
A nonadjuvanted polypeptide nanoparticle vaccine confers long-lasting protection against rodent...
11
See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/38091002 A Nonadjuvanted Polypeptide Nanoparticle Vaccine Confers Long-Lasting Protection against Rodent Malaria Article in The Journal of Immunology · November 2009 DOI: 10.4049/jimmunol.0901957 · Source: PubMed CITATIONS 71 READS 55 9 authors, including: Some of the authors of this publication are also working on these related projects: Nanobiology and Nanomedicine View project Clara Brando Johns Hopkins University 19 PUBLICATIONS 457 CITATIONS SEE PROFILE Christian Mittelholzer Redbiotec AG 124 PUBLICATIONS 1,266 CITATIONS SEE PROFILE Ueli Aebi University of Basel 427 PUBLICATIONS 24,441 CITATIONS SEE PROFILE All content following this page was uploaded by Christian Mittelholzer on 04 December 2016. The user has requested enhancement of the downloaded file. All in-text references underlined in blue are added to the original document and are linked to publications on ResearchGate, letting you access and read them immediately.
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of A nonadjuvanted polypeptide nanoparticle vaccine confers long-lasting protection against rodent...

Seediscussions,stats,andauthorprofilesforthispublicationat:https://www.researchgate.net/publication/38091002
ANonadjuvantedPolypeptideNanoparticleVaccineConfersLong-LastingProtectionagainstRodentMalaria
ArticleinTheJournalofImmunology·November2009
DOI:10.4049/jimmunol.0901957·Source:PubMed
CITATIONS
71
READS
55
9authors,including:
Someoftheauthorsofthispublicationarealsoworkingontheserelatedprojects:
NanobiologyandNanomedicineViewproject
ClaraBrando
JohnsHopkinsUniversity
19PUBLICATIONS457CITATIONS
SEEPROFILE
ChristianMittelholzer
RedbiotecAG
124PUBLICATIONS1,266CITATIONS
SEEPROFILE
UeliAebi
UniversityofBasel
427PUBLICATIONS24,441CITATIONS
SEEPROFILE
AllcontentfollowingthispagewasuploadedbyChristianMittelholzeron04December2016.
Theuserhasrequestedenhancementofthedownloadedfile.Allin-textreferencesunderlinedinblueareaddedtotheoriginaldocumentandarelinkedtopublicationsonResearchGate,lettingyouaccessandreadthemimmediately.

A Nonadjuvanted Polypeptide Nanoparticle Vaccine ConfersLong-Lasting Protection against Rodent Malaria1
Stephen A. Kaba,* Clara Brando,* Qin Guo,* Christian Mittelholzer,† Senthilkumar Raman,†
David Tropel,† Ueli Aebi,† Peter Burkhard,‡ and David E. Lanar2*
We have designed and produced a prototypic malaria vaccine based on a highly versatile self-assembling polypeptide nanoparticle(SAPN) platform that can repetitively display antigenic epitopes. We used this platform to display a tandem repeat of the B cellimmunodominant repeat epitope (DPPPPNPN)2D of the malaria parasite Plasmodium berghei circumsporozoite protein. Admin-istered in saline, without the need for a heterologous adjuvant, the SAPN construct P4c-Mal conferred a long-lived, protectiveimmune response to mice with a broad range of genetically distinct immune backgrounds including the H-2b, H-2d, and H-2k
alleles. Immunized mice produced a CD4� T cell-dependent, high-titer, long-lasting, high-avidity Ab response against the B cellepitope. Mice were protected against an initial challenge of parasites up to 6 mo after the last immunization or for up to 15 moagainst a second challenge after an initial challenge of parasites had successfully been cleared. Furthermore, we demonstrate thatthe SAPN platform not only functions to deliver an ordered repetitive array of B cell peptide epitopes but operates as a classicalimmunological carrier to provide cognate help to the P4c-Mal-specific B cells. The Journal of Immunology, 2009, 183: 7268–7277.
M alaria, an infectious disease that is manifest in hun-dreds of millions of people each year, has no effectivevaccine on the market. Many potential vaccines have
been investigated, using various vaccine platforms including re-combinant proteins, nucleic acid, multiple antigenic peptides, vi-rus-like particles (VLP),3 and both bacterial and viral attenuatedmicroorganisms (1, 2). Recombinant-protein vaccines and DNAvaccines, unlike live-vector vaccines, often need the coadminis-tration of an immunostimulator or adjuvant to sufficiently activatethe immune effector mechanisms. In human trials, the nucleic acidand live-vector vaccines have not been efficacious on their own norwhen given in a prime-boost regimen (3–6). Given the scarcity ofhuman-use approved adjuvants, the broad development of protein-based vaccines for malaria has not significantly advanced (7). Onlythe Plasmodium falciparum vaccine RTS,S has proven effectivewhen combined with one of a series of adjuvants (AS01B, AS02A,AS02D) developed by the maker of RTS,S, GlaxoSmithKline (8).Clinical and field trials of other malaria recombinant proteins co-
administered with the GlaxoSmithKline adjuvants have not dem-onstrated the same protective efficacy (1). Although these resultsmay be more a choice of Ag than adjuvant, it has clearly beenshown that choice of adjuvant does affect the level of the immuneresponse (9).
Recently, a few nanoparticle-based vaccines have shown to beeffective in the induction of immune responses in animal modelswithout the need for an adjuvant (10–15). However, most of theseparticles are based on polymers that encapsulate Ag or create asolid support to which protein Ags are chemically coupled. Theactual physical array of epitopes on the Ag-presenting platformappears to influence the immune response and this cannot be wellcontrolled in solid-support platforms. Well ordered, repetitive dis-plays on VLPs (16) are among the strongest immunogens known(17, 18). A case for ordered Ag display was made more than 20years ago during an attempt to produce high-titer/high-affinitymonospecific Abs against the different capsid proteins of bacterio-phage T4 (19–23). More recently, Liu and Chen (24) have shownthat Ab affinity constants are as much as 2 logs higher when Agsare displayed in optimal density arrays. It is possible that Agspresented in an optimum configuration could induce a powerfulimmune response in the absence of extraneous adjuvant. For arecent review see Jennings and Bachmann (25).
Raman et al. have developed mechanically and chemically sta-ble nanoparticles using the intrinsic property of peptides and pro-teins to self-assemble to specific oligomers, which then leads to theformation of particles of well defined size and shape (26–28). Weuse these self-assembling polypeptide nanoparticles (SAPN) as thebasis for delivery of antigenic peptides of Plasmodium berghei asa model system for the development of a new vaccine platform.
The structural foundation of the SAPN is the molecular designof a monomeric linear polypeptide (LP) of �100 amino acids (Fig.1A). Each LP contains both a pentameric and a trimeric coiled-coiloligomerization domain, joined by a two-glycine residue flexiblelinker. The pentameric coiled-coil oligomerization domain is de-rived from the cartilage oligomeric matrix protein (COMP) (29)whereas the trimeric domain is a de novo-designed coiled-coildomain (30). Upon refolding from denaturing conditions, the
*Division of Malaria Vaccine Development, Walter Reed Army Institute of Research,Silver Spring, MD 20910; †M.E. Muller Institut, Basel, Switzerland; and ‡Institute ofMaterials Science, Department of Molecular and Cell Biology, University of Con-necticut, Storrs, CT 06269
Received for publication July 7, 2009. Accepted for publication October 2, 2009.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 C.M., D.T., and S.R. were supported by the Grants 8433.1 and Peptide NP from theKommission fur Technologie und Innovation (Switzerland) and the Swiss Nano-science Institute.2 Address correspondence and reprint requests to Dr. David E. Lanar, Division ofMalaria Vaccine Development, Walter Reed Army Institute of Research, 503 RobertGrant Ave., Silver Spring, MD 20910. E-mail address: [email protected] Abbreviations used in this paper: VLP, virus-like particle; AI, avidity index;COMP, cartilage oligomeric matrix protein; DC, dendritic cell; DLS, dynamiclight scattering; KO, knockout; LAL, Limulus amebocyte lysate; LP, linearpolypeptide; MHC II, MHC class II; NaSCN, sodium isothiocyanate; P4c-Mal/M,P4c-Mal in Montanide ISA-720; PbCSP, Plasmodium berghei circumsporozoiteprotein; R-PbCSP, recombinant-PbCSP; R-PbCSP/M, R-PbCSP protein in Mon-tanide ISA-720; SAPN, self-assembling polypeptide nanoparticle; TEM, trans-mission electron microscopy.
Copyright © 2009 by The American Association of Immunologists, Inc. 0022-1767/09/$2.00
The Journal of Immunology
www.jimmunol.org/cgi/doi/10.4049/jimmunol.0901957

peptides coassemble into SAPN (Fig. 1B). The assembly isdriven by the multiple coiled-coils formed by the two oligomer-ization domains. Ideally, such SAPN will have icosahedralsymmetry and thus resemble VLPs in their architecture (26).Because both the N- and C-terminal ends of the peptide areexposed on the surface, the SAPN are nicely suited for thepresentation of B cell epitopes in a repetitive-Ag display.
