
Leishmania amazonensis META2 protein confers protection against heat shock and oxidative stress
11
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
Transcript of Leishmania amazonensis META2 protein confers protection against heat shock and oxidative stress

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Leishmania amazonensis META2 protein confers protection against heat shockand oxidative stress
Camila S. Ramos a,1, Jenicer K.U. Yokoyama-Yasunaka a, Cristina Guerra-Giraldez b,2, Helen P. Price b,Renato A. Mortara c, Deborah F. Smith b, Silvia R.B. Uliana a,⇑a Departamento de Parasitologia, Instituto de Ciências Biomédicas, Universidade de São Paulo, USP, São Paulo, SP, Brazilb Centre for Immunology and Infection, Department of Biology and Hull York Medical School, University of York, Heslington, York, United Kingdomc Departamento de Microbiologia, Imunologia e Parasitologia, Universidade Federal de São Paulo, São Paulo, SP, Brazil
a r t i c l e i n f o
Article history:Received 22 March 2010Received in revised form 29 July 2010Accepted 2 August 2010Available online 14 August 2010
Keywords:Leishmania amazonensisMetacyclogenesisOxidative stressHeat shockCalpain-likeTrypanosoma brucei
a b s t r a c t
The META cluster of Leishmania amazonensis contains both META1 and META2 genes, which are upregu-lated in metacyclic promastigotes and encode proteins containing the META domain. Previous studiesdefined META2 as a 48.0-kDa protein, which is conserved in other Leishmania species and in Trypanosomabrucei. In this work, we demonstrate that META2 protein expression is regulated during the Leishmanialife cycle but constitutive in T. brucei. META2 protein is present in the cytoplasm and flagellum ofL. amazonensis promastigotes. Leishmania META2-null replacement mutants are more sensitive to oxida-tive stress and, upon heat shock, assume rounded morphology with shortened flagella. The increasedsusceptibility of null parasites to heat shock is reversed by extra-chromosomal expression of the META2gene. Defective Leishmania promastigotes exhibit decreased ability to survive in macrophages. By contrast,META2 expression is decreased by 80% in RNAi-induced T. brucei bloodstream forms with no measurableeffect on survival or resistance to heat shock.
� 2010 Elsevier Inc. All rights reserved.
1. Introduction
Parasites of the Leishmania genus cause a disease known asleishmaniasis, with a wide spectrum of clinical manifestationsranging from single cutaneous lesions to a systemic infectionthat can be fatal if untreated. According to the World HealthOrganisation, there are 12 million people infected and about 2.3million new cases every year. The life cycle of the parasite includesan extracellular phase within the digestive tract of the invertebratevector and an intracellular stage inside the macrophages of a ver-tebrate host (Lainson and Shaw, 1987). During development inthe sandfly, promastigotes undertake a process known as metacy-clogenesis, when procyclic promastigotes, which are dividing,non-infective forms, differentiate into non-dividing infective meta-cyclic promastigotes (Sacks, 1989). In this process, a variety ofgenes show differential expression patterns, some of which encodevirulence factors.
Both META1 and META2 genes of Leishmania amazonensis areupregulated in metacyclic promastigotes and conserved in otherLeishmania species and trypanosomatids (Nourbakhsh et al.,1996; Ramos et al., 2004b; Uliana et al., 1999). Both META1 andMETA2 proteins contain the META domain (PF03724) present inseveral bacterial proteins of unknown function and linked, insome instances, to protein secretion and heat shock. While METAdomain is present in a single copy in META1 peptide sequence,the META2 protein is composed of three copies of the META do-main and a carboxyl-terminus calpain-like domain (Ramoset al., 2004b).
L. amazonensis mutants overexpressing the META1 gene showincreased virulence in vivo (Uliana et al., 1999). The structuralsimilarities between META1 and META2 genes led us to investigatewhether the latter is also involved in parasite virulence.
In this study, we describe META2 functional characterisation.The localisation of the native Leishmania protein has been deter-mined by immunofluorescence using an antiserum producedagainst recombinant META2 protein, while the role of the META2gene has been assessed by the study of META2-null mutants ofL. amazonensis. Since three conserved copies of the META2 geneare present in the Trypanosoma brucei genome, we have also lookedat their expression pattern during the parasite life cycle and usedRNA interference to modulate META2 expression in bloodstreamtrypanosomes.
0014-4894/$ - see front matter � 2010 Elsevier Inc. All rights reserved.doi:10.1016/j.exppara.2010.08.004
⇑ Corresponding author. Address: Departamento de Parasitologia, Instituto deCiências Biomédicas, Universidade de São Paulo, Av. Professor Lineu Prestes, 1374,CEP 05508-900 São Paulo, SP, Brazil. Fax: +55 11 30917417.
E-mail address: [email protected] (S.R.B. Uliana).1 Present address: Centro de Ciências Biológicas e da Saúde (CCBS), Universidade
Presbiteriana Mackenzie, São Paulo, Brazil.2 Present address: Departamento de Microbiología, Facultad de Ciencias y Filosofía,
Universidad Peruana Cayetano Heredia, Lima, Peru.
Experimental Parasitology 127 (2011) 228–237
Contents lists available at ScienceDirect
Experimental Parasitology
journal homepage: www.elsevier .com/locate /yexpr

Author's personal copy
2. Materials and methods
2.1. Parasites
Promastigotes of L. amazonensis MHOM/BR/73/M2269 weregrown at 25 �C in medium 199 (Invitrogen) supplemented with10% foetal calf serum (FCS) (Invitrogen), as previously described(Uliana et al., 1999). T. brucei bloodstream form (BSF) strain427–1313 (Alibu et al., 2005) and procyclic (PCF) strain 449(Biebinger et al., 1997) were provided by Christine Clayton, ZMBH,Universitat Heidelberg, Germany. Cells were grown in the presenceof 0.2 lg/ml (BSF) or 0.5 lg/ml phleomycin (PCF).
2.2. Expression and purification of the recombinant protein
The 1.3-kb fragment containing the L. amazonensis META2coding sequence was amplified using primers 50-cgggatccATGACCACCGCTGACGATG-30 and 50-ggaattcGCGTGCACGCCACATTCG-30,digested with BamHI and EcoRI (restriction sites underlined) andcloned into pAE vector. The expression of the recombinant proteinwith the predicted molecular mass of 48.0 kDa was obtained inEscherichia coli as described previously (Ramos et al., 2004a). Thetarget protein was affinity purified by column chromatographyusing Ni-NTA resins (QIAGEN) with urea/imidazol buffer (8 M urea,500 mM imidazol, 0.1 M NaH2PO4, 0.01 M Tris–HCl, pH 8.0). Thepurified protein was quantified using the Bio-Rad Protein Assay(Bio-Rad).
The recombinant protein was used to raise polyclonalantibodies in mice. Briefly, BALB/c mice were immunized withthree doses administered subcutaneously, at 15-day intervals, with25 lg of the recombinant META2 protein, emulsified in completeor incomplete Freund’s adjuvant.
2.3. Immunoblotting
Total protein extracts were prepared from log and stationary-phase promastigotes as described (Uliana et al., 1999). Samplesequivalent to 2 � 107 promastigotes were fractionated in SDS–PAGE and transferred to nitrocellulose membranes (Hybond-ECL,Amersham-Pharmacia). Membranes were incubated in TBST-5%milk (10 mM Tris, pH 8.0, 150 mM NaCl, 0.1% Tween 20, 5% nonfatpowdered milk) for 1 h followed by incubation with anti-META2serum diluted 1:200 in TBST-3% milk for 1 h. After three washesof 10 min each with TBST, the membranes were incubated withanti-mouse polyvalent IgG conjugated to alkaline phosphatase(Sigma), diluted in TBST 1:3000. Membranes were then washedthree times with TBST and developed with AP buffer (100 mMTris–HCl, pH 9.5, 100 mM NaCl, 1.0 mM MgCl2) containing350 lg/ml nitroblue tetrazolium and 175 lg/ml 5-bromo-4-chloro-3-indolyl phosphate. All the procedures were done at roomtemperature.
