
Detection of MHC class II transcripts in lymphoid tissues of the common carp ( Cyprinus carpio L.)
Upload
independentCategory
view
0download
0
Journal of Fish Diseases 1996, 19, 199-207
The histopathology of carp, Cyprinus carpio L., exposed tomicrocystins by gavage, immersion and intraperitonealadministration
C R Carbis', G T Rawlin^ G F MitchelP, J W Anderson' and 1 McCauley^
1 Department of Botany, La Trobe University, Bundoora, Australia2 Victorian Institute of Animal Science, Attwood, Australia3 Royal Melbourne Zoological Gardens, Parkville, Victoria, Australia
Abstract
Carp liver, gills, intestine, kidneys, heart andspleen were studied by histology after the fishwere exposed to microcystins by gavage, immersionand intraperitoneal administration, Intraperitonealinoculation with microcystins caused necrosis or dose-dependent degeneration in the liver, gills and kid-neys. Gavaging v ith microcystins caused changes inthe histopathology of the liver and gills. Cellulardegeneration and necrosis occurred in the liver, gillsand kidneys when carp were introduced to a tankcontaining 1-7 fJ% ml"' of microcystins. Lesions werenot observed in the heart, spleen or intestines fromatiy of the treated catp. Microcystins adtninisteredby intraperitoneal injection at a concentration of50 (^ kg"' were lethal to all fish within 8 h, whilegavaging with 250 /^ kg"' of microcystins causedminimal damage in the tissues studied.
Introduction
Cyanobacteria (blue-green algae) frequently dominatethe epilimnion of eutrophic bodies of water duringsummer (Scvrin-Reyssac & Pletikosic 1990). Manyspecies of cyanobacteria produce potent hcpatotox-ins, which are collectively known as microcystins(Skulberg, Carmichaei, Codd & Skulberg 1993).Toxic blooms of cyanobacteria are frequently associa-ted with fish mortalities, and may be caused by citherdirect toxicosis or changes in the water chemistry as
Correspondence DrC. R. Carbis. Department of Botany, LaTrobe University, Plenty Road, Bundoora, Victoria, Australia3083
a consequence of the bloom; for example, oxygendepletion or H^S production when the bloomcollapses (Mackenthun, Herman & Barcsch 1948;Prescott 1948; Schwitnmer & Schwimmer 1968;Watanabe & Oishi 1983; Persson, Sivonen, Keto,Kononen, Niemi & Viljamaa 1984; Eriksson,Meriluoto & Lindhoim 1986; Langdon 1988;Toranzo, Nieto & Barja 1990; Rodger, Turnbull,Edwards & Codd 1994).
In fish, intraperitoneal (i.p.) exposure to micro-cystins causes tissue damage in the liver (Phillips,Roberts, Stewart & Codd 1985; Rabergh, Bylund &Eriksson 1991; Andersen, Luu, Chen, Holmes, Kent,Le Blanc,Taylor & Williams 1993), kidneys (Raberghetal. 1991), and cerebellar and optic neurons (Phillipset al. 1985). Recent studies on the histopathologyof wild populations of trout, Salmo trutta L., exposedto microcystins at Loch Leven, Scotland (Rodgeret al. 1994), and bteam, Abramis brama (L.), androach, Rutilus rutilus (L.), at Lake Langsjon, southwest Finland (Eriksson et al. 1986), indicatethat microcystins may also damage the gillepithelium.
At certain times of the year, cyanobacteria arethe major component of cyprinid diets (Bittetlich1985;Spatatu&Cophen 1985). Rabergh ff /. (1991)have shown that carp are sensitive to microcystirisby intraperitoneal injection. However, cyprinid mor-taliries associated with cyanobacterial blooms areonly reported occasionally (Mackenthun etal. 1948;Schwimmer & Schwimmer 1968; Vtvsson etal 1984;Eriksson et al. 1986). Moriarty &: Moriarty (1973)repotted that the efficient digestion of Microcystisaeruginosa cells by Tilapia nilotica and Haplochrornisnigriptrmisw3.s largely dependenc on the highly acidic
Lrd 199

Journal ot Fish Diseases 1996, 19, 199-207 C R Carbis et al. Microcystin histopathology of carp
environments of their stomachs. Since the entiredigestive tract of carp is netitml or slightly basic witha pH range of6-12-7-72 (Ai-Hussaini i 949), this mayaccount tor the relative rarity of cyprinid deaths.
Experimental poisoning offish with microcystinshas been confined, ahnost exclusively, to rhe perito-neal route (Phillips etal. 1985; Rabergh etal. 1991;Andersen et al. 1993), which provides limited infor-mation regarding piscine toxicosis from M. aeruginosain environments containing a heavy bloom of thisorganism. This papet reports a study of the potentialof peritoneal, oral and branchial routes for the uptakeand toxicit)^ ot microcystins in carp, Cyprinus carpioL. The histopathology of carp that were exposed tosoluble microcysrins by i.p. injection was comparedto those in vt'hich microcystins were administeredin various other forms, including whole cells ofM. aeruginosa. This was done to replicate the type ofexposure that carp might experience under fieldconditions. Toxicity was evaluated by histologicalexamination ot the liver, intestine, kidney, gill, heartand spleen.
