
Variation in the Group B Streptococcus CsrRS Regulon and Effects on Pathogenicity
11
Published Ahead of Print 18 January 2008. 2008, 190(6):1956. DOI: 10.1128/JB.01677-07. J. Bacteriol. Cieslewicz, Hervé Tettelin and Michael R. Wessels Manuela Puliti, Luciana Tissi, Nikhil Kumar, Michael J. Sheng-Mei Jiang, Nadeeza Ishmael, Julie Dunning Hotopp, Pathogenicity CsrRS Regulon and Effects on Streptococcus Variation in the Group B http://jb.asm.org/content/190/6/1956 Updated information and services can be found at: These include: SUPPLEMENTAL MATERIAL Supplemental material REFERENCES http://jb.asm.org/content/190/6/1956#ref-list-1 at: This article cites 30 articles, 17 of which can be accessed free CONTENT ALERTS more» articles cite this article), Receive: RSS Feeds, eTOCs, free email alerts (when new http://journals.asm.org/site/misc/reprints.xhtml Information about commercial reprint orders: http://journals.asm.org/site/subscriptions/ To subscribe to to another ASM Journal go to: on December 12, 2013 by guest http://jb.asm.org/ Downloaded from on December 12, 2013 by guest http://jb.asm.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Variation in the Group B Streptococcus CsrRS Regulon and Effects on Pathogenicity

Published Ahead of Print 18 January 2008. 2008, 190(6):1956. DOI: 10.1128/JB.01677-07. J. Bacteriol.
Cieslewicz, Hervé Tettelin and Michael R. WesselsManuela Puliti, Luciana Tissi, Nikhil Kumar, Michael J. Sheng-Mei Jiang, Nadeeza Ishmael, Julie Dunning Hotopp, Pathogenicity CsrRS Regulon and Effects on
StreptococcusVariation in the Group B
http://jb.asm.org/content/190/6/1956Updated information and services can be found at:
These include:
SUPPLEMENTAL MATERIAL Supplemental material
REFERENCEShttp://jb.asm.org/content/190/6/1956#ref-list-1at:
This article cites 30 articles, 17 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on Decem
ber 12, 2013 by guesthttp://jb.asm
.org/D
ownloaded from
on D
ecember 12, 2013 by guest
http://jb.asm.org/
Dow
nloaded from

JOURNAL OF BACTERIOLOGY, Mar. 2008, p. 1956–1965 Vol. 190, No. 60021-9193/08/$08.00�0 doi:10.1128/JB.01677-07Copyright © 2008, American Society for Microbiology. All Rights Reserved.
Variation in the Group B Streptococcus CsrRS Regulon and Effectson Pathogenicity�†
Sheng-Mei Jiang,1,2 Nadeeza Ishmael,3 Julie Dunning Hotopp,3 Manuela Puliti,4 Luciana Tissi,4Nikhil Kumar,3 Michael J. Cieslewicz,1,2 Herve Tettelin,3 and Michael R. Wessels1,2*
Division of Infectious Diseases, Children’s Hospital Boston,1 and Harvard Medical School,2 Boston, Massachusetts 02115;The J. Craig Venter Institute, Rockville, Maryland 208503; and Microbiology Section, Department of
Experimental Medicine and Biochemical Sciences, University of Perugia, Perugia, Italy4
Received 16 October 2007/Accepted 8 January 2008
CsrRS (or CovRS) is a two-component regulatory system that controls expression of multiple virulencefactors in the important human pathogen group B Streptococcus (GBS). We now report global gene expressionstudies in GBS strains 2603V/R and 515 and their isogenic csrR and csrS mutants. Together with data reportedpreviously for strain NEM316, the results reveal a conserved 39-gene CsrRS regulon. In vitro phosphorylation-dependent binding of recombinant CsrR to promoter regions of both positively and negatively regulated genessuggests that direct binding of CsrR can mediate activation as well as repression of target gene expression.Distinct patterns of gene regulation in csrR versus csrS mutants in strain 2603V/R compared to 515 wereassociated with different hierarchies of relative virulence of wild-type, csrR, and csrS mutants in murine modelsof systemic infection and septic arthritis. We conclude that CsrRS regulates a core group of genes includingimportant virulence factors in diverse strains of GBS but also displays marked variability in the repertoire ofregulated genes and in the relative effects of CsrS signaling on CsrR-mediated gene regulation. Such variationis likely to play an important role in strain-specific adaptation of GBS to particular host environments andpathogenic potential in susceptible hosts.
Many bacterial species utilize two-component systems(TCS) as a means to regulate gene expression in response tosignals from the environment (1, 27). While several variationsexist, the basic model of a TCS consists of a sensor histidinekinase that usually is positioned at the cell surface or periplas-mic space to interact with external stimuli. Contact with anappropriate stimulus triggers a conformational change in thesensor protein that alters the autokinase activity of its cyto-plasmic domain. Subsequent transfer of the phosphate groupfrom the sensor to a cognate regulator component, in turn,modulates the activity of the regulator as a transcriptionalrepressor or activator of one or more target genes. Coordinateregulation of gene expression in response to environmentalcues may be especially important for an organism like group BStreptococcus (GBS [S. agalactiae]) that exists in a commensalrelationship with its human host as an asymptomatic colonizerof the genital and gastrointestinal tracts but has the potentialto cause invasive infection during pregnancy and childbirth, inthe colonized infant during the first weeks of life, or in thesetting of concomitant chronic illness or advanced age (7, 26).Regulated changes in expression of virulence factors and met-abolic pathways enhance the organism’s adaptation to survivein the varied niches encountered in its existence as a commen-sal or as an invasive pathogen.
In keeping with the adaptation of GBS to diverse host en-
vironments, the genome sequences of GBS strains 2603V/R(hereafter referred to as 2603) and NEM316 revealed 17 and20 predicted TCS, respectively (9, 30). Of these, the CsrRS (orCovRS) system has been investigated most thoroughly. Twoindependent studies reported that inactivation of csrR or csrRSresulted in increased expression of the cyl operon encoding theGBS �-hemolysin/cytolysin and a corresponding increase inhemolytic activity as well as a marked decrease in expression ofcfb and its product, CAMP factor, that enhances the hemolyticactivity of staphylococcal sphingomyelinase (12, 15). Bothgroups demonstrated, as well, that csrR mutants were attenu-ated in rodent models of GBS infection, a finding that sup-ported the importance of CsrRS in pathogenesis.
Transcriptional profiling experiments using genomic mac-roarrays found evidence that CsrRS influenced expression ofmore than 100 genes in the type III GBS strain NEM316 (15).In contrast to the orthologous CsrRS (CovRS) system in Strep-tococcus pyogenes strain MGAS5005, in which CsrR is reportedto act chiefly as a repressor, studies in GBS revealed similarnumbers of activated and repressed genes (10, 15).
These initial studies provided important insights into thefunction of the CsrRS system as a global regulator of GBSgene expression that is likely to play a critical role in patho-genesis of GBS infection. However, many key features of thisimportant regulatory system remain obscure. While the genesencoding CsrRS are highly conserved, the repertoires of genesregulated by the system appear not to be identical among GBSstrains. For example, the cps operon that directs capsular poly-saccharide biosynthesis was regulated to a modest degree inNEM316 (15). In contrast, measurement of transcripts of cpsEand of type-specific capsular polysaccharide revealed no sig-nificant difference between csrR mutants and their respective
* Corresponding author. Mailing address: Division of InfectiousDiseases, Children’s Hospital Boston, 300 Longwood Ave., Boston,MA 02115. Phone: (617) 919-2900. Fax: (617) 730-0254. E-mail:[email protected].
† Supplemental material for this article may be found at http://jb.asm.org/.
� Published ahead of print on 18 January 2008.
1956
on Decem
ber 12, 2013 by guesthttp://jb.asm
.org/D
ownloaded from

wild-type parent strains in either 2603 or 515 (12). A compar-ison of csrS and csrR mutants in strains 2603 and 515 revealedsimilar but less extreme changes in expression of three regu-lated genes in csrS mutants compared to csrR mutants, butwhether both CsrS and CsrR have similar relative effects onexpression of the entire repertoire of regulated genes has notbeen investigated previously (12). Finally, Lamy et al. demon-strated binding of the CsrR protein to the promoter regions ofthree genes whose expression is repressed by CsrR (15). How-ever, it remains to be determined whether genes for whichexpression is activated by CsrR also are regulated by directbinding of CsrR or rather by repression of intermediate regu-lator(s).
