
Photodynamic vaccination of hamsters with inducible suicidal mutants of Leishmania amazonensis...
14
Photodynamic vaccination of hamsters with inducible suicidal mutants of Leishmania amazonensis elicits immunity against visceral leishmaniasis Shraddha Kumari 1 , Mukesh Samant 1 , Prashant Khare 1 , Pragya Misra 1 , Sujoy Dutta 2 , Bala Krishna Kolli 2 , Sharad Sharma 3 , Kwang Poo Chang 2 and Anuradha Dube 1 1 Division of Parasitology, Central Drug Research Institute, Lucknow, India 2 Department of Microbiology/Immunology, Chicago Medical School, Rosalind Franklin University, North Chicago, IL, USA 3 Division of Toxicology, Central Drug Research Institute, Lucknow, India Leishmania, naturally residing in the phagolysosomes of macrophages, is a suitable carrier for vaccine delivery. Genetic complementation of these trypanosomatid protozoa to partially rectify their defective heme-biosynthesis renders them inducible with d-amino- levulinate to develop porphyria for selective photolysis, leaving infected host cells unscathed. Delivery of released ‘‘vaccines’’ to antigen-presenting cells is thus expected to enhance immune response, while their self-destruction presents added advantages of safety. Such suicidal L. amazonensis was found to confer immunoprophylaxis and immunotherapy on hamsters against L. donovani. Neither heat-killed nor live parasites without suicidal induction were effective. Photodynamic vaccination of hamsters with the suicidal mutants reduced the parasite loads by 99% and suppressed the development of disease. These suppressions were accompanied by an increase in Leishmania-specific delayed-type hypersensitivity and lymphoproliferation as well as in the levels of splenic iNOS, IFN-c, and IL-12 expressions and of Leishmania-specific IgG2 in the serum. More- over, a single intravenous administration of T cells from vaccinated hamsters was shown to confer on naı ¨ve animals an effective cellular immunity against L. donovani challenges. The absence of lesion development at vaccination sites and parasites in the draining lymphnodes, spleen and liver further indicates that the suicidal mutants provide a safe platform for vaccine delivery against experimental visceral leishmaniasis. Key words: Porphyrinogenic mutant . Suicidal vaccination . T-cell-adoptive transfer . Visceral leishmaniasis Introduction World Health Organization (WHO) considers leishmaniasis to be one of the most serious, epidemic-prone parasitic diseases of the poor and disadvantaged. An estimated 350 million people are at risk of Leishmania infection worldwide with 1.5 million new cases of cutaneous leishmaniasis (CL) and 500 000 cases of visceral leishmaniasis (VL) reported each year. Recent epidemics of VL or kala-azar in Sudan and India have resulted in over 100 000 deaths [1]. With the advent of the HIV epidemics, VL has emerged as an important opportunistic infection in AIDS patients [2, 3]. In India, high incidence of kala-azar has been reported Correspondence: Dr. Anuradha Dube e-mail: [email protected] & 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu DOI 10.1002/eji.200838389 Eur. J. Immunol. 2009. 39: 178–191 Shraddha Kumari et al. 178
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Photodynamic vaccination of hamsters with inducible suicidal mutants of Leishmania amazonensis...

Photodynamic vaccination of hamsters with induciblesuicidal mutants of Leishmania amazonensis elicitsimmunity against visceral leishmaniasis
Shraddha Kumari1, Mukesh Samant1, Prashant Khare1, Pragya Misra1,
Sujoy Dutta2, Bala Krishna Kolli2, Sharad Sharma3, Kwang Poo Chang2
and Anuradha Dube1
1 Division of Parasitology, Central Drug Research Institute, Lucknow, India2 Department of Microbiology/Immunology, Chicago Medical School, Rosalind Franklin
University, North Chicago, IL, USA3 Division of Toxicology, Central Drug Research Institute, Lucknow, India
Leishmania, naturally residing in the phagolysosomes of macrophages, is a suitable carrier
for vaccine delivery. Genetic complementation of these trypanosomatid protozoa to
partially rectify their defective heme-biosynthesis renders them inducible with d-amino-
levulinate to develop porphyria for selective photolysis, leaving infected host cells
unscathed. Delivery of released ‘‘vaccines’’ to antigen-presenting cells is thus expected to
enhance immune response, while their self-destruction presents added advantages of
safety. Such suicidal L. amazonensis was found to confer immunoprophylaxis and
immunotherapy on hamsters against L. donovani. Neither heat-killed nor live parasites
without suicidal induction were effective. Photodynamic vaccination of hamsters with the
suicidal mutants reduced the parasite loads by 99% and suppressed the development of
disease. These suppressions were accompanied by an increase in Leishmania-specific
delayed-type hypersensitivity and lymphoproliferation as well as in the levels of splenic
iNOS, IFN-c, and IL-12 expressions and of Leishmania-specific IgG2 in the serum. More-
over, a single intravenous administration of T cells from vaccinated hamsters was shown
to confer on naı̈ve animals an effective cellular immunity against L. donovani challenges.
The absence of lesion development at vaccination sites and parasites in the draining
lymphnodes, spleen and liver further indicates that the suicidal mutants provide a safe
platform for vaccine delivery against experimental visceral leishmaniasis.
Key words: Porphyrinogenic mutant . Suicidal vaccination . T-cell-adoptive transfer .
Visceral leishmaniasis
Introduction
World Health Organization (WHO) considers leishmaniasis to be
one of the most serious, epidemic-prone parasitic diseases of the
poor and disadvantaged. An estimated 350 million people are at
risk of Leishmania infection worldwide with �1.5 million new
cases of cutaneous leishmaniasis (CL) and 500 000 cases of
visceral leishmaniasis (VL) reported each year. Recent epidemics
of VL or kala-azar in Sudan and India have resulted in over
100 000 deaths [1]. With the advent of the HIV epidemics, VL has
emerged as an important opportunistic infection in AIDS patients
[2, 3]. In India, high incidence of kala-azar has been reportedCorrespondence: Dr. Anuradha Dubee-mail: [email protected]
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
DOI 10.1002/eji.200838389 Eur. J. Immunol. 2009. 39: 178–191Shraddha Kumari et al.178

from the states of Bihar, Assam, West Bengal and Eastern Uttar
Pradesh. A recent survey in Bihar has recorded an alarming
100 000 cases, of which 10 000 are unresponsive to antimonials.
The numerous relapses due to the increasing incidence of drug
resistance are also associated with the toxic anti-leishmanial
drugs in use [4]. Hence, the development of an effective vaccine
becomes all the more urgent in the absence of suitable anti-
leishmanial drugs to control this disease.
Effective vaccines for VL require their activity to elicit cell-
mediated immunity (CMI) capable of activating macrophages to
a microbicidal state [5]. Further, it is desirable to have a suitable
animal model for human kala-azar adequate for realistic
evaluation of vaccine safety and efficacy. Golden hamster
(Mesocricetus auratus) is the best animal model of human VL in
having the same clinico-pathological features, including a
relentless increase in visceral parasitic burdens, progressive
cachexia, hepatosplenomegaly, pancytopenia, hyper-gamma-
globulinemia and ultimately death [6, 7].
Several vaccination strategies using live, killed and defined
vaccines have been attempted with particular emphasis on their
efficacy against CL instead of VL [3]. The only successful immuni-
zation strategy in humans has been the ‘‘leishmanization’’, which is
based on the development of durable immunity after recovery from
infection at a chosen site, usually the arm, with viable non-attenuated
parasites [8, 9]. The use of live vaccines is very promising since they
most closely mimic the natural course of infection and, therefore,
may elicit similarly effective immunity after cure. However, its use
has been restricted or abandoned entirely due to the safety concerns.
Nevertheless, WHO/Tropical Disease Research (WHO/TDR) has very
recently revived the consideration of using live, but genetically atte-
nuated vaccines [10]. Other potentially usable live vaccines include
Leishmania administered at a low dose [11], drug-attenuated Leish-
mania [12], recombinant Leishmania expressing cytokines [13] or
suicide markers [14] and parasites with CpG-oligodeoxynucleotide
motifs [15]. However, these experimental vaccines have yet to reach
the clinical trial stage. While each of them produced some level of
protective efficacy, the completeness and longevity of this protection
has not been clearly demonstrated. Hence, no effective vaccine
currently exists against any form of human leishmaniasis.
