
Kinetics and Pathogenicity of Equine Herpesvirus9 Infection following Intraperitoneal Inoculation in...
11
EXPERIMENTALLY-INDUCED DISEASE Kinetics and Pathogenicity of Equine Herpesvirus-9 Infection following Intraperitoneal Inoculation in Hamsters E. El-Nahass *,‡ , N. El-Habashi *,x , M. Nayel †,{ , S. Kasem † , H. Fukushi † , Y. Suzuki * , A. Hirata * , H. Sakai * and T. Yanai * *Department of Veterinary Pathology, † Department of Veterinary Microbiology, Faculty of Applied Biological Science, Gifu University, 1-1 Yanagido, Gifu 501-1193, Japan, ‡ Departments of Veterinary Pathology, Beni-Suef University, x Kafr El-Shikh University, and { Department of Animal Medicine and Infectious Diseases, Faculty of Veterinary Medicine, Sadat City, Menofiya University, Egypt Summary The kinetics of infection and pathogenicity of equine herpesvirus-9 (EHV-9) was studied in a hamster model. Five-week-old Syrian hamsters and 5-day-old suckling hamsters were inoculated intraperitoneally with 10 5 and 4 10 4 plaque-forming units of EHV-9, respectively. EHV-9 antigens were detected by immunocyto- chemistry in the peritoneal macrophages, which may be the primary site of virus attachment and propagation at 6 h post inoculation (hpi). At 12 hpi, viral antigen was observed in the abdominal nerves and ganglia (mainly the coeliac ganglia). Virus antigen was detected in the dorsal root (spinal) ganglia, in parts of the spinal cord (particularly the mid-lumbar area) and in the myenteric plexuses at 36, 48 and 72 hpi, respectively. At 96 hpi, virus antigen was detected in the most caudal part of the brain. Polymerase chain reaction conducted on samples of the blood, spinal cord and brain revealed EHV-9 DNA in the spinal cord at 36 hpi and in the blood at 48 hpi and for 4 days after this initial detection. It is suggested that after initial propagation in the abdominal macrophages, EHV-9 infected the abdominal ganglia or myenteric plexuses and then travelled to the brain via the peripheral nerves and spinal cord. Examination of other organs also revealed the presence of EHV-9, suggesting that the virus might infect tissues other than those of the nervous system. Ó 2011 Elsevier Ltd. All rights reserved. Keywords: EHV-9; hamster model; neuropathogenesis Introduction Equine herpesvirus-9 (EHV-9), a varicellovirus in the subfamily Alphaherpesvirinae, is the newest member of the equine herpesviruses and is closely related to equine herpesvirus-1 (EHV-1). EHV-9 was first described in an outbreak of disease in Thomson’s gazelles (Gazella thomsoni) in a Japanese zoo (Fukushi et al., 1997) and subsequently in a giraffe (Giraffe camelopardalis reticulata) with signs of encephalitis (Borchers et al., 2005; Kasem et al., 2008). Recently, EHV-9 was detected in a polar bear with progressive encephalitis (Donovan et al., 2009), raising fears of emerging infections in various wild and zoo animal species. Emerging EHV-9 infection is also a concern with respect to domestic animals, because the virus has exhibited a wide range of susceptible hosts and has been shown to be easily transmittable via the nasal route in goats (Taniguchi et al., 2000), pigs (Narita et al., 2000), dogs and cats (Yanai et al., 2003a, b) and common marmosets (Kodama et al., 2007). The pathogenesis of EHV-9 following nasal infec- tion has been studied in a suckling hamster model. EHV-9 propagated in the olfactory epithelium 12e24 h post inoculation (hpi) and then gained ac- cess to the brain through the olfactory and trigeminal nerves (El-Habashi et al., 2010a). Correspondence to: T. Yanai (e-mail: [email protected]). 0021-9975/$ - see front matter Ó 2011 Elsevier Ltd. All rights reserved. doi:10.1016/j.jcpa.2011.01.009 J. Comp. Path. 2011, Vol. 145, 271e281 Available online at www.sciencedirect.com www.elsevier.com/locate/jcpa
Transcript of Kinetics and Pathogenicity of Equine Herpesvirus9 Infection following Intraperitoneal Inoculation in...

J. Comp. Path. 2011, Vol. 145, 271e281 Available online at www.sciencedirect.com
www.elsevier.com/locate/jcpa
EXPERIMENTALLY-INDUCED DISEASE
Kinetics and Pathogenicity of EquineHerpesvirus-9Infection following Intraperitoneal Inoculation in
Hamsters
Cor
002
doi
E. El-Nahass*,‡, N. El-Habashi*,x, M. Nayel†,{, S. Kasem†, H. Fukushi†,Y. Suzuki*, A. Hirata*, H. Sakai* and T. Yanai*
*Department of Veterinary Pathology, †Department of Veterinary Microbiology, Faculty of Applied Biological Science, Gifu
University, 1-1 Yanagido, Gifu 501-1193, Japan, ‡Departments of Veterinary Pathology, Beni-Suef University, xKafrEl-Shikh University, and {Department of Animal Medicine and Infectious Diseases, Faculty of Veterinary Medicine, Sadat
City, Menofiya University, Egypt
resp
1-99
:10.1
Summary
The kinetics of infection and pathogenicity of equine herpesvirus-9 (EHV-9) was studied in a hamster model.Five-week-old Syrian hamsters and 5-day-old suckling hamsters were inoculated intraperitoneally with 105
and 4� 104 plaque-forming units of EHV-9, respectively. EHV-9 antigens were detected by immunocyto-chemistry in the peritoneal macrophages, which may be the primary site of virus attachment and propagationat 6 h post inoculation (hpi). At 12 hpi, viral antigen was observed in the abdominal nerves and ganglia(mainly the coeliac ganglia). Virus antigen was detected in the dorsal root (spinal) ganglia, in parts of thespinal cord (particularly the mid-lumbar area) and in the myenteric plexuses at 36, 48 and 72 hpi, respectively.At 96 hpi, virus antigenwas detected in themost caudal part of the brain. Polymerase chain reaction conductedon samples of the blood, spinal cord and brain revealed EHV-9 DNA in the spinal cord at 36 hpi and in theblood at 48 hpi and for 4 days after this initial detection. It is suggested that after initial propagation in theabdominal macrophages, EHV-9 infected the abdominal ganglia or myenteric plexuses and then travelledto the brain via the peripheral nerves and spinal cord. Examination of other organs also revealed the presenceof EHV-9, suggesting that the virus might infect tissues other than those of the nervous system.
� 2011 Elsevier Ltd. All rights reserved.
