
High-value products from transgenic maize
14
Research review paper High-value products from transgenic maize Shaista Naqvi a , Koreen Ramessar a , Gemma Farré a , Maite Sabalza a , Bruna Miralpeix a , Richard M. Twyman b , Teresa Capell a , Changfu Zhu a , Paul Christou a,c, ⁎ a Departament de Producció Vegetal i Ciència Forestal, Universitat de Lleida, Av. Alcalde Rovira Roure, 191, Lleida, 25198, Spain b Department of Biological Sciences, University of Warwick, Coventry, CV4 7AL, United Kingdom c Institucio Catalana de Recerca I Estudis Avancats, Passeig Lluís Companys, 23, 08010 Barcelona, Spain abstract article info Article history: Received 3 May 2010 Received in revised form 17 August 2010 Accepted 17 August 2010 Available online 15 September 2010 Keywords: Maize Corn Transgenic Molecular pharming Metabolic engineering Recombinant protein Processing Regulation Biosafety Maize (also known as corn) is a domesticated cereal grain that has been grown as food and animal feed for tens of thousands of years. It is currently the most widely grown crop in the world, and is used not only for food/feed but also to produce ethanol, industrial starches and oils. Maize is now at the beginning of a new agricultural revolution, where the grains are used as factories to synthesize high-value molecules. In this article we look at the diversity of high-value products from maize, recent technological advances in the field and the emerging regulatory framework that governs how transgenic maize plants and their products are grown, used and traded. © 2010 Elsevier Inc. All rights reserved. Contents 1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 41 2. Food versus industrial products — maize on the front line . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 41 3. Gene transfer in maize — from single genes to multiple traits and pathways . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 41 3.1. Maize transformation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 41 3.2. Multiple gene transfer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 43 3.3. Strategies used to synthesize value added products in maize . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 43 4. Value added products. Specific targets — primary metabolism . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 44 4.1. Carbohydrates . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 44 4.2. Essential amino acids and nutritional proteins . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 44 4.3. Fatty acids . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 45 5. Value added products. Specific targets — vitamins and minerals . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 45 5.1. Micronutrient levels in maize . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 45 5.2. Conventional and transgenic strategies to enhance carotenoid levels . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 46 5.3. Vitamins B9, C and E . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 46 6. Value added products. Specific targets — recombinant proteins . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 46 6.1. Pharmaceutical proteins . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 46 6.2. Industrial proteins . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 48 7. Regulatory, economic and political aspects of high-value products from maize . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 48 7.1. Economics . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 48 7.2. Regulatory challenges . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 49 Biotechnology Advances 29 (2011) 40–53 ⁎ Corresponding author. Departament de Producció Vegetal i Ciència Forestal, Universitat de Lleida, Avenue Alcalde Rovira Roure 191, 25198 Lleida, Spain. Tel.: + 34 973702693; fax: + 34 973238264. E-mail address: [email protected] (P. Christou). 0734-9750/$ – see front matter © 2010 Elsevier Inc. All rights reserved. doi:10.1016/j.biotechadv.2010.08.009 Contents lists available at ScienceDirect Biotechnology Advances journal homepage: www.elsevier.com/locate/biotechadv
Transcript of High-value products from transgenic maize

Biotechnology Advances 29 (2011) 40–53
Contents lists available at ScienceDirect
Biotechnology Advances
j ourna l homepage: www.e lsev ie r.com/ locate /b iotechadv
Research review paper
High-value products from transgenic maize
Shaista Naqvi a, Koreen Ramessar a, Gemma Farré a, Maite Sabalza a, Bruna Miralpeix a, Richard M. Twyman b,Teresa Capell a, Changfu Zhu a, Paul Christou a,c,⁎a Departament de Producció Vegetal i Ciència Forestal, Universitat de Lleida, Av. Alcalde Rovira Roure, 191, Lleida, 25198, Spainb Department of Biological Sciences, University of Warwick, Coventry, CV4 7AL, United Kingdomc Institucio Catalana de Recerca I Estudis Avancats, Passeig Lluís Companys, 23, 08010 Barcelona, Spain
⁎ Corresponding author. Departament de Producció Vfax: +34 973238264.
E-mail address: [email protected] (P. Christou).
0734-9750/$ – see front matter © 2010 Elsevier Inc. Aldoi:10.1016/j.biotechadv.2010.08.009
a b s t r a c t
a r t i c l e i n f oArticle history:Received 3 May 2010Received in revised form 17 August 2010Accepted 17 August 2010Available online 15 September 2010
Keywords:MaizeCornTransgenicMolecular pharmingMetabolic engineeringRecombinant proteinProcessingRegulationBiosafety
Maize (also known as corn) is a domesticated cereal grain that has been grown as food and animal feed fortens of thousands of years. It is currently the most widely grown crop in the world, and is used not only forfood/feed but also to produce ethanol, industrial starches and oils. Maize is now at the beginning of a newagricultural revolution, where the grains are used as factories to synthesize high-value molecules. In thisarticle we look at the diversity of high-value products from maize, recent technological advances in the fieldand the emerging regulatory framework that governs how transgenic maize plants and their products aregrown, used and traded.
egetal i Ciència Forestal, Universitat de Lleida, Avenue Alc
l rights reserved.
© 2010 Elsevier Inc. All rights reserved.
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 412. Food versus industrial products — maize on the front line . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 413. Gene transfer in maize — from single genes to multiple traits and pathways . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 41
3.1. Maize transformation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 413.2. Multiple gene transfer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 433.3. Strategies used to synthesize value added products in maize . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 43
4. Value added products. Specific targets — primary metabolism . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 444.1. Carbohydrates . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 444.2. Essential amino acids and nutritional proteins . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 444.3. Fatty acids. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 45
5. Value added products. Specific targets — vitamins and minerals . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 455.1. Micronutrient levels in maize . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 455.2. Conventional and transgenic strategies to enhance carotenoid levels . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 465.3. Vitamins B9, C and E . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 46
6. Value added products. Specific targets — recombinant proteins . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 466.1. Pharmaceutical proteins . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 466.2. Industrial proteins . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 48
7. Regulatory, economic and political aspects of high-value products from maize . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 487.1. Economics . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 487.2. Regulatory challenges . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 49
alde Rovira Roure 191, 25198 Lleida, Spain. Tel.: +34 973702693;

41S. Naqvi et al. / Biotechnology Advances 29 (2011) 40–53
8. Outlook . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 51Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 51References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 51
Table 1World maize production in thousands of metric tons (adapted from USDA-FAS, 2008).
Production 2004/05 2005/06 2006/07 2007/08 2008/09a
Argentina 20,500 15,800 22,500 20,850 18,000Brazil 35,000 41,700 51,000 58,600 55,000Canada 8837 9361 8990 11,650 9900China 130,290 139,365 151,600 152,300 156,000Egypt 5840 5932 6149 6174 6175EU-27 66,471 60,668 53,829 47,519 59,486India 14,180 14,710 15,100 19,310 18,000Indonesia 7200 6500 6700 7500 7750Mexico 22,050 19,500 22,350 22,650 24,000Nigeria 6500 7000 7800 6500 7800Philippines 5050 5884 6231 7250 6500Russian Federation 3500 3200 3600 3950 6500
1. Introduction
Maize or corn1 (Zea mays) is the most widely grown cereal crop inthe world with 792 million tons produced in 2007/2008, of which774 million tons was consumed as food or feed (Tables 1 and 2). Aswell as its food/feed use,maize has a number of industrial applications,including the production of bioethanol and starch (Fig. 1). Morerecently, maize has attracted interest as a production vehicle for high-value products such as recombinant pharmaceutical proteins andspecialty chemicals.
The diverse uses of maize reflect its long history as a domesticatedcrop and its wide gene pool (Gewin, 2003). Interest in maize as aplatform for high-value products reflects a number of uniqueadvantages over other plants (Ramessar et al., 2008a). These includeits GRAS (Generally Regarded As Safe) status, the establishedagricultural infrastructure, its well characterized genetic properties,its amenability to in vitro manipulation and gene transfer, and itsefficient biomass production resulting from the C4 photosyntheticpathway. More specific for the production of high-value recombinantproteins, maize seeds are excellent production vehicles because oftheir large size compared to other cereals, with 82% of the seed madeup of endosperm (Watson, 1987) (Fig. 2). Furthermore, the maturemaize seed is desiccated, lacks active proteases and contains a richmix of molecular chaperones and disulfide isomerases, helping toensure correct protein folding, assembly and enhanced stability.Maize endosperm tissue can perform post-translational modifications(e.g. glycosylation) that occur in animal cells, thus ensuring thatheterologous proteins expressed in seeds retain their biologicalactivity. The stability of recombinant proteins produced in maizeseeds is favored because the maize endosperm is a natural proteinstorage organ, thus proteins in dry maize seed remain stable for morethan 6 years at ambient temperatures, and for months in crackedand flaked maize seeds stored at up to 10 °C.
Maize has few potential drawbacks as a production platform andthose that have been proposed are mainly concerned with biosafetyand regulatory issues affecting maize crops grown in the field. Forexample, maize is a wind-pollinated, outcrossing crop, which raisesissues surrounding gene flow to non-transgenic maize and wildrelatives. This is discussed later in the section dealing with regulatorychallenges. The advantages and potential drawbacks of maize asa production platform for high-value products are summarized inTable 3.
2. Food versus industrial products — maize on the front line
The cost of food in real terms fell by 75% between 1974 and 2005,but increased sharply from mid-2005 onwards. As the most widelygrown food crop in the world, the tripling of the price of maizebetween 2005 and 2008 was shocking and unprecedented. It alsocoincided with a sharp increase in the use of maize to producebioethanol, resulting in a passionate debate focusing on the role ofbiofuels in food pricing and food security (Bailey, 2008). AWorld Bankreport published in 2008 concluded that biofuels produced from
1 The scientific community uses these terms interchangeably. In common usage,maize is the preferred term in Europe, whereas corn is preferred in North America andAustralia.
grains were responsible for part of the increase in grain prices,although many additional factors were involved as outlined in Fig. 3(Mitchell, 2008). The OECD came to a similar conclusion: “…theimpact of current biofuel policies onworld crop prices, largely throughincreased demand for cereals and vegetable oils, is significant butshould not be overestimated. Current biofuel support measures aloneare estimated to increase average wheat prices by about 5 percent,maize by around 7 percent and vegetable oil by about 19 percent overthe next 10 years” (OECD, 2008). It is also important to note thatethanol is made from field corn, a grain that is indigestible by humansin its raw form and that requires processing before it can be consumed.Very little field corn is actually used for human food ingredientprocessing. The overwhelmingmajority of US corn (including exports)is used to feed livestock rather than humans. Additionally, the ethanolprocess creates feed and food ingredients — the starch portion ofthe kernel is converted to ethanol, while the protein, fat and othernutrients, vitamins and minerals are passed through to the feed co-products or human food ingredients. Indeed there is more foodper capita today on a global scale than ever before and lack ofinfrastructure, access to capital, general poverty and other issuesare the more likely causes of hunger — not scarcity of food (reviewedby NCGA, 2008). A balanced assessment of the impact of biofuelcrops on food security has been published recently (Martindale andTrewavas, 2008).
3. Gene transfer in maize — from single genes to multiple traitsand pathways
3.1. Maize transformation
In the 1980s, Agrobacterium-mediated gene transfer emerged asthe most straightforward way to generate transgenic dicotyledonousplants, but few monocots were amenable to this process. Efforts totransform maize therefore began with physical delivery systemstargeting protoplasts (Fromm et al., 1985; Klein et al., 1988). Thefirst transgenic maize plants produced by protoplast transformationwere sterile (Rhodes et al., 1988) but fertile transgenic plants were
Serbia nr nr 6415 4054 6000South Africa 11,716 6935 7300 12,500 11,500Ukraine 8800 7150 6400 7400 9500United States 299,914 282,311 267,598 332,092 305,319Others 69,922 73,149 68,880 71,660 73,929World total 715,770 699,165 712,442 791,959 781,359
a Projected values.