2.4. Indirect immunofluorescence assays
Promastigotes were washed in PBS (10 mM Na2PO4, 1.76 mMKH2PO4 (pH 7.4), 137 mM NaCl and 2.7 mM KCl), fixed in 4% para-formaldehyde for 30 min, resuspended in PBS and applied ontoglass slides. BALB/c resident peritoneal macrophages were platedin round glass coverslips inside the wells of 24-well culture dishes,at a density of 4 � 105 cells per coverslip. Cells were infected withstationary-phase promastigotes in a ratio of 7 parasites:1 cell at33 �C for 3 h. Cultures were washed, kept at 33 �C in 5% CO2 for48 h and fixed in methanol. Fixed promastigotes or infected macro-phages were preincubated at room temperature for 45 min in PBScontaining 3% bovine serum albumin and 0.1% Triton X-100. The
slides were then incubated with anti-META2 serum diluted inthe same solution (1:50) for 1 h at room temperature and washedfour times in PBS containing 0.1% Triton X-100. Anti-mousepolyvalent IgG conjugated to Alexa Fluor 488 (Molecular Probes),diluted in blocking solution in a ratio of 1:500, was then appliedonto the slides for 30 min at room temperature. After two washesin PBS containing Triton X-100, followed by two washes in PBSalone, the cells were stained for DNA content with DAPI 2.5lg/ml for 5 min and washed again with PBS. Finally, a 2.5% sodiumazide solution was applied and the cover slides were sealed.
The cells were visualised using a Zeiss Axiovert or a Bio-Rad1024-UV confocal system (Bio-Rad) attached to a Zeiss Axiovert100 microscope, using a 100� immersion objective N.A. 1.2 plan-apochromatic lens. The images acquired were merged using ImageJsoftware (http://www.rsb.info.nih.gov/ij/) and represent a uniqueoptical axis.
2.5. Leishmania plasmids and constructs
The plasmid pNEO (Uliana et al., 1999), obtained by cloning theneomycin phosphotransferase gene (NEO) in pBlueScript II SK (�)(Stratagene), was used in the constructs for the knock-out of theMETA2 gene. Restriction fragments containing META2 50-UTR(0.8 kb) and 30-UTR (2.2 kb) were cloned, respectively, betweenHindIII/HincII and XbaI sites of pNEO.
To complement the deficient line, a 2.5-kb fragment containingthe META2 gene flanked by its UTR regions was amplified using thepair of primers 50-ccccccgggGCGTATAGGGATGAGAGTGG-30 and50-ccccccgggAAGCGCGCCTAAGAAACAGG-30 and cloned into theXmaI site of pX63HYG vector (Cruz et al., 1991). Cloning was con-firmed by restriction digest and sequencing.
2.6. DNA purification and analysis
Leishmania genomic DNA was purified as described (Medina-Acosta and Cross, 1993). DNA restriction fragments were separatedin agarose gels, analysed using an Eagle Eye (Stratagene) and trans-ferred to nylon membranes (Hybond-XL, Amersham-Pharmacia).For pulse field gel electrophoresis (PFGE) analysis, chromosomalDNA was prepared in low-melting temperature agarose plugs(Coburn et al., 1991). Chromosomes were separated by PFGE ranon a contour-clamped homogeneous electric field apparatus (CHEFDR II, Bio-Rad). Gels were run at 14 �C, at 6.0 V/cm for 30 h with a24–240-s pulse ramp time. The electrophoresed DNAs were trans-ferred to nylon membranes (Hybond-XL, Amersham-Pharmacia).Hybridization experiments with radiolabeled probes were per-formed at 42 �C, using 50% formamide buffer, and washed twicein 0.1� SSC, 0.1% SDS solution at 60 �C. Probe M2 is a 1.0-kbfragment from the META2 gene and NEO probe contains the entirecoding sequence of this selection marker gene.
2.7. Transfection and selection of mutants
XhoI/NotI digestions of clone pNEO/50/30M2 were separated inagarose gels and purified by electroelution, while circular DNAfrom pX63/META2 was precipitated with ethanol. DNA was resus-pended in sterile water and quantified spectrophotometrically.
DNA electroporation in L. amazonensis was done as described(Coburn et al., 1991) and the cells were then plated in medium199/agar 1% with either hygromycin (32 lg/ml) or G418 (20lg/ml). After 2 weeks, colonies were picked, expanded in liquidmedia and drug concentrations were gradually increased to150 lg/ml of hygromycin or 160 lg/ml of G418 (knock-out clones).Cultures grown on 160 lg/ml of G418 for 8–10 passages were pla-ted and 6 clones were picked, expanded and analysed.
C.S. Ramos et al. / Experimental Parasitology 127 (2011) 228–237 229

Author's personal copy
2.8. Determination of the number of metacyclic promastigotes usingmonoclonal antibody 3A1
The number of metacyclic promastigotes in stationary phasepromastigote cultures (6 days) of wild-type and mutant parasiteswas determined by negative selection with the 3A1 monoclonalantibody (mAb) (kindly provided by Dr. Elvira M. Saraiva, Institutode Microbiologia, UFRJ, Brazil) as described (Pinto-da-Silva et al.,2005). Briefly, promastigotes were washed in PBS and incubatedwith mAb 3A1-La ascites (1:4000) at room temperature (RT) for30 min. The parasite suspension was then centrifuged at 40g for5 min, and the non-agglutinated promastigotes were collectedfrom the supernatant. The percentage of metacyclics purified fromwild-type and mutant parasites was determined by counting.
2.9. Sensitivity to oxidative stress and heat shock
Sensitivity to oxidative stress was tested by incubating para-sites in increasing concentrations of hydrogen peroxide for 3, 6or 24 h. Cell viability was evaluated by MTT [3-(4,5-dimethylthia-zol-2-yl)-2,5-diphenyl tetrazolium bromide (Sigma–Aldrich)] test(Mosmann, 1983). In brief, cells were incubated in (MOPS)-buffered saline containing MTT 0.83 mg/ml for 2 h. MTT cleavagewas assessed by using a multiwell scanning spectrophotometer(Labsystems; Multiskan EX) with a reference wavelength of690 nm and a test wavelength of 595 nm. Assays were performedin triplicate and the results are expressed as the percentage ofparasite survival compared with controls incubated in (MOPS)-buf-fer alone. Cell morphology was analysed by staining methanol-fixed parasites with the Instant-Prov kit (Newprov, Pinhais, Brazil)and examined by optical microscopy.
Promastigotes in culture media were incubated at 25 or 33 �Cfor 3, 6, 9, 12 or 24 h. In some experiments, after 24 h of heat shock,cultures were returned to incubation at 25 �C for additional 24 h.At various time points, aliquots were taken and deposited onmicroscope slides, fixed and stained. In parallel, cultures were eval-uated for viability by counting, by the MTT test (as describedabove) or by trypan blue exclusion.
Sensitivity to oxidative stress and heat shock experiments wereperformed with transgenic parasites cultured in the presence ofthe selective drug and with parasites kept in medium without drugfor 72–96 h.
2.10. Macrophage infections
Resident peritoneal macrophages were obtained from BALB/cmice by peritoneal lavage with RPMI-1640 medium. The collectedcell suspension was centrifuged at 300g for 10 min at 4 �C, the pel-let was resuspended in RPMI-1640 supplemented with 10% FCSand 50 lg/ml gentamicin and plated in round glass coverslips in-side the wells of 24-well culture dishes, at a density of 4 � 105 cellsper coverslip. After 2 h of incubation at 37 �C in an atmosphere of5% CO2, L. amazonensis stationary-phase promastigotes (in a ratioof 7 parasites:1 macrophage) were added to the wells, and the cul-tures were incubated at 33 �C in a 5% CO2 atmosphere. After 2 h,the parasites were removed by extensive washing with RPMImedium without FCS, and the infected cultures were incubatedfor further 16 or 48 h. The monolayers were washed, fixed andstained with the Instant-Prov kit (Newprov, Pinhais, Brazil). Thepercentage of infected macrophages and the number of intracellu-lar amastigotes were assessed by light microscopy observation,counting at least 100 cells in triplicate coverslips. Infections withtransgenic parasites were performed with parasites cultured with-out G418 and hygromycin for 72 h as well as with parasites grownin the presence of the selective drug.
2.11. Mice infections
Female BALB/c mice (4–5 weeks old) were inoculated subcuta-neously with 1 � 106 stationary-phase parasites at the left hindfootpad. Groups of five animals were infected with L. amazonensiswild-type or transfected lines. Footpad thickness was evaluatedweekly and the lesion size was calculated as the differencebetween the thickness of infected and contralateral uninfectedfootpads. Results shown are the mean and standard deviation foreach group in one experiment, representative of two independentexperiments.
2.12. Sensitivity to drugs
Cell viability after incubation with novobiocin was evaluatedin vitro by cultivating promastigotes (5 � 106) in 199 medium con-taining increasing concentrations of the drug for 24 h. Quantifica-tion of viable cells was assessed by measuring the cleavage ofMTT as described above.
