
MvfR, a key Pseudomonas aeruginosa pathogenicity LTTR-class regulatory protein, has dual ligands

Clin Exp Immunol 1994; 97:113-119
Pathogenicity and immunogenicity of Paracoccidioides brasiliensis isolates inthe human disease and in an experimental murine model
L. M. SINGER-VERMES, E. BURGER, V. L. G. CALICH, L. H. MODESTO-XAVIER,T. N. SAKAMOTO, M. F. SUGIZAKI*, D. A. MEIRA* & R. P. MENDES* Depto. de Imunologia,
Instituto de Ciincias Biomedicas, Universidade de Sdo Paulo, Sdo Paulo, and *Faculdade de Medicina e Instituto de Biociencias,UNESP, Botucatu, Brasil
(Acceptedfor publication 17 March 1994)
SUMMARY
The pathogenicity and immunogenicity of six recently isolated Paracoccidioides brasiliensissamples derived from patients presenting distinct and well defined clinical forms of paracocci-dioidomycosis (PCM) were compared as to their virulence, tropism to different organs and abilityto induce specific cellular and humoral immune response in susceptible (BIO.A) inbred mice.Isolates Pb44 and Pb47 were obtained from acute cases, Pb5O from a chronic severe form, Pb45from a chronic moderate case and both Pb56 and Pb57 from chronic mild forms of PCM.Pathogenicity and tropism of each fungal sample were evaluated by LD50% estimation,examination of gross lesions on various organs at 2, 4, 12 and 16 weeks post-infection, and bycolony-forming unit (CFU) counts in the lungs at week 16 post-infection of mice. Fungal tropismin human PCM and in B1O.A mice was always dissociated. A well defined relationship betweenvirulence of the fungal sample and the clinical findings of the correspondent patient was notevident, although a tendency to higher LD50% and less intense paracoccidioidic lesions was
observed in mice infected with Pb56 and Pb57. The specific DTH response patterns variedaccording to the infectant sample, but positive DTH reactions at the beginning of the infectionand a tendency to anergy or low DTH responses at week 12 and/or week 16 post-infection were
always observed. A correspondence between the DTH response in humans and in mice was
noticeable only when the isolates from the most benign cases (Pb56 and Pb57) were considered.The specific antibody patterns in mice and in the correspondent patients were also not analogous.Collectively, these results indicate that an association between the fungal pathogenicity andimmunogenicity in the human disease and in susceptible mice was discernible only when isolatesobtained from very mild cases (Pb56 and Pb57) were considered.
Keywords paracoccidioidomycosis isogenic murine model fungal virulenceantibody response DTH reactions
INTRODUCTION
Paracoccidioidomycosis (PCM), a human systemic mycosiscaused by the thermally dimorphic fungus Paracoccidioidesbrasiliensis, is confined to Latin America. Brazil, Venezuela,Colombia, Ecuador and Argentina are the main endemic areas.Most exposed subjects develop only an asymptomatic infec-tion, although some individuals present clinical manifestationswhen infected by P. brasiliensis. A wide spectrum of clinical,
* Correspondence: Lucia Mary Singer-Vermes, Depto. de Imuno-logia, Instituto de Ciencias Biomedicas, Universidade de Sao Paulo,Av. Prof. Lineu Prestes, 2415, CEP 05508-900 - Cidade Universitiria,SAo Paulo - S.P., Brasil.
pathological and immunological manifestations can beobserved in these patients. Lungs, lymph nodes, upperdigestive and respiratory tract mucous membranes, adrenals,liver and skin are usually the most affected organs [1,2]. Twomain forms of the disease are distinguished: the acute orsubacute (juvenile type), and the chronic form (adult type).The former runs a rapid course and is usually more severe thanthe latter.
Acute cases are characterized by a marked involvement ofthe reticuloendothelial system, i.e. lymph node hypertrophy,hepatomegaly, and/or splenomegaly. In this clinical form,mucosal involvement is infrequent and pulmonary involve-ment rare. Antibody titres are usually high and severe depres-sion of the cellular immunity is commonly observed.
113

114 L. M. Singer- Vermes et al.
Table 1. Identification, clinical characteristics, magnitude of the DTH responses to paracoccidioidin and levels of serum specific antibody at the timeof the fungal isolation from paracoccidioidomycosis (PCM) patients
DTH SpecificP. brasiliensis Patient Site of Organs/tissues responset antibody Clinical form ofisolate no. Sex isolation* affected* (mm) titrest PCM
Pb44 1 F LN LN; intestines Neg. Neg. Acute(severe)
Pb47 2 M Blood LN; MM; skin Neg. 1:512 Acute(severe)
Pb5O 3 M LN Lungs; MM; skin Neg. 1:64 Chronic(severe)
Pb45 4 M MM Lungs; MM Neg. 1:16 Chronic(moderate)
Pb56 5 M MM Lungs; MM; LN 5 x 5 1:16 Chronic(mild)
Pb57 6 M MM Lungs; MM 12 x 12 1:16 Chronic(mild)
* LN, Lymph nodes; MM, oral mucous membrane.t Cutaneous response to paracoccidioidin prepared according to Fava Netto's method [11].t Measured by immunodiffusion tests using a culture filtrate antigen.