Materials and methods
Fish
Mature, healthy carp (approximate weight 3-4 kg)were transported to outdoor pools from LakeWellington, Victoria, Australia, and divided into 12groups of five individuals. The groups were dividedamong three experiments {A, B and C), each con-sisting of four treatments per experiment (i.e. « = 5for each treatment, and n = 20 for each experiment).The fish were subsequently weighed and tagged, sothat precise dose rates could be calculated for eachfish. A separate batch of eight fish, each of similarweight (456+ 12 g), were used for experiment D. Thefish were weighed, tagged and divided into two groupsof four (i.e. n = 4 for each treatment, and n = 8 forthe experiment). Eor experiments A, B and C, thefish were kept outdoors in pools (6500-1 capacity).For the duration of the experiment, the fish weremaintained in water containing 8-5 gl~' of coarseocean salt, which was added to the pools to assistthe carp with osmoregulation and reduce the riskof infection. In experiment D, fish were kept inoutdoor pools for approximately one week, andthen transferred to the 45-1 treatment tanks at thebeginning of the experiment. Exposure to chlorinewas minimized by filling the pools and tanks ar least3 days prior to introducing the fish. All pools and
tanks were fitted with air-stones to ensure an adequatesupply ot oxygen. The carp were ted pelleted foodand given one week to adjust to the experimentalconditions before the trials began.
Experimental treatments
The treatments are summarized in Table L In ex-periment A, various doses of microcystins wereadministered by i.p. injection as a single inoculum,in 0-4 ml of sterile saline (0-9% w/v NaCl), at5 0 / ^ kg"' of body weight (i.p. high), 25 jL% kg"'(i.p. medium), 2 . 5 / ^ kg"' (i.p. low) and O / ^ k g '(i.p. control). High-dose fish were killed 8 h afterinoculation (for ethical reasons); medium dose, lowdose and control fish were killed 4 days afterinoculation.
In experiment B, a single oral dose of microcystinswas administered by gavage in 6 ml of saline, at250 /^ kg"' (oral high), 25 | ^ kg"' (oral medium),2-5 ^ kg"' (oral low) andO jt^ kg"' (oral control). Allfish from experitnent B wete necropsied after 7 days.
In experiment C, microcystins were prepared bydifferent methods, and administered to fish in eachtreatment by gavage at a dose rate of 2-5 fJ% kg"' bodyweight in 6 ml of saline (multiple oral). The treat-ments included microcystins purified by glass columnchromatography (pure multiple oral), microcystinscontained within a matrix of lysed Af. aeruginosa cells(lysed multiple oral), mictocystins contained withina matrix of whole cells (whole multiple oral), and dailygavage with saline (control multiple oral). All fish fromexperiment C were necropsied 16 days after the firstgavage was administered.
In experiment D, carp were exposed to solublemicrocystins in a way that was designed to replicatethe type of exposure that may occur when a bloom ofM. aeruginosa undergoes lysis under field conditions.Two carp wete released into each of two 45-1 tankscontaining microcystins at a concentration of ap-proximately 1-7 /Jg ml"' (immersion), and t\\ o carpwere placed in each of two additional tanks withoutmicrocystins (immersion control). All of the fishin experiment D were necropsied 7 days after theimmersion treatment began.
Microcystins used in the experiment
A cell suspension of M aeruginosa from Lake Mokoanwas concentrated, frozen and thawed three times,and filtered (0-45 /im). The supernatant was passed
Ltd 200

Journal of Fish Diseases 1996, 19, 199-207 C R Carbis ec al. Microcysiiri histopathology of carp
Table 1 Summary of the experimental treatments (A-D). the estimated amount of microcystins adminisrcred
with each trciumcnt and the number offish {n) in e;ich trtatment.
Experiment
AAAA
BBBB
o O
O o
DD
n
5555
5555
5555
44
Treatment
i.p. highLp. mediumi.p. lowi.p. control
oral highoral mediumoral loworal control
whole multiple orallysed multiple oralpure multiple oralcontrol multiple oral
immersioncontrol immersion
Concentration of microcystins
50 /ig kg-'25 ixQ kg-'2-5 /vg kg-'0 A/g kg-'
250 /jg kg-'25 ^g kg-'2-5 ;/g kg-'0 /ig kg-'
2-5 /ig kg-' day2-5/ig kg-'day2-5 ^g kg-' dayO/igkg-'day
1-7Mgml-'0 /jg kg-'
through a glass column using previously describedmethods (Carbis, Simons, Mitchell, Anderson &McCauley 1994) to yield a partially purified extract.Microcystins were identified using a Waters 996photo-diode array detector, and the toral concen-tration was calculated by comparing the peak areas ofthe Lake Mokoan microcystins with that for a givenamount of authentic microcystin-LR (Calbiochem).The partially purified extract produced 11 chroma-tographic peaks, and five of these were identified asmicrocystins on the basis of their spectra.
