
Histidine-mediated xylem loading of zinc is a species-wide character in Noccaea caerulescens
12
Histidine-mediated xylem loading of zinc is a species-wide character in Noccaea caerulescens Anna D. Kozhevnikova 1 , Ilya V. Seregin 1 , Nadezhda T. Erlikh 1 , Taisiya A. Shevyreva 2 , Igor M. Andreev 2 , Rudo Verweij 3 and Henk Schat 4 1 Laboratory of Root Physiology, Timiryazev Institute of Plant Physiology, Russian Academy of Sciences, Botanicheskaya str. 35, 127276 Moscow, Russia; 2 Laboratory of Plant Cell Membranes, Timiryazev Institute of Plant Physiology, Russian Academy of Sciences, Botanicheskaya str. 35, 127276 Moscow, Russia; 3 Department of Animal Ecology, Faculty of Earth and Life Sciences, Vrije Universiteit Amsterdam, de Boelelaan 1085, 1081 HV Amsterdam, the Netherlands; 4 Department of Genetics, Faculty of Earth and Life Sciences, Vrije Universiteit Amsterdam, de Boelelaan 1085, 1081 HV Amsterdam, the Netherlands Author for correspondence: Anna D. Kozhevnikova Tel: +7 499 231 83 24 Email: [email protected] Received: 17 February 2014 Accepted: 17 March 2014 New Phytologist (2014) doi: 10.1111/nph.12816 Key words: histidine, hyperaccumulation, Noccaea caerulescens, Thlaspi arvense, tissue distribution, xylem loading, zinc (Zn). Summary Histidine plays a crucial role in nickel (Ni) translocation in Ni-hyperaccumulating plants. Here, we investigated its role in zinc (Zn) translocation in four accessions of the Zn hyperaccu- mulator, Noccaea caerulescens, using the related non-hyperaccumulator, Thlaspi arvense, as a reference. We compared the effects of exogenous histidine supply on Zn xylem loading, and of Zn– histidine complex formation on Zn uptake in energized tonoplast vesicles. The Zn distribution patterns over root tissues were also compared. Exogenous histidine supply enhanced Zn xylem loading in all the N. caerulescens accessions, but decreased it in T. arvense. Zn distribution patterns over root tissues were similar, apart from the accumulation in cortical and endodermal cells, which was much lower in N. caerulescens than in T. arvense. Zn uptake in energized tonoplast vesicles was inhibited sig- nificantly in N. caerulescens, but not affected significantly in T. arvense, when Zn was supplied in combination with histidine in a 1 : 2 molar ratio. Histidine-mediated Zn xylem loading seems to be a species-wide character in N. caerulescens. It may well have evolved as a component trait of the hyperaccumulation machinery for Zn, rather than for Ni. Introduction A small minority of vascular plant species accumulate heavy met- als at extremely high, normally toxic, concentrations in their foliage. These so-called hyperaccumulators are characterized by high rates of metal root to shoot translocation, typically resulting in shoot to root metal concentration ratio’s above unity, which are, for most metals, c. 10-fold higher than in normal plants (Brooks et al., 1977; Baker et al., 2000). The mechanisms under- lying these strongly enhanced translocation rates are incompletely known to date. The superior rate of zinc (Zn) root to shoot trans- location in the Zn/cadmium (Cd) hyperaccumulator, Arabidopsis halleri, in comparison with A. thaliana, was completely abolished by RNAi-mediated silencing, down to the A. thaliana expression level, of HMA4 (Hanikenne et al., 2008), which encodes a 1b P-type heavy metal-transporting ATPase previously shown to be responsible for the major part of Zn and Cd xylem loading in A. thaliana (Hussain et al., 2004; Wong & Cobbett, 2009). However, the expression of hyperaccumulator HMA4 cDNAs from their endogenous promoters in A. thaliana or tomato did not yield considerable Zn or Cd translocation phenotypes, although it strongly enhanced the gene’s expression level, in comparison with wild-type A. thaliana (Hanikenne et al., 2008; Barabasz et al., 2012; Iqbal et al., 2013). These results suggest that enhanced HMA4 expression is essential, but probably not sufficient, for hyperaccumulator-like metal translocation rates. In other words, the effect of enhanced HMA4 expression may depend on whether the gene functions in a hyperaccumulator or a non-hyperaccumulator background. The genetic determinants of the apparent background depen- dence of the effects of enhanced HMA4 expression on heavy metal translocation have been elusive to date, but there is circum- stantial evidence that such factors may be involved in the control of metal retention in root cell vacuoles. It is generally assumed that vacuolar storage in root cells will decrease a metal’s availabil- ity for radial transport to the root stele and its subsequent loading into the xylem. There are a number of reported cases of decreased metal accumulation in root vacuoles in hyperaccumulators in comparison with non-hyperaccumulators. Using compartmental flux analysis, Lasat et al. (1998) showed that the fraction of absorbed 65 Zn that was stored in root vacuoles was 2.4-fold higher in the non-hyperaccumulator Thlaspi arvense than in the related Zn/Cd/nickel (Ni) hyperaccumulator Noccaea caerulescens (formerly Thlaspi caerulescens). Moreover, the rate of 65 Zn efflux Ó 2014 The Authors New Phytologist Ó 2014 New Phytologist Trust New Phytologist (2014) 1 www.newphytologist.com Research
Transcript of Histidine-mediated xylem loading of zinc is a species-wide character in Noccaea caerulescens

Histidine-mediated xylem loading of zinc is a species-widecharacter in Noccaea caerulescens
Anna D. Kozhevnikova1, Ilya V. Seregin1, Nadezhda T. Erlikh1, Taisiya A. Shevyreva2, Igor M. Andreev2,
Rudo Verweij3 and Henk Schat4
1Laboratory of Root Physiology, Timiryazev Institute of Plant Physiology, Russian Academy of Sciences, Botanicheskaya str. 35, 127276 Moscow, Russia; 2Laboratory of Plant Cell Membranes,
Timiryazev Institute of Plant Physiology, Russian Academy of Sciences, Botanicheskaya str. 35, 127276 Moscow, Russia; 3Department of Animal Ecology, Faculty of Earth and Life Sciences,
Vrije Universiteit Amsterdam, de Boelelaan 1085, 1081 HV Amsterdam, the Netherlands; 4Department of Genetics, Faculty of Earth and Life Sciences, Vrije Universiteit Amsterdam,
de Boelelaan 1085, 1081 HV Amsterdam, the Netherlands
Author for correspondence:Anna D. Kozhevnikova
Tel: +7 499 231 83 24Email: [email protected]
Received: 17 February 2014
Accepted: 17 March 2014
New Phytologist (2014)doi: 10.1111/nph.12816
Key words: histidine, hyperaccumulation,Noccaea caerulescens, Thlaspi arvense, tissuedistribution, xylem loading, zinc (Zn).
Summary
� Histidine plays a crucial role in nickel (Ni) translocation in Ni-hyperaccumulating plants.
Here, we investigated its role in zinc (Zn) translocation in four accessions of the Zn hyperaccu-
mulator, Noccaea caerulescens, using the related non-hyperaccumulator, Thlaspi arvense, as
a reference.� We compared the effects of exogenous histidine supply on Zn xylem loading, and of Zn–
histidine complex formation on Zn uptake in energized tonoplast vesicles. The Zn distribution
patterns over root tissues were also compared.� Exogenous histidine supply enhanced Zn xylem loading in all the N. caerulescens accessions,
but decreased it in T. arvense. Zn distribution patterns over root tissues were similar, apart
from the accumulation in cortical and endodermal cells, which was much lower in
N. caerulescens than in T. arvense. Zn uptake in energized tonoplast vesicles was inhibited sig-
nificantly in N. caerulescens, but not affected significantly in T. arvense, when Zn was supplied
in combination with histidine in a 1 : 2 molar ratio.� Histidine-mediated Zn xylem loading seems to be a species-wide character in
N. caerulescens. It may well have evolved as a component trait of the hyperaccumulation
machinery for Zn, rather than for Ni.