In the present investigation, we used genomic microarrays toperform transcriptional profiling of csrR and csrS mutants inthe background of strains 2603 and 515. We compared therepertoire of CsrRS-regulated genes in these strains with thatdescribed previously for strain NEM316 to determine the ex-tent of conservation and diversity of the CsrRS regulon amongGBS strains. We found considerable diversity in the CsrRSregulons and evidence for both activation and repression oftarget genes. Direct binding studies using recombinant CsrRrevealed binding to both positively and negatively regulatedpromoters. Finally, we found that divergent patterns of regu-lation in csrS and csrR mutants were associated with strain-specific alterations in virulence. The results imply that variabil-ity in the CsrRS regulon may contribute to adaptation ofparticular GBS strains to specific host niches.
MATERIALS AND METHODS
Bacterial strains, plasmids, and growth conditions. GBS strains used in thisstudy included type Ia strain 515 (32) and type V strain 2603 (2603V/R) (30) andtheir derivative �csrR and �csrS mutants (12). GBS was grown in liquid culturein Todd-Hewitt broth (THB; Difco), on trypticase soy agar (TSA) supplementedwith 5% defibrinated sheep blood (PML Microbiologicals), or on Todd-Hewittagar supplemented with antibiotics and 5% defibrinated sheep blood. Escherichiacoli DH5� and E. coli M15(pREP4) were grown in Luria-Bertani broth or onLuria-Bertani agar. When appropriate, antibiotics were added at the followingconcentrations: ampicillin, 100 �g/ml; and kanamycin, 25 �g/ml for E. coli. GBSwas grown without shaking in liquid culture. E. coli was grown with shaking at37°C. Plasmid pGEM-T (Promega) was used for the direct cloning of PCRproducts; plasmid pQE30 was used for the expression of recombinant His6-tagged CsrR (Qiagen).
RNA isolation. GBS strains grown overnight on TSA blood agar plates wereinoculated in 10 ml THB broth and collected by centrifugation (3,200 � g, 5 min)at mid-exponential-phase growth (optical density at 650 nm of 0.3). The pelletwas resuspended in 0.5 ml 0.9% NaCl and 1 ml RNA Protect buffer (Qiagen) andkept at room temperature for 5 min. After centrifugation, the bacterial pellet wastreated for 15 min at 37°C with 100 U mutanolysin (Sigma) and 15 mg/mllysozyme (Sigma) in Tris-EDTA buffer, pH 8.0, in a final volume of 100 �l. Totalbacterial RNA was then isolated using an RNeasy mini kit (Qiagen) accord-ing to the manufacturer’s instructions. RNA samples were treated withDNase I (Invitrogen) for 30 min at 37°C to remove any contaminating DNA.The RNA concentration was adjusted to 100 ng/�l, and samples were storedat �80°C until use.
Microarray fabrication. GBS amplicon microarrays were prepared as de-scribed previously (30) using DNA fragments of the annotated open readingframes (ORFs) from GBS strain 2603 (30). Additional primer pairs were de-signed to replace unresponsive 2603 amplicons and to include additional ORFsfrom strain A909, a serotype Ia strain (29). PCR primer pairs were designed withPrimer3 (23) and locally developed Perl scripts. DNA fragments were amplifiedwith these primers using a final concentration of 1� AmpliTaq buffer (AppliedBiosystems, Blanchburg, NJ), 2.5 mM MgCl2, 0.8 mM deoxynucleoside triphos-phate (dNTP) mix (Applied Biosystems), 1.25 U AmpliTaq DNA polymerase(Applied Biosystems), 0.15 �M each primer, and 20 ng/�l DNA with denatur-ation at 95°C for 5 min followed by amplification with 35 cycles at 95°C for 45 s,
55°C for 45 s, and 72°C for 45 s and a final elongation step at 72°C for 10 min.Amplicons were purified using Montage 96 well SEQ plates (Millipore, Billerica,MA) and spotted onto UltraGAPS aminosilane-coated glass slides (Corning,Corning, NY) using 50% dimethyl sulfoxide as the spotting buffer. Ampliconswere bound to the slides by UV cross-linking at 25,000 �J/cm2. Printed slideswere stored until use in a benchtop desiccator. The final array contains ampliconsfor 2,086 genes from strain 2603 and 206 genes from strain A909, representing96.4% and 97.0% of the annotated ORFs in the 2603 and A909 strains, respec-tively. The slide design has been deposited in ArrayExpress as A-TIGR-25,A-TIGR-26, and A-TIGR-27.
Probe labeling and hybridization. Total RNA (2 �g) from each sample wasreverse transcribed into single-stranded cDNA using 1� first strand buffer (In-vitrogen), 10 mM dithiothreitol (DTT), 6 �g random hexamers (Invitrogen), 0.5mM dATP, 0.5 mM dCTP, 0.5 mM dGTP, 0.3 mM dTTP, 0.2 mM aminoallyl-dUTP (Invitrogen), and 400 U SuperScript II reverse transcription (RT) enzyme(Invitrogen). Cy dyes were chemically coupled to the incorporated aminoallyl-dUTP using Cy3- or Cy5-NHS-ester fluorescent dyes (Amersham-Pharmacia,Piscataway, NJ).
Slides were prehybridized in 8% goat serum or 1% bovine serum in 5� SSC(1� SSC is 0.15 M NaCl plus 0.015 M sodium citrate), 0.1% sodium dodecylsulfate (SDS) for 60 min at 42°C, washed in water and then isopropanol, anddried by centrifugation (20). Cy dye-labeled probes from mutant and wild-typestrain RNAs were resuspended in hybridization buffer containing 50% formam-ide, 5� SSC, 0.1% SDS, and 0.6 �g/�l salmon sperm DNA (Applied Biosystems)and hybridized to the microarray slide at 42°C for 16 to 20 h in a sealed,humidified chamber (Corning) (20). Following hybridization, slides were sequen-tially washed in 2� SSC and 0.1% SDS for 10 min at 55°C, 0.1� SSC and 0.1%SDS for 10 min at room temperature, 0.1� SSC for 10 min at room temperature,and deionized water for 5 min at room temperature and then dried by centrif-ugation (20). RNA was separately isolated twice, and probes were prepared (seeabove) and hybridized to the array, resulting in two biological replicates for eachexperimental condition. Technical replication consisted of (i) �3-fold spottedreplication on a single slide; (ii) hybridization of each RNA sample �4 or moretimes, including two dye-swap replicates; and (iii) duplicate or triplicate ampli-cons for a subset of genes. This level of replication resulted in �28 data pointsfor each gene per experimental condition.
Scanning and image analysis. Slides were scanned using an Axon 4000Bmicroarray scanner (Axon Instruments, Union City, CA) at 10-�m resolution.Data were saved as two independent 16-bit TIFF files corresponding to the Cy3and Cy5 channels and were analyzed using TIGR Spotfinder to assess relativeexpression levels (24). Data from TIGR Spotfinder were stored in MAD, arelational database designed to effectively capture microarray data (24). The datafrom this study have been deposited in ArrayExpress (E-TIGR-131).
Data normalization and analysis. To adjust for differences in labeling anddetection efficiencies of the fluorescent labels, the data were normalized usingthe MIDAS software tool (24). A low-intensity filter (100 K) was used to elim-inate background fluorescence; data were normalized with iterative log meancentering (3). Ratios were calculated as the log2 (mutant/wild-type) for all spotswith fluorescence above background. The mean and standard deviation werecalculated from all of the log ratios for a given gene across all technical andbiological replicates using local Perl scripts.
Identification of regulated genes potentially organized in operons. Chromo-somal DNA sequences flanking CsrRS-regulated genes were inspected to iden-tify potential operons on the basis of gene orientation and the presence ofpredicted Rho-independent terminators of transcription. Genes were added tothe final list of regulated genes displayed in Table S1 in the supplementalmaterial if they completed putative CsrRS-regulated operons but did not meetthe threshold value to be classified as up- or down-regulated in the CsrR and/orCsrS mutant(s) or had fewer than 18 hybridization data values.
Search for potential promoter motifs upstream of regulated genes/operons.Intergenic regions upstream of CsrRS-regulated single genes or operons weresearched for the presence of conserved motifs using AlignAce (http://atlas.med.harvard.edu/) and BEST (4). BEST (Binding-site Estimation Suite of Tools)runs, optimizes, and compares the results of four motif-finding programs: Align-Ace, BioProspector, CONSENSUS, and MEME. Intergenic regions were sub-mitted to the programs as follows: (i) all up- and down-regulated genes fromeach experiment, (ii) all up-regulated genes from each experiment, and (iii) alldown-regulated genes from each experiment. Overrepresented motifs were iden-tified and used to search the entire genome sequence of strains 2603 and 515. Foreach motif, the frequency of occurrence and location with regard to adjacentgenes/operons were determined, as well as the extent of CsrRS regulation ofgenes downstream.