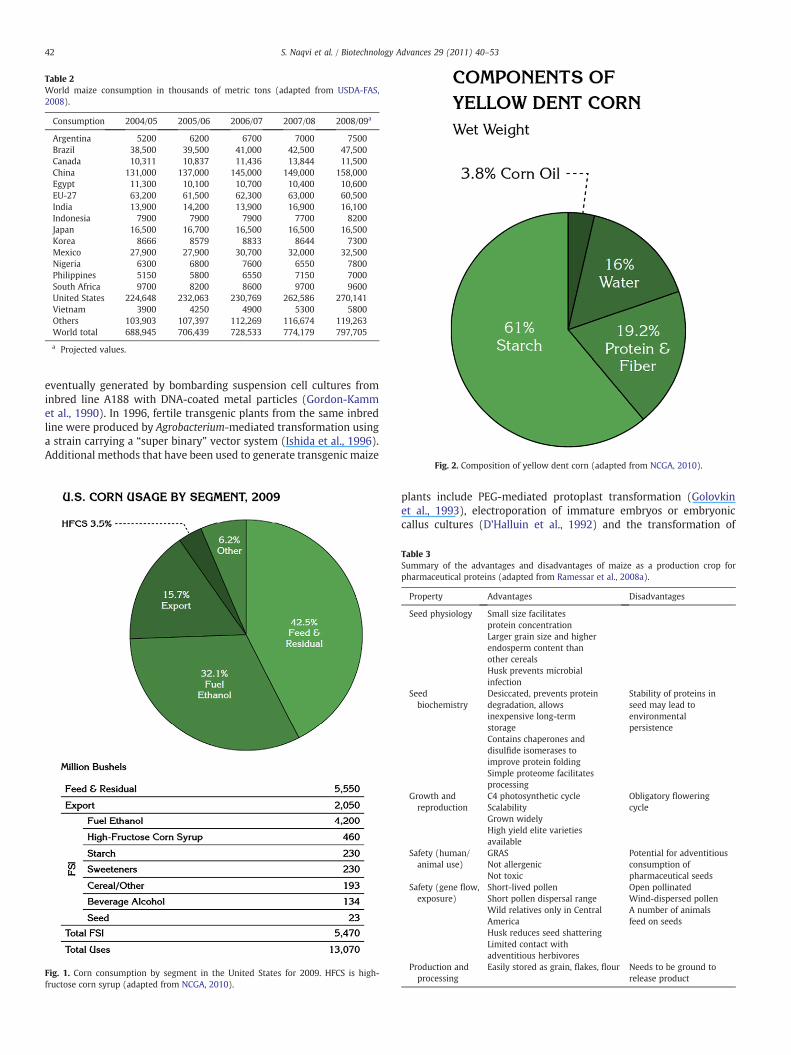
Table 2World maize consumption in thousands of metric tons (adapted from USDA-FAS,2008).
Consumption 2004/05 2005/06 2006/07 2007/08 2008/09a
Argentina 5200 6200 6700 7000 7500Brazil 38,500 39,500 41,000 42,500 47,500Canada 10,311 10,837 11,436 13,844 11,500China 131,000 137,000 145,000 149,000 158,000Egypt 11,300 10,100 10,700 10,400 10,600EU-27 63,200 61,500 62,300 63,000 60,500India 13,900 14,200 13,900 16,900 16,100Indonesia 7900 7900 7900 7700 8200Japan 16,500 16,700 16,500 16,500 16,500Korea 8666 8579 8833 8644 7300Mexico 27,900 27,900 30,700 32,000 32,500Nigeria 6300 6800 7600 6550 7800Philippines 5150 5800 6550 7150 7000South Africa 9700 8200 8600 9700 9600United States 224,648 232,063 230,769 262,586 270,141Vietnam 3900 4250 4900 5300 5800Others 103,903 107,397 112,269 116,674 119,263World total 688,945 706,439 728,533 774,179 797,705
a Projected values.
42 S. Naqvi et al. / Biotechnology Advances 29 (2011) 40–53
eventually generated by bombarding suspension cell cultures frominbred line A188 with DNA-coated metal particles (Gordon-Kammet al., 1990). In 1996, fertile transgenic plants from the same inbredline were produced by Agrobacterium-mediated transformation usinga strain carrying a “super binary” vector system (Ishida et al., 1996).Additional methods that have been used to generate transgenic maize
Fig. 1. Corn consumption by segment in the United States for 2009. HFCS is high-fructose corn syrup (adapted from NCGA, 2010).
Fig. 2. Composition of yellow dent corn (adapted from NCGA, 2010).
plants include PEG-mediated protoplast transformation (Golovkinet al., 1993), electroporation of immature embryos or embryoniccallus cultures (D'Halluin et al., 1992) and the transformation of
Table 3Summary of the advantages and disadvantages of maize as a production crop forpharmaceutical proteins (adapted from Ramessar et al., 2008a).
Property Advantages Disadvantages
Seed physiology Small size facilitatesprotein concentrationLarger grain size and higherendosperm content thanother cerealsHusk prevents microbialinfection
Seedbiochemistry
Desiccated, prevents proteindegradation, allowsinexpensive long-termstorage
Stability of proteins inseed may lead toenvironmentalpersistence
Contains chaperones anddisulfide isomerases toimprove protein foldingSimple proteome facilitatesprocessing
Growth andreproduction
C4 photosynthetic cycle Obligatory floweringcycleScalability
Grown widelyHigh yield elite varietiesavailable
Safety (human/animal use)
GRAS Potential for adventitiousconsumption ofpharmaceutical seeds
Not allergenicNot toxic
Safety (gene flow,exposure)
Short-lived pollen Open pollinatedShort pollen dispersal range Wind-dispersed pollenWild relatives only in CentralAmerica
A number of animalsfeed on seeds
Husk reduces seed shatteringLimited contact withadventitious herbivores
Production andprocessing
Easily stored as grain, flakes, flour Needs to be ground torelease product

Fig. 3. The short- and long-term factors contributing to food price increases in the 21st century (adapted from Trostle, 2008).
43S. Naqvi et al. / Biotechnology Advances 29 (2011) 40–53
embryogenic cell suspensions (Frame et al., 1994) or callus cultures(Petolino et al., 2000) using silicon carbide whiskers. Agrobacterium-mediated transformation and particle bombardment remain themost widely used methods for transformation. A list of markergenes used for the selection and regeneration of transgenic maizeplants is provided in Table 4.
3.2. Multiple gene transfer
The novel traits introduced into commercial cultivars of maizewere initially limited to herbicide tolerance and pest or diseaseresistance, primarily for use in the agriculture systems of theindustrialized world. However, horizons have expanded morerecently following research into the metabolic pathways that produceprimary and secondary metabolites, particularly for compoundsimportant for human health and nutrition including amino acids(Galili and Höfgen, 2002), very long chain polyunsaturated fatty acids
Table 4Properties of screenable and selectable marker genes used in maize.
Selectable/screenablemarker genes Characteristics
bar Phosphinothricin and bialaphos resistanceaphIV Hygromycin resistanceepsps synthase Glyphosate resistancenptII Kanamycin resistanceALS Sulfonylurea resistancemdhfr Inhibition of mDHFR enzymecat Chloramphenicol resistancepmi Converts mannose-6-phosphate to fructose-6-
phosphateipt ESP phenotype and loss of apical dominanceluc Emits yellow-green light (560 nm) with
luciferin substrategfp Green fluorescent proteindsRED Red fluorescent proteingusA Encodes β-glucuronidaseLC Anthocyanin biosynthetic geneC1 Anthocyanin biosynthetic geneB-Peru Anthocyanin biosynthetic geneB-I Anthocyanin biosynthetic gene
(Napier et al., 2006), vitamins (Giuliano et al., 2008; Zhu et al., 2008;Chen et al., 2006; Ishikawa et al., 2006) and minerals (Drakakakiet al., 2005). The complexity of such pathways means that single genetransfer strategies are becoming less appropriate to modify targettraits. Metabolic pathways may have multiple branches, multifunc-tional enzymes, cell-type-specific and compartmentalized enzymes,and complex feedback mechanisms (Verpoorte et al., 2000). Underthese circumstances single gene transfer is often unhelpful becausepathways are regulated at multiple steps. More recently, researchershave developed new transformation methods that recognize thedesire to introduce multiple transgenes into plants and express themin a coordinated manner (Naqvi et al., 2009a). Multiple gene transfercan augment, reconstruct or extend complex biosynthetic pathwaysin plants, including maize (Zhu et al., 2008) and can allow thesynthesis of complex multimeric proteins (Capell and Christou, 2004;Twyman et al., 2005).
The simultaneous transfer of multiple genes has made it possibleto develop a combinational transformation system that not onlyallows libraries of transgenic metabolic variants to be generated, andnovel metabolic traits to be selected (Zhu et al., 2008) but also has apotential to generate transgenic plants with multiple traits (Naqviet al., 2009b). The multiplex transgenic plants were generated byparticle bombardment, coating the particles with different vectorsand screening for plants expressing different combinations oftransgenes. In the future, the most complex strategies for genetransfer to plants will only be achievable using multiple gene transfertechniques (Altpeter et al., 2005).
3.3. Strategies used to synthesize value added products in maize
When discussing value added products we can draw a distinctionbetween two broad classes of compounds — proteins and low-molecular-weight metabolites. This distinction is important becausemost transgenes encode proteins, and in some cases the proteinencoded by the transgene is itself the desired molecule, while in othercases the protein is only the first step in the pathway towards thedesired compound, e.g. it may encode an enzyme that helps the plantto produce a particular chemical compound. In much the same way,transgenes can also be used to express functional RNA molecules to

44 S. Naqvi et al. / Biotechnology Advances 29 (2011) 40–53
affect the outcome of a metabolic pathway to encourage the synthesisof a specific, desired compound. The first approach (where the proteinitself is the desired molecule) can be termed molecular farming, ormolecular pharming if the protein of interest has a medical use; thesecond (where the transgene alters the metabolic phenotype of theplant and leads to the production of a low-molecular-weightcompound) can be termed metabolic engineering.
Molecular pharming in plants has been developed for 20 years andit is now possible to express virtually any protein of interest in largequantities and extract it directly from plant tissue. Maize was the firstcereal to be developed into an expression platform thanks largely tothe efforts of scientists at Prodigene Inc., College Station, TX, and it hasbeen used to produce vaccines, antibodies, industrial enzymes andhigh-value pharmaceutical polypeptides (reviewed by Ramessar et al.,2008a). Maize seeds can also be used as edible vaccines, asdemonstrated by the expression of a Newcastle disease virus fusionprotein in transgenic maize seeds which, when fed to chickens,induced the production of antibodies conferring protection against aviral challenge (Guerrero-Andrade et al., 2006). Due to its status as amolecular pharming crop, much research has focused on strategies toimprove recombinant protein expression levels in maize seeds andtarget proteins to ensure correct folding, assembly and post-translational modification. Proteomic analysis has been carried outto elucidate protein storage and accumulation kinetics in maizeendosperm to maximize recombinant protein stability. Bespokedownstream purification processes have been developed for maize,to achieve efficient recovery and high levels of purity, as shown forthe monoclonal anti-HIV antibodies 2F5 and 2G12, where N90% ofthe antibody was recovered at N95% purity using a modified affinitychromatography process (Platis and Labrou, 2008; Ramessar et al.,2008b).
Whereas efforts in molecular pharming aim to increase proteinyields, stability and recovery, metabolic engineering is a diverse fieldin which multiple strategies can be employed to achieve the goal ofincreasing the recovery of a specific target compound. Such strategiesinclude overexpressing rate-limiting enzymes to enhance the syn-thesis of a target compound (Zhu et al., 2008), silencing genesencoding enzymes that degrade or convert a target compound orsynthesize a competitive compound using a common precursor(Houmard et al., 2007; Frizzi et al., 2008), expressing enzymes alteredto avoid feedback control (Huang et al., 2005; Frizzi et al., 2008), andexpressing multiple enzymes to extend a truncated metabolicpathway or completely import a heterologous pathway (Zhu et al.,2008). It may be necessary to ensure that the desired metaboliteaccumulates in a certain tissue (e.g. tubers and seeds) or in specificsubcellular compartments (e.g. the vacuole) to avoid toxicity or off-target effects, thus regulation of transgene expression and proteintargeting is every bit as important in metabolic engineering as it is inmolecular pharming.
4. Value added products. Specific targets — primary metabolism
4.1. Carbohydrates
Plants make both polymeric carbohydrates (e.g. glucans andfructans) and individual sugars (e.g. glucose and fructose). Carbohy-drates are not only ubiquitous in human and animal diets, they arealso used in food processing and in the manufacture of adhesives,cosmetics, detergents, paper and textiles (Ellis et al., 1998). Over halfthe mass of a maize seed is carbohydrate, predominantly starch(Fig. 2).
Starch, which is stored in the form of water insoluble granules, iscomposed primarily of two types of α-glucan: amylose and amylo-pectin. Amylose is a predominantly linear polymer of α-1,4-linkedglucose residues with rare α-1,6-linkages, whereas amylopectinconsists of α-1,4-linked glucan chains and is highly branched with
5–6% α-1,6-bonds (Buleon et al., 1998). Typical amylose levels instarches are 15–25% (Manners, 1979) but starch from the wx (waxy)mutant of maize is virtually amylose free (Shure et al., 1983) whereasamylose extender (ae) mutants (Boyer and Preiss, 1978) comprise50–85% amylose (Manners, 1979).
The ratio of amylose to amylopectin in starch determines itsphysical and chemical properties and thereby its end-use, soengineering starch synthesis in maize can be used to tailor the cropfor different applications (Fergason, 1994). The suppression ofendogenous starch branching enzyme 2 (SEB 2) by RNAi increasedthe amylose content up to 50% although the total starch contentremained the same (Chai et al., 2005). Another approach that may beworth considering is overexpression of the plastidial adenylatetransporter, since Bt-1 mutants lacking this protein have low starchlevels (Shannon et al., 1998; Creech, 1965).