The sensitivity to meglumine antimoniate was assayed by incu-bating promastigote cultures in the presence of increasing concen-trations of the drug for 96 h. Cell numbers were counted using aNeubauer haemocytometer chamber. The 50% inhibitory concen-trations (IC50) were determined from sigmoidal regression of theconcentration–response curves using Scientific Graphing andAnalysis Software ORIGIN 7.5.
2.13. Cloning of RNAi fragment, transfection and analysis of T. bruceimutants
A META2 450-bp fragment was amplified from T. brucei genomicDNA with the oligonucleotides TbM2523/542 XhoI/XbaI50-ccgctcgagtctagaCGATGTGCGAGGTGGATGG-30 and TbM2 973/954 PstI/HindIII 50-cccaagcttctgcagCAAGAGTCAGCACCGTGTCG-30.The amplified fragment was cloned into XbaI and PstI sites ofpBluescript. A stuffer fragment was obtained from the PstI andEcoRI digestion of vector pHD1188 (Guerra-Giraldez et al., 2002)and transferred to the META2 construct. A second inverted copyof META2 fragment was cloned into the HindIII and XhoI sites.The cassette containing the two inverted copies of the META2 frag-ment plus the stuffer was isolated through NotI/XhoI digestions ofthe plasmid and this filled-in fragment was cloned into the HpaIsite of pHD677, generating TbMETA2pHD677.
Mid-log phase T. brucei BSF strain 427–1313 parasites wereelectroporated with 10 lg of EcoRV-digested construct TbME-TA2pHD677 using methods described previously (Price et al.,2005). Expression of META2-specific dsRNA was induced in stablecell lines by incubating parasites in 100 ng/ml tetracycline.
Expression of META2 was monitored by quantitative PCR, usingSYBR Green Mastermix (Applied Biosystems). Total RNA was ex-tracted from parasites using TRIzol Reagent (Invitrogen), treatedwith DNase I (Ambion) and reverse-transcribed using OmniscriptRT (Qiagen). An 82 bp fragment of META2 (residues 987–1068)was amplified using SYBR Green Mastermix (Applied Biosystems)on a Prism7000 (Applied Biosystems) and compared to levels ofthe constitutive control, a-tubulin.
The effects of META2 on heat shock were investigated bygrowing cells in the absence or presence of tetracycline for 48 h,then seeding new cultures at 5 � 105 ml�1 and incubating at42 �C for up to 6 h. Motile cell numbers were monitored using ahaemocytometer.
2.14. Statistical analysis
Results were compared for statistical significance usingtwo-tailed Student’s t-test for paired samples or two-way ANOVA
230 C.S. Ramos et al. / Experimental Parasitology 127 (2011) 228–237
Author's personal copy
followed by Bonferroni’s multiple comparison test. A result wasconsidered significant at P < 0.05.
2.15. Ethics of experimentation
All the experiments involving animals were approved by theEthics Committee for Animal Experimentation of the Instituto deCiências Biomédicas, University of São Paulo.
3. Results
3.1. Expression and sub-cellular localisation of L. amazonensis META2protein
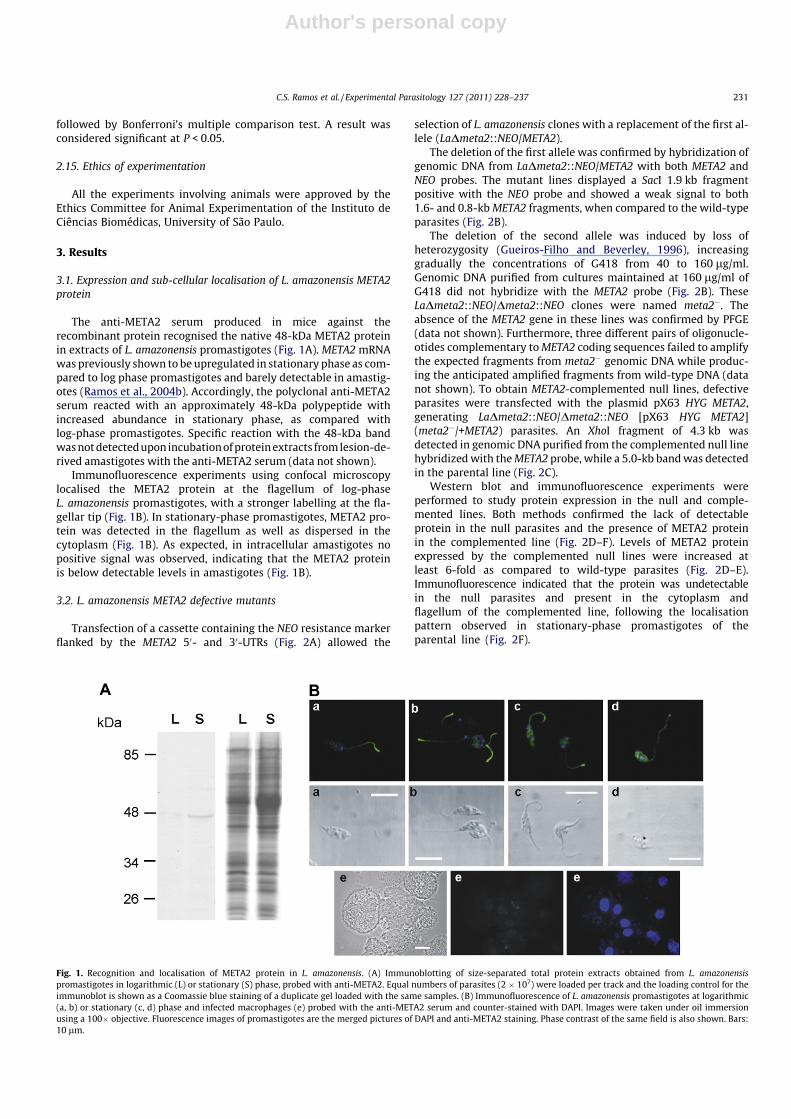
The anti-META2 serum produced in mice against therecombinant protein recognised the native 48-kDa META2 proteinin extracts of L. amazonensis promastigotes (Fig. 1A). META2 mRNAwas previously shown to be upregulated in stationary phase as com-pared to log phase promastigotes and barely detectable in amastig-otes (Ramos et al., 2004b). Accordingly, the polyclonal anti-META2serum reacted with an approximately 48-kDa polypeptide withincreased abundance in stationary phase, as compared withlog-phase promastigotes. Specific reaction with the 48-kDa bandwas not detected upon incubation of protein extracts from lesion-de-rived amastigotes with the anti-META2 serum (data not shown).
Immunofluorescence experiments using confocal microscopylocalised the META2 protein at the flagellum of log-phaseL. amazonensis promastigotes, with a stronger labelling at the fla-gellar tip (Fig. 1B). In stationary-phase promastigotes, META2 pro-tein was detected in the flagellum as well as dispersed in thecytoplasm (Fig. 1B). As expected, in intracellular amastigotes nopositive signal was observed, indicating that the META2 proteinis below detectable levels in amastigotes (Fig. 1B).
