
Intraperitoneal administration and other modifications of the 2-deoxy-d-glucose technique
10
Brain Research, 195 (1980) 167-176 167 © Elsevier/North-HollandBiomedicalPress INTRAPERITONEAL ADMINISTRATION AND OTHER MODIFICATIONS OF THE 2-DEOXY-D-GLUCOSE TECHNIQUE RICHARD C. MEIBACH, STANLEY D. GLICK, DAVID A. ROSS, RUSSELL D. COX and SAUL MAAYANI Department of Pharmacology, Mount Sinai School of Medicine of the City University of New York, New York, N. Y. 10029 (U.S.A.) (Accepted February 6th, 1980) Key words: 2-deoxyglucose technique - - functional neuroanatomy SUMMARY Experiments were conducted to determine the optimal conditions necessary for implementing modifications of the 2-deoxyglucose (2-DDG) technique. Substitution of tritium-labeled 2-DDG with subsequent microdissection of selected brain regions and liquid scintillation counting produced results that were highly correlated with both [14C]radioautograms and glucose utilization values as obtained by Sokoloff et al. The route of administration of isotope was also varied. Whole brain uptake at maximal levels of incorporation was the same for both intravenously and intraperitoneally injected animals. Radioautograms from i.p. and i.v. injected animals were indistin- quishable. Densitometric analyses of the i.p. radioautograms were highly correlated with glucose utilization values. Thus, relative indices of functional activity may be obtained when experimental circumstances preclude arteriovenous cannulations and restraint. The use of naive, unrestrained animals, therefore, makes the 2-DDG technique applicable to a broader range of studies. INTRODUCTION The [14C]deoxyglucose technique as developed by Sokoloff et al. 21-23 has become widely accepted among neuroscientists. Dramatic changes in glucose utilization in anatomically and electrophysiologically defined areas following either olfactory stimulation 20, peripheral nerve stimulation 9, or visual deprivation 9 have clearly established the close relationship between functional activity and energy metabolism. Thus, over the past two years we have witnessed a rapid proliferation of studies utilizing the technique1-4,6-s,10-z9,24, 26.
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Intraperitoneal administration and other modifications of the 2-deoxy-d-glucose technique

Brain Research, 195 (1980) 167-176 167 © Elsevier/North-Holland Biomedical Press
INTRAPERITONEAL ADMINISTRATION AND OTHER MODIFICATIONS OF THE 2-DEOXY-D-GLUCOSE TECHNIQUE
RICHARD C. MEIBACH, STANLEY D. GLICK, DAVID A. ROSS, RUSSELL D. COX and SAUL MAAYANI
Department o f Pharmacology, Mount Sinai School of Medicine of the City University of New York, New York, N. Y. 10029 (U.S.A.)
(Accepted February 6th, 1980)
Key words: 2-deoxyglucose technique -- functional neuroanatomy
SUMMARY
Experiments were conducted to determine the optimal conditions necessary for implementing modifications of the 2-deoxyglucose (2-DDG) technique. Substitution of tritium-labeled 2-DDG with subsequent microdissection of selected brain regions and liquid scintillation counting produced results that were highly correlated with both [14C]radioautograms and glucose utilization values as obtained by Sokoloff et al. The route of administration of isotope was also varied. Whole brain uptake at maximal levels of incorporation was the same for both intravenously and intraperitoneally injected animals. Radioautograms from i.p. and i.v. injected animals were indistin- quishable. Densitometric analyses of the i.p. radioautograms were highly correlated with glucose utilization values. Thus, relative indices of functional activity may be obtained when experimental circumstances preclude arteriovenous cannulations and restraint. The use of naive, unrestrained animals, therefore, makes the 2-DDG technique applicable to a broader range of studies.
INTRODUCTION
The [14C]deoxyglucose technique as developed by Sokoloff et al. 21-23 has become widely accepted among neuroscientists. Dramatic changes in glucose utilization in anatomically and electrophysiologically defined areas following either olfactory stimulation 20, peripheral nerve stimulation 9, or visual deprivation 9 have clearly established the close relationship between functional activity and energy metabolism. Thus, over the past two years we have witnessed a rapid proliferation of studies utilizing the technique1-4,6-s,10-z9,24, 26.