Introduction
A small minority of vascular plant species accumulate heavy met-als at extremely high, normally toxic, concentrations in theirfoliage. These so-called hyperaccumulators are characterized byhigh rates of metal root to shoot translocation, typically resultingin shoot to root metal concentration ratio’s above unity, whichare, for most metals, c. 10-fold higher than in normal plants(Brooks et al., 1977; Baker et al., 2000). The mechanisms under-lying these strongly enhanced translocation rates are incompletelyknown to date. The superior rate of zinc (Zn) root to shoot trans-location in the Zn/cadmium (Cd) hyperaccumulator, Arabidopsishalleri, in comparison with A. thaliana, was completely abolishedby RNAi-mediated silencing, down to the A. thaliana expressionlevel, of HMA4 (Hanikenne et al., 2008), which encodes a 1bP-type heavy metal-transporting ATPase previously shown to beresponsible for the major part of Zn and Cd xylem loading inA. thaliana (Hussain et al., 2004; Wong & Cobbett, 2009).However, the expression of hyperaccumulator HMA4 cDNAsfrom their endogenous promoters in A. thaliana or tomato didnot yield considerable Zn or Cd translocation phenotypes,although it strongly enhanced the gene’s expression level, in
comparison with wild-type A. thaliana (Hanikenne et al., 2008;Barabasz et al., 2012; Iqbal et al., 2013). These results suggestthat enhanced HMA4 expression is essential, but probably notsufficient, for hyperaccumulator-like metal translocation rates. Inother words, the effect of enhanced HMA4 expression maydepend on whether the gene functions in a hyperaccumulator ora non-hyperaccumulator background.
The genetic determinants of the apparent background depen-dence of the effects of enhanced HMA4 expression on heavymetal translocation have been elusive to date, but there is circum-stantial evidence that such factors may be involved in the controlof metal retention in root cell vacuoles. It is generally assumedthat vacuolar storage in root cells will decrease a metal’s availabil-ity for radial transport to the root stele and its subsequent loadinginto the xylem. There are a number of reported cases of decreasedmetal accumulation in root vacuoles in hyperaccumulators incomparison with non-hyperaccumulators. Using compartmentalflux analysis, Lasat et al. (1998) showed that the fraction ofabsorbed 65Zn that was stored in root vacuoles was 2.4-foldhigher in the non-hyperaccumulator Thlaspi arvense than in therelated Zn/Cd/nickel (Ni) hyperaccumulator Noccaea caerulescens(formerly Thlaspi caerulescens). Moreover, the rate of 65Zn efflux
� 2014 The Authors
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014) 1
www.newphytologist.com
Research

from the root vacuolar compartment was approximately two-foldhigher in N. caerulescens than in T. arvense. Yang et al. (2006)found a 2.7-fold higher root vacuolar 65Zn fraction in the non-hyperaccumulating ecotype of Sedum alfredii than in the hyperac-cumulating one. Xing et al. (2008), comparing twoN. caerulescens accessions with contrasting rates of Cd and Znroot to shoot translocation, observed a much higher fraction ofvacuolar 109Cd in the accession with the lowest translocation rate,although the half-lives for vacuolar efflux were no different. It isremarkable that the levels of HMA4 expression found in the latterstudy were almost the same in both accessions, suggesting that atleast part of the variation in translocation efficiency amonghyperaccumulator species or accessions may be determined byvariation in the root vacuolar storage capacity, rather than HMA4expression. In addition, Richau et al. (2009) compared Ni accu-mulation in vitro in energized tonoplast vesicles isolated fromN. caerulescens and T. arvense. In the case of T. arvense, theyfound much faster accumulation in root-derived vesicles than inshoot-derived ones, but the opposite was found in the case ofN. caerulescens, irrespective of the speciation of Ni in the bathingsolution. Moreover, Ni accumulation in root-derived vesicles washigher in T. arvense than in N. caerulescens, but the opposite wasfound for shoot-derived vesicles, except when Ni was supplied asan Ni–histidine (Ni–His) complex. These results suggest thatorgan-specific alterations of tonoplast metal transport capacitiesprobably have played a role in the evolution of thehyperaccumulation syndrome.
There are strong indications that differences in tonoplast metaltransport capacities between hyperaccumulators and non-hyper-accumulators are dependent on a metal’s speciation in the cyto-plasm. For example, when Ni was supplied as a 1 : 1 Ni–citratecomplex, root-derived tonoplast vesicles of T. arvense andN. caerulescens accumulated Ni at comparable rates, but, when Niwas supplied as NiSO4 or as an Ni–His (1 : 1) complex, vesiclesfrom T. arvense accumulated c. three-fold or seven-fold more Ni,respectively, than those from N. caerulescens (Richau et al., 2009).This may be taken to suggest that the strongly enhanced constitu-tive free His concentrations in the roots of N. caerulescens (c. 10-fold higher than in T. arvense) (Richau et al., 2009) and in the Nihyperaccumulator Alyssum lesbiacum (c. five-fold higher than innon-hyperaccumulators) (Kerkeb & Kr€amer, 2003) serve toprevent the vacuolar sequestration of Ni in root cells, thus keep-ing it available for loading into the xylem (Richau et al., 2009).However, although it seems that Ni may well be bound to His inhyperaccumulator roots, such as demonstrated for Zn (Salt et al.,1999), there is no evidence, as yet, concerning the subcellularlocalization of this complex. Regardless of the underlying mecha-nism, an exogenous His supply enhanced significantly Ni xylemloading in N. caerulescens, particularly under high Ni exposure,and more so in high-Ni-accumulating accessions than in low-Ni-accumulating ones (Richau et al., 2009). In non-hyperaccu-mulators, the reported effects of an exogenous His supply andtransgenic upregulation of the His biosynthetic pathway arecontradictory. An exogenous His supply was without effect, oreven decreased Ni xylem loading in T. arvense, as expected(Richau et al., 2009), but strongly increased it in Alyssum
montanum (Kr€amer et al., 1996) and Brassica juncea (Kerkeb &Kr€amer, 2003). Transgenic upregulation of His biosynthesisenhanced the free His concentrations in roots and shoots, but didnot affect Ni root to shoot translocation in A. thaliana (Wycisket al., 2004). It is conceivable that these contradictory resultsmight reflect variation amongst non-hyperaccumulators withregard to their capacity to transport the Ni–His complex acrossthe tonoplast, but direct evidence is lacking to date (see earlier).
In most of the studies available to date, enhanced cellular Hisconcentrations have been specifically associated with the toleranceand root to shoot transport of Ni, usually in naturally Ni-hyper-accumulating species or accessions from serpentine soil (Kr€ameret al., 1996; Persans et al., 1999; Kerkeb & Kr€amer, 2003;Wycisk et al., 2004; Ingle et al., 2005; Callahan et al., 2008). InN. caerulescens, however, natural Ni hyperaccumulation is con-fined to a relatively small number of local serpentine accessions,whereas His-dependent Ni xylem loading and high root His con-centrations seem to be species-wide properties (Richau et al.,2009). Therefore, it is unlikely, at least for N. caerulescens, thatHis would exclusively function in the hyperaccumulation of Ni.As N. caerulescens is primarily a hyperaccumulator of Zn, it isplausible to suppose that His also functions in the Zn hyperaccu-mulation machinery. In the present study, we tested the hypothe-sis that His-mediated xylem loading of Zn is a species-wideproperty in N. caerulescens. To this end, we compared the effectsof an exogenous His supply on Zn xylem loading in fourN. caerulescens accessions, using T. arvense as a non-hyperaccu-mulator reference. To check for potential effects of differentialZn loading efficiencies on the root-internal accumulation pat-terns, we also compared the Zn distribution over tissues and celltypes in roots using histochemical techniques. To establish theeffect of Zn speciation on its vacuolar sequestration, Zn uptakewas compared in MgATP-energized purified root- and shoot-derived tonoplast vesicles, with Zn supplied as a Zn–His complex(1 : 1), Zn–His complex (1 : 2) or as a sulfate salt.
Materials and Methods
Plant material and growth conditions
Seeds were collected from four accessions of Noccaea caerulescensF.K. Mey (formerly Thlaspi caerulescens J. & C. Presl), two origi-nating from Zn/Cd/lead (Pb)-enriched calamine soil at La Cala-mine, Belgium (LC) and Saint F�elix de Pailli�eres, France (SF),one from Ni-enriched serpentine soil at Monte Prinzera, Italy(MP), and one from a non-metalliferous site near Lellingen, Lux-embourg (LE) (Assunc�~ao et al., 2003). Thlaspi arvense L. seedswere collected from a non-metalliferous site near Amsterdam, theNetherlands. The seeds were sown on moist vermiculite, and2-wk-old seedlings were transferred to 1-l polyethylene pots(three seedlings per pot) filled with modified half-strength Hoa-gland’s nutrient solution (Assunc�~ao et al., 2003). Zn was addedat appropriate concentrations as Zn(NO3)2. The solutions wererenewed weekly. The plants were grown in a climate chamber(21°C : 18°C day : night; light intensity at plant level,200 lmol m�2 s�1; 14 h d�1; relative humidity (RH), 70%).