VOL. 190, 2008 DIVERSITY OF THE GROUP B STREPTOCOCCAL CsrRS REGULON 1957
on Decem
ber 12, 2013 by guesthttp://jb.asm
.org/D
ownloaded from

qRT-PCR. For real-time quantitative RT-PCR (qRT-PCR), cDNA was gen-erated using 2 �g of total RNA, 2.5 ng random hexamers (Invitrogen), 0.5 mMdNTP mix, and 200 U SuperScript III reverse transcriptase (Invitrogen) in a20-�l reaction mixture with RT at 25°C for 10 min followed by 50°C for 50 minand termination at 85°C for 5 min. Gene-specific primers (Operon Technologies)were designed to amplify �150-bp fragments and to be 100% identical in both2603 and 515 using Primer3 (23). Transcript levels were quantified using theQuantitect SYBR green PCR kit (Qiagen). Briefly, cDNA (2 �l of a 1:100dilution of the above mixture) was used as template in a reaction containing 1�QuantiTect SYBR green mix (Qiagen) and gene-specific primers (see Table S2in the supplemental material). The standard curve for each transcript was gen-erated using a serial dilution of 515 csrS cDNA and gene-specific primers forSAG_0944. Reactions were run on an iCycler iQ (Bio-Rad). The reactions weredenatured at 95°C for 15 min followed by amplification with 45 cycles at 94°C for15 s, 55°C for 30 s, and 72°C for 30 s. Reactions were followed by a melt curveanalysis with a disassociation step at 95°C for 1 min and 55°C for 1 min plus0.5°C/cycle for 80 cycles.
Microarray analysis. A curated set of regulated data (see Table S1 in thesupplemental material) was assembled from (i) genes �2-fold regulated in 2603or 515 with n � 18 and genes more than 2-fold regulated in NEM316; (ii) genes�2-fold regulated in at least two strains with n � 10; and (iii) manual curationto add likely genes cotranscribed with those in the previous two conditions. Therole category composition of this data set was compared to that of the populationof amplicons on the microarray using Fisher’s exact test with a Bonferronistepdown correction for multiple experiments implemented in TIGR MeV (24).A P value of 10�4 was considered significant.
This subset and the complete data set were clustered using various algorithmsin the TIGR MeV software to facilitate analysis of the data. Support trees wereconstructed using Euclidean distance and average linkage with resampling ofexperiment and sample trees by bootstrapping. K-means clustering was per-formed using Euclidean distance and calculated means. Ten clusters were se-lected and converged after eight iterations. Clustering of the subset and thecomplete data set resulted in similar conclusions.
Expression and purification of His6-CsrR. Plasmid vector pQE-30 was usedfor GBS CsrR protein overexpression in E. coli. A 687-bp PCR-generatedBamHI/HindIII DNA fragment corresponding to the CsrR coding sequence wasamplified using specific primers #880 and #881 (see Table S2 in the supplemen-tal material) and then cloned between the BamHI and HindIII sites of pQE-30to give pQE30CsrR. The resulting clone was first transformed into E. coli DH5�for amplification and then isolated and introduced into the E. coli M15 bearingthe plasmid encoding the lac repressor, pREP-4. The transformants were inoc-ulated into LB medium containing both ampicillin (100 �l/ml) and kanamycin(25 �g/ml) with 500 �l of the overnight culture and grown at 37°C until A600 0.5 to 0.7. Expression of His6-CsrR was induced by the addition of isopropyl-�-D-thiogalactopyranoside (IPTG) to 1 mM and then followed by 3 h of incubationat 37°C. Cells were lysed by sonication, and insoluble cell debris was removed bycentrifugation (20,800 � g, 20 min). The cell lysate was then passed over anNi2�-nitrilotriacetic acid agarose column for purification according to the man-ufacturer’s instructions (Qiagen). His6-CsrR was suspended in 50% glycerol andstored at �20°C. The concentration of purified His6-CsrR was estimated bycomparison with a bovine serum albumin standard using the bicinchoninic acidkit (Pierce).
EMSA. For electrophoretic mobility-shift assays (EMSA), pairs of specificoligonucleotide primers were used for the PCR amplification of 250- to 305-bpDNA fragments representing the potential promoter regions of target genes cyl(�-hemolysin), cfb (CAMP factor), and scpB (C5a peptidase) (see Table S2 in thesupplemental material). The purified PCR products were end labeled with �-[S-32P]dATP in the presence of T4 polynucleotide kinase using the Promega gelshift assay system kit. Labeled probes were purified from free nucleotides on aG-25 spin column (Amersham). As a negative control, a similar size DNAfragment was also amplified from the promoter region of the capsule synthesis(cps) locus. His6-CsrR was serially diluted and added to 10 ng of probe DNA inbinding buffer [20 mM Tris (pH 7.5), 1 mM CaCl2, 1 mM DTT, 10 �g/mlpoly(dI-dC) and 100 �g/ml bovine serum albumin] in a total volume of 10 �l.DNA and protein were incubated for 15 min at room temperature. The reactionproducts were mixed with 2 �l of 50% glycerol and loaded onto a 5% Tris-borate-EDTA (TBE) polyacrylamide gel (Bio-Rad; TBE is 90 mM Tris-borate,2 mM EDTA [pH 8.0]). After loading samples, the gel was run at room tem-perature in 0.5� TBE buffer at 350 V for 15 to 20 min and then dried andexposed to X-ray film. In some experiments, purified His6-CsrR was subjected toin vitro phosphorylation before addition to the assay mixture: 10 �g His6-CsrRprotein was incubated for 90 min at 37°C with 32 mM acetyl phosphate in freshly
made phosphorylation buffer (20 mM NaH2PO4, pH 8.0, 10 mM MgCl2, 1 mMDTT) in a total volume of 100 �l.
Mouse virulence studies. Adult outbred male CD-1 mice were obtained fromCharles River Breeding Laboratories (Calco, Italy). The animals were 6 to 8weeks of age at the beginning of each experiment. GBS strains were grownovernight at 37°C in THB (Oxoid, Ltd., Basingstoke, England), washed, anddiluted in RPMI 1640 medium (Gibco Life Technologies, Milan, Italy). Theinoculum size was confirmed by quantitative cultures. Mice were inoculatedintravenously via the tail vein with different infecting doses of GBS in a volumeof 0.5 ml. Control mice were injected by the same route with 0.5 ml of RPMI1640 medium. Mortality was recorded at 24-h intervals for 30 days. The 50%lethal dose (LD50) was calculated by the method of Reed and Muench (22).
GBS-infected mice were evaluated for signs of arthritis and mortality. Afterchallenge, mice were examined daily by two independent observers for 1 monthto evaluate the presence of joint inflammation. Arthritis was defined as visibleerythema and/or swelling of at least one joint. Clinical severity of arthritis wasgraded on a scale of 0 to 3 for each paw, according to changes in erythema andswelling (0 no change; 1 point mild swelling and/or erythema; 2 points moderate swelling and erythema; 3 points marked swelling, erythema, and/orankylosis). Thus, a mouse could have a maximum score of 12. The arthritis index(mean � standard deviation) was calculated by dividing the total score (cumu-lative value of all paws) by the number of animals in each experimental group.
Microarray data accession numbers. The slide design for this study has beendeposited in ArrayExpress as A-TIGR-25, A-TIGR-26, and A-TIGR-27. Themicroarray data from this study have been deposited in ArrayExpress (E-TIGR-131).
RESULTS
To investigate the function of the CsrRS TCS in GBS, wepreviously constructed mutants in the background of GBS typeIa strain 515 and type V strain 2603 (12). A nonpolar inacti-vating mutation was introduced into csrR (515�csrR or2603�csrR) or csrS (515�csrS or 2603�csrS). Analysis of ex-pression patterns of a limited number of genes revealed in-creased expression of cylE (�-hemolysin/cytolysin) and scpB(C5a peptidase) and reduced expression of cfb (CAMP factor)in the csrR mutants in both strain backgrounds and similar,though less extreme, changes in the csrS mutants (12). In thepresent study, we used GBS genomic microarrays as a morecomprehensive means to investigate genome-wide changes ingene expression that result from inactivation of csrR or csrS inthe same two GBS strain backgrounds.