To test the vaccine efficacy of the platform to display peptidesof malaria parasites, we selected the tandem repeat(DPPPPNPN)2D of the B cell immunodominant repeat epitope ofthe malaria parasite Plasmodium berghei circumsporozoite protein(PbCSP) (31) to be displayed on the SAPN surface. It has previ-ously been shown that this tandem repeat, when displayed as amultiple antigenic peptide and administered in CFA, can conferprotection in mice against a lethal challenge of live P. bergheisporozoites (32). In this paper, we show that P4c-Mal, the malariaepitope displaying SAPN, when administered in PBS, can confer along-lived protective immune response to mice without the need ofa heterologous adjuvant and that the Abs produced in response tothe SAPN are of higher avidity than Abs produced against a nearfull-length recombinant PbCSP (R-PbCSP) delivered in the adju-vant Montanide ISA-720. Mice that received P4c-Mal were pro-tected against an initial challenge of parasites as much as 6 moafter the last immunization and for up to 15 mo against a secondchallenge after an initial challenge of parasites had successfullybeen cleared. Furthermore, we show that immunization with P4c-Mal SAPN generate CD4� T cell-dependent Abs which accountfor this protection.
Materials and MethodsStrains of mice
Six-to-eight-week old female C57BL/6 (H-2b) and BALB/c (H-2d) micewere used for most protection experiments. In addition, CD8 knockoutstrain B6/129S2-Cd8atm1Mak/J, the MHC class II (MHC II) knockout (KO)strain B6.129S2-H2dlAb1-Ea/J, the CD4 knockout strain B6.129S2-Cd4tm1Mak/J, or the athymic nude strain B6.Cg-Foxn1�/J of mice were usedto dissect the immune response to the SAPN. Two strains of TLR4-defi-cient (C57BL/10ScNJ and C3H/He3) mice were used to test for the pres-ence of bound endotoxin. All mice were obtained from The JacksonLaboratory.
Gene cloning
The sequence encoding the SAPN peptide was ligated into the BamHI/EcoRI restriction sites of mpPEP-T expression vector, which correspondsto pPEP-T lacking the laminin oligomerization domain (33). Oligonucle-otides encoding PbCSP B cell epitope (DPPPPNPN)2D were then clonedinto the XmaI/EcoRI restriction sites of the SAPN expression construct toyield the final amino acid sequence MGHHHHHHGDWKWDGGLVPRGSDEMLRELQETNAALQDVRELLRQQVRQITFLRCLLMGGRLLCRLEELERRLEELERRLEELERAINTVDLELAALRRRLEELARGGSGDPPPPNPNDPPPPNPND (bold is the CSP peptide). A construct, P4c-RD, was made that contained a random peptide sequence, IPSTAFTDIAWVRLPNHY, at the C-terminal end in place of the PbCSP epitope.
Protein purification, refolding, and analysis of the nanoparticlepolypeptide
Recombinant protein was expressed, purified, and refolded essentially aspreviously described (26). Briefly, the LP was expressed as a recombinantprotein in Escherichia coli and purified by metal affinity chromatographyfollowed by two polishing ion-exchange chromatography steps. The first,Q-Sepharose, captured the endotoxin, and the second, SP-Sepharose, con-centrated the protein. Throughout the purification protocol the LP was keptin a denatured state in 8 M urea, then the LP was slowly folded by stepwisedialysis to remove the urea. The shape and size of the P4c-Mal nanopar-ticles were studied by using transmission electron microscopy (TEM) anddynamic light scattering (DLS) (27). TEM analysis was preformed as pre-viously described (29) and photographed on a Zeiss EM910 transmissionelectron microscope (Carl Zeiss). The hydrodynamic diameter of the SAPNwas measured using a Zetasizer Nano S (Malvern Instruments) DLS in-strument in PBS at 25°C and pH 7.5.
SAPN containing different ratios of C-terminal peptides
To determine the percent of PbCSP peptide in a SAPN that was needed tostill be protective, we prepared a series of SAPN containing different ratiosof LP containing either the PbCSP repeat peptide epitope or the randompeptide sequence RD. After each LP-PbCSP or LP-RD was expressed andpurified, they were mixed in the ratio of 0:100, 10:90: 25:75, 50:50, 75:25,or 100:0 (LP-PbCSP:LP-RD). Urea was then removed by dialysis and finalSAPN analyzed as described (27).
Endotoxin analysis of the final product
Endotoxin levels in the final product were determined using the Pyro-chrome Limulus amebocyte lysate (LAL) test (Associates of Cape Cod)following the manufacturer’s instructions and as reported previously (34).
Immunization of mice
All animal study protocols were reviewed and approved by the WalterReed Army Institute of Research (WRAIR) Institutional Animal Care andUse Committee, conducted in compliance with the Animal Welfare Actand other federal statutes and regulations relating to animals and experi-ments involving animals, and adhered to principles stated in the Guide forthe Care and Use of Laboratory Animals. Mice were randomly divided intogroups of 5 or 10 and immunized i.p. three times at 14-day intervals. Whereindicated, a positive control group was immunized with irradiated P.berghei sporozoites according to methods described in Ref. 35.
Passive transfer of serum and splenocytes
Ten BALB/c or C57BL/6 mice were immunized three times at 14 daysintervals with 10 �g of P4c-Mal per mouse per injection (11) in the pres-ence or absence of Montanide ISA-720. One week after the third immu-nization, animals were killed and bled by cardiac puncture for processingof serum. Spleens were surgically removed for harvesting of splenocytes.Serum or splenocytes from the 10 animals were pooled. Groups of 10 naiveBALB/c or C57BL/6 mice were transfused with 100 �l of pooled serum(nondiluted), 1 � 107 cells, or both. The infectivity control group receivedneither cells nor serum. Animals were challenged with live sporozoites16 h after the transfer.
Challenge of mice with live P. berghei sporozoites
P. berghei sporozoites (ANKA strain), maintained by cyclical transmissionin mice and Anopheles stephensi, were dissected from mosquitoes 21–23days after their infectious blood meal and used within 6 h. Fourteen daysafter the final immunization or at other specific times on long-term memoryexperiments, mice were challenged with a lethal dose of live P. bergheisporozoites by i.v. inoculation. C57BL/6, MHC KO, and nude mice wereinjected with 1000 sporozoites and BALB/c mice were injected with 4000sporozoites per mouse.
Determination of infection following parasite challenge
Parasitemia was determined by examining Giemsa-stained thin smears pre-pared with blood from each mouse from days 6–15 after challenge. Para-sitemic animals were euthanized within 24–48 h of detecting blood stageparasites and an animal was considered fully protected if no parasites weredetected by 20 days post challenge. In no case did an animal developlow-grade parasitemia and self-cure.
Cellular response (ELISPOT)
Splenocytes from P4c-Mal immunized or naive animals were resuspendedat a concentration of 4 � 106/ml in complete medium. One hundred mi-croliters of cell suspension was distributed into 96-well ELISPOT platesfor IL-2 release determination and stimulated with P4c-Mal protein (5 �g/well) or Con-A (5 �g/ml) or left in medium alone with or without anti-CD4�-blocking Ab (2 �g/ml). To activate memory T cells, all culturescontained 4 �g/ml each of anti-CD28 and anti-CD49d Abs (36). The num-ber of cells secreting IL-2 was determined.
Ab titer in response to vaccination
Ab responses to the PbCSP peptide (DPPPPNPN)2D were measured byELISA as previously described (37). Briefly, 96-well microplates (Dynax)were coated with 100 ng of the synthetic PbCSP peptide (GenScript) perwell overnight at 4°C. The Ag was blocked for 1 h at 37°C with PBS (pH7.4) containing 0.05% Tween 20 and 1% casein (Sigma-Aldrich). Plateswere washed three times with PBS-T and incubated for 2 h at room tem-perature with individual mouse sera in triplicate wells per serum sample.Plates were washed again and incubated for 1 h at room temperature with
7269The Journal of Immunology
1/5000 diluted (PBS) secondary anti-mouse Ig (total IgG) or the differentIg subclasses (IgG1, IgG2a, IgG2b, and IgG2c) Abs labeled with HRP(Southern Biotechnology Associates). Plates were washed and developedby adding ABTS substrate (Kirkegaard & Perry Laboratories). Color re-action was measured in a Dynatech MR5000 microplate reader by deter-mining OD at 405 nm (OD(405 nm)). The results were calculated as meanOD
(405 nm)readings of duplicate assays � SDs.
Ab avidity index (AI) determination
An initial concentration of sodium isothiocyanate (NaSCN) that eluted50% of the total bound IgG was determined by the method described byPullen et al. (38). Then two side-by-side 4-fold serial dilutions for a secondELISA were done (in triplicate). After three washes with PBS (pH 7.4)containing 0.05% Tween 20 and one wash with PBS, 100 �l of the con-centration of NaSCN (1.5 M dissolved in PBS) measured in the initialavidity determination was added to one set of the serial dilutions on theplate and plain PBS was added to the other dilution series. The same elu-tion concentration of NaSCN determined for total IgG was used for elutionof IgG subclasses. After 15-min incubation, all wells were washed anddeveloped as described above. The AI was calculated based on the ratioof the areas derived from under the curves obtained by the plot of OD(OD(405 nm)) and log of the sera dilution in the ELISA experiment withand without NaSCN treatment as described (39).
Statistics
Student’s t tests were performed using Excel software (Microsoft). Theassociation of Ab titer and protection was determined using logistic re-gression analysis in the program SAS (SAS Institute).