New Phytologist (2014) � 2014 The Authors
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist2

Experimental procedures
The effect of exogenous His on Zn xylem loading To assess theeffect of exogenous His supply on Zn xylem loading, 8-wk-oldplants, grown at 2 lM Zn in the nutrient solution, were placedfor 4 h with their roots in 1 mM L-His in a 2-mM Mes/KOHbuffer (pH 5.5), always at noon. A Mes/KOH-buffered 1-mML-alanine solution and Mes/KOH-buffered demineralized water(pH 5.5) were used as controls. After treatment, the root systemswere rinsed in demineralized water, and the leaf rosettes were cutoff with a razor blade, just below the lowest leaf. Then, the rootsystems were transferred to a fresh nutrient solution amendedwith 5, 50 or, only for N. caerulescens, 500 lM Zn(NO3)2. Rootpressure exudates were collected overnight in 2-ml vials con-nected with the excised stems via silicon tubing. The exudateswere collected at 10:00 h, their volumes were measured and thesamples were frozen at �20°C until analysis. The roots weredesorbed in Na2-EDTA (20 mM) for 10 min, rinsed in deminer-alized water and dried superficially with filter paper. They werethen dried in a stove at 80°C for 24 h, weighed and digested asdescribed later under ‘Measurement of Zn and His’.
An additional experiment was executed in order to checkwhether pretreatment with His primarily affected the loading ofnewly taken up Zn or (also) of the Zn burden that was alreadyaccumulated before the pretreatment. To that purpose, plantswere grown in nutrient solution containing 5 lM Zn(NO3)2(T. arvense andN. caerulescens, SF accession) or 50 lMZn(NO3)2(only N. caerulescens, SF accession) for 8 wk. Half were then pre-treated with 1 mM L-His in a 2-mM Mes/KOH buffer (pH 5.5)and the other half with Mes/KOH-buffered demineralized water(pH 5.5) for 4 h (as already described). After the pretreatment, theroot systems were rinsed with demineralized water, and the leafrosettes were cut off with a razor blade, just below the lowest leaf.Then, half of the His- or water-treated root systems were trans-ferred to fresh nutrient solution without Zn and the other half tofresh nutrient solution with 5 or 50 lM Zn(NO3)2, that is, thesame Zn concentrations in which the plants had been raised. Rootpressure exudates were collected overnight as described above.The volume of the exudates was measured and the root sampleswere prepared for Zn analysis as already described.
The amount of Zn loaded into the xylem was calculated, on aroot weight basis, as the product of the Zn concentration in theroot pressure exudate and the volume of the root pressure exu-date, divided by the root dry weight. The ‘total Zn burden’ of theroot system and the root pressure exudate together, at the end ofthe experiment, was calculated, on a root weight basis, as the sumof the products of the total amounts of Zn present in the rootpressure exudate and the root system, respectively. Zn xylemloading was then recalculated as a percentage of the total root Znburden.
Tonoplast vesicle isolation To establish the effect of His on Zntonoplast transport, 8-wk-old N. caerulescens (La Calamineecotype) and T. arvense plants grown in nutrient solution wereharvested. Tonoplast vesicles were obtained from root and shootmaterials according to Maeshima & Yoshida (1989) with slight
modifications. All procedures were conducted at 4°C. Plant mate-rial (100 g) was homogenized with a blender (Moulinex, GroupeSEB, Ecully, France) in a total of 400 ml of homogenizationmedium containing 250 mM sorbitol, 5 mM EGTA, 1 mMphenylmethylsulfonyl fluoride, 1.5% polyvinylpyrrolidone K30,1% ascorbic acid, 5 mM potassium metabisulfite, 5 mM dith-iothreitol (DTT), 1 lM E64, 200 lM benzamidine and 100 mMTris/Mes buffer, pH 7.8. The homogenate was filtered and centri-fuged at 10 000 g for 15 min in a 5.8.10 R centrifuge (Eppendorf,Hamburg, Germany). The supernatant was centrifuged at100 000 g for 30 min in a Beckman centrifuge (Beckman Coulter,Brea, CA, USA). The precipitate was suspended in 6 ml of 10 mMTris/Mes (pH 7.5), 500 mM sucrose, 2 mM EDTA and 2 mMDTT, and poured into a centrifugation tube. The suspension wasoverlayered with 6 ml of 10 mM Bis-Tris propane (BTP)/Mes(pH 7.2), 250 mM sorbitol, 0.5 mM EDTA and 1 mM DTT.After centrifugation at 120 000 g for 60 min in a Beckman centri-fuge (rotor SW 40), the interface portion was collected and dilutedin medium containing 10 mM BTP/Mes (pH 7.2), 250 mM sor-bitol, 0.5 mM EDTA and 1 mM DTT. The suspension was thencentrifuged at 100 000 g for 30 min, and the resulting white pellet(tonoplast vesicles) was suspended in medium containing250 mM sorbitol, 10 mM BTP (pH 7.5, set with Mes), 10 mMDTT and 1.1M glycerol. The protein concentration was deter-mined according to Bradford (1976), using BSA as the standard.
Vesicle integrity and purity Before the experiments, the integrityof the vesicles was checked by testing their interior acidificationtriggered by the addition of ATP (0.9mM Na2ATP) in the pres-ence of Mg2+ (2 mM MgSO4) in the assay medium by recordingthe differential absorbance changes at 492–540 nm undergone bythe membrane-penetrating ΔpH probe acridine orange (Palmgren,1991), using a Hitachi 557 (Tokyo, Japan) dual-wavelength spec-trophotometer. The degrees of inhibition of ATPase activity by ba-filomycin A1 (2.5 nM) and KNO3 (50mM), as well as byNa3VO4 (100 lM) and NaN3 (100 lM), were measured to esti-mate the contributions of vacuolar V-type, plasma membrane P-type and mitochondrial/chloroplastic F-type ATPases, respectively.
Metal uptake assay All uptake experiments were performedaccording to Richau et al. (2009) with minor modifications. Vesi-cle suspension aliquots (containing 25 lg of protein) were incu-bated in bathing medium containing 250 mM sorbitol, 10 mMBTP (pH 7.5, set with Mes), 10 mM DTT, 1.1 M glycerol and3 mM ATP. A proton gradient was induced by the addition ofMgSO4 at a final concentration of 6 mM. Zn was added to thebathing medium at a final concentration of 100 lM, as ZnSO4,Zn–His (1 : 1) or Zn–His (1 : 2) complex. After 100 s of incuba-tion with Zn, the vesicles were filtered over a nitrocellulosemembrane Protran BA 85 (Whatman GmbH, Dassei, Germany).After five washings with 5 ml sorbitol/Tris buffer, the residue waswashed off with 1.5 ml of 0.1 M HNO3 (Ultrex II Ultrapurereagent; Avantor™ Performance Materials BV, Deventer, theNetherlands), and Zn concentrations were measured using agraphite furnace atomic absorption spectrophotometer (PerkinElmer 5100 PC; Perkin Elmer Nederland BV, Groningen,
� 2014 The Authors
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014)
www.newphytologist.com
NewPhytologist Research 3

the Netherlands). The values were corrected for non-specificbinding of Zn to the vesicle membrane, established in a parallelincubation with vesicles that had been inactivated by heating for5 min at 90°C. Another parallel incubation was performed with-out additional Zn supply to the bathing medium to correct thevalues for Zn content in intact vesicles before exposure.
Measurement of Zn and His Zn concentrations in root pressureexudates and plant digests were measured using a flame atomicabsorption spectrophotometer (Perkin Elmer 2100). Dried plantmaterials were digested in 2 ml of a 4 : 1 (v/v) mixture of HNO3
(65%, v/v) and HCl (37%, v/v) in Teflon bombs at 140°C for7 h. Before drying, the roots were desorbed in 20 mM Na2EDTAfor 10 min at room temperature. His concentrations in plantmaterials were measured using high-performance liquid chroma-tography (HPLC), as described in Richau et al. (2009).
Histochemical detection of Zn in root tissues Roots of 8-wk-old plants grown at different Zn exposure levels were used tovisualize Zn distribution patterns over root tissues. To maximizethe chances of detecting quantitative differences, we applied twoZn staining techniques with different sensitivities, using Zinconand Zinpyr-1, respectively. Zincon is a relatively Zn-selectiveindicator, forming a blue-colored complex with Zn. To maximizesensitivity, we adopted the procedure described in Seregin &Kozhevnikova (2011) and Seregin et al. (2011). In short, 0.013 gZincon (sodium salt, C20H15N4NaO6S; Sigma-Aldrich ChemieGmbH) and 0.19 g borax (Na2B4O7.10H2O) were dissolved in0.2 ml of 1M NaOH (pH 9.8–10.4) and the volume wasadjusted to 10 ml with super-demineralized water. The solutionwas then heated to 90°C and used after cooling to room tempera-ture. Thin transverse sections of roots were prepared using asafety razor blade. Fine root segments and sections were incu-bated on a glass slide in a few drops of the reagent for 10–15 min,covered with a cover glass and examined using an OlympusCX41 microscope (Olympus, Shinjuku, Tokyo, Japan). Photo-graphs were taken using a color video camera Altra 20 (Olym-pus). Six to eight plants per treatment were inspected.