Recently, a xenotransgenic-L. amazonensis was constructed by a
novel suicidal design, which was predicted to effect its specific and
selective destruction in infected macrophages [16]. This inducible
suicidal mutant was produced transgenically to partially rectify its
natural defects in heme-biosynthesis. The second and third enzymes
(ALAD: d-aminolevulinate dehydratase, PBGD: porphobilinogen
deaminase) of the heme-biosynthetic pathway were episomally
expressed in Leishmania. This transfectant was thus responsive to
d-aminolevulinate (ALA), the product of the first enzyme in this
pathway, for accumulation of photosensitive uroporphyrin. This
porphyrin can be excited by light to produce leishmanolytic oxidative
species. With a complete heme-metabolic pathway, the host cells
only develop a very low level of transient porphyria in response to
ALA. This parasite versus host-cell difference in accumulation of
porphyrins was exploited to achieve selective photolysis of Leishma-
nia in the phagolysosomes of the antigen-presenting cells. The basic
principle for using the suicidal mutants as vaccine carriers is that they
retain infection molecules for homing to phagolysosomes, thereby
delivering vaccines to the desirable destination. Induction of their
photolysis therein achieves not only vaccine release for appropriate
processing and presentation but also their self-destruction.
An ideal live vaccine should elicit long-term effective immune
responses without undue persistence in the vaccinated host, as
safety is an important criterion for the use of live vaccines in
humans. The potential utility of the suicidal mutants as a vaccine
candidate was explored here against Indian kala-azar in hamster
model. Results obtained demonstrated the efficacy of such
photodynamic vaccination. In addition, a long-lasting protective
immunity was found to develop by a single intravenous injection
of lymphnode T cells from porphyrinogenic vaccinated hamsters.
This adoptive transfer is capable of protecting naı̈ve hamsters to
L. donovani challenges.
ResultsAbsence of cutaneous lesion and amastigotes inporphyrinogenic vaccinated hamsters
The site of vaccination was inspected in all groups for evidence of
any cutaneous lesions – a characteristic of CL. The lesions of
0.3–0.65 cm in diameter appeared at the site of injection after
3 wk of vaccination, but only in animals of the control non-
porphyrinogenic groups, viz. DT�ALA and ST1ALA (see Materi-
als and methods). In contrast, porphyrinogenic (DT1ALA)
produced no visible lesion throughout the experiment period till
day 180 post-challenges (p.c.), indicative of an excellent safety
margin. This was further confirmed by histopathological studies
of the skin in separate groups of hamsters 3 wk after vaccination
(Fig. 1). In skin sections of porphyrinogenic vaccinated animals,
only uninfected macrophages were observed without inflamma-
tory cell infiltrates (Fig. 1A and B). In contrast, the control
groups, i.e. DT�ALA and ST1ALA, produced lesions, which
clearly resulted from inflammatory responses due to cell
infiltrates, consisting mainly of lymphocytes, plasma cells, and
Leishmania-infected macrophages (Fig. 1C–F). No infected cells
were seen in histological sections of spleens, livers, and
lymphnodes from DT1ALA hamsters. Cultures of tissues from
the cutaneous sites of vaccination, their draining lymphnodes,
spleens, and livers yielded no promastigotes after incubation for 1
month at 261C, providing further evidence for the absence of
detectable parasites in these animals.
Vaccination with porphyrinogenic-L. amazonensisinduced optimum protection against L. donovanichallenges
The porphyrinogenic-mutant (DT1ALA) vaccinated hamsters
were protected from the challenge infection of L. donovani, as
indicated by their weight gain with time, like non-vaccinated and
Eur. J. Immunol. 2009. 39: 178–191 Immunity to infection 179
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

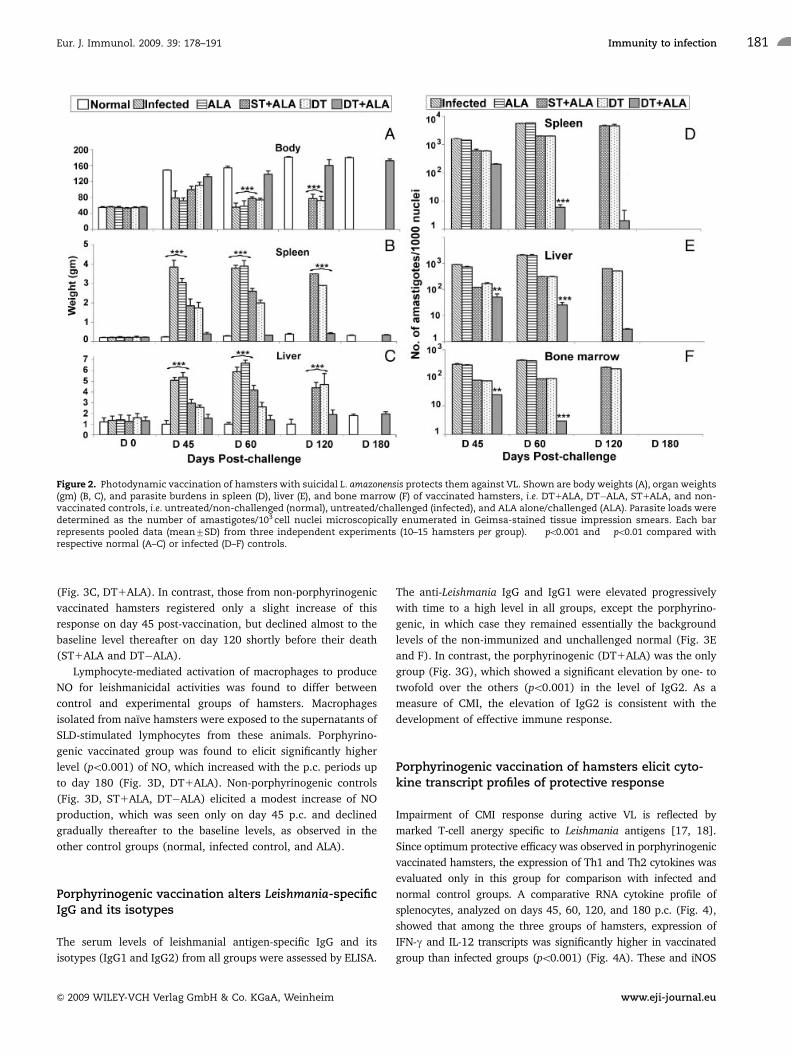
non-challenged group (Fig. 2A, solid bar versus blank bar) in
contrast to the significant weight loss (po0.001) of all the control
groups, i.e. infected, ALA, ST1ALA, DT�ALA (bars in between
the solid and the blank bars). Hepatosplenomegaly, normally
associated with the challenge infection, was absent in the
porphyrinogenic vaccinated group (Fig. 2B and C, solid bar).
This phenotype was evident, albeit much less pronounced in the
beginning, in the two groups vaccinated with non-porphyrino-
genic controls, indicative of their ineffective protection (Fig. 2B
and C, stippled and hatched bars). Parasite loads were positively
correlated with splenomegaly and hepatomegaly observed among
different groups: an increase from �103 to 4104 in all the
groups, except the porphyrinogenic vaccinated groups (Fig. 2D,
solid bar versus the rest), as seen in Geimsa-stained splenic
smears, from day 45 to day 120 p.c. In this last group, parasite
loads decreased from �2� 102 on day 45 to a negligible level
(po0.001) of o10 subsequently, rendering them difficult to
discern by microscopy (not shown). Similarly, in liver and bone
marrow, parasite loads sharply decreased after day 45 p.c. and
parasites were essentially absent by day 180 p.c. in the same fully
vaccinated group in contrast to all the control groups (Fig. 2E and
F, solid bars versus the rest). Cultivation of the spleen, liver, and
lymphnode tissues from the fully vaccinated hamsters in vitro
yielded no promastigotes after prolonged incubation for 3 wk.
Porphyrinogenic-L. amazonensis (DT1ALA) vaccinated
hamsters survived the challenges of L. donovani and remained
healthy thereafter until the termination of the experiment 6
months p.c. In contrast, hamsters vaccinated with non-porphyr-
inogenic mutants, i.e. DT�ALA or ST1ALA survived for only 3–4
months, while all those untreated or treated with ALA alone died
of kala-azar within �2 months.
Immunization of hamsters with killed DT mutants under the
same vaccination conditions gave only limited prophylactic effi-
cacy. This was demonstrated in a separate set of experiments, in
which three groups of hamsters at five males each were vacci-
nated for comparison with autoclaved DT, DT1ALA, and
DT�ALA, respectively. Evaluation of splenic amastigote loads 45
days p.c. showed parasite suppression of 95, 43, and 31% for DT
1ALA, DT�ALA, and killed DT, respectively. Photodynamic
vaccination with live DT1ALA was shown again to give the most
complete protection that is unmatched by either live DT alone or
killed DT. Since the killed mutants were least effective, they were
not included for further investigation.