Depending on the degree of dissemination, this form can besubtyped into moderate or severe [1-4].
The chronic and more common form ofPCM can affect oneor more organs/systems. Pulmonary or mucosal involvement isvery frequent. Lymph node enlargement can also be observed,but is not a dominant finding. Depending on the patient'sgeneral condition and on clinical findings, chronic cases areclassified as mild, moderate or severe.
Mild cases are characterized by absent or mild pulmonaryand/or tegumentary involvement. Lymph node hypertrophy,when present, is limited to lymphatic chains of the cephalicsegment, and is of the inflammatory, non-suppurative type.These patients do not exhibit clinical involvement of otherorgans/systems, and their specific serum antibody titres are lowwhile their responses to paracoccidioidin are usually marked[1-4].
At the other extreme, the chronic severe form of the diseaseis characterized by extensive pulmonary lesions. Tegumentarylesions are often present and can be severe. Lymph nodehypertrophy, when present, is not limited to the lymphaticchains of the cephalic segment, and is of the tumoral orsuppurative type. Involvement of other systems/organs, suchas the digestive tract, adrenals, central nervous system andbones, can also be present. These patients usually present highserum specific antibody levels accompanied by negative para-coccidioidin skin test [1-4].
The chronic moderate form of PCM occupies an inter-mediate position between these two poles: specific antibodytitres are moderate, DTH reaction to paracoccidioidin isusually discrete, but involvement of other organs, tracts orsystems is not found.
The broad spectrum of the disease depends on severalfactors associated with the host response. However, character-istics of the infecting agent, mainly its virulence, may alsoinfluence the outcome of PCM [2,3].
Our previous experimental studies using susceptible inbredmice (B1O.A) revealed that distinct P. brasiliensis samples vary
in virulence, immunogenicity and tropism to the differentorgans/systems [5]. These studies, however, were done usingfungal samples maintained by subcultures for many years(some of them for more than 50 years), and it is well knownthat repeated in vitro maintainance alters various characteris-tics of P. brasiliensis, mainly its virulence [6-9].
In this study, the pathogenicity and immunogenicity ofsix recently isolated P. brasiliensis samples obtained frompatients with distinct clinical forms ofPCM were characterizedusing Bl0.A mice. We investigated whether similarities existamong some features of the human disease and those ofinfected mice that could be ascribed to the infecting organ-ism. The pathogenicity and tropism of these fungal samples aswell as the cellular and humoral immune response patternselicited by them in patients and in susceptible inbred mice werecompared.
PATIENTS AND METHODS
Patients andfungal isolatesSix P. brasiliensis samples were isolated from patients living inan endemic area (Botucatu surroundings, Sao Paulo state) andall presented active PCM at the time the fungal samples werecollected. The identification, clinical characteristics, magnitudeof the DTH responses to paracoccidioidin, and levels of serumspecific antibody (evaluated by immunodiffusion tests) at thetime of the fungal isolation of each patient are summarized inTable 1. Pb44 and Pb47 were obtained from patients with acutesevere forms, isolate Pb5O from a chronic severe case, Pb45from a chronic moderate form, while Pb56 and Pb57 wereisolated from benign cases of chronic PCM.
MiceEleven-week-old male B10.A mice, a strain susceptible toP. brasiliensis i.p. infection [10], were used in all experiments.Mice were bred in the University of Sao Paulo animal facilities,and were provided with acidified water and sterilized food.

Comparison between human and experimental paracoccidioidomycosis
Fungal growth conditionsThe six recently isolated fungal samples were used within2 months of being obtained. During this period theP. brasiliensis cells were kept in potato-agar medium overlayedwith mineral oil, to avoid subcultures that could change thefungal characteristics. At the time of animal inoculation, theisolates were cultured and expanded in Fava Netto's medium[11] and maintained in their yeast phase at 36°C for no longerthan 2 weeks.
Preparation of the inocula and infection ofmiceYeast cell suspensions from 7-day-old cultures were washed inPBS and the cells were counted in a haemacytometer. Fungalviability was determined with the Janus Green [12] vital stainand was always > 80%. Mice were infected intraperitoneallywith the appropriate dose of P. brasiliensis yeast cellssuspended in 0-5 ml of PBS.