As a more sensitive reagent, we used the membrane-permeableZn fluorophore Zinpyr-1. Zinpyr-1 is highly selective for Zn overother metals (Kd = 0.7� 0.1 nM) (Burdette et al., 2001). Weapplied the procedure described in Sinclair et al. (2007), withslight modifications (Seregin & Kozhevnikova, 2011; Sereginet al., 2011). In short, we used a 10 lM solution of Zinpyr-1(C46H36Cl2N6O5; Fluka, St Louis, MO, USA) in super-demin-eralized water, diluted from a 1 mM stock made up in dimethylsulfoxide (DMSO). Before investigation, roots were washed indemineralized water, placed in 10 mM Na2EDTA for 5 min andthen washed again with demineralized water. Root samples wereimmersed in Zinpyr-1 solution (10 lM) and incubated at roomtemperature in darkness for 1 h. Then, the samples were washedin ultrapure water. We then prepared transverse sections using asafety razor blade, placed them on a glass slide in a few drops ofultrapure water and covered them with a cover glass. Intact fineroot segments were also examined. Images were taken on a confo-cal laser scanning microscope Nikon Eclipse TE2000-S (laser
scanning system Radiance 2000; Nikon, Chiyoda, Tokyo,Japan), using excitation at 488 nm. To prove that the observedfluorescence was caused by Zinpyr-1 binding to Zn, we alsoexposed root samples to the Zn-chelator, TPEN (N,N,N 0,N 0-tetrakis(2-pyridylmethyl)ethylenediamine, 1 mM solution), for2 h in darkness at room temperature, before exposure to Zinpyr-1. Zn-dependent fluorescence was not observed in this case.
Statistical analysis
All experiments were replicated at least three times. Quantitativedata were analyzed statistically using two-way and one-wayANOVA, if necessary after log transformation. A posteriori com-parison of individual means was performed using the minimumsignificant range (MSR) as a statistic (Sokal & Rohlf, 1981). Forhistochemical analyses, we used six to eight plants per treatment.
Results
Effects of exogenous His supply on Zn xylem loading
His concentrations in roots and shoots were as in Richau et al.(2009), meaning that His concentrations were c. 10-fold higherin N. caerulescens than in T. arvense, although only in roots, andthe root and shoot His concentrations were unaffected by the Znexposure level (data not shown).
The volume of exuded xylem sap was not influenced signifi-cantly by the exogenous amino acid supply in the majority of theexperiments (Fig. 1). His treatment had a significant positiveeffect on the exudate volume in the SF and MP accessions ofN. caerulescens at 500 lM Zn, and in the LC accession at 5 lMZn (Fig. 1). Alanine treatment decreased significantly the exudatevolume in T. arvense at 5 lM Zn, but increased the volume in SFat 500 lM Zn (Fig. 1).
Exogenous alanine supply did not influence significantly, andoccasionally decreased slightly, the Zn concentrations in the rootpressure exudates in both species, in comparison with the watercontrol treatment (Fig. 2a–c). Exogenous His supply enhancedsignificantly the Zn concentrations in the root pressure exudatesin all accessions of N. caerulescens, irrespective of the Zn concen-tration in the nutrient solution after the amino acid treatment,except for the accessions LE and LC at the exposure level of 5 lMZn (Fig. 2a–c). The Zn concentrations in the root pressure exu-dates of T. arvense, however, were not affected, or were decreasedslightly, by both the His and alanine treatments, in comparisonwith the water control treatment (Fig. 2a,b). In all the treatments,including the water controls, there was an overall increase in thexylem Zn concentration with the Zn concentration in the nutri-ent solution applied after the treatments. The relative effects ofthe His treatment, in comparison with the water controls, werenot significantly dependent on the Zn supply rate after the treat-ments, except for the LE N. caerulescens accession, where the Hiseffect increased strongly with increasing Zn supply (Fig. 2a–c).
Recalculation of the amount of Zn loaded into the xylem as apercentage of the total Zn burden of the root itself and the rootpressure exudate together, at the end of the experiment,
New Phytologist (2014) � 2014 The Authors
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist4

completely abolished the significant effect of the external Zn con-centration, suggesting that both species loaded a more or less con-stant fraction of their root Zn burden into the xylem (Fig. 2d–f).Remarkably, when expressed as a fraction of the total burden,there was no longer any significant difference between LC and SF(Fig. 2d–f), which had very different Zn concentrations in theirroot pressure exudates (Fig. 2a–c), apparently entirely caused bydifferential total Zn burdens. However, when expressed as a frac-tion of the total burden, Zn xylem loading remained low in thecase of accessions LE and MP (Fig. 2d–f), but the significantincrease in the effect of His pretreatment on the Zn concentra-tion in the root pressure exudate with the external Zn concentra-tion characteristic of LE (Fig. 2a–c) was no longer apparent. Theeffect of the His treatment, however, in comparison with thewater control, remained significant in T. arvense, as well as in allof the N. caerulescens accessions, except for LE at 5 lM and MPat 50 and 500 lM Zn.
As xylem Zn concentrations often increased only slightly withthe Zn supply after the His treatment, particularly when compar-ing the supply rates of 5 and 50 lM (Fig. 3a,b), we performed anadditional experiment with T. arvense and N. caerulescens, acces-sion SF, grown at 5 lM Zn (T. arvense) and 5 or 50 lM Zn(N. caerulescens) for 8 wk. After the His treatment, the root sys-tems were either exposed to the same Zn concentrations or to aZn-free nutrient solution. It appeared that the presence orabsence of external Zn after the treatments did not affect signifi-cantly the Zn concentrations in the root pressure exudates(Fig. 3a), nor did it affect significantly the relative xylem Znloads, expressed as a percentage of the total Zn burden of the rootand the exudate together (Fig. 3b). The interaction between Znsupply after the amino acid treatment and the relative xylem Znload, or the xylem Zn concentration, was consistently insignifi-cant in both species (Table 1). However, the His effect was con-sistently significant in the SF N. caerulescens accession (Fig. 3,
Fig. 1 Volumes of root pressure exudates ofdifferent Noccaea caerulescens accessions(N.c. SF, LC, LE and MP) treated with 5, 50or 500 lM zinc (Zn) and Thlaspi arvense(T.a.) treated with 5, 25 or 50 lM Znovernight after pretreatment with 1 mM L-histidine (L-His) (white bars), L-alanine (graybars) or Mes/KOH-buffered demineralizedwater (pH 5.5; unpretreated, black bars).Values are the means of three replicates withsix plants per replicate� SE. Significantlydifferent from control: *, P < 0.05; **,P < 0.01.
� 2014 The Authors
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014)
www.newphytologist.com
NewPhytologist Research 5
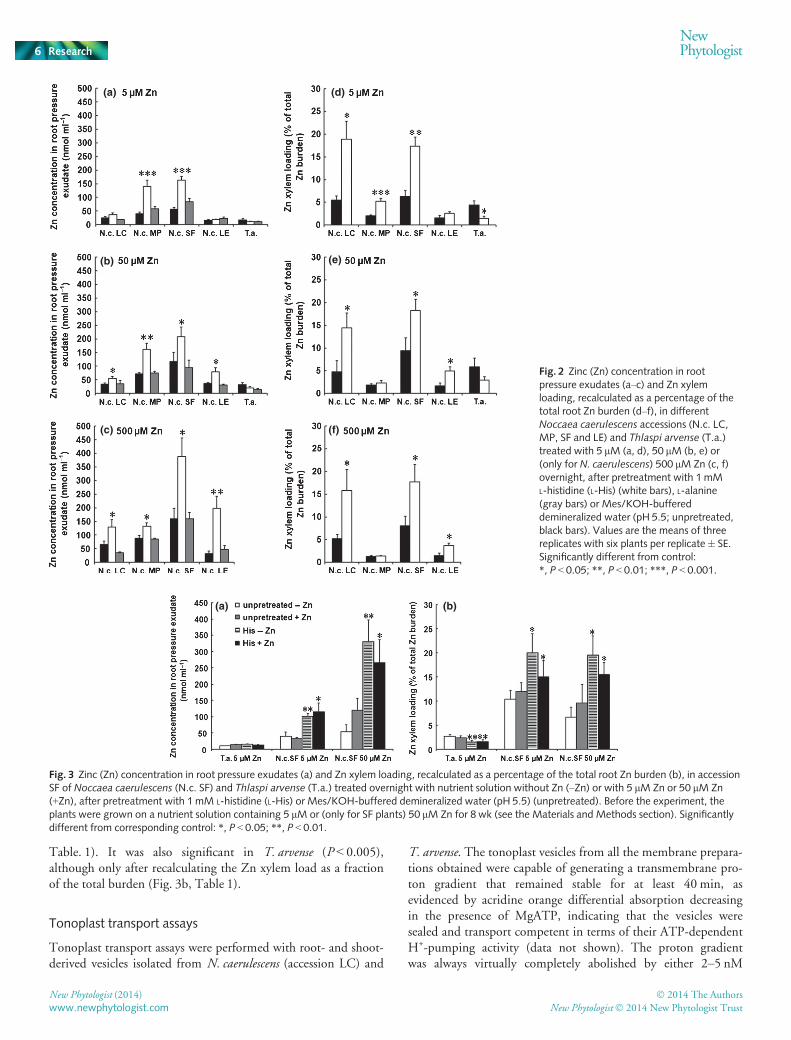
Table. 1). It was also significant in T. arvense (P < 0.005),although only after recalculating the Zn xylem load as a fractionof the total burden (Fig. 3b, Table 1).