ResultsPreparation and characterization of P4c-Mal SAPN
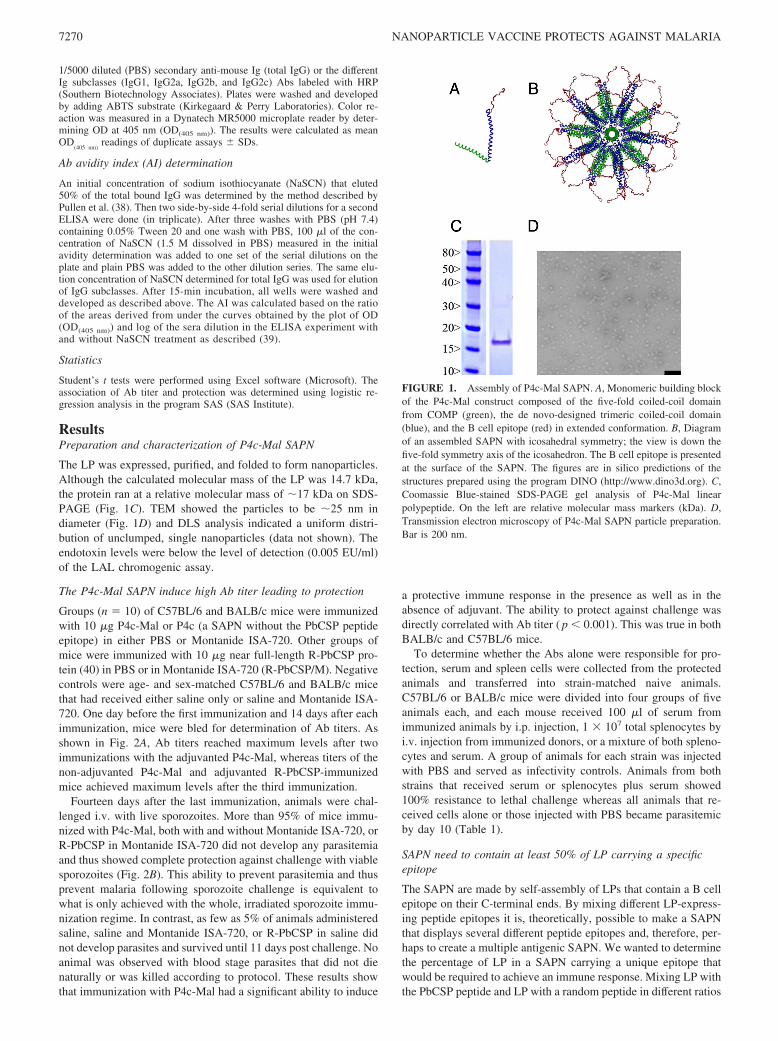
The LP was expressed, purified, and folded to form nanoparticles.Although the calculated molecular mass of the LP was 14.7 kDa,the protein ran at a relative molecular mass of �17 kDa on SDS-PAGE (Fig. 1C). TEM showed the particles to be �25 nm indiameter (Fig. 1D) and DLS analysis indicated a uniform distri-bution of unclumped, single nanoparticles (data not shown). Theendotoxin levels were below the level of detection (0.005 EU/ml)of the LAL chromogenic assay.
The P4c-Mal SAPN induce high Ab titer leading to protection
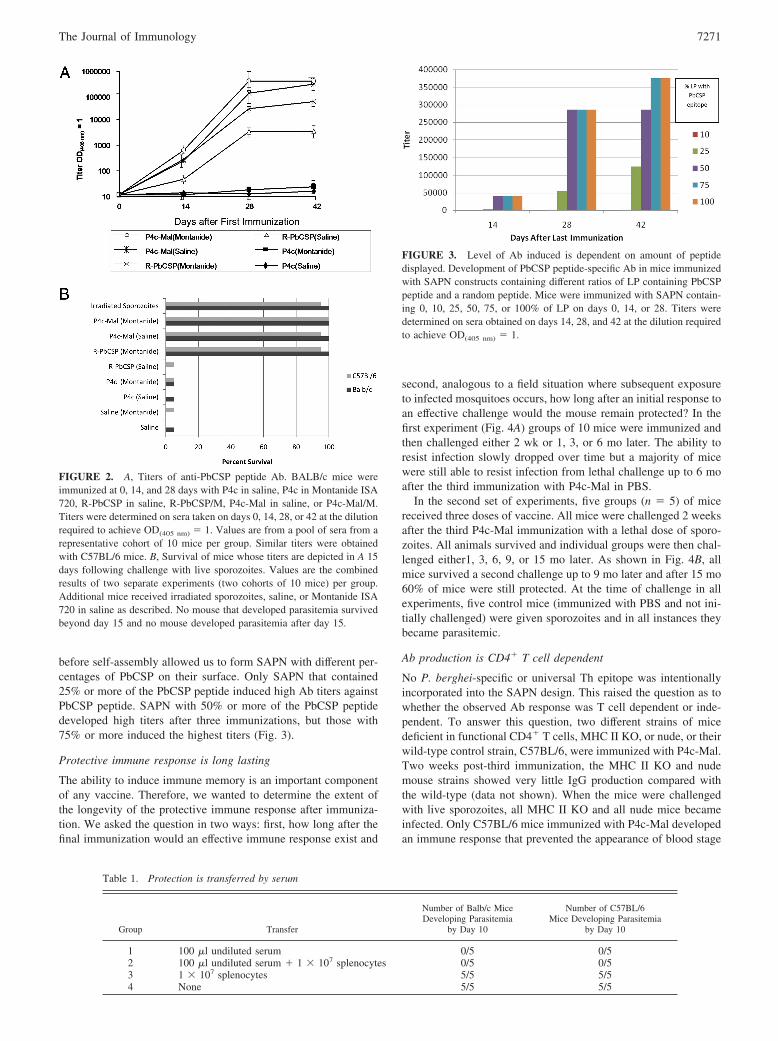
Groups (n � 10) of C57BL/6 and BALB/c mice were immunizedwith 10 �g P4c-Mal or P4c (a SAPN without the PbCSP peptideepitope) in either PBS or Montanide ISA-720. Other groups ofmice were immunized with 10 �g near full-length R-PbCSP pro-tein (40) in PBS or in Montanide ISA-720 (R-PbCSP/M). Negativecontrols were age- and sex-matched C57BL/6 and BALB/c micethat had received either saline only or saline and Montanide ISA-720. One day before the first immunization and 14 days after eachimmunization, mice were bled for determination of Ab titers. Asshown in Fig. 2A, Ab titers reached maximum levels after twoimmunizations with the adjuvanted P4c-Mal, whereas titers of thenon-adjuvanted P4c-Mal and adjuvanted R-PbCSP-immunizedmice achieved maximum levels after the third immunization.
Fourteen days after the last immunization, animals were chal-lenged i.v. with live sporozoites. More than 95% of mice immu-nized with P4c-Mal, both with and without Montanide ISA-720, orR-PbCSP in Montanide ISA-720 did not develop any parasitemiaand thus showed complete protection against challenge with viablesporozoites (Fig. 2B). This ability to prevent parasitemia and thusprevent malaria following sporozoite challenge is equivalent towhat is only achieved with the whole, irradiated sporozoite immu-nization regime. In contrast, as few as 5% of animals administeredsaline, saline and Montanide ISA-720, or R-PbCSP in saline didnot develop parasites and survived until 11 days post challenge. Noanimal was observed with blood stage parasites that did not dienaturally or was killed according to protocol. These results showthat immunization with P4c-Mal had a significant ability to induce
a protective immune response in the presence as well as in theabsence of adjuvant. The ability to protect against challenge wasdirectly correlated with Ab titer ( p � 0.001). This was true in bothBALB/c and C57BL/6 mice.
To determine whether the Abs alone were responsible for pro-tection, serum and spleen cells were collected from the protectedanimals and transferred into strain-matched naive animals.C57BL/6 or BALB/c mice were divided into four groups of fiveanimals each, and each mouse received 100 �l of serum fromimmunized animals by i.p. injection, 1 � 107 total splenocytes byi.v. injection from immunized donors, or a mixture of both spleno-cytes and serum. A group of animals for each strain was injectedwith PBS and served as infectivity controls. Animals from bothstrains that received serum or splenocytes plus serum showed100% resistance to lethal challenge whereas all animals that re-ceived cells alone or those injected with PBS became parasitemicby day 10 (Table 1).
SAPN need to contain at least 50% of LP carrying a specificepitope
The SAPN are made by self-assembly of LPs that contain a B cellepitope on their C-terminal ends. By mixing different LP-express-ing peptide epitopes it is, theoretically, possible to make a SAPNthat displays several different peptide epitopes and, therefore, per-haps to create a multiple antigenic SAPN. We wanted to determinethe percentage of LP in a SAPN carrying a unique epitope thatwould be required to achieve an immune response. Mixing LP withthe PbCSP peptide and LP with a random peptide in different ratios
FIGURE 1. Assembly of P4c-Mal SAPN. A, Monomeric building blockof the P4c-Mal construct composed of the five-fold coiled-coil domainfrom COMP (green), the de novo-designed trimeric coiled-coil domain(blue), and the B cell epitope (red) in extended conformation. B, Diagramof an assembled SAPN with icosahedral symmetry; the view is down thefive-fold symmetry axis of the icosahedron. The B cell epitope is presentedat the surface of the SAPN. The figures are in silico predictions of thestructures prepared using the program DINO (http://www.dino3d.org). C,Coomassie Blue-stained SDS-PAGE gel analysis of P4c-Mal linearpolypeptide. On the left are relative molecular mass markers (kDa). D,Transmission electron microscopy of P4c-Mal SAPN particle preparation.Bar is 200 nm.