168
The technique, as originally described, involves the determination of brain glucose utilization according to an operational equation z2. This requires that for each experiment plasma [14C]deoxyglucose levels, plasma glucose levels, brain deoxyglu- cose-6-phosphate, and rate constants (rate of transport of 2-deoxyglucose from plasma to tissue and back, and the rate of phosphorylation within the tissue) must be determined. The preparation of radioautograms was developed for the purpose of assaying deoxyglucose-6-phosphate within the brain. The radioautograms alone often presented striking evidence of differences in deoxyglucose-6-phosphate concentrations within the brain. Since determination of brain glucose utilization requires exhaustive procedures, modifications of the procedure have appeared which attempt quantifi- cation in lieu of the operational equation. In studies in which [14C]deoxyglucose radioautograms were presented, relative optical densities from experimental and control animals were compared2,1ms, 2°. In other studies, [14C]deoxyglucose had been substituted with [aH]deoxyglucose and brain regions were dissected and counted in a liquid scintillation counter 4,6,13. Further, modifications employing different routes of administration of the label have also been reported4,13. However, there is presently no data available regarding the effects of these modifications on the experimental outcome of the technique. Therefore, the present study examines in detail the validity of these modifications and presents evidence that extends the applicability of the 2- deoxyglucose technique.
MATERIALS AND METHODS
Female Sprague-Dawley strain rats, approximately 3 months old, were used in all experiments. [l,2-aH]2-deoxy-D-glucose (specific activity 40.0 Ci/mmol), [1-14C]2 - deoxy-D-glucose (specific activity activity 53.0 mCi/mmol), and [l-a4C]2-deoxygluco - se-6-phosphate, disodium salt (specific activity 52.2 mCi/mmol) were obtained from New England Nuclear, Boston, Mass. [14C]Polymethylmethacrylate standards were obtained from Amersham. All animals unless noted received intravenous injections of 2-deoxyglucose (2-DDG) through chronically implanted cannula placed into the internal jugular vein 2~.
To determine whether incorporation of 2-DDG is independent of isotope, animals were injected with a combination of 25 #Ci of [14C]2DDG and 50 #Ci of [3H]2DDG. Thirty minutes later animals were sacrificed by decapitation, their brains removed and dissected into 12 regions. Each region was homogenized in 1.0 ml 0.01 M sodium phosphate buffer, pH 7.4. Four aliquots (each 100 #1) were removed and counted in a Beckman LS9000 liquid scintillation counter programmed for both 14C and 3H with automatic quench correction using NEN formula 950A liquid scintil- lation cocktail.
In another series of experiments the distribution of [aH]2DDG-6-P as deter- mined by liquid scintillation counting of dissected specimens was compared with the distribution of [14C]2DDG-6-P seen in radioautograms. Fifty #Ci of [3H]2DDG was injected into rats and processed as detailed above. Another group of animals received injections of 25 #Ci [14C]2DDG. The animals were sacrificed by decapitation after 30

169
min, their brains quickly removed, cooled in liquid freon (--25 °C) and mounted onto a cryostat chuck with Lipshaw embedding medium. Sections of 36 #m were cut at - -26 °C and thaw-mounted onto 25 mm square coverglass. The sections were dried on a slide warmer for 5-10 minutes at 60 °C before being glued to a 10 in. × 14 in. sheet of cardboard. When all of the sections had been collected a set of deoxyglucose standards was added and the coverglass covered cardboard sheet was apposed to Kodak SB-5 X- ray film and exposed for 7 days. The X-ray film was developed for 5 min in Kodak X- ray developer at 27 °C, fixed in Kodak fixer with hardener and washed for 30 min. The X-ray radioautograms were then assayed for 2-DDG-6-P in discrete areas of brain using a Photovolt model 520A densitometer, fitted with a 0.25 mm aperture.