Keywords: EHV-9; hamster model; neuropathogenesis
Introduction
Equine herpesvirus-9 (EHV-9), a varicellovirus in thesubfamily Alphaherpesvirinae, is the newest memberof the equine herpesviruses and is closely related toequine herpesvirus-1 (EHV-1). EHV-9 was firstdescribed in an outbreak of disease in Thomson’sgazelles (Gazella thomsoni) in a Japanese zoo (Fukushiet al., 1997) and subsequently in a giraffe (Giraffecamelopardalis reticulata) with signs of encephalitis(Borchers et al., 2005; Kasem et al., 2008). Recently,EHV-9 was detected in a polar bear with progressive
ondence to: T. Yanai (e-mail: [email protected]).
75/$ - see front matter
016/j.jcpa.2011.01.009
encephalitis (Donovan et al., 2009), raising fears ofemerging infections in various wild and zoo animalspecies. Emerging EHV-9 infection is also a concernwith respect to domestic animals, because the virushas exhibited a wide range of susceptible hosts andhas been shown to be easily transmittable via the nasalroute in goats (Taniguchi et al., 2000), pigs (Naritaet al., 2000), dogs and cats (Yanai et al., 2003a, b)and common marmosets (Kodama et al., 2007).
The pathogenesis of EHV-9 following nasal infec-tion has been studied in a suckling hamster model.EHV-9 propagated in the olfactory epithelium12e24 h post inoculation (hpi) and then gained ac-cess to the brain through the olfactory and trigeminalnerves (El-Habashi et al., 2010a).
� 2011 Elsevier Ltd. All rights reserved.

272 E. El-Nahass et al.
EHV-9 may also infect hamsters by the ocular, oraland intraperitoneal routes (El-Habashi et al., 2010b).Animals infected via the oral and peritoneal routesexhibitedmilder lesions and fewer viral antigen-positivecells in the glomerular and mitral cell layers of theolfactory bulb than animals infected via the nasal route.This observation suggests that the virus most likely doesnot enter the brain through the olfactory nerve, butmay enter through other nerves. Further investigationsare needed to determine exactly how EHV-9 reachesthe brain following infection by non-nasal routesand to identify the pathogenesis of EHV-9-induced encephalitis following these routes ofadministration. In particular, the primary site of prop-agation of the virus has not yet been identified, nor is itknown how the virus gains access to the brain by theseroutes. Another unanswered question is the time re-quired for the virus to travel to the brain. The patho-genesis of EHV-1 induced encephalitis and myelitisfollowing intraperitoneal inoculation has been studiedpreviously (Hasebe et al., 2002), and other neurotropicviruses, such asTheiler’s virus andpoliovirus, have beenstudied in the CBA mouse model using different routesof infection in order to elucidate their detailed neuropa-thogenicity (Villarreal et al., 2006).
The aims of the present study were (1) to determinethe primary site of virus propagation following intra-peritoneal inoculation of hamsters with EHV-9, (2) tocharacterize the pathway by which EHV-9 accessesthe central nervous system (CNS), (3) to define thekinetics of development of encephalitis induced byEHV-9 and (4) to determine whether EHV-9 hasany effect on the internal organs of infected animals.
Materials and Methods
Virus Culture
Madin-Darby bovine kidney (MDBK) cells were usedfor the propagation of EHV-9. The inocula were pre-paredby culturing the virus from the original seed stocksof EHV-9 (P19, 5th passage inMDBK cells). The viruswas titrated by plaque-forming assay on MDBK cells.
Animals
Four 5-week-old male and twelve 10-week-old femaleSyrian hamsters that were 10 days pregnant were pur-chased from a breeder (SLC Inc, Hamamatsu,Japan). The animals were housed in plastic cagesand kept in an isolated biohazard cabinet for approx-imately 1 week of acclimatization. The animals wereprovided with a basal pellet diet (Oriental MF, Ori-ental Yeast Co., Tokyo, Japan) and bottled waterad libitum. The experiment was conducted in accor-dance with laws and Standard Operating Procedures
related to the use of laboratory animals. The experi-mental protocol was approved by the Animal Exper-iment Committee of the Faculty of Applied BiologicalScience at Gifu University.
Virus Inoculation
Four 5-week-old hamsters were injected intraperitone-ally with a single dose of 105 plaque-forming units(PFUs) of EHV-9 virus and were maintained until theend of the experiment (15 days post inoculation [dpi])or the appearance of neurological signs. The 12 preg-nant dams were left to give birth, after which fifty 5-day-old suckling hamsters were inoculated intraperito-neally with 4� 104 PFU of EHV-9 virus. Of the fivesuckling hamsters that were killed at intermediate inter-vals of 6, 12, 24, 36, 48, 72, 96, 120, 144 and168 hpi, fourwere subjected to histopathological and immunohisto-chemical analysis and one provided samples for poly-merase chain reaction (PCR) studies. Five uninfectedanimals were kept as a control group. The animalswere checked for clinical signs at least three times daily.
Collection of Peritoneal Cavity Cells
Cells were collected from the peritoneal cavity as de-scribed by Goafa et al. (1996). After death, the ab-dominal skin was treated with 70% ethanol andthen the outer skin layer was opened and 10 ml ofcold phosphate buffered saline (PBS; pH 7.2,0.01 M) containing 3% fetal calf serum was injectedinto the abdominal cavity using a 27-gauge needle at-tached to a 5 ml syringe. The abdomen was gentlymassaged and then the PBS solution was carefully as-pirated using a 25-gauge needle attached to a 5 ml sy-ringe. The cell suspension obtained was centrifuged at400g for 8 min. The supernatant was discarded andthe cells were resuspended in PBS in order to preparesmears for immunocytochemistry (ICC).
Pathological Investigations
The brain, different levels of spinal cord, lungs, liver,spleen and small and large intestines were taken fromthe juvenile hamsters and fixed in 10%neutral bufferedformalin. The suckling hamsters were bisected in thesagittal plane and were fixed similarly. Samples wereprocessed routinely and embedded in paraffinwax. Sec-tions (5 mm) were stained with haematoxylin and eosin(HE). The suckling hamsters were not decalcified.
Fixed tissue sections were also subjected to immu-nohistochemistry (IHC) by the avidinebiotincomplex (ABC) method as described previously(Yanai et al., 1998). The primary antibody wasa rabbit antibody specific for EHV-9 (1 in 800dilution; Veterinary Microbiology Laboratory, Gifu
Equine Herpesvirus-9 Infection of Hamsters 273
University) and the secondary antibody wasbiotinylated anti-rabbit immunoglobulin (Ig) G(DAKO Cytomation, Fort Collins, Colorado) with‘visualization’ of the reaction by application of liquid3,30-diaminobenzidine substrate chromogen system(DAKO Cytomation) and haematoxylin counter-stain. Tissue sections from the EHV-9-infectedhamsters and sera from a non-immunized rabbitand goat were used as controls.
For ICC, cytological preparations of the abdomi-nal cell suspensions were air dried for 30 min, fixedin cooled acetone and immunolabelled with theEHV-9 antiserum as described above.