3.2. L. amazonensis META2 defective mutants
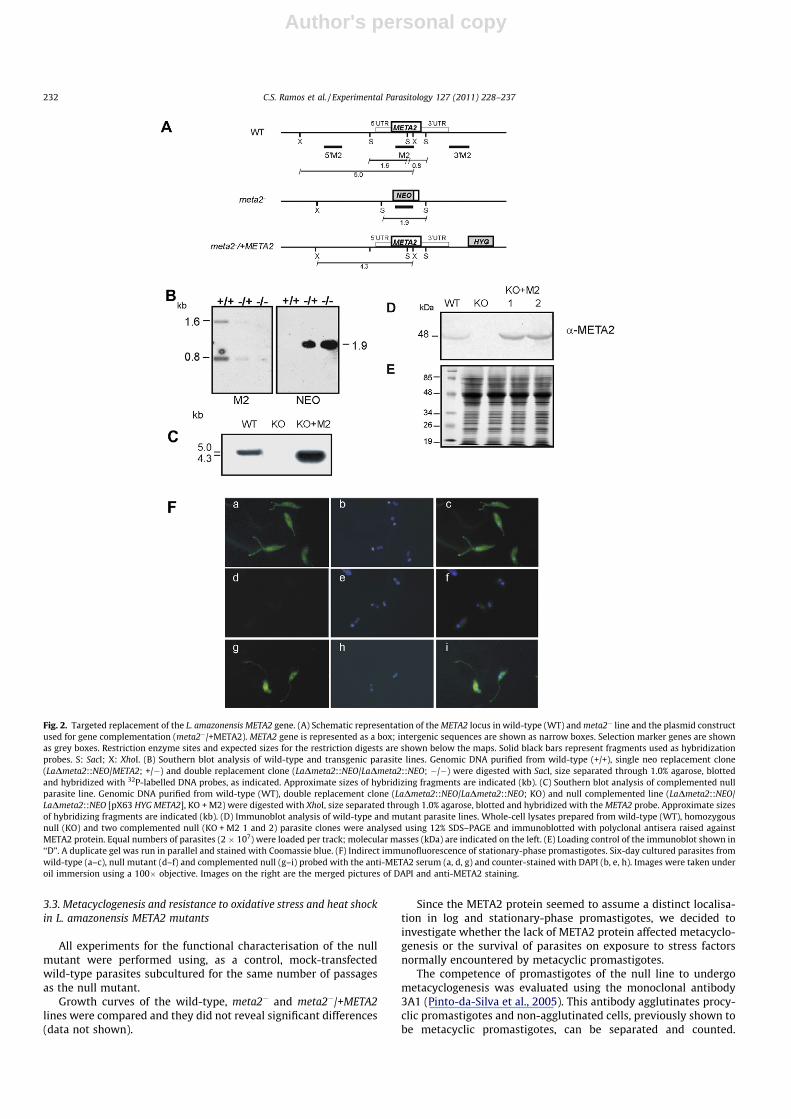
Transfection of a cassette containing the NEO resistance markerflanked by the META2 50- and 30-UTRs (Fig. 2A) allowed the
selection of L. amazonensis clones with a replacement of the first al-lele (LaDmeta2::NEO/META2).
The deletion of the first allele was confirmed by hybridization ofgenomic DNA from LaDmeta2::NEO/META2 with both META2 andNEO probes. The mutant lines displayed a SacI 1.9 kb fragmentpositive with the NEO probe and showed a weak signal to both1.6- and 0.8-kb META2 fragments, when compared to the wild-typeparasites (Fig. 2B).
The deletion of the second allele was induced by loss ofheterozygosity (Gueiros-Filho and Beverley, 1996), increasinggradually the concentrations of G418 from 40 to 160 lg/ml.Genomic DNA purified from cultures maintained at 160 lg/ml ofG418 did not hybridize with the META2 probe (Fig. 2B). TheseLaDmeta2::NEO/Dmeta2::NEO clones were named meta2�. Theabsence of the META2 gene in these lines was confirmed by PFGE(data not shown). Furthermore, three different pairs of oligonucle-otides complementary to META2 coding sequences failed to amplifythe expected fragments from meta2� genomic DNA while produc-ing the anticipated amplified fragments from wild-type DNA (datanot shown). To obtain META2-complemented null lines, defectiveparasites were transfected with the plasmid pX63 HYG META2,generating LaDmeta2::NEO/Dmeta2::NEO [pX63 HYG META2](meta2�/+META2) parasites. An XhoI fragment of 4.3 kb wasdetected in genomic DNA purified from the complemented null linehybridized with the META2 probe, while a 5.0-kb band was detectedin the parental line (Fig. 2C).
Western blot and immunofluorescence experiments wereperformed to study protein expression in the null and comple-mented lines. Both methods confirmed the lack of detectableprotein in the null parasites and the presence of META2 proteinin the complemented line (Fig. 2D–F). Levels of META2 proteinexpressed by the complemented null lines were increased atleast 6-fold as compared to wild-type parasites (Fig. 2D–E).Immunofluorescence indicated that the protein was undetectablein the null parasites and present in the cytoplasm andflagellum of the complemented line, following the localisationpattern observed in stationary-phase promastigotes of theparental line (Fig. 2F).
Fig. 1. Recognition and localisation of META2 protein in L. amazonensis. (A) Immunoblotting of size-separated total protein extracts obtained from L. amazonensispromastigotes in logarithmic (L) or stationary (S) phase, probed with anti-META2. Equal numbers of parasites (2 � 107) were loaded per track and the loading control for theimmunoblot is shown as a Coomassie blue staining of a duplicate gel loaded with the same samples. (B) Immunofluorescence of L. amazonensis promastigotes at logarithmic(a, b) or stationary (c, d) phase and infected macrophages (e) probed with the anti-META2 serum and counter-stained with DAPI. Images were taken under oil immersionusing a 100� objective. Fluorescence images of promastigotes are the merged pictures of DAPI and anti-META2 staining. Phase contrast of the same field is also shown. Bars:10 lm.
C.S. Ramos et al. / Experimental Parasitology 127 (2011) 228–237 231
Author's personal copy
3.3. Metacyclogenesis and resistance to oxidative stress and heat shockin L. amazonensis META2 mutants
All experiments for the functional characterisation of the nullmutant were performed using, as a control, mock-transfectedwild-type parasites subcultured for the same number of passagesas the null mutant.
Growth curves of the wild-type, meta2� and meta2�/+META2lines were compared and they did not reveal significant differences(data not shown).
Since the META2 protein seemed to assume a distinct localisa-tion in log and stationary-phase promastigotes, we decided toinvestigate whether the lack of META2 protein affected metacyclo-genesis or the survival of parasites on exposure to stress factorsnormally encountered by metacyclic promastigotes.
The competence of promastigotes of the null line to undergometacyclogenesis was evaluated using the monoclonal antibody3A1 (Pinto-da-Silva et al., 2005). This antibody agglutinates procy-clic promastigotes and non-agglutinated cells, previously shown tobe metacyclic promastigotes, can be separated and counted.
Fig. 2. Targeted replacement of the L. amazonensis META2 gene. (A) Schematic representation of the META2 locus in wild-type (WT) and meta2� line and the plasmid constructused for gene complementation (meta2�/+META2). META2 gene is represented as a box; intergenic sequences are shown as narrow boxes. Selection marker genes are shownas grey boxes. Restriction enzyme sites and expected sizes for the restriction digests are shown below the maps. Solid black bars represent fragments used as hybridizationprobes. S: SacI; X: XhoI. (B) Southern blot analysis of wild-type and transgenic parasite lines. Genomic DNA purified from wild-type (+/+), single neo replacement clone(LaDmeta2::NEO/META2; +/�) and double replacement clone (LaDmeta2::NEO/LaDmeta2::NEO; �/�) were digested with SacI, size separated through 1.0% agarose, blottedand hybridized with 32P-labelled DNA probes, as indicated. Approximate sizes of hybridizing fragments are indicated (kb). (C) Southern blot analysis of complemented nullparasite line. Genomic DNA purified from wild-type (WT), double replacement clone (LaDmeta2::NEO/LaDmeta2::NEO; KO) and null complemented line (LaDmeta2::NEO/LaDmeta2::NEO [pX63 HYG META2], KO + M2) were digested with XhoI, size separated through 1.0% agarose, blotted and hybridized with the META2 probe. Approximate sizesof hybridizing fragments are indicated (kb). (D) Immunoblot analysis of wild-type and mutant parasite lines. Whole-cell lysates prepared from wild-type (WT), homozygousnull (KO) and two complemented null (KO + M2 1 and 2) parasite clones were analysed using 12% SDS–PAGE and immunoblotted with polyclonal antisera raised againstMETA2 protein. Equal numbers of parasites (2 � 107) were loaded per track; molecular masses (kDa) are indicated on the left. (E) Loading control of the immunoblot shown in‘‘D”. A duplicate gel was run in parallel and stained with Coomassie blue. (F) Indirect immunofluorescence of stationary-phase promastigotes. Six-day cultured parasites fromwild-type (a–c), null mutant (d–f) and complemented null (g–i) probed with the anti-META2 serum (a, d, g) and counter-stained with DAPI (b, e, h). Images were taken underoil immersion using a 100� objective. Images on the right are the merged pictures of DAPI and anti-META2 staining.
232 C.S. Ramos et al. / Experimental Parasitology 127 (2011) 228–237

Author's personal copy
In stationary-phase cultures, 25.0 ± 5.3% of wild-type parasiteswere not agglutinated by the antibody. Significant differences werenot observed in the number of metacyclics in cultures of meta2�
(18.1 ± 4.4%) or meta2�/+META2 (18.7 ± 1.8), indicating that meta-cyclogenesis or, at least the expression of surface molecules typicalof metacyclics, was not prevented by the lack of META2 protein.