The route of administration of the labeled 2-DDG was also examined in an attempt to determine whether the method could be used with intraperitoneal (i.p.) injection. The first experiments compared the whole brain uptake of [3H]2-DDG after i.p. and i.v. injections. [3H]2-DDG in 0.5 ml saline was injected (at a dose of 200 /~Ci/kg) i.p. and the animals were decapitated I, 2.5, 5, 10, 20, 30 and 45 min later. Control animals receiving the same dose of [SH]2-DDG through internal jugular vein cannula were also sacrificed along the same time intervals. An index of brain uptake (dpm isotope in whole brain/dpm of injected isotope × 100) was determined in all animals. In an additional group of three animals sacrificed at 30 min, the brains were dissected into 12 regions and these were compared to those of i.v. injected animals.
The use of i.p. injection of labeled isotope was also tested with [14C]2-DDG. Three animals were injected with 25/~Ci [14C]2DDG i.p. and decapitated 30 min later. The brains were removed and processed for radioautography as detailed above. Densitometric readings were made from selected regions throughout the brain and the amount of label was compared to that of i.v. injected animals. The densitometric values obtained for each structure were divided by the optical density of the corpus callosum to obtain a grey/white matter ratio. These ratios were determined for both i.p. and i.v. injected animals and compared.
The basic assumption which permits correlation of 2-DDG uptake with functional activity rests on the enzymatic conversion by hexokinase of 2-DDG to 2- DDG-6-P. The latter metabolite, by virtue of its polarity remains essentially trapped within the neuron. We therefore deemed it important to determine the exact ratio of 2- D D G to 2-DDG-6-P. This was accomplished using a slight modification of the procedure described for i.v. vs i.p. administration. Three rats were injected i.p. with [3H]2DDG and at 30 min post-injection were sacrificed by decapitation. Their brains were removed and homogenized in 5 vols of a 5 ~ TCA solution in 0.01 M sodium phosphate buffer and the homogenates spun for 10 min at 5000 rpm on a small bench centrifuge. The clear acidic supernatant was poured off and neutralized to pH 7.2 with 1 N NaOH.
The assay, a modification'of Diamond and Fishman 5, was standardized by preparing 0.5 × 6.0 cm columns of Dowex AGIX 8 100-200 mesh (C1- form) in water which were washed with two volumes of water prior to the addition of approximately 104 dpm [aH]2-DDG and/or [14C]2-DDG-6-P. The D D G was eluted with 5 ml of water in 0.5 ml fractions collected separately in vials for counting. The 2-DDG-6-P was

170
collected identically except 1.5 N HC1 was used to elute the polar 2 -DDG-6-P . To
ensure that the TCA, N a O H or substituents o f the supernatant had no effect on the
elution profile o f 2 - D D G and 2-DDG-6-P , two subsequent control experiments were
carried out adding, first, the 5 ~ T C A to the system (with appropr ia te N a O H ) and
then a cold homogena te supernatant , to which a s tandard amoun t o f dpms were
added.
Fol lowing this s tandardizat ion procedure, supernatants f rom brains o f experi-
mental animals were spiked with 104 dpm of [14C]2-DDG and 104 dpm of [14C]2-
D D G - 6 - P as internal standards. One hundred microl i ter al iquots of the supernatant
were counted in triplicate for the subsequent determinat ion o f co lumn recovery which
ranged between 90 and 100 ~ . After addi t ion o f 1 ml of the supernatant, the columns
were eluted exactly as control columns with 0.5 ml fractions collected and counted.