DNA Extraction and Polymerase Chain Reaction
Blood and fresh tissues (including the spinal cord andbrain) were subjected to DNA extraction with a Sepa-gene kit for virus DNA detection (Sanko Pharmaco-logicals, Tokyo, Japan). Virus DNA was detectedusing primers for the open-reading frame (ORF)76-F (50-TTT CCC TCT CAG CGA TCA CTT
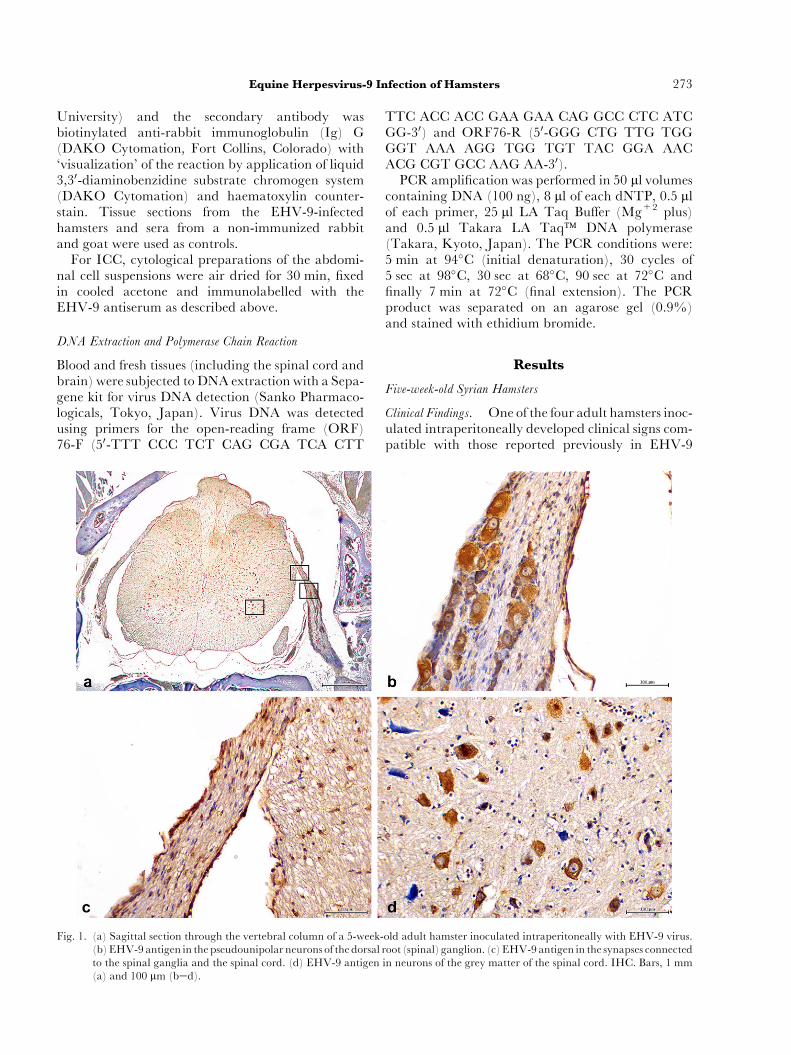
Fig. 1. (a) Sagittal section through the vertebral column of a 5-week-(b)EHV-9 antigen in thepseudounipolar neurons of the dorsal rto the spinal ganglia and the spinal cord. (d) EHV-9 antigen(a) and 100 mm (bed).
TTC ACC ACC GAA GAA CAG GCC CTC ATCGG-30) and ORF76-R (50-GGG CTG TTG TGGGGT AAA AGG TGG TGT TAC GGA AACACG CGT GCC AAG AA-30).
PCR amplification was performed in 50 ml volumescontaining DNA (100 ng), 8 ml of each dNTP, 0.5 mlof each primer, 25 ml LA Taq Buffer (Mg+2 plus)and 0.5 ml Takara LA Taq� DNA polymerase(Takara, Kyoto, Japan). The PCR conditions were:5 min at 94�C (initial denaturation), 30 cycles of5 sec at 98�C, 30 sec at 68�C, 90 sec at 72�C andfinally 7 min at 72�C (final extension). The PCRproduct was separated on an agarose gel (0.9%)and stained with ethidium bromide.
Results
Five-week-old Syrian Hamsters
Clinical Findings. One of the four adult hamsters inoc-ulated intraperitoneally developed clinical signs com-patible with those reported previously in EHV-9
old adult hamster inoculated intraperitoneally with EHV-9 virus.oot (spinal) ganglion. (c)EHV-9antigen in the synapses connectedin neurons of the grey matter of the spinal cord. IHC. Bars, 1 mm

274 E. El-Nahass et al.
infection. These included weight loss, crouching pos-ture, nasal and lachrymal discharges and salivation.By the second day, this animal became hyper-reactiveto stimuli and displayed occasional tremors associatedwith uncoordinated movements of the limbs.
Pathological Findings. At necropsy examination, noneof the inoculated animals exhibited gross abnormali-ties. Microscopically, all of the hamsters inoculatedwith EHV-9 had lymphocytic meningomyelitis andmeningoencephalitis characterized by neuronaldegeneration and necrosis, varying degrees of gliosis,perivascular aggregates of lymphocytes, plasma cellsand neutrophils, with mild focal or diffuse lympho-cytic infiltration of the meninges. The affectedneuronal cells displayed severe chromatolysis orkaryorrhexis. The degenerate neuronal cells oftenhad intranuclear inclusion bodies. No abnormalitieswere detected in other organs except for moderateinterstitial pneumonia in the animals inoculatedwith EHV-9. The pulmonary changes consisted ofdiffusely thickened alveolar septa with varyingdegrees of macrophage and neutrophil infiltration.
Immunohistochemistry. EHV-9 antigen was detectedin the pseudounipolar neurons of the dorsal root(spinal) ganglia (Fig. 1a) and their synapses withinthe spinal cord (Fig. 1b) and in neurons in thegrey matter of the spinal cord (Fig. 1c). The pres-ence of EHV-9 antigen was confirmed at most levelsof the spinal cord in all inoculated animals. Strongpositive immunolabelling was detected in the me-dulla oblongata, pons and cerebellum. There wasmoderate intensity of labelling in the hippocampus,
Table
Histopathological changes in suckling
Tissue Lesion
12 (4) 24 (4)
Peripheral ganglia and
nerves
Degenerative change,
necrosis and
inflammation
+ +
Spinal ganglia Degenerative change,
necrosis and
inflammation
� +
Spinal cord Myelitis � � +Brain Encephalitis � �Myenteric plexuses Degenerative change,
necrosis and
inflammation
� �
Liver Degenerative change,
necrosis and
inflammation
� �
�, negative; +, positive; ND, not detected.
cerebral cortex and granular and mitral layers ofthe olfactory bulb and weak labelling of the glomer-ular layer of the olfactory bulb.