Inactivation of csrR or of csrS was associated with alteredexpression of a large number of genes in both strain back-grounds (Table 1). Using as a threshold a twofold change ingene expression between the mutant and wild type, we foundevidence of CsrRS regulation of 134 genes in strain 2603 and
TABLE 1. Numbers of regulated genes in csrR and csrS mutantsof GBS strain 2603 or 515 relative to their respective
wild-type parent strainsa
Mutant genotype Regulation pattern
No. of genes inGBS strain
background:
2603 515
�csrR Up 94 57Down 13 12Total 107 69
�csrS Up 36 27Down 18 7Total 54 34
a The threshold is a twofold change in transcript abundance.
1958 JIANG ET AL. J. BACTERIOL.
on Decem
ber 12, 2013 by guesthttp://jb.asm
.org/D
ownloaded from

80 genes in strain 515. One significant difference between the2603 and 515 strains was the presence of 12 IS1381 transposasesubunits found to be up-regulated in 2603�csrR. A glycosyl-transferase (SAG_1548, SAG_1551) in the up-regulated andcotranscribed SAG_1548 to -1555 is disrupted by an IS1381transposase (SAG_1549, SAG_1550). Although the IS1381transposase genes are on the opposite strand relative to thecotranscribed unit, the amplicon-based microarray queriesboth the transcript and its reverse complement. Therefore,up-regulation of genes SAG_1548 to -1555 results in what islikely to be artifactual “up-regulation” of the inserted trans-posase. Further, the high nucleotide identity of all the trans-posases results in positive results for all chromosomal loca-tions. Therefore, the IS1381 transposases were removed fromsubsequent analyses, as it is unlikely that this apparent up-regulation is biologically relevant.
Bioinformatics software was used to search for putativeCsrR-binding motifs associated with CsrRS-regulated individ-ual genes and operons in the genome sequences of strains 2603and 515. Among a number of candidate motifs, we failed toidentify any that were both overrepresented in intergenic re-gions and preferentially located upstream of CsrRS-regulatedgenes. Therefore, this analysis did not support the earlier sug-gestion of a distinct CsrR binding sequence (15) but suggestedrather that CsrR recognizes regions of DNA that are notreadily identified by a canonical nucleotide sequence motif.
Validation of microarray results by qRT-PCR. To confirmthe changes in transcript abundance observed by microarrayhybridization, we performed qRT-PCR for a subset of regu-lated genes using RNA samples from mutant and wild-typestrains. Nine genes were selected for qRT-PCR testing usingRNA samples from 515�csrR, 515�csrS, 2603�csrR, and2603�csrS, and their respective wild-type parent strains. Thechange (fold) in gene expression by qRT-PCR correlated wellwith those calculated from the microarray experiments in eachof the mutant strains (R2 0.94; see Fig. S1 and Table S3 inthe supplemental material).
Characterization of the CsrRS regulon in GBS strains 2603and 515. Transcriptional analysis using genomic microarraysconfirmed the previously reported regulation of known or pu-tative virulence factors including the cyl operon (SAG_0662 to-0673) encoding the GBS �-hemolysin/cytolysin, scpB (SAG_1236; C5a peptidase), and cfb (SAG_2043, CAMP factor), aswell as a second gene transcriptionally linked to cfb (SAG_2042) that is predicted to encode a rhodanese-like protein ofunknown function (see Table S1 in the supplemental material).A second locus encoding a predicted protein with 67% aminoacid identity to C5a peptidase was also regulated by CsrRS(SAG_0416). As reported previously, and in contrast to theearlier analysis of GBS strain NEM316, we did not find aconsistent pattern of regulation of the cps capsular polysaccha-ride synthetic operon in strain 2603. In 515�csrR, there was atrend of down-regulation of the cps operon, but this trend onlyreached the twofold threshold for cps1aJ (SAL_1283) andcps1aH (SAL_1285) among the 16 genes in the cps operon.
Regulated genes in one or both strains encoded proteinsinvolved in a wide range of cell functions including known orpredicted virulence factors, transporters of amino acids, pep-tides, sugars, and metals, and proteins that mediate adaptationto environmental stresses (see Table S1 in the supplemental
material). Among the various functional categories of regu-lated genes, transporters of amino acids, peptides, and amineswere most significantly overrepresented in the study (P 10�7), followed by transport and binding proteins (P 10�5)and pathogenesis genes (P 10�5). We found evidence forregulation of expression of several proteins that are predictedto be secreted or surface associated, including SAG_0297(aminopeptidase C), SAG_1002 (putative protease), SAG_1890 (putative endopeptidase O), and two operons predictedto encode membrane proteins (SAG_0364 to -365 and SAG_0798 to -799).
Genes encoding proteins involved in transport of a variety ofsubstrates constituted a large group of regulated genes. Theseincluded the oppA1-F operon (SAG_0148 to -0152), encodingan oligopeptide ABC transporter that has been implicated inmodulating the attachment of GBS to host cells and the adcoperon (SAG_0154 to -0156), homologs of which encode azinc/manganese transporter in Streptococcus pneumoniae and amanganese acquisition and homeostasis system in Streptococ-cus gordonii (6, 16, 25). Also regulated by CsrRS are homologsof a system involved in iron transport (SAG_1007–1010).
In keeping with the inferred role of CsrRS in adaptation tochanging environments, the system regulates several stress re-sponse mechanisms in GBS. These included homologs ofAphC and AphF (SAG_1833 to -1834), two components of thealkyl hydroperoxide reductase of S. pyogenes. In that species,the alkyl hydroperoxide reductase system contributes to scav-enging endogenous hydrogen peroxide and has been linked tovirulence in a murine infection model (2). Homologs of theenterococcal and lactococcal general stress protein Gls24(SAG_1135 and SAG_1137) were also regulated by CsrRS inGBS. Gls24 has been implicated in stress response and viru-lence in Enterococcus faecalis (18, 28). The GBS CsrRS alsoregulates expression of two separate operons predicted to en-code components of a glycine/betaine osmoregulation system(SAG_1796 to -1797 and SAG_0241 to -0244), a system thatmediates adaptation to osmotic stress in Bacillus subtilis andLactococcus lactis (13, 14, 19, 31). Although their role in GBSis undefined, expression of two predicted transcriptional reg-ulators is also controlled by CsrRS: SAG_0712 encoding aputative regulator of the OmpR family and SAG_0938 encod-ing a predicted GntR family transcriptional regulator (11, 27).
Comparison of the CsrRS regulons in three different GBSstrains. The availability of genome-wide transcriptional profil-ing data in strains 2603 and 515 together with the previouslydescribed results in strain NEM316 provided the opportunityto compare the CsrRS regulons in independent GBS isolatesrepresenting the three most important capsular serotypes inhuman infection, types Ia (strain 515), III (strain NEM316),and V (strain 2603). This analysis revealed a core group of 39genes whose expression was changed as a result of inactivationof csrR and/or csrS in all three strain backgrounds (Fig. 1 andTable 2). Two-way comparisons showed further overlap in therepertoire of CsrRS-regulated genes, with 16 genes regulatedin both 2603 and 515, 18 in 2603 and NEM316, and 3 in 515and NEM316. For each of the three strains, certain CsrRS-regulated genes were regulated in only one strain background.A higher number of uniquely regulated genes were identifiedin NEM316, but this may be a result of the differences in arrayplatform and mutant design between the two studies. These
VOL. 190, 2008 DIVERSITY OF THE GROUP B STREPTOCOCCAL CsrRS REGULON 1959
on Decem
ber 12, 2013 by guesthttp://jb.asm
.org/D
ownloaded from

combined results suggest that CsrRS regulates a conservedcore group of genes in multiple GBS strains, including thosecoding for the important virulence factors �-hemolysin, C5apeptidase, and CAMP factor, as well as a large repertoire ofgenes whose regulation varies among GBS isolates.
Differential effects on gene regulation of inactivating CsrRversus CsrS. Mutation of csrR in strain 2603 was associatedwith increased expression of 94 genes and reduced expressionof 13 genes. This pattern suggests that CsrR acts predomi-nantly as a transcriptional repressor but that it can also activategene transcription, directly or indirectly. Inactivation of csrSresulted in expression changes of a smaller number of geneswith 36 genes up-regulated in 2603�csrS and 18 genes down-regulated (Table 1). For 27 genes, we observed altered expres-sion in both 2603�csrR and in 2603�csrS (Fig. 2). In 23 of these27 genes, gene expression was up-regulated in both 2603�csrRand 2603�csrS. For most regulated genes, mutation of csrRproduced a greater effect than did mutation of csrS (see Fig. S2in the supplemental material). We observed a change in ex-pression in 2603�csrR but not in 2603�csrS for 80 genes (Fig.2). For the majority of these genes, the change in expression in2603�csrS was in the same direction as that in 2603�csrR, butit did not reach the twofold threshold. For two linked genes,SAG_1706 (hypothetical protein) and SAG_1707 (putativedrug resistance transporter), expression was increased in2603�csrR and decreased in 2603�csrS.