RESULTS
Substitution o f [14C]2-DDG with tritium
In two animals which received i.v. injections of both al l - and 14C-labeled D D G
TABLE I
Regional comparisons of 2-deoxyglucose uptake with glucose utilization values
14 C a H
Glucose i.v.** i.p.** i.v.*** i.p.*** utilization*
Inferior colliculus 4.70 Cingulate gyrus Auditory cortex 3.73 Thalamus 2.45 Caudoputamen 2.64 Frontal cortex Retrosplenial cortex Hypothalamus 1.50 Hippocampus 1.88 Cerebellum 1.57 ZH-i.p./ZH-i.v. r -- 0.99 3H-i.p./14C-i.p. r -- 0.97 3H-i.v./14C-i.v. r -- 0.95 3H-i.v.#4C-GU§ r 0.93 14C-i.v./laC-i.p. r -- 0.93 14C-GU#aC-i.v. r - - 0.92 14C-GU/14C-i.p. r -- 0.95
2.83 3.25 89.2 - - 2.48 3.21 77.5 80.8 2.52 2.58 - - - - 2.42 2.58 70.8 74.1 2.06 2.29 64.8 64.2 2.16 2.29 62.6 65.1 2.06 2.58 60.0 - - 1.54 1.75 55.4 54.3 1.67 1.92 47.8 50.7 1.38 1.56 50.2 48.3
P < 0.001 P < 0.001 P < 0.001 P < 0.01 P < 0.001 P < 0.01 P < 0.01
* The values expressed are grey/white matter ratios obtained by dividing the glucose utilization (in/~mol/100 g/min) for each structure by the glucose utilization of the corpus callosum as de- termined by Sokoloff et al. 22.
* * The values expressed are grey/white matter ratios determined by dividing the densitometric value of each structure by the densitometric value of the corpus callosum.
* * * The values expressed are the dpm/mg. § 14C-GU = glucose utilization values as determined in *.

171
100'
75'
~ 5 0 '
25
~'o :~o ~o .;o ~o TIME (rain.)
Fig. 1. Graph plotting ratio of uptake of intraperitoneally injected animals to intravenously injected animals with increasing time.
we found an extremely high correlation among 32 individually dissected brain regions (r = 0.9954, P < 0.001). Therefore either isotope can be used depending on the needs of the investigator. Six out of the 12 areas which were included in our microdissections were anatomically matched to areas for which Sokoloff et al. z2 obtained glucose utilization values. Therefore, only these regions were used for comparing our data with theirs. These included the inferior colliculus, thalamus, caudoputamen, hypothalamus, hippocampus and cerebellum. Although the possibility for error appeared large because our microdissections included an entire region while Sokoloff's glucose utilization values were obtained from pinpoint densitometric readings we found that our values were highly correlated with theirs (r = 0.93, P < 0.01) (Table I). From densitometric values obtained from our own radioautograms in which we carefully selected the regions to include all the areas that were microdissected we were able to anatomically compare 9 regions. From this larger group we found an even greater degree of correlation (r = 0.95, P < 0.001). Comparison of 7 areas taken from our [14C]2-DDG radioautograms with Sokoloff's values for glucose utilization obtained from [14C]2-DDG radioautograms were also highly correlated (r = 0.92, P < 0.01).
.40-
~.20-
.10=
:'o 1"5 ~o 2"5 ~o ~s ;o ;s TIME (rain)
Fig. 2. Graph plotting uptake of [3H]2-deoxyglucose following intraperitoneal administration in whole brain with increasing time.
172
Route of administration of 2-DDG In 3 animals which had received an i.p. injection of [zI-t]2-DDG and were
sacrificed 30 min later, we found a correlation of 0,99 to that of i.v. administered [3H]2-DDG. At 30 min either route of administration yielded equal uptake (0.23 ~) and the ratio of i.p./i.v, became 1 (Fig. 1). I.v. injections attained maximal absorption in less than 10 min, however it took 30 min for the i.p. injections to reach the same plateau (Fig. 2). Longer survival times (45 rain) showed no further increase or decrease in 2-DDG levels.