Tonoplast transport assays
Tonoplast transport assays were performed with root- and shoot-derived vesicles isolated from N. caerulescens (accession LC) and
T. arvense. The tonoplast vesicles from all the membrane prepara-tions obtained were capable of generating a transmembrane pro-ton gradient that remained stable for at least 40 min, asevidenced by acridine orange differential absorption decreasingin the presence of MgATP, indicating that the vesicles weresealed and transport competent in terms of their ATP-dependentH+-pumping activity (data not shown). The proton gradientwas always virtually completely abolished by either 2–5 nM
(a)
(b)
(c)
(d)
(e)
(f)
Fig. 2 Zinc (Zn) concentration in rootpressure exudates (a–c) and Zn xylemloading, recalculated as a percentage of thetotal root Zn burden (d–f), in differentNoccaea caerulescens accessions (N.c. LC,MP, SF and LE) and Thlaspi arvense (T.a.)treated with 5 lM (a, d), 50 lM (b, e) or(only for N. caerulescens) 500 lM Zn (c, f)overnight, after pretreatment with 1 mML-histidine (L-His) (white bars), L-alanine(gray bars) or Mes/KOH-buffereddemineralized water (pH 5.5; unpretreated,black bars). Values are the means of threereplicates with six plants per replicate� SE.Significantly different from control:*, P < 0.05; **, P < 0.01; ***, P < 0.001.
(a) (b)
Fig. 3 Zinc (Zn) concentration in root pressure exudates (a) and Zn xylem loading, recalculated as a percentage of the total root Zn burden (b), in accessionSF of Noccaea caerulescens (N.c. SF) and Thlaspi arvense (T.a.) treated overnight with nutrient solution without Zn (–Zn) or with 5 lM Zn or 50 lM Zn(+Zn), after pretreatment with 1mM L-histidine (L-His) or Mes/KOH-buffered demineralized water (pH 5.5) (unpretreated). Before the experiment, theplants were grown on a nutrient solution containing 5 lM or (only for SF plants) 50 lM Zn for 8 wk (see the Materials and Methods section). Significantlydifferent from corresponding control: *, P < 0.05; **, P < 0.01.
New Phytologist (2014) � 2014 The Authors
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist6

bafilomycin or 50 mM KNO3, two well-known specific inhibi-tors of vacuolar V-type H+-ATPase (Bowman et al., 1988; White,1994). However, the ATPase activity was not inhibited signifi-cantly by the specific P-type ATPase inhibitor, Na3VO4 (0–1.1%inhibition), or the F-type ATPase inhibitor, NaN3 (0–1.3% inhi-bition). These results suggest that all of the vesicle preparationswere, at most, slightly contaminated with non-vacuolar mem-brane fractions.
Significant Zn uptake was found in all the vesicle preparations(Fig. 4). The root-derived tonoplast vesicles of T. arvense took upsignificantly more Zn than did the leaf-derived ones, irrespective ofwhether Zn was supplied as Zn-His or ZnSO4. In root-derivedtonoplast vesicles ofT. arvense, Zn uptake was higher, albeit insignif-icantly, when Zn was supplied as Zn-His, in comparison withZnSO4. In N. caerulescens, the Zn uptake was unaffected in root-derived vesicles, and decreased insignificantly in leaf-derived ones,when Zn andHis were supplied in a 1 : 1 molar ratio, in comparisonwith the ZnSO4 supply. When Zn and His were supplied in a 1 : 2molar ratio, Zn uptake was inhibited significantly inN. caerulescens,but not affected significantly in T. arvense, in comparison with theZnSO4 supply, in both root- and shoot-derived vesicles (Fig. 4).Two-way analysis of the results obtained with tonoplast vesiclesyielded a significant (P < 0.05) Zn supply9 species interaction(Table 2).
Zn accumulation in roots and leaves
At 2 lM Zn in the nutrient solution, the Zn concentration in theroots was comparable in T. arvense and N. caerulescens (Fig. 5a),whereas, in the leaves, it was much higher in N. caerulescens, withaccession LE showing the highest value (Fig. 5b). At 50 lM Znin the nutrient solution, T. arvense accumulated up to 16 timesmore Zn in the roots than did N. caerulescens (Fig. 5a), whereasthe Zn concentration in the leaves was higher in the hyperaccu-mulator, except for accession LC (Fig. 5b).
Table 1 Two-way analysis of variance of the zinc (Zn) xylem loading (% oftotal root Zn burden) in Noccaea caerulescens (SF accession) and Thlaspi
arvense
VariantSource ofvariation df SS MS F P
SF, 5 lM Zn � Zn 1 47.870 47.870 1.011 ns�His 1 332.893 332.893 7.030 0.015Zn9His 1 106.866 106.866 2.257 nsWithin 20 947.113 47.356
SF, 50 lM Zn � Zn 1 1.444 1.444 0.027 ns�His 1 431.536 431.536 8.074 0.012Zn9His 1 58.198 58.198 1.089 nsWithin 16 855.205 53.450
T. arvense,5 lM Zn
� Zn 1 0.026 0.026 0.032 ns�His 1 7.649 7.649 9.367 0.005Zn9His 1 0.369 0.369 0.451 nsWithin 28 22.864 0.817
His, histidine; ns, not significant; difference considered to be significant atP < 0.05.
(a) (b)
Fig. 4 Zinc (Zn) concentrations in MgATP-energized root-derived (a) and leaf-derived(b) tonoplast vesicles of Thlaspi arvense(T.a.) and Noccaea caerulescens, accessionLC (N.c. LC) after 100 s of incubation with100 lM ZnSO4 (black bars), Zn–histidine(Zn–His) (1 : 1; white bars) or Zn-His (1 : 2;gray bars) (means� SE of three independentbatches of plants). Values were corrected fornon-specific binding. Significantly differentfrom ZnSO4 supply: ***, P < 0.001.
Table 2 Two-way analysis of variance of the zinc (Zn) concentration in root- and shoot-derived tonoplast vesicles of Noccaea caerulescens (LC accession)and Thlaspi arvense
Variant Source of variation df SS MS F P
Roots Species 1 315.178 315.178 36.274 0.00002***Zn treatment 2 160.034 80.017 9.209 0.00789**Species9 Zn treatment 2 63.443 31.721 3.651 0.04942*Within 20 173.774 8.689
Shoots Species 1 25.503 25.503 27.330 0.00021***Zn treatment 2 48.573 24.287 26.027 0.00004***Species9 Zn treatment 2 11.332 5.666 6.072 0.01508*Within 12 11.198 0.933
ns, not significant; *, P < 0.05; **, P < 0.01;***, P < 0.001.
� 2014 The Authors
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014)
www.newphytologist.com
NewPhytologist Research 7

Equal root-internal Zn concentrations were achieved at 70 lMZn in the solution for T. arvense and at 1000 lM Zn forN. caerulescens, accession LE (Fig. 5a). Therefore, these plantswere compared for the distribution of Zn over root tissues.
Zn distribution in roots
In control plants (grown at 2 lM Zn), Zn was found in all theroot tissues. In the rhizodermis, cortex, endodermis and xylem,
(a) (b)Fig. 5 Zinc (Zn) concentration in roots (a)and leaves (b) of Noccaea caerulescensaccessions (N.c. SF, LE, LC, MP) and Thlaspi
arvense (T.a.) after 8 wk of exposure todifferent Zn concentrations in the nutrientsolution. Values are the means of threereplicates with six plants per replicate� SE.Treatments assigned different letters indicatea significant difference between the means(P < 0.05). ANOVA post-hoc comparisonswere performed separately for roots andleaves, as well as for species/accessions.