Porphyrinogenic vaccination stimulates delayed-typehypersensitivity, mitogenic, and Leishmania-specificcellular responses
Porphyrinogenic and non-porphyrinogenic immunizations
elicited different CMI, as determined by assessing the delayed-
type hypersensitivity (DTH) and lymphoblast proliferation
responses to specific and/or non-specific antigens in these
hamsters after L. donovani challenges at different time periods.
Porphyrinogenic vaccinated hamsters displayed significant DTH
response, which increased progressively throughout the entire
p.c. period (Fig. 3A, DT1ALA) and reached the levels that were
significantly higher than those of the control groups (po0.001) at
all time points for the duration of the experiments for up to 180
days. The DTH response increased in the non-porphyric control
groups (DT�ALA, ST1ALA), but this increase was rather modest.
In vitro stimulation of the lymphocytes with Concanavalin A
(Con A) showed comparable proliferative response at high levels
in all the groups when assayed before challenges (Fig. 3B, day 0).
Con A induced lymphoproliferative response remained elevated
in porphyrinogenic vaccinated animals as much as those of the
normal hamsters throughout the entire p.c. period (Fig. 3B,
normal and DT1ALA bars), but it decreased precipitously with
time in all the other control groups (bars between normal and DT
1ALA). In soluble leishmania donovani (SLD)-specific re-stimu-
lation assays, lymphoproliferative response was negative
for all groups on pre-vaccination day 0 (Fig. 3C, blank bars)
and for the non-vaccinated control groups throughout
the p.c. period (normal, infected, and ALA). Cells from
porphyrinogenic vaccinated hamsters produced significantly
higher response (po0.001), which reached almost to the maxi-
mum on day 45 post-vaccination and increased further thereafter
Figure 1. Histopathology of the skin of hamsters vaccinated withtransgenic-L. amazonensis. Tissues were collected 3 wk post-vaccina-tion from hamsters immunized with DT1ALA (A and B), DT�ALA(C and D), and ST1ALA (E and F). Sections were processed and stainedwith H&E. Micrographs taken under 40� and 100� objectives shownin the left panel (A, C, E) and the right panel (B, D, F), respectively. Note:the normal microarchitecture of the skin (A) and the absence ofamastigote-infected macrophages (B) in the porphyrinogenic DT1ALAvaccinated hamsters; inflammatory responses due to cell infiltrationof the skin (C, E) and abundant vacuolated macrophages containingLeishmania-amastigotes (D, F circles and arrows) in non-porphyrino-genic DT�ALA and ST1ALA vaccinated hamsters.
Eur. J. Immunol. 2009. 39: 178–191Shraddha Kumari et al.180
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
(Fig. 3C, DT1ALA). In contrast, those from non-porphyrinogenic
vaccinated hamsters registered only a slight increase of this
response on day 45 post-vaccination, but declined almost to the
baseline level thereafter on day 120 shortly before their death
(ST1ALA and DT�ALA).
Lymphocyte-mediated activation of macrophages to produce
NO for leishmanicidal activities was found to differ between
control and experimental groups of hamsters. Macrophages
isolated from naı̈ve hamsters were exposed to the supernatants of
SLD-stimulated lymphocytes from these animals. Porphyrino-
genic vaccinated group was found to elicit significantly higher
level (po0.001) of NO, which increased with the p.c. periods up
to day 180 (Fig. 3D, DT1ALA). Non-porphyrinogenic controls
(Fig. 3D, ST1ALA, DT�ALA) elicited a modest increase of NO
production, which was seen only on day 45 p.c. and declined
gradually thereafter to the baseline levels, as observed in the
other control groups (normal, infected control, and ALA).
Porphyrinogenic vaccination alters Leishmania-specificIgG and its isotypes
The serum levels of leishmanial antigen-specific IgG and its
isotypes (IgG1 and IgG2) from all groups were assessed by ELISA.
The anti-Leishmania IgG and IgG1 were elevated progressively
with time to a high level in all groups, except the porphyrino-
genic, in which case they remained essentially the background
levels of the non-immunized and unchallenged normal (Fig. 3E
and F). In contrast, the porphyrinogenic (DT1ALA) was the only
group (Fig. 3G), which showed a significant elevation by one- to
twofold over the others (po0.001) in the level of IgG2. As a
measure of CMI, the elevation of IgG2 is consistent with the
development of effective immune response.
Porphyrinogenic vaccination of hamsters elicit cyto-kine transcript profiles of protective response
Impairment of CMI response during active VL is reflected by
marked T-cell anergy specific to Leishmania antigens [17, 18].
Since optimum protective efficacy was observed in porphyrinogenic
vaccinated hamsters, the expression of Th1 and Th2 cytokines was
evaluated only in this group for comparison with infected and
normal control groups. A comparative RNA cytokine profile of
splenocytes, analyzed on days 45, 60, 120, and 180 p.c. (Fig. 4),
showed that among the three groups of hamsters, expression of
IFN-g and IL-12 transcripts was significantly higher in vaccinated
group than infected groups (po0.001) (Fig. 4A). These and iNOS
Figure 2. Photodynamic vaccination of hamsters with suicidal L. amazonensis protects them against VL. Shown are body weights (A), organ weights(gm) (B, C), and parasite burdens in spleen (D), liver (E), and bone marrow (F) of vaccinated hamsters, i.e. DT1ALA, DT�ALA, ST1ALA, and non-vaccinated controls, i.e. untreated/non-challenged (normal), untreated/challenged (infected), and ALA alone/challenged (ALA). Parasite loads weredetermined as the number of amastigotes/103 cell nuclei microscopically enumerated in Geimsa-stained tissue impression smears. Each barrepresents pooled data (mean7SD) from three independent experiments (10–15 hamsters per group). ���po0.001 and ��po0.01 compared withrespective normal (A–C) or infected (D–F) controls.
Eur. J. Immunol. 2009. 39: 178–191 Immunity to infection 181
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

transcripts were all elevated by three- to fourfold after vaccination
(Fig. 4B). The expression levels of Th1-suppressive cytokines, i.e.
TGF-b, IL-10, and IL-4 were up-regulated in infected group, but not
in porphyrinogenic vaccinated hamsters throughout the p.c. periods
from day 45 to day 180 (Fig. 4B).
Adoptive transfer of T cells from porphyrinogenicvaccinated group protects naı̈ve hamsters againstL. donovani challenges by generating specific cell-mediated immune response
Protection of hamsters against L. donovani challenges by
porphyrinogenic vaccination (Figs. 1 and 2) was unequivocally
confirmed by adoptive transfer of T lymphocytes from immunized
animals to naı̈ve hamsters (Fig. 5, DT1ALA/R). The recipients of
immune T cells were observed to gain as much weight as the
normal hamsters, both groups being 2–3 times heavier than the
animals in the group, which received no T cells, but challenged
with L. donovani (Fig. 5A, LD/R). In the protected group
(DT1ALA/R), hepatosplenomegaly was totally absent through-
out the entire p.c. period; parasite loads were significantly
suppressed in all visceral organs from day 45 to day 180, i.e. a
decrease by 41 log in the spleen and to negligible levels in the
liver and bone marrow (Fig. 5D–F). All the hamsters in this group
survived and remained healthy for 1 year when the experiment
was terminated. On the other hand, the animals receiving cells
from non-vaccinated, but infected hamsters (LD/R) developed
visceral disease and died of the challenging infection in �2
months. Although some non-porphyrinogenic control groups
were precluded from this study due to their death, the results
obtained provide significant evidence, indicating that the
protective immunity elicited by photodynamic vaccination is
transferable and thus long lasting.