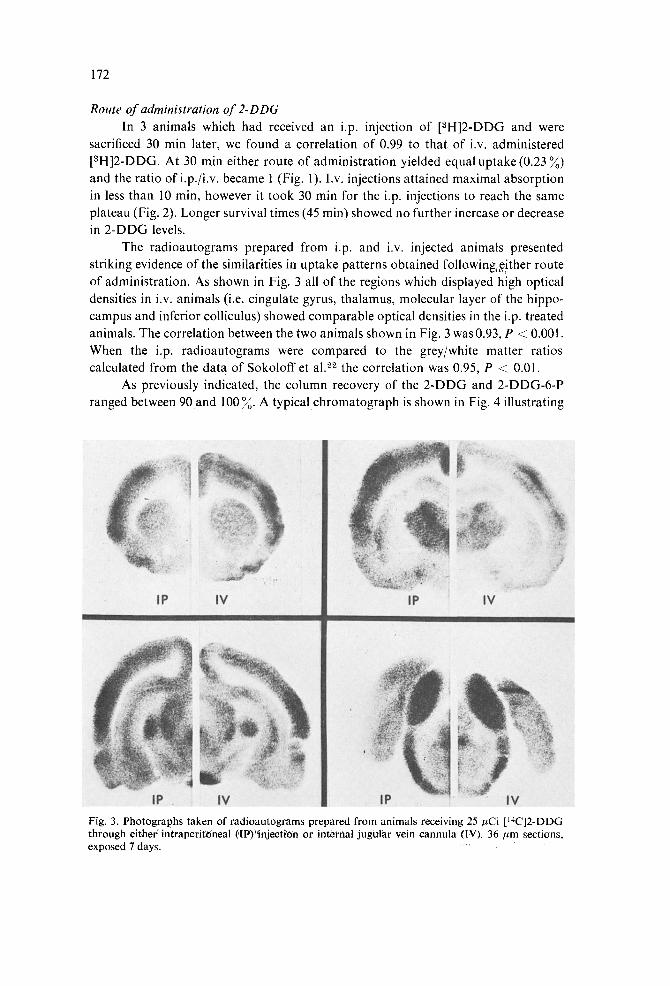
The radioautograms prepared from i.p. and i.v. injected animals presented striking evidence of the similarities in uptake patterns obtained following~,~!ther route of administration. As shown in Fig. 3 all of the regions which displayed high optical densities in i.v. animals (i.e. cingulate gyrus, thalamus, molecular layer of the hippo- campus and inferior colliculus) showed comparable optical densities in the i.p. treated animals. The correlation between the two animals shown in Fig. 3 was 0.93, P < 0.001. When the i.p. radioautograms were compared to the grey/white matter ratios calculated from the data of Sokoloff et al. ~2 the correlation was 0.95, P < 0.01.
As previously indicated, the column recovery of the 2-DDG and 2-DDG-6-P ranged between 90 and 100 ~. A typical chromatograph is shown in Fig. 4 illustrating
Fig. 3. Photographs taken of radioautograms prepared from animals receiving 25/zCi [14C]2-DDG through eitherintraperit'Oneal flF)"injecti'On or internal jugular vein cannula (IV). 36/tm sections, exposed 7 days.

173
f-
la. z
c 3
601
5 0 I
4 0 1
3 0 1
2 0 ,
101
0
e
k ........... 14 C DG
i - - :~ H DG
/
1.0 2 0 3.0 4.0 5.0 f4L. H 0
t9
z
IK
~2~
60 '
50~
40~
30a
20~
I0~
0
/ J L .....
- - - 3 H DGP
........ 24C DGP
• ,'o" ~o" ~.o" ~o" ~.o ML. HCt_ ~l.SN~
Fig. 4. Column chromatography of 2-DDG and 2-DDG-6-P. Double-label scintillation counting of the two internal standards ([14C]2-DDG, 64,623 dpm; [laC]2-DDG-6-P, 29,560 dpm) showed that 100 % of the 2-DDG and 92 % of the 2-DDG-6-P was recovered. Based upon these values, 5 % of the tritiated material co-chromatographed with [14C]2-DDG and 95 ~ with [14C]2-DDG-6-P. The average value was 96.3 + 2.2%, n = 4.
the separation and relative abundance of administered [aH]2-DDG and, its metabolic product [3H]2-DDG-6-P. In addition, the separation of [14C]2-DDG and [a4C]2- DDG-6-P, added as internal standards, is also shown. Greater than 95% of the administered [3H]2-DDG eluted as [aH]2-DDG-6-P and the [3H]2-DDG and [3H]2- DDG-6-P co-chromatographed with their respective carbon-14 radionuclides.