Pathogenicity studiesThe pathogenicity of each P. brasiliensis isolate was evaluatedby estimation of the 50% lethal dose (LDS0%), enumeration ofviable yeasts in the lungs, and by anatomopathological exami-nation. All analyses were performed blind with respect to thecorresponding clinical form of the patient.
The LD50% value for each fungal sample was estimated bythe probit method [13], using mortality rates obtained with atleast five different doses of yeast cells (ranging from 8 x 104 to18 x 106 viable cells) and groups of 8-12 mice /dose. Mortalitywas scored daily over a 120-day period.
The enumeration of yeast cells in the lungs was done bycounting the number of colony-forming units (CFU) in miceinfected with the distinct P. brasiliensis samples. Five to 12animals infected with 5 x 106 yeast cells were killed at week 16,a time post-infection which clearly discriminates susceptiblefrom resistant animals by the number of CFU in the lungs [14].The lungs from each mouse were collected, weighed anddisrupted using a tissue grinder. The cellular suspensions were
plated on BHI agar supplemented with 4% (v/v) horse serum
and 5% Pb192 culture filtrate, the latter constituting a source
of growth-promoting factor, as described elsewhere [15,16].The plates were sealed, incubated at 37°C and colonies were
counted daily, until no increase in counts was observed. Theresults were expressed in logarithmic values of the CFUcounts/g of tissue.
For the anatomopathological studies, six groups of atleast 30 mice each were infected with 5 x 106 yeast cellscontained in 0-5 ml of PBS. Six to 16 mice from each group
were bled and killed at 2, 4, 12 and 16 weeks after infection, andtheir organs (spleen, omentum, liver, lungs, lymph nodes anddiaphragm) were examined for gross paracoccidioidomycoticgranulomata. The severity of the lesions was graded semi-quantitatively from 0 to 3+, taking into account the numberand size of granulomata.
Cellular immune response
The cellular immune response was always evaluated just beforeautopsy of the same mice used in the anatomopathologicalsurvey by measurement of their DTH reactions. Theseevaluations were done by the footpad test according to pre-
viously described conditions [17] where 25 til of the FavaNetto's antigen [18] were inoculated in the left hind footpad
of the mice. The footpad thickness was measured using acalliper (Mitutoyo, Japan) immediatly before and 24h afterantigen inoculation, and the increase in thickness wascalculated. Non-infected BlO.A mice submitted to the footpadtest were used as controls.
Specific humoral immune responseAntibody titres to P. brasiliensis were determined in the sera ofthe same mice submitted to anatomopathological and DTHstudies. An ELISA as previously described [19] employing aculture filtrate antigen and commercial anti-mouse totalimmunoglobulin peroxidase-labelled (Sigma, St Louis, MO)was used in all experiments.
Statistical analysisThe median scores for the peritoneal lesions were compared bya multisample median test followed by a multiple comparisontest [20,21]. The number of CFU/g of tissue and the antibodytitres were transformed into logarithmic units before statisticalanalysis. One- and two-way analysis of variance followed bymultiple comparisons according to the Tukey method wereapplied to compare respectively the CFU counts in the lungsand the antibody titres induced by the six P. brasiliensis isolatesat the four assayed times post-infection [21]. DTH responses ofinfected animals which were higher than the upper limit of the95% confidence interval of control mice (mean + 2 s.d.) wereconsidered significant.
Correlation studies among the scores for the peritoneallesions of the distinct organs, cellular and humoral immuneresponses were done using Spearman's non-parametricmultiple correlation test [21]. Probability P < 0 05 was consid-ered significant.
RESULTS
Pathogenicity studiesAll P. brasiliensis isolates were lethal to BIO.A mice, althoughan association between the LD50% of each fungal sample formice and the severity of the human disease was not clear. Asshown in Table 2, isolates Pb44 and Pb47, both derived fromacute forms of the disease, presented LD50% of 0-99 x 106 and2-46 x 106, respectively; the LD50% of PbMO, isolated from achronic severe case, was 2 45 x 106, whereas that from thesample derived from a chronic moderate form (Pb45) was1 18 x 106. Mice infected with isolates PbS6 and PbS7, bothobtained from patients with mild PCM, displayed the highestLD50% values: 9-60 x 106 and 2 93 x 106, respectively.
As shown in Table 2, mice infected with Pb5O and Pb57presented significantly higher fungal CFU in the lungs thanthose infected with the other studied isolates, whereas animalsinfected with Pb56 never displayed viable yeasts in this organ.Both Pb56 and Pb57 originated from patients with mildpulmonary forms of PCM and induced respectively thelowest and the highest numbers ofCFU in the lungs of infectedmice. Thus in our experimental model, the fungal tropism topulmonary tissues and the severity of the disease, whenconsidering CFU counts in the lungs, are not parallel tothose observed in the corresponding patient.