Although only limited success has been achieved by the directmodification of starch biosynthesis, alternative glucans can beproduced from sucrose through the expression of glucosyltransferases(GTFs). These are bacterial enzymes that catalyze the polymerizationof sucrose, which is abundant in the developing maize seed, leading tothe production of high molecular weight glucans with α-1,3/α-1,6linkages that do not normally exist in higher plants. Zhang et al.(2007) fused amutant form of the gtfD gene from Sreptococcus mutansto the maize plastid Brittle-1 transit peptide for amyloplast targeting.Overexpression of gtfD resulted in the accumulation of a novel, solubleα-(1–6)-linked glucan at high levels in the mature kernels (up to 14%of their dry weight) (Zhang et al., 2007).
Phytoglycogen is a highly branched, water-soluble glucan pro-duced in addition to starch in the sugary-1 mutants of maize (Creech,1965) and rice (Nakamura et al., 1997). In both species, thephytoglycogen content of mutant grains is approximately 25–30% oftotal carbohydrates. Increasing the phytoglycogen content above 30%could produce polysaccharides with unique functional propertiesincluding reduced viscosity, gel formation and retrogradation rate, butincreased water retention, making it easier for livestock to digest(Johnson et al., 2001a,b). Researchers have attempted to increasephytoglycogen levels by stacking different mutant alleles in maize buthave been unsuccessful thus far (Wang et al., 1993).
Finally, fructans are fructose polymers localized, unlike starch, invacuoles rather than amyloplasts. Fructans are important componentsof functional foods because they have probiotic properties and offerprotection against certain types of cancer (Menne et al., 2000;Roberfroid, 2000). Tissue-specific expression and targeting of Bacillusamyloliquefaciens SacB (levan sucrase) to maize endosperm vacuolesresulted in the accumulation of high molecular weight fructan,representing 4–8% of the dry weight of mature seeds (Caimi et al.,1996).
4.2. Essential amino acids and nutritional proteins
In humans, eight of the twenty standard amino acids are describedas essential because they cannot be synthesized de novo and must beobtained from thediet.However,most staple food crops are deficient forcertain essential amino acids, with the levels of lysine and tryptophanbeing particularly low in maize (Galili and Höfgen, 2002; Huang et al.,2006). Thus tryptophan and lysine deficiency are prevalent indeveloping countries where maize is the sole or predominant sourceof food.
The naturally occurring opaque-2 mutant has higher lysine andtryptophan levels than wild type maize but has a soft, chalky kernelwhich is unsatisfactory for cooking (Mertz et al., 1964). The highercontent of essential amino acids in the mutant results from a reducedlevel of certain storage proteins (e.g. α- and β-zeins) allowing otherproteins to replace them and increase the level of lysine andtryptophan. Breeding programs have resulted in the development ofQuality Protein Maize (QPM) which benefits from the higher lysine

Table 5Selected nutritional properties of maize (data from UDSA nutrient database, www.ars.usda.gov).
Micronutrient Amount in 100 g maize seed Percentage of RDI
Vitamin A 10 μg equiv 1%Niacin (B3) 1.7 mg 11%Folate (B9) 46 μg 12%Vitamin C 7 mg 12%Iron 0.5 mg 4%
45S. Naqvi et al. / Biotechnology Advances 29 (2011) 40–53
and tryptophan levels of the opaque-2mutant but combines this witha hard kernel with superior cooking qualities (Prasanna et al., 2001).Further investigation has shown that although the QPM has lowerlevels of α- and β-zeins compared to wild type maize, it has higherlevels of the γ-zein. Zarkadas et al. (2000) showed that high lysineQPM may provide up to 73% of human protein requirements,compared to 28–50% for common maize. The laborious breedingprogram that gave rise to QPM can be replicated much more rapidlywith RNAi. Segal et al. (2003) transformed maize with an RNAiconstruct resulting in a phenocopy of the opaque-2 phenotype.Transgenic seeds contained less α-zein than normal (and lower levelsof leucine, alanine and glutamine) but higher levels of lysine.Recently, Huang et al. (2006) obtained similar results by transformingmaize with constructs expressing chimeric double-stranded RNA.
Two transgenic strategies have been reported in the literature toincrease the content of essential amino acids in maize. The firstinvolves increasing the free amino acid pools throughmanipulation ofthe corresponding biosynthetic pathways, although as a sole approachthis can result in the amino acids leaching during food processing andcooking. The second is to enhance the level of proteins containingessential amino acids either by modifying the sequence of seedstorage protein genes to incorporate more lysine and tryptophancodons, or by importing heterologous or completely syntheticproteins containing tryptophan and lysine and expressing suchproteins in the seeds. Although leaching is not a problem with thisstrategy, high lysine/tryptophan proteins will not accumulate if theamino acid pools are depleted, so the best overall approach is toenhance the free amino acid pools and express proteins with highessential amino acid contents. Another novel approach is the over-expression of Arabidopsis thaliana lysyl tRNA synthetase (AtKRS),which results in the increased insertion of lysine residues in place ofother amino acids. Stable, elevated expression of AtKRS in transgenicmaize resulted in significant lysine recoding in zein proteins normallylacking lysine, increasing the lysine content of transgenic grain byup to 26% (Wu et al., 2007). Stable, elevated expression of AtKRS intransgenic maize causes significant recoding of lysine in zeins whichnormally have little to no lysine, enriching grain (Wu et al., 2007). Thegrain of transgenic maize contained up to 26% elevated lysine content(Wu et al., 2007). Crossing a maize variety with reduced levels of α-zein and a transgenic line expressing a lysine biosynthetic enzymereleased from feedback inhibition (CordapA), resulted in a doubling ofthe lysine content in F1 progeny (Huang et al., 2005). Houmard et al.(2007) reported that maize seeds accumulated more lysine whenlysine catabolism was repressed using RNAi to inhibit the key enzymelysine-ketoglutarate reductase/saccharopine dehydrogenase (LKR/SDH). Frizzi et al. (2008) combined both approaches, expressing thederegulated version of CordapA and a LKR-SDH-specific RNAi (fromthe same construct), resulting in an unprecedented 40-fold increase inlysine levels.
The expression of lysine-rich proteins such as hordothionine(HT12) containing 28% lysine and barley high lysine protein 8 (BHL8)containing 24% lysine has achieved significant increases in total lysinelevels in maize seeds, but coexpression with the bacterial enzymeDHPS elevated the total lysine content to over 0.7% compared to 0.2%in wild type seeds (Jung and Carl, 2000). Transgenic maize expressingthe sb401 gene encoding a protein with a very high lysine content,increased lysine levels up to 54.8% and total protein content up to 39%.These levels were maintained over six generations (Yu et al., 2004).Lysine levels could also be increased by expressing animal nutritionalproteins, such asmilk proteins. The expression of porcineα-lactalbuminin maize was tested with two targeting strategies, one constructcontaining a zein signal sequence and one containing an endoplasmicreticulum (ER) retention signal, but individual amino acid levels werenot reported (Yang et al., 2002). Recently, Bicar et al. (2008) carried outa similar experiment and showed that α-lactalbumin expression inmaize did indeed increase the lysine content.
4.3. Fatty acids
Although higher plants synthesize over 300 different types of fattyacids (Aitzetmuller et al., 2003), only a few are common to all plantse.g. the saturated fatty acids palmitic acid (16:0) and stearic acid(18:0), and the unsaturated fatty acids oleic acid (18:1Δ9), linoleicacid (LA) (18:2Δ9, 12), and α-linolenic acid (ALA) (18:3Δ9,12,15)(Broun et al., 1999). The relative proportions of the different types offatty acids impact on cooking properties, and monounsaturated fattyacids are considered to be healthier than saturated and polyunsatu-rated fatty acids (PUFAs) (Kinney et al., 2002).
Most fatty acids can be synthesized de novo by humans but certainPUFAs cannot and must be obtained from the diet. These includethe longer chain ω-3 fatty acids normally found in fish andmarine microorganisms, eicosapentaenoic acid (EPA; C20:5n−3)and docosahexaenoic acid (DHA; C22:6n−3), which play an impor-tant role in the regulation of inflammatory responses, blood pressureand brain development. Stearidonic acid (SDA; C18:4n−3), EPA andDHA also help to prevent certain cancers (Christensen et al., 1999;Smuts et al., 2003; Reiffel and McDonald, 2006).
For many years, scientists have been looking for healthy replace-ments for palm and hydrogenated oils that have similar propertiessuch as solidity and resistance to oxidation. Oils rich in oleic acid meetsome of these needs, and high oleic mutants, with an oleic acidcontent ranging from 60% to 90%, have been developed in maize.These are FAD2mutants (Belo et al., 2008), the FAD2 gene encoding anomega-6 desaturase that converts oleic acid to linoleic acid. Morerecently, Zheng et al. (2008) showed that a high-oil QTL (qHO6)affecting maize seed oil and oleic acid contents encodes an acyl-CoA:diacylglycerol acyltransferase (DGAT1–2), which catalyzes the finalstep of oil synthesis and has been implicated as a rate-limiting step(Jako et al., 2001; Lung and Weselake, 2006). A phenylalanineinsertion in DAGT1–2 at position 469 is responsible for the increasedoil and oleic acid contents (Zheng et al., 2008). Ectopic expression ofthe high-oil DAGT1–2 allele increases oil and oleic acid contents byup to 41% and 107%, respectively (Zheng et al., 2008). This is thefirst report of direct metabolic engineering for fatty acid content inmaize, a strategy that has been employed in model plants and oilcropsextensively (Truksa et al., 2009).
5. Value added products. Specific targets — vitamins and minerals
5.1. Micronutrient levels in maize
Cereals including maize are generally deficient in most vitaminsand minerals, which means that people relying on maize as a staplefood risk malnutrition even if they obtain sufficient calories from theirdiet. Typical nutritional values for maize seeds are listed in Table 5,which is sourced from the USDA nutrient database. A typical serving of100 g of seeds contains approximately 10–15% of the recommendeddaily intake of the principal B vitamins and vitamin C (ascorbate) butonly 1% of the RDI of vitamin A. Similarly, maize seeds are poorsources of iron, selenium, magnesium and potassium. In theindustrialized world, micronutrient deficiency is avoided by varieddiets, supplementation, and the fortification of processed cereals and

46 S. Naqvi et al. / Biotechnology Advances 29 (2011) 40–53
bread (Zhu et al., 2007; Gómez-Galera et al., 2010). In developingcountries where such methods are limited in impact, the directfortification of cereal crops at source may be the only way to reducethe incidence of micronutrient deficiency in the long term.
5.2. Conventional and transgenic strategies to enhance carotenoid levels
Because maize has such a diverse gene pool, significant variationexists in different cultivars and varieties with respect to the levels ofvitamins and minerals. For this reason, conventional breedingapproaches have been used to select more nutritious varietiesalthough this has met with only limited success. For example, manyresearchers have studied variation in β-carotene levels in maize, sincethis affects the kernel color and is an easily scored phenotype(Kurilich and Juvik, 1999; Islam, 2004). Humans can synthesizevitamin A from β-carotene, so a sufficient intake of β-carotene canprevent the diseases caused by vitamin A deficiency, which includeblindness, immune dysfunction and skin disorders. Harjes et al.(2008) described four polymorphisms at the lycE locus in maize,encoding the enzyme lycopene-ε cyclase, which determines the fluxin the α-carotene and β-carotene branches of the carotenoidsynthesis pathway. However, only marginal success was achievedwith the selection of lines containing up to 13.6 μg/g dry weight ofβ-carotene (a 30–40% improvement). It would take years of conven-tional breeding to introgress this trait into locally adapted maizevarieties grown by subsistence farmers in the developing world.
Transgenic approaches offer more rapid progress towards nutri-tional enhancement since long breeding programs are unnecessary.Genes encoding the relevant biosynthetic enzymes can be transferreddirectly into the cultivar of choice allowing that single trait to bemodified without affecting the genetic background, and hence theagronomic properties of the crop. With respect to the carotenoidpathways, Aluru et al. (2008) introduced the bacterial genes crtB(encoding phytoene synthase) and crtI (which catalyzes the fourdesaturation steps in the carotenoid pathway, a process requiring twoenzymes in plants) into maize under the control of a ‘super γ-zeinpromoter’ for endosperm-specific expression. This increased thetotal carotenoid content to 33.6 μg/g dry weight and raised the levelsof β-carotene.
Using a novel combinatorial genetic transformation approach, thecarotenoid biosynthetic pathway has been modified and extended toproduce not only β-carotene but also other important carotenoidssuch as lutein, zeaxanthin, lycopene and astaxanthin, which areknown to have nutritional andmedical benefits (Zhu et al., 2008). Thisstudy, mentioned briefly earlier, involved combinatorial genetictransformation to generate a library of metabolic variants expressingdifferent combinations of five carotenogenic genes (Zmpsy1, PacrtI,Gllycb, Glbch and ParacrtW). Plants from this library expressingZmpsy1 and PacrtI under the control of an endosperm-specificpromoter contained extraordinarily high levels of β-carotene, in thebest cases reaching 56 μg/g dry weight.
