1 (factor 1) and PC2 (factor 2) explained 54.62and 14.46% of total variance respectively.
Effect of drying treatment on the content of different low molecular weightphenolic compounds in Cabernet Sauvignon and Cabernet Francwinemaking by-products
The freeze-drying (FD) techniquewas used to achieveminimal ther-mal degradation of phenolic compounds present in analyzed samples.The confirmation of this hypothesis was based on the monitoring ofthe total content of anthocyanins (Table 1). The fermented skins ofCabernet Sauvignon (only from the year 2011) dried at 50 °C (D50)only contained 0.94 g/kg of total anthocyanins, thus representing aloss of around 47% when compared with the freeze-drying (FD)technique. This loss of anthocyanins was accompanied by changes inthe profile of anthocyanins (Fig. 5A), where there was an increasein the molar percentage of pyranoanthocyanins and a decrease inthe molar proportions of other anthocyanins, except for mv-3-glc, mv-3-acglc, dp-3-acglc, pn-3-acglc, and mv-3-cis cmglc which remainedstatistically unchanged (p b 0.05)when dried at 50 °C (D50). This resultsuggests that pyranoanthocyanins should be more resistant towardsthermal degradation than their corresponding anthocyanins.
The D50 lees (only from the year 2011) contained 0.49 g/kg(Cabernet Sauvignon) and 0.41 g/kg (Cabernet Franc) of total anthocy-anins, with significant losses of 67 and 72% respectively with regard toFD samples. In contrast, the spray-drying treatment (SD) did not affectthe total anthocyanin content of the lees compared with FD. The dryingtreatments evaluated (D50 and SD) caused significant differences in theprofiles of anthocyanins and derived compounds (Fig. 4C and D). Someof these pigments maintained their respective molar percentagesalmost unchanged (mv-3-glc, mv-3-cis cmglc, pn-3-cmglc, and pycy-3-glc for the lees of Cabernet Sauvignon; pn-3-cmglc for the lees ofCabernet Franc). In the lees subjected to D50, a significant increase inthe content of pyranoanthocyanins was observed, and also noticeablewas the absence of some minor anthocyanins in the lees from D50and SD treatments, which were detected in the lees treated by FD.