Anatomopathological examinations revealed the presenceof peritoneal lesions in all mice infected with Pb44, Pb47, Pb5O,Pb45 and PbS7. A lower percentage ofmice (75%) infected with
115
L. M. Singer-Vermes et al.
Table 2. Pathogenicity of the six recently isolated P. brasiliensis samples, defined by 50% lethal dose (LD50%) estimations,anatomopathology and colony-forming unit (CFU) counts in Bl0.A mice intraperitoneally infected with yeast cells
Cumulative percentage ofmice presenting intense Overall median Mean CFU/g of
P. brasiliensis LD50% paracoccidioidic scores of the pulmonary tissueisolate (x 106 viable yeasts)* granulomatat severity of lesions (log I0)T
Pb44 0.99 100 3 0 502 ± 0-328Pb47 2 46 80 1 0 656 ± 0 286Pb5O 245 90 2 2 527±0 850Pb45 1-18 87 2 NDPb56 9-60 41 1 00±00Pb57 2-93 76 2 3 357 ± 0-523
* Estimated 120 days after infection of B10.A mice with at least five doses of yeast cells, ranging from 8 x 104 to 18 x 106 cells.t Intense (score 2) and/or very intense (score 3) lesions found on at least one out of the six examined organs.t CFU counts were done at 16 weeks after infection.
enc0
U)
.O0e
a)
0
-0)
3.5
3-0
2-5
2.0
I- 5
*0
0.5
0
2 weeks
Fhi4 weeks
16 weeks
~ 4O
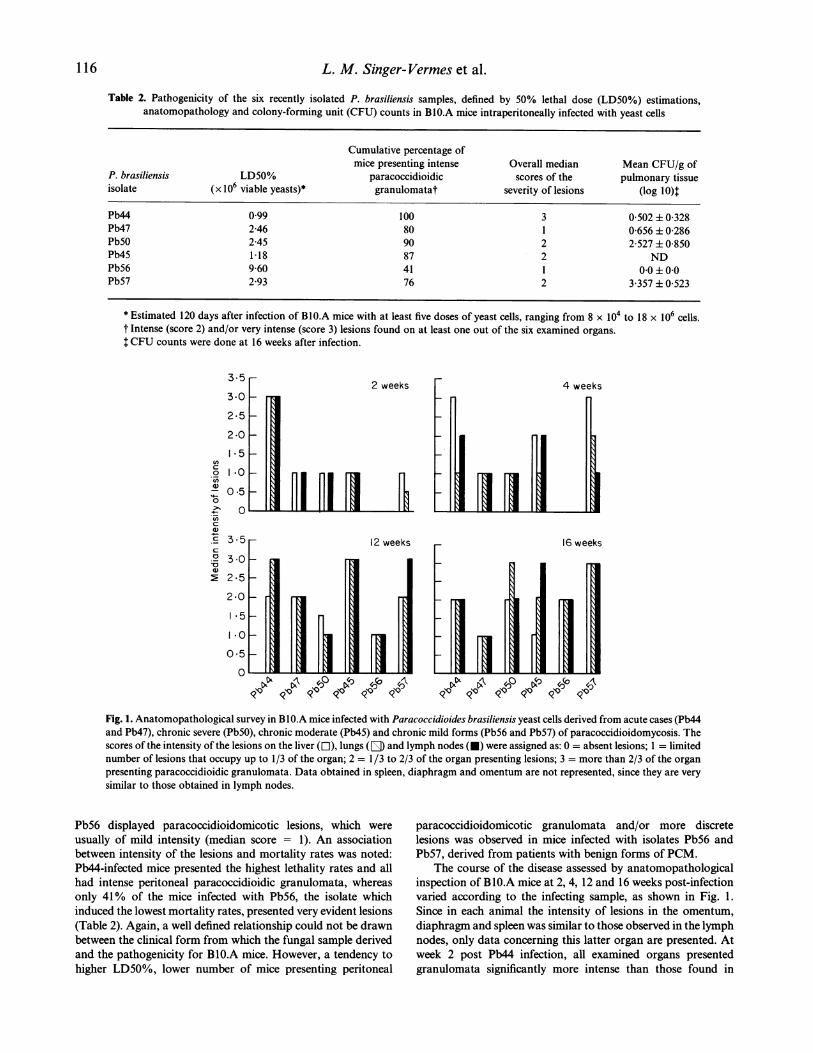
Fig. 1. Anatomopathological survey in BIO.A mice infected with Paracoccidioides brasiliensis yeast cells derived from acute cases (Pb44and Pb47), chronic severe (Pb5O), chronic moderate (Pb45) and chronic mild forms (Pb56 and Pb57) of paracoccidioidomycosis. Thescores of the intensity of the lesions on the liver (El), lungs (E) and lymph nodes (0) were assigned as: 0 = absent lesions; 1 = limitednumber of lesions that occupy up to 1/3 of the organ; 2 = 1/3 to 2/3 of the organ presenting lesions; 3 = more than 2/3 of the organpresenting paracoccidioidic granulomata. Data obtained in spleen, diaphragm and omentum are not represented, since they are verysimilar to those obtained in lymph nodes.