Another approach taken to functionally evaluate the stationary-phase population was to test the response of the META2-null linesto stress factors such as oxidative conditions and temperature.Their ability to survive oxidative stress was investigated byincubating parasites with increasing concentrations of hydrogenperoxide for 3 h. The IC50 for H2O2 in wild-type parasites was810.40 ± 31.30 lM, while the IC50 calculated for the meta2� was312.16 ± 75.98 lM. The sensitivity of meta2�/+META2 parasitesremained similar to that of the null parasites, with IC50 of290.01 ± 3.80 lM. META2-null parasites were therefore more sen-sitive to oxidative shock than wild-type lines (P = 0.049) and thisphenotype was not reversed by adding back the META2 gene. Toascertain that this increased sensitivity was not related to the drugpressure on mutant parasites, these were also tested after 72 h inculture in the absence of the selective drug (G418, or G418 plushygromycin). No differences were observed in the behaviour oftransgenic parasites irrespective of the presence or absence ofthe drug (data not shown). Morphological analysis of parasitessubjected to oxidative shock did not reveal differences betweenwild-type and transgenic parasites (data not shown).
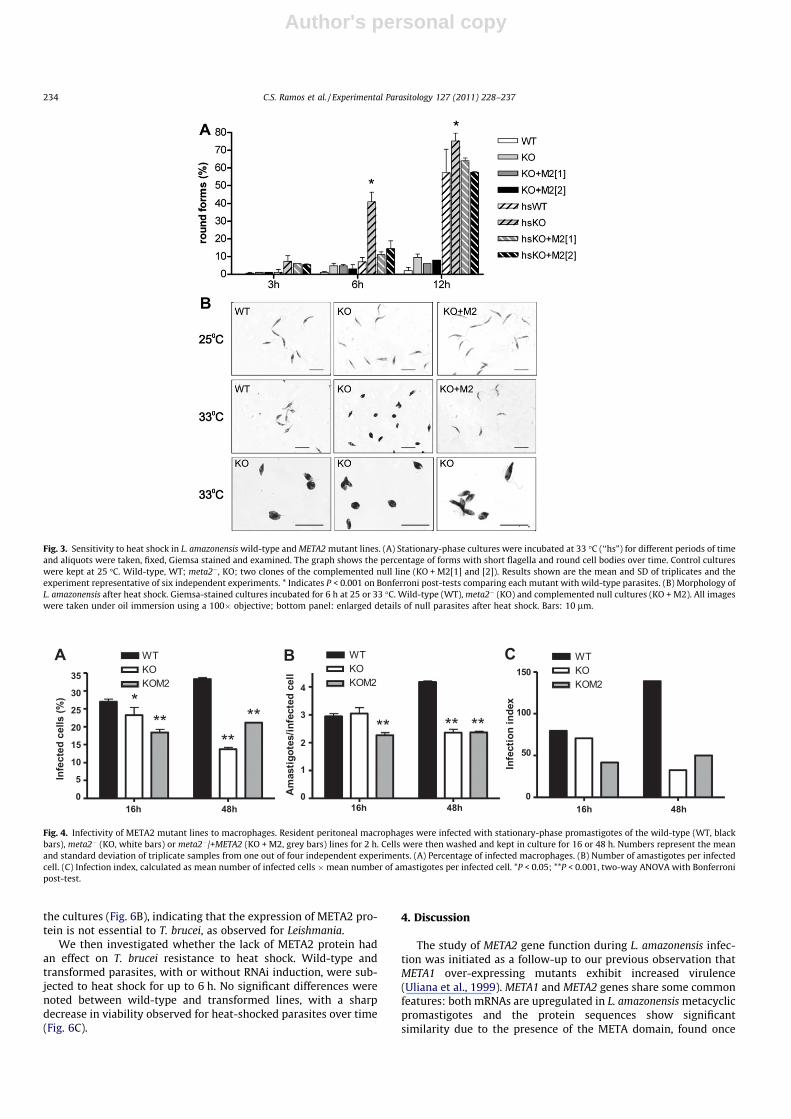
Viability of wild-type and meta2� parasites was also testedupon heat shock for variable periods of time. As expected, growthof parasites at 33 �C was impaired as compared to cultures kept at25 �C, but there was no difference in the growth rate betweenwild-type, defective or complemented null parasites in these con-ditions (Table 1). During heat shock we also did not observe anyimportant changes in viability, evaluated by trypan blue exclusion(Table 1) or MTT (data not shown). This was confirmed by incubat-ing cultures that had been subjected to heat shock for 24 h for arecovery period of 24 h at 25 �C; the number of cells in all linestested was equivalent by the end of this period (data not shown).Although no viability differences were noted, a clear morphologicalchange was observed in null parasites subjected to various incuba-tion times at 33 �C. The null line was morphologically more sensi-tive to temperature than the wild-type parasites. After 3 h ofincubation at 33 �C, a larger proportion of parasites in theMETA2-null line exhibited shortening of the flagellum androunding of the cell body (Fig. 3A and B). By 6 h of heat shock,the difference in the number of ‘‘short/round” cells between theknock out and wild-type lines was maximum (P < 0.001). Theseforms did not represent dying parasites, since they were not
stained by trypan blue and were able to divide (Fig. 3B, KO33 �C). These observations were confirmed with clones obtainedfrom two independent transfection experiments and with trans-genic parasites kept in the absence of drug for 72 h. The same mor-phological changes were also detected in the wild-type parasitesbut only after prolonged heat shock (data not shown). So, parasiteslacking META2 protein developed an increased sensitivity to tem-perature, translated into morphological changes without loss ofviability. The complementation of these clones with the plasmidcontaining the META2 gene restored the wild-type phenotype(Fig. 3A and B).
3.4. Infectivity of L. amazonensis META2 mutants in vitro and in vivo
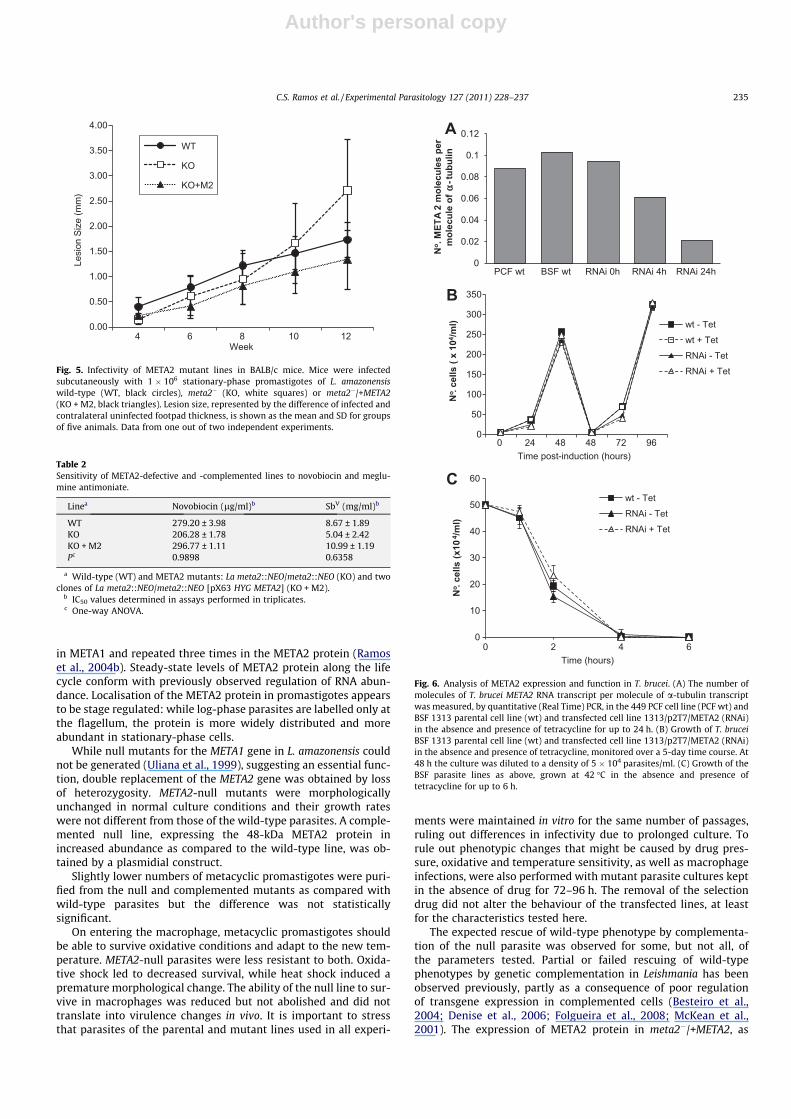
Having observed that META2-null parasites were more sensi-tive to oxidative and heat stress, we then looked at their abilityto invade macrophages and multiply intracellularly, as expressedby number of infected cells, mean number of amastigotes per in-fected cell and infection indexes (Fig. 4). At early time points, thepercentage of infected cells was reduced for both the null andcomplemented lines. Compared with the wild-type infections,intra-macrophagic multiplication and survival of the null line weresignificantly compromised as evidenced by the number of infectedcells (P < 0.001) as well as by the number of amastigotes per cell(P < 0.001) after 48 h of infection (Fig. 4). The number of cells in-fected by the wild-type parasites increased by 24% after 48 h whilethe percentage of infection by the null line was reduced by 59%(Fig. 4A). Survival of meta2�/+META2 parasites at 48 h was partiallyrestored and the percentage of infected cells rose by 15% from 16to 48 h. The same behaviour was apparent in the average numberof amastigotes per infected cell and in the overall infection index(Fig. 4B and C).