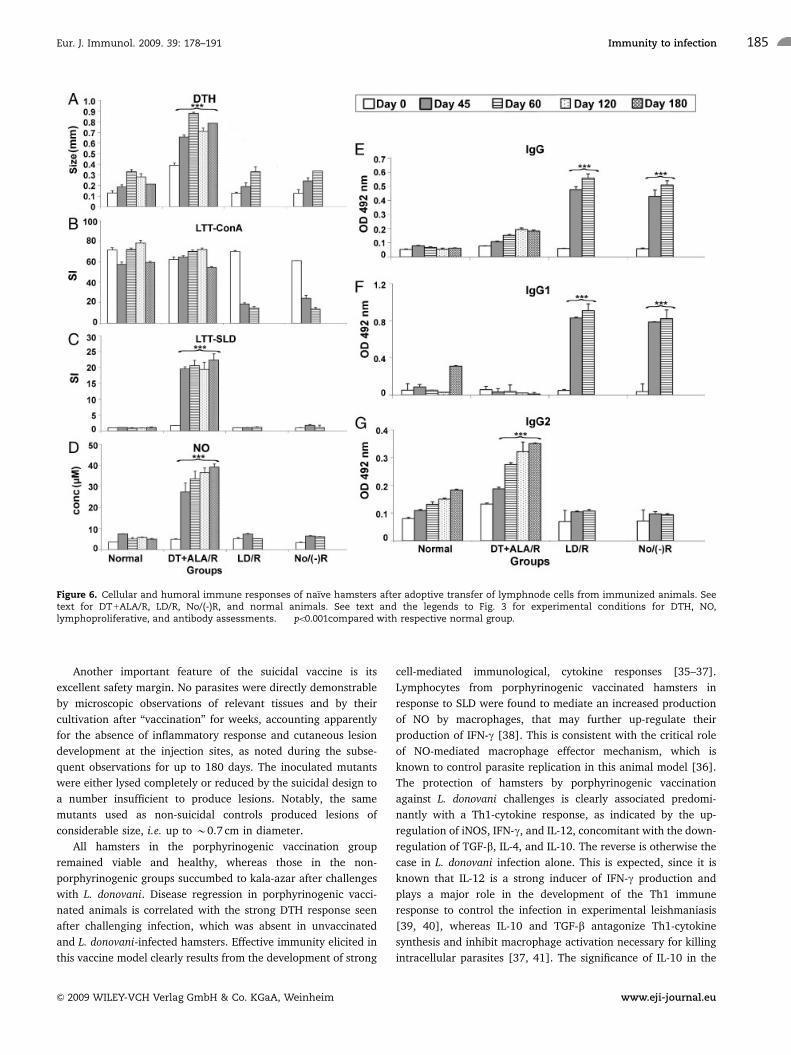
The transfer of protective immunity from vaccinated animals
to the recipients was indicated by their cellular responses
(Fig. 6A–D, DT1ALA/R), which were significantly higher than
those of the control groups (po0.001), i.e. DTH, lymphoproli-
ferative responses, and NO production against SLD throughout
the p.c. periods from day 0 to day 180.
As expected, while the Leishmania-specific IgG and IgG1 were
of the background levels in the DT1ALA/R group, its IgG2 level
Figure 3. Cellular and humoral immune responses of photodynamically vaccinated hamsters. These were assessed at the same time points fromthe same groups of hamsters, as described in the legend to Fig. 2. Shown are DTH response (mm) to SLD as footpad swelling at 24 h (A) andlymphoproliferative response (SI) of lymphnode cells to Con A (B) and SLD (C). NO production (mg/mL) of macrophages measured in samples (seetext) by the absorbance of the reaction products at 540 nm using Greiss reagent (D); anti-leishmanial IgG levels (OD) (E) and its isotypes, i.e. IgG1 (F)and IgG2 (G), determined by ELISA. Each bar represents the pooled data (mean7SD) of three independent experiments each with 10–15 hamsters.���po0.001 compared with respective normal group.
Eur. J. Immunol. 2009. 39: 178–191Shraddha Kumari et al.182
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

became increasingly elevated with the p.c. periods; the reverse
was observed in all the control groups, e.g. LD/R (Fig. 6E–G).
There was significant up-regulation of iNOS, IFN-g, and IL-12,
concomitant with down-regulation of Th2 cytokines (IL-10, IL-4,
and TGF-b) (po0.001) in DT1ALA/R versus the infected control
groups (Fig. 7). All these findings corroborate our earlier results
with porphyrinogenic vaccination (Fig. 4).
DiscussionMildly virulent Leishmania (from the lesions of diseased patients)
have been used for centuries to vaccinate children (known as
‘‘leishmanization’’) with proven effectiveness for immunoprophy-
laxis against human simple CL – a disease known to heal
spontaneously, resulting in the development of life-long immunity
[19]. ‘‘Leishmanization’’ becomes more feasible for consideration,
with the advances in Leishmania molecular genetics to knockout
virulence genes or to express transgenic suicidal genes [20–24]. In
the present study, a transgenic mutant line of L. amazonensis was
demonstrated to have the potential as a suicidal and thus safe
vaccine candidate to facilitate antigen presentation, thereby
enhancing the magnitude and quality of T-cell immune responses.
A single subcutaneous injection of such a mutant was reported
here to elicit a protective immune response of hamster against
otherwise fatal challenges of L. donovani.
The doubly transgenic-Leishmania used become porphyric
only when exposed to ALA [16] – a water soluble inexpensive
metabolite that is in clinical use for photodynamic therapy
against cancers [25], but not for vaccination using Leishmania in
the manner as presented in this study. The closest is the bacterial
mutants of Legionella whose DNA repair genes were obliterated,
thereby rendering them sensitive to psoralin-dependent and UV-
sensitive lysis [26]. The potential use of suicidal Leishmania for
live-cell vaccination finds support from previous reports on the
immunization of laboratory animals with drug or genetically
attenuated mutants for avirulence [22, 27, 28]. A major differ-
ence of the present suicidal design from the previous ones is the
use of two very different suicidal signals in our model, i.e. ALA
Figure 4. Th1/Th2 cytokine and iNOS mRNA in protected porphyrinogenic-L. amazonensis vaccinated hamsters. (A) Splenic iNOS and cytokinemRNA expression profile analysis of normal, infected, and porphyrinogenic vaccinated hamsters challenged with L. donovani (DD8). Spleens ofhamsters were analyzed on days 45, 60, 120, and 180 p.c. for RT-PCR. Experiments were carried out in triplicate for each of the three representativehamsters from individual experimental groups. (B) Densitometry analysis of the data in (A) showing the relative mean % change in iNOS andcytokine mRNA expression7SD normalized against control (HGPRT). The differences between various groups are significant, as indicated(��po0.01 and ���po0.001 compared with respective infected control).
Eur. J. Immunol. 2009. 39: 178–191 Immunity to infection 183
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

and light. Both have long been used clinically for different
purposes with excellent margin of safety and therapeutic
values. The use of two different signals in combination, although
laborious and cumbersome in practice, offers the modality of
fine-tuning the strengths and timing to deliver adequate
cytolysis of the live vaccines. This is further amenable to further
regulation, if necessary, by application of an additional photo-
sensitizer externally [29]. We have found no evidence for
‘‘escape’’ of the live mutants from the ‘‘treatment’’ for cytolysis by
leaving the vaccination sites prematurely. This is further
preventable in the future by using strictly cutanotropic Leishma-
nia and/or by rendering them porphyric before injection.
The suicidal design is thus particularly suitable for assessing
challenging parasite ‘‘persistence’’ [27, 30, 31] versus inanimated
vaccines, e.g. killed-microorganisms, peptide antigens, and
cDNA [3, 32], which may be used in combination or separately
for comparison in their ability to elicit memory T cells for lasting
immunity.
An ‘‘ideal’’ anti-leishmanial vaccine is the one, which can be
effective against both CL and VL, considering their co-endemicity
in many places. Cross-species protection has been reported
previously in experimental leishmaniasis [19, 33, 34]. Here, we
demonstrated this for the first time that a suicidal mutant
L. amazonensis induces cross-protective immune response against
a virulent L. donovani challenge. This suicidal design for photo-
dynamic vaccination makes it possible not only to elicit protective
immunity against L. donovani but also to let the live vaccine
undergo self-destruction. Although our results demonstrated the
utility of the mutants for eliciting protection against leishma-
niasis, they have the potential to serve more universally as a
platform to receive add-on vaccines for immunization against
other infectious and non-infectious diseases.
Figure 5. Protection of naı̈ve hamsters by adoptive transfer of lymphnode cells from immunized animals. Shown are body weights (A), organweights (gm) (B, C), and parasite burdens in spleen (D), liver (E), and bone marrow (F) of the recipient hamsters, i.e. DT1ALA/R, LD/R, No/(-)R, andnormal animals determined on different p.c. days as shown. See text and the legend to Fig. 2 for the assessment of parasite burdens. Each barrepresents pooled data (mean7SD) from three independent experiments (10–15 hamsters per group). ���po0.001 compared with respective normal(A–C) or infected control (D–F).
Eur. J. Immunol. 2009. 39: 178–191Shraddha Kumari et al.184
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
Another important feature of the suicidal vaccine is its
excellent safety margin. No parasites were directly demonstrable
by microscopic observations of relevant tissues and by their
cultivation after ‘‘vaccination’’ for weeks, accounting apparently
for the absence of inflammatory response and cutaneous lesion
development at the injection sites, as noted during the subse-
quent observations for up to 180 days. The inoculated mutants
were either lysed completely or reduced by the suicidal design to
a number insufficient to produce lesions. Notably, the same
mutants used as non-suicidal controls produced lesions of
considerable size, i.e. up to �0.7 cm in diameter.
All hamsters in the porphyrinogenic vaccination group
remained viable and healthy, whereas those in the non-
porphyrinogenic groups succumbed to kala-azar after challenges
with L. donovani. Disease regression in porphyrinogenic vacci-
nated animals is correlated with the strong DTH response seen
after challenging infection, which was absent in unvaccinated
and L. donovani-infected hamsters. Effective immunity elicited in
this vaccine model clearly results from the development of strong
cell-mediated immunological, cytokine responses [35–37].