DISCUSSION
Correlation of 2-deoxyglucose-6-phosphate distribution with glucose utilization Although many studies employing the 2 - D D G technique attempt quantification
of their data on the basis o f densitometry alone, or from liquid scintillation counting of dissected brain regions, it would appear from the operational equation that these methods cannot be employed for comparing 2 - D D G incorporation among different animals. This is due to the fact that plasma glucose levels play a significant role in determining the kl, k~, and ks rate constants. Therefore, since plasma glucose levels probably vary among animals (especially under experimental conditions) it would be incorrect to assume that the differences seen either in the radioautograms or in the counts are relative to actual differences in 2 -DDG incorporation (i.e. glucose utiliza- tion). However, in all o f our control animals in both this study and in two previous communications 8,12 we never found a significant variation in any structure among the groups of animals studied. While we did observe differences in actual counts or optical densities, these were always found to be attributable to variability of activity in our injected bolus o f radioisotope. In addition, both our 3H and 14C experiments were highly correlated with one another and even without determining plasma glucose levels, both were highly correlated with glucose utilization values determined in Soko-

174
loft's control group. Therefore it appears that possible variations in plasma glucose levels do not measureably alter 2-DDG incorporation. This is a critical point for two reasons. Firstly, it allows relative comparisons of 2-DDG incorporation between animals without prior determination of plasma glucose levels and glucose utilization values. Secondly, it allows comparisons to be made between control and experimental animals. The fact that, in animals treated with either amphetamine 6, or phencycli- dine 12, we found insignificant changes from control animals in some regions of the brain further suggests that small changes in blood glucose do not alter 2-DDG uptake. One should use caution, however, as procedures which can greatly alter cerebral blood flow (e.g. lesions, strokes) must receive special consideration. At the present time we would believe that new rate constants would have to be determined under these con- ditions. Without knowledge of plasma glucose levels, comparisons could not be made among animals since we have no way of knowing whether the differences in 2-DDG incorporation are due to changes in metabolism or are merely reflective of changes in blood glucose. Glucose utilization must be determined in these cases as well as new rate constants. In some instances however, GU determinations may still not accurately reflect metabolic changes. If there were localized changes in blood flow (e.g. middle cerebral artery occlusions) with compensatory changes in structures immediately peripheral to the site of injury, plasma glucose levels would appear unaltered. The changes then, in 2-DDG levels, as seen in the autoradiograms, would incorrectly appear as changes in GU when it could very well be due instead to a differential distribution of cerebral glucose. Nevertheless, for the majority of studies, it appears perfectly valid to compare 2-DDG uptake. For 14C studies using a densitometer, the methacrylate standards must be used and the optical densities must be converted to /~Ci/g. This assures that changes in developing procedure (i.e. length of exposure, age of developer, time of development) do not produce artifacts. In addition, by presenting the data as grey/white matter ratios using the same white matter structure (i.e. corpus callosum) differences in amount of label injected, or section thickness would be elimin- ated. In studies employing [aH]2-DDG, counts in the amount injected or plasma 2- DDG levels should be obtained. By expressing the data as percent uptake for each region different animals could be compared and the variability that occurs when in- jecting the isotope would be eliminated.