In vivo infections were carried out in susceptible BALB/c mice byinoculation of stationary-phase promastigotes of wild-type, nulland complemented lines. No significant differences were observedin the pattern of lesion development showing that META2, null andcomplemented parasites infect animals at the same rate and to thesame extent as their counterpart wild-type controls (Fig. 5).
3.5. Sensitivity to novobiocin and antimonials
Proteins containing the META domain, which is repeated threetimes in the META2 protein, have been associated in bacteria withresistance to novobiocin (Lilic et al., 2003). On the other hand, thecalpain-like C-terminus of META2 protein shows similarity to theLeishmania major small kinetoplastid calpain-related protein(LmjF14.0850, SKCRP14.1), which has been linked to resistanceagainst antimonials (Vergnes et al., 2007). The sensitivity of wild-type, META2-null and complemented null parasites was reducedsimilarly in response to stepwise increase in the concentration ofboth drugs without significant changes in the IC50 of these para-sites to either novobiocin or meglumine antimoniate (Table 2).
3.6. Characterisation of META2 expression in T. brucei
To investigate whether META2 expression influences similarprocesses in T. brucei, we used real time PCR to determine mRNAabundance in procyclic and bloodstream parasite forms. The levelsof META2 mRNA were comparable in both life cycle stages (Fig. 6A),revealing that the stage-specific regulation found in L. amazonensisis not conserved in T. brucei, at least at the RNA level.
The functional role of META2 protein in this organism wasprobed by depleting META2 RNA by RNAi. Upon induction of RNAifor 24 h, META2 mRNA levels were reduced by 80% (Fig. 6A). Thegrowth kinetics of transformed BSF parasites were indistinguish-able from the wild-type, irrespective of tetracycline addition to
Table 1Growth and viability of wild-type and mutant lines upon heat shock.
Linea Temperature(�C)
0 h 3 h 6 h 12 h
WT 25 5.00b 4.48 (98.35)c 5.33 (98.61) 6.28 (98.82)KO 5.00 4.30 (97.73) 5.85 (98.32) 7.45 (99.00)KO + M2(1) 5.00 5.93 (98.75) 6.10 (97.99) 9.58 (98.97)KO + M2(2) 5.00 4.78 (98.45) 5.65 (98.26) 6.48 (98.48)
WT 33 5.00 4.90 (98.99) 5.60 (96.55) 3.50 (96.71)KO 5.00 6.15 (97.62) 5.30 (97.25) 3.05 (96.97)KO + M2(1) 5.00 5.33 (98.61) 7.35 (98.66) 5.70 (98.30)KO + M2(2) 5.00 5.55 (99.11) 6.60 (98.51) 3.20 (98.51)
a Wild-type (WT) and META2 mutants: La meta2::NEO/meta2::NEO (KO) and twoclones of La meta2::NEO/meta2::NEO [pX63 HYG META2] (KO + M2).
b Promastigote cultures were seeded with 5.00 � 106 parasites/ml and incubatedat 25 or 33 �C for the indicated times.
c After 3, 6 and 12 h triplicate aliquots were stained with trypan blue andcounted. Shown are: mean of total number of cells (�106) and percentage of viablecells (in parenthesis). The standard deviations did not exceed 10% in any sample.
C.S. Ramos et al. / Experimental Parasitology 127 (2011) 228–237 233
Author's personal copy
the cultures (Fig. 6B), indicating that the expression of META2 pro-tein is not essential to T. brucei, as observed for Leishmania.
We then investigated whether the lack of META2 protein hadan effect on T. brucei resistance to heat shock. Wild-type andtransformed parasites, with or without RNAi induction, were sub-jected to heat shock for up to 6 h. No significant differences werenoted between wild-type and transformed lines, with a sharpdecrease in viability observed for heat-shocked parasites over time(Fig. 6C).
4. Discussion
The study of META2 gene function during L. amazonensis infec-tion was initiated as a follow-up to our previous observation thatMETA1 over-expressing mutants exhibit increased virulence(Uliana et al., 1999). META1 and META2 genes share some commonfeatures: both mRNAs are upregulated in L. amazonensis metacyclicpromastigotes and the protein sequences show significantsimilarity due to the presence of the META domain, found once
Fig. 3. Sensitivity to heat shock in L. amazonensis wild-type and META2 mutant lines. (A) Stationary-phase cultures were incubated at 33 �C (‘‘hs”) for different periods of timeand aliquots were taken, fixed, Giemsa stained and examined. The graph shows the percentage of forms with short flagella and round cell bodies over time. Control cultureswere kept at 25 �C. Wild-type, WT; meta2�, KO; two clones of the complemented null line (KO + M2[1] and [2]). Results shown are the mean and SD of triplicates and theexperiment representative of six independent experiments. * Indicates P < 0.001 on Bonferroni post-tests comparing each mutant with wild-type parasites. (B) Morphology ofL. amazonensis after heat shock. Giemsa-stained cultures incubated for 6 h at 25 or 33 �C. Wild-type (WT), meta2� (KO) and complemented null cultures (KO + M2). All imageswere taken under oil immersion using a 100� objective; bottom panel: enlarged details of null parasites after heat shock. Bars: 10 lm.
16h 48h0
5
10
15
20
25
30
35
WTKOKOM2
Infe
cted
cel
ls (%
)
16h 48h0
1
2
3
4
WTKOKOM2
Am
astig
otes
/infe
cted
cel
l
16h 48h0
50
100
150WTKOKOM2
Infe
ctio
n in
dex
A B C
Fig. 4. Infectivity of META2 mutant lines to macrophages. Resident peritoneal macrophages were infected with stationary-phase promastigotes of the wild-type (WT, blackbars), meta2� (KO, white bars) or meta2�/+META2 (KO + M2, grey bars) lines for 2 h. Cells were then washed and kept in culture for 16 or 48 h. Numbers represent the meanand standard deviation of triplicate samples from one out of four independent experiments. (A) Percentage of infected macrophages. (B) Number of amastigotes per infectedcell. (C) Infection index, calculated as mean number of infected cells �mean number of amastigotes per infected cell. *P < 0.05; **P < 0.001, two-way ANOVA with Bonferronipost-test.
234 C.S. Ramos et al. / Experimental Parasitology 127 (2011) 228–237
Author's personal copy
in META1 and repeated three times in the META2 protein (Ramoset al., 2004b). Steady-state levels of META2 protein along the lifecycle conform with previously observed regulation of RNA abun-dance. Localisation of the META2 protein in promastigotes appearsto be stage regulated: while log-phase parasites are labelled only atthe flagellum, the protein is more widely distributed and moreabundant in stationary-phase cells.
While null mutants for the META1 gene in L. amazonensis couldnot be generated (Uliana et al., 1999), suggesting an essential func-tion, double replacement of the META2 gene was obtained by lossof heterozygosity. META2-null mutants were morphologicallyunchanged in normal culture conditions and their growth rateswere not different from those of the wild-type parasites. A comple-mented null line, expressing the 48-kDa META2 protein inincreased abundance as compared to the wild-type line, was ob-tained by a plasmidial construct.
Slightly lower numbers of metacyclic promastigotes were puri-fied from the null and complemented mutants as compared withwild-type parasites but the difference was not statisticallysignificant.