The D50 and SD techniques caused a negative effect in the totalcontent of flavonols, hydroxycinnamic acid derivatives (HCAD), andstilbenes, which was greater than that observed for the total anthocya-nin content (Fig. 4B). In the fermented skins of Cabernet Sauvignon, thetotal contents of flavonols (343.4 mg/kg) and HCADs (18.18 mg/kg) inD50 samples represented 57 and 72% of reduction, respectively,when compared with FD (Table 2). The molar percentages of flavonolsQ-3-rut, Q-3-rha, free I, and free S were not affected by D50 treatment.Regarding HCAD, a total loss of cis-fertaric acid was observed, togetherwith 10-fold increase in the molar percentage of trans-fertaric acid.The other HCADs were apparently not affected qualitatively.
The FD and SD lees showed similar flavonol profiles (Fig. 4D and F).However, the SD lees of Cabernet Sauvignon (total flavonols,1111.1 mg/kg) and Cabernet Franc (total flavonols, 712.9 mg/kg)showed losses of 25 and 42% in their respective total content of flavo-nols. The decreases in the concentration of flavonols in samplesof D50 lees were 78% for Cabernet Sauvignon (total flavonols,324.2 mg/kg) and 90% for Cabernet Franc (total flavonols, 121.6 mg/kg),mainly due to losses of free quercetin and free myricetin. The behaviorof HCADs was similar, with losses ranging between 90 and 91% in the
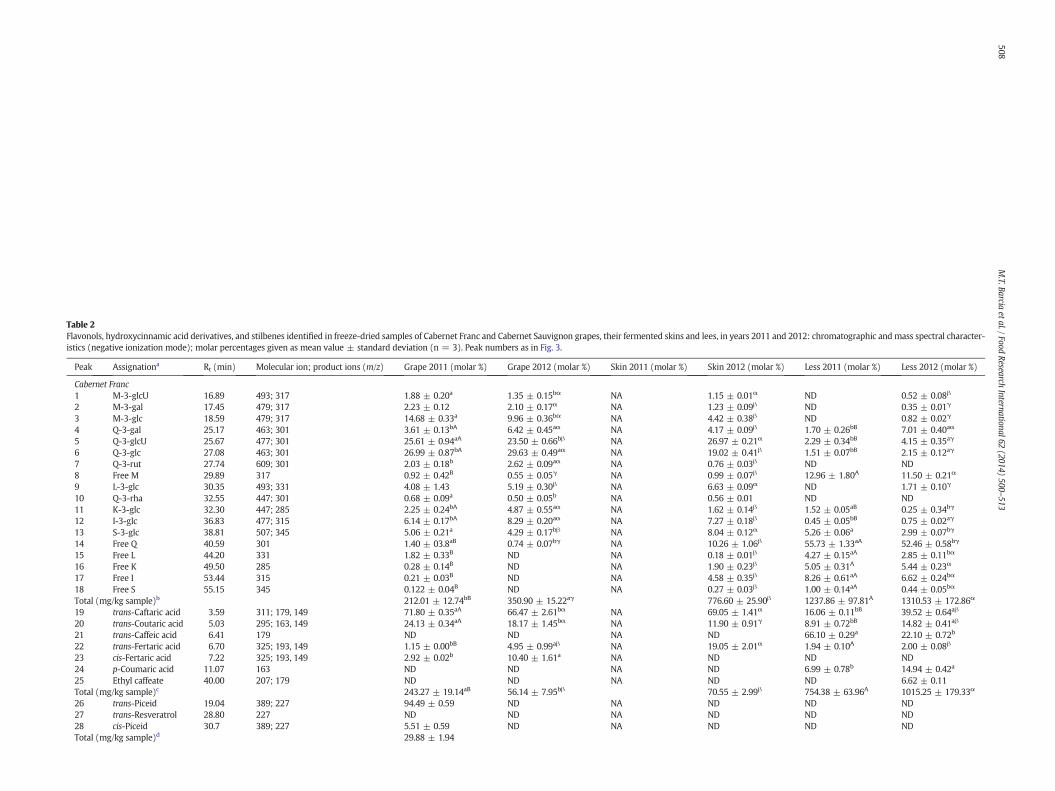
Table 2Flavonols, hydroxycinnamic acid derivatives, and stilbenes identified in freeze-dried samples of Cabernet Franc and Cabernet Sauvignon grapes, their fermented skins and lees, in years 2011 and 2012: chromatographic and mass spectral character-istics (negative ionization mode); molar percentages given as mean value ± standard deviation (n = 3). Peak numbers as in Fig. 3.
Peak Assignationa Rt (min) Molecular ion; product ions (m/z) Grape 2011 (molar %) Grape 2012 (molar %) Skin 2011 (molar %) Skin 2012 (molar %) Less 2011 (molar %) Less 2012 (molar %)
Cabernet Franc1 M-3-glcU 16.89 493; 317 1.88 ± 0.20a 1.35 ± 0.15bα NA 1.15 ± 0.01α ND 0.52 ± 0.08β
2 M-3-gal 17.45 479; 317 2.23 ± 0.12 2.10 ± 0.17α NA 1.23 ± 0.09β ND 0.35 ± 0.01γ