Suckling Hamsters
Clinical Signs. All of the inoculated suckling hamstershad varying degrees of clinical signs that consisted ofdepression and uncoordinated movements starting at96 hpi. At 168 hpi all animals exhibited markedlyuncoordinated movement.
Pathological Findings. At necropsy examination, nogross abnormalities were observed in any of the ani-mals examined. Sagittal sections of the suckling ham-sters enabled examination of the entire body ina single histological section. Microscopical findingsin suckling hamsters inoculated with EHV-9 are sum-marized in Table 1.
At 6 hpi there were no significant abnormalities ex-cept for mild neutrophil and lymphocyte infiltrationof the peritoneal cavity.
At 12 hpi there was mild degenerative change andvacuolation of the abdominal and peripheral ganglia(particularly the coeliac ganglia). There was alsomild perineuritis and neuritis with mild neutrophiland lymphocyte infiltration around the axons ofnerve fibres within the abdominal cavity (Fig. 2).
At 24 hpi, lymphocytic infiltration was observed inthe peripheral ganglia and nerves in the abdominalcavity, in addition to early vacuolation of the pseu-dounipolar neurons of the dorsal root ganglia.
At 36 hpi there were more prominent degenerativechanges and necrosis of the peripheral ganglia as wellas the pseudounipolar neurons of the spinal ganglia,
1
hamsters inoculated with EHV-9
hpi (number of animals)
36 (4) 48 (4) 72 (4) 96 (4) 120 (4) 144 (4) 168 (4)
+ + + + + + ND
+ + + + + + +
(meningitis) + + + + + +� � � + + + +
� + + + + + +
+ + + + + + +

Fig. 2. Inflammatory cells (predominantly lymphocytes)surround a peripheral nerve fibre adjacent to the kidney,resulting in perineuritis (12 hpi). HE. Bar, 100 mm.
Equine Herpesvirus-9 Infection of Hamsters 275
but there were no microscopical abnormalities in thespinal cord except for mild focal lymphocytic menin-gitis. There were degenerative changes in hepatocytesnear Glisson’s capsule in the liver.
At 48 hpi there was frequent necrosis of the neuronsin the spinal ganglia. In the grey matter of the spinalcord, themost characteristic findingwasmultifocalme-ningomyelitis, which presented as neuronal degenera-tion accompanied by gliosis and perivascularlymphocyte aggregation.Therewasmultifocal hepato-cellular degeneration and necrosis. Within the myen-teric plexuses of the intestine there was degenerativechange accompanied by moderate infiltration of lym-phocytes and plasma cells.
At 72 hpi there were more diffuse and prominentinflammatory aggregates in the spinal ganglia, as
Fig. 3. Severe meningomyelitis in a sagittal section of the spinalcord from a suckling hamster. There ismarked gliosis, peri-vascular lymphocytic aggregation and haemorrhage(72 hpi). HE. Bar, 200 mm.
well as intranuclear inclusions in the pseudounipolarneurons. Other findings included extension of menin-gomyelitis in the spinal cord (Fig. 3) and extension ofthe affected areas of the liver.
At 96 hpi there was more severe meningomyelitisextending to the anterior segments of the spinal cord,in addition tomarkednecrosis of thedorsal root ganglia(Fig. 4). There was meningoencephalitis affecting thebrain and this was particularly prominent in the poste-rior area of the brain at the level of the pons, medullaoblongata and cerebellum. These lesions were charac-terized by neuronal necrosis, mixed cell perivascularaggregation and gliosis. Intranuclear inclusions wereidentified in the hepatocytes (Fig. 5) and there was in-creased severity of degenerative change and inflamma-tory cell infiltration of the myenteric plexuses.
At 120 hpi, severe meningoencephalitis was pres-ent. This was characterized by neuronal necrosis,gliosis and perivascular lymphocyte aggregation. In-tranuclear inclusions were observed in the brain.These changes extended to the more anterior partsof the cerebral cortex and hippocampus.
At 144 and 168 hpi there was severe meningomye-litis and meningoencephalitis affecting most of theCNS, including the olfactory bulb, and severe necro-sis and inflammation of the hepatic parenchyma.
No abnormalities were observed in organs otherthan the CNS and liver except for moderate intersti-tial pneumonia in the lungs (at 36 hpi). The pulmo-nary changes consisted of diffusely thickenedalveolar septa with infiltration of these by varyingnumbers of macrophages and neutrophils. In addi-tion to the presence of bronchitis/bronchiolitis withmultifocal mucosal necrosis, findings included degen-eration and desquamation of the epithelial cells withabundant inflammatory cell infiltration of themucosaextending to the lumina of airways.
Immunohistochemistry. The distribution of EHV-9 an-tigens in the different parts of the body and theCNS is summarized in Table 2. A representative sag-ittal section is shown in Fig. 6.
At 6 hpi, EHV-9 antigen was observed in the peri-toneal macrophages (Fig. 7) and this labelling be-came more intense in cells collected at 12 hpi. Virusantigen was also detected in peripheral nerves withinthe abdominal cavity at 12 hpi (Fig. 8). At 24 hpithere was immunoreactivity in the pseudounipolarnuclei of the peripheral ganglia of the abdominal cav-ity (particularly the coeliac ganglia; Fig. 9) and in theaxons of the peripheral nerves. At 36 hpi, virus anti-gen was detected in the dorsal root ganglia and therewas light labelling of the meninges of the spinal cord.At 48e72 hpi, EHV-9 antigen was found in most ofthe neurons (either in the cytoplasm or in the nuclei)
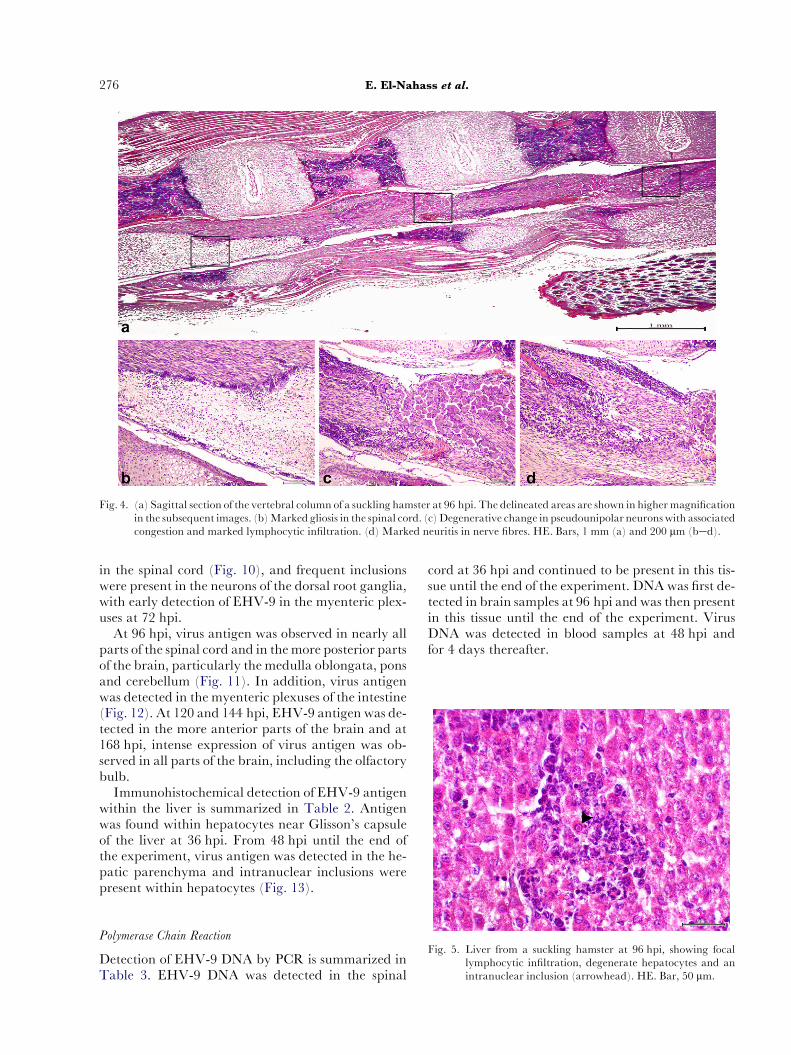
Fig. 4. (a) Sagittal section of the vertebral column of a suckling hamster at 96 hpi. The delineated areas are shown in higher magnificationin the subsequent images. (b)Marked gliosis in the spinal cord. (c)Degenerative change in pseudounipolar neuronswith associatedcongestion and marked lymphocytic infiltration. (d) Marked neuritis in nerve fibres. HE. Bars, 1 mm (a) and 200 mm (bed).