Pb56 displayed paracoccidioidomicotic lesions, which were
usually of mild intensity (median score = 1). An associationbetween intensity of the lesions and mortality rates was noted:Pb44-infected mice presented the highest lethality rates and allhad intense peritoneal paracoccidioidic granulomata, whereasonly 41% of the mice infected with Pb56, the isolate whichinduced the lowest mortality rates, presented very evident lesions(Table 2). Again, a well defined relationship could not be drawnbetween the clinical form from which the fungal sample derivedand the pathogenicity for BIO.A mice. However, a tendency tohigher LD50%, lower number of mice presenting peritoneal
paracoccidioidomicotic granulomata and/or more discretelesions was observed in mice infected with isolates Pb56 andPb57, derived from patients with benign forms of PCM.
The course of the disease assessed by anatomopathologicalinspection of BlO.A mice at 2, 4, 12 and 16 weeks post-infectionvaried according to the infecting sample, as shown in Fig. 1.Since in each animal the intensity of lesions in the omentum,diaphragm and spleen was similar to those observed in the lymphnodes, only data concerning this latter organ are presented. Atweek 2 post Pb44 infection, all examined organs presentedgranulomata significantly more intense than those found in
116
. N=

Comparison between human and experimental paracoccidioidomycosis
(b)
I . I
8 12 16 0 2 4 8 12 16Weeks after infection
I0 2 4 8 12 16
Fig. 2. DTH reaction in BlO.A mice (n = 6-16) infected intraperitoneally with Paracoccidioides brasiliensis yeast cells derived from:(a) acute cases (0 Pb44; A, Pb47); (b) chronic severe (I, PbMO), chronic moderate (*, Pb45); and (c) chronic mild forms (El, Pb56;V, Pb57) of human paracoccidioidomycosis. Range bars represent s.e.m. The horizontal dotted lines denote mean + 2 s.d. (confidenceinterval 95%) of control non-infected mice submitted to the footpad test (n = 22).
- 320ev o 160 Iw_ 207% .0
.0Z:E 20
5 12
0 2 4 8 12 16 2 4 8 12 16Weeks after infection
0 2 4 8 12 16
Fig. 3. Specific antibody production measured by ELISA in BlO.A mice infected intraperitoneally with Paracoccidioides brasiliensisyeast cells derived from acute (0O Pb44; A, Pb47), chronic severe (V, PbMO), chronic moderate (*, Pb45) and from mild cases
(El, Pb56; V, Pb57) of human paracoccidioidomycosis (n = 6 to 16). Range bars denote s.e.m.
mice infected with the other isolates, except Pb45, indicating thatPb44 presents a faster dissemination to all organs compared withthe other fungal samples. Pb56, however, disseminated very
slowly, since no lesions were found at week 2 post-infection,and only 30% of the animals displayed discrete granulomata,restricted to the diaphragm or lymph nodes at week 4.
Specific cellular immunityThe isolates collected from patients with the acute form ofPCM induced distinct DTH profiles in mice (Fig. 2a). Pb44infection did not provoke DTH response at week 2 post-infection, but was able to elicit significant responses at thesubsequent post-infection times, compared with control non-
infected mice. On the other hand, Pb47 induced cellularimmune responses at all assayed points but not at week 12.
Mice infected with Pb5O, obtained from a chronic severe
form of PCM, displayed significant DTH responses at allassayed data points, and Pb45, derived from a chronic moder-ate case, also induced significant responses at all points, exceptat week 16 post-infection (Fig. 2b).
The highest DTH responses were observed in mice infectedwith Pb56, originally obtained from a benign PCM case and theless virulent isolate for B10.A mice. This fungal sample alwaysinduced significant DTH reactions in mice. However, theanimals infected with Pb57, which also derived from a mildPCM form of the human disease, became anergic at week 16post-infection (Fig. 2c).
Independently from the infecting fungal isolate, a tendencyto low or absent DTH responses at the late data points (weeks12 and/or 16 post-infection) was constantly observed.
Specific humoral response
The specific antibody levels induced by infection with the sixP. brasiliensis isolates were much alike until week 12. However,
at week 16 Pb57-infected animals presented significantly lowerantibody levels compared with all other experimental groups
(Fig. 3).
Relationship between the dissemination of the infection, humoraland cellular immune response
Correlation analyses were conducted to evaluate possiblerelationships between the dissemination of the infection,humoral and cellular immune responses, since all mentionedmeasurements were taken in the same animals.