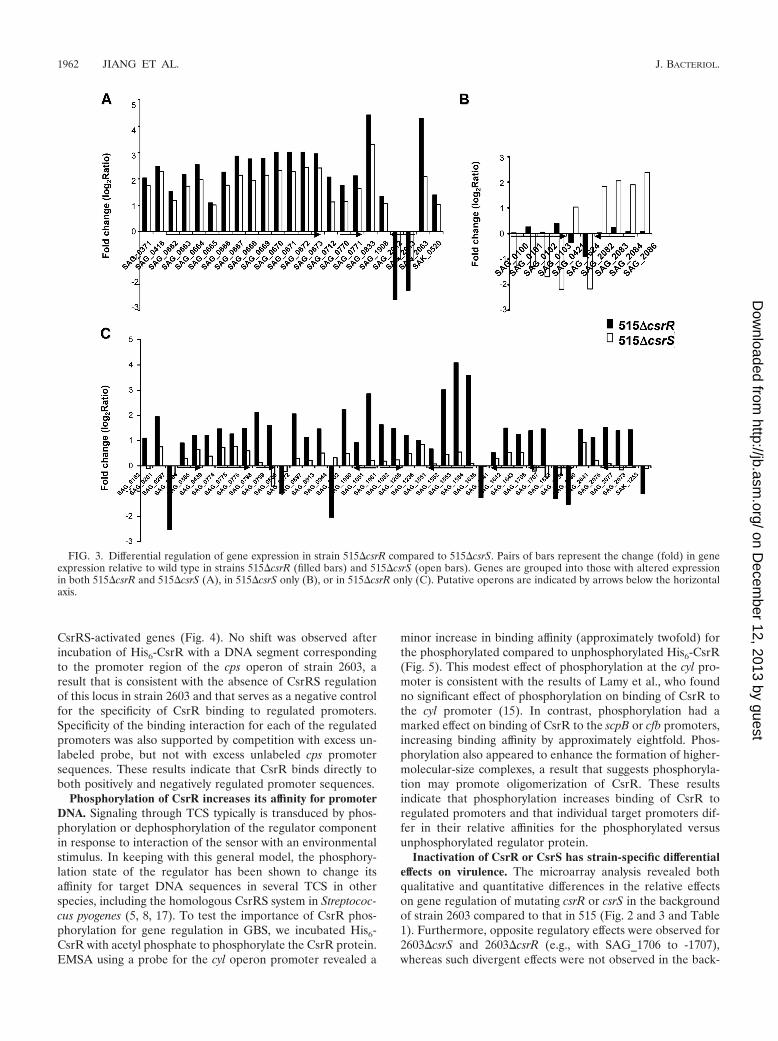
While the total number of CsrRS-regulated genes was some-what lower in strain 515 than in 2603 (Table 1), we observed asimilar pattern with respect to the relative effects of mutationin csrR versus csrS. That is, the predominant pattern was agreater effect on target gene expression in 515�csrR than in515�csrS, but for several genes, the effect was greater in515�csrS than in 515�csrR (Fig. 3).
CsrR binds directly to promoter regions of both activatedand repressed genes. An earlier investigation of GBS strainNEM316 used DNase I protection and EMSA to demonstratedirect binding of recombinant CsrR to a DNA segment up-stream of the cyl operon (15). DNase I protection experimentsalso suggested direct binding to the promoter regions of twoother genes whose expression, like that of the cyl operon, is
repressed by CsrRS. To further characterize the interaction ofCsrR with regulated promoters, we expressed CsrR as a His6
fusion in Escherichia coli and purified the recombinant proteinby Ni2�-affinity chromatography. His6-CsrR was used inEMSA with DNA probes from strain 2603 that correspondedto the promoter regions of the cyl operon and scpB (C5apeptidase), two genes whose expression is repressed by CsrRS;and cfb (CAMP factor), a gene whose expression is activatedby CsrRS. For all three promoters, band shifts were observedin the presence of His6-CsrR, indicating direct binding of CsrRto DNA sequences upstream of both CsrRS-repressed and
TABLE 2. Conserved genes of the CsrRS regulon in threeGBS strains
Locus Gene product name or function
Relative geneexpression (CsrR orCsrRS mutant/wild
type)
2603 515 NEM316
SAG_0201 Oxidoreductase, putative 12.76 3.87 3.61SAG_0267 Conserved hypothetical protein 6.47 6.1 4.47SAG_0297 Aminopeptidase C 0.12 0.17 0.29SAG_0371 Hypothetical protein 9.93 4.08 25.81SAG_0416 Serine peptidase, S8/S53 family 6.81 5.54 2.14SAG_0662 CylX protein 3.45 2.86 6.28SAG_0663 CylD protein 7.93 4.46 5.50SAG_0664 CylG protein 10.94 5.79 5.06SAG_0665 Acyl carrier protein AcpC 5.39 2.13 7.01SAG_0666 CylZ protein 7.93 4.75 5.66SAG_0667 CylA protein 12.41 7.25 4.41SAG_0668 CylB protein 9.6 6.75 4.11SAG_0669 CylE protein 9.95 6.84 1.84SAG_0670 CylF protein 12.13 7.99 4.82SAG_0671 CylI protein 12.54 8.05 3.84SAG_0672 CylJ protein 11.92 8.03 3.92SAG_0673 CylK protein 12.73 7.73 4.69SAG_0712 DNA-binding response
regulator6.76 4.2 4.23
SAG_0770 Membrane protein, putative 5.62 3.36 1.54SAG_0771 Cell wall surface anchor family
protein6.82 4.31 2.69
SAG_0913 Chloramphenicolacetyltransferase
3.36 2.75 3.84
SAG_1002 Protease, putative 5.65 4.68 3.73SAG_1050 Ribonucleotide reductase,
truncation2.86 1.88 2.36
SAG_1051 Aspartate-semialdehydedehydrogenase
10.95 7.21 2.81
SAG_1206 Conserved domain protein 4.01 2.29 6.28SAG_1548 Glycosyl transferase, group 2
family protein, interruption C17.52 16.68 10.48
SAG_1551 Glycosyl transferase, group 2family protein, interruption N
3.5 1.59 10.48
SAG_1552 Conserved hypothetical protein 10.68 8.09 8.94SAG_1553 Hypothetical protein 20.43 16.92 16.45SAG_1554 Hypothetical protein 13.61 11.98 9.92SAG_1641 YaeC family protein 1.99 1.43 3.32SAG_1642 ABC transporter, substrate-
binding protein3.91 2.82 3.84
SAG_1643 Glutamine amidotransferase,class I
3.43 2.37 2.81
SAG_1706 Conserved hypothetical protein 2.08 2.61 2.81SAG_1890 Endopeptidase O 3.87 2.73 3.01SAG_1908 Hypothetical protein 15.31 2.52 31.13SAG_2042 Rhodanese-like domain protein 0.13 0.17 0.44SAG_2043 CAMP factor 0.18 0.21 0.06SAG_2063 Pathogenicity protein, putative 23.78 19.57 57.28
FIG. 1. Conserved and strain-specific gene regulation by CsrRS.Numbers within the circle corresponding to each GBS strain representthe number of genes that are CsrRS regulated in one, two, or all threestrains. The number of regulated genes shown here is lower than thatin Table 1 and elsewhere in the text because this analysis only includedgenes for which expression data were evaluable in all three strainbackgrounds.