276 E. El-Nahass et al.
in the spinal cord (Fig. 10), and frequent inclusionswere present in the neurons of the dorsal root ganglia,with early detection of EHV-9 in the myenteric plex-uses at 72 hpi.
At 96 hpi, virus antigen was observed in nearly allparts of the spinal cord and in themore posterior partsof the brain, particularly the medulla oblongata, ponsand cerebellum (Fig. 11). In addition, virus antigenwas detected in the myenteric plexuses of the intestine(Fig. 12). At 120 and 144 hpi, EHV-9 antigen was de-tected in the more anterior parts of the brain and at168 hpi, intense expression of virus antigen was ob-served in all parts of the brain, including the olfactorybulb.
Immunohistochemical detection of EHV-9 antigenwithin the liver is summarized in Table 2. Antigenwas found within hepatocytes near Glisson’s capsuleof the liver at 36 hpi. From 48 hpi until the end ofthe experiment, virus antigen was detected in the he-patic parenchyma and intranuclear inclusions werepresent within hepatocytes (Fig. 13).
Fig. 5. Liver from a suckling hamster at 96 hpi, showing focallymphocytic infiltration, degenerate hepatocytes and anintranuclear inclusion (arrowhead). HE. Bar, 50 mm.
Polymerase Chain Reaction
Detection of EHV-9 DNA by PCR is summarized inTable 3. EHV-9 DNA was detected in the spinal
cord at 36 hpi and continued to be present in this tis-sue until the end of the experiment. DNAwas first de-tected in brain samples at 96 hpi and was then presentin this tissue until the end of the experiment. VirusDNA was detected in blood samples at 48 hpi andfor 4 days thereafter.

Table 2
Virus detection by ICC and IHC in suckling hamsters inoculated with EHV-9
Tissue Virus antigen labelling pattern hpi (number of animals)
6(4) 12(4) 24(4) 36(4) 48(4) 72(4) 96(4) 120(4) 144 (4) 168(4)
Peritoneal macrophages Attachment, migrationand propagation
+ + + ND ND ND ND ND ND ND
Peripheral ganglia
and nerves
Migration � + + + + + + + + ND
Spinal ganglia Migration and propagation � � � + + + + + + +Spinal cord Migration � � � + (meninges) + + + + + +
Brain Migration � � � � � � + + + +
Myenteric plexuses Migration and propagation � � � � � + + + + +Liver Attachment and propagation � � � + + + + + + +
�, negative; +, positive; ND, not detected.
Equine Herpesvirus-9 Infection of Hamsters 277
Discussion
Numerous studies have suggested that infection withEHV-9 may be an emerging infectious disease be-cause the virus produces a fulminant encephalitisthat is easily transmitted via nasal inoculation in var-ious animal species including mice and hamsters(Fukushi et al., 2000), goats (Taniguchi et al., 2000),dogs and cats (Yanai et al., 2003a, b) and commonmarmosets (Kodama et al., 2007). In addition, natu-ral infections with EHV-9 have been reported re-cently in a polar bear (Donovan et al., 2009) anda giraffe (Schrenzel et al., 2008).
Previous studies have shown that EHV-9 encepha-litis may be induced experimentally in hamsters fol-lowing inoculation of virus by the ocular, oral orintraperitoneal routes. However, these investigationshave not defined the primary site of virus attachmentand propagation or the detailed pathogenesis of infec-tion (El-Habashi et al., 2010b).
In the present study, we first inoculated young adultSyrianhamsters intraperitoneallywithEHV-9 to study
Fig. 6. Sagittal section of the entire body of a suckling hamsterfrom the neck to the tail showing all internal organs.Note marked antigen expression in the liver. IHC. Bar,10 mm.
the infectivity and pathogenesis, but it was difficult toexamine the spinal cord and the associated peripheralnerves and ganglia because of the calcified vertebraeand skull. Therefore, we used a suckling hamstermodel, which had been the subject of a previous studyelucidating thepathogenesis ofEHV-9 infection follow-ing nasal inoculation (El-Habashi et al., 2010a).
To confirm virus propagation at the site of inocula-tion, peritoneal macrophages were collected at6e24 hpi. These cells were shown to express EHV-9antigen by ICC and this was interpreted to meanthat these cells may be the primary site of virus attach-ment and propagation after intraperitoneal inocula-tion. Moreover, the virus may then spread from theperitoneal macrophages to the peripheral nervesand ganglia in the abdominal and pelvic cavities.The role of peritoneal macrophages in the transmis-sion of viruses has been described in studies of herpessimplex virus type 1 (Hirsch et al., 1970; Letterio et al.,1983). After this virus replicated in the peritonealmacrophages it extended to the gut plexuses
Fig. 7. Expression of EHV-9 antigen in peritoneal macrophages(arrow) at 6 hpi. ICC. Bar, 100 mm.