7270 NANOPARTICLE VACCINE PROTECTS AGAINST MALARIA
before self-assembly allowed us to form SAPN with different per-centages of PbCSP on their surface. Only SAPN that contained25% or more of the PbCSP peptide induced high Ab titers againstPbCSP peptide. SAPN with 50% or more of the PbCSP peptidedeveloped high titers after three immunizations, but those with75% or more induced the highest titers (Fig. 3).
Protective immune response is long lasting
The ability to induce immune memory is an important componentof any vaccine. Therefore, we wanted to determine the extent ofthe longevity of the protective immune response after immuniza-tion. We asked the question in two ways: first, how long after thefinal immunization would an effective immune response exist and
second, analogous to a field situation where subsequent exposureto infected mosquitoes occurs, how long after an initial response toan effective challenge would the mouse remain protected? In thefirst experiment (Fig. 4A) groups of 10 mice were immunized andthen challenged either 2 wk or 1, 3, or 6 mo later. The ability toresist infection slowly dropped over time but a majority of micewere still able to resist infection from lethal challenge up to 6 moafter the third immunization with P4c-Mal in PBS.
In the second set of experiments, five groups (n � 5) of micereceived three doses of vaccine. All mice were challenged 2 weeksafter the third P4c-Mal immunization with a lethal dose of sporo-zoites. All animals survived and individual groups were then chal-lenged either1, 3, 6, 9, or 15 mo later. As shown in Fig. 4B, allmice survived a second challenge up to 9 mo later and after 15 mo60% of mice were still protected. At the time of challenge in allexperiments, five control mice (immunized with PBS and not ini-tially challenged) were given sporozoites and in all instances theybecame parasitemic.
Ab production is CD4� T cell dependent
No P. berghei-specific or universal Th epitope was intentionallyincorporated into the SAPN design. This raised the question as towhether the observed Ab response was T cell dependent or inde-pendent. To answer this question, two different strains of micedeficient in functional CD4� T cells, MHC II KO, or nude, or theirwild-type control strain, C57BL/6, were immunized with P4c-Mal.Two weeks post-third immunization, the MHC II KO and nudemouse strains showed very little IgG production compared withthe wild-type (data not shown). When the mice were challengedwith live sporozoites, all MHC II KO and all nude mice becameinfected. Only C57BL/6 mice immunized with P4c-Mal developedan immune response that prevented the appearance of blood stage
FIGURE 2. A, Titers of anti-PbCSP peptide Ab. BALB/c mice wereimmunized at 0, 14, and 28 days with P4c in saline, P4c in Montanide ISA720, R-PbCSP in saline, R-PbCSP/M, P4c-Mal in saline, or P4c-Mal/M.Titers were determined on sera taken on days 0, 14, 28, or 42 at the dilutionrequired to achieve OD(405 nm) � 1. Values are from a pool of sera from arepresentative cohort of 10 mice per group. Similar titers were obtainedwith C57BL/6 mice. B, Survival of mice whose titers are depicted in A 15days following challenge with live sporozoites. Values are the combinedresults of two separate experiments (two cohorts of 10 mice) per group.Additional mice received irradiated sporozoites, saline, or Montanide ISA720 in saline as described. No mouse that developed parasitemia survivedbeyond day 15 and no mouse developed parasitemia after day 15.
FIGURE 3. Level of Ab induced is dependent on amount of peptidedisplayed. Development of PbCSP peptide-specific Ab in mice immunizedwith SAPN constructs containing different ratios of LP containing PbCSPpeptide and a random peptide. Mice were immunized with SAPN contain-ing 0, 10, 25, 50, 75, or 100% of LP on days 0, 14, or 28. Titers weredetermined on sera obtained on days 14, 28, and 42 at the dilution requiredto achieve OD(405 nm) � 1.
Table 1. Protection is transferred by serum
Group Transfer
Number of Balb/c MiceDeveloping Parasitemia
by Day 10
Number of C57BL/6Mice Developing Parasitemia
by Day 10
1 100 �l undiluted serum 0/5 0/52 100 �l undiluted serum � 1 � 107 splenocytes 0/5 0/53 1 � 107 splenocytes 5/5 5/54 None 5/5 5/5
7271The Journal of Immunology

parasites (Table 2). Thus, the absence of functional CD4� T cellsimpairs the ability of mice to generate protective Abs when im-munized with P4c-Mal.
Ag-specific CD4� cells are generated in response to SAPNimmunization
Determining that CD4� T cells are needed to initiate the humoralimmune response raised the question of how these cognate CD4�
helper cells are generated. The SAPN is a complex protein struc-ture and we suspected that a CD4� stimulatory epitope was likelyto be part of the LP that makes up the SAPN. To test this hypoth-esis, a group of C57BL/6 mice (n � 5) were immunized withP4c-Mal and mice and control groups were euthanized 2 weekspost-third immunization, spleens were removed, and splenocyteswere pooled and cultured in ELISPOT plates. During culture,splenocytes were incubated with medium only or medium contain-ing P4c-Mal (10 �g/ml) or Con A (5 �g/ml) in the presence orabsence of 2 �g/ml the CD4� T cell-specific Abs. The incubationof splenocytes from mice immunized with P4c-Mal resulted in theinduction of IL-2 production whereas incubation of P4c-Mal withsplenocytes from naive mice did not (Fig. 5). The IL-2 productionwas abrogated by inclusion of blocking anti-CD4� Abs in the me-dium, thus indicating that the IL-2 producing cells were CD4� T
cells. Inclusion of Con A in the culture caused IL-2 production insplenocytes from naive and immunized mice but was not affectedby anti-CD4� Abs, showing that the ablation of the CD4� T cellresponse was not an artifact of the Abs used. Thus, the SAPNplatform functions not only to deliver an ordered repetitive array ofB cell peptide epitopes but operates as a classical immunologicalcarrier to provide cognate help to the P4c-Mal-specific B cells.
Testing for potentially bound endotoxin using TLR4�/� mice
It is possible that endotoxin could be tightly bound in the crevicesbetween the adjacent coiled-coils of the SAPN and because boundendotoxin is not detected by the LAL assay we needed to rule outthe possibility that the adjuvant-like capacity of the SAPN was dueto endogenously bound bacterial LPS. Because LPS is a TLR4agonist, we reasoned that if the immunostimulatory activity ob-served with SAPN immunizations was due to contamination with
FIGURE 5. P4c-Mal induces the production of IL-2 in CD4� T cells.Splenocytes from P4c-Mal immunized (IM) or naive C57BL/6 mice werecultured with P4c-Mal nanoparticles, Con A, or medium with or withoutanti-CD4� T cell-blocking Abs. After 18 h the numbers of cells secretingIL-2 were determined. Values are representative of one of four separateexperiments. �, p � 0.001.
FIGURE 4. P4c-Mal induces long-term immunity that can be boosted bychallenge (Ch) with live sporozoites.A, Groups of mice (n � 10) receivedthree doses of P4c-Mal or saline at 0,2, and 4 weeks. One group each waschallenged with live sporozoites ei-ther 14 days, or 1, 3, or 6 mo later. B,Groups of mice (n � 10) receivedthree doses of P4c-Mal or saline at 0,2, and 4 weeks. All mice that receivedP4c-Mal were challenged with livesporozoites two weeks post-thirddose of vaccine. All mice survivedchallenge. One group (n � 5) each ofvaccinated and challenged mice ormice receiving saline only were chal-lenged a second time either 1, 3, 6, 9,or 15 mo later with live sporozoites.
Table 2. T cell dependency of protective immune response withP4c-Mal immunization
Mice Immunized with% Developing Parasitemia
After Challengea
C57BL/6 PBS 100C57BL/6 P4c-Mal 0Nude PBS 100Nude P4c-Mal 100MHC II KO PBS 100MHC II KO P4c-Mal 100
a By day 10 post-challenge with 1000 sporozoites.
7272 NANOPARTICLE VACCINE PROTECTS AGAINST MALARIA

bound endotoxin we would not be able to protect TLR4-null micewith a P4c-Mal preparation. Therefore, we immunized two differ-ent TLR4-null, endotoxin-resistant mouse strains C57BL/6/10ScNJ (TLR4�/�) and C3H/He3 (defective LPS-response alleleTlr4Lps-d) as well as their wild-type equivalent C57BL/6 and C3Hmice, respectively, with P4c-Mal. After immunization with P4c-Mal in PBS and sporozoite challenge, three of five TLR4�/�- andfour of five Tlr4Lps-d-deficient mice and five of five of each of theirwild-type counterparts were protected. All mice that survived de-veloped moderate to high Ag-specific Ab titers whereas all micethat died failed to develop any significant Ag-specific Ab. The factthat we were not able to protect 100% of all the TLR4-null miceis not surprising. It has been shown that there is an increased sus-ceptibly to pathogens when TLR4 signaling is impaired (41). How-ever, the ability to protect 70% of TLR4-null mice strongly indi-cates that LPS is not involved in the stimulation of the immunesystem in response to SAPN immunization.