(a) (e) (i)
(b) (f) (j)
(c) (g) (k)
(d) (h) (l)
Fig. 6 Zinc (Zn) distribution over root tissuesin Noccaea caerulescens (accession LE)exposed to 2 lM (a, b), 50 lM (e, f) or1000 lM (c, i, j) Zn and Thlaspi arvenseexposed to 2 lM (d), 50 lM (g, h) or 70 lM(k, l) Zn for 8 wk. Green fluorescenceindicates the location of the Zn–Zinpyr-1complex. (a, c, g, i, k) Root hair zone. (b, d,e, f, h, j, l) Mature zone of the root. Eachimage is representative of a selection of sixplants per plant type per treatment.Designations: Atr, atrichoblast; C, cortex;CC, central cylinder; E, endodermis; LRP,lateral root primordium; P, pericycle; Ph,phloem; R, rhizodermis; RH, root hair; Tr,trichoblast; X, xylem vessel; XP, xylemparenchyma.
New Phytologist (2014) � 2014 The Authors
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist8

the fluorescence of the Zn–Zinpyr-1 complex was detectedmainly in the cell walls, whereas, in the pericycle, phloem andxylem parenchyma, it was more prominent in the protoplasts(Fig. 6b,d). In T. arvense, the fluorescence of the protoplasts washigher than in N. caerulescens, and it was occasionally alsodetected in the endodermis and the inner cortical layer (Fig. 6d).In both species, fluorescence was also intense in the protoplasts ofthe trichoblasts, especially in growing root hairs (Fig. 6a). Thehighest intensity of fluorescence was found in the root apex andthe lateral root primordia (data not shown).
At 50 lM Zn in the nutrient solution, the intensity of fluores-cence of the Zn–Zinpyr-1 complex was higher overall in bothspecies, in comparison with the exposure at 2 lM, but more so inT. arvense than in N. caerulescens (Fig. 6e–h). The pattern of Zndistribution remained largely the same (Fig. 6e–h; SupportingInformation Fig. S1), although, in the pericycle, phloem andxylem parenchyma, Zn was also detected in the cell walls. Znaccumulated in the root apex, lateral root primordia and growingroot hairs (Fig. 6e,g). The Zn-dependent fluorescence of the pro-toplasts of the root cells, especially those of the inner corticallayer and the endodermis, was less intense in N. caerulescens thanin T. arvense (Fig. 6f,h).
When grown at high Zn, and compared at equal root-internalZn concentrations, that is in plants grown at 1000 lM Zn forN. caerulescens (accession LE) and 70 lM Zn for T. arvense(Fig. 5a), both species exhibited very similar patterns of Zn distri-bution over root tissues. In the root apex, Zn was found in all thetissues and accumulated in the meristem, in both the cell wallsand protoplasts (Fig. 7a,b,e). Zn was unevenly distributed overrhizodermal cell types: it accumulated much more in trichoblaststhan in atrichoblasts (Fig. 7b). Intense fluorescence of theZn–Zinpyr-1 complex and Zincon staining were detected ingrowing root hairs (Figs 6, 7c,g). Interestingly, high Zn exposureinfluenced the development of root hairs in both species: theystarted to branch (Fig. 7c,g). In the root hair zone and the matureroot sections of both N. caerulescens and T. arvense, Zn wasdetected in all the tissues and accumulated in the central cylinder,mainly in the conducting tissues and the pericycle, where the
fluorescence and staining were more intense in the protoplaststhan in the cell walls (Figs 6i–l, 7d,f,h). In the outer cortical layer,the fluorescence and staining were more intense in the cell wallsthan in the protoplasts (Figs 6j–l, 7d,f,h). Zn also often accumu-lated in the protoplasts of the endodermis and the cells of theinner cortical layer (Figs 6i,k, 7d,f,h).
Discussion
Our results clearly demonstrate a strong effect of exogenouslysupplied His on the Zn concentration in the root pressure exu-date. The increase observed in N. caerulescens is certainly notattributable to a change in exudate volume, but is evidentlycaused by an enhanced rate of Zn loading into the xylem. Theeffects on Zn xylem loading are qualitatively in full agreementwith earlier results obtained for Ni xylem loading in the samespecies (Richau et al., 2009). Overall, in N. caerulescens, the xylemloading of Zn seems to be impacted more strongly than that ofNi by exogenous His, for which more than two-fold enhancedxylem loading rates were found in only one accession, that is MP,which is by far the best Ni-hyperaccumulating accession (Richauet al., 2009). As suggested by the experiment in which we did notrestore the Zn supply after the His treatment, this may be relatedto a His-mediated mobilization of Zn for xylem loading frompre-existing root-internal burdens. In the experiment of Richauet al. (2009), however, such a burden was not present, becausethe plants were grown in Ni-free nutrient solution until shootexcision. The negative effect of exogenous His on Zn xylem load-ing in T. arvense was also relatively strong, in comparison withthat on Ni loading, and also apparent both with and without Znin the nutrient solution, suggesting that exogenous His mightimmobilize a significant part of the pre-existing root Zn burdenin this species.
We also performed the above experiments with Cd, althoughonly at a concentration of 1 lM in the nutrient solution, becausehigher concentrations often decreased strongly the exudate vol-umes. In contrast with Zn, regardless of whether xylem Cd wasexpressed as a concentration or as a fraction of the total root
(a) (b) (d)
(e) (f) (h)(g)
(c)Fig. 7 Zinc (Zn) distribution over root tissuesin Noccaea caerulescens (LE accession)exposed to 1000 lM Zn (a–d) and in Thlaspiarvense exposed to 70 lM Zn (e–h) for 8 wk.Blue staining indicates location of the Zn–Zincon complex. (a, b, e) Apical part of theroot. (d, f, h) Mature zone of the root. (c, g)Root hairs. Bars: (a–h) 25 lm. Each image isrepresentative of a selection of six plants perplant type per treatment. Designations: Atr,atrichoblast; C, cortex; CC, central cylinder;E, endodermis; M, meristematic cells; P,pericycle; Ph, phloem; R, rhizodermis; RC,root cap; Tr, trichoblast; X, xylem vessel; XP,xylem parenchyma.
� 2014 The Authors
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014)
www.newphytologist.com
NewPhytologist Research 9

burden, there was never any significant effect of His pretreatmenton Cd xylem loading (A. D. Kozhevnikova, I. V. Seregin, R.Verweij & H. Schat, unpublished data). This could be becauseCd, as a sulfur-seeking metal, preferentially binds to sulfur-basedchelators, rather than nitrogen/oxygen-based ones, at least atcytoplasmic pH levels (Singel & Singel, 1986; Seregin & Ivanov,2001).
With regard to the potential role of His in vacuolar Zn seques-tration, our results obtained with tonoplast vesicles are in linewith the hypothesis that Zn–His complex formation inhibits Zntransport across the tonoplast in N. caerulescens, but not inT. arvense. The supply of Zn in combination with His (1 : 1)enhanced slightly Zn uptake in T. arvense vesicles, in comparisonwith ZnSO4 supply, such as found previously for Ni–His. How-ever, it did not, or only slightly, lower the Zn uptake inN. caerulescens vesicles, which is different from the resultsobtained with Ni (Richau et al., 2009). However, when Zn andHis were supplied in a 1 : 2 molar ratio, Zn uptake was stronglyand significantly suppressed in vesicles of N. caerulescens, but notsignificantly affected in vesicles of T. arvense. Admittedly, our evi-dence that His-mediated Zn xylem loading relies on Zn–Hiscomplex formation in the cytoplasm is circumstantial, rather thandirect. There is strong evidence that Zn is largely bound to His inN. caerulescens roots (Salt et al., 1999), but the precise subcellularlocalization of this His-bound Zn pool has been elusive to date.
The Zn distribution patterns over root tissues and root celltypes were virtually the same in both species, at least when com-pared in plants grown at high Zn concentrations in the nutrientsolution. In both species, Zn accumulated strongly in the rootapex and lateral root primordia, which was also found for Ni(Richau et al., 2009). Intracellular Zn accumulation was apparentin all the stelar tissues, the endodermis and the cortex. Withinthe root hair zone, intracellular Zn also accumulated in the rhizo-dermis, particularly in root hair-bearing cells (‘trichoblasts’),rather than in cells without a root hair (‘atrichoblasts’).