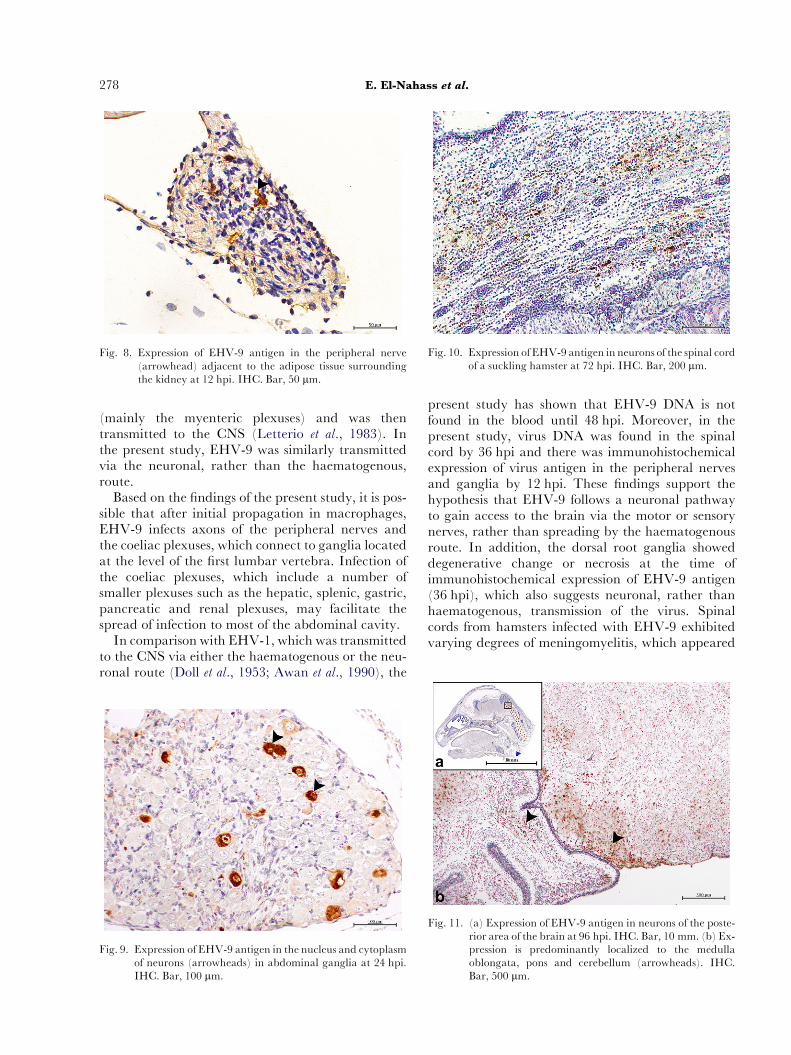
Fig. 8. Expression of EHV-9 antigen in the peripheral nerve(arrowhead) adjacent to the adipose tissue surroundingthe kidney at 12 hpi. IHC. Bar, 50 mm.
Fig. 10. Expression of EHV-9 antigen in neurons of the spinal cordof a suckling hamster at 72 hpi. IHC. Bar, 200 mm.
278 E. El-Nahass et al.
(mainly the myenteric plexuses) and was thentransmitted to the CNS (Letterio et al., 1983). Inthe present study, EHV-9 was similarly transmittedvia the neuronal, rather than the haematogenous,route.
Based on the findings of the present study, it is pos-sible that after initial propagation in macrophages,EHV-9 infects axons of the peripheral nerves andthe coeliac plexuses, which connect to ganglia locatedat the level of the first lumbar vertebra. Infection ofthe coeliac plexuses, which include a number ofsmaller plexuses such as the hepatic, splenic, gastric,pancreatic and renal plexuses, may facilitate thespread of infection to most of the abdominal cavity.
In comparison with EHV-1, which was transmittedto the CNS via either the haematogenous or the neu-ronal route (Doll et al., 1953; Awan et al., 1990), the
Fig. 9. Expression of EHV-9 antigen in the nucleus and cytoplasmof neurons (arrowheads) in abdominal ganglia at 24 hpi.IHC. Bar, 100 mm.
present study has shown that EHV-9 DNA is notfound in the blood until 48 hpi. Moreover, in thepresent study, virus DNA was found in the spinalcord by 36 hpi and there was immunohistochemicalexpression of virus antigen in the peripheral nervesand ganglia by 12 hpi. These findings support thehypothesis that EHV-9 follows a neuronal pathwayto gain access to the brain via the motor or sensorynerves, rather than spreading by the haematogenousroute. In addition, the dorsal root ganglia showeddegenerative change or necrosis at the time ofimmunohistochemical expression of EHV-9 antigen(36 hpi), which also suggests neuronal, rather thanhaematogenous, transmission of the virus. Spinalcords from hamsters infected with EHV-9 exhibitedvarying degrees of meningomyelitis, which appeared
Fig. 11. (a) Expression of EHV-9 antigen in neurons of the poste-rior area of the brain at 96 hpi. IHC. Bar, 10 mm. (b) Ex-pression is predominantly localized to the medullaoblongata, pons and cerebellum (arrowheads). IHC.Bar, 500 mm.
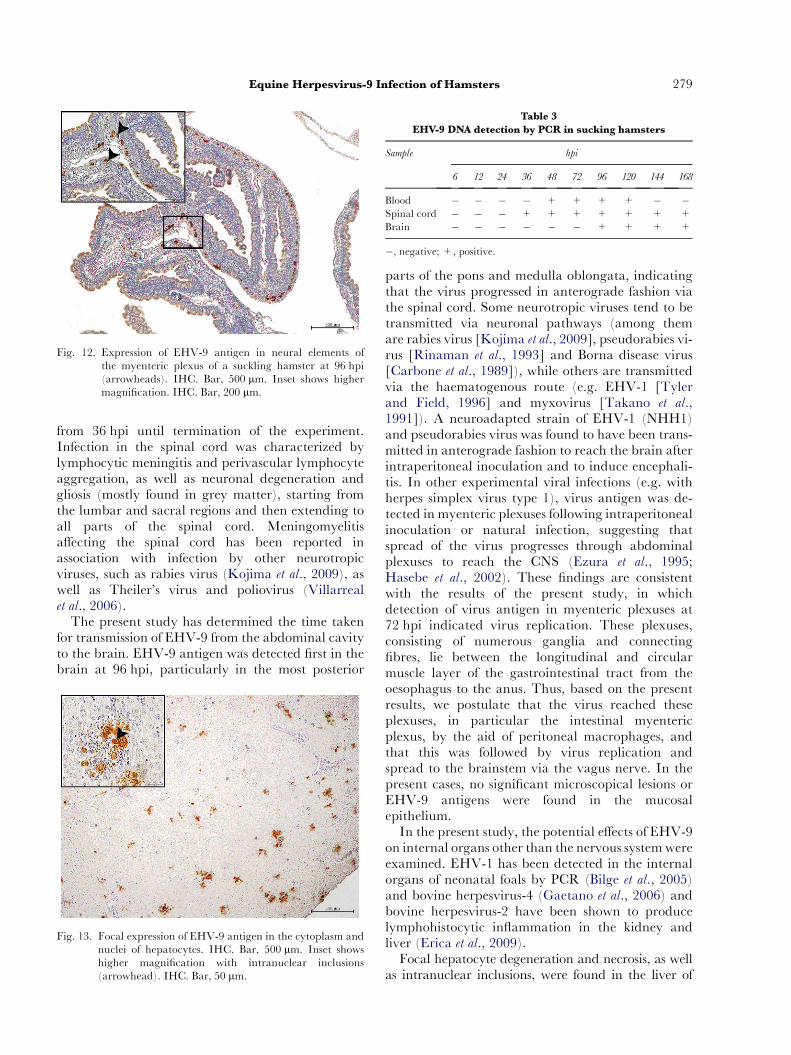
Fig. 12. Expression of EHV-9 antigen in neural elements ofthe myenteric plexus of a suckling hamster at 96 hpi(arrowheads). IHC. Bar, 500 mm. Inset shows highermagnification. IHC. Bar, 200 mm.