IgG subclass Ab responses
Anti-PbCSP peptide Ab levels were determined in serum fromC57BL/6 and BALB/c mice immunized with P4c-Mal. The geo-metric mean values of the total IgG anti-PbCSP Abs or IgG sub-class Abs in both strains are illustrated in Fig. 6. We measuredIgG2a in BALB/c mice and IgG2c in C57BL/6 mice. C57BL/6mice lack the Igh-1a allele that codes for the IgG2a but insteadexpress IgG2c from the Igh-1b allele (42, 43). Sera were pooledfrom 10 mice in each group 2-week post-third immunization. Im-munization with P4c-Mal and P4c-Mal/M resulted predominantlyin the induction of specific IgG1 and IgG2a/c Abs. C57BL/6 miceimmunized with R-PbCSP/M resulted in the induction of onlyIgG1 Abs (Figs. 6B and 7A).
In C57BL/6 mice, P4c-Mal induced a significantly largeramount of IgG1 Ab than P4c-Mal/M ( p � 0.001; Fig. 6B). Al-though over four times the amount of IgG1 was produced inBALB/c mice than in C57BL/6 ( p � 0.0002), no significant dif-ference was observed in the amount of IgG1 subclass Abs whenBALB/c were immunized with either P4c-Mal or P4c-Mal/M ( p �0.2). The formulation of P4c-Mal with Montanide ISA-720 signif-icantly affected IgG2a responses in BALB/c mice ( p � 0.001).IgG2c Ab levels produced in C57BL/6 mice immunized with ei-ther P4c-Mal or P4c-Mal/M are statistically the same ( p � 0.4).
Determination of AI of Abs made against PbCSP repeat epitope
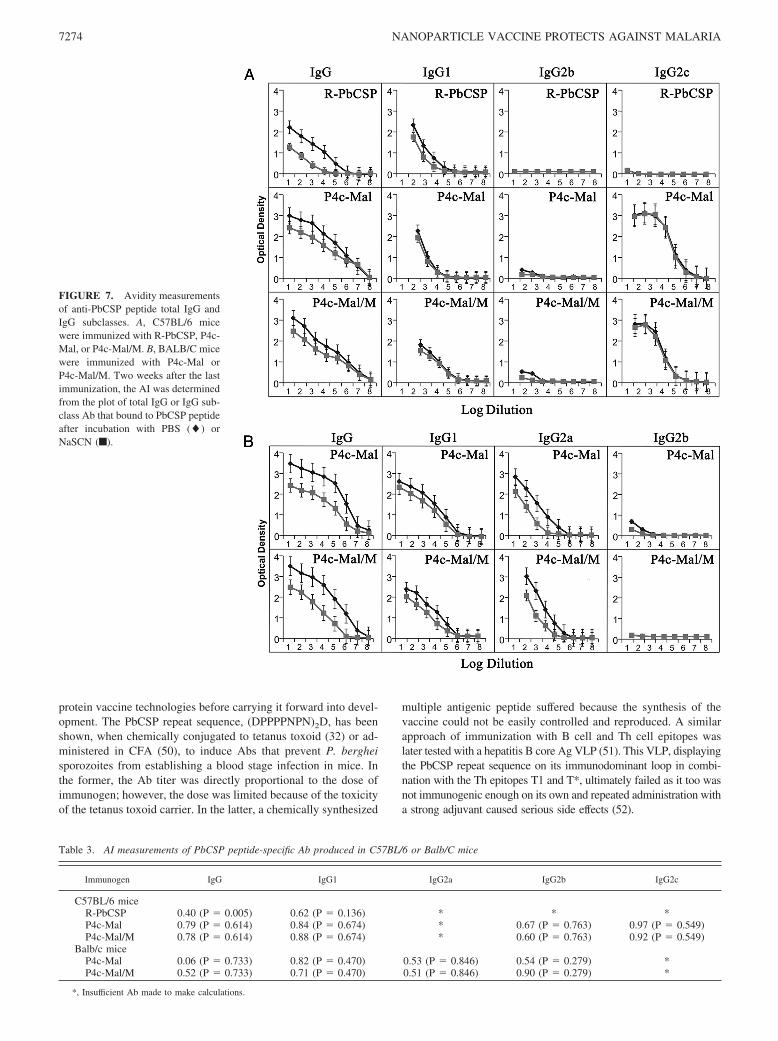
The chaotropic ELISA elution assay was used for determination ofthe AI of the Abs induced by SAPN-presented or recombinantprotein-presented epitopes. In Fig. 7, the OD(405 nm) values of theELISA from each group of mice were plotted against the respec-tive log of the sera dilutions. Rather than using a single specificpoint on the curve to determine the AI, the area under the curvewas used (39). Table 3 shows the calculated AI for sera fromR-PbCSP/M, P4c-Mal, and P4c-Mal/M Ags. Immunization withR-PbCSP/M induced only IgG1 subclass Ab. The AI value (0.4)for total IgG for R-PbCSP/M was significantly lower ( p � 0.005)than for any SAPN-PbCSP peptide-induced IgG, regardless ofwhether the epitope was delivered in adjuvant ( p � 0.5) or saline( p � 0.5). AI values for the total IgG induced by P4c-Mal orP4c-Mal/M were statistically similar. For IgG1, there was no sig-nificant difference among the AI values for R-PbCSP/M ( p � 0.1)and the P4c-Mal ( p � 0.5) or P4c-Mal/M ( p � 0.5). For IgG2a/cand IgG, there was no significant difference between the AI valuesAbs induced by either of the SAPN-PbCSP peptides. Remarkably,the AI for IgG2c was close to 1, indicating that this particular IgGsubclass had an extremely high avidity for the circumsporozoiteprotein peptide epitope.
DiscussionDespite the recent limited successes of RTS,S in field trials (8, 44),the development of a vaccine for malaria has been a disappointingone. Neither live attenuated viral vectors nor DNA-based vaccines,alone or in combination, have worked effectively (3–5, 45–48). Inaddition, the effectiveness of protein based vaccines, even RTS,S,has been limited by their selection of a coadministered adjuvant(49). We present here a new platform technology based on self-assembling polypeptides that form nanoparticles (27) to induce aprotective immune response against whole organism challenge invivo. We have found that the SAPN functions as a vehicle for thepresentation of selected antigenic epitopes and acts as a powerfulimmunostimulatory entity.
To test the feasibility of this new platform to induce an immuneresponse against an otherwise lethal infectious agent, we chose toconstruct a vaccine that presents a known protective B cell epitopeas a test Ag. We reasoned that the platform would have to besubstantially better in the induction of an immune response and indemonstrating protective efficacy than many of the other existing
FIGURE 6. P4c-Mal induces amixed TH1/TH2 IgG subclass profile.C57BL/6 or BALB/C mice were im-munized with P4c-Mal in saline orP4c-Mal/M. A group of C57BL/6mice also received R-PbCSP/M. To-tal IgG (A) and IgG subclass titers (B)were determined on pooled sera ob-tained 2 weeks after the last immuni-zation. Error bars depict the SD of thegeometric mean values.
7273The Journal of Immunology
protein vaccine technologies before carrying it forward into devel-opment. The PbCSP repeat sequence, (DPPPPNPN)2D, has beenshown, when chemically conjugated to tetanus toxoid (32) or ad-ministered in CFA (50), to induce Abs that prevent P. bergheisporozoites from establishing a blood stage infection in mice. Inthe former, the Ab titer was directly proportional to the dose ofimmunogen; however, the dose was limited because of the toxicityof the tetanus toxoid carrier. In the latter, a chemically synthesized
multiple antigenic peptide suffered because the synthesis of thevaccine could not be easily controlled and reproduced. A similarapproach of immunization with B cell and Th cell epitopes waslater tested with a hepatitis B core Ag VLP (51). This VLP, displayingthe PbCSP repeat sequence on its immunodominant loop in combi-nation with the Th epitopes T1 and T*, ultimately failed as it too wasnot immunogenic enough on its own and repeated administration witha strong adjuvant caused serious side effects (52).
Table 3. AI measurements of PbCSP peptide-specific Ab produced in C57BL/6 or Balb/C mice
Immunogen IgG IgG1 IgG2a IgG2b IgG2c
C57BL/6 miceR-PbCSP 0.40 (P � 0.005) 0.62 (P � 0.136) * * *P4c-Mal 0.79 (P � 0.614) 0.84 (P � 0.674) * 0.67 (P � 0.763) 0.97 (P � 0.549)P4c-Mal/M 0.78 (P � 0.614) 0.88 (P � 0.674) * 0.60 (P � 0.763) 0.92 (P � 0.549)
Balb/c miceP4c-Mal 0.06 (P � 0.733) 0.82 (P � 0.470) 0.53 (P � 0.846) 0.54 (P � 0.279) *P4c-Mal/M 0.52 (P � 0.733) 0.71 (P � 0.470) 0.51 (P � 0.846) 0.90 (P � 0.279) *
*, Insufficient Ab made to make calculations.
FIGURE 7. Avidity measurementsof anti-PbCSP peptide total IgG andIgG subclasses. A, C57BL/6 micewere immunized with R-PbCSP, P4c-Mal, or P4c-Mal/M. B, BALB/C micewere immunized with P4c-Mal orP4c-Mal/M. Two weeks after the lastimmunization, the AI was determinedfrom the plot of total IgG or IgG sub-class Ab that bound to PbCSP peptideafter incubation with PBS (�) orNaSCN (f).