Ail of the microcystin treatments for experimentsA and B involved the administration of microcystinsin the form of partially purified extract (Table I).This extract was also used as the source of microcystinsfor the pure multiple oral dose in experiment C.For experiment C, the lysed cells used in the multipleoral lysed treatment (Table 1) were obtained by threecycles of freezing and thawing from the same stocko£M. aeruginosa used to prepare the partially purifiedextract. The concentration of microcystins in thispreparation was approximately 1-2 ^g ml"' of sus-pension. The whole cells used for the multiple oralwhole treatment in experiment C were collectedfrom Lake Mokoan immediately prior to commenc-ing the experiment. These cells were not from theM. aeruginosa stock used to prepare the partiallypurified extract, but an analysis o^ an extract o^ thecells by HPLC indicated that the elutioa profile wasqualitatively similar to that of the partially purifiedextract used in experiments A-C. The concentrationof microcystins in the whole cell suspension was
adjusted to approximately 1-2 ml '. In experimentD (immersion), a crude extract of microcystins wasprepared from the same stock of Lake MokoanM. aeruginosa used to prepare the partially purifiedmicrocystin preparation. The crude extract was pre-pared by sonicating M. aeruginosa cells for 4 min, andthen removing the cell debris by centrifugation(1256 g for 30 min) and filtration (0-45 jltn). Theextract of M. aeruginosa was then added to theappropriate tanks to give a final concentrationof 1-7 jUgml"'. The concentration was verified byanalysing the tank water containing the crude toxinpreparation, by HPLC after the tank was filled. Sincethe crude extract also contained the water solublepigment (phycocyanin) from A/, aeruginosa, the waterin the tank treated with crude extract appearedstrongly blue at this stage. After 7 days, 100 ml ofthe tank water was concentrated through a sep-pak(Waters C ^ sep-pak cartridge), using methodsdescribed by Harada, Matsuura & Suzuki (1988),and the eluate was reanalysed for microcystins byHPLC.
Sampling procedures and histology
Cill samples were taken from all fish (withoutanaesthesia) by lifting the operculum with forceps,and cutting the tips of two or three gill filaments withfine scissors (Clark 1990). The sampling process wasrestricted to the filament tips for ethical reasons.Filament samples were Immediately fixed in 10%phosphate-buffered formalin. Cill samples were taken
Ltd 201

Journai ot Fish Diseases 1996, 19, 199-207 C R Carbis et al. Microcystin histopathology of carp
each day trom carp in the immersion treatment(experiment D). In experiments A, B and C, gillsamples were taken Irom live fish from each treatment,without anaesthesia, at the end of the study period.The fish were subsequently killed, and samples ofhepatopancreas, kidney, intestine, heart and spleenwere immediately fixed in 10% phosphate-bufferedformalin, and processed using standard methods.Paraffin-embedded tissue sections (5 /im) were stainedwith haematoxylin and eosin for light microscopy(Disbrey & Rack 1970).
Results
Clinical assessment
Immersion, i.p. and oral exposure to microcystitiscaused a variety of behavioural and physical aber-rations in carp. Compared to controls, swimmingactivity and schooling behaviour were markedlyreduced within 4 h in carp from experiment A (i.p.high and medium) and experiment B (oral high andmedium). Swimming activity and schooling behaviourslowly declined throughout the treatment period inall multiple oral treatments (experiment C).
In contrast, fish immersed in microcystins (exper-iment D) were hyperactive for the First 3 h; they hadfrenetic swimming movements and made severalattempts to jump out of the water. Hyperactivitydeclined within 12 h, after which time the fish beganto gape open-mouthed at the surface. After 30 h, theblue colouration attributable to phycocyanin in thecrude microcystin sample was no longer evident, andthe fish stopped gaping at the surface. For theremainder of the observation period, the behaviourof the carp that were immersed in microcystins(experiment D) was clinically indistinguishable fromthe control fish.
In carp from experiment A (i.p. high and medium)and carp from experiment B (oral high and medium),the opercular beat races became more frequent com-pared to the controls, and movement ofwater throughthe oral cavity was maintained with irregular gapes.Compared to the controls, fish in the i.p. high andmedium dose treatments (experiment A) had highopercular beat rates until the fish were killed. In oralhigh and medium dose treatments (experiment B),the opercular beat rates were similar to those of thecontrol carp after 2-3 days.
In all fish exposed to microcystins, except theimmersion treatment (experiment D), melanodermiawas observed after approximately 24 h. Pigmentation
was dose-related and far more obvious in the high-dose treatments.
Death was imminent when fish dropped their headsbelow horizontal and floated near the surface withthe dorsal section of their caudal fins out of the water.This only occurred in fish from experiment A in thei.p. high dose (after 5 h) and i.p. medium dose (after2 days).