Lymphocytes from porphyrinogenic vaccinated hamsters in
response to SLD were found to mediate an increased production
of NO by macrophages, that may further up-regulate their
production of IFN-g [38]. This is consistent with the critical role
of NO-mediated macrophage effector mechanism, which is
known to control parasite replication in this animal model [36].
The protection of hamsters by porphyrinogenic vaccination
against L. donovani challenges is clearly associated predomi-
nantly with a Th1-cytokine response, as indicated by the up-
regulation of iNOS, IFN-g, and IL-12, concomitant with the down-
regulation of TGF-b, IL-4, and IL-10. The reverse is otherwise the
case in L. donovani infection alone. This is expected, since it is
known that IL-12 is a strong inducer of IFN-g production and
plays a major role in the development of the Th1 immune
response to control the infection in experimental leishmaniasis
[39, 40], whereas IL-10 and TGF-b antagonize Th1-cytokine
synthesis and inhibit macrophage activation necessary for killing
intracellular parasites [37, 41]. The significance of IL-10 in the
Figure 6. Cellular and humoral immune responses of naı̈ve hamsters after adoptive transfer of lymphnode cells from immunized animals. Seetext for DT1ALA/R, LD/R, No/(-)R, and normal animals. See text and the legends to Fig. 3 for experimental conditions for DTH, NO,lymphoproliferative, and antibody assessments. ���po0.001compared with respective normal group.
Eur. J. Immunol. 2009. 39: 178–191 Immunity to infection 185
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

pathogenesis of human VL is supported by the observations that it
down-regulates parasite-specific T-cell responses [42] and inhi-
bits NO production of human macrophages to kill L. infantum
[43]. These observations corroborate our data presented here,
indicating that the progressive disease in this kala-azar model is
associated with a defect in the generation of NO.
Apart from diminished cellular responses, VL is associated
with the production of high levels of antibodies, which have been
observed prior to detection of parasite-specific T-cell response
[44]. Unlike mice where IL-4 and IL-12 direct IgG subclass
switching of IgG1 and IgG2a, respectively, such distinct IgG
classes remain obscure in hamsters [45, 46]. It is believed that
hamster IgG1 and IgG2 correspond to mouse IgG1 and IgG2a/
IgG2b, respectively. It has been well established that IgG and
IgG1 antibodies increase in titer with the L. donovani loads [47].
The virtual absence of these antibodies is thus consistent with the
decreasing parasite loads seen in porphyrinogenic vaccinated
group. The significant increase in the IgG2 levels only in
porphyrinogenic vaccinated animals is indicative of enhanced
CMI.
Protective immunity can be passively transferred to naı̈ve
recipients with T-cell enriched preparations from donor animals
that have resolved infection [48]. Evidence obtained in human
and in experimental murine leishmaniasis indicates that the
healing of cutaneous lesions is associated with the development
of specific T-cell-dependent cellular response [49, 50], the role of
which has been further established by experiments showing that
resistance to infection was achieved by the transfer of syngeneic
immune T lymphocytes [51]. Adoptive transfer of the splenocytes
from immunized animals has been shown previously to achieve a
certain level of protection against L. donovani infection [52–54].
Successful protection was achieved in the present study by a
single intravenous administration of T cells from porphyrinogenic
vaccinated hamsters, conferring an effective control of infection
on hamsters against L. donovani challenges, as indicated both at
the level of restricting parasite loads and induction of protective
Figure 7. Th1/Th2 cytokine and iNOS mRNA of hamsters adoptively transferred with lymphnode cells from porphyrinogenic-L. amazonensisvaccinated hamsters. See legend to Fig. 5 for group designation. See text and the legend to Fig. 4 for experimental and other details. (A) SpleniciNOS and cytokine mRNA expression profiles; (B) densitometric analyses of data in part A, as described in the legend to Fig. 4 (��po0.01 and���po0.001 compared with respective infected control).
Eur. J. Immunol. 2009. 39: 178–191Shraddha Kumari et al.186
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

cellular responses. In animals of the DT1ALA/R group, indeed,
the virtual absence of parasites on day 180 p.c. is accompanied by
a strong induction of favorable immune parameters, i.e. antigen-
specific T-cell proliferation, production of NO, increased mRNA
expression of iNOS, IL-12, IFN-g, robust DTH responses, and low
antibody titers. Clearly, donor T cells are skewed toward a
protective Th1-type response, indicative of durable immunity.
Interestingly, the overall protective and immunological profiles
are more pronounced in DT1ALA/R group than the porphyr-
inogenic vaccinated ones. Thus, passive transfer of immunity in
the absence of parasites may favor the development of central
memory T cells for long-lasting protection.
Evidence presented in this study clearly indicates that the
porphyrinogenically lysed L. amazonensis after delivery in vivo is
protective against experimental VL. Worthy of emphasis are two
major findings. Firstly, porphyrinogenic-L. amazonensis appears
to be a promising live candidate vector for development of an
effective vaccine against Leishmania infections. Secondly, transfer
of T cells from effectively immunized animals alone, in the
absence of parasites, to the naı̈ve animals protects the latter with
robust and lasting immunity, suggestive of the development of
central memory T cells. It will be highly advantageous to obtain
such lasting immunity against VL by using porphyrinogenic
mutants from strictly cutaneous Leishmania, as their residence in
the skin makes them readily accessible to suicidal induction.
The work presented offers a new avenue for photodynamic
immunoprophylaxis and immunotherapy of other infectious
diseases [16]. Further studies may be envisaged to simplify the
applications, for example, by one-time administration of the
mutants followed by inducing their porphyria using commercially
available ALA ointment and photolysis with bandaged light
emitting diodes (heatless light emitting diodes).
Materials and methods
Animal
Laboratory bred male golden hamsters (M. auratus, 45–50 g)
from the Institute’s Animal Facility were used as the experimental
host. They were housed in climatically controlled room and fed
with standard rodent food pellet (Lipton India, Mumbai, India)
and water ad libitum. The usage of the animals was approved by
the Institute’s Animal Ethical Committee.
Parasites
Leishmania donovani
Wild-type (MHOM/IN/80/DD8) has been maintained by amas-
tigote to amastigote serial passages every �2 months in
hamsters. The parasites were isolated and purified from the
spleen of infected hamsters for challenging infection.
For the preparation of SLD, promastigotes were grown in L-15
medium (Sigma, USA) for 3–4 days and processed as before [55].
The protein content of the SLD was estimated [56] and stored at
�701C.
Leishmania amazonensis mutants
The construction, selection, and maintenance of L. amazonensis
non-porphyrinogenic-single transfectants (ST) and porphyrino-
genic-double transfectants (DT), used for vaccination, were
described elsewhere [16]. Briefly, wild-type L. amazonensis
(LV78) promastigotes (clone 12-1) were grown at 251C in
Medium 199 HEPES-buffered to pH 7.4 and supplemented with
10% heat-inactivated fetal bovine serum. Transfectants were
grown under similar conditions with different concentrations of
selective pressures, i.e. G418 and/or tunicamycin at 100 and/or
20 mg/mL, respectively. Cultures were grown for one cycle in
drug-free medium before use. These mutants (109 cells/mL) were
also heat-killed by autoclaving for 15 min and used in a separate
set of experiments for comparison of their prophylactic potentials
with porphyrinogenic and non-porphyrinogenic cells.
Vaccination with transgenic-L. amazonensis inhamsters
In three separate experiments, hamsters were vaccinated on the
back on shaved areas to facilitate their exposure to white light
illumination from the top after ALA treatment (20mL of 100 mM
ALA in Hank’s Balanced Salt Solution, pH 7.4) to induce specific
uroporphyria of intracellular transgenic-Leishmania. A total of 90
hamsters were divided into six groups at 10–15 per group. Hamsters
of the main experimental group or porphyrinogenic vaccinated
group (DT1ALA) were each vaccinated first with DT and then
administered with ALA 2–3 days later. The hamsters of second
group (DT�ALA) were vaccinated with DT without ALA and those
of the third group were given ST and ALA (ST1ALA) similarly as
described for the first group. The hamsters of the fourth group were
treated with ALA alone and those of the fifth were kept as
unvaccinated control. All animals in above-mentioned five groups
were challenged with L. donovani amastigotes as described [38].
The sixth and the last group comprised of unvaccinated and
unchallenged control animals. The second and third groups were
designated as the non-porphyrinogenic control groups. The experi-
mental schedules for vaccination are given in Table 1.