1960 JIANG ET AL. J. BACTERIOL.
on Decem
ber 12, 2013 by guesthttp://jb.asm
.org/D
ownloaded from
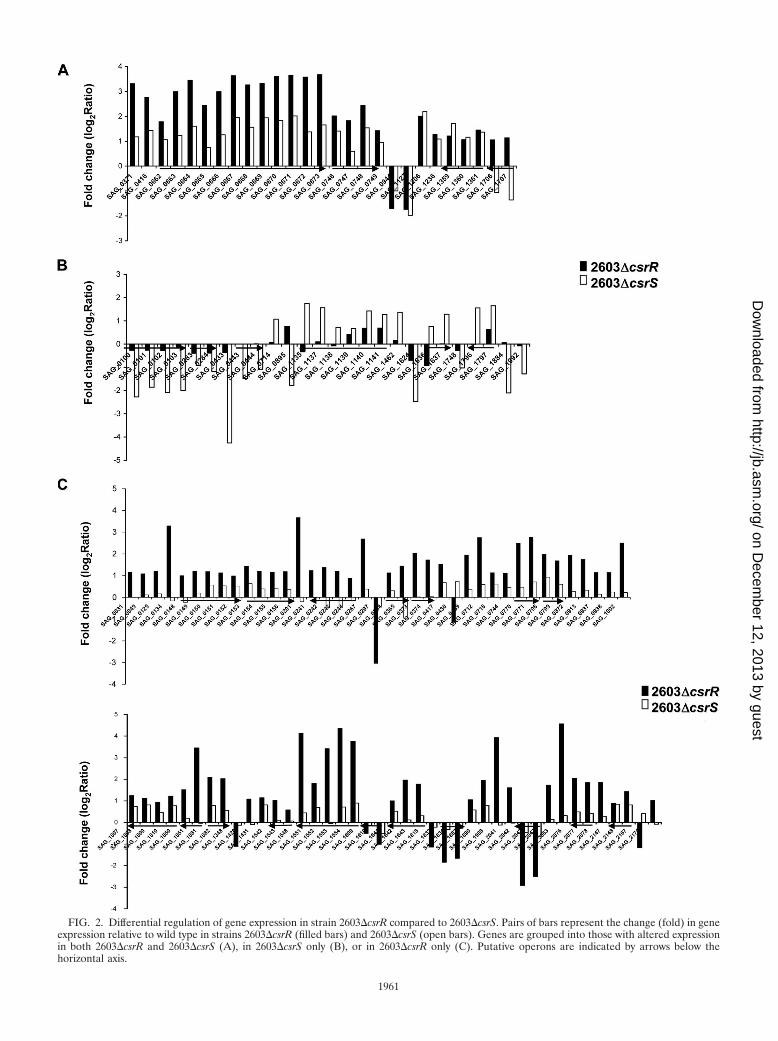
FIG. 2. Differential regulation of gene expression in strain 2603�csrR compared to 2603�csrS. Pairs of bars represent the change (fold) in geneexpression relative to wild type in strains 2603�csrR (filled bars) and 2603�csrS (open bars). Genes are grouped into those with altered expressionin both 2603�csrR and 2603�csrS (A), in 2603�csrS only (B), or in 2603�csrR only (C). Putative operons are indicated by arrows below thehorizontal axis.
1961
on Decem
ber 12, 2013 by guesthttp://jb.asm
.org/D
ownloaded from
CsrRS-activated genes (Fig. 4). No shift was observed afterincubation of His6-CsrR with a DNA segment correspondingto the promoter region of the cps operon of strain 2603, aresult that is consistent with the absence of CsrRS regulationof this locus in strain 2603 and that serves as a negative controlfor the specificity of CsrR binding to regulated promoters.Specificity of the binding interaction for each of the regulatedpromoters was also supported by competition with excess un-labeled probe, but not with excess unlabeled cps promotersequences. These results indicate that CsrR binds directly toboth positively and negatively regulated promoter sequences.
Phosphorylation of CsrR increases its affinity for promoterDNA. Signaling through TCS typically is transduced by phos-phorylation or dephosphorylation of the regulator componentin response to interaction of the sensor with an environmentalstimulus. In keeping with this general model, the phosphory-lation state of the regulator has been shown to change itsaffinity for target DNA sequences in several TCS in otherspecies, including the homologous CsrRS system in Streptococ-cus pyogenes (5, 8, 17). To test the importance of CsrR phos-phorylation for gene regulation in GBS, we incubated His6-CsrR with acetyl phosphate to phosphorylate the CsrR protein.EMSA using a probe for the cyl operon promoter revealed a
minor increase in binding affinity (approximately twofold) forthe phosphorylated compared to unphosphorylated His6-CsrR(Fig. 5). This modest effect of phosphorylation at the cyl pro-moter is consistent with the results of Lamy et al., who foundno significant effect of phosphorylation on binding of CsrR tothe cyl promoter (15). In contrast, phosphorylation had amarked effect on binding of CsrR to the scpB or cfb promoters,increasing binding affinity by approximately eightfold. Phos-phorylation also appeared to enhance the formation of higher-molecular-size complexes, a result that suggests phosphoryla-tion may promote oligomerization of CsrR. These resultsindicate that phosphorylation increases binding of CsrR toregulated promoters and that individual target promoters dif-fer in their relative affinities for the phosphorylated versusunphosphorylated regulator protein.
Inactivation of CsrR or CsrS has strain-specific differentialeffects on virulence. The microarray analysis revealed bothqualitative and quantitative differences in the relative effectson gene regulation of mutating csrR or csrS in the backgroundof strain 2603 compared to that in 515 (Fig. 2 and 3 and Table1). Furthermore, opposite regulatory effects were observed for2603�csrS and 2603�csrR (e.g., with SAG_1706 to -1707),whereas such divergent effects were not observed in the back-
FIG. 3. Differential regulation of gene expression in strain 515�csrR compared to 515�csrS. Pairs of bars represent the change (fold) in geneexpression relative to wild type in strains 515�csrR (filled bars) and 515�csrS (open bars). Genes are grouped into those with altered expressionin both 515�csrR and 515�csrS (A), in 515�csrS only (B), or in 515�csrR only (C). Putative operons are indicated by arrows below the horizontalaxis.
1962 JIANG ET AL. J. BACTERIOL.
on Decem
ber 12, 2013 by guesthttp://jb.asm
.org/D
ownloaded from

ground of strain 515. To investigate whether such differentialeffects of CsrR compared to CsrS might be reflected in therelative pathogenic potential of the mutant strains, we testedthe virulence of csrR and csrS mutants and wild-type strains2603 and 515 in a murine model of systemic infection andseptic arthritis. Mice were challenged intravenously with vari-ous doses of GBS and observed for development of signs ofarthritis and for mortality. Wild-type strain 515 was the mostvirulent in these studies, with an LD50 of 7.2 � 104. The LD50
for 515�csrR was 320-fold higher at 2.3 � 107, whereas515�csrS had an intermediate level of virulence (LD50 of 8.5 �106). This hierarchy of relative virulence is the same as thatreported previously for these strains in a murine intraperito-neal challenge model (12). Similarly, strain 2603�csrR wasattenuated in virulence (no deaths at challenge doses up to 108
CFU) relative to wild-type strain 2603 (LD50 of 2.8 � 107
CFU), as reported previously for the intraperitoneal challengemodel (12). In striking contrast, strain 2603�csrS was morevirulent (LD50 of 2.4 � 106 CFU) than wild-type 2603. Thisvirulence hierarchy was reflected not only in the relative le-thality of the three strains in the 2603 background but also inseverity of arthritis, whether scored by number of affectedjoints or by clinical severity index (Fig. 6).
These results demonstrate that differential patterns of reg-ulation by CsrRS in strains 2603 and 515 are associated withstriking differences in the overall relative virulence of csrSmutants in the two strain backgrounds. In particular, we ob-served increased expression of certain genes in 2603�csrS, butnot in 2603�csrR or in either mutant in the 515 background. Of
these, possible virulence genes include SAG_1135 and SAG_1137 that encode homologs of Gls24, a stress response proteinshown to contribute to virulence in experimental enterococcalinfection (18, 28). A similar pattern of regulation was noted forSAG_1796 and SAG_1797, which are predicted to encode aglycine/betaine osmoregulation system implicated in adapta-tion to osmotic stress in other species (13, 14, 19, 31). Differ-ential CsrS-dependent regulation of these loci in the 2603strain background may account for the unexpectedly high vir-ulence of strain 2603�csrS.
FIG. 4. Binding of purified His6-CsrR to promoter regions ofCsrRS-regulated genes. EMSA were performed using 32P-labeled DNAfragments corresponding to the promoter region of the indicated gene oroperon and purified His6-CsrR protein. Lanes contain 32P-labeled pro-moter sequence only (lane 1), 32P-labeled promoter sequence plus His6-CsrR protein (lane 2), 32P-labeled promoter sequence plus His6-CsrRprotein plus excess unlabeled promoter sequence (lane 3), or 32P-labeledpromoter sequence plus His6-CsrR protein plus excess negative con-trol promoter sequence (from the cpsA promoter in panels A to C andfrom the recA promoter in panel D). Band shifts indicating CsrRbinding are observed for promoter regions of genes whose expressionis up-regulated (cfb, CAMP factor) or down-regulated (cyl operon,�-hemolysin; and scpB, C5a peptidase), but not for the promoter of theunregulated cps operon encoding capsular polysaccharide biosyntheticproteins.