Histopathology of the liver
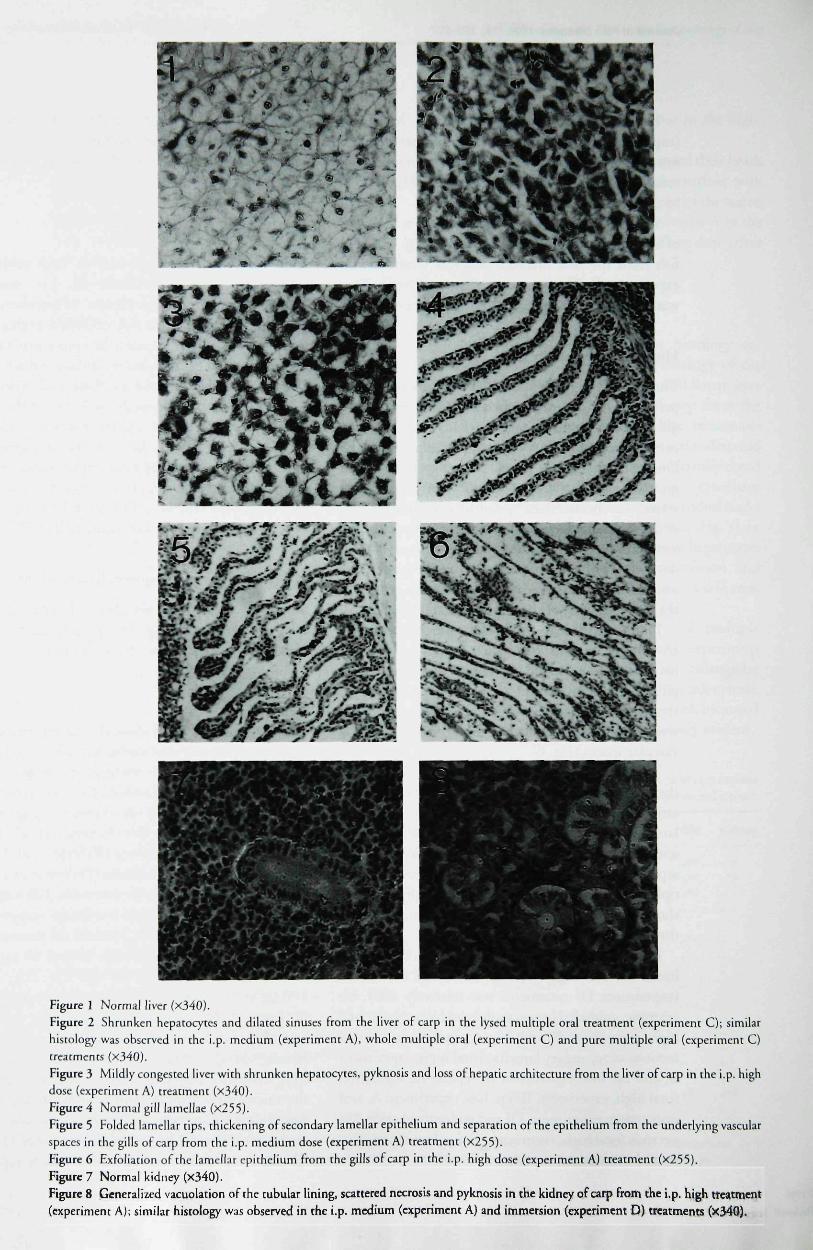
The results of the hepatopancreas histology aresummarized in Table 2. The histopathology of theliver was characterized by dose-dependent hepatocytenecrosis and pyknosis. The livers of carp from thei.p. high (Fig. 3) and medium dose treatments(experiment A) were congested, and showed widespreaddissociation and hepatocyte shrinkage. In lethally dosedcarp from experiment A (i.p. high), the cytoplasmdisintegrated and many cells persisted as individual nucleiattached to fragments of the cell's cytoplasm (Fig. 3). Insublethally poisoned carp, degeneration of hepatocyteswas characterized by cytoplasmic vacuolation andhepatocyte shrinkage, which was more common in areasclose to the arteriaj blood supply (Fig. 2).
The sinuses of carp livers from the whole multipleoral and lysed multiple oral treatments (experimentC) were dilated (Fig. 2), and mild cholangiolathyperplasia was frequently observed, MegaJocytosis,typified by individual large hepatocytes with increasednuclear/cytoplasmic ratio, was occasionally evident.
Table 2 Summary of the histopachology findings in carp exposed
to microcystins by gavage, immersion and intraperitoneal routes*.
Experiment
AAAA
BBB8
CCCC
DD
Treatment
(p. highi.p. mediumi.p. lowi.p. control
oral highoral mediumoral loworal control
whole multiple orallysed multiple oralpure multiple oralcontrol multiple orai
immersioncontrol immersion
Liver
^ A Q . C
. A,B,C
, 6
-
-
-
-
+B,C
+B.C
-
-
Giil
^ B , C
+s,c
+B.C
-
+s,c
--
-
—
-
-—
—
Kidney
+A,D
-
-
—
-
-
-
—
-
-
—
-
* {+") congestion; {widespread necrosis;
loss of structural architecture;focal necrosis; (-) no change.
Ltd 202

Journal of Fish Diseases 1996, 19, 199-207 C R Carbis et al. Microcystin htstopaiholtigy of carp
The livers of carp from the i.p. low dose(experiment A), oral high dose (expeiiment B) andimmersion (experiment D) treatments were charac-terized by cytoplasmic vacuolation and mild shrinkageof the hepatocytes.
Significant lesions were not observed in the liversof control carp from all experiments (Fig. 1), or infish from the oral medium and low treatments inexperiment B. The pancreatic tissue within the liverwas normal in all treatments from ail four experiments-
Histopathology of the gills
The gill histology for each of the treatments for ex-periments A—D is summarized in Table 2. Samples ofgill filament tips were taken at daily intervals fromcarp in both of the treatments in experiment D.During the first 2 days, the gill pathology in the treatedgroup (i.e. 1 -7^ mi ' of microcystins in the tankwater) was characterized by clubbing of distal sectionsofthe secondary lamellae. However, no abnormalitieswere observed during the following 3 days. After 6days, the epithelium separated from the underlyingvascular spaces and epithelial necrosis was evident atthe distal sections ofthe secondary lamellae (Fig. 5).
