
The crystal structures of trout Hb I in the deoxy and carbonmonoxy forms
12
J. Mol. Biol. (1996) 259, 749–760 The Crystal Structures of Trout Hb I in the Deoxy and Carbonmonoxy Forms Jeremy R. H. Tame 1 *, Julie C. Wilson 1 and Roy E. Weber 2 We have determined the X-ray crystallographic structure of trout Hb I in 1 Department of Chemistry both the deoxy and carbonmonoxy forms to resolution limits of 2.3 Å and University of York 2.5 Å, respectively. The overall fold of the molecule is highly similar to that Heslington, York YO1 5DD UK of human HbA despite the low level of sequence identity between these proteins. Trout Hb I is unusual in displaying almost no pH dependence 2 Zoophysiology Department of oxygen binding affinity, and (at most) very weak interactions with Institute of Biological heterotropic effector ligands such as organic phosphates. Comparison of Sciences, Building 131 the two quaternary states of the protein indicates how such effects are Aarhus University minimised and how the low-affinity T state of the protein is stabilised in 8000 Aarhus C, Denmark the absence of heterotropic interactions. 7 1996 Academic Press Limited *Corresponding author Keywords: crystallography; haemoglobin; Bohr effect Introduction Unlike the majority of mammals, which produce only a single major haemoglogin (Hb) component (>90% of the Hb content of the red cell), many fish species have multiple Hb components which show considerable differences in amino acid residue sequence and functional properties (di Prisco & Tamburrini, 1992; Riggs, 1979; Weber, 1982). Despite the considerable effort which has been devoted to characterising a number of fish Hbs, it remains unclear in many cases what advantage is bestowed by this multiplicity. Clearly, however, functional heterogeneity of several Hb types within the red cell could extend the range of conditions under which oxygen can be transported effectively around the blood stream, and permits a division of labour between the various components which can each fulfil a specific role (Weber, 1990). Blood from the trout Oncorhyncus mykiss (formerly Salmo irideus ) contains four distinct Hbs which can be separated by ion-exchange chromatography (Binotti et al ., 1971; Brunori et al ., 1973). Hb I (pI = 8.2) and Hb IV (pI = 6.2 to 6.5) make up roughly 20% and 65% of the total Hb content, respectively. These differ markedly in their response to changes of pH. Under physiological conditions the oxygen affinity of human Hb decreases with pH, an effect known as the Bohr effect (Kilmartin et al ., 1973). Trout Hb IV is characterised by the Root effect, an unusually strong dependence of oxygen affinity on pH. Below pH 6 the low-affinity T state of the molecule is so strongly stabilised that oxygen binding becomes non-cooperative. This allows discharge of oxygen into the swim-bladder against the concentration gradient by the secretion of lactic acid into the blood in the gas gland. Trout Hb I, in contrast, shows no change in oxygen affinity or cooperativity with pH (Binotti et al ., 1971; Weber et al ., 1976). The function of trout Hb I appears to be to maintain a viable level of oxygenation during hyperactivity acidosis which would prevent oxygen uptake by Hb IV (Brunori, 1975), although active fish like tench also lack Hb I. It may also play a role in temperature acclimation (Weber et al ., 1976). Carp and the Antarctic fish Pagothenia bernachii , which are both much more sedentary than trout, have only Root effect Hbs, indicating that Hb I may be necessary only to support an active life-style (Camardella et al ., 1992; Tan et al ., 1972). As well as lacking a Bohr effect, Hb I shows no interaction with heterotropic effector ligands such as ATP and DPG (diphosphoglycerate) or chloride ions. Stripped of such ligands human Hb shows an oxygen affinity too high for efficient oxygen transport throughout the body, but their presence in the red cell stabilises the low oxygen affinity T state of the molecule. The amino acid residues principally responsible for the Bohr effect and other heterotropic interactions in human Hb are known from structural and functional studies of native and modified Hb (Perutz, 1980), and many of these are found to be altered in trout Hb I (Bossa et al ., 1978; Barra et al ., 1983). Unlike human Hb and trout Hb IV, trout Hb I shows no change in circular dichroism on ligation, Abbreviations used: Hb, haemoglobin; DPG, diphosphoglycerate; HbPb, Pagothenia bernachii Hb; PEG, polyethylene glycol. 0022–2836/96/240749–12 $18.00/0 7 1996 Academic Press Limited