3 M-3-glc 18.59 479; 317 14.68 ± 0.33a 9.96 ± 0.36bα NA 4.42 ± 0.38β ND 0.82 ± 0.02γ
4 Q-3-gal 25.17 463; 301 3.61 ± 0.13bA 6.42 ± 0.45aα NA 4.17 ± 0.09β 1.70 ± 0.26bB 7.01 ± 0.40aα
5 Q-3-glcU 25.67 477; 301 25.61 ± 0.94aA 23.50 ± 0.66bβ NA 26.97 ± 0.21α 2.29 ± 0.34bB 4.15 ± 0.35aγ
6 Q-3-glc 27.08 463; 301 26.99 ± 0.87bA 29.63 ± 0.49aα NA 19.02 ± 0.41β 1.51 ± 0.07bB 2.15 ± 0.12aγ
7 Q-3-rut 27.74 609; 301 2.03 ± 0.18b 2.62 ± 0.09aα NA 0.76 ± 0.03β ND ND8 Free M 29.89 317 0.92 ± 0.42B 0.55 ± 0.05γ NA 0.99 ± 0.07β 12.96 ± 1.80A 11.50 ± 0.21α
9 L-3-glc 30.35 493; 331 4.08 ± 1.43 5.19 ± 0.30β NA 6.63 ± 0.09α ND 1.71 ± 0.10γ
10 Q-3-rha 32.55 447; 301 0.68 ± 0.09a 0.50 ± 0.05b NA 0.56 ± 0.01 ND ND11 K-3-glc 32.30 447; 285 2.25 ± 0.24bA 4.87 ± 0.55aα NA 1.62 ± 0.14β 1.52 ± 0.05aB 0.25 ± 0.34bγ
12 I-3-glc 36.83 477; 315 6.14 ± 0.17bA 8.29 ± 0.20aα NA 7.27 ± 0.18β 0.45 ± 0.05bB 0.75 ± 0.02aγ
13 S-3-glc 38.81 507; 345 5.06 ± 0.21a 4.29 ± 0.17bβ NA 8.04 ± 0.12α 5.26 ± 0.06a 2.99 ± 0.07bγ
14 Free Q 40.59 301 1.40 ± 03.8aB 0.74 ± 0.07bγ NA 10.26 ± 1.06β 55.73 ± 1.33aA 52.46 ± 0.58bγ
15 Free L 44.20 331 1.82 ± 0.33B ND NA 0.18 ± 0.01β 4.27 ± 0.15aA 2.85 ± 0.11bα
16 Free K 49.50 285 0.28 ± 0.14B ND NA 1.90 ± 0.23β 5.05 ± 0.31A 5.44 ± 0.23α
17 Free I 53.44 315 0.21 ± 0.03B ND NA 4.58 ± 0.35β 8.26 ± 0.61aA 6.62 ± 0.24bα
18 Free S 55.15 345 0.122 ± 0.04B ND NA 0.27 ± 0.03β 1.00 ± 0.14aA 0.44 ± 0.05bα
Total (mg/kg sample)b 212.01 ± 12.74bB 350.90 ± 15.22aγ 776.60 ± 25.90β 1237.86 ± 97.81A 1310.53 ± 172.86α
19 trans-Caftaric acid 3.59 311; 179, 149 71.80 ± 0.35aA 66.47 ± 2.61bα NA 69.05 ± 1.41α 16.06 ± 0.11bB 39.52 ± 0.64aβ
20 trans-Coutaric acid 5.03 295; 163, 149 24.13 ± 0.34aA 18.17 ± 1.45bα NA 11.90 ± 0.91γ 8.91 ± 0.72bB 14.82 ± 0.41aβ
21 trans-Caffeic acid 6.41 179 ND ND NA ND 66.10 ± 0.29a 22.10 ± 0.72b
22 trans-Fertaric acid 6.70 325; 193, 149 1.15 ± 0.00bB 4.95 ± 0.99aβ NA 19.05 ± 2.01α 1.94 ± 0.10A 2.00 ± 0.08β
23 cis-Fertaric acid 7.22 325; 193, 149 2.92 ± 0.02b 10.40 ± 1.61a NA ND ND ND24 p-Coumaric acid 11.07 163 ND ND NA ND 6.99 ± 0.78b 14.94 ± 0.42a
25 Ethyl caffeate 40.00 207; 179 ND ND NA ND ND 6.62 ± 0.11Total (mg/kg sample)c 243.27 ± 19.14aB 56.14 ± 7.95bβ 70.55 ± 2.99β 754.38 ± 63.96A 1015.25 ± 179.33α