For histological studies, five hamsters were ‘‘vaccinated’’ as
described for the first group and sacrificed 3 wk after the vacci-
nation, the skin tissues at the site of injection, draining lymph-
nodes, and spleens were removed for histology sections and
assessed microscopically after staining with H&E.
Peritoneal exudates, lymphnodes, spleens, and blood were
collected from the hamsters of all groups sacrificed at different
time points as mentioned in Table 1 to obtain cells and sera for
evaluation of cellular and antibody responses. The criterion for
prophylactic efficacy included the assessment of parasite loads as
Eur. J. Immunol. 2009. 39: 178–191 Immunity to infection 187
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

the number of amastigotes/103 splenic cell nuclei in Geimsa-
stained touch-blots of spleens, livers, and bone marrows. The
percentage inhibition was assessed as described elsewhere [38].
Various tissues were also incubated in vitro in NNN tubes with
RPMI-1640 as an overlay to assess the presence or absence of
viable amastigotes therein based on their conversion into
promastigotes.
Adoptive transfer of T-cell immunity from porphyr-inogenic vaccinated hamsters
For this experiment, a total of 40–60 hamsters at 10–15 per group
were used. The experimental groups were categorized as follows:
DT1ALA/R (animals receiving lymphocytes from porphyrino-
genic vaccinated group sacrificed on day 180 p.c.); LD/R
(animals receiving lymphocytes from non-vaccinated but
L. donovani challenged group); No/(-)R (naı̈ve animals receiving
no lymphocytes). Lymphocytes used were isolated from lymph-
nodes (inguinal and mesenteric) of porphyrinogenic vaccinated
(DT1ALA) and/or L. donovani challenged animals (on day 180
p.c.), and the isolated cells (107/100 mL) were suspended in
complete RPMI medium [38]. Hamsters were injected with 107
lymphocytes per animal intracardially and were challenged 21
days later with L. donovani (DD8) amastigotes (107 per animal),
except the normal group, which received neither lymphocytes nor
L. donovani challenges. Progression of disease and immunological
responses were monitored periodically up to 180 days p.c. when
the experiment was terminated.
Post-challenge survival assessments
Animals of both the experimental and control groups were given
proper care and observed for their survival which lasted for more
than 12 months p.c. Survival of individual hamsters was recorded
and mean survival period was calculated.
Lymphoproliferative assays
Lymphnodes of hamsters were excised aseptically and processed
for the isolation of lymphocytes [38]. The lymphocyte were
suspended to 106/mL and cultured at 105 cells/well in 96-well flat
bottom tissue culture plates (Nunc, Denmark). One hundred
microliters of Con A (10mg/mL; Sigma) or SLD (10mg proteins/
mL) were added to each well in triplicate. Wells without stimulants
served as negative controls. Cultures were incubated at 371C in a
CO2 incubator for 3 days in case of mitogen and for 5 days in case
of SLD antigens. Eighteen hours prior to termination of culture,
0.5mCi of [3H] thymidine (BARC, India) was added to each well
and then cells were harvested on glass fiber mats (Whatman);
radioactivity counted in a liquid scintillation counter. Results were
expressed as stimulation index (SI), which was calculated as mean
cpm of stimulated culture/mean cpm of unstimulated control. SI
values of more than 2.5 were considered as positive response.
NO assays
The presence of NO was assessed in the culture supernatants of
peritoneal macrophages from naı̈ve hamsters after exposure to
the supernatants of stimulated lymphocyte’s cultures by using
Griess reagent [57]. Isolated peritoneal macrophages [55] were
suspended in culture medium for plating at 106 cells/well and
exposed to the supernatants of above-described stimulated
lymphocyte’s culture supernatants from all the study groups.
The supernatants (100 mL) collected from macrophage cultures
24 h after incubation were each mixed with an equal volume of
Griess reagent (Sigma) and left for 10 min at room temperature.
Table 1. Experimental schedules for vaccination
Step Day (p.v./p.c.) Group Treatment
1 0 1–3 Intradermal inoculation of a shaved area (�2 cm2) on the back of the animals, with
porphyrinogenic DT, non-porphyrinogenic ST and heat-killed DT (1.0–1.5�107 cells/100mL
each), respectively
4–6 No vaccination controls
2 4–8 p.v. 1,3,
and 4
ALA administration four times daily to the vaccination site of each animal for ‘‘induction of
porphyria’’ with 20 mL of 100 mM ALA in HBSS, pH 7.4
2,5,
and 6
No ALA treatment
3 8–12 p.v. 1–6 Illumination of caged animals for�8 h daily � 5 days under white light (four fluorescent bulbs
of 100 W (Phillips) 1 two fluorescent tube lights of 40 W) placed 2 ft above the cages, thereby
maintaining a constant temperature of 26–271C
4 26 p.v. 1–5 Challenge intracardially with 107 DD8 strain of L. donovani amastigotes/animal
6 No challenge
5 0, 45, 60, 120, and
180 p.c.
1–6 Examination of 2–3 hamsters for pathological, parasitological, and immunological evaluation
p.v., post-vaccination; p.c., post-challenge.
Eur. J. Immunol. 2009. 39: 178–191Shraddha Kumari et al.188
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

The absorbance of the reaction products was measured at 540 nm
in an ELISA reader [57].
Measurements of DTH
DTH was performed by injecting intradermally 50 mg/50 mL of
SLD in PBS into one footpad and PBS alone into the other footpad
of each of the vaccinated and unvaccinated controls. The
response was evaluated 24 h later by measuring the difference
in footpad swelling between the two with and without SLD for
each animal [45].
RT-PCR of mRNA for cytokines and iNOS
RT-PCR was performed to assess the expression of mRNA for
various cytokines and iNOS in splenic cells. Splenic tissues
were taken from each of the three individual animals randomly
chosen from different groups. Total RNA were isolated using Tri-
reagent (Sigma) on days 45, 60, 120, and 180 p.c., and quantified
by using Gene-quant (BioRad). The primer sequences of cytokine
and iNOS primers used to amplify their respective cDNA were as
described [37] (Table 2). Housekeeping hypoxanthine-guanine
phosphoribosyltransferase (HGPRT) was used as a control.
One microgram of total RNA was used for the synthesis of
cDNA using first strand cDNA synthesis kit (Fermentas). An
amount of 0.5 mg of cDNA was amplified by PCR under the
following condition: Initial denaturation at 951C for 2 min
followed by 40 cycles, each consisting of denaturation at 951C for
30 s, annealing at 551C for 40 s and extension at 721C for 40 s per
cycle. The final extension step was carried out at 721C for 10 min.
RT-PCR products were quantitatively assessed by densitometry of
ethidium bromide-stained bands using the software AlphaIma-
gerTM 2200 (Alfainnotech). The mean percentage expression
values were presented as the intensities of PCR products relative
to those of their corresponding HGPRT.
Determination of Leishmania-specific anti-leishmanialantibody levels
The levels of anti-leishmanial antibodies – IgG and its isotypes –
IgG1 and IgG2 in sera samples from hamsters of different
groups were measured as described [58]. The 96-well ELISA
plates (Nunc) were coated in each well with SLD (2 mg/mL in
PBS) overnight at 41C and blocked with 1% BSA at room
temperature for 1 h. The optimum dilution of sera was
standardized at 1:200 for IgG, and 1:100 for IgG1 and IgG2,
respectively for 2 h at room temperature. HRP-conjugated goat
anti-hamster IgG (H1L) (Serotec, USA) and biotin-conjugated
mouse anti-Armenian and anti-Syrian hamster IgG1 (for IgG1)
(BD, Pharmingen) as well as mouse anti-Syrian hamster IgG2
(BD, Pharmingen) was added for 1 h at room temperature. IgG1
and IgG2 plates were further incubated with sterptavidin-
conjugated peroxidase (Sigma) for 1 h. Finally, plates were
developed using O-phenylene diamine dihydrochloride as the
substrate (Sigma) and the reaction products read at 492 nm in an
ELISA reader (BioTek, USA).
Statistical analysis
Results were expressed as meanSD. The results (pooled data of
three independent experiments) were analyzed by one-way
ANOVA test and comparisons with control data were made with
Dunnett’s post-test using Graph Pad Prism software program. The
upper level of significance was chosen as po0.001.
Acknowledgements: We expressed our sincere gratitude to the
Director CDRI for his keen interest and for providing facilities for
the experiments. Financial support for senior research fellowship
to S.K. and M.S. from CSIR, New Delhi and technical support of
S. M. Verma, Division of Toxicology are gratefully acknowledged.
This has CDRI communication no. 7449. Supported in part by
USA-NIH AI-68835 to K.P.C.
Conflict of interest: The authors declare no financial or
commercial conflict of interest.