5.3. Vitamins B9, C and E
Metabolic engineering is a promising alternative to artificialfortification with folate, which is particularly important during neuraldevelopment in the human fetus and has a profound effect on theincidence of neural tube defects such as spina bifida (Scott et al., 2000;Bouis, 2003; DellaPenna, 1999). There have been successful attemptsto increase folate levels in tomato fruits (Diaz de la Garza et al., 2004,2007) and rice grains (Storozhenko et al., 2007) using transgenicstrategies, and using a similar strategy in maize, Naqvi et al. (2009b)increased the total folate content up to 2-fold. In the case of vitamin C,constitutive expression of the enzyme dehydroascorbate reductase(DHAR) in maize doubled the vitamin C level (Chen et al., 2003). Thisenzyme recycles oxidized ascorbate, and similar results were obtained
when endosperm-specific expression of DHAR resulted in a 6-foldincrease in vitamin C content in the transgenicmaize plants comparedto the wild type (Naqvi et al., 2009b).
Vitamin E is a collection of eight related vitamers comprisingfour tocopherols and four tocotrienols, collectively known astocochromanols (DellaPenna and Pogson, 2006). The most potent ofthese isα-tocopherolwhich has 100-fold the activity of other vitamers(Traber and Sies, 1996). Plants vary in their tocochromanol contentand in the proportions contributed by the different compounds,with α-tocopherols more abundant in leaves and γ-tocopherolpredominating in seeds (Tan, 1989; Demurin et al., 1996).The analysisof tocochromanols in different maize inbred lines has revealed thatseeds show substantial natural variation in the levels of α-, γ- and δ-tocopherols (Grams et al., 1970; Egesel et al., 2003). The maizetocopherol synthesis pathway remains to be fully elucidated, but onerelevant gene (VTE1, encoding tocopherol cyclase) has been clonedand characterized (Provencher et al., 2001; Sattler et al., 2003). In amore recent study, chromosomal regions associated with quantitativevariation of tocopherol content have been identified inmaize. Chanderet al. (2008) used QTL mapping combined with the candidate geneapproach in recombinant inbred lines (RIL) with different geneticbackgrounds to study tocopherol accumulations in maize. This studyidentified six candidate genes in the tocopherol biosynthesis pathway(HPPD, VTE1, VTE3, VTE4, P3VTE5, and P4VTE5), and 31 QTLs associatedwith quantitative variation of tocopherol content and compositionwere identified by composite interval mapping (CIM).
Metabolic engineering could be used either to increase totaltocochromanol levels or shift the balance of vitamers in favor of γ-tocopherol (DellaPenna, 1999; Savidge et al., 2002). The expression oftocochromanol synthesis genes in model plants and crops such ascanola and soybean (Karunanandaa et al., 2005) has boosted vitaminE levels in transgenic plants primarily by increasing the amount ofα-tocopherol. Limited progress has been made in maize, which hasthe second highest tocopherol content after soybean and a greaterproportion of α-tocopherol (Grusak and DellaPenna, 1999). Cahoonet al. (2003) created transgenic maize plants expressing barleyhomogentisate geranylgeranyl transferase, the first committed en-zyme in the tocochromanol pathway, under the control of the embryo-specific maize 16-kDa oleosin gene promoter. This increased totaltocochromanol levels 6-fold, mainly due to an increase in tocotrienollevels (tocopherol levels remained largely unchanged). Simultaneousexpression of ρ-hydroxyphenylpyruvate dioxygenase and MPBQmethyltransferase in maize resulted in a 3-fold increase in totaltocopherol content (Naqvi et al., 2010). This increase in tocopherolcontent in transgenic seeds was due to an increase in γ-tocopherolwhereas other tocopherol isomers remained undetectable (Naqviet al., 2010).
6. Value added products. Specific targets — recombinant proteins
6.1. Pharmaceutical proteins
The biopharmaceutical industry is responsible for a new wave ofdrugs based on mammalian proteins, including a burgeoning industrybased on monoclonal antibodies. Furthermore, the expression ofpathogen subunit proteins as vaccines provides a safe and effectiveway to protect humans and animals against emerging and re-emerging diseases. The industry currently relies on microbialproduction systems (mostly E. coli and yeast) and cultured mamma-lian cells, but both are fermenter-based and hence expensive andlimited in capacity (Torres et al., 1999). Plants have all the advantagesof mammalian cells (i.e. they can express complex proteins such asantibodies and can fold and assemble them properly) yet they are aseconomical as microbial systems but unlimited in capacity, and haveadditional advantages such as the lack of human pathogens and adiversity of platforms offering niche concepts (e.g. long-term storage

Table 7Pharmaceutical proteins produced in maize, intended for veterinary applications(adapted from Ramessar et al., 2008a).
Product Indication Status
VaccinesE. coli heat labile toxin(ETC LtB)
Diarrhea Two independent phase Itrials
PTGEV, porcine transmissiblegastroenteritis virus(oral vaccine)
Piglet gastroenteritis Phase I completed
Newcastle disease virus(NCDV) (oral vaccine)
Protection againstviral changes
Tested in chickens.Determination of theappropriate dose
AntibodiesMonoclonal antibody E559 Rabies Experimental studies
47S. Naqvi et al. / Biotechnology Advances 29 (2011) 40–53
in seeds, administration of oral vaccines in fruit, unique purificationsystems in oilseeds) (Twyman et al., 2003, 2005; Stoger et al., 2005a,b; Ma et al., 2005; Basaran and Rodriguez-Cerezo, 2008).
Cereal seeds, particularly maize, are useful for pharmaceuticalproteins because they can be stored for years, even at roomtemperature, without the recombinant protein losing activity (Stogeret al., 2002a). This means that drugs expressed in seeds can bedistributed and stored in countries that lack a cold chain, and can beextracted in local facilities rather than relying on central facilities andnational distribution networks. Seeds are natural protein storageorgans with an appropriate biochemical environment to promotestable protein accumulation with no detectable loss of activity. Inaddition, cereal seeds lack compounds such as the phenolics andalkaloids present in tobacco leaves, which complicate downstreamprocessing and purification. Therefore, cereal seeds are an idealvehicle for the stable accumulation of recombinant proteins (Rames-sar et al., 2008c).
Functional recombinant monoclonal antibodies have been suc-cessfully produced in plants since 1989 and are now one of the mostadvanced biopharmaceutical categories produced in plants (reviewedby Twyman et al., 2005; Nicholson et al., 2005; Stoger et al., 2002b).Several antibodies have been produced successfully in maize seeds(Tables 6 and 7), some of which have been difficult to produceeconomically in mammalian cells (Chintalacharuvu and Morrison,1997). One of the main benefits of using plants for antibodyproduction is the economy of scale. This is an important factor forantibodies intended for high-volume use, such as those destined to beincluded in topical microbicides where the doses aremuch larger thanantibodies intended for injection. The anti-HIVmonoclonal antibodies2F5 and 2G12 are good examples, since effective neutralizationrequires the application of milligram amounts of antibody. To meetthe global demand, both antibodies would need to be produced on theton scale, which in a fermenter-based platform would be economi-cally unfeasible. However, both antibodies have been produced in
Table 6Pharmaceutical proteins produced in maize, intended for administration to humans(adapted from Ramessar et al., 2008a).
Product Indication Status
VaccinesE. coli heat labile toxin(ETEC LtB)
Diarrhea Two independentphase I trials
AntibodiesAvicidin Colorectal cancer Withdrawn from
phase II in 1998Humanized IgG Therapy use (inhalation):
respiratory syncytial virusTested in animalmodels
Humanized IgG Therapy use (oral): Clostridiumdifficile
Tested in animalmodels
Humanized IgG Sperm (topical): contraceptive Tested in animalmodels
2G12 HIV Neutralization ofthe virus
Monomeric hIgA1 Herpes simplex virus (HSV)(anti-herpes)
Experimental studies
Monomeric hIgA1 SAGA-1 antigen Experimental studies
Other productsGastric lipase Cystic fibrosis, pancreatitis Phase II trialsLactoferrin Gastrointestinal infections Phase I trialsAvidin Diagnostic use Available in Sigma
catalogueTrypsin Wound care/insulin
manufactureAvailable in Sigmacatalogue
Aprotinin Protease inhibitor in tissuecultures and cardiac surgery
Marketed byProdigene
Recombinant humanproinsulin (rhProinsulin)
Diabetes Experimental studies
plants where the global demand could bemet with just a few hectaresof maize producing the antibodies in the seeds. The successfulexpression of 2G12 in maize, and the extraction of a pure andfunctional antibody, has been documented (Ramessar et al., 2008b;Rademacher et al., 2008).
Antibodies for ‘passive’ immunization are one important targetfor molecular pharming, but ‘active’ immunization with antigensproduced in plants is another emerging field. The use of plants tosynthesize pathogen antigens as potential vaccines carries all theadvantages of safety and scalability listed earlier for antibodies, butstudies have shown that the antigens produced by plants can induceimmune responses at both the mucous and serum levels, and canbe administered by injection as well as orally, the latter providingsignificant cost benefits by removing the necessity for extensivedownstream processing. Several clinical studies have been carriedout with antigens expressed in a variety of plant tissues, showingoverwhelmingly positive results in terms of both antibody produc-tion and response to pathogen challenge (Yusibov and Rabindran,2008).
A number of different platforms have been developed for antigenexpression including tomato, tobacco, potato and other species whichfeatured in early clinical trials. However, maize has emerged as apromising candidate because of the production advantages citedearlier, the stability of the antigen in situ and the fact that manyanimals will eat maize allowing it to be used to administer veterinaryvaccines (Table 7). Maize has been used as a seed-based vaccineantigen production system (Streatfield and Howard, 2003) andresearchers have found that the environment of the seed provides“bioencapsulation” for recombinant proteins, thus delaying digestionand prolonging the time window during which immune stimulationcan occur in the gut (Zhang et al., 2006). Of particular note is thesuccess of maize-based vaccines in tackling Newcastle disease inpoultry. The Newcastle disease virus infects a wide range of bothdomestic and wild birds in most countries and has a devastating effecton commercial poultry production (Alexander, 1997; Seal et al.,2000). Feeding chickens with maize seeds containing the virus Fprotein induced the production of antibodies that conferred protec-tion against a viral challenge equivalent to a commercial vaccine thatneeds to be injected into individual chickens.
Other interesting pharmaceutical proteins produced in maizeseeds include trypsin, a pancreatic serine protease that is usuallyproduced from bovine carcasses. Because of fears surrounding thepossibility of transmissible spongiform encephalopathy, non-animalsources of trypsin are now preferred and the high-volume productionpossible in maize is ideal (Woodard et al., 2003). Similarly, aprotininwhich is sourced predominantly from bovine lung has also beenexpressed in maize to reduce the risk of contamination with prions(Zhong et al., 1999).

Table 8Industrial enzymes and non-medical protein reagents produced in maize.
Enzyme Gene Use Expression levels
Laccase (peroxidase, lignolytic enzyme) Trametes versicolorlaccase I
To improve lignin processing, used as a bleachingagent and anti-microbial; beverage clarification andwaste stream detoxification
0.20% of dry weight of seed
Cellulase/glucanase(endo-1,4-β-D-glucanase)
Acidothermuscellulolyticus
Enzymatic conversion of cellulose to fermentable sugars 2.1% of total soluble protein with enzymatic activityof 0.845 nmol/mg/min in leaf and 0.835 nmol/mg/minin root
Phytase Aspergillus nigerphyA2
Improve phosphorus availability in animal feed 2200 phytase units (FTU) per kg seed
β-Glucuronidase E. coli gusA Technical reagent 0.7% of total soluble protein
2 A neutraceutical is a substance found naturally in food which has been shown tohave a physiological or medical benefit, e.g. resveratrol and flavonoids.
48 S. Naqvi et al. / Biotechnology Advances 29 (2011) 40–53
6.2. Industrial proteins
As well as pharmaceutical proteins, which generally need to behighly purified before use (oral vaccines being the major exception),plants can also be used for the production of bulk industrial enzymesused in the food and feed industries, as well as paper, textiles andmanufacturing. As for pharmaceutical proteins, the main advantageof using plants is economical, since large scale production can beachieved by sowing additional fields rather than commissioning newfermenter facilities or hiring contract manufacturing organizations.For example, fungal laccases have been expressed in a variety ofheterologous microbes including Saccharomyces cerevisiae, Pichiapastoris, Trichoderma reesei and Aspergillus oryzae, albeit at relativelylow concentrations. Transgenic maize plants expressing high levels oflaccase as an immobilized enzyme could be grown on a large scale,and the enzyme could be extracted using a simple procedure withoutfurther enrichment or purification, thereby decreasing the productioncosts (Bailey et al., 2004).
As discussed earlier, animal-derived proteins are falling out offavor in the biopharmaceutical industry because of the risk of prioninfections, and alternative sources include transgenic plants. As wellas trypsin and aprotitin, which havemedical as well as industrial uses,a range of laboratory reagents can be produced in plants to ensure thatproducts used in cell and tissue culture are disease free. Such reagentswould include trypsin, as discussed earlier, but also avidin and β-glucuronidase both of which have been produced commercially inmaize seeds and perform as well as their counterparts extracted fromtraditional sources.