26 trans-Piceid 19.04 389; 227 94.49 ± 0.59 ND NA ND ND ND27 trans-Resveratrol 28.80 227 ND ND NA ND ND ND28 cis-Piceid 30.7 389; 227 5.51 ± 0.59 ND NA ND ND NDTotal (mg/kg sample)d 29.88 ± 1.94
508M.T.Barcia
etal./FoodResearch
International62(2014)
500–513
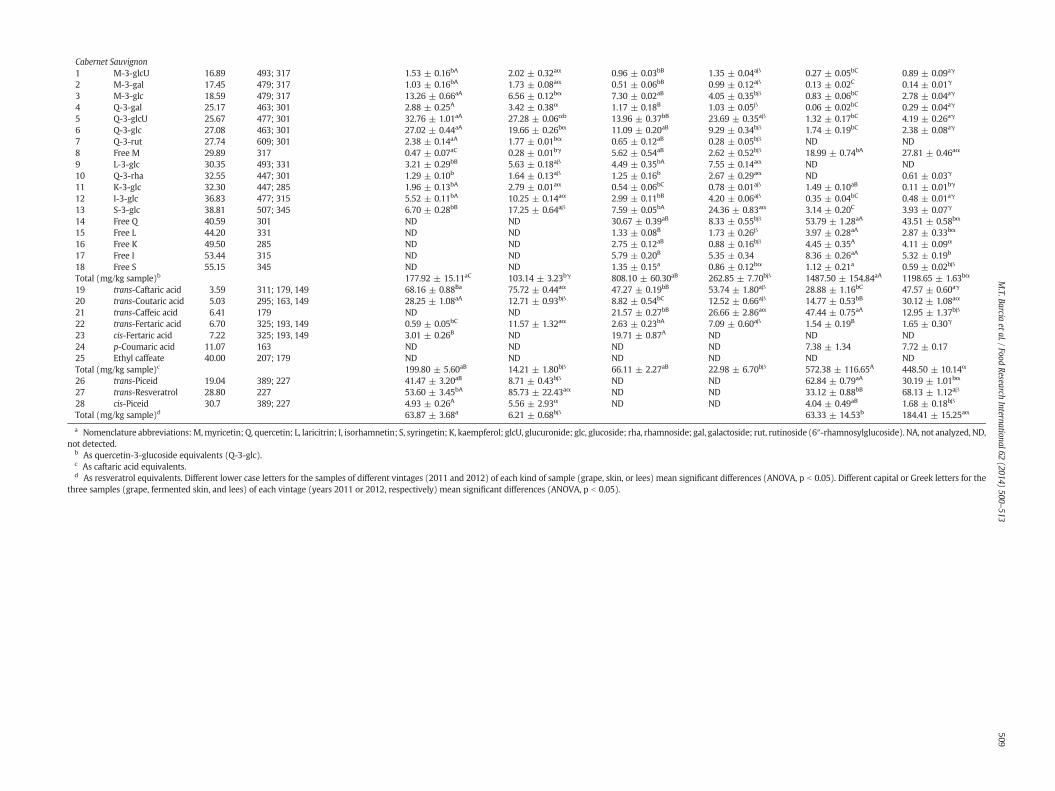
Peak
Assignationa Rt (min) Molecular ion; product ions (m/z) Grape 2011 (molar %) Grape 2012 (molar %) Skin 2011 (molar %) Skin 2012 (molar %) Less 2011 (molar %) Less 2012 (molar %)
Cabernet Sauvignon1 M-3-glcU 16.89 493; 317 1.53 ± 0.16bA 2.02 ± 0.32aα 0.96 ± 0.03bB 1.35 ± 0.04aβ 0.27 ± 0.05bC 0.89 ± 0.09aγ
2 M-3-gal 17.45 479; 317 1.03 ± 0.16bA 1.73 ± 0.08aα 0.51 ± 0.06bB 0.99 ± 0.12aβ 0.13 ± 0.02C 0.14 ± 0.01γ
3 M-3-glc 18.59 479; 317 13.26 ± 0.66aA 6.56 ± 0.12bα 7.30 ± 0.02aB 4.05 ± 0.35bβ 0.83 ± 0.06bC 2.78 ± 0.04aγ
4 Q-3-gal 25.17 463; 301 2.88 ± 0.25A 3.42 ± 0.38α 1.17 ± 0.18B 1.03 ± 0.05β 0.06 ± 0.02bC 0.29 ± 0.04aγ
5 Q-3-glcU 25.67 477; 301 32.76 ± 1.01aA 27.28 ± 0.06αb 13.96 ± 0.37bB 23.69 ± 0.35aβ 1.32 ± 0.17bC 4.19 ± 0.26aγ
6 Q-3-glc 27.08 463; 301 27.02 ± 0.44aA 19.66 ± 0.26bα 11.09 ± 0.20aB 9.29 ± 0.34bβ 1.74 ± 0.19bC 2.38 ± 0.08aγ
7 Q-3-rut 27.74 609; 301 2.38 ± 0.14aA 1.77 ± 0.01bα 0.65 ± 0.12aB 0.28 ± 0.05bβ ND ND8 Free M 29.89 317 0.47 ± 0.07aC 0.28 ± 0.01bγ 5.62 ± 0.54aB 2.62 ± 0.52bβ 18.99 ± 0.74bA 27.81 ± 0.46aα
9 L-3-glc 30.35 493; 331 3.21 ± 0.29bB 5.63 ± 0.18aβ 4.49 ± 0.35bA 7.55 ± 0.14aα ND ND10 Q-3-rha 32.55 447; 301 1.29 ± 0.10b 1.64 ± 0.13aβ 1.25 ± 0.16b 2.67 ± 0.29aα ND 0.61 ± 0.03γ
11 K-3-glc 32.30 447; 285 1.96 ± 0.13bA 2.79 ± 0.01aα 0.54 ± 0.06bC 0.78 ± 0.01aβ 1.49 ± 0.10aB 0.11 ± 0.01bγ
12 I-3-glc 36.83 477; 315 5.52 ± 0.11bA 10.25 ± 0.14aα 2.99 ± 0.11bB 4.20 ± 0.06aβ 0.35 ± 0.04bC 0.48 ± 0.01aγ
13 S-3-glc 38.81 507; 345 6.70 ± 0.28bB 17.25 ± 0.64aβ 7.59 ± 0.05bA 24.36 ± 0.83aα 3.14 ± 0.20C 3.93 ± 0.07γ
14 Free Q 40.59 301 ND ND 30.67 ± 0.39aB 8.33 ± 0.55bβ 53.79 ± 1.28aA 43.51 ± 0.58bα
15 Free L 44.20 331 ND ND 1.33 ± 0.08B 1.73 ± 0.26β 3.97 ± 0.28aA 2.87 ± 0.33bα
16 Free K 49.50 285 ND ND 2.75 ± 0.12aB 0.88 ± 0.16bβ 4.45 ± 0.35A 4.11 ± 0.09α