In the other treatments from the other experiments(A—C), gill sections were taken immediately beforenecropsy. Dose-dependent lesions were seen in thegills. Carp from the i.p. high dose (experiment A)treatment had segments of gill where the epitheliumwas completely exfoliated from the underlyingvascular spaces (Fig. 6).
The histopathology of the branchial tissue fromthe i.p. medium dose (experiment A) carp was charac-terized by progressive thickening of the secondarylamellar epithelium, widespread epithelial necrosisand epithelial cell pyknosis. The epithelium showedseparation from underlying vascular spaces and mul-tiple lamellar telangiectasis. Less severely affected areasshowed folded lamellar tips. Congestion was commonthroughout the gills (Fig. 5).
Branchial damage in carp from the i.p. low (exper-iment A), oral high (experiment B) and immersion(experiment D) treatments was relatively mild; forexample, epithelial hypertrophy and hyperplasia (oralhigh, experiment B), mucus accumulations at distalsections of secondary lamellae (oral high, experimentB; and i.p. low, experiment A), epithelial separation(oral high, experiment B; i.p. low, experiment A; andimmersion, experiment D) and widespread epithelialnecrosis (oral high, experiment B; i.p. low experimentA; and immersion, experiment D).
No significant lesions were observed in branchialtissue from any of the multiple oral treatments(experiment C) or any of the control treatments(experiments A, B, C and D) (Fig. 4).
Histopathology of the kidney
Kidney lesions were seen only in carp subjectedto the i.p. high (experiment A), i.p. medium(experiment A) and immersion (experiment D)treatments (Fig. 8). The fish exhibited evidence oftoxic nephrosis characterized by severe generalizedvacuolation of the tubular lining, which wasassociated with scattered necrosis and pyknosis.These lesions occurred in about 2—5% ofthe renaltubules in affected fish, and thete was no effect onhaematopoietic tissues. Lesions were not observedin the renal tissue of carp from exper-iments A (i.p.low), B (all mictocystin treatments), C (allmicrocystin treatments), D (immersion) or in thecontrol fish for all four experiments (Fig. 7).
Histopathoiogy of the spleen, heart and intestines
Significant lesions were not observed in preparationsfrom the heart, spleen and intestines from any ofthetreatments in experiments A, B, C and D.
Discussion
The results of this study show that carp hepatocytes,branchial epithelial cells, and to a much lesser degree,renal tubular cells are sensitive to microcystins.Previous investigations in which fish were exposed tomicrocystins found evidence of tissue damage in theliver (Phillips et al. 1985; Rabergh et al. 1991;Andersen etal. 1993), kidney (Rabergh etal. 1991),and cerebellar and optic neurons (Phillips etal. 1985).
The LD Q for carp was not determined directly, buti.p. high- and medium-dose treatments (experimentA) indicate that the i.p. LD ^ vaJue for the microcystinsused in this study was between 20 and 50 /Jg kg"'.These results are similar to the i.p. LD (50-100 g kg-') values reported for rats and mice (Codd& Poon 1988), and considerably lower than thei.p. LD^^ for carp (300-550/^/kg"') reported byRabergh et at. (1991). The difference between i.p.LD^^ values for carp may reflect a higher potency ofthe mictocystin sample from Lake Mokoan, whichwas shown to contain five microcystins, comparedto the microcystin-LR used by Rabergh etal. (1991).Alternatively, the lower i.p. LD^^ values reported
ience Ltd 203
m^'i^
Figure I Normal liver fx340).Figure 2 Shrunken hepacocytcs and dilated sinuses from the liver of carp in the lysed multiple oral rreatment (experiment C); similarhistology was observed in the i.p. medium (experiment A), whole multiple oral (experiment C) and pure multiple oral (experiment C)treatments (xiAO).Figure 3 Mildly congested HVCT with shrunken hepatocyre.s, pyknosis and loss of hepatic architecture from the liver of carp in the i.p. highdose (experiment A) rreatment (x340).Figure 4 Normal gill lamellae (x255).Figure 5 Folded lamellar tips, thickening of secondary lamellar epithelium and separation of the epithelium from the underlying vascularspaces in the gills of carp from the i.p. medium dose (experiment A) treatment (X253).Figure 6 Exfoliation of the lamellar epirheiium from the gills of carp in the i.p. high dose (experiment A) treatment (X255).Figure 7 Normal kidney (x340).Figure 8 Generalized vacuolation of rhe tubular lining, scattered necrosis and pyknosis in the kidney of carp from the i.p. high treatment(experiment A); similar hisrology was observed in the i.p. medium (experiment A) and immersion (experiment D) treatments (X34Q).

Journal of Fish Diseases 1996, 19, 199-207 C R Carbis et al. Microcystin hiitopathology of carp
Ltd
in this study may result from differences in thenutritional and physiological state of the ftsh (RaberghetaL 1991).