When compared at a normal nutritional Zn supply in thenutrient solution (2 lM), the fluorescence of the Zn–Zinpyr-1complex was largely associated with cell walls. Intracellular fluo-rescence was apparent in the inner cortical cell layer and theendodermis of mature root sections in T. arvense, but not inN. caerulescens. At 50 lM in the nutrient solution, at which theoverall root-internal Zn concentration was higher in T. arvensethan in N. caerulescens, the difference between the two speciesbecame more pronounced: Zn-dependent fluorescence of theprotoplasts of the root cells, especially in the inner cortical layerand endodermis, was more intense in T. arvense, in comparisonwith N. caerulescens. In addition, Zn treatment- or accession-dependent variation in intracellular Zn staining in N. caerulescenswas also mainly apparent in the cortex and the endodermis (datanot shown). This may be taken to suggest that the high root Hisconcentration in N. caerulescens serves to counteract the vacuolarretention of Zn in mature peripheral root tissues, particularly thecortex and the endodermis. Richau et al. (2009) obtained verysimilar results for Ni, that is less intracellular Ni storage in thecortex and the endodermis in N. caerulescens in comparison withT. arvense.
In summary, in many respects, there is a strong resemblancebetween Ni and Zn with regard to their distribution patterns overroot tissues and the effects of His on their loading into the xylemin N. caerulescens and T. arvense. This suggests that the underly-ing mechanism is the same for both metals, that is inhibition inN. caerulescens and enhancement in T. arvense, of vacuolar storagein root cortical and endodermal cells. Regardless of the underly-ing mechanism, it is evident that His-mediated Zn xylem loadingis a species-wide property in N. caerulescens. This suggests thatthe property may have evolved as a component part of the Znhyperaccumulation machinery, because the hyperaccumulationof Zn is species wide, whereas Ni hyperaccumulation is accessionspecific in N. caerulescens (Assunc�~ao et al., 2003). In this scenario,the capacity for His-mediated Ni xylem loading could well repre-sent a by-product of the Zn hyperaccumulation trait. The evolu-tion of Ni hyperaccumulation in serpentine accessions, such asMP, would then merely require an enhanced capacity for Niuptake (Richau et al., 2009), because the translocation machineryis already there. Of course, it cannot be excluded that theancestral Noccaea species was an Ni hyperaccumulator. If so, thenHis-mediated Zn xylem loading in N. caerulescens might, in fact,reflect a case of ‘inadvertent hyperaccumulation’ as a result of anincompletely metal-specific ancestral mechanism for Ni xylemloading.
It is interesting to compare the role of His in Ni-hyperaccumu-lating Alyssum and Zn-hyperaccumulating Noccaea, particularlybecause N. caerulescens consistently shows high root to shoottranslocation rates for both Ni and Zn (Richau et al., 2009), butNi-hyperaccumulating Alyssum species only exclusively for Ni(deVarennes et al., 1996; Schat et al., 2000). The reason for thismay be that root His accumulation in Alyssum requires furtherinduction by, specifically, Ni exposure (Kr€amer et al., 1996),although the constitutive concentration is already high, in com-parison with non-hyperaccumulators (Kerkeb & Kr€amer, 2003).More likely, root His accumulation is obviously not sufficient forZn hyperaccumulation, which also requires, as shown in bothA. halleri and N. caerulescens, enhanced expression of the Zn-transporting 1b P-type ATPase, HMA4, which transports Zn,but also Cd, from the xylem parenchyma into the xylem vessels(Hanikenne et al., 2008; Lochlainn et al., 2011; Iqbal et al.,2013); this might, in turn, explain the correlation between Znand Cd translocation among Noccaea species and accessions(Xing et al., 2008). It seems unlikely that HMA4 is also directlyinvolved in the xylem loading of Ni, although direct evidence islacking at this point.
Finally, whatever the underlying mechanism of His-imposedenhanced root to shoot metal translocation, it is almost certainlynot based on the chelation of the metals in the xylem itself, whichis apparently not required for hyperaccumulation (Ueno et al.,2008; Alves et al., 2011; Monsant et al., 2011; Centofani et al.,2013). It seems likely that His contributes to the radial transportof Zn or Ni across the root from the root surface into the stele, oreven the xylem, such as postulated for nicotianamine in A. halleri(Deinlein et al., 2012). It is not unlikely that nicotianamine playsa comparable role in N. caerulescens (van de Mortel et al., 2006;Gendre et al., 2007; Trampczynska et al., 2010). Thus, it is
New Phytologist (2014) � 2014 The Authors
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist10

conceivable that His and nicotianamine both function to enhancethe radial transport of Ni and Zn across the root inN. caerulescens. It is not yet known whether these compoundsserve distinct, specific functions in this process, or whether theyact in a merely additive manner.
Acknowledgements
This work was supported by grants from the Dutch Organizationfor Scientific Research (NWO, 047.017.008) and the RussianFoundation for Basic Research (RFBR, no. 11-04-00513).The authors would like to thank Dr Francesca Quattrocchiofrom the Department of Genetics and Dr Yves J. M. Bollen fromthe Department of Structural Biology at the Free UniversityAmsterdam for assistance, Professor Victor Ivanov andDr Marina S. Trofimova from the Institute of Plant Physiologyand Riet Vooijs, Henk Hakvoort and Richard van Logtestijnfrom the Free University Amsterdam for fruitful discussions,Dr Kerstin Helga Richau from Ume�a University for advice andsupport, and Dr Elena I. Bystrova, Nina E. Starovskaya, Dr IlyaBaklanov, Dr Mikhail Piotrovsky and Vladislava Gracheva fromthe Institute of Plant Physiology for technical support.
References
Alves S, Nabais C, Gonc�alves MDS, dos Santos MMC. 2011. Nickel speciation
in the xylem sap of the hyperaccumulator Alyssum serpyllifolium ssp lusitanicumgrowing on serpentine soils of north-east Portugal. Journal of Plant Physiology168: 1715–1722.
Assunc�~ao AGL, Ten BookumWM, Nelissen HJM, Vooijs R, Schat H, Ernst
WHO. 2003. Differential metal tolerance and accumulation patterns among
Thlaspi caerulescens populations originating from different soil types. NewPhytologist 159: 411–419.
Baker AJM, McGrath SP, Reeves DR, Smith JAC. 2000.Metal
hyperaccumulator plants: a review of the ecology and physiology of a biological
resource of phytoremediation of metal-polluted soils. In: Terry N, Banuelos G,
eds. Phytoremediation of contaminated soils and water. Boca Raton, FL, USA:
CRC Press, 171–188.Barabasz A, Wilkowska A, Ruszczy~nska A, Bulska E, Hanikenne M, Czarny M,
Kr€amer U, Antosiewicz DM. 2012.Metal response of transgenic tomato
plants expressing P1B-ATPase. Physiologia Plantarum 145: 315–331.Bowman EJ, Siebers A, Altendorf K. 1988. Bafilomicins: a class of
inhibitors of membrane ATPases from microorganisms, animal cells, and
plant cells. Proceedings of the National Academy of Sciences, USA 85:
7972–7976.Bradford MM. 1976. A rapid and sensitive method for the quantitation of
microgram quantities of protein utilizing the principle of protein–dye binding.Analytical Biochemistry 72: 248–254.
Brooks RR, Lee J, Reeves RD, Jaffr�e T. 1977. Detection of nickelliferous rocks
by analysis of herbarium specimens of indicator plants. Journal of GeochemicalExploration 7: 49–57.
Burdette SC, Walkup GK, Spingler B, Tsien RY, Lippard SJ. 2001. Fluorescent
sensors for Zn2+ based on a fluorescein platform: synthesis, properties and
intracellular distribution. Journal of the American Chemical Society 123:7831–7841.
Callahan DL, Spas DK, O’Hair RAJO, Salt DE, Baker AJM. 2008.
Relationships of nicotianamine and other amino acids with nickel, zinc and
iron in Thlaspi caerulescens. New Phytologist 176: 836–848.Centofani T, Sayers Z, Cabello-Conejo MI, Kidd P, Nishizawa NK, Kakei Y,
Davies AP, Sicher AC, Chaney RL. 2013. Xylem exudate composition and
root-to-shoot nickel translocation in Alyssum species. Plant and Soil 373:59–75.
Deinlein U, Weber M, Schmidt H, Rensch S, Trampczynska A, Hansen TH,
Husted S, Schjoerring JK, Talke IN, Kr€amer U et al. 2012. Elevatednicotianamine levels in Arabidopsis halleri roots play a key role in zinc
hyperaccumulation. Plant Cell 24: 708–723.Gendre D, Czernic P, Con�ej�ero G, Pianelli K, Briat J-F, Lebrun M, Mari S.
2007. TcYSL3, a member of the YSL gene family from the hyper-accumulator
Thlaspi caerulescens, encodes a nicotianamine-Ni/Fe transporter. Plant Journal49: 1–15.