On entering the macrophage, metacyclic promastigotes shouldbe able to survive oxidative conditions and adapt to the new tem-perature. META2-null parasites were less resistant to both. Oxida-tive shock led to decreased survival, while heat shock induced apremature morphological change. The ability of the null line to sur-vive in macrophages was reduced but not abolished and did nottranslate into virulence changes in vivo. It is important to stressthat parasites of the parental and mutant lines used in all experi-
ments were maintained in vitro for the same number of passages,ruling out differences in infectivity due to prolonged culture. Torule out phenotypic changes that might be caused by drug pres-sure, oxidative and temperature sensitivity, as well as macrophageinfections, were also performed with mutant parasite cultures keptin the absence of drug for 72–96 h. The removal of the selectiondrug did not alter the behaviour of the transfected lines, at leastfor the characteristics tested here.
The expected rescue of wild-type phenotype by complementa-tion of the null parasite was observed for some, but not all, ofthe parameters tested. Partial or failed rescuing of wild-typephenotypes by genetic complementation in Leishmania has beenobserved previously, partly as a consequence of poor regulationof transgene expression in complemented cells (Besteiro et al.,2004; Denise et al., 2006; Folgueira et al., 2008; McKean et al.,2001). The expression of META2 protein in meta2�/+META2, as
0.00
0.50
1.00
1.50
2.00
2.50
3.00
3.50
4.00
4 6 8 10 12
Lesi
on S
ize
(mm
)
Week
WT
KO
KO+M2
Fig. 5. Infectivity of META2 mutant lines in BALB/c mice. Mice were infectedsubcutaneously with 1 � 106 stationary-phase promastigotes of L. amazonensiswild-type (WT, black circles), meta2� (KO, white squares) or meta2�/+META2(KO + M2, black triangles). Lesion size, represented by the difference of infected andcontralateral uninfected footpad thickness, is shown as the mean and SD for groupsof five animals. Data from one out of two independent experiments.
Table 2Sensitivity of META2-defective and -complemented lines to novobiocin and meglu-mine antimoniate.
Linea Novobiocin (lg/ml)b SbV (mg/ml)b
WT 279.20 ± 3.98 8.67 ± 1.89KO 206.28 ± 1.78 5.04 ± 2.42KO + M2 296.77 ± 1.11 10.99 ± 1.19Pc 0.9898 0.6358
a Wild-type (WT) and META2 mutants: La meta2::NEO/meta2::NEO (KO) and twoclones of La meta2::NEO/meta2::NEO [pX63 HYG META2] (KO + M2).
b IC50 values determined in assays performed in triplicates.c One-way ANOVA.
0
0.02
0.04
0.06
0.08
0.1
0.12
PCF wt BSF wt RNAi 0h RNAi 4h RNAi 24h
No .
MET
A 2
mol
ecul
es p
er
mol
ecul
e of
- tub
ulin
0
50
100
150
200
250
300
350
0 24 48 48 72 96N
o . ce
lls (
x 10
4 /ml)
Time post-induction (hours)
wt - Tet
wt + Tet
RNAi - Tet
RNAi + Tet
0
10
20
30
40
50
60
0 2 4 6
No .
cells
(x10
4 /ml)
Time (hours)
wt - Tet
RNAi - Tet
RNAi + Tet
A
B
C
Fig. 6. Analysis of META2 expression and function in T. brucei. (A) The number ofmolecules of T. brucei META2 RNA transcript per molecule of a-tubulin transcriptwas measured, by quantitative (Real Time) PCR, in the 449 PCF cell line (PCF wt) andBSF 1313 parental cell line (wt) and transfected cell line 1313/p2T7/META2 (RNAi)in the absence and presence of tetracycline for up to 24 h. (B) Growth of T. bruceiBSF 1313 parental cell line (wt) and transfected cell line 1313/p2T7/META2 (RNAi)in the absence and presence of tetracycline, monitored over a 5-day time course. At48 h the culture was diluted to a density of 5 � 104 parasites/ml. (C) Growth of theBSF parasite lines as above, grown at 42 �C in the absence and presence oftetracycline for up to 6 h.
C.S. Ramos et al. / Experimental Parasitology 127 (2011) 228–237 235

Author's personal copy
judged by immunofluorescence, had the same distribution patternas the stationary-phase parental line (data not shown), but proteinsteady-state levels were increased approximately 6-fold. The par-tial functional rescue could also indicate the existence of otherproducts interacting with the META2 protein, which may havetheir concentration unbalanced by the excess of META2.
Aberrant forms in promastigote cultures have been observedafter disruption of other genes, such as the thymidine kinase ofL. major (Thiel et al., 2008) and HSP70-II gene in Leishmania infantum(Folgueira et al., 2008). However, these mutants were constitutivelyaltered, while META2-null parasites displayed normal shape andsize under regular culture conditions.
The META2 protein carboxyl-terminus contains the PF09149domain, found in the cysteine protease calpain. Several trypanoso-matid proteins containing this domain have been identified andnamed calpain-like proteins (Ersfeld et al., 2005). In fact, due toits C-terminal domain, the L. major META2 protein (GeneDBLmjF17.0870) has been included into a sub-family within the cal-pain-like proteins of trypanosomatids called small kinetoplastidcalpain-related protein (SKCRP) family. These are usually smallproteins (about 100 amino acids) and some have been found tobe modified by fatty acids (Ersfeld et al., 2005). They have also beenassociated with the cytoskeleton (Hertz-Fowler et al., 2001) andthe flagellum (Tull et al., 2004). Several SKCRPs are stage regulated,as is META2. The function of these proteins is, however, unknown.T. brucei CAP5.5, an 883 amino acid, stage-regulated calpain-related protein, has been recently shown to be essential for correctmorphogenesis during cell division. Ablation of CAP5.5 in procyclictrypanosomes results in aberrant organelle segregation and cytoki-nesis (Olego-Fernandez et al., 2009). Interestingly, other calpain-like proteins, such as T. brucei CALP1.3 and SKCRP7.2, were alsoshown to localise to the flagellar tip leading to a suggestion thatthese proteins might have sensory functions (Liu et al., 2010). Onthe other hand, a stress-related function has been suggested forthe Trypanosoma cruzi epimastigote-specific calpain-like proteinTcCAPPx.11 since increased temperature and pH shifts are associ-ated with an increase in TcCALPx.11 abundance (Giese et al., 2008).
The observation of a relationship between META2 and heatshock is interesting. The META domain is found in more than 300bacterial proteins as a single copy motif or repeated 2–3 times inthe same protein. It is defined as a domain found in proteins ofunknown function, some of which are described as heat shockproteins. Amongst those, the E. coli HslJ protein of 14 kDa has beenexperimentally shown to be a heat-inducible protein (Chuang andBlattner, 1993), to be upregulated in more pathogenic strains(Dowd and Ishizaki, 2006) and to be directly correlated with novo-biocin resistance (Vergnes et al., 2007). In eukaryotic organismsthis domain has been found so far only in the Kinetoplastidae,represented by the META proteins of Leishmania spp. and theirhomologues in Trypanosoma and Crithidia (www.genedb.org).
These observations prompted us to look at META2 expression inT. brucei. The stage-regulated RNA expression observed inLeishmania was absent in trypanosomes, where the same levelsof META2 transcript were found both in procyclic and bloodstreamforms by quantitative PCR. One of the three copies of the META2homologue genes in T. brucei, Tb927.5.2190, was recently de-scribed as upregulated in bloodstream forms (Liu et al., 2010). Thisdiscrepancy cannot be explained at present, but it will be interest-ing to verify whether the three copies of META2 in T. brucei aresimilarly regulated. Reducing the META2 mRNA levels inbloodstream forms through RNAi did not lead to decreased survivalunder heat stress. It will be interesting to investigate whether thelack of META2 protein gives rise to a different phenotype in procy-clic trypanosomes. On the other hand, since T. brucei is an extracel-lular parasite for the duration of its life cycle, the META2 proteincould perform a different task in these organisms.
In conclusion, we present evidence that the META2 protein isfunctionally related to stress responses in L. amazonensis. Uponentry into the mammalian host, resistance to heat and oxidativeshock are vital to parasite survival and the META2 protein seemsto be part of Leishmania’s arsenal to overcome the initial steps ofhost infection. On the other hand, at present we cannot rule outa role of META2 in the insect vector.