Xenobiotics that enter the body through che oralroute are transported directly to the Hver (Frimmer& Zieglet 1988). Previous studies have shown thatthe liver is the most sensitive organ to microcystins(Phillips et al. 1985; Rabergh et aL 1991). In thecurrent study, the fish from the oral medium-(experimervt B) and the oral low-dose (experimentB) treatments, and the control fish from experimentsA, B, C and D were the only carp that had normallivers. In most of the fish from the other treatments(Table 2), microcystins caused histological changesin the liver, which confirms the earlier findings ofPhillips et al. (1985) and Rabergh et aL (1991).However, carp subjected to the i.p. high (experimentA), and, to a lesser degree, i.p. medium (experimentA) treatments had dark friable livers in which thesinusoids were congested, and blood had infiltratedthe liver parenchyma (Fig. 3). The livers of thesefish were similar to those of mammals suppliedwith microcystins, in which massive intrahepatichaemorrhaging is followed by haemodynamic shock(Falconer, Jackson, Langley & Runnegar 1981).
Toxins which enter the liver via the hepatic portalvein are removed by a process known as presystemichepatic elimination, which eliminates toxins duringa single pass through the liver under optimal con-ditions. This process prevents, or at least minimizes,the distribution of foreign chemicals to other partsof the body (Klaassen & Watkins 1984). However,other organs or tissues may be exposed to microcystinswhen the process of presystemic hepatic eliminationis overwhelmed or bypassed by alternative routes.The results of the curtent study indicate that thehepatopancreas of carp was not readily overwhelmed,since the only gavage treatment in which microcystinsentered the systemic circulation in sufficiently highconcentrations to cause branchial injury was the oralhigh dose (experiment B).
Branchial injury was more severe when theperitoneal route was used to administer microcystins,particularly in i.p. high (experiment A) and i.p.medium (experiment A) treatments. When toxinis injected into the peritoneal cavity, it enters thelymphatic system and is directed to the heart via theposterior cardinal vein (Clark 1990). The blood is thenpumped directly to the gills for the uptake of oxygen,so the concentration of microcystins is not reducedby presystemic hepatic elimination. Therefore,microcystins are far more accessible to the gillepithelium via the peritoneal route.
205
Branchial damage also occurred when carp wereimmersed in a toxic cell-free lysate of M aeruginosacontaining soluble microcystins (experiment D).Phycocyanin was present as a visible blue solute forapproximately 2 days, and may have been responsiblefor epithelial damage during this period (i.e. clubbingof secondary lamellae). After 2 days, the blue colourwas no longer visible, and the branchial epitheliumappeared normal for the following 3 days. However,widespread epithelial necrosis occurred after 6 days,and eill lesions in these fish were similar to i.p.!ow-dose (experiment A) carp. It seems unlikelythat these lesions ate attributable to phycocyanin.Rather, the delayed branchial injury is attributableto microcystins, which do not readily infiltrate thegill epithelium from the water. These results are con-sistent with the possibility chat delayed branchialdamage occurs after microcystins are taken up by theintestines. If the respiratory function of carp iscompromised by exposure to microcystins when abloom collapses, then the increase in bacterial res-piration that occurs when the cyanobacterial biomassis biodegraded may cause hypoxia and increase themortality rate.
Microcystins are eventually removed from thegeneral circulation by renal or subsequent hepaticelimination. Toxic renal nephrosis was observed incarp exposed to microcystins by i.p. high (exper-iment A), medium (experiment A) and immersion(experiment D) routes, which is similar to the fmdingsof Rabergh et aL (1991). The kidneys are potentiallyexposed to greater concentrations of microcystinswhen the toxins are not subjected to presystemichepatic elimination. This may occur when micro-cystins enter the body by intraperitoneal injection,or if water soluble microcystins enter the bodydirectly through branchial, dermal or conjunctivalroutes.
Microcystins did not appear to have a significanteffect on heart, spleen or intestines, which is similarto the findings of Phillips et aL (1985) and RaberghetaL (1991).
Carp will be exposed to microcystins via the oralroute when they ingest M. aeruginosa cells. Microcystisis an major component of cyptinid diets and is utilizedas the principle source of food when Microcystisdominates the phytoplankton community (Bitterlich1985; Spataru & Gophen 1985). However, grazingrates are apparently suppressed when cyanobacteriaproduce hepatotoxins (Beveridge, Baitd, Rahmatullah,Lawton, Beattie & Codd 1993). In addition,M. aeruginosa cells are not readily digested by cyprinids,so most of the toxin may be contained within

Journal of Fish Diseases 1996, 19, 199-207 C R Carbis et al. Microcystin histopathology of carp
Microcy'stis cells as they pass through the carp intestine(Bitterlich 1985).
The results of experiment A (i.e. microcystinsadministered by i.p. injection) indicate that thehepatopancreas, gills and kidneys of carp are highlysusceptible to microcystins, but the experimentsdesigned to simulate the type of exposure that carpmight experience under field conditions indicate thatlethal toxicoses are unlikely to occur.
It is difficult to estimate the volume o^ M.aeruginosa cells that might be ingested by carp underfield conditions. However, the whole multiple oraltreatment (experiment C), in which the maximumpossible volume of A/, aeruginosa cells were repeatedlyadministered by gavage, indicate that lethal toxicosesare unlikely to occur under field conditions, even ifM. aeruginosa is a major component of their diet.Nonetheless, the histological evidence of hepaticinjury in carp that were repeatedly gavaged with toxicM. aeruginosa cells, albeit relatively insignificantby comparison with the histological evidence ofhepatic injury in carp that were exposed to micro-cystins by i.p. injection, is consistent with the pos-sibility that microcystins could accumulate in thehepatopancreas of sublethally poisoned carp undernatural conditions.