Hanikenne M, Talke IN, Haydon MJ, Lanz C, Nolte A, Motte P, Kroymann J,
Weigel D, Kr€amer U. 2008. Evolution of metal hyperaccumulation required
cis-regulatory changes and triplication of HMA4. Nature 453: 391–396.Hussain D, Haydon MJ, Wang Y, Wong E, Sherson SE, Young Y, Camakaris J,
Harper JF, Cobbett CF. 2004. P-type ATPase heavy metal transporters with
roles in essential zinc homeostasis in Arabidopsis. Plant Cell 16: 1327–1339.Ingle RA, Mugford ST, Rees JD, Campbell MM, Smith JAC. 2005.
Constitutively high expression of the histidine biosynthetic pathway contributes
to nickel tolerance in hyperaccumulator plants. Plant Cell 17: 2089–2106.Iqbal M, Nawaz I, Hassan Z, Hakvoort HWJ, Bliek M, Aarts MAM, Schat H.
2013. Expression of HMA4 cDNAs of the zinc hyperaccumulator Noccaeacaerulescens from endogenous NcHMA4 promoters does not complement the
zinc-deficiency phenotype of the Arabidopsis thaliana hma2hma4 doublemutant. Frontiers in Plant Science 4: article 404.
Kerkeb L, Kr€amer U. 2003. The role of free histidine in xylem loading of nickel
in Alyssum lesbiacum and Brassica juncea. Plant Physiology 131: 716–724.Kr€amer U, Cotter-Howells JD, Charnock JM, Baker AJM, Smith JAC. 1996.
Free histidine as a metal chelator in plants that hyperaccumulate nickel. Nature379: 635–638.
Lasat MM, Baker AJM, Kochian LV. 1998. Altered zinc compartmentation in
the root symplasm and stimulated Zn absorption in the leaves as mechanisms
involved in Zn hyperaccumulation in Thlaspi caerulescens. Plant Physiology 118:875–883.
Lochlainn SO, Bowen HC, Fray RG, Hammond JP, King GJ, White PJ,
Graham NS, Broadley MP. 2011. Tandem quadruplication of HMA4 in the
zinc (Zn) and cadmium (Cd) hyperaccumulator Noccaea caerulescens. PLoSONE 6: e17814.
Maeshima M, Yoshida S. 1989. Purification and properties of vacuolar
membrane proton-translocating inorganic pyrophosphatase from mung bean.
Journal of Biological Chemistry 264: 20068–20073.Monsant AC, Kappen P, Wang YD, Pigram PJ, Baker AJM, Tang CX. 2011.
In vivo speciation of zinc in Noccaea caerulescens in response to nitrogen form
and zinc exposure. Plant and Soil 348: 167–183.van de Mortel JE, Almar Villanueva L, Schat H, Kwekkeboom J, Coughlan S,
Moerland PD, Ver Loren, van Themaat E, Koornneef M, Aarts MGM. 2006.
Large expression differences in genes for iron and zinc homeostasis, stress
response and lignin biosynthesis distinguish roots of Arabidopsis thaliana andthe related metal hyperaccumulator Thlaspi caerulescens. Plant Physiology 142:1127–1147.
Palmgren MG. 1991. Acridine orange as a probe for measuring pH gradient
across membranes: mechanism and limitations. Analytical Biochemistry 192:316–321.
Persans MW, Yan XG, Patnoe JMML, Kr€amer U, Salt DE. 1999.Molecular
dissection of the role of histidine in nickel hyperaccumulation in Thlaspigoesingense (H�al�acsy). Plant Physiology 121: 1117–1126.
Richau KH, Kozhevnikova AD, Seregin IV, Vooijs R, Koevoets PLM, Smith
JAC, Ivanov VB, Schat H. 2009. Chelation by histidine inhibits the vacuolar
sequestration of nickel in roots of the hyperaccumulator Thlaspi caerulescens.New Phytologist 183: 106–116.
Salt DE, Prince RC, Baker AJM, Raskin I, Pickering IJ. 1999. Zinc ligands in
the metal hyperaccumulator Thlaspi caerulescens as determined using X-ray
absorption spectroscopy. Environmental Science and Technology 33: 713–717.Schat H, Llugany M, Bernhard R. 2000.Metal-specific patterns of tolerance,
uptake, and transport of heavy metals in hyperaccumulating and
non-hyperaccumulating metallophytes. In: Terry N, Banuelos G, eds.
Phytoremediation of contaminated soils & water. Boca Raton, FL, USA: CRCPress, 171–188.
Seregin IV, Ivanov VB. 2001. Physiological aspects of cadmium and lead toxic
effects on higher plants. Russian Journal of Plant Physiology 48: 523–544.
� 2014 The Authors
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014)
www.newphytologist.com
NewPhytologist Research 11

Seregin IV, Kozhevnikova AD. 2011.Histochemical methods for detection of
heavy metals and strontium in the tissues of higher plants. Russian Journal ofPlant Physiology 58: 721–727.
Seregin IV, Kozhevnikova AD, Gracheva VV, Bystrova EI, Ivanov VB. 2011.
Tissue zinc distribution in maize seedling roots and its action on growth.
Russian Journal of Plant Physiology 56: 109–117.Sinclair SA, Sherson SM, Jarvis R, Camakaris J, Cobbett CS. 2007. The use of
the zinc-fluorophore, Zinpyr-1, in the study of zinc homeostasis in Arabidopsisroots. New Phytologist 174: 39–45.
Singel H, Singel A. 1986.Metal ions in biological systems. Concepts on metal iontoxicity. Vol. 20. New York, NY, USA: Marcel Dekker.
Sokal RR, Rohlf FJ. 1981. Biometry, 2nd edn. San Francisco, CA, USA: W.H.
Freeman & Co.
Trampczynska A, K€upper H, Meyer-Klaucke M, Schmidt H, Clemens S.
2010. Nicotianamine forms complexes with Zn(II) in vivo. Metallomics 2:57–66.
Ueno D, Iwashita T, Zhao FJ, Ma JF. 2008. Characterization of Cd
translocation and identification of the Cd form in the xylem sap of the
Cd-hyperaccumulator Arabidopsis halleri. Plant and Cell Physiology 49:540–548.
deVarennes A, Torres MO, Coutinho JF, Rocha MMGS, Neto MMPM. 1996.
Effects of heavy metals on the growth and the mineral composition of a nickel
hyperaccumulator. Journal of Plant Nutrition 19: 669–676.White PJ. 1994. Bafilomycin A1 is a non-competitive inhibitor of the tonoplast
H+-ATPase of maize coleoptiles. Journal of Experimental Botany 45: 1397–1402.
Wong CKE, Cobbett CF. 2009.HMA P-type ATPases are the major mechanism
for root-to-shoot Cd translocation in Arabidopsis thaliana. New Phytologist 181:71–78.
Wycisk K, Kim EJ, Schroeder JI, Kr€amer U. 2004. Enhancing the first step in
the histidine biosynthesis pathway increases the free histidine pool and nickel
tolerance in Arabidopsis thaliana. FEBS Letters 587: 128–134.Xing JP, Jiang RF, Ueno D, Ma JF, Schat H, McGrath SP, Zhao FJ. 2008.
Variation in root-to-shoot translocation of cadmium and zinc among different
accessions of the hyperaccumulators Thlaspi caerulescens and Thlaspi praecox.New Phytologist 178: 315–325.
Yang X, Li TQ, Yang JC, He ZL, Lu LL, Meng FH. 2006. Zinc
compartmentation in root, transport into xylem, and absorption into leaf cells
in the hyperaccumulating species of Sedum alfrediiHance. Planta 224:185–195.
Supporting Information
Additional supporting information may be found in the onlineversion of this article.
Fig. S1 Staining of zinc (Zn) with Zinpyr-1, staining of cell wallswith propidium iodide (PI) and merged picture of their co-locali-zation in the roots of Noccaea caerulescens (accession LE) exposedto 50 lM Zn.
Please note: Wiley Blackwell are not responsible for the contentor functionality of any supporting information supplied by theauthors. Any queries (other than missing material) should bedirected to the New Phytologist Central Office.
New Phytologist is an electronic (online-only) journal owned by the New Phytologist Trust, a not-for-profit organization dedicatedto the promotion of plant science, facilitating projects from symposia to free access for our Tansley reviews.
Regular papers, Letters, Research reviews, Rapid reports and both Modelling/Theory and Methods papers are encouraged. We are committed to rapid processing, from online submission through to publication ‘as ready’ via Early View – our average timeto decision is <25 days. There are no page or colour charges and a PDF version will be provided for each article.
The journal is available online at Wiley Online Library. Visit www.newphytologist.com to search the articles and register for tableof contents email alerts.
If you have any questions, do get in touch with Central Office ([email protected]) or, if it is more convenient,our USA Office ([email protected])
For submission instructions, subscription and all the latest information visit www.newphytologist.com
New Phytologist (2014) � 2014 The Authors
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist12




