References
1 Desjeux, P., Leishmaniasis: current situation and new perspectives. Comp.
Immunol. Microbiol. Infect. Dis. 2004. 27: 305–318.
Table 2. Primer sequences for RT-PCR amplification of hamstertranscripts
Transcript
of
Primer sequence
(forward/reverse)
Expected
size (bp)
HGPRT 50-ATCACATTATGGCCCTCTGTG-30 125
50-CTGATAAAATCTACAGTYATGG-30a)
iNOS 50-GCAGAATGTGACCATCATGG-30 198
50-CTCGAYCTGGTAGTAGTAGAA-30
IFN-g 50-GGATATCTGGAGGAACTGGC-30 309
50-CGACTCCTTTTCCGCTTCCT-30
IL-12 50-GTACACCTGYCACAAAGGAG-30 430
50-GATGTCCCTGATGAAGAAGC-30
TGF-b 50-CCCTGGAYACCAACTATTGC-30 310
50-ATGTTGGACARCTGCTCCAC-30
IL-4 50-CATTGCATYGTTAGCRTCTC-30a) 463
50-TTCCAGGAAGTCTTTCAGTG-30
IL-10 50-ACAATAACTGCACCCACTTC-30 432
50-AGGCTTCTATGCAGTTGATG-30
a) Sequences with base degeneracy (Y 5 C and T; R 5 A and G).
Eur. J. Immunol. 2009. 39: 178–191 Immunity to infection 189
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

2 Alvar, J., Aparicio, P., Aseffa, A., Den Boer, M., Canavate, C., Dedet, J. P.,
Gradoni, L. et al., The relationship between leishmaniasis and AIDS: the
second 10 years. Clin. Microbiol. Rev. 2008. 21: 334–359.
3 Handman, E., Leishmaniasis: current status of vaccine development. Clin.
Microbiol. Rev. 2001. 14: 229–243.
4 Sundar, S., Drug resistance in Indian visceral leishmaniasis. Trop. Med. Int.
Health 2001. 6: 849–854.
5 Reed, S. G. and Scott, P., T-cell and cytokine responses in leishmaniasis.
Curr. Opin. Immunol. 1993. 5: 524–531.
6 Melby, P. C., Tryon, V. V., Chandrasekar, B. and Freeman, G. L., Cloning of
Syrian hamster (Mesocricetus auratus) cytokine cDNAs and analysis of
cytokine mRNA expression in experimental visceral leishmaniasis. Infect.
Immun. 1998. 66: 2135–2142.
7 Garg, R. and Dube, A., Animal models for vaccine studies for visceral
leishmaniasis. Indian J. Med. Res. 2006. 123: 439–454.
8 Greenblatt, C. L., The present and future of vaccination for cutaneous
leishmaniasis. Prog. Clin. Biol. Res. 1980. 47: 259–285.
9 Breton, M., Tremblay, M. J., Ouellette, M. and Papadopoulou, B., Live
nonpathogenic parasitic vector as a candidate vaccine against visceral
leishmaniasis. Infect. Immun. 2005. 73: 6372–6382.
10 WHO/TDR, Coming Back: WHO/TDR initiative to support live attenuated
vaccines. TDR News September 2005. No. 75.
11 Bretscher, P. A., Menon, J. N. and Ogunremi, O., Towards a strategy of
universally efficacious vaccination against pathogens uniquely suscep-
tible to cell-mediated attack. J. Biotechnol. 1996. 44: 1–4.
12 Daneshvar, H., Coombs, G. H., Hagan, P. and Phillips, R. S., Leishmania
mexicana and Leishmania major: attenuation of wild-type parasites
and vaccination with the attenuated lines. J. Infect. Dis. 2003. 187:
1662–1668.
13 Dumas, C., Muyombwe, A., Roy, G., Matte, C., Ouellette, M., Olivier, M.
and Papadopoulou, B., Recombinant Leishmania major secreting biologi-
cally active granulocyte-macrophage colony-stimulating factor survives
poorly in macrophages in vitro and delays disease development in mice.
Infect. Immun. 2003. 71: 6499–6509.
14 Muyombwe, A., Olivier, M., Harvie, P., Bergeron, M. G., Ouellette, M. and
Papadopoulou, B., Protection against Leishmania major challenge infection
in mice vaccinated with live recombinant parasites expressing a
cytotoxic gene. J. Infect. Dis. 1998. 177: 188–195.
15 Mendez, S., Tabbara, K., Belkaid, Y., Bertholet, S., Verthelyi, D., Klinman,
D., Seder, R. A. and Sacks, D. L., Coinjection with CpG-containing
immunostimulatory oligodeoxynucleotides reduces the pathogenicity of
a live vaccine against cutaneous leishmaniasis but maintains its potency
and durability. Infect. Immun. 2003. 71: 5121–5129.
16 Sah, J. F., Ito, H., Kolli, B. K., Peterson, D. A., Sassa, S. and Chang, K. P.,
Genetic rescue of Leishmania deficiency in porphyrin biosynthesis
creates mutants suitable for analysis of cellular events in
uroporphyria and for photodynamic therapy. J. Biol. Chem. 2002. 277:
14902–14909.
17 Haldar, J. P., Ghose, S., Saha, K. C. and Ghose, A. C., Cell-mediated
immune response in Indian kala-azar and post-kala-azar dermal
leishmaniasis. Infect. Immun. 1983. 42: 702–707.
18 Gifawesen, C. and Farrell, J. P., Comparison of T-cell responses in self-
limiting versus progressive visceral Leishmania donovani infections in
golden hamsters. Infect. Immun. 1989. 57: 3091–3096.
19 Kedzierski, L., Zhu, Y. and Handman, E., Leishmania vaccines: progress
and problems. Parasitology 2006. 133: S87–S112.
20 Titus, R. G., Gueiros-Filho, F. J., de Freitas, L. A. and Beverley, S. M.,
Development of a safe live Leishmania vaccine line by gene replacement.
Proc. Natl. Acad. Sci. USA 1995. 92: 10267–10271.
21 Alexander, B., Lozano, C., Barker, D. C., McCann, S. H., and Adler, G. H.,
Detection of Leishmania (Viannia) braziliensis complex in wild mammals
from Colombian coffee plantations by PCR and DNA hybridization. Acta
Trop. 1998. 69: 41–50.
22 Papadopoulou, B., Roy, G., Breton, M., Kundig, C., Dumas, C., Fillion, I.,
Singh, A. K. et al., Reduced infectivity of a Leishmania donovani biopterin
transporter genetic mutant and its use as an attenuated strain for
vaccination. Infect. Immun. 2002. 70: 62–68.
23 Spath, G. F., Garraway, L. A., Turco, S. J. and Beverley, S. M., The role(s) of
lipophosphoglycan (LPG) in the establishment of Leishmania major
infections in mammalian hosts. Proc. Natl. Acad. Sci. USA 2003. 100:
9536–9541.
24 Spath, G. F., Lye, L. F., Segawa, H., Sacks, D. L., Turco, S. J. and Beverley, S.
M., Persistence without pathology in phosphoglycan-deficient Leishmania
major. Science 2003. 301: 1241–1243.
25 Kelty, C. J., Brown, N. J., Reed, M. W. and Ackroyd, R., The use of
5-aminolaevulinic acid as a photosensitiser in photodynamic therapy
and photodiagnosis. Photochem. Photobiol. Sci. 2002. 1: 158–168.
26 Brockstedt, D. G., Bahjat, K. S., Giedlin, M. A., Liu, W., Leong, M., Luckett,
W., Gao, Y. et al., Killed but metabolically active microbes: a new vaccine
paradigm for eliciting effector T-cell responses and protective immunity.
Nat. Med. 2005. 11: 853–860.
27 Uzonna, J. E., Spath, G. F., Beverley, S. M. and Scott, P., Vaccination with
phosphoglycan-deficient Leishmania major protects highly susceptible
mice from virulent challenge without inducing a strong Th1 response.
J. Immunol. 2004. 172: 3793–3797.
28 Selvapandiyan, A., Duncan, R., Debrabant, A., Lee, N., Sreenivas, G., Salotra,
P. and Nakhasi, H. L., Genetically modified live attenuated parasites as
vaccines for leishmaniasis. Indian J. Med. Res. 2006. 123: 455–466.
29 Dutta, S., Ray, D., Kolli, B. K. and Chang, K. P., Photodynamic sensitization
of Leishmania amazonensis in both extracellular and intracellular stages
with aluminum phthalocyanine chloride for photolysis in vitro. Anti-
microb. Agents Chemother. 2005. 49: 4474–4484.
30 Scott, P., Immunologic memory in cutaneous leishmaniasis. Cell. Micro-
biol. 2005. 7: 1707–1713.