Acknowledgments
We thank Dr. Stephen Beverley for pX63Hyg, Dr. Paulo Ho forpAE vector, Dr. Elvira Saraiva for 3A1 monoclonal antibody, Dr. LuisCarlos de Souza Ferreira for providing novobiocin, Dr. Luiz Tosi andDr. Angela K. Cruz for helpful suggestions. We are also grateful toDanilo Ciccone Miguel for suggestions and support. We also thankthe Wellcome Trust Sanger Institute for making their data freelyavailable for analyses. This work was funded by Fundação deAmparo à Pesquisa do Estado de São Paulo (FAPESP) and ConselhoNacional de Desenvolvimento Científico e Tecnológico (CNPq).
References
Alibu, V.P., Storm, L., Haile, S., Clayton, C., Horn, D., 2005. A doubly inducible systemfor RNA interference and rapid RNAi plasmid construction in Trypanosomabrucei. Molecular and Biochemical Parasitology 139, 75–82.
Besteiro, S., Coombs, G.H., Mottram, J.C., 2004. A potential role for ICP, a leishmanialinhibitor of cysteine peptidases, in the interaction between host and parasite.Molecular Microbiology 54, 1224–1236.
Biebinger, S., Wirtz, L.E., Lorenz, P., Clayton, C., 1997. Vectors for inducibleexpression of toxic gene products in bloodstream and procyclic Trypanosomabrucei. Molecular and Biochemical Parasitology 85, 99–112.
Chuang, S.E., Blattner, F.R., 1993. Characterization of 26 new heat-shock genes ofEscherichia coli. Journal of Bacteriology 175, 5242–5252.
Coburn, C.M., Otteman, K.M., McNeely, T., Turco, S.J., Beverley, S.M., 1991. StableDNA transfection of a wide-range of trypanosomatids. Molecular andBiochemical Parasitology 46, 169–180.
Cruz, A., Coburn, C.M., Beverley, S.M., 1991. Double targeted gene replacement forcreating null mutants. Proceedings of the National Academy of Sciences of theUnited States of America 88, 7170–7174.
Denise, H., Poot, J., Jimenez, M., Ambit, A., Herrmann, D.C., Vermeulen, A.N., Coombs,G.H., Mottram, J.C., 2006. Studies on the CPA cysteine peptidase in theLeishmania infantum genome strain JPCM5. BMC Molecular Biology 7, 42.
Dowd, S.E., Ishizaki, H., 2006. Microarray based comparison of two Escherichia coliO157:H7 lineages. BMC Microbiology 6, 30.
Ersfeld, K., Barraclough, H., Gull, K., 2005. Evolutionary relationships and proteindomain architecture in an expanded calpain superfamily in kinetoplastidparasites. Journal of Molecular Evolution 61, 742–757.
Folgueira, C., Carrion, J., Moreno, J., Saugar, J.M., Canavate, C., Requena, J.M., 2008.Effects of the disruption of the HSP70-II gene on the growth, morphology, andvirulence of Leishmania infantum promastigotes. International Microbiology 11,81–89.
Giese, V., Dallagiovanna, B., Marchini, F.K., Pavoni, D.P., Krieger, M.A., Goldenberg, S.,2008. Trypanosoma cruzi: a stage-specific calpain-like protein is induced aftervarious kinds of stress. Memorias do Instituto Oswaldo Cruz 103, 598–601.
Gueiros-Filho, F.J., Beverley, S.M., 1996. Selection against the dihydrofolatereductase-thymidylate synthase (DHFR-TS) locus as a probe of geneticalterations in Leishmania major. Molecular and Cellular Biology 16, 5655–5663.
Guerra-Giraldez, C., Quijada, L., Clayton, C.E., 2002. Compartmentation of enzymesin a microbody, the glycosome, is essential in Trypanosoma brucei. Journal of CellScience 115, 2651–2658.
Hertz-Fowler, C., Ersfeld, K., Gull, K., 2001. CAP5.5, a life-cycle-regulated,cytoskeleton-associated protein is a member of a novel family of calpain-related proteins in Trypanosoma brucei. Molecular and Biochemical Parasitology116, 25–34.
Lainson, R., Shaw, J.J., 1987. Evolution, classification and geographical distribution.In: Peters, W., Killick-Kendrick, R. (Eds.), The Leishmaniases in Biology andMedicine. Academic Press, London, pp. 1–20.
Lilic, M., Jovanovic, M., Jovanovic, G., Savic, D.J., 2003. Identification of the CysB-regulated gene, hslJ, related to the Escherichia coli novobiocin resistancephenotype. FEMS Microbiology Letters 224, 239–246.
Liu, W., Apagyi, K., McLeavy, L., Ersfeld, K., 2010. Expression and cellular localisationof calpain-like proteins in Trypanosoma brucei. Molecular and BiochemicalParasitology 169, 20–26.
McKean, P.G., Denny, P.W., Knuepfer, E., Keen, J.K., Smith, D.F., 2001. Phenotypicchanges associated with deletion and overexpression of a stage-regulated genefamily in Leishmania. Cellular Microbiology 3, 511–523.
Medina-Acosta, E., Cross, G.A.M., 1993. Rapid isolation of DNA from trypanosomatidprotozoa using a simple mini-prep procedure. Molecular and BiochemicalParasitology 59, 327–329.
236 C.S. Ramos et al. / Experimental Parasitology 127 (2011) 228–237

Author's personal copy
Mosmann, T., 1983. Rapid colorimetric assay for cellular growth and survival –application to proliferation and cytotoxicity assays. Journal of ImmunologicalMethods 65, 55–63.
Nourbakhsh, F., Uliana, S.R., Smith, D.F., 1996. Characterisation and expression of astage-regulated gene of Leishmania major. Molecular and BiochemicalParasitology 76, 201–213.
Olego-Fernandez, S., Vaughan, S., Shaw, M.K., Gull, K., Ginger, M.L., 2009. Cellmorphogenesis of Trypanosoma brucei requires the paralogous, differentiallyexpressed calpain-related proteins CAP5.5 and CAP5.5v. Protist 160, 576–590.
Pinto-da-Silva, L.H., Fampa, P., Soares, D.C., Oliveira, S.M.P., Souto-Padron, T.,Saraiva, E.M., 2005. The 3a1-la monoclonal antibody reveals key features ofLeishmania (L.) amazonensis metacyclic promastigotes and inhibits procyclicsattachment to the sand fly midgut. International Journal for Parasitology 35,757–764.
Price, H.P., Panethymitaki, C., Goulding, D., Smith, D.F., 2005. Functional analysis offunctional analysis of TbARL1, an N-myristoylated Golgi protein essential forviability in bloodstream trypanosomes. Journal of Cell Science 118, 831–841.
Ramos, C.R.R., Abreu, P.A.E., Nascimento, A., Ho, P.L., 2004a. A high-copy T7Escherichia coli expression vector for the production of recombinant proteinswith a minimal N-terminal His-tagged fusion peptide. Brazilian Journal ofMedical and Biological Research 37, 1103–1109.
Ramos, C.S., Franco, F.A., Smith, D.F., Uliana, S.R., 2004b. Characterisation of a newLeishmania META gene and genomic analysis of the META cluster. FEMSMicrobiology Letters 238, 213–219.
Sacks, D.L., 1989. Metacyclogenesis in Leishmania promastigotes. ExperimentalParasitology 69, 100–103.
Thiel, M., Harder, S., Wiese, M., Kroemer, M., Bruchhaus, I., 2008. Involvement of aLeishmania thymidine kinase in flagellum formation, promastigote shape andgrowth as well as virulence. Molecular and Biochemical Parasitology 158, 152–162.
Tull, D., Vince, J.E., Callaghan, J.M., Naderer, T., Spurck, T., McFadden, G.I., Currie, G.,Ferguson, K., Bacic, A., McConville, M.J., 2004. SMP-1, a member of a new familyof small myristoylated proteins in kinetoplastid parasites, is targeted to theflagellum membrane in Leishmania. Molecular Biology of the Cell 15, 4775–4786.
Uliana, S.R., Goyal, N., Freymuller, E., Smith, D.F., 1999. Leishmania: overexpressionand comparative structural analysis of the stage-regulated meta 1 gene.Experimental Parasitology 92, 183–191.
Vergnes, B., Gourbal, B., Girard, I., Sundar, S., Drummelsmith, J., Ouellette, M., 2007.A proteomics screen implicates HSP83 and a small kinetoplastid calpain-relatedprotein in drug resistance in Leishmania donovani clinical field isolates bymodulating drug-induced programmed cell death. Molecular and CellularProteomics 6, 88–101.
C.S. Ramos et al. / Experimental Parasitology 127 (2011) 228–237 237




