Differences in route of administration of isotope When the operational equation is employed for quantification of the method the
animals must be restrained and cannula implanted to obtain both arterial and venous blood samples. Obviously, this places limits on the application of the methodology as it could not be extended to behavioral studies which would require freely moving animals. Even in studies which would not require mobility the procedure places the animal under unusually stressful conditions. Since we were able to quantify the tech- nique without the use of the operational equation we found that the i.v. route of administration of the labeled 2-DDG was also not necessary. Our results indicated that both [all]- and [14C]2-DDG i.p. injections were highly correlated with i.v. injected animals. Further, the values obtained after i.p. administration were also highly

175
correlated with the values taken from Sokoloff et al.2L Striking evidence was presented in the radioautograms in which the two routes of administration were indistin- quishable from one another. It was also shown that after 30 min both i.p. and i.v. animals reached equal incorporation. In addition there was almost complete enzyma- tic conversion of the i.p. administered 2-DDG to 2-DDG-6-P, as shown in the chro- matographic separation. Differences between the two routes of administration appear- ed in the rate of uptake. While i.v. injected animals took up the label in less than 10 min it required 30 min for the i.p. injected animals to attain the same level of incorpora- tion. This difference allows the investigator to choose his route of injection according to the time profile of his stimulus. For example, in pharmacological studies, in which drug-induced changes occur along a slow time sequence it might be advantageous to use the i.p. route. Alternatively, for stimulation experiments which are immediate cause and effect situations, one would prefer to employ the i.v. route so that stimula- tion would only have to be maintained for a few minutes.
In conclusion, the results of the present series of experiments increases the possible applications of the 2-DDG technique. First, the less expensive tritium-labeled isotope can be used in studies in which selected areas are being investigated. Second, densitometric values obtained from 14C radioautograms from different animals and/or experimental conditions can be compared. Finally, awake, freely moving animals under normal physiological conditions can be given a pulse of labeled 2-DDG without relying on chronically implanted cannula. This last point now allows the 2-DDG tech- nique to be used in a wide range of behavioral studies.
ACKNOWLEDGEMENTS
The authors wish to thank Ms. Linda Weaver for preparation of the figures, and Dr. Robert Katzman, Dr. Lucy Brown, and Dr. Leslie Wolfson for generously allow- ing us use of their densitometer. This research was supported in part by Grant DA01044 and by Research Scientist Development Award DA70082 to S.D.G.
REFERENCES
1 Basinger, S. F., Gordon, W. C. and Lam, D. M. K., Differential labeling of retinal neurones by aH- 2-deoxyglucose, Nature (Lond.), 280 (1979) 682-684.
2 Brown, L. L. and Wolfson, L. I., Apomorphine increases glucose utilization in the substantia nigra, subthalamic nucleus and corpus striatum of the rat, Brain Research, 140 (1978) 188-193.
3 Buchner, E., Buchner, S. and Hengstenberg, R., 2-Deoxy-D-glucose maps movement-specific nervous activity in the second visual ganglion of Drosophila, Science, 205 (1979) 687-688.
4 Delanoy, R. L. and Dunn, A. J., Mouse brain deoxyglucose uptake after footshock, ACTH analogs, a-MSH, corticosterone or lysine vasopressin, PharmacoL Biochem. Behav., 9 (1978) 21-26.
5 Diamond, I. and Fishman, R. A., High-affinity transport and phosphorylation of 2-deoxy-D- glucose in synaptosomes, J. Neurochem., 20 (1973) 1533-1542.
6 Glick, S. D., Meibach, R. C., Cox, R. D. and Maayani, S., Multiple and interrelated functional asymmetries in rat brain, Life Sci., 25 (1979) 395400.
7 Hendrickson, A. E. and Wilson, J. R., A difference in [14Cldeoxyglucose autoradiographic patterns in striate cortex between Macaca and Saimiri monkeys following monocular stimulation, Brain Research, 170 (1979) 353-358.

176
8 Horn, G., McCabe, B. J. and Bufeson, P. P. G., An autoradiographic study of the chick brain after imprinting, Brain Research, 168 (1979) 361-373.
9 Kennedy, C., DesRosiers, M. H., Jehle, J. W., Reivich, M., Sharp, F. and Sokoloff, L., Mapping of functional neural pathways by autoradiographic survey of local metabolic rate with [14C]deoxyglu- cose, Science, 187 (1975) 850-853.