31 Tabbara, K. S., Peters, N. C., Afrin, F., Mendez, S., Bertholet, S., Belkaid, Y.
and Sacks, D. L., Conditions influencing the efficacy of vaccination with
live organisms against Leishmania major infection. Infect. Immun. 2005. 73:
4714–4722.
32 Coler, R. N. and Reed, S. G., Second-generation vaccines against
leishmaniasis. Trends Parasitol. 2005. 21: 244–249.
33 Dube, A., Sharma, P., Srivastava, J. K., Misra, A., Naik, S. and Katiyar, J. C.,
Vaccination of langur monkeys (Presbytis entellus) against Leishmania
donovani with autoclaved L. major plus BCG. Parasitology 1998. 116:
219–221.
34 Misra, A., Dube, A., Srivastava, B., Sharma, P., Srivastava, J. K., Katiyar,
J. C. and Naik, S., Successful vaccination against Leishmania donovani
infection in Indian langur using alum-precipitated autoclaved Leishmania
major with BCG. Vaccine 2001. 19: 3485–3492.
35 Howard, J. G. and Liew, F. Y., Mechanisms of acquired immunity in
leishmaniasis. Philos. Trans. R. Soc. London B: Biol. Sci. 1984. 307: 87–98.
36 Armijos, R. X., Weigel, M. M., Calvopina, M., Hidalgo, A., Cevallos, W. and
Correa, J., Safety, immunogenecity, and efficacy of an autoclaved
Leishmania amazonensis vaccine plus BCG adjuvant against New World
cutaneous leishmaniasis. Vaccine 2004. 22: 1320–1326.
Eur. J. Immunol. 2009. 39: 178–191Shraddha Kumari et al.190
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

37 Melby, P. C., Chandrasekar, B., Zhao, W. and Coe, J. E., The hamster as a
model of human visceral leishmaniasis: progressive disease and
impaired generation of nitric oxide in the face of a prominent Th1-like
cytokine response. J. Immunol. 2001. 166: 1912–1920.
38 Garg, R., Gupta, S. K., Tripathi, P., Hajela, K., Sundar, S., Naik, S. and
Dube, A., Leishmania donovani: identification of stimulatory soluble
antigenic proteins using cured human and hamster lymphocytes for
their prophylactic potential against visceral leishmaniasis. Vaccine 2006.
24: 2900–2909.
39 Scharton-Kersten, T. and Scott, P., The role of the innate immune
response in Th1 cell development following Leishmania major infection.
J. Leukoc. Biol. 1995. 57: 515–522.
40 Trinchieri, G., Interleukin-12: a proinflammatory cytokine with immu-
noregulatory functions that bridge innate resistance and antigen-specific
adaptive immunity. Annu. Rev. Immunol. 1995. 13: 251–276.
41 Kenney, R. T., Sacks, D. L., Gam, A. A., Murray, H. W. and Sundar, S.,
Splenic cytokine responses in Indian kala-azar before and after treat-
ment. J. Infect. Dis. 1998. 177: 815–818.
42 Ghalib, H. W., Piuvezam, M. R., Skeiky, Y. A., Siddig, M., Hashim, F. A., el-
Hassan, A. M., Russo, D. M. and Reed, S. G., Interleukin 10 production
correlates with pathology in human Leishmania donovani infections. J. Clin.
Invest. 1993. 92: 324–329.
43 Vouldoukis, I., Becherel, P. A., Riveros-Moreno, V., Arock, M., da Silva, O.,
Debre, P., Mazier, D. and Mossalayi, M. D., Interleukin-10 and interleukin-
4 inhibit intracellular killing of Leishmania infantum and Leishmania major
by human macrophages by decreasing nitric oxide generation. Eur. J.
Immunol. 1997. 27: 860–865.
44 Ghose, A. C., Haldar, J. P., Pal, S. C., Mishra, B. P. and Mishra, K. K.,
Serological investigations on Indian kala-azar. Clin. Exp. Immunol. 1980.
40: 318–326.
45 Bhowmick, S., Ravindran, R. and Ali, N., Leishmanial antigens in
liposomes promote protective immunity and provide immunotherapy
against visceral leishmaniasis via polarized Th1 response. Vaccine 2007.
25: 6544–6556.
46 Rodrigues, V., Jr., Da Silva, J. S. and Campos-Neto, A., Selective inability
of spleen antigen presenting cells from Leishmania donovani infected
hamsters to mediate specific T cell proliferation to parasite antigens.
Parasite Immunol. 1992. 14: 49–58.
47 Basu, R., Bhaumik, S., Basu, J. M., Naskar, K., De, T. and Roy, S.,
Kinetoplastid membrane protein-11 DNA vaccination induces complete
protection against both pentavalent antimonial-sensitive and -resistant
strains of Leishmania donovani that correlates with inducible nitric
oxide synthase activity and IL-4 generation: evidence for mixed Th1-
and Th2-like responses in visceral leishmaniasis. J. Immunol. 2005. 174:
7160–7171.
48 Rezai, H. R., Farrell, J. and Soulsby, E. L., Immunological responses
of L. donovani infection in mice and significance of T cell in
resistance to experimental leishmaniasis. Clin. Exp. Immunol. 1980. 40:
508–514.
49 Convit, J., Leishmaniasis: immunological and clinical aspects and
vaccines in Venezuela. Clin. Dermatol. 1996. 14: 479–487.
50 Howard, J. G., Hale, C. and Chan-Liew, W. L., Immunological regulation of
experimental cutaneous leishmaniasis. 1. Immunogenetic aspects of
susceptibility to Leishmania tropica in mice. Parasite Immunol. 1980. 2:
303–314.
51 Scott, P., Natovitz, P., Coffman, R. L., Pearce, E. and Sher, A., Immunor-
egulation of cutaneous leishmaniasis. T cell lines that transfer
protective immunity or exacerbation belong to different T helper subsets
and respond to distinct parasite antigens. J. Exp. Med. 1988. 168:
1675–1684.
52 Jarecki-Black, J. C., Glassman, A. B. and James, E. R., Adoptive transfer of
vaccine-induced resistance to Leishmania donovani. Am. J. Trop. Med. Hyg.
1985. 34: 1095–1097.
53 Ulczak, O. M., Ghadirian, E., Skamene, E., Blackwell, J. M. and
Kongshavn, P. A., Characterization of protective T cells in the
acquired response to Leishmania donovani in genetically determined cure
(H-2b) and noncure (H-2d) mouse strains. Infect. Immun. 1989. 57:
2892–2899.
54 Polley, R., Stager, S., Prickett, S., Maroof, A., Zubairi, S., Smith, D. F. and
Kaye, P. M., Adoptive immunotherapy against experimental visceral
leishmaniasis with CD81 T cells requires the presence of cognate
antigen. Infect. Immun. 2006. 74: 773–776.
55 Garg, R., Gupta, S. K., Tripathi, P., Naik, S., Sundar, S. and Dube, A.,
Immunostimulatory cellular responses of cured Leishmania-infected
patients and hamsters against the integral membrane proteins and
non-membranous soluble proteins of a recent clinical isolate of
Leishmania donovani. Clin. Exp. Immunol. 2005. 140: 149–156.
56 Lowry, O. H., Rosebrough, N. J., Farr, A. L. and Randall, R. J., Protein
measurement with the Folin phenol reagent. J. Biol. Chem. 1951. 193:
265–275.
57 Ding, A. H., Nathan, C. F. and Stuehr, D. J., Release of reactive nitrogen
intermediates and reactive oxygen intermediates from mouse peritoneal
macrophages. Comparison of activating cytokines and evidence for
independent production. J. Immunol. 1988. 141: 2407–2412.
58 Voller, A., Bidwell, D. E. and Bartlett, A., The Enzyme Linked Immunosorbent
Assay (ELISA): A Guide with Abstracts of Microplate Application, Fileroline
Press, Guernsey, CI 1979.
Abbreviations: ALA: d-aminolevulinate � CL: cutaneous leishmaniasis �CMI: cell-mediated immunity � DT: double transfectants � DTH:
delayed-type hypersensitivity � HGPRT: hypoxanthine-guanine
phosphoribosyltransferase � p.c.: post-challenges � SI: stimulation
index � ST: single transfectants � SLD: soluble leishmania donovani � VL:
visceral leishmaniasis � WHO: World Health Organization
Full correspondence: Dr. Anuradha Dube, Division of Parasitology,
Central Drug Research Institute, Chattar Manzil Palace, MG Road,
Lucknow 226001, India
Fax: 191-0522-2623938
e-mail: [email protected]
Received: 2/4/2008
Revised: 30/8/2008
Accepted: 1/10/2008
Eur. J. Immunol. 2009. 39: 178–191 Immunity to infection 191
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu




