The high cost of enzymes make some industrial processes expen-sive or unfeasible on a large scale, as is the case for the conversion ofcellulosic biomass into ethanol by cellulases. Commercial cellulasesare produced in microbial systems, but the costs are prohibitive.Transgenic plants producing cellulases bring the same economics ofscale as seen for other products but the additional advantage in maizeis that recombinant cellulases can be activated in situ to beginthe conversion process without any requirement for extraction orpurification. Examples of cellulases expressed in maize are listed inTable 8.
7. Regulatory, economic and political aspects of high-valueproducts from maize
The main drivers for the sustainable use of high-value productsfrom maize are demand, economics, efficiency and safety. Thedemand for these products already exists as there is a lack ofadequate, cost-effective bioprocess manufacturing facilities for suchimportant molecules. The lower cost of goods is the primary source ofpotential benefits and therefore, the economic viability and commer-cialization of such production platforms are of great importance(Fig. 4).
Although the cost of downstream processing accounts for N85% ofexpenditure (Twyman et al., 2003) in the production of value added
molecules and nutraceuticals2 in maize, the ‘upstream productionphase’ is just as important because it influences the yield of proteinavailable for downstream processing. This yield is influenced by thechoice of genotype/explant for production and expression levels(reviewed in Ramessar et al., 2008c). Certain genotypes expressproteins at higher levels than others, and the choice of genotype oftenreflects grain morphology and the propensity to store protein withinthe endosperm. Maize is particularly advantageous over the othercereals because it has the largest annual grain yield (approximately8300 kg/ha) but lower overall production costs (Giddings et al., 2000)and also a relatively high seed protein content (10%). The choiceof variety is also influenced by the ease of transformation and the easeof purification (e.g. varieties with higher oil contents are morelikely to clog filters and chromatography resins, while those with highcarbohydrate contents support the growth of microorganisms(Farinas et al., 2005)).
High protein expression levels are beneficial because this allows agreater amount of recombinant protein to be extracted from a givenamount of plant biomass (making downstream processing andpurification more efficient), and means that fewer hectares of landare required to achieve a particular yield target. Thereforemaximizingthe accumulation of recombinant protein is an important goal (Maet al., 2003; Streatfield, 2007). The key factors that affect recombinantprotein expression levels include expression construct design, wherehigh levels of recombinant protein can be achieved by choosingcomponents that maximize the rates of transcription and proteinsynthesis, and promote transcript and protein stability (Delaney,2002; Stoger et al., 2002a; Hood et al., 2003; Tada et al., 2003).However, protein targeting is important to optimize stability, andthis is achieved by subcellular targeting which influences the folding,assembly and post-translational modification of proteins, thusaffecting their stability and final yield (Schillberg et al., 2002). Otherfactors that may be involved include zygosity, which has been shownto boost transgene expression, and the inclusion of purification tags tofacilitate extraction and purification.
7.1. Economics
The purity of the recombinant protein for its application ofteninfluences the economics of production platforms. Different puritylevels are required for different applications (Table 9), i.e. higherpurity implies more complicated and costly downstream processing.The use of maize seeds for production is advantageous as the GRASstatus means that products that do not require extensive purificationcan be administered as whole seeds, flakes, flour or paste, withminimal purification.
Because downstream processing represents such a high propor-tion of production costs in plants, it is important to ensure thatoptimal purification and processing methods are used in order to
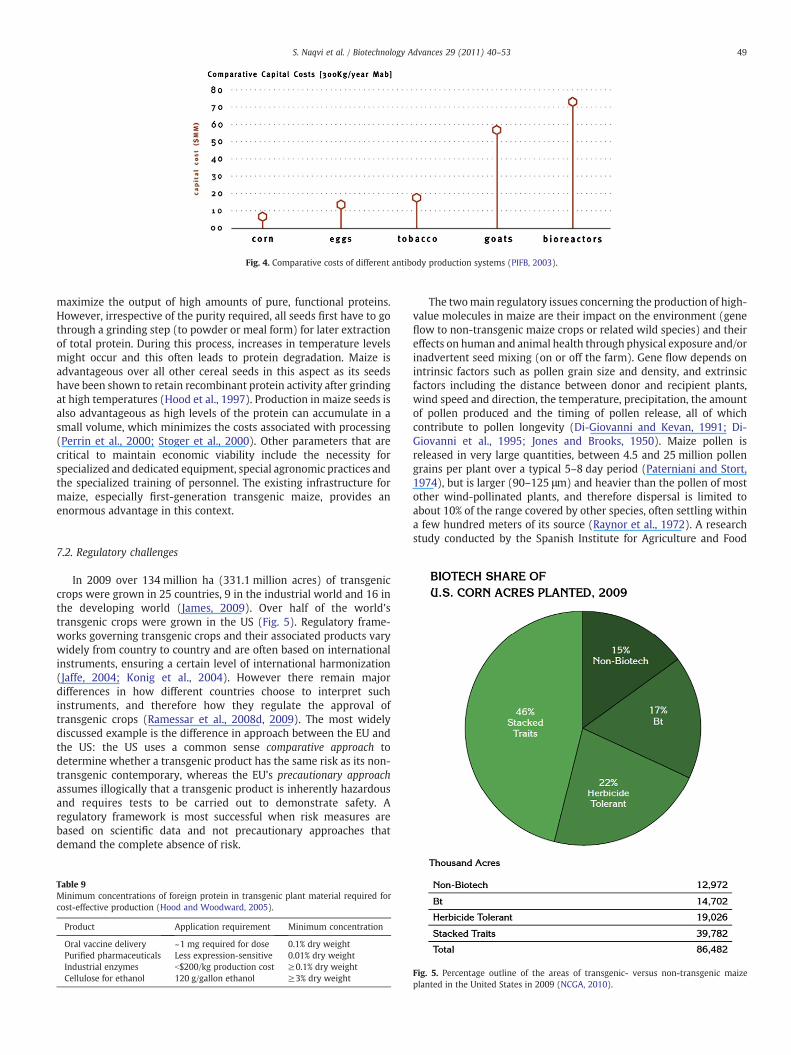
Fig. 4. Comparative costs of different antibody production systems (PIFB, 2003).
49S. Naqvi et al. / Biotechnology Advances 29 (2011) 40–53
maximize the output of high amounts of pure, functional proteins.However, irrespective of the purity required, all seeds first have to gothrough a grinding step (to powder or meal form) for later extractionof total protein. During this process, increases in temperature levelsmight occur and this often leads to protein degradation. Maize isadvantageous over all other cereal seeds in this aspect as its seedshave been shown to retain recombinant protein activity after grindingat high temperatures (Hood et al., 1997). Production in maize seeds isalso advantageous as high levels of the protein can accumulate in asmall volume, which minimizes the costs associated with processing(Perrin et al., 2000; Stoger et al., 2000). Other parameters that arecritical to maintain economic viability include the necessity forspecialized and dedicated equipment, special agronomic practices andthe specialized training of personnel. The existing infrastructure formaize, especially first-generation transgenic maize, provides anenormous advantage in this context.
7.2. Regulatory challenges
In 2009 over 134 million ha (331.1 million acres) of transgeniccrops were grown in 25 countries, 9 in the industrial world and 16 inthe developing world (James, 2009). Over half of the world'stransgenic crops were grown in the US (Fig. 5). Regulatory frame-works governing transgenic crops and their associated products varywidely from country to country and are often based on internationalinstruments, ensuring a certain level of international harmonization(Jaffe, 2004; Konig et al., 2004). However there remain majordifferences in how different countries choose to interpret suchinstruments, and therefore how they regulate the approval oftransgenic crops (Ramessar et al., 2008d, 2009). The most widelydiscussed example is the difference in approach between the EU andthe US: the US uses a common sense comparative approach todetermine whether a transgenic product has the same risk as its non-transgenic contemporary, whereas the EU's precautionary approachassumes illogically that a transgenic product is inherently hazardousand requires tests to be carried out to demonstrate safety. Aregulatory framework is most successful when risk measures arebased on scientific data and not precautionary approaches thatdemand the complete absence of risk.
Table 9Minimum concentrations of foreign protein in transgenic plant material required forcost-effective production (Hood and Woodward, 2005).
Product Application requirement Minimum concentration
Oral vaccine delivery ~1 mg required for dose 0.1% dry weightPurified pharmaceuticals Less expression-sensitive 0.01% dry weightIndustrial enzymes b$200/kg production cost ≥0.1% dry weightCellulose for ethanol 120 g/gallon ethanol ≥3% dry weight
The twomain regulatory issues concerning the production of high-value molecules in maize are their impact on the environment (geneflow to non-transgenic maize crops or related wild species) and theireffects on human and animal health through physical exposure and/orinadvertent seed mixing (on or off the farm). Gene flow depends onintrinsic factors such as pollen grain size and density, and extrinsicfactors including the distance between donor and recipient plants,wind speed and direction, the temperature, precipitation, the amountof pollen produced and the timing of pollen release, all of whichcontribute to pollen longevity (Di-Giovanni and Kevan, 1991; Di-Giovanni et al., 1995; Jones and Brooks, 1950). Maize pollen isreleased in very large quantities, between 4.5 and 25 million pollengrains per plant over a typical 5–8 day period (Paterniani and Stort,1974), but is larger (90–125 μm) and heavier than the pollen of mostother wind-pollinated plants, and therefore dispersal is limited toabout 10% of the range covered by other species, often settling withina few hundred meters of its source (Raynor et al., 1972). A researchstudy conducted by the Spanish Institute for Agriculture and Food
Fig. 5. Percentage outline of the areas of transgenic- versus non-transgenic maizeplanted in the United States in 2009 (NCGA, 2010).
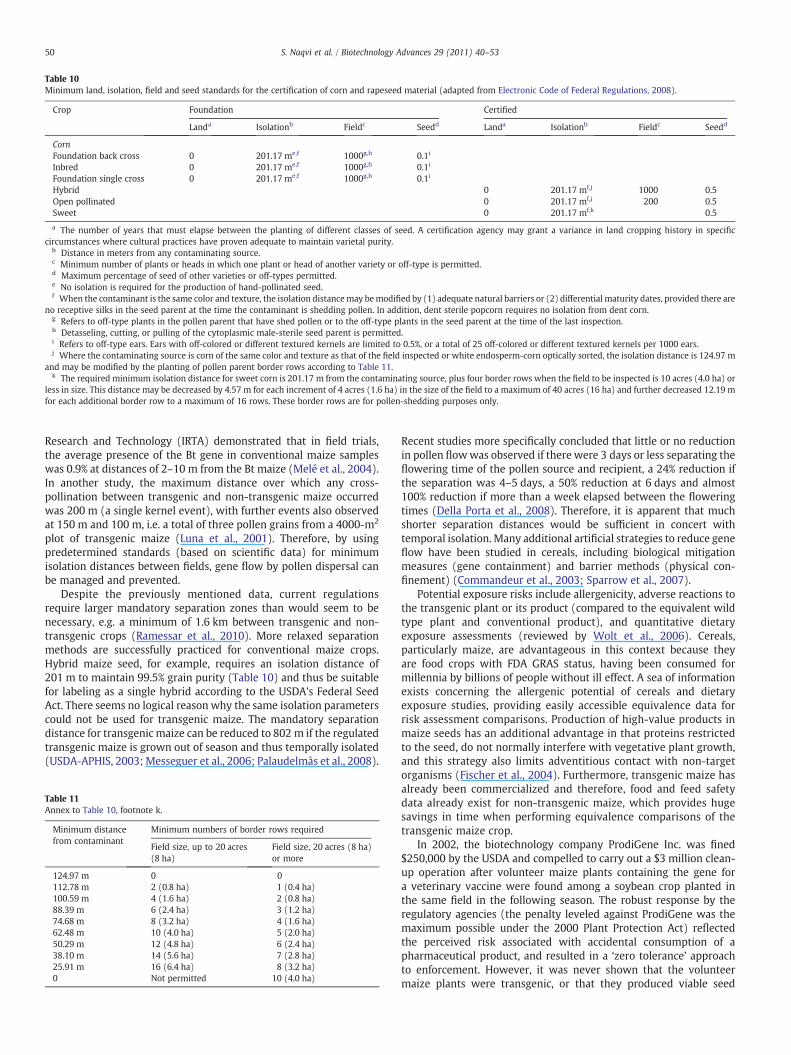
Table 10Minimum land, isolation, field and seed standards for the certification of corn and rapeseed material (adapted from Electronic Code of Federal Regulations, 2008).