7274 NANOPARTICLE VACCINE PROTECTS AGAINST MALARIA

Immunization of mice with the P4c-Mal SAPN in saline pro-vided complete protection to a lethal challenge of sporozoites 1month after the third dose of vaccine. Subsequent challenge of thesame mice showed a sustained immune response capable of pro-tective efficacy against an otherwise lethal challenge of live sporo-zoites as late as 15 mo later. This level of protection has only beendemonstrated previously with the whole cell irradiated sporozoitemodel of immunization (35). We have shown that Abs were theeffector of protection as serum was required to transfer protectionwhereas cells alone could not and that the response was T cell-dependent as MHC II KO animals were incapable of mounting aprotective response.
The T cells were demonstrated to be SAPN-specific, providingcognate help to the P4c-Mal peptide-specific B cells. It is wellestablished that to achieve a T cell-dependent B cell response, Bcells need to interact with the cognate T cells that provide help forthe Ab production. At the same time, only activated cognate Tcells can provide help, and T cell activation only occurs uponsimultaneous stimulation of the TCR and the ligand for costimu-latory molecules. Conventionally, adjuvant induces expression ofhigh levels of costimulatory molecules on the dendritic cells (DC)presenting Ag to T cells, thus enabling them to simultaneouslyprovide signal 1 (Ag) and signal 2 (costimulatory molecule). Withthis platform, we did not need to use adjuvant, raising the question:What was the initializing DC maturation to provide signal 2? Stud-ies in our laboratory have shown that this T cell help came from asequence lying with the COMP pentameric oligomerization do-main. When this region was replaced with another pentameric oli-gomerization domain, T cell help was greatly diminished. How-ever, inclusion of a universal Th epitope (PADRE) restored theactivation potential of the SAPN (our manuscript in preparation).
Other reports using nanoparticles to present Ag have used inertsubstrates such as gold particles coated with azobenzene disulfidedye (53) or polymers of proteinaceous materials as the substrate.Polypeptides containing T or B cell epitopes bound to polymers ofglutamic acid or poly-L-lysine-coated polystyrene (10, 54–56)have been used to enhance DNA vaccine efficacy (12). Most of theimmunostimulatory effects of those nanoparticles could be attrib-uted to their interaction with DC in the skin or the lymph nodes.Recent studies have presented convincing evidence that small size(20–30 nm) particles are preferentially transported to the lymphnodes where the particles are presented to immature DC (14, 15,57, 58). Once internalized in the DC, the nanoparticles are pro-cessed and antigenic epitopes are released to activate MHC class Iand II pathways, leading to induction of both CD4� and CD8� Tcell activation.
The high levels of IgG1 and IgG2a/c, as well as a considerableIgG2b subclass of Ig, reflect a mixed Th1/Th2 response. In mice,IgG1 responses are usually associated with Th2 responses,whereas high levels of IgG2a, sometimes associated with IgG2band IgG3, are thought to reflect Th1 responses. Several parametersinfluence the IgG subclass responses to proteins, including thedose of Ag used, use of adjuvants or routes of immunization, andthe intrinsic immunogenicity of the protein itself (59–65). We didnot see major differences in IgG subclasses with different routes ofimmunization (data not shown) and adjuvants added to SAPN didnot influence IgG subclass distribution. Therefore, the major ef-fector of the IgG subclass appears to be associated with an intrinsicimmunogenic property of the SAPN.
A surprising finding in these studies was the high avidity andsubclass specificity of the Abs produced. Of note is the extremelyhigh avidity of the IgG2c subclass Abs. Immunization with R-PbCSP in the adjuvant Montanide ISA-720 did not induce anydetectable IgG2c Abs, whereas high titer, high avidity IgG2c Abs
were generated against P4c-Mal with or without adjuvant. Clearlythe effectiveness of a vaccine that works predominately by induc-tion of neutralizing Abs, as is the mode of action of the immuneresponse induced in these studies, is dependent not only on theamount of Ab induced but also the avidity of those Abs. The com-plex cellular events that are needed to generate high-affinity Absecreting cells are not clearly understood. B cells in the germinalcenters undergo somatic hypermutation of V(D)J Ig genes andcompetition for Ag retained on the germinal center follicular DCresults in the selection of B cells secreting the highest affinity Abs(66). Many things influence the combined affinity (avidity) of Absto antigenic determinants, including the valence of the Ab and thevalence of the Ag. The physical Ag properties or adjuvant assetsthat influence the maturation process, which may be essential forthe protective efficacy, have not been characterized following im-munization with T cell-dependent protein Ags but is clearly im-portant in vaccine development (67). The coiled-coil nature of theSAPN may influence the presentation of the Ag and maturation ofthe Ab affinity because linear recombinant protein with the sameepitope given without adjuvant (R-PbCSP) or a modified P4c-MalLP that would not form a SAPN (data not shown) did not induceany significant Ab and failed to provide a protective immuneresponse.
The nanoparticles used in this study appear to display B cellepitopes in a very immunostimulatory array. The particles aresmall, �25 nm in size, and contain strong hydrophobic oligomer-ization domains that dictate their self-assembly. Recently we haveshown that they are efficiently taken up by and activate mousemacrophages and bone marrow-derived DC (manuscript in prep-aration). The platform has recently been used to effectively presentan otherwise poor antigenic determinant to make mAbs against aconserved actin domain (28), as well as to induce in vitro neutral-izing Abs against a SARS virus epitope (26). We have now ex-tended the use of this platform to induce an in vivo immune re-sponse that is protective against challenge with a lethal parasite.When administered in saline alone, the SAPN are capable of in-ducing maturation of a long-term memory response to a B cellepitope. The induced Abs protect against an otherwise lethal chal-lenge of sporozoites. It should be noted that when the leadingmalaria vaccine candidate, RTS,S, itself a VLP vaccine formula-tion, was administered to mice in saline, no significant anti-circumsporozoite protein repeat or HBsAg Ab was detected (68).This clearly shows that nanoparticles or VLP, a priori, are not thekey to effective immune responses. We are now determiningwhether the SAPN platform can be used to make a vaccine againstP. falciparum and P. vivax malaria; whether it can induce a CD8�-mediated immune response, and whether it is effective in higherorder mammals such as rabbits and nonhuman primates. Further-more, to make a vaccine for human use, autoimmune reactionscould develop due to the immune responses induced against theCOMP sequence in the SAPN scaffold because, although the se-quence was derived from mouse COMP, it also has a high homol-ogy to human COMP. Therefore, in advanced designs, we havereplaced the COMP with a de novo designed sequence withouthomology to any human sequences in the protein databank.
AcknowledgmentsR-PbCSP was a gift from E. Angov, WRAIR, Silver Spring, MD.
DisclosuresThis work was performed while S.A.K. held a National Research CouncilResearch Associate award at WRAIR. D.E.L. received funding from theMilitary Infectious Disease Research Program. U.A. and P.B. hold shares
7275The Journal of Immunology

in the company Alpha-O Peptides. The other authors have no financialconflict of interest.
References1. Ballou, W. R., M. Arevalo-Herrera, D. Carucci, T. L. Richie, G. Corradin,
C. Diggs, P. Druilhe, B. K. Giersing, A. Saul, D. G. Heppner, et al. 2004. Updateon the clinical development of candidate malaria vaccines. Am. J. Trop. Med.Hyg. 71: 239–247.
2. Epstein, J. E., B. Giersing, G. Mullen, V. Moorthy, and T. L. Richie. 2007.Malaria vaccines: are we getting closer? Curr. Opin. Mol. Ther. 9: 12–24.
3. Bejon, P., J. Mwacharo, O. Kai, T. Mwangi, P. Milligan, S. Todryk, S. Keating,T. Lang, B. Lowe, C. Gikonyo, et al. 2006. A phase 2b randomised trial of thecandidate malaria vaccines FP9 ME-TRAP and MVA ME-TRAP among childrenin Kenya. PLoS Clin. Trials 1: e29.
4. Dunachie, S. J., M. Walther, J. M. Vuola, D. P. Webster, S. M. Keating,T. Berthoud, L. Andrews, P. Bejon, I. Poulton, G. Butcher, et al. 2006. A clinicaltrial of prime-boost immunisation with the candidate malaria vaccines RTS,S/AS02A and MVA-CS. Vaccine 24: 2850–2859.
5. Moorthy, V. S., E. B. Imoukhuede, P. Milligan, K. Bojang, S. Keating, P. Kaye,M. Pinder, S. C. Gilbert, G. Walraven, B. M. Greenwood, and A. S. Hill. 2004.A randomised, double-blind, controlled vaccine efficacy trial of DNA/MVA ME-TRAP against malaria infection in Gambian adults. PLoS Med. 1: e33.
6. Walther, M., F. M. Thompson, S. Dunachie, S. Keating, S. Todryk, T. Berthoud,L. Andrews, R. F. Andersen, A. Moore, S. C. Gilbert, et al. 2006. Safety, im-munogenicity, and efficacy of prime-boost immunization with recombinant pox-virus FP9 and modified vaccinia virus Ankara encoding the full-length Plasmo-dium falciparum circumsporozoite protein. Infect. Immun. 74: 2706–2716.