17 Free I 53.44 315 ND ND 5.79 ± 0.20B 5.35 ± 0.34 8.36 ± 0.26aA 5.32 ± 0.19b
18 Free S 55.15 345 ND ND 1.35 ± 0.15a 0.86 ± 0.12bα 1.12 ± 0.21a 0.59 ± 0.02bβ
Total (mg/kg sample)b 177.92 ± 15.11aC 103.14 ± 3.23bγ 808.10 ± 60.30aB 262.85 ± 7.70bβ 1487.50 ± 154.84aA 1198.65 ± 1.63bα
19 trans-Caftaric acid 3.59 311; 179, 149 68.16 ± 0.88Ba 75.72 ± 0.44aα 47.27 ± 0.19bB 53.74 ± 1.80aβ 28.88 ± 1.16bC 47.57 ± 0.60aγ
20 trans-Coutaric acid 5.03 295; 163, 149 28.25 ± 1.08aA 12.71 ± 0.93bβ 8.82 ± 0.54bC 12.52 ± 0.66aβ 14.77 ± 0.53bB 30.12 ± 1.08aα
21 trans-Caffeic acid 6.41 179 ND ND 21.57 ± 0.27bB 26.66 ± 2.86aα 47.44 ± 0.75aA 12.95 ± 1.37bβ
22 trans-Fertaric acid 6.70 325; 193, 149 0.59 ± 0.05bC 11.57 ± 1.32aα 2.63 ± 0.23bA 7.09 ± 0.60aβ 1.54 ± 0.19B 1.65 ± 0.30γ
23 cis-Fertaric acid 7.22 325; 193, 149 3.01 ± 0.26B ND 19.71 ± 0.87A ND ND ND24 p-Coumaric acid 11.07 163 ND ND ND ND 7.38 ± 1.34 7.72 ± 0.1725 Ethyl caffeate 40.00 207; 179 ND ND ND ND ND NDTotal (mg/kg sample)c 199.80 ± 5.60aB 14.21 ± 1.80bβ 66.11 ± 2.27aB 22.98 ± 6.70bβ 572.38 ± 116.65A 448.50 ± 10.14α
26 trans-Piceid 19.04 389; 227 41.47 ± 3.20aB 8.71 ± 0.43bβ ND ND 62.84 ± 0.79aA 30.19 ± 1.01bα
27 trans-Resveratrol 28.80 227 53.60 ± 3.45bA 85.73 ± 22.43aα ND ND 33.12 ± 0.88bB 68.13 ± 1.12aβ
28 cis-Piceid 30.7 389; 227 4.93 ± 0.26A 5.56 ± 2.93α ND ND 4.04 ± 0.49aB 1.68 ± 0.18bβ
Total (mg/kg sample)d 63.87 ± 3.68a 6.21 ± 0.68bβ 63.33 ± 14.53b 184.41 ± 15.25aα
a Nomenclature abbreviations:M,myricetin; Q, quercetin; L, laricitrin; I, isorhamnetin; S, syringetin; K, kaempferol; glcU, glucuronide; glc, glucoside; rha, rhamnoside; gal, galactoside; rut, rutinoside (6″-rhamnosylglucoside). NA, not analyzed, ND,not detected.
b As quercetin-3-glucoside equivalents (Q-3-glc).c As caftaric acid equivalents.d As resveratrol equivalents. Different lower case letters for the samples of different vintages (2011 and 2012) of each kind of sample (grape, skin, or lees) mean significant differences (ANOVA, p b 0.05). Different capital or Greek letters for the
three samples (grape, fermented skin, and lees) of each vintage (years 2011 or 2012, respectively) mean significant differences (ANOVA, p b 0.05).