Significant inverse correlations among DTH responses andintensity of the lesions on all examined organs were found inPb56 and in Pb57-infected mice (P varying from < 0 01 to0-014). The intensity of lesions in the lungs of Pb44-infectedanimals was also inversely correlated with their DTH reactions(P = 0 04).
Association among the antibody titres and the intensity ofthe lesions was only found in Pb45-infected animals, in whichlungs and liver presented significant positive correlations withthe antibody levels (P < 0 01 and P = 0 04, respectively).
DISCUSSION
In fungal as in all microbial infections, the genotypes andphenotypes of both hosts and parasites play a crucial role inthe host-fungi interactions, which in turn determine the out-come of the disease. Various experimental studies using murinemodels showed that distinct P. brasiliensis strains vary invirulence [5, 22-27], a fact that may explain at least partiallythe gamut of clinical manifestations of PCM. The knowledgethat can be drawn from experimental models, however, isfrequently hampered by lack of information as to whether themodel mimics the natural disease. To evaluate if there are
similarities among the pathological and immunological
.sEo E0
0
u uO.rcDO
0 2 4
117

L. M. Singer- Vermes et al.
features of human PCM and those found in susceptible B10.A
mice that could be ascribed to the infecting organism, we
compared six P. brasiliensis samples recently isolated from
patients with distinct and well defined clinical forms of PCM.
The degree of virulence of the different P. brasiliensis
isolates for inbred mice can be defined based on both the
LD50% values and the anatomopathological patterns, since
these parameters were shown to be associated. Considering
these data, the relative virulence of the isolates could roughly be
classified as: Pb44 = Pb45 = Pb5O = Pb47 > Pb57 > Pb56. Thus,
an association between the clinical form of the disease from
which the sample derived and the virulence to mice was evident
only when we compared the fungal isolates obtained from the
very mild cases (Pb57 and Pb56) with the samples from more
severe forms of the disease. Our data resemble those obtained
by studies with Coccidioides immitis [28] and Blastomyces
dermatitidis isolates [29], in which the survival times of infected
mice were not fully associated with the severity of the corre-
spondent patient's disease. Although we studied a limited
number of fungal samples, our results suggest that further
investigations using other isolates, especially those obtained
from very mild cases, should be encouraged to confirm whether
an association between human and experimental PCM exists in
these cases.
In previous studies the LD50% of P. brasiliensis samples
maintained for many years in culture were estimated 200 days
post i.p. infection in B10.A mice. The highly virulent Pbl8 and
Pb2052 samples presented LD50% below 2-0 x 106 viable
yeasts; the non-lethal isolates were defined as slightly virulent,
whereas some samples behaved as intermediate and their
LD50% ranged from 9 to 22 x 106 viable yeasts [5]. Due to
the high mortality ratio induced by most fungal samples here
investigated, the ideal post-infection time to compare the
LD50% of all isolates is 120 days. However, we were able to
verify that the LD50% at 200 days after Pb56 infection was
3-8 x 106 viable yeasts, a figure close to those obtained with
Pbl8 and Pb2052. These data confirmed that all recently
isolated samples are highly virulent compared with the fungal
cells maintained for long periods under in vitro conditions.
The fastest dissemination of the disease to all organs and the
lowest LD50% were observed in Pb44-infected mice, whereas
Pb56 induced the highest LD50% and disseminated very
slowly. Thus, the lethality of P. brasiliensis cells in our experi-
mental model may be linked to its precocious dissemination to
vital organs/systems, and the initial weeks of the infection may
be determinant to the outcome of the disease.
The tropism of P. brasiliensis cells in the human host and in
our experimental model is not parallel. Isolates derived from
patients with pulmonary involvement induced distinct evolu-
tionary profiles of paracoccidioidic lesions and extremely
different counts of viable yeasts in the lungs of infected mice.
The fungal load in the lungs was previously shown to be a
reliable discriminant parameter to categorize susceptible and
resistant animals infected with Pbl8 [14]. However, the data
obtained here demonstrated that an association between
lethality and fungal growth in the lungs may not be a general
phenomenon.Neither the cellular nor the humoral specific response
patterns found in infected mice were parallel to those found
in the correspondent patients: Pb57 obtained from a patient
presenting very strong reaction to paracoccidioidin induced
DTH responses in mice which were not higher than thoseobserved in mice infected with isolates collected from anergicpatients. Likewise, the ability to evoke specific humoralimmune response cannot be ascribed to differences in theinfectant agent, since our patients presented distinct levels ofspecific antibody, whereas the antibody titres found in thecorrespondent mice were basically the same, independentlyfrom the infecting isolate.