The histology of tissues from carp that wereimmersed in tanks of water that contained dissolvedmicrocystins at a concentration of 1 -7 jWg ml ' (treat-ment D) were relatively insignificant by comparisonwith the carp that were exposed to microcystins byi.p. injection. Theoretical calculations, based on thedata of Jones, Falconer & Wilkins (1995) and Jackson,Mclnnes, Falconer & Runnegar (1984), indicate thatthe concentration of microcystins supplied inexperiment D (1-7 ^g ml"') was relatively small incomparison to the maximum concentrations thatmight be attained under field conditions. Jones etal.(1995) showed that each gram of dried M. aeruginosascum from Lake Mokoan produced 3-5 mg of micro-cystins, and Jackson etal. (1984) reported that 650 mlof surface scum, as it occurred at Sunnyside Dam,Australia, produced 30 g of dried scum material. Thus,if ail the cells lysed simultaneously the water wouldcontain up to 175 /Jg ml'' of microcystins. Since theconcentration of microcystins used in experiment Dwas approximately 100-fold less than fish are likelyto experience under field conditions, it is possible thatcarp will be exposed to a lethal toxic dose when thecyanobacterial bloom collapses.
The cyclic pentapeptidcs (nodularins) produced byNodularia spumigena (a cyanobacterium generally
found in brackish water) exhibit similar hepatotoxicactivity to the mictocystins (Falconer 1993). However,the effects of soluble microcystins on carp in freshwater, as described in this paper, may not becomparable to the effects of nodularins on fish undersaline conditions, since marine and estuarine fish,unlike freshwater fish, drink large amounts of waterto maintain hydration. Therefore, marine andestuarine fish may be exposed to extremely largeamounts of soluble nodularins by ingestion when thecells have lysed (Munday 1990). Thus, the impact ofnodularins on marine and esruarine fish might begreater than the impact of microcystins on freshwaterfish.
Acknowledgments
We would like to thank Keith Bell (commercialfisherman at Lake Wellington) and LachlanMcKinnon (Kyalla Fisheries Research Station,Department of Conservation and Natural Resources)for supplying carp, and Peter Bury, Phillip Jackson,Donna Waldron and Dr John Humphrey forassistance. This project was funded by Tbe VictorianInstitute of Animal Science, LaTrobe University andThe Royal Melbourne Zoological Gardens. Theseexperiments were performed with approval of theAnimal Ethics Committee at the Victorian Instituteof Animal Science (protocol numbers 1032 and1238).
References
Al-Hussaini A. H. (1949) On the functional morphology ofthealimentary tract of some fish in relation to theit feedinghabits: cytology and physiology. Quarterly Journal ofMicroscopical Science 90, 323-354.
Andersen R. J., Luu H. A., Chen D. Z. X., Holmes C. F. B.,Kent M. L, Le Blanc M., Taylot F. J. R. M. & Williams D. E.(1993) Chemical and biological evidence links mjcrocystins tosalmon netpen liver disease. Toxicon 31. 1315-1323.
Beveridge M. C. M., Baird D. J., Rahmarullah S. M., LawtonL. A., Beattie K. A. & Codd C. A. (1993) Crazing rates ontoxic and non-toxic strains of cyanobacceria byHypophthdlmichthyi motitrix and Qreochromis niloticus. Joumaiof Fish Biology AX 901-907.
Bitterlich G. (1985) The nutrition of stomachlessphytoplanktivorous fish in compatison with tilapia.Hydrobiologia 121, 173-179.
Carbis C. R., Simons J. A., Mitchell C. R, Anderson J. W. &McCauley I. (1994) A biochemical profile in sheep forpredicting the chronic exposure of sheep to Microcystisaeriigitiosa, an hepatotoxic species oi blue-green alga. Researchin Veterinary Science 57, 310-316.
: Ltd 206

Journal ot Fish Diseases 1996, 19, 199-207 C R Carbis et al. Microcystin hhiopuihohgy of carp
Clark A. (1990) Gross examination in fish health work. In: Fin
Fish Diseases (cd. by Post Craduate Committee in Veterinary
Science), pp. 9-45. Post Graduate Conimirtee in Veterinary
Science. University of Sydney.
Codd G. A. & Poon G. K. (1988) Cyanobacterial toxins. In:
Biochemistry of the Algae and Cyanobacteria (ed. by L. J. Rogers
& J. R. Gallon), pp. 283-296. Oxford Science Publications,
Oxford.
Disbrey B. D. & Rack J. H. (1970) Curtis's picro-ponceau stainfor collagen and Mayers haematoxylin and eosin stain. In:Histological Laboratory Methods, pp. 93-110. E & SLivingstone, London.
Eriksson J. E., Medluoto J. A. O. & LindholmT. (1986) Cancyanobacterial toxins accumulate in aquatic food chains. In:Perspectives in Microbial Ecology, Proceedings of the 4thInternational Symposium on Microbial Ecology (ed. by F.Megusar & M. Gantar), pp. 655-658. Slovene Society forMicrobiology, Ljubliana.