FIG. 5. Phosphorylation of CsrR enhances binding to regulatedpromoters. EMSA were performed using 32P-labeled DNA fragmentscorresponding to the promoter region of the indicated gene or operonand increasing concentrations of purified His6-CsrR protein withoutprior treatment (untreated) or after incubation with acetyl phosphate(phosphorylated). Band shifts indicating binding of CsrR were ob-served at an approximately twofold lower concentration of CsrR afterphosphorylation for the cyl operon promoter, but at approximatelyeightfold lower concentration after phosphorylation for the cfb andscpB promoters.
FIG. 6. Relative virulence of wild-type strain 2603, 2603�csrS, or2603�csrR in a murine arthritis model. The values shown represent theclinical arthritis index after intravenous challenge with 1 � 107 CFU ofthe indicated GBS strain. Data represent means � standard deviationsfor three independent experiments, each using 10 animals per exper-imental group.
VOL. 190, 2008 DIVERSITY OF THE GROUP B STREPTOCOCCAL CsrRS REGULON 1963
on Decem
ber 12, 2013 by guesthttp://jb.asm
.org/D
ownloaded from

DISCUSSION
Results of the present investigation provide several new in-sights into the CsrRS TCS in GBS and its potential functionsduring infection. Transcriptional profiling studies usinggenomic microarrays yielded a comprehensive picture of globalgene regulation by CsrRS in GBS strains 2603 and 515 inaddition to that reported previously for strain NEM316. Wefound evidence for an extensive regulon in all three strains,including genes that encode products known or predicted toenhance bacterial survival under varied conditions and/or tocontribute to pathogenicity in the human or animal host. Mi-croarray hybridization experiments and qRT-PCR confirmedCsrRS regulation of three major virulence determinants iden-tified in earlier studies: �-hemolysin, CAMP factor, and C5a-peptidase. There was also evidence of regulation of severalpredicted surface or secreted proteins of unknown functionthat may participate in GBS adherence or in modification ofthe local host environment.
The largest functional class of CsrRS-regulated genes en-coded transport systems for various small molecules includingpeptides, amino acids, sugars, and metals. This broad regula-tion of small molecule transporters is consistent with the pos-tulated role of CsrRS in mediating adaptation of GBS to variedenvironmental and nutritional circumstances encounteredwithin the colonized or infected host. We also found evidencefor CsrRS regulation of stress response systems such as theOpuA betaine uptake osmoprotection system. Adaptation toosmotic stress may be especially important for GBS survival atmucosal sites in which fluid and solute shifts result in changingosmotic conditions. Similarly, CsrRS appears to regulate ex-pression of genes encoding alkyl hydroperoxide reductase, anenzyme implicated in resistance to endogenous hydrogen per-oxide stress in S. pyogenes. Such resistance may be adaptiveduring interaction of GBS with host phagocytes or under othercircumstances of oxidative stress.
Comparison of the repertoire of genes regulated by CsrRSin three different GBS strains suggests that there is a “coreregulon” of genes that are regulated by CsrRS in multiplestrains, but that there is substantial diversity in the remainderof the regulon. For the three strains studied to date, the coreregulon consists of 39 genes, including the virulence factorsdescribed in earlier reports. Thirty-seven genes show evidenceof CsrRS regulation in two of the three strains, while 155 genesare regulated in only one strain. Such variation in CsrRS regu-lons is likely to be a reflection of the overall genomic variabilityin this species (29). Heterogeneity in gene regulation amongindividual isolates may account, in part, for differences in ad-aptation to specific host environments and for the pathogenicpotential of particular strains. The molecular basis for variabil-ity in CsrRS repertoire remains to be determined and mayinvolve multiple factors, including strain-specific variation inpromoter sequences of regulated genes and the presence orabsence of additional interacting regulators.
By characterizing changes in gene regulation in csrS mutantsas well as csrR mutants, we uncovered another level of com-plexity in CsrRS-mediated gene regulation. For most regulatedgenes, we observed a similar trend in gene expression in thecsrS mutant as in the csrR mutant, but the change in geneexpression was of a lower magnitude in the csrS mutant. How-
ever, in a significant minority of cases, mutation in csrS, but notin csrR, resulted in a change in gene expression, or the degreeof change in expression was greater in the csrS mutant than inthe csrR mutant. In two cases, we observed divergent effects ongene regulation in the csrS compared to csrR mutants. Themost common pattern of regulation—a greater effect by inac-tivating CsrR than CsrS—is consistent with the basic TCSmodel: removal of the transcriptional regulator has a maximaleffect, usually by derepressing target gene transcription. Inac-tivation of the sensor may have a similar, but often lesser, effectby preventing signaling that activates or inactivates the regu-lator. However, other patterns of responses are also possible.An equivalent effect of inactivating CsrS might be expected forgenes whose promoter regions bind phospho-CsrR with muchhigher affinity than the unphosphorylated CsrR, assumingCsrR phosphorylation is dependent on CsrS signaling. Inagreement with this formulation, we found heterogeneity inthe importance of CsrR phosphorylation in determining therelative binding affinity of CsrR for different target promotersin vitro. Alternatively, or in addition, differential effects couldbe mediated by interaction of either CsrS or CsrR with othertranscriptional regulators with various specificities for CsrRS-regulated genes. For example, evidence has been presentedthat the serine/threonine kinase Stk1 interacts with CsrR toregulate expression of �-hemolysin and CAMP factor in GBS(21).
Strain-to-strain variation in the CsrRS regulon and in therelative regulatory effects of the sensor and regulator compo-nents implies that GBS strains vary not only in gene contentbut also in their dynamic capacity to adapt to changing envi-ronments in the host. A particular repertoire of CsrRS generegulation may confer a survival advantage or pathogenic po-tential in a specific microenvironment. This conclusion is sup-ported by our finding that the virulence of csrR and csrS mu-tants relative to their respective parent strains differs in strains2603 and 515. Thus, an individual strain may be better adaptedfor survival in a particular host site such as the bovine mam-mary gland or the human gastrointestinal or genitourinarytract. In this way, the species as a whole has enhanced adap-tation to varied host niches. Together, these results providefurther evidence that CsrRS serves as a global regulatory sys-tem in GBS that functions in both conserved and variable waysto enhance adaptation of this important pathogen for survivalin the host.
ACKNOWLEDGMENTS
We thank Dennis L. Kasper for helpful advice.This work was supported in part by NIH grant AI59502.
REFERENCES
1. Beier, D., and R. Gross. 2006. Regulation of bacterial virulence by two-component systems. Curr. Opin. Microbiol. 9:143–152.
2. Brenot, A., K. Y. King, and M. G. Caparon. 2005. The PerR regulon inperoxide resistance and virulence of Streptococcus pyogenes. Mol. Micro-biol. 55:221–234.
3. Causton, H. C., J. Quackenbush, and A. Brazma. 2003. Microarray geneexpression data analysis: a beginner’s guide. Blackwell Science Ltd., Oxford,United Kingdom.
4. Che, D., S. Jensen, L. Cai, and J. S. Liu. 2005. BEST: binding-site estimationsuite of tools. Bioinformatics 21:2909–2911.
5. Dalton, T. L., and J. R. Scott. 2004. CovS inactivates CovR and is requiredfor growth under conditions of general stress in Streptococcus pyogenes. J.Bacteriol. 186:3928–3937.
6. Dintilhac, A., and J. P. Claverys. 1997. The adc locus, which affects compe-
1964 JIANG ET AL. J. BACTERIOL.
on Decem
ber 12, 2013 by guesthttp://jb.asm
.org/D
ownloaded from

tence for genetic transformation in Streptococcus pneumoniae, encodes anABC transporter with a putative lipoprotein homologous to a family ofstreptococcal adhesins. Res. Microbiol. 148:119–131.
7. Doran, K. S., and V. Nizet. 2004. Molecular pathogenesis of neonatal groupB streptococcal infection: no longer in its infancy. Mol. Microbiol. 54:23–31.
8. Gao, J., A. A. Gusa, J. R. Scott, and G. Churchward. 2005. Binding of theglobal response regulator protein CovR to the sag promoter of Streptococ-cus pyogenes reveals a new mode of CovR-DNA interaction. J. Biol. Chem.280:38948–38956.