Table 3
EHV-9 DNA detection by PCR in sucking hamsters
Sample hpi
6 12 24 36 48 72 96 120 144 168
Blood � � � � + + + + � �Spinal cord � � � + + + + + + +
Brain � � � � � � + + + +
�, negative; +, positive.
Equine Herpesvirus-9 Infection of Hamsters 279
from 36 hpi until termination of the experiment.Infection in the spinal cord was characterized bylymphocytic meningitis and perivascular lymphocyteaggregation, as well as neuronal degeneration andgliosis (mostly found in grey matter), starting fromthe lumbar and sacral regions and then extending toall parts of the spinal cord. Meningomyelitisaffecting the spinal cord has been reported inassociation with infection by other neurotropicviruses, such as rabies virus (Kojima et al., 2009), aswell as Theiler’s virus and poliovirus (Villarrealet al., 2006).
The present study has determined the time takenfor transmission of EHV-9 from the abdominal cavityto the brain. EHV-9 antigen was detected first in thebrain at 96 hpi, particularly in the most posterior
Fig. 13. Focal expression of EHV-9 antigen in the cytoplasm andnuclei of hepatocytes. IHC. Bar, 500 mm. Inset showshigher magnification with intranuclear inclusions(arrowhead). IHC. Bar, 50 mm.
parts of the pons and medulla oblongata, indicatingthat the virus progressed in anterograde fashion viathe spinal cord. Some neurotropic viruses tend to betransmitted via neuronal pathways (among themare rabies virus [Kojima et al., 2009], pseudorabies vi-rus [Rinaman et al., 1993] and Borna disease virus[Carbone et al., 1989]), while others are transmittedvia the haematogenous route (e.g. EHV-1 [Tylerand Field, 1996] and myxovirus [Takano et al.,1991]). A neuroadapted strain of EHV-1 (NHH1)and pseudorabies virus was found to have been trans-mitted in anterograde fashion to reach the brain afterintraperitoneal inoculation and to induce encephali-tis. In other experimental viral infections (e.g. withherpes simplex virus type 1), virus antigen was de-tected in myenteric plexuses following intraperitonealinoculation or natural infection, suggesting thatspread of the virus progresses through abdominalplexuses to reach the CNS (Ezura et al., 1995;Hasebe et al., 2002). These findings are consistentwith the results of the present study, in whichdetection of virus antigen in myenteric plexuses at72 hpi indicated virus replication. These plexuses,consisting of numerous ganglia and connectingfibres, lie between the longitudinal and circularmuscle layer of the gastrointestinal tract from theoesophagus to the anus. Thus, based on the presentresults, we postulate that the virus reached theseplexuses, in particular the intestinal myentericplexus, by the aid of peritoneal macrophages, andthat this was followed by virus replication andspread to the brainstem via the vagus nerve. In thepresent cases, no significant microscopical lesions orEHV-9 antigens were found in the mucosalepithelium.
In the present study, the potential effects of EHV-9on internal organs other than the nervous systemwereexamined. EHV-1 has been detected in the internalorgans of neonatal foals by PCR (Bilge et al., 2005)and bovine herpesvirus-4 (Gaetano et al., 2006) andbovine herpesvirus-2 have been shown to producelymphohistocytic inflammation in the kidney andliver (Erica et al., 2009).
Focal hepatocyte degeneration and necrosis, as wellas intranuclear inclusions, were found in the liver of

280 E. El-Nahass et al.
the hamsters in the present study. Immunohistochemi-cal labelling in the same areas suggested that the liveris a possible target organ for EHV-9 when inoculatedvia the intraperitoneal route. Previous studies of suck-ling mice have shown that the liver plays an importantrole in virus replication, especially during the initialstages of infection (Plummer et al., 1973).
In conclusion, the present study has characterizedthe pathogenesis of EHV-9 infection following intra-peritoneal inoculation of the virus into young adultand suckling hamsters. After inoculation, EHV-9may have propagated in the peritoneal macrophagesbefore being transmitted to the peritoneal peripheralnerves. The virus may then gain access to the abdom-inal ganglia and the dorsal root (spinal) ganglia,where it invades the spinal cord, resulting inmeningo-myelitis. At the same time, the virus appears to betransmitted to the myenteric plexuses in the intestine.In the final stage of infection, EHV-9 reaches thebrain (where it induces encephalitis) by means of as-cending transmission via the spinal cord. The presentstudy is also the first to describe localization of EHV-9in the liver following inoculation of hamsters by theintraperitoneal route.
Acknowledgments
This study was supported in part by a grant-in-aid(Emerging-General) for scientific research from theMinistry of Health, Labor and Welfare of Japanand a grant from the Ono Pharmaceutical Co., Ltd.We also thank Miss C. Swift for her conscientiousproofreading of the manuscript.
Conflict of Interest
None of the authors of this paper has a financial orpersonal relationship with other persons or organiza-tions that could inappropriately influence or bias thecontent of the paper.
References
AwanAR, Chong YC, Field HJ (1990) The pathogenesis ofequine herpesvirus type 1 in the mouse: a new model forstudying host responses to infection. Journal of GeneralVirology, 71, 1131e1140.
Bilge K, Tomio M, Nuri T, Hayri E, Huseyin Y (2005)Detection of equine herpesvirus type 1 (EHV-1) DNAin organs of neonatal dead foals in Turkey. Journal ofEquine Science, 17, 23e26.
Borchers K, Wiik H, Frolich K, Ludwig H, East M (2005)Antibodies against equine herpesviruses and equinearteritis virus in Burchell’s zebra (Equus burchelli) fromthe Serengeti ecosystem. Journal of Wildlife Diseases, 41,80e86.