Falconer I. R. (1993) Mechanism of toxicity of cyclic peptidetoxins from the blue-green algae. In: Algal Toxins in Seafoodand Drinking Water (ed. by I. R. Falconer), pp. 165-176.Academic Press, London.
Falconer I. R., Jackson A. R. B., Langley J. & Runnegar M. T. C.(1981) Liver pathology in mice in poisoning by the blue-greenalga Microcystis aeruginosa. Australian Journal of BiologicalScience 34, 179-187.
Frimmer M. & Ziegler K. (1988) The transport of bile acids in
liver cells. Biochimica et Biophysica Acta 947, 75-99.
Harada K., Matsuura K. & Suzuki M. (1988) Analysis and
purification of toxic peptides from cyanobacteria hy reverscd-
phase high-performance liquid chTormto^x^phy. Journal of
Chromatography 488, 275-283.
Jackson A. R. B., Mclnnes A., Falconet L R. &; Runnegar M. T.C. (1984) Clinical and pathological changes in sheepexperimentally poisoned by the blue-green alga Microcystisaeruginosa. Veterinary Pathology 2\, 102-113.
Jones G. J., Falconer L R. & Wiikins R. M, (1995) Persistence ofcyclic peptide toxins m dried Microcystis aeruginosa crustsfrom Lake Mokoan, Australia. Environmental Toxicology andWater Quality 10, 9-24.
Klaassen C D. &: Watkins J. B. (1984) Mechanisms of bileformation, hepatic uptake, and biliary excretion.
Pharmacological Reviews 36, \—67.
Langdon J. S. (1988) Investigation offish kills. In: Fish Diseases
(ed. by Post Graduate Committee in Veterinary Science), pp.
167-223. Post Graduate Committee in Veterinary Science,
University of Sydney.
MackentKun K. M., Herman E. E & Bartsch A. E (1948) A
heavy mortality of fishes resulting from the decomposition of
algae in the Yahara River, Wisconsin. Transactions of the
American Fisheries Society 75, 175-180,
Moriarty D. J. W. & Moriarty C. M. (1973) Quantitative
estimation of the daily ingestion rate of phytoplankton by
Tilapia nilotica and Haplochromis nigripinnis in Lake George,
Vga^ndi. Journal of Zoology (London) 17L 41-55-
Munday B. L. (1990) Fish physiology. In: Fin Fish Diseases:
Rejresher Course for Veterinarians Proceedings 128 (ed. by Post
Graduate Committee in Veterinary Science), pp. 109-134.
University of Sydney.
Persson P £., Sivonen K., Keto J., Kononen K., Niemi M. &
Viljamaa H. (1984) Potentially toxic blue-green algae
(cyanobacteria) in Finnish natural waters. A/^ua Fennica 14,
147-154.
Phillips M. ] . , Roberts R. J., Stewart ] . A. & Codd G. A. (1985)
The toxicity of the cyanobacterlum Microcystis aeruginosa to
rainbow trout, Salmo gairdneri Richardson. Joumal of Fish
Diseases 8, 339-334.
Prescott G. W. (1948) Objectionable algae with reference to the
killing offish and other animals. Hydrobiologia 1, 1-13.
Rabergh C M. L, Bylund G. & Eriksson J. E. (1991)Histopathological effects of microcyscin-LR, a cyclic peptide
toxin from rhe cyanobacterium (blue-green alga) Microcystis
aeruginosa, on common catp. Aquatic Toxicology 20, 1 3 1 -
146.
Rodger H. D., Turnbuli T , Edwards C. & C o d d G . A. (1994)
Cyanobacterial (blue-green algal) bloom associated pathology
in brown trout, Salmo trutta L., in Loch Leven, Scotland.
Joumal of Fish Diseases 17. 177-181.
Schwimmer M. &c Schwimmer D. (1968) Medical aspects ofphycology. In: Algae, Man and the Environment (ed. by D. F.Jackson), pp. 279-358. Syracuse University Press. New York,NY.
Sevrin-Reyssac J. & Pletikosic M. (1990) Cyanobacteria in fishpoaAs. A(juaculture%?>, 1-20.
Skulberg O. M., Carmichael W W., Codd G. A. & Skulberg R.(1993) Taxonomy of toxic cyanophyceae. \T\: Algal Toxins inSeajhod and Drinking Water {cd. by I. R. Falconer), pp. 145-164. Academic Press, London.
Spataru P & Gophen M. (1985) Feeding behaviour of silver carp
Hypophthalmichthys molitrixV^. and its impact on the food
web of Lake Kinneret, Israel. Hydrobiologia 120, 53-61.
Toranzo A. E.. Nieto F. &Bar)aJ. L. (1990) Mortality associatedwith q'anobactetiai bloom in farmed rainbow trout Jn Galicia(North-Western Spain). Bulletin of the European Association ofFish Pathologists 10, 106-107.
Watanabe M. E & Oishi S. (1983) A highly toxic strain of the
blue-green alga Microcystis aeruginosa isolated from Lake
Suwz. Japanese Society for Scientific Fisheries 49, 1759-1765.
Ltd 207

Copyright © 2022 FDOKUMEN




