Crop Foundation Certified
Landa Isolationb Fieldc Seedd Landa Isolationb Fieldc Seedd
CornFoundation back cross 0 201.17 me,f 1000g,h 0.1i
Inbred 0 201.17 me,f 1000g,h 0.1i
Foundation single cross 0 201.17 me,f 1000g,h 0.1i
Hybrid 0 201.17 mf,j 1000 0.5Open pollinated 0 201.17 mf,j 200 0.5Sweet 0 201.17 mf,k 0.5
a The number of years that must elapse between the planting of different classes of seed. A certification agency may grant a variance in land cropping history in specificcircumstances where cultural practices have proven adequate to maintain varietal purity.
b Distance in meters from any contaminating source.c Minimum number of plants or heads in which one plant or head of another variety or off-type is permitted.d Maximum percentage of seed of other varieties or off-types permitted.e No isolation is required for the production of hand-pollinated seed.f When the contaminant is the same color and texture, the isolation distancemay bemodified by (1) adequate natural barriers or (2) differential maturity dates, provided there are
no receptive silks in the seed parent at the time the contaminant is shedding pollen. In addition, dent sterile popcorn requires no isolation from dent corn.g Refers to off-type plants in the pollen parent that have shed pollen or to the off-type plants in the seed parent at the time of the last inspection.h Detasseling, cutting, or pulling of the cytoplasmic male-sterile seed parent is permitted.i Refers to off-type ears. Ears with off-colored or different textured kernels are limited to 0.5%, or a total of 25 off-colored or different textured kernels per 1000 ears.j Where the contaminating source is corn of the same color and texture as that of the field inspected or white endosperm-corn optically sorted, the isolation distance is 124.97 m
and may be modified by the planting of pollen parent border rows according to Table 11.k The required minimum isolation distance for sweet corn is 201.17 m from the contaminating source, plus four border rows when the field to be inspected is 10 acres (4.0 ha) or
less in size. This distance may be decreased by 4.57 m for each increment of 4 acres (1.6 ha) in the size of the field to a maximum of 40 acres (16 ha) and further decreased 12.19 mfor each additional border row to a maximum of 16 rows. These border rows are for pollen-shedding purposes only.
50 S. Naqvi et al. / Biotechnology Advances 29 (2011) 40–53
Research and Technology (IRTA) demonstrated that in field trials,the average presence of the Bt gene in conventional maize sampleswas 0.9% at distances of 2–10 m from the Bt maize (Melé et al., 2004).In another study, the maximum distance over which any cross-pollination between transgenic and non-transgenic maize occurredwas 200 m (a single kernel event), with further events also observedat 150 m and 100 m, i.e. a total of three pollen grains from a 4000-m2
plot of transgenic maize (Luna et al., 2001). Therefore, by usingpredetermined standards (based on scientific data) for minimumisolation distances between fields, gene flow by pollen dispersal canbe managed and prevented.
Despite the previously mentioned data, current regulationsrequire larger mandatory separation zones than would seem to benecessary, e.g. a minimum of 1.6 km between transgenic and non-transgenic crops (Ramessar et al., 2010). More relaxed separationmethods are successfully practiced for conventional maize crops.Hybrid maize seed, for example, requires an isolation distance of201 m to maintain 99.5% grain purity (Table 10) and thus be suitablefor labeling as a single hybrid according to the USDA's Federal SeedAct. There seems no logical reason why the same isolation parameterscould not be used for transgenic maize. The mandatory separationdistance for transgenic maize can be reduced to 802 m if the regulatedtransgenic maize is grown out of season and thus temporally isolated(USDA-APHIS, 2003; Messeguer et al., 2006; Palaudelmàs et al., 2008).
Table 11Annex to Table 10, footnote k.
Minimum distancefrom contaminant
Minimum numbers of border rows required
Field size, up to 20 acres(8 ha)
Field size, 20 acres (8 ha)or more
124.97 m 0 0112.78 m 2 (0.8 ha) 1 (0.4 ha)100.59 m 4 (1.6 ha) 2 (0.8 ha)88.39 m 6 (2.4 ha) 3 (1.2 ha)74.68 m 8 (3.2 ha) 4 (1.6 ha)62.48 m 10 (4.0 ha) 5 (2.0 ha)50.29 m 12 (4.8 ha) 6 (2.4 ha)38.10 m 14 (5.6 ha) 7 (2.8 ha)25.91 m 16 (6.4 ha) 8 (3.2 ha)0 Not permitted 10 (4.0 ha)
Recent studies more specifically concluded that little or no reductionin pollen flowwas observed if there were 3 days or less separating theflowering time of the pollen source and recipient, a 24% reduction ifthe separation was 4–5 days, a 50% reduction at 6 days and almost100% reduction if more than a week elapsed between the floweringtimes (Della Porta et al., 2008). Therefore, it is apparent that muchshorter separation distances would be sufficient in concert withtemporal isolation. Many additional artificial strategies to reduce geneflow have been studied in cereals, including biological mitigationmeasures (gene containment) and barrier methods (physical con-finement) (Commandeur et al., 2003; Sparrow et al., 2007).
Potential exposure risks include allergenicity, adverse reactions tothe transgenic plant or its product (compared to the equivalent wildtype plant and conventional product), and quantitative dietaryexposure assessments (reviewed by Wolt et al., 2006). Cereals,particularly maize, are advantageous in this context because theyare food crops with FDA GRAS status, having been consumed formillennia by billions of people without ill effect. A sea of informationexists concerning the allergenic potential of cereals and dietaryexposure studies, providing easily accessible equivalence data forrisk assessment comparisons. Production of high-value products inmaize seeds has an additional advantage in that proteins restrictedto the seed, do not normally interfere with vegetative plant growth,and this strategy also limits adventitious contact with non-targetorganisms (Fischer et al., 2004). Furthermore, transgenic maize hasalready been commercialized and therefore, food and feed safetydata already exist for non-transgenic maize, which provides hugesavings in time when performing equivalence comparisons of thetransgenic maize crop.
In 2002, the biotechnology company ProdiGene Inc. was fined$250,000 by the USDA and compelled to carry out a $3 million clean-up operation after volunteer maize plants containing the gene fora veterinary vaccine were found among a soybean crop planted inthe same field in the following season. The robust response by theregulatory agencies (the penalty leveled against ProdiGene was themaximum possible under the 2000 Plant Protection Act) reflectedthe perceived risk associated with accidental consumption of apharmaceutical product, and resulted in a ‘zero tolerance’ approachto enforcement. However, it was never shown that the volunteermaize plants were transgenic, or that they produced viable seed

51S. Naqvi et al. / Biotechnology Advances 29 (2011) 40–53
containing the vaccine. This was indeed a case where the companywas punished for its failure to comply with regulations, not becausethere was evidence of actual risk. It is notable that the USDA has sinceabandoned its zero tolerance policy, demonstrating clearly that theregulations are still evolving (Ramessar et al., 2008a).
8. Outlook
Maize is the world's most widely grown food crop so it is perhapsunsurprising that it should now be at the forefront of a new greenrevolution in which plants are used not only as food, feed orcommodities, but as advanced biochemical factories to produce thevaluable molecules we need as pharmaceuticals, industrial enzymes,nutrients, chemical precursors and fuels. The economic case for usingmaize has beenmade, and several case studies now show conclusivelythat it is a viable production platform for high-value molecules, evenwhen onerous regulations are taken into account. The regulatoryframework for plant-made pharmaceutical and industrial products isevolving (Spök et al., 2008) but it is imperative to base the emergingregulations on scientific principles and not risks that are poorlyunderstood and largely unquantified. We must avoid fearing the‘bogeyman under the bed’, and must instead study the impact of plantfactories rigorously and carefully. It has been suggested that a morepractical and productive approach might be to base risk assessmentson ecotoxicological principles, in which research problems refer torelevant assessment endpoints and risk hypotheses predict noadverse effects of transgenic plants on the assessment endpoints(Raybould, 2007). Maize presents a number of important advantagesas a production platform for high-value molecules, including thebenefits of its unique physiology, the ability to express recombinantproteins in seeds, the GRAS status, its widespread and well-developedinfrastructure, its genetic diversity, well-understood genetics and lackof allergenicity and toxicity. It can also be grown on all continents and,with the exception of Central America, has no co-localized weedyrelatives, and is thus ideal for adoption in local production settings inrural environments. The regulatory framework needs to be builtaround these solid scientific facts rather than media-fueled specula-tion, and we should focus on the benefits that transgenic maize canbring to both the industrial world and the poor in developingcountries, where the fruits of the technology are needed the most.
Acknowledgements
Research in our laboratory is supported by Ministry of Scienceand Innovation (MEI, Spain) Grant BFU2007-61413; the Ramon yCajal program (MICINN, Spain); the Pharma-Planta FP6 EU projectLSH-2002-1.2.5-2; Center Consolider, MICINN, Spain; Acciones Com-plementarias (MICINN, Spain) Grant BIO2005-24826; ERC AdvancedGrant BIOFORCE to PC; SN and GF are recipients of MICINN PhDFellowships.
References
Aitzetmuller K, Matthaus B, Friedrich H. A new database for seed oil fatty acids — thedatabase SOFA. Eur J Lipid Sci Technol 2003;105:92-103.
Alexander DJ. Newcastle disease and other avian paramixoviridae infections. In: CalnekBW, Barnes HJ, Beard CW, Reid WM, Yoder HW, editors. Diseases of poultry. Iowa:State University Press; 1997. p. 541–70.
Altpeter F, Baisakh N, Beachy R, Bock R, Capell T, Christou P, et al. Particle bombardmentand the genetic enhancement of crops: myths and realities. Mol Breed 2005;15:305–27.
Aluru M, Xu Y, Guo R, Wang Z, Li S, White W, et al. Generation of transgenic maize withenhanced provitamin A content. J Exp Bot 2008;59:3551–62.
Bailey R. Another inconvenient truth— how biofuel policies are deepening poverty andaccelerating climate change. Oxfam Briefing Paper. Oxfam International; 2008. 25June.
Bailey MR, Woodard SL, Callaway E, Beifuss K, Magallanes-Lundback M, Lane JR, et al.Improved recovery of active recombinant laccase from maize seed. Appl MicrobiolBiotechnol 2004;63:390–7.
Basaran P, Rodriguez-Cerezo E. Plant molecular farming: opportunities and challenges.Crit Rev Biotechnol 2008;28:153–72.
Belo A, Zheng P, Luck S, Shen B, Meyer DJ, Li B, et al. Whole genome scan detects anallelic variant of fad2 associated with increased oleic acid levels inmaize. Mol GenetGenomics 2008;279:1-10.
Bicar E, Woodman-Clikeman W, Sangtong V, Peterson J, Yang S, Lee M, Scott M.Transgenic maize endosperm containing a milk protein has improved amino acidbalance. Transgenic Res 2008;17:59–71.
Bouis HE. Micronutrient fortification of plants through plant breeding: can it improvenutrition in man at low cost? Proc Nutr Soc 2003;62:403–11.
Boyer CD, Preiss J. Multiple forms of starch branching enzyme of maize. Evidence forindependent genetic control. Biochem Biophys Res Commun 1978;80:169–75.
Broun P, Gettner S, Somerville C. Genetic engineering of plant lipids. Annu Rev Nutr1999;19:197–216.
Buleon A, Colonna P, Planchot V, Ball S. Starch granules: structure and biosynthesis. Int JBiol Macromol 1998;23:85-112.
Cahoon EB, Hall SE, Ripp KG, Ganzke TS, Hitz WD, Coughlan SJ. Metabolic redesign ofvitamin E biosynthesis in plants for tocotrienol production and increasedantioxidant content. Nat Biotechnol 2003;21:1082–7.
Caimi PG, McCole LM, Klein TM, Kerr PS. Fructan accumulation and sucrose metabolismin transgenic maize endosperm expressing a Bacillus amyloliquefaciens SacB gene.Plant Physiol 1996;110:355–63.
Capell T, Christou P. Progress in plant metabolic engineering. Curr Opin Biotechnol2004;15:148–54.
Chai XJ, Wang PW, Guan SY, Xu YW. Reducing the maize amylopectin content throughRNA interference manipulation. Zhi Wu Sheng Li Yu Fen Zi Sheng Wu Xue Xue Bao2005;31:625–30.
Chander S, Guo YQ, Yang XH, Yan JB, Zhang YR, Song TM, Li JS. Genetic dissection oftocopherol content and composition in maize grain using quantitative trait locianalysis and the candidate gene approach. Mol Breed 2008;22:353–65.
Chen Z, Young TE, Ling J, Chang SC, Gallie DR. Increasing vitamin C content of plantsthrough enhanced ascorbate recycling. Proc Natl Acad Sci USA 2003;100:3525–30.
Chen S, Li H, Liu G. Progress of vitamin E metabolic engineering in plants. TransgenicRes 2006;15:655–65.
Chintalacharuvu KA, Morrison SL. Production of secretory immunoglobulin A by asingle mammalian cell. Proc Natl Acad Sci USA 1997;94:6364–8.
Christensen JH, Christensen MS, Dyerberg J, Schmidt EB. Heart rate variability and fattyacid content of blood cell membranes: a dose response study with n-3 fatty acids.Am J Clin Nutr 1999;70:331–7.
Commandeur U, Twyman RM, Fischer R. The biosafety of molecular pharming in plants.AgBiotechNet 2003;5:110.
Creech RG. Genetic control of carbohydrate synthesis in maize. Genetics 1965;52:1175–86.
D'Halluin K, Bonne E, Bossut M, Beuckeleer M, Leemans J. Transgenic maize plants bytissue electroporation. Plant Cell 1992;4:1495–505.