9. Glaser, P., C. Rusniok, C. Buchrieser, F. Chevalier, L. Frangeul, T. Msadek,M. Zouine, E. Couve, L. Lalioui, C. Poyart, P. Trieu-Cuot, and F. Kunst.2002. Genome sequence of Streptococcus agalactiae, a pathogen causinginvasive neonatal disease. Mol. Microbiol. 45:1499–1513.
10. Graham, M. R., L. M. Smoot, C. A. Migliaccio, K. Virtaneva, D. E. Sturdevant,S. F. Porcella, M. J. Federle, G. J. Adams, J. R. Scott, and J. M. Musser. 2002.Virulence control in group A Streptococcus by a two-component gene regula-tory system: global expression profiling and in vivo infection modeling. Proc.Natl. Acad. Sci. USA 99:13855–13860.
11. Haydon, D. J., and J. R. Guest. 1991. A new family of bacterial regulatoryproteins. FEMS Microbiol. Lett. 63:291–295.
12. Jiang, S.-M., M. J. Cieslewicz, D. L. Kasper, and M. R. Wessels. 2005.Regulation of virulence by a two-component system in group B Streptococ-cus. J. Bacteriol. 187:1105–1113.
13. Kappes, R. M., B. Kempf, and E. Bremer. 1996. Three transport systems forthe osmoprotectant glycine betaine operate in Bacillus subtilis: characteriza-tion of OpuD. J. Bacteriol. 178:5071–5079.
14. Kappes, R. M., B. Kempf, S. Kneip, J. Boch, J. Gade, J. Meier-Wagner, andE. Bremer. 1999. Two evolutionarily closely related ABC transporters me-diate the uptake of choline for synthesis of the osmoprotectant glycinebetaine in Bacillus subtilis. Mol. Microbiol. 32:203–216.
15. Lamy, M. C., M. Zouine, J. Fert, M. Vergassola, E. Couve, E. Pellegrini, P.Glaser, F. Kunst, T. Msadek, P. Trieu-Cuot, and C. Poyart. 2004. CovS/CovR of group B streptococcus: a two-component global regulatory systeminvolved in virulence. Mol. Microbiol. 54:1250–1268.
16. Loo, C. Y., K. Mitrakul, I. B. Voss, C. V. Hughes, and N. Ganeshkumar.2003. Involvement of the adc operon and manganese homeostasis in Strep-tococcus gordonii biofilm formation. J. Bacteriol. 185:2887–2900.
17. Miller, A. A., N. C. Engleberg, and V. J. Dirita. 2001. Repression of virulencegenes by phosphorylation-dependent oligomerization ofCsrR at target pro-moters in S. pyogenes. Mol. Microbiol. 40:976–990.
18. Nannini, E. C., F. Teng, K. V. Singh, and B. E. Murray. 2005. Decreasedvirulence of a gls24 mutant of Enterococcus faecalis OG1RF in an experi-mental endocarditis model. Infect. Immun. 73:7772–7774.
19. Obis, D., A. Guillot, J.-C. Gripon, P. Renault, A. Bolotin, and M.-Y. Mistou.1999. Genetic and biochemical characterization of a high-affinity betaineuptake system (BusA) in Lactococcus lactis reveals a new functional organi-zation within bacterial ABC transporters. J. Bacteriol. 181:6238–6246.
20. Peterson, S., R. T. Cline, H. Tettelin, V. Sharov, and D. A. Morrison. 2000.Gene expression analysis of the Streptococcus pneumoniae competence regu-lons by use of DNA microarrays. J. Bacteriol. 182:6192–6202.
21. Rajagopal, L., A. Vo, A. Silvestroni, and C. E. Rubens. 2006. Regulation of
cytotoxin expression by converging eukaryotic-type and two-component sig-nalling mechanisms in Streptococcus agalactiae. Mol. Microbiol. 62:941–957.
22. Reed, L. J., and H. Muench. 1938. A simple method of estimating fifty percent endpoints. Am. J. Hyg. 27:493–497.
23. Rozen, S., and H. Skaletsky. 2000. Primer3 on the WWW for general usersand for biologist programmers, p. 365–386. In S. Krawetz and S. Misener(ed.), Bioinformatics methods and protocols: methods in molecular biology.Humana Press, Totowa, NJ.
24. Saeed, A. I., V. Sharov, J. White, J. Li, W. Liang, N. Bhagabati, J. Braisted,M. Klapa, T. Currier, M. Thiagarajan, A. Sturn, M. Snuffin, A. Rezantsev,D. Popov, A. Ryltsov, E. Kostukovich, I. Borisovsky, Z. Liu, A. Vinsavich, V.Trush, and J. Quackenbush. 2003. TM4: a free, open-source system formicroarray data management and analysis. BioTechniques 34:374–378.
25. Samen, U., B. Gottschalk, B. J. Eikmanns, and D. J. Reinscheid. 2004.Relevance of peptide uptake systems to the physiology and virulence ofStreptococcus agalactiae. J. Bacteriol. 186:1398–1408.
26. Schuchat, A., and J. Wenger. 1994. Epidemiology of group B streptococcaldisease. Epidemiol. Rev. 16:374–402.
27. Stock, A. M., V. L. Robinson, and P. N. Goudreau. 2000. Two-componentsignal transduction. Annu. Rev. Biochem. 69:183–215.
28. Teng, F., E. C. Nannini, and B. E. Murray. 2005. Importance of gls24 invirulence and stress response of Enterococcus faecalis and use of the Gls24protein as a possible immunotherapy target. J. Infect. Dis. 191:472–480.
29. Tettelin, H., V. Masignani, M. J. Cieslewicz, C. Donati, D. Medini, N. L.Ward, S. V. Angiuoli, J. Crabtree, A. L. Jones, A. S. Durkin, R. T. Deboy,T. M. Davidsen, M. Mora, M. Scarselli, I. Margarit y Ros, J. D. Peterson,C. R. Hauser, J. P. Sundaram, W. C. Nelson, R. Madupu, L. M. Brinkac,R. J. Dodson, M. J. Rosovitz, S. A. Sullivan, S. C. Daugherty, D. H. Haft, J.Selengut, M. L. Gwinn, L. Zhou, N. Zafar, H. Khouri, D. Radune, G.Dimitrov, K. Watkins, K. J. O’Connor, S. Smith, T. R. Utterback, O. White,C. E. Rubens, G. Grandi, L. C. Madoff, D. L. Kasper, J. L. Telford, M. R.Wessels, R. Rappuoli, and C. M. Fraser. 2005. Genome analysis of multiplepathogenic isolates of Streptococcus agalactiae: implications for the micro-bial “pan-genome.” Proc. Natl. Acad. Sci. USA 102:13950–13955.
30. Tettelin, H., V. Masignani, M. J. Cieslewicz, J. A. Eisen, S. Peterson, M. R.Wessels, I. T. Paulsen, K. E. Nelson, I. Margarit, T. D. Read, L. C. Madoff,A. M. Wolf, M. J. Beanan, L. M. Brinkac, S. C. Daugherty, R. T. DeBoy, A. S.Durkin, J. F. Kolonay, R. Madupu, M. R. Lewis, D. Radune, N. B. Fedorova,D. Scanlan, H. Khouri, S. Mulligan, H. A. Carty, R. T. Cline, S. E. Van Aken,J. Gill, M. Scarselli, M. Mora, E. T. Iacobini, C. Brettoni, G. Galli, M.Mariani, F. Vegni, D. Maione, D. Rinaudo, R. Rappuoli, J. L. Telford, D. L.Kasper, G. Grandi, and C. M. Fraser. 2002. Complete genome sequence andcomparative genomic analysis of an emerging human pathogen, serotype VStreptococcus agalactiae. Proc. Natl. Acad. Sci. USA 99:12391–12396.
31. van der Heide, T., and B. Poolman. 2000. Osmoregulated ABC-transportsystem of Lactococcus lactis senses water stress via changes in the physicalstate of the membrane. Proc. Natl. Acad. Sci. USA 97:7102–7106.
32. Wessels, M. R., L. C. Paoletti, A. K. Rodewald, F. Michon, J. DiFabio, H. J.Jennings, and D. L. Kasper. 1993. Stimulation of protective antibodiesagainst type Ia and Ib group B streptococci by a type Ia polysaccharide-tetanus toxoid conjugate vaccine. Infect. Immun. 61:4760–4766.
VOL. 190, 2008 DIVERSITY OF THE GROUP B STREPTOCOCCAL CsrRS REGULON 1965
on Decem
ber 12, 2013 by guesthttp://jb.asm
.org/D
ownloaded from




