7. Plotkin, S. A. 2005. Vaccines: past, present and future. Nat. Med. 11: S5–S11.8. Alonso, P. L., J. Sacarlal, J. J. Aponte, A. Leach, E. Macete, J. Milman,
I. Mandomando, B. Spiessens, C. Guinovart, M. Espasa, et al. 2004. Efficacy ofthe RTS,S/AS02A vaccine against Plasmodium falciparum infection and diseasein young African children: randomised controlled trial. Lancet 364: 1411–1420.
9. Rafi-Janajreh, A., J. E. Tongren, C. Kensil, C. Hackett, F. Candal, A. Lal, andV. Udhayakumar. 2002. Influence of adjuvants in inducing immune responses todifferent epitopes included in a multiepitope, multivalent, multistage Plasmodiumfalciparum candidate vaccine (FALVAC-1) in outbred mice. Exp. Parasitol. 101:3–12.
10. Akagi, T., X. Wang, T. Uto, M. Baba, and M. Akashi. 2007. Protein directdelivery to dendritic cells using nanoparticles based on amphiphilic poly(aminoacid) derivatives. Biomaterials 28: 3427–3436.
11. Kalkanidis, M., G. A. Pietersz, S. D. Xiang, P. L. Mottram, B. Crimeen-Irwin,K. Ardipradja, and M. Plebanski. 2006. Methods for nano-particle based vaccineformulation and evaluation of their immunogenicity. Methods 40: 20–29.
12. Minigo, G., A. Scholzen, C. K. Tang, J. C. Hanley, M. Kalkanidis, G. A. Pietersz,V. Apostolopoulos, and M. Plebanski. 2007. Poly-L-lysine-coated nanoparticles:a potent delivery system to enhance DNA vaccine efficacy. Vaccine 25:1316–1327.
13. Mottram, P. L., D. Leong, B. Crimeen-Irwin, S. Gloster, S. D. Xiang, J. Meanger,R. Ghildyal, N. Vardaxis, and M. Plebanski. 2007. Type 1 and 2 immunity fol-lowing vaccination is influenced by nanoparticle size: formulation of a modelvaccine for respiratory syncytial virus. Mol. Pharm. 4: 73–84.
14. Reddy, S. T., A. Rehor, H. G. Schmoekel, J. A. Hubbell, and M. A. Swartz. 2006.In vivo targeting of dendritic cells in lymph nodes with poly(propylene sulfide)nanoparticles. J. Control Release 112: 26–34.
15. Reddy, S. T., M. A. Swartz, and J. A. Hubbell. 2006. Targeting dendritic cellswith biomaterials: developing the next generation of vaccines. Trends Immunol.27: 573–579.
16. Noad, R., and P. Roy. 2003. Virus-like particles as immunogens. Trends Micro-biol. 11: 438–444.
17. Bachmann, M. F., U. H. Rohrer, T. M. Kundig, K. Burki, H. Hengartner, andR. M. Zinkernagel. 1993. The influence of antigen organization on B cell re-sponsiveness. Science 262: 1448–1451.
18. Bachmann, M. F., and R. M. Zinkernagel. 1996. The influence of virus structureon antibody responses and virus serotype formation. Immunol. Today 17:553–558.
19. Aebi, U., B. ten Heggeler, L. Onorato, J. Kistler, and M. K. Showe. 1977. Newmethod for localizing proteins in periodic structures: Fab fragment labeling com-bined with image processing of electron micrographs. Proc. Natl. Acad. Sci. USA74: 5514–5518.
20. Buhle, E. L., Jr., and U. Aebi. 1984. Specific labeling of protein domains withantibody fragments. J. Ultrastruct. Res. 89: 165–178.
21. Greenwood, J., A. E. Willis, and R. N. Perham. 1991. Multiple display of foreignpeptides on a filamentous bacteriophage: peptides from Plasmodium falciparumcircumsporozoite protein as antigens. J. Mol. Biol. 220: 821–827.
22. Kistler, J., U. Aebi, L. Onorato, B. ten Heggeler, and M. K. Showe. 1978. Struc-tural changes during the transformation of bacteriophage T4 polyheads: charac-terization of the initial and final states by freeze-drying and shadowing Fab-fragment-labelled preparations. J. Mol. Biol. 126: 571–589.
23. Sanchez, J., and J. Holmgren. 1989. Recombinant system for overexpression ofcholera toxin B subunit in Vibrio cholerae as a basis for vaccine development.Proc. Natl. Acad. Sci. USA 86: 481–485.
24. Liu, W., and Y. H. Chen. 2005. High epitope density in a single protein moleculesignificantly enhances antigenicity as well as immunogenicity: a novel strategyfor modern vaccine development and a preliminary investigation about B celldiscrimination of monomeric proteins. Eur. J. Immunol. 35: 505–514.
25. Jennings, G. T., and M. F. Bachmann. 2008. The coming of age of virus-likeparticle vaccines. Biol. Chem. 389: 521–536.
26. Pimentel, T. A., Z. Yan, S. A. Jeffers, K. V. Holmes, R. S. Hodges, andP. Burkhard. 2009. Peptide nanoparticles as novel immunogens: design and anal-ysis of a prototypic severe acute respiratory syndrome vaccine. Chem. Biol. DrugDes. 73: 53–61.
27. Raman, S., G. Machaidze, A. Lustig, U. Aebi, and P. Burkhard. 2006. Structure-based design of peptides that self-assemble into regular polyhedral nanoparticles.Nanomedicine 2: 95–102.
28. Schroeder, U., A. Graff, S. Buchmeier, P. Rigler, U. Silvan, D. Tropel,B. M. Jockusch, U. Aebi, P. Burkhard, and C. A. Schoenenberger. 2009. Peptidenanoparticles serve as a powerful platform for the immunogenic display of poorlyantigenic actin determinants. J. Mol. Biol. 386: 1368–1381.
29. Malashkevich, V. N., R. A. Kammerer, V. P. Efimov, T. Schulthess, and J. Engel.1996. The crystal structure of a five-stranded coiled coil in COMP: a prototypeion channel? Science 274: 761–765.
30. Burkhard, P., M. Meier, and A. Lustig. 2000. Design of a minimal protein oli-gomerization domain by a structural approach. Protein Sci. 9: 2294–2301.
31. Eichinger, D. J., D. E. Arnot, J. P. Tam, V. Nussenzweig, and V. Enea. 1986.Circumsporozoite protein of Plasmodium berghei: gene cloning and identificationof the immunodominant epitopes. Mol. Cell. Biol. 6: 3965–3972.
32. Zavala, F., J. P. Tam, P. J. Barr, P. J. Romero, V. Ley, R. S. Nussenzweig, andV. Nussenzweig. 1987. Synthetic peptide vaccine confers protection against mu-rine malaria. J. Exp. Med. 166: 1591–1596.
33. Brandenberger, R., R. A. Kammerer, J. Engel, and M. Chiquet. 1996. Nativechick laminin-4 containing the �2 chain (s-laminin) promotes motor axongrowth. J. Cell Biol. 135: 1583–1592.
34. Dutta, S., P. V. Lalitha, L. A. Ware, A. Barbosa, J. K. Moch, M. A. Vassell,B. B. Fileta, S. Kitov, N. Kolodny, D. G. Heppner, J. D. Haynes, and D. E. Lanar.2002. Purification, characterization, and immunogenicity of the refolded ectodo-main of the Plasmodium falciparum apical membrane antigen 1 expressed inEscherichia coli. Infect. Immun. 70: 3101–3110.
35. Nussenzweig, R. S., J. Vanderberg, H. Most, and C. Orton. 1967. Protectiveimmunity produced by the injection of x-irradiated sporozoites of Plasmodiumberghei. Nature 216: 160–162.
36. Waldrop, S. L., K. A. Davis, V. C. Maino, and L. J. Picker. 1998. Normal humanCD4� memory T cells display broad heterogeneity in their activation thresholdfor cytokine synthesis. J. Immunol. 161: 5284–5295.
37. Brando, C., L. A. Ware, H. Freyberger, A. Kathcart, A. Barbosa, S. Cayphas,M. A. Demoitie, P. Mettens, D. G. Heppner, and D. E. Lanar. 2007. Murineimmune responses to liver-stage antigen 1 protein FMP011, a malaria vaccinecandidate, delivered with adjuvant AS01B or AS02A. Infect. Immun. 75:838–845.
38. Pullen, G. R., M. G. Fitzgerald, and C. S. Hosking. 1986. Antibody aviditydetermination by ELISA using thiocyanate elution. J. Immunol. Methods 86:83–87.
39. Perciani, C. T., P. S. Peixoto, W. O. Dias, F. S. Kubrusly, and M. M. Tanizaki.2007. Improved method to calculate the antibody avidity index. J. Clin. Lab.Anal. 21: 201–206.
40. Scheiblhofer, S., D. Chen, R. Weiss, F. Khan, S. Mostbock, K. Fegeding,W. W. Leitner, J. Thalhamer, and J. A. Lyon. 2001. Removal of the circum-sporozoite protein (CSP) glycosylphosphatidylinositol signal sequence from aCSP DNA vaccine enhances induction of CSP-specific Th2 type immune re-sponses and improves protection against malaria infection. Eur. J. Immunol. 31:692–698.
41. Miller, S. I., R. K. Ernst, and M. W. Bader. 2005. LPS, TLR4 and infectiousdisease diversity. Nat. Rev. Microbiol. 3: 36–46.