Delaney DE. Choice of crop species and development of transgenic product lines. In:Hood EE, Howard JA, editors. Plants as factories for protein production. Dordrecht:Kluwer; 2002. p. 55–78.
Della Porta G, Ederle D, Bucchini L, Prandi M, Verderio A, Pozzi C. Maize pollen mediatedgene flow in the Po valley (Italy): source–recipient distance and effect of floweringtime. Eur J Agron 2008;28:255–65.
DellaPenna D. Nutritional genomics: manipulating plant micronutrients to improvehuman health. Science 1999;285:375–9.
DellaPenna D, Pogson BJ. Vitamin synthesis in plants: tocopherols and carotenoids.Annu Rev Plant Biol 2006;57:711–38.
Demurin Y, Skorie D, Karlovic D. Genetic variability of tocopherol composition insunflower seeds as a basis of breeding for improved oil quality. Plant Breed1996;115:33–6.
Diaz de la Garza RI, Quinlivan EP, Klaus SM, Basset GJ, Gregory III JF, Hanson AD. Folatebiofortification in tomatoes by engineering the pteridine branch of folate synthesis.Proc Natl Acad Sci USA 2004;101:13720–5.
Diaz de la Garza RI, Gregory III JF, Hanson AD. Folate biofortification of tomato fruit. ProcNatl Acad Sci USA 2007;104:4218–22.
Di-Giovanni F, Kevan PG. Factors affecting pollen dynamics and its importance to pollencontamination: a review. Can J For Res 1991;21:1155–70.
Di-Giovanni F, Kevan PG, Nasr ME. The variability in settling velocities of some pollenand spores. Grana 1995;34:39–44.
Drakakaki G, Marcel S, Glahn RP, Lund L, Periagh S, Fischer R, et al. Endosperm specificco-expression of recombinant soybean ferritin and Aspergillus phytase in maizeresults in significant increases in the levels of bioavailable iron. Plant Mol Biol2005;59:869–80.
Egesel CO, Wong JC, Lambert RJ, Rocheford TR. Combining ability of maize inbreds forcarotenoids and tocopherols. Crop Sci 2003;43:818–23.
Electronic Code of Federal Regulations. US Department of Agriculture Regulations: Title7 Agriculture. Part #201.76 Minimum land, isolation, field, and seed standards. USFederal Seed Act Regulations (Certified Seed); 2008. Available at http://ecfr.gpoaccess.gov Accessed 30th November 2008.
Ellis RP, Cochrane MP, Dale MFB, Duffus CM, Lynn A, Morrison IM, et al. Starchproduction and industrial use. J Sci Food Agric 1998;77:289–311.
Farinas CS, Leite A, Miranda EA. Aqueous extraction of maize endosperm: insights forrecombinant protein hosts based on downstream processing. Process Biochem2005;40:3327–36.
FergasonV.High amylose andwaxy corns. In: Hallauer AR, editor. Specialty corns. London:CRC Press; 1994. p. 55–7.
Fischer R, Stoger E, Schillberg S, Christou P, Twyman RM. Plant-based production ofbiopharmaceuticals. Curr Opin Plant Biol 2004;7:152–8.

52 S. Naqvi et al. / Biotechnology Advances 29 (2011) 40–53
Frame BR, Drayton PR, Bagnall SV, Lewnau CJ, Bullock WP, Frame HM, et al. Productionof fertile transgenic maize plants by silicon carbide whisker-mediated transfor-mation. Plant J 1994;6:941–8.
Frizzi A, Huang S, Gilbertson LA, Armstrong TA, Luethy MH, Malvar TM. Modifyinglysine biosynthesis and catabolism in corn with a single bifunctional expression/silencing transgene cassette. Plant Biotechnol J 2008;6:13–21.
FrommM, Taylor LP, Walbot V. Expression of genes transferred into monocot and dicotplant cells by electroporation. Proc Natl Acad Sci USA 1985;82:5824–8.
Galili G, Höfgen R. Metabolic engineering of amino acids and storage proteins in plants.Metab Eng 2002;4:3-11.
Gewin V. Genetically modified corn — environmental benefits and risks. PLoS Biol2003;1:15–9.
Giddings G, Allison G, Brooks D, Carter A. Transgenic plants as factories forbiopharmaceuticals. Nat Biotechnol 2000;18:1151–5.
Giuliano G, Tavazza R, Diretto G, Beyer P, Taylor MA. Metabolic engineering ofcarotenoid biosynthesis in plants. Trends Biotechnol 2008;26:139–45.
GolovkinM, AbrahamM,Morocz S, Bottka S, Feher A, Dudits D. Production of transgenicmaize plants by direct DNA uptake into embryogenic protoplasts. Plant Sci1993;90:41–52.
Gómez-Galera S, Rojas E, Sudhakar D, Zhu C, Pelacho AM, Capell T, Christou P. Criticalevaluation of strategies for mineral fortification of staple food crops. Transgenic Res2010;19:65–80.
Gordon-Kamm WJ, Spencer TM, Mangano ML, Adams TR, Daines RJ, Start WG, et al.Transformation of maize cells and regeneration of fertile transgenic plants. PlantCell 1990;2:603–18.
Grams GW, Blessin CW, Inglett GE. Distribution of tocopherols within the corn kernel.J Am Oil Chem Soc 1970;47:337–9.
Grusak MA, DellaPenna D. Improving the nutrient composition of plants to enhancehuman nutrition and health. Annu Rev Plant Physiol Plant Mol Biol 1999;50:133–61.
Guerrero-Andrade O, Loza-Rubio E, Olivera-Flores T, Fehervari-Bone T, Gomez-LimMA.Expression of the Newcastle disease virus fusion protein in transgenic maize andimmunological studies. Transgenic Res 2006;15:455–63.
Harjes CE, Rocheford TR, Bai L, Brutnell TP, Kandianis CB, Sowinski SG, et al. Naturalgenetic variation in lycopene epsilon cyclase tapped for maize biofortification.Science 2008;319:330–3.
Hood EE,Woodward SL. Commercialization of a protein product from transgenic maize.Agricultural biotechnology: beyond food and energy to health and the environ-ment, National Agricultural Biotechnology Council (NABC) Report 17; 2005.p. 147–58.
Hood EE, Witcher DR, Maddock S, Meyer T, Baszczynski C, Bailey M, et al. Commercialproduction of avidin from transgenic maize: characterization of transformant,production, processing, extraction and purification. Mol Breed 1997;3:291–306.
Hood EE, Bailey MR, Beifuss K, Magallanes-Lundback M, Horn ME, Callaway E, et al.Criteria for high-level expression of a fungal laccase gene in transgenic maize. PlantBiotechnol J 2003;1:129–40.
Houmard NM, Mainville JL, Bonin CP, Huang S, Luethy MH, Malvar TM. High-lysine corngenerated by endosperm-specific suppression of lysine catabolism using RNAi.Plant Biotechnol J 2007;5:605–14.
Huang S, Kruger DE, Frizzi A, D'Ordine RL, Florida CA, AdamsWR, et al. High-lysine cornproduced by the combination of enhanced lysine biosynthesis and reduced zeinaccumulation. Plant Biotechnol J 2005;3:555–69.
Huang S, Frizzi A, Florida C, Kriger D, Luethy M. High lysine and high tryptophantransgenic maize resulting from the reduction of both 19- and 22-kDα-zeins. PlantMol Biol 2006;61:525–35.
Ishida Y, Saito H, Ohta S, Hiei Y, Komari T, Kumashiro T. High efficiency transformationof maize (Zea mays L.) mediated by Agrobacterium tumefaciens. Nat Biotechnol1996;14:745–50.
Ishikawa T, Dowdle J, Smirnoff N. Progress in manipulating ascorbic acid biosynthesisand accumulation in plants. Physiol Plant 2006;126:343–55.
Islam, SN. MSc thesis. Survey of carotenoid variation and quantitative trait loci mappingfor carotenoid and tocopherol variation in maize. University of Illinois at Urbana-Champaign, 2004.
Jaffe G. Regulating transgenic crops: a comparative analysis of different regulatoryprocesses. Transgenic Res 2004;13:5-19.
Jako C, Kumar A, Wei Y, Zou J, Barton DL, Giblin EM, et al. Seed-specific over-expressionof an Arabidopsis cDNA encoding a diacylglycerol acyltransferase enhances seed oilcontent and seed weight. Plant Physiol 2001;126:861–74.
James C. Global status of commercialized biotech/GM crops: 2009. ISAAA Brief No. 41.Ithaca NY: ISAAA; 2009.
Johnson LA, Hardy CL, Baumel CP, White PJ. Identifying valuable corn quality traits forstarch production. Cereal Foods World 2001a;46:417–23.
Johnson LA, Hardy CL, Baumel CP, Yu TH, Sell JL. Identifying valuable corn quality traitsfor livestock feed. Cereal Foods World 2001b;46:472–81.
Jones MD, Brooks JS. Effectiveness of distance and border rows in preventingoutcrossing in corn. Okla Agr Exp Sta Techn Bul 1950:T-38.
Jung R, Carl F. Transgenic corn with an improved amino acid composition. In: Müntz K,Ulrich W, editors. 8th International Symposium on Plant Seeds. Gatersleben,Germany: Institute of Plant Genetics and Crop Plant Research; 2000.
Karunanandaa B, Qi Q, Hao M, Baszis SR, Jensen PK, Wong YH, et al. Metabolicallyengineered oilseed crops with enhanced seed tocopherol. Metab Eng 2005;7:384–400.
Kinney AJ, Cahoon EB, Hitz WD. Manipulating desaturase activities in transgenic cropplants. Biochem Soc Trans 2002;30:1099–103.
Klein TM, Gradziel T, Fromm ME, Sanford JC. Factors influencing gene delivery into Zeamays cells by high-velocity microprojectiles. Bio/Technol 1988;6:559–63.
Konig A, Cockburn A, Crevel RW, Debruyne E, et al. Assessment of the safety of foodderived from genetically modified GM crops. Food Chem Toxicol 2004;42:1047–88.
Kurilich AC, Juvik JA. Quantification of carotenoid and tocopherol antioxidants in Zeamays. J Agr Food Chem 1999;47:1948–55.
Luna VS, Figueroa JM, Baltazar BM, Gomez RL, Townsend R, Schoper JB. Maize pollenlongevity and distance isolation requirements for effective pollen control. Crop Sci2001;41:1551–7.
Lung SC, Weselake RJ. Diacylglycerol acyltransferase: a key mediator of planttriacylglycerol synthesis. Lipids 2006;41:1073–86.
Ma JK, Drake PM, Christou P. The production of recombinant pharmaceutical proteins inplants. Nat Rev Genet 2003;4:794–805.
Ma JK, Barros E, Bock R, Christou P, Dale PJ, Dix PJ, et al. The production ofpharmaceutical proteins in transgenic plants— current perspectives. EMBO Reports2005;6:593–9.
Manners DJ. The enzymic degradation of starches. In: Blanshard JMV, Mitchell JR,editors. Polysaccharides in food. London: Butterworths; 1979. p. 75–91.
Martindale W, Trewavas A. Fuelling the 9 billion. Nat Biotechnol 2008;26:1068–70.Melé E, Ballester J, Peñas G, Folch I, Olivar J, Alcalde E, Messeguer J. First results of co-
existence study. Eur Biotechnol Sci Indust News 2004;3:8.Menne E, Guggenbuhl N, Roberfroid MB. Fn-type chicory inulin hydrolysate has a
prebiotic effect in humans. J Nutr 2000;130:1197–9.Mertz ET, Bates LS, Nelson OE. Mutant gene that changes protein composition for grain
quality traits. Science 1964;258:372–5.Messeguer J, Peñas G, Ballester J, Bas M, Serra J, et al. Pollen-mediated gene flow in
maize in real situations of coexistence. Plant Biotechnol J 2006;4:633–45.Mitchell D. A note on rising food prices. World Bank policy research working paper
4682. Washington DC: World Bank; 2008.Nakamura Y, Kubo A, Shimamune T, Matsuda T, Harada K, Satoh H. Correlation between
activities of starch debranching enzyme andα-polyglucan structure in endospermsof sugary-1 mutants of rice. Plant J 1997;12:143–53.
Napier JA, Haslam R, Caleron MV, Michaelson LV, Beaudoin F, Sayanova O. Progresstowards the production of very long-chain polyunsaturated fatty acid in transgenicplants: plant metabolic engineering comes of age. Physiol Plant 2006;126:398–406.
Naqvi S, Farre G, Sanahuja G, Capell T, Zhu C, Christou P. When more is better:multigene engineering in plants. Trends Plants Sci 2009a;15:48–56.
Naqvi S, Zhu C, Farre G, Ramessar K, Bassie L, Breitenbach J, Conesa D, Ros G, SandmannG, Capell T, Christou P. Transgenic multivitamin corn through biofortification ofendosperm with three vitamins representing three distinct metabolic pathways.Proc Natl Acad Sci USA 2009b;106:7762–7.