509M.T.Barcia
etal./FoodResearch
International62(2014)
500–513

Fig. 3. DAD-chromatograms corresponding to the profiles of flavonols (detection at 360 nm), hydroxycinnamic acid derivatives and stilbenes (figure enlargements with detection at 320nm) of Cabernet Franc samples: A) grapes; (B) fermented skins, (C) wine lees). Identification of the peaks referred to Table 2.
Fig. 4.Molar profiles (%) of low molecular weight phenolic compounds identified in winemaking by-products from year 2011 subjected to different drying processes (FD, freeze-drying;D50, oven-drying at 50 °C; SP, spray-drying): A) Anthocyanin and pyranoanthocyanin profiles of Cabernet Sauvignon fermented skins dried by D50 and FD; B) Flavonol andhydroxycinnamic acid derivative (HCAD) profiles of Cabernet Sauvignon fermented skins dried by D50 and FD; C) Anthocyanin and pyranoanthocyanin profiles of Cabernet Sauvignonwine lees dried by SP, D50, and FD; D) Flavonol, stilbene, and HCAD profiles of Cabernet Sauvignon wine lees dried by SP, D50, and FD; E) Anthocyanin and pyranoanthocyanin profilesof Cabernet Franc wine lees dried by SP, D50, and FD; F) Flavonol and HCAD profiles of Cabernet Franc wine lees dried by SP, D50, and FD. Abbreviations like in Tables 1 and 2.
510 M.T. Barcia et al. / Food Research International 62 (2014) 500–513

512 M.T. Barcia et al. / Food Research International 62 (2014) 500–513
total content of HCADs. For the D50 lees, the final concentrations were54.37 and 77.80 mg/kg for Cabernet Sauvignon and Cabernet Francrespectively. In the case of SD lees, the final concentrations were 260.5and 183.6 mg/kg for the Cabernet Sauvignon and Cabernet Francrespectively. This is due to the significant loss of trans-caffeic in thelees treated by D50 and losses of all other types of HCADs in the leestreated by SD. It is noticeable that for Cabernet Sauvignon lees therewas a total loss of stilbenes.
Thus, the freeze-drying (FD) technique seems to be the mostsuitable for preserving the phenolic compounds and, subsequently, toprocess winemaking by-product samples. However, the spray-drying(SD) technique, a less expensive alternative to freeze-drying, appearsto be the most adequate for preservation of the anthocyanins. On thebasis of the results previously discussed, oven-drying at 50 °C underforced air flow (D50), even though it is a cheap process, is not a recom-mended process for drying of winemaking by-products.
Conclusions
The winemaking by-products of V. vinifera grape cultivars CabernetFranc and Cabernet Sauvignon contained high and significant concen-trations of lowmolecularweight phenolic compounds, having function-al and bioactive properties. Thus, the winemaking by-products couldbecome promissory sources of valuable functional and bioactivephenolic compounds. In addition to original grape phenolics, thesewinemaking by-products accounted for newly developed phenoliccompounds by the action of enzymatic (yeast mediated) and chemicalreactions occurring during winemaking, including formation ofanthocyanin-derived pigments (pyranoanthocyanins) and hydrolysisof flavonol glycosides and tartaric esters of hydroxycinnamicacids. In total, 19 anthocyanins (including the complete series ofnon-acylated, acetylated and p-coumaroylated derivatives of fivecommon anthocyanidins), 9 pyranoanthocyanins (6 vitisin type and 3hydroxyphenyl-type), 18 flavonols (12 glycosides and 6 aglycones),7 hydroxycinnamic acid derivatives (4 tartaric esters, 2 free hydro-xycinnamic acids, and 1 ethyl ester), and 3 resveratrol-based stilbeneswere identified. It is noticeable that wine lees have been found to be aninteresting source of released free flavonol aglycones and pyrano-anthocyanins. However, due to variations between sample years, furtherstudies focused on the winemaking process and the vintage effect areneeded in order to understand the parameters controlling the phenoliccomposition of winemaking by-products and, perhaps, to find a way tomake standardized by-products for further good use. In addition to therichness of phenolic compounds found in winemaking by-products, thesample conservation technique used prior to the extraction of phenoliccompounds played an important role in the preservation of these com-pounds and hence in their recovery yield. Recovery yields were higherusing freeze-drying, although spray-drying appeared to be the mostadequate technique in the case of lees because it combined low costand similar degree of preservation of the anthocyanins. In contrast,oven-drying was not recommended at all because thermal degradationof phenolic compounds strongly decreased their recovery yields.