Carbone KM, Trapp BD, Griffin JW, Duchala CS,Narayan O (1989) Astrocytes and Schwann cells arevirus host cells in nervous system of rats with Borna dis-ease. Journal of Neuropathology & Experimental Neurology,48, 631e644.
Doll ER, Richards MG,WallaceME (1953) Adaptation ofthe equine abortion virus to suckling Syrian hamsters.Cornell Veterinarian, 42, 551e558.
Donovan TA, Schrenzel MD, Tucker T, Pessier AP,Bicknese B et al. (2009) Meningoencephalitis in a polarbear caused by equine herpesvirus-9 (EHV-9).VeterinaryPathology, 46, 1138e1143.
El-Habashi N, El-Nahass E, Fukushi H, Hibi D, Skai Het al. (2010a) Experimental intranasal infection ofequine herpesvirus-9 (EHV-9) in suckling hamsters:kinetics of viral transmission and inflammation in thenasal cavity and brain. Journal of Neurovirology, 16, 1e7.
El-Habashi N, Murakami M, El-Nahass E, Hibi D,Sakai H et al. (2010b) Study on infectivity of equine her-pesvirus-9 (EHV-9) by different routes of inoculation inhamsters. Veterinary Pathology, 47, 1e7.
Erica A, Maria Q, Bomfim G, Da Fonseca P, Drumond Cet al. (2009) Bovine herpesvirus-2 infection in foals inBrazil. Emerging Infectious Diseases, 15, 844e845.
Ezura K, Usami Y, Tajima K, Komaniwa H, Nagai S et al.(1995) Gastrointestinal and skin lesions in piglets natu-rally infected with pseudorabies virus. Journal of Veteri-nary Diagnostic Investigation, 7, 451e455.
Fukushi H, Taniguchi A, Yasuda K, Yanai T, Masegi Tet al. (2000) A hamster model of equine herpesvirus-9-induced encephalitis.Journal ofNeurovirology, 6, 314e319.
Fukushi H, Tomita T, Taniguchi A, Ochiai Y, Kirisawa Ret al. (1997) Gazelle herpesvirus-1: a new neurotropicherpesvirus immunologically related to equine herpesvi-rus-1. Virology, 227, 34e44.
Gaetano D, EugenioM, Enzo P, Claudia L, FilippoM et al.(2006) Bovine herpes-4-based vector interaction withliver cells in vitro and in vivo. Journal of Virological Methods,136, 126e136.
Goafa F, Cheryl C, Ari S, Reinhard K, Susan E et al. (1996)Endogenous substance P mediates cold water stress-in-duced increase in interleukin-6 secretion from peritonealmacrophages. Journal of Neuroscience, 16, 3745e3752.
Hasebe R, Kimura T, Sato E, Okazaki K, Ochiai K et al.(2002) Equine herpesvirus-1 induced encephalomyelitisin mice: a comparative study of neuroadapted virus andits parental strain. Journal of Comparative Pathology,127, 118e125.
Hirsch MB, Zisman B, Allison AC (1970) Macrophagesand age dependent resistance to herpes simplex virusin mice. Journal of Immunology, 104, 1160e1165.
Kasem S, Yamada S, KiupelM,Woodruff M, OhyaK et al.(2008) Equine herpesvirus type 9 in a giraffewith enceph-alitis. Emerging Infectious Diseases Journal, 14, 1948e1949.
Kodama A, Yanai T, Yomemaru K, Sakai H, Masegi Tet al. (2007) Acute neuropathogenicity with experimen-tal infection of equine herpesvirus-9 in common marmo-sets (Callithrix jacchus). Journal of Medical Primatology,36, 335e342.

Equine Herpesvirus-9 Infection of Hamsters 281
Kojima D, Park CH, Satoh Y, Inoue S, Noguchi A et al.(2009) Pathology of spinal cord of C57BL/6J miceinfected with rabies virus (CVS-11 strain). Journal ofVeterinary Medical Science, 71, 319e324.
Letterio B, Iannello D,Merendino R, Aren A, Mastroeni P(1983) Tumor-dependent resistance of rat peritonealmacrophages to herpes simplex virus. Infection and
Immunity, 39, 575e579.Narita M, Uchimura A, Kimura K, Tanimura N, Yanai T
et al. (2000) Brain lesions and transmission of experimen-tal equine herpesvirus type 9 in pigs. Veterinary Pathology,37, 476e479.
Plummer G, Coleman P, Cleveland P, Henson D (1973)Neurovirulence of equine herpes type 1 in mice of differ-ent ages. Journal of Infectious Diseases, 128, 202e210.
Rinaman L, Card JP, Enquist LW (1993) Spatiotemporalresponses of astrocytes, ramified microglia and brainmacrophages to central neuronal infection with pseu-dorabies virus. Journal of Neuroscience, 13, 685e702.
Schrenzel MD, Tucker TA, Donovan TA, Martin DM,Busch MD et al. (2008) New hosts for equine herpesvi-rus-9.Emerging Infectious Diseases Journal, 14, 1616e1619.
Takano T, Ohno M, Yamano T, Shimada M (1991)Experimental hydrocephalus in suckling hamsters inducedby myxovirus infection: pathogenesis of hydrocephaluscaused bymumps virus. Congenital Anomalies, 31, 115e128.
Taniguchi A, Fukushi H, Yanai T, Masegi T, Hirai K(2000) Equine herpesvirus-9-induced lethal encephalitis
in experimentally infected goats. Archives of Virology,145, 2619e2627.
Tyler KL, Field BN (1996) Pathogenesis of viral infec-tion. In: Virology, 3rd Edit., Vol. I, BN Fields,DM Knipe, PM Howley, Eds., Raven Press, NewYork, pp. 173e218.
Villarreal D, Young CR, Storts R, Tinng JW, Welsh CJ(2006) A comparison of the neurotropism of Thelier’svirus and poliovirus in CBA mice.Microbial Pathogenesis,41, 149e156.
Yanai T, Fujishima N, Fukushi H, Hirata A, Sakai H et al.(2003a) Experimental infection of equine herpesvirus-9in dogs. Veterinary Pathology, 40, 263e267.
Yanai T, Sakai T, Fukushi H, Hirai K, Narita M et al.(1998) Neuropathological study of gazelle herpesvirus-9 (equine herpesvirus-9) infection in Thomson’s gazelles(Gazella thomsoni). Journal of Comparative Pathology, 119,159e168.
Yanai T, Tsujioka S, Sakai H, Fukushi H, Hirai K et al.(2003b) Experimental infection with equine herpesvi-rus-9 (EHV-9) in cats. Journal of Comparative Pathology,128, 113e118.
½ R
A
eceived, October 18th, 2010
ccepted, January 13th, 2011
�



