42. Jouvin-Marche, E., M. G. Morgado, C. Leguern, D. Voegtle, F. Bonhomme, andP. A. Cazenave. 1989. The mouse Igh-1a and Igh-1b H chain constant regions arederived from two distinct isotypic genes. Immunogenetics 29: 92–97.
43. Martin, R. M., J. L. Brady, and A. M. Lew. 1998. The need for IgG2c specificantiserum when isotyping antibodies from C57BL/6 and NOD mice. J. Immunol.Methods 212: 187–192.
44. Abdulla, S., R. Oberholzer, O. Juma, S. Kubhoja, F. Machera, C. Membi,S. Omari, A. Urassa, H. Mshinda, A. Jumanne, et al. 2008. Safety and immuno-genicity of RTS,S/AS02D malaria vaccine in infants. N. Eng. J. Med. 359:2533–2544.
45. Dunachie, S. J., M. Walther, J. E. Epstein, S. Keating, T. Berthoud, L. Andrews,R. F. Andersen, P. Bejon, N. Goonetilleke, I. Poulton, et al. 2006. A DNA prime-modified vaccinia virus ankara boost vaccine encoding thrombospondin-relatedadhesion protein but not circumsporozoite protein partially protects healthy ma-laria-naive adults against Plasmodium falciparum sporozoite challenge. Infect.Immun. 74: 5933–5942.
46. Gilbert, S. C., M. Plebanski, S. J. Harris, C. E. Allsopp, R. Thomas, G. T. Layton,and A. V. Hill. 1997. A protein particle vaccine containing multiple malariaepitopes. Nat. Biotechnol. 15: 1280–1284.
47. Li, S., E. Locke, J. Bruder, D. Clarke, D. L. Doolan, M. J. Havenga, A. V. Hill,P. Liljestrom, T. P. Monath, H. Y. Naim, et al. 2007. Viral vectors for malariavaccine development. Vaccine 25: 2567–2574.
48. Moorthy, V. S., E. B. Imoukhuede, S. Keating, M. Pinder, D. Webster,M. A. Skinner, S. C. Gilbert, G. Walraven, and A. V. Hill. 2004. Phase 1 eval-uation of 3 highly immunogenic prime-boost regimens, including a 12-monthreboosting vaccination, for malaria vaccination in Gambian men. J. Infect. Dis.189: 2213–2219.
49. Stoute, J. A., M. Slaoui, D. G. Heppner, P. Momin, K. E. Kester, P. Desmons,B. T. Wellde, N. Garcon, U. Krzych, M. Marchand, et al. 1997. A preliminary
7276 NANOPARTICLE VACCINE PROTECTS AGAINST MALARIA

evaluation of a recombinant circumsporozoite protein vaccine against Plasmo-dium falciparum malaria. N. Eng. J. Med. 336: 86–91.
50. Romero, P. J., J. P. Tam, D. Schlesinger, P. Clavijo, H. Gibson, P. J. Barr,R. S. Nussenzweig, V. Nussenzweig, and F. Zavala. 1988. Multiple T helper cellepitopes of the circumsporozoite protein of Plasmodium berghei. Eur. J. Immu-nol. 18: 1951–1957.
51. Birkett, A., K. Lyons, A. Schmidt, D. Boyd, G. A. Oliveira, A. Siddique,R. Nussenzweig, J. M. Calvo-Calle, and E. Nardin. 2002. A modified hepatitis Bvirus core particle containing multiple epitopes of the Plasmodium falciparumcircumsporozoite protein provides a highly immunogenic malaria vaccine in pre-clinical analyses in rodent and primate hosts. Infect. Immun. 70: 6860–6870.
52. Oliveira, G. A., K. Wetzel, J. M. Calvo-Calle, R. Nussenzweig, A. Schmidt,A. Birkett, F. Dubovsky, E. Tierney, C. H. Gleiter, G. Boehmer, et al. 2005.Safety and enhanced immunogenicity of a hepatitis B core particle Plasmodiumfalciparum malaria vaccine formulated in adjuvant Montanide ISA 720 in a phaseI trial. Infect. Immun. 73: 3587–3597.
53. Ishii, N., F. Fitrilawati, A. Manna, H. Akiyama, Y. Tamada, and K. Tamada.2008. Gold nanoparticles used as a carrier enhance production of anti-hapten IgGin rabbit: a study with azobenzene-dye as a hapten presented on the entire surfaceof gold nanoparticles. Biosci. Biotechnol. Biochem. 72: 124–131.
54. Matsuo, K., T. Yoshikawa, A. Oda, T. Akagi, M. Akashi, Y. Mukai, Y. Yoshioka,N. Okada, and S. Nakagawa. 2007. Efficient generation of antigen-specific cel-lular immunity by vaccination with poly(�-glutamic acid) nanoparticles entrap-ping endoplasmic reticulum-targeted peptides. Biochem. Biophys. Res. Commun.362: 1069–1072.
55. Okamoto, S., H. Yoshii, T. Akagi, M. Akashi, T. Ishikawa, Y. Okuno,M. Takahashi, K. Yamanishi, and Y. Mori. 2007. Influenza hemagglutinin vac-cine with poly(�-glutamic acid) nanoparticles enhances the protection againstinfluenza virus infection through both humoral and cell-mediated immunity. Vac-cine 25: 8270–8278.
56. Okamoto, S., H. Yoshii, T. Ishikawa, T. Akagi, M. Akashi, M. Takahashi,K. Yamanishi, and Y. Mori. 2008. Single dose of inactivated Japanese enceph-alitis vaccine with poly(�-glutamic acid) nanoparticles provides effective protec-tion from Japanese encephalitis virus. Vaccine 26: 589–594.
57. Manolova, V., A. Flace, M. Bauer, K. Schwarz, P. Saudan, and M. F. Bachmann.2008. Nanoparticles target distinct dendritic cell populations according to theirsize. Eur. J. Immunol. 38: 1404–1413.
58. Reddy, S. T., A. J. van der Vlies, E. Simeoni, V. Angeli, G. J. Randolph, C. P.O’Neil, L. K. Lee, M. A. Swartz, and J. A. Hubbell. 2007. Exploiting lymphatic
transport and complement activation in nanoparticle vaccines. Nat. Biotechnol.25: 1159–1164.
59. Ahlborg, N., I. T. Ling, A. A. Holder, and E. M. Riley. 2000. Linkage of exog-enous T-cell epitopes to the 19-kilodalton region of Plasmodium yoelii merozoitesurface protein 1 (MSP1(19)) can enhance protective immunity against malariaand modulate the immunoglobulin subclass response to MSP1(19). Infect. Im-mun. 68: 2102–2109.
60. de Oliveira, G. A., P. Clavijo, R. S. Nussenzweig, and E. H. Nardin. 1994.Immunogenicity of an alum-adsorbed synthetic multiple-antigen peptide based onB- and T-cell epitopes of the Plasmodium falciparum CS protein: possible vac-cine application. Vaccine 12: 1012–1017.
61. Guy, B., T. Krell, V. Sanchez, A. Kennel, C. Manin, and R. Sodoyer. 2005. DoTh1 or Th2 sequence motifs exist in proteins?: identification of amphipatic im-munomodulatory domains in Helicobacter pylori catalase. Immunol. Lett. 96:261–275.
62. Hisaeda, H., K. Yasutomo, and K. Himeno. 2005. Malaria: immune evasion byparasites. Int. J. Biochem. Cell Biol. 37: 700–706.
63. Lee, E. A., D. R. Palmer, K. L. Flanagan, W. H. Reece, K. Odhiambo, K. Marsh,M. Pinder, M. B. Gravenor, W. A. Keitel, K. E. Kester, et al. 2002. Induction ofT helper type 1 and 2 responses to 19-kilodalton merozoite surface protein 1 invaccinated healthy volunteers and adults naturally exposed to malaria. Infect.Immun. 70: 1417–1421.
64. Li, H., S. B. Willingham, J. P. Ting, and F. Re. 2008. Cutting edge: inflamma-some activation by alum and alum’s adjuvant effect are mediated by NLRP3.J. Immunol. 181: 17–21.
65. Livingston, B., C. Crimi, M. Newman, Y. Higashimoto, E. Appella, J. Sidney,and A. Sette. 2002. A rational strategy to design multiepitope immunogens basedon multiple Th lymphocyte epitopes. J. Immunol. 168: 5499–5506.
66. McHeyzer-Williams, L. J., D. J. Driver, and M. G. McHeyzer-Williams. 2001.Germinal center reaction. Curr. Opin. Hematol. 8: 52–59.
67. Siegrist, C. A., M. Pihlgren, C. Tougne, S. M. Efler, M. L. Morris,M. J. AlAdhami, D. W. Cameron, C. L. Cooper, J. Heathcote, H. L. Davis, andP. H. Lambert. 2004. Co-administration of CpG oligonucleotides enhances thelate affinity maturation process of human anti-hepatitis B vaccine response. Vac-cine 23: 615–622.
68. Mettens, P., P. M. Dubois, M. A. Demoitie, B. Bayat, M. N. Donner,P. Bourguignon, V. A. Stewart, D. G. Heppner, Jr., N. Garcon, and J. Cohen.2008. Improved T cell responses to Plasmodium falciparum circumsporozoiteprotein in mice and monkeys induced by a novel formulation of RTS,S vaccineantigen. Vaccine 26: 1072–1082.
7277The Journal of Immunology




