Our previous investigations demonstrated that BIO.A miceinfected with Pbl 8 are able to mount DTH responses at week 2and week 4 after infection, becoming anergic at subsequenttimes [14,30]. These animals present an increase in the antibodylevels with time, the fungal load in various organs correlateswith the antibody titres and is inversely correlated with theintensity of the DTH reactions [14]. Mice infected with therecently isolated samples were also able to mount an early DTHresponse, and presented depressed DTH responses thereafter.In addition, significant inverse correlation between the severityof pulmonary lesions and the intensity of the DTH reaction wasalso shown in Pb44, Pb56 and Pb57-infected mice. This generalbehaviour of susceptible mice infected with virulent P. brasi-liensis cells indicates that both the capacity to mount an earlyDTH response and the subsequent anergy development may bea host-inherent characteristic, whereas the infecting fungalsample may play a secondary role. On the contrary, thehumoral immune response patterns elicited by the fungalsamples investigated here were not similar to those displayedby Pbl8-infected mice.
The diversity in virulence and immunogenicity displayed bythe six fungal samples assayed here under extremely controlledhomogeneous in vivo conditions, indicates that these isolatesmay constitute different strains of P. brasiliensis. In fact,morphological studies and growth curve determinationsrevealed that these fungi can be classified into two maingroups: isolates with good in vitro growth and markedmorphological heterogeneity (Pb44 and Pb45), and samplesthat present relatively low growth rates and show the charac-teristic morphology of P. brasiliensis (Pb47, Pb5O, Pb56 andPb57) ([31], Burger et al., unpublished data). Furthermore,ongoing studies in our laboratory to classify isolates ofP. brasiliensis by use of mitochondrial and ribosomalenzyme-restricted DNA polymorphisms and by randomlyamplified polymorphic DNA markers suggest that theserecently obtained P. brasiliensis isolates belong to at least twodifferent classes (Delouya et al., unpublished data ).
When comparisons between the human disease and theexperimental model are to be made, we have to consider thatPCM usually presents a very slow evolution. Thus, it is almostimpossible to ascertain for how long the patient is infected andwhat the stage of the disease is at the moment medical assistanceis sought. Furthermore, the experimental route of infection (i.p.)used in this investigation does not mimic the natural disease,since it is usually accepted that humans may be infected byinhalation of airborne conidia or mycelial fragments [1-3]. Evenso, the experimental controlled conditions used in this investiga-tion allowed us to conclude that differences in virulence andimmunogenicity exist among recently isolated P. brasiliensissamples. However, these differences are not sufficient to explainthe severity of the human disease or the tropism to a given organ/system. Host-related factors, such as genetic pattern, sex, age,nutritional and immunological status, as well as the intensity of
118

Comparison between human and experimental paracoccidioidomycosis 119
the fungal load to which humans are submitted, may be ofgreater relevance in the evolution and outcome of PCM.
ACKNOWLEDGMENTS
This work was supported by Fundasao de Amparo a Pesquisa do Estadode Sao Paulo (FAPESP) and Conselho Nacional De Pesquisas (CNPq).We are grateful to Mr R. N. P. Silva for taking care of the animals.
REFERENCES
1 Brummer E, Castafieda E, Restrepo A. Paracoccidioidomycosis: anupdate. Clin Microbiol Revl993; 6:89-117.
2 Franco M, Mendes RP, Moscardi-Bacchi et al. Paracoccidioido-mycosis. Bailliere's Clin Trop Med Commun Dis 1989; 4:185-220.
3 Franco M. Host-parasite relationships in paracoccidioidomycosis.J Med Vet Mycol 1987; 25:5-18.
4 Mendes RP. The gamut of clinical manifestations. In: Franco M,Lacaz CS, Restrepo-Moreno A et al. Paracoccidioidomycosis. BocaRaton: CRC Press, 1994:233-58.
5 Singer-Vermes LM, Burger E, Franco MF et al. Evaluation of thepathogenicity and immunogenicity of seven Paracoccidioides brasi-liensis isolates in susceptible inbred mice. J Med Vet Mycol 1989;27:71-82.
6 San-Blas G, San-Blas F, Serrano LE. Host-parasite relationship inthe yeastlike form of Paracoccidioides brasiliensis strain IVIC Pb 9.Infect Immun 1977; 15:343-6.
7 Kashino SS, Singer-Vermes LM, Calich VLG et al. Alterations inthe pathogenicity of one Paracoccidioides brasiliensis isolate do notcorrelate with its in vitro growth. Mycopathol 1990; 111:173-80.
8 Brummer E, Restrepo A, Hanson LH et al. Virulence of Paracocci-dioides brasiliensis: the influence of in vitro passage and storage.Mycopathol 1990; 109:13-17.