10 Kliot, M. and Poletti, C., Hippocampal aftercharges: Differential spread of activity shown by the [14C] deoxyglucose technique, Science, 204 (1979) 641-643.
11 Kohsaka, S.-I., Takamatsu, K., Aoki, E. and Tsukada, Y., Metabolic mapping of the chick brain after imprinting using [14C]2-deoxyglucose technique, Brain Research, 172 (1979) 539-544.
12 Meibach, R. C., Glick, S. D., Cox, R. and Maayani, S., Localisation of phencyclidine-induced changes in brain energy metabolism, Nature (Lond.), 282 (1979) 625-626.
13 Reinstein, D. K., Isaacson, R. L. and Dunn, A. J., Regional changes in 2-deoxyglucose uptake after neocortical and hippocampal destruction, Brain Research, 175 (1979) 392-397.
14 Sakurada, O., Sokoloff, L. and Jacquet, Y. F., Local cerebral glucose utilization following injection of fl-endorphin into periaqueductal gray matter in the rat, Brain Research, 153 (1978) 403-407.
15 Savak, H. E., Kadekaro, M., Jehle, J. and Sokoloff, L., a- and fl-adrenoreceptor blockers have opposite effects on energy metabolism of the central auditory system, Nature (Lond.), 276 (1978) 521-523.
16 Schwartz,W. J., A role for the dopaminergic nigrostriatal bundle in the pathogenesis of altered brain glucose consumption after lateral hypothalamic lesions. Evidence using the 14C-labeled deoxygluco- se technique, Brain Research, 158 (1978) 129-147.
17 Schwartz, W. J. and Sharp, F. R., Autoradiographic maps of regional brain glucose consumption in resting awake rats using [~4C]2-deoxyglucose, J. camp. Neurol., 177 (1978) 335-360.
18 Schwartz, W. J., Sharp, F. R., Gunn, R. H. and Evarts, E. V., Lesions of ascending dopaminergic pathways decrease forebrain glucose uptake, Nature (Lond.), 261 (1976) 155-157.
19 Schwartz, W. J., Smith, C. B., Davidsen, L., Savaki, H., Sokoloff, L., Mata, M., Fink, D. J. and Gainer, H., Metabolic mapping of functional activity in the hypothalamo-neurohypophysial system of the rat, Science, 205 (1979) 723-725.
20 Sharp, F. R., Kaver, K. S. and Shepherd, G. M., Local sites of activity-related glucose metabolism in rat olfactory bulb during olfactory stimulation, Brain Research, 98 (1975) 596-600.
21 Sokoloff, L., Relation between physiological function and energy metabolism in the central nervous system, J. Neurochem., 29 (1977) 13-26.
22 Sokoloff, L., Reivich, M., Kennedy, C., Des Rosiers, M. H., Patlak, C. S., Pettigrew, K. D., Sakurada, O. and Shinohara, M., The [~4C]deoxyglucose method for the measurement of local cerebral glucose utilization: theory, procedure and normal values in the conscious and anesthetized albino rat, J. Neurochem., 28 (1977) 897-916.
23 Sokoloff, L., Reivich, M., Patlak, C. S., Pettigrew, K. D., Des Rosiers, M. and Kennedy, C., The [14C]deoxyglucose technique for the quantitative determination of local cerebral glucose con- sumption. In Proc. 5th Meet. Amer. Sac. Neurochem., 1974 p. 86.
24 Wechster, L. R., Savak, H. E. and Sokoloff, L., Effects of D- and L-amphetamine on local cerebral glucose utilization in the conscious rat, J. Neurochem., 32 (1979) 15-22.
25 Weeks, J. R., Long-term intravenous infusion. In R. D. Myers (Ed.), Methods in Psychobiology, Vol. 2, Academic Press, 1972, pp. 155-168.
26 Wolfson, L. I., Sakurada, O. and Sokoloff, L., Effects of 7-butyrolactone on local cerebral glucose utilization in the rat, J. Neurochem., 29 (1977) 777-783.




