Acknowledgments
Authors thank the Coordenação de Aperfeiçoamento de Pessoal deNível Superior (CAPES). Author Sergio Gómez Alonso thanks theFondo Social Europeo and the Junta de Comunidades de Castilla-LaMancha for co-funding his contract through the INCRECYT program.This work was partially financed by Junta de Comunidades de Castilla-La Mancha (POII10-0061-4432) and Spanish Ministerio de Economía yCompetitividad (AGL2011-29708-C02-02). Authors also thank Msc.Fernando José de Góes, the owner of Góes winery (São Roque, Brazil)for supplying the samples.
Appendix A. Supplementary data
Supplementary data to this article can be found online at http://dx.doi.org/10.1016/j.foodres.2014.03.051.
References
Amico, V., Napoli, E.M., Renda, A., Ruberto, G., Spatafora, C., & Tringali, C. (2004). Constituentsof grape pomace from the Sicilian cultivar ‘Nerello Mascalese’. Food Chemistry, 88,599–607.
Babbar, N., Oberoi, H. S., Uppal, D. S., & Patil, R. T. (2011). Total phenolic content andantioxidant capacity of extracts obtained from six important fruit residues. FoodResearch International, 44, 391–396.
Blanco-Vega, D., López-Bellido, F. J., Alía-Robledo, J. M., & Hermosín-Gutiérrez, I. (2011).HPLC–DAD–ESI–MS/MS characterization of pyranoanthocyanins pigments formedin model wine. Journal of Agricultural and Food Chemistry, 59, 9523–9531.
Boulton, R. (2001). The copigmentation of anthocyanins and its role in the color of redwine: A critical review. American Journal of Enology and Viticulture, 52, 67–87.
Castillo-Muñoz, N., Fernández-Gonzalez, M., Gómez-Alonso, S., García-Romero, E., &Hermosín-Gutiérrez, I. (2009). Red-color related phenolic composition of GarnachaTintorera (Vitis vinifera L.) grapes and red wines. Journal of Agricultural and FoodChemistry, 57, 7883–7891.
Castillo-Muñoz, N., Gómez-Alonso, S., García-Romero, E., & Hermosín-Gutiérrez, I. (2007).Flavonol profiles of Vitis vinifera red grapes and their single-cultivar wines. Journal ofAgricultural and Food Chemistry, 55, 992–1002.
Cheng, V. J., Bekhit, A. E. A., McConnell, M., Mros, S., & Zhao, J. (2012). Effect of extractionsolvent, waste fraction and grape variety on the antimicrobial and antioxidantactivities of extracts from wine residue from cool climate. Food Chemistry, 134,474–482.
Cortés, S., Rodríguez, R., Salgado, J. M., & Domínguez, J. M. (2011). Comparative studybetween Italian and Spanish grapemarc spirits in terms of major volatile compounds.Food Control, 22, 673–680.
Ferrandino, A., Carra, A., Rolle, L., Schneider, A., & Schubert, A. (2012). Profiling ofhydroxycinnamoyl tartrates and acylated anthocyanins in the skin of 34 Vitis viniferagenotypes. Journal of Agricultural and Food Chemistry, 60, 4931–4945.
Hermosín-Gutiérrez, I., Castillo-Muñoz, N., Gómez-Alonso, S., & García-Romero, E. (2011Chapter 8). Flavonol profiles for grape and wine authentication. In S. E. Ebeler, G.R. Takeoka, & P. Winterhalter (Eds.), Progress in authentication of food and wine(pp. 113–129). Washington, DC: American Chemical Society, ACS Symposium Series.
Kammerer, D., Claus, A., Carle, R., & Schieber, A. (2004). Polyphenol screening of pomacefrom red and white grape varieties (Vitis vinifera L.) by HPLC–DAD–MS/MS. Journal ofAgricultural and Food Chemistry, 52, 4360–4367.
Kammerer, D., Claus, A., Schieber, A., & Reinhold, C. (2005). A novel process for therecovery of polyphenols from grape (Vitis vinifera L.) pomace. Journal of FoodScience, 70(2), 157–163.
Koundouras, S., Hatzidimitriou, E., Karamolegkou, M., Dimopoulou, E., Kallithraka, S.,Tsialtas, J. T., et al. (2009). Irrigation and root stockeffects on thephenolic concentrationand aroma potential of Vitis vinifera L. cv. Cabernet Sauvignon grapes. Journal ofAgricultural and Food Chemistry, 57, 7805–7813.