9 Castafieda E, Brummer E, Pappagianis D et al. Chronic pulmonaryand disseminated paracoccidioidomycosis in mice: quantitation ofprogression and chronicity. J Med Vet Mycol 1987; 25:277-87.
10 Calich VLG, Singer-Vermes LM, Siqueira AM et al. Susceptibilityand resistance of inbred mice to Paracoccidioides brasiliensis. Br JExp Pathol 1985;66: 585-94.
11 Fava Netto C. Contribuicalo para o estudo da blastomicose de Lutz.Rev Inst A Lutz (Sao Paulo) 1961; 2:99-194.
12 Berliner MD, Reca ME. Vital staining of Histoplasma capsulatumwith Janus Green B. Sabouraudia 1969;5:26-29.
13 Finney DJ. Probit Analysis, 3rd edn. Cambridge: CambridgeUniversity Press, 1977.
14 Singer-Vermes LM, Caldeira CB, Burger E et al. Experimentalmurine paracoccidioidomycosis: relationship among the dissemina-tion of the infection, humoral and cellular immune responses. ClinExp Immunol 1993; 94:75-79.
15 Castafieda E, Brummer E, Perlman AM et al. A culture medium forParacoccidioides brasiliensis with high plating efficiency, and theeffect of siderophores. J Med Vet Mycol 1988; 26:351-8.
16 Singer-Venues LM, Ciavaglia MC, Kashino SS et al. The source ofthegrowth-promoting factor(s) affects the plating efficiency of Paracoc-cidioides brasiliensis. J Med Vet Mycol 1992; 30:261-4.
17 Fazioli RA, Singer-Vermes LM, Kashino SS et al. Standardizationof the intradermal test in experimental murine paracoccidioidomy-cosis. Braz J Med Biol Res 1986; 19:641A.
18 Fava Netto C, Vega VS, Scianamea I et al. Antigeno polissacaridicodo Paracoccidioides brasiliensis. Estudo do tempo de cultivo doP. brasiliensis necessirio ao preparo do antigeno. Rev Inst MedTrop Sao Paulo 1969; 11:177-81
19 Mendes-Giannini MJS, Camargo, ME, Lacaz CS et al. Immuno-enzymatic absorption test for serodiagnosis of paracoccidioidomy-cosis. J Clin Microbiol 1984; 20:103-8.
20 Levy KJ. Pairwise comparisons associated with the K independentsample median test. Amer Statistics 1979; 33:138-9.
21 Zar JH. Biostatistical analysis, 2nd edn, Englewood Cliffs: Prentice-Hall, 1984.
22 Linares LI, Friedman L. Experimental paracoccidioidomycosis inmice. Infect Immun 1972; 5:681-7.
23 Manocha MS, San-Blas G, Centeno C. Lipid composition ofParacoccidioides brasiliensis: possible correlation with virulence ofdifferent strains. J Gen Microbiol 1980; 117:147-54.
24 Robledo MA, Graybill JR, Ahrens J et al. Host defense againstparacoccidioidomycosis. Am Rev Resp Dis 1982; 125:563-7.
25 Zaccharias D, Ueda A, Moscardi-Bacchi M et al. A comparativehistopathological, immunological and biochemical study ofintravenous paracoccidioidomycosis induced in mice by threeParacoccidioides brasiliensis isolates. J Med Vet Mycol 1986;24:445-54.
26 Acosta MV, Melo L, Castafieda E. Estudio del comportamiento invivo e in vitro de seis cepas de Paracoccidioides brasiliensis. RevistaIberoamericana de Micologia 1992; 9:7-11.
27 Sano A, Miyaji M, Nishimura K et al. Studies on the relationshipbetween pathogenicity of Paracoccidioides brasiliensis in mice andits growth under different oxygen atmospheres. Mycopathol 1991;114:93-101.
28 Friedman L, Smith CE. The comparison of four strains of Cocci-dioides immitis with diverse histories. Mycopathol et Mycol Applic1957; 8:47-53.
29 Moser SA, Koker PJ, Williams JE. Fungal-strain-dependent altera-tions in the time course and mortality of chronic pulmonaryblastomycosis. Infect Immun 1988; 56:34-39.
30 Calich VLG, Fazioli RA, Teixeira HC et al. Mechanisms of hostresistance to Paracoccidioides brasiliensis. In: Torres-Rodrigues,JM, ed. Proceedings of the X Congress of the International Societyfor Human and Animal Mycology. Barcelona. J. R. Prous ScienceEd., 1988:154-9.
31 Burger E, Sugizaki, MF, Kashino SS et al. "In vitro" studies ofP. brasiliensis (Pb) samples recently isolated from patients.Abstracts of the XI Congress of the International Society forHuman and Animal Mycology. Montreal, 1991:173.
Copyright © 2022 FDOKUMEN




