Naqvi S, Farré Zhu C, Sandmann G, Capell C, Christou C. Simultaneous expression ofArabidopsis ρ-hydroxyphenylpyruvate dioxygenase and MPBQ methyltransferasein transgenic corn kernels triples the tocopherol content. Transgenic Res 2010,doi:10.1007/s11248-010-9393-6.
NCGA National Corn Growers Association. U.S. corn growers: producing food and fuel.Chesterfield,MO:NCGA;2008. http://www.ncga.com/files/pdf/FoodandFuelPaper10-08.pdf.
NCGA National Corn Growers Association. Meeting the challenge of production; 2010.http://www.ncga.com/files/pdf/WOC2010.pdf.
Nicholson L, Gonzalez-Melendi L, van Dolleweerd C, Tuck H, Perrin Y, Ma JKC, et al. Arecombinant multimeric immunoglobulin expressed in rice shows assembly-dependent subcellular localization in endosperm cells. Plant Biotech J 2005;3:115–27.
OECD. Biofuel support policies: an economic assessment. Paris: OECD; 2008.Palaudelmàs M, Melé E, Peñas G, Pla M, Nadal A, et al. Sowing and flowering delays can
be an efficient strategy to improve coexistence of genetically modified andconventional maize. Crop Sci 2008;48:2404–13.
Paterniani E, Stort AC. Effective maize pollen dispersal in the field. Euphytica 1974;23:129–34.
Perrin Y, Vaquero C, Gerrard I, SackM, Drossard J, Stoger E, et al. Transgenic pea seeds asbioreactors for the production of a single-chain Fv fragment (scFV) antibody usedin cancer diagnosis and therapy. Mol Breed 2000;6:345–52.
Petolino JF, Hopkins NL, Kosegi BD, Skokut M. Whisker-mediated transformation ofembryogenic callus of maize. Plant Cell Rep 2000;19:781–6.
PIFB. Pharming the field: a look at the benefits and risks of bioengineering plants toproduce pharmaceuticals.Washington DC: Pew Initiative on Food and Biotechnology;2003.
Platis D, Labrou NE. Affinity chromatography for the purification of therapeutic proteinsfrom transgenic maize using immobilized histamine. J Sep Sci 2008;31:636–45.
Prasanna BM, Vasal SK, Singh NN. Quality protein maize. Curr Sci 2001;81:1308–19.Provencher LM, Miao L, Sinha N, Lucas WJ. Sucrose export defective1 encodes a novel
protein implicated in chloroplast-to-nucleus signalling. Plant Cell 2001;13:1127–41.
Rademacher T, Sack M, Arcalis E, Stadlmann J, et al. Recombinant antibody 2G12produced in maize endosperm efficiently neutralizes HIV-1 and containspredominantly single-GlcNAc N-glycans. Plant J 2008;6:189–201.
Ramessar K, Sabalza M, Capell T, Christou P. Maize plants: an ideal production platformfor effective and safe molecular pharming. Plant Sci 2008a;174:409–19.
Ramessar K, Rademacher T, Sack M, Stadlmann J, Platis D, Stiegler G, et al. Cost-effectiveproduction of a vaginal protein microbicide to prevent HIV transmission. Proc NatlAcad Sci USA 2008b;105:3727–32.
Ramessar K, Capell T, Christou P. Molecular pharming in cereal crops. Phytochem Rev2008c;7:579–92.
Ramessar K, Capell T, Twyman RM, Quemada H, Christou P. Calling the tunes ontransgenic crops: the case for regulatory harmony. Mol Breed 2008d;23:99-112.
Ramessar K, Capell T, Twyman RM, Quemada H, Christou P. Trace and traceability — acall for regulatory harmony. Nat Biotechnol 2009;26:975–8.

53S. Naqvi et al. / Biotechnology Advances 29 (2011) 40–53
Ramessar K, Capell T, Twyman RM, Christou P. Going to ridiculous lengths — Europeancoexistence regulations for GM crops. Nat Biotechnol 2010;28:133–6.
Raybould A. Ecological versus ecotoxicological methods for assessing the environmen-tal risks of transgenic crops. Plant Sci 2007;173:589–602.
Raynor GS, Eugene CO, Janet VH. Dispersion and deposition of corn pollen fromexperimental sources. Agron J 1972;64:420–7.
Reiffel JA, McDonald A. Anti-arrhythmic effects of omega-3 fatty acids. Am J Cardiol2006;98:50i–60i.
Rhodes CA, Pierce DA, Mettler IJ, Mascarenhas D, Detmer JJ. Genetically transformedmaize plants from protoplasts. Science 1988;240:204–7.
Roberfroid MB. Chicory fructooligosaccharides and the gastrointestinal tract. Nutrition2000;16:677–9.
Sattler SE, Cahoon EB, Coughlan SJ, DellaPenna D. Characterization of tocopherolcyclases from higher plants and cyanobacteria. Evolutionary implications fortocopherol synthesis and function. Plant Physiol 2003;132:2184–95.
Savidge B, Weiss JD, Wong YH, Lassner MW, Mitsky TA, Shewmaker CK, et al. Isolationand characterization of homogentisate phytyltransferase genes from Synechocystissp. PCC 6803 and Arabidopsis. Plant Physiol 2002;129:321–32.
Schillberg S, Emans N, Fischer R. Antibodymolecular pharming in plants and plant cells.Phytochem Rev 2002;1:45–54.
Scott J, Rébeillé F, Fletcher J. Folic acid and folates: the feasibility for nutritionalenhancement in plant foods. J Sci Food Agric 2000;80:795–824.
Seal BS, King DJ, Sellers HL. The avian response to Newcastle disease virus. DevelopComp Immunol 2000;24:257–68.
Segal G, Song R, Messing J. A new opaque variant of maize by a single dominant RNA-interference-inducing transgene. Genetics 2003;165:387–97.
Shannon JC, Pien FM, Cao H, Liu KC. Brittle-1, an adenylate translocator, facilitatestransfer of extraplastidial synthesized ADP-glucose into amyloplasts. Plant Physiol1998;117:1235–52.
Shure M, Wessler S, Fedoroff N. Molecular identification and isolation of thewaxy locusin maize. Cell 1983;35:225–33.
Smuts CM, Huang M, Mundy D, Plasse T, Major S, Carlson SE. A randomized trial ofdocosahexaenoic acid supplementation during the third trimester of pregnancy.Obstet Gynecol 2003;101:469–79.
Sparrow PAC, Irwin JA, Dale PJ, Twyman RM, Ma JKC. Pharma-Planta: road testing thedeveloping regulatory guidelines for plant-made pharmaceuticals. Transgenic Res2007;16:147–61.
Spök A, Twyman RM, Fischer R, Ma JKC, Sparrow PAC. Evolution of a regulatoryframework for plant-made pharmaceuticals. Trends Biotechnol 2008;26:506–17.
Stoger E, Vaquero C, Torres E, Sack M, Nicholson L, Drossard J, et al. Cereal crops asviable production and storage systems for pharmaceutical scFv antibodies. PlantMol Biol 2000;42:583–90.
Stoger E, Sack M, Fischer R, Christou P. Plantibodies: applications, advantages andbottlenecks. Curr Opin Biotechnol 2002a;13:161–6.
Stoger E, Sack M, Perrin Y, Vaquero C, Torres E, Twyman RM, et al. Practicalconsiderations for pharmaceutical antibody production in different crop systems.Mol Breed 2002b;9:149–58.
Stoger E, Sack M, Nicholson L, Fischer R, Christou P. Recent progress in plantibodytechnology. J Curr Pharmaceut Design 2005a;11:2439–57.
Stoger E, Ma JK-C, Fischer R, Christou P. Sowing the seeds of success: pharmaceuticalproteins from plants. Curr Opin Biotechnol 2005b;16:167–73.
Storozhenko S, De Brouwer V, Volckaert M, Navarrete O, Blancquaert D, Zhang GF, et al.Folate fortification of rice by metabolic engineering. Nat Biotechnol 2007;25:1277–9.
Streatfield SJ. Approaches to achieve high-level heterologous protein production inplants. Plant Biotechnol J 2007;5:2-15.
Streatfield S, Howard J. Plant based vaccines. Int J Parisitol 2003;33:5–6.Tada Y, Utsumi S, Takaiwa F. Foreign gene products can be enhanced by introduction
into storage protein mutants. Plant Biotechnol J 2003;1:411–22.Tan B. Palm carotenoids, tocopherols and tocotrienols. J Am Oil Chemists Soc 1989;66:
770–6.
Torres E, Vaquero C, Nicholson L, Sack M, Stöger E, Drossard J, Christou P, Fischer R,Perrin Y. Rice cell culture as an alternative production system for functionaldiagnostic and therapeutic antibodies. Transgenic Res 1999;8:441–9.
Traber M, Sies H. Vitamin E in humans: demand and delivery. Annu Rev Nutr 1996;16:321–47.
Trostle R. Global agricultural supply and demand: factors contributing to the recentincrease in food commodity prices. USDA Economic Research Service OutlookReport WRS-0801. Washington DC: USDA; 2008.
Truksa M, Vrinten P, Qiu X. Metabolic engineering of plants for polyunsaturated fattyacid production. Mol Breed 2009;23:1-11.
Twyman RM, Stoger E, Schillberg S, Christou P, Fischer R. Molecular farming in plants:host systems and expression technology. Trends Biotechnol 2003;21:570–8.
Twyman RM, Schillberg S, Fischer R. Transgenic plants in the biopharmaceuticalmarket. Expert Opin Emerg Drugs 2005;10:185–218.
USDA-APHIS. Field testing and plants engineered to produce pharmaceutical andindustrial compounds. Fed Regis 2003;68.
USDA-FAS. World corn production, consumption and stocks report. Washington DC:UDSA; 2008.
Verpoorte R, van der Heijden R, Memelink J. Engineering the plant cell factory forsecondary metabolite production. Transgenic Res 2000;9:323–43.
Wang YJ, White P, Pollak L, Jane JL. Characterization of starch structures of 17 maizeendosperm mutant genotypes with Oh43 inbred line background. Cereal Chem1993;70:171–9.
Watson SA. Structure and composition. In: Watson SA, Ramstad PT, editors. Corn:chemistry and technology. St. Paul, MN: American Association of Cereal Chemists;1987. p. 53–82.
Wolt JD, Wang K, Peterson RKD. Assessing risk of unintended antigen occurrence infood: a case instance for maize-expressed LT-B. Hum Ecol Risk Assess 2006;12:1-15.
Woodard S, Mayor J, Bailey M, Barker D, Love R, Lane J, et al. Maize (Zea mays)-derivedbovine trypsin: characterization of the first large-scale, commercial protein productfrom transgenic plants. Biotechnol Appl Biochem 2003;38:123–30.
Wu X, Kenzior A, Willmot D, Scanlon S, Chen Z, Topin A, et al. Altered expression ofplant lysyl tRNA synthetase promotes tRNA misacylation and translation recodingof lysine. Plant J 2007;50:627–63.
Yang S, Moran D, Jia H, Bicar E, Lee M, Scott M. Expression of a synthetic porcine α-lactoalbumin gene in the kernels of transgenic maize. Transgenic Res 2002;11:11–20.
Yu J, Peng P, Zhang X, Zhao Q, Zhy D, Sun X, et al. Seed-specific expression of a lysinerich protein sb401 gene significantly increases both lysine and total protein contentin maize seeds. Mol Breed 2004;14:1–7.
Yusibov V, Rabindran S. Recent progress in the development of plant derived vaccines.Expert Rev Vaccines 2008;7:1173–83.
Zarkadas C, Hamilton R, Yu Z, Choi V, Khanizadeh S, Rose N, Pattison P. Assessment ofthe protein quality of 15 new northern adapted cultivars of quality protein maizeusing amino acid analysis. J Agric Food Chem 2000;48:5351–61.
Zhang C, Meng X, Halbur P. Development of edible vaccines against PRRSV: a proof ofconcept study. National Pork Board; 2006.
Zhang S, Dong JG, Wang T, Guo S, Glassman K, Ranch J, Nichols SE. High levelaccumulation of alpha-glucan in maize kernels by expressing the gtfD gene fromStreptococcus mutans. Transgenic Res 2007;16:467–8.
Zheng P, Allen WB, Roesler K, Williams ME, Zhang S, Li J, et al. A phenylalanine in DGATis a key determinant of oil content and composition in maize. Nat Genet 2008;40:367–72.
Zhong G, Peterson D, Delaney D, Bailey M, Witcher D, Register III J, et al. Commercialproduction of aprotinin in transgenic maize seeds. Mol Breed 1999;5:345–56.
Zhu C, Naqvi S, Gomez-Galera S, Pelacho AM, Capell T, Christou P. Transgenic strategiesfor the nutritional enhancement of plants. Trends Plant Sci 2007;12:548–55.
Zhu C, Naqvi S, Breitenbach J, Sandmann G, Christou P, Capell T. Combinatorial genetictransformation generates a library of metabolic phenotypes for the carotenoidpathway in maize. Proc Natl Acad Sci USA 2008;105:18232–7.




