Lago-Vanzela, E. S., Da-Silva, R., Gomes, E., García-Romero, E., & Hermosín-Gutiérrez, I.(2011a). Phenolic composition of the edible parts (flesh and skin) of Bordô grape(Vitis labrusca) using HPLC–DAD–ESI–MS/MS. Journal of Agricultural and FoodChemistry, 59, 13136–13146.
Lago-Vanzela, E. S., Da-Silva, R., Gomes, E., García-Romero, E., & Hermosín-Gutiérrez, I.(2011b). Phenolic composition of the Brazilian seedless table grape varieties BRSClara and BRS Morena. Journal of Agricultural and Food Chemistry, 59, 8314–8323.
Maragkoudakis, P. A., Nardi, T., Bovo, B., D'Andrea, M., Howell, K. S., Giacomini, A., et al.(2013). Biodiversity, dynamics and ecology of bacterial community duringgrape marc storage for the production of grappa. International Journal of FoodMicrobiology, 162(2), 143–151.
Mendes, J. A. S., Prozil, S. O., Evtuguin, D.V., & Lopes, L. P. C. (2013). Towards comprehensiveutilization of winemaking residues: Characterization of grape skins from red grapepomaces of variety Touriga Nacional. Industrial Crops and Products, 43, 25–32.
Morata, A., Gómez-Cordovés, M. C., Colomo, B., & Suárez, J. A. (2005). CellWall anthocyaninadsorption by different Saccharomyces strains during the fermentation ofVitis vinifera L.cv Graciano grapes. European Food Research and Technology, 220, 341–346.
Nixdorf, S. L., & Hermosín-Gutiérrez, I. (2010). Brazilian red wines made from the hybridgrape cultivar Isabel: Phenolic composition and antioxidant capacity. AnalyticaChimica Acta, 659, 208–215.
Paradelo, R., Moldes, A.B.., & Barral, M. T. (2010). Utilization of a factorial design to studythe composting of hydrolyzed grape marc and vinification lees. Journal of Agriculturaland Food Chemistry, 58, 3085–3092.
Poudel, P. R., Mochioka, R., Beppu, K., & Kataoka, I. (2009). Influence of temperature onberry composition of interspecific hybrid wine grape ‘Kadainou R-1’ (Vitis ficifoliavar. ganebu × V. vinifera ‘Muscat of Alexandria’). Journal of the Japanese Society forHorticultural Science, 78, 169–174.
Rebello, L. P. G., Lago-Vanzela, E. S., Barcia, M. T., Ramos, A.M., Stringheta, P. C., Da-Silva, R.,et al. (2013). Phenolic composition of the berry parts of hybrid grape cultivar BRSVioleta (BRS Rubea × IAC 1398-21) using HPLC–DAD–ESI–MS/MS. Food ResearchInternational, 54, 354–366.
Rockenbach, I. I., Gonzaga, L. V., Rizelio, V. M., Gonçalves, A. E. S. S., Genovese, M. I., & Fett,R. (2011). Phenolic compounds and antioxidant activity of seed and skin extracts ofred grape (Vitis vinifera and Vitis labrusca) pomace from Brazilian winemaking.Food Research International, 44, 897–901.

513M.T. Barcia et al. / Food Research International 62 (2014) 500–513
Rubilar, M., Pinelo, M., Shene, C., Sineiro, J., & Nuñez, M. J. (2007). Separation and HPLC-MS identification of phenolic antioxidants from agricultural residues: Almond hullsand grape pomace. Journal of Agricultural and Food Chemistry, 55, 10101–10109.
Ryan, J., & Revilla, E. (2003). Anthocyanin composition of Cabernet Sauvignon andTempranillo grapes at different stages of ripening. Journal of Agricultural and FoodChemistry, 51, 3372–3378.
Shrikhande, A. J. (2000). Wine by-products with health benefits. Food ResearchInternational, 33, 469–474.
Slier, E. O., Neira, A. P., Solís, R. L., Marín, F. Z., Da-Silva, J. M. R., & Laureano, O. (2010).Comparative study of the phenolic composition of seeds and skins from Carménèreand Cabernet Sauvignon grape varieties (Vitis vinifera L.) during ripening. Journal ofAgricultural and Food Chemistry, 58, 3591–3599.




