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of The crystal structures of trout Hb I in the deoxy and carbonmonoxy forms

J. Mol. Biol. (1996) 259, 749–760
The Crystal Structures of Trout Hb I in the Deoxyand Carbonmonoxy Forms
Jeremy R. H. Tame 1*, Julie C. Wilson 1 and Roy E. Weber 2
We have determined the X-ray crystallographic structure of trout Hb I in1Department of Chemistryboth the deoxy and carbonmonoxy forms to resolution limits of 2.3 Å andUniversity of York2.5 Å, respectively. The overall fold of the molecule is highly similar to thatHeslington, York YO1 5DD
UK of human HbA despite the low level of sequence identity between theseproteins. Trout Hb I is unusual in displaying almost no pH dependence2Zoophysiology Department of oxygen binding affinity, and (at most) very weak interactions with
Institute of Biological heterotropic effector ligands such as organic phosphates. Comparison ofSciences, Building 131 the two quaternary states of the protein indicates how such effects areAarhus University minimised and how the low-affinity T state of the protein is stabilised in8000 Aarhus C, Denmark the absence of heterotropic interactions.
7 1996 Academic Press Limited
*Corresponding author Keywords: crystallography; haemoglobin; Bohr effect
Introduction
Unlike the majority of mammals, which produceonly a single major haemoglogin (Hb) component(>90% of the Hb content of the red cell), many fishspecies have multiple Hb components which showconsiderable differences in amino acid residuesequence and functional properties (di Prisco &Tamburrini, 1992; Riggs, 1979; Weber, 1982).Despite the considerable effort which has beendevoted to characterising a number of fish Hbs, itremains unclear in many cases what advantage isbestowed by this multiplicity. Clearly, however,functional heterogeneity of several Hb types withinthe red cell could extend the range of conditionsunder which oxygen can be transported effectivelyaround the blood stream, and permits a division oflabour between the various components which caneach fulfil a specific role (Weber, 1990). Blood fromthe trout Oncorhyncus mykiss (formerly Salmoirideus) contains four distinct Hbs which can beseparated by ion-exchange chromatography(Binotti et al., 1971; Brunori et al., 1973). Hb I(pI = 8.2) and Hb IV (pI = 6.2 to 6.5) make uproughly 20% and 65% of the total Hb content,respectively. These differ markedly in theirresponse to changes of pH. Under physiologicalconditions the oxygen affinity of human Hbdecreases with pH, an effect known as the Bohreffect (Kilmartin et al., 1973). Trout Hb IV ischaracterised by the Root effect, an unusually
strong dependence of oxygen affinity on pH. BelowpH 6 the low-affinity T state of the molecule is sostrongly stabilised that oxygen binding becomesnon-cooperative. This allows discharge of oxygeninto the swim-bladder against the concentrationgradient by the secretion of lactic acid into the bloodin the gas gland. Trout Hb I, in contrast, shows nochange in oxygen affinity or cooperativity with pH(Binotti et al., 1971; Weber et al., 1976). The functionof trout Hb I appears to be to maintain a viable levelof oxygenation during hyperactivity acidosis whichwould prevent oxygen uptake by Hb IV (Brunori,1975), although active fish like tench also lack Hb I.It may also play a role in temperature acclimation(Weber et al., 1976). Carp and the Antarctic fishPagothenia bernachii, which are both much moresedentary than trout, have only Root effect Hbs,indicating that Hb I may be necessary only tosupport an active life-style (Camardella et al., 1992;Tan et al., 1972). As well as lacking a Bohr effect, HbI shows no interaction with heterotropic effectorligands such as ATP and DPG (diphosphoglycerate)or chloride ions. Stripped of such ligands humanHb shows an oxygen affinity too high for efficientoxygen transport throughout the body, but theirpresence in the red cell stabilises the low oxygenaffinity T state of the molecule. The amino acidresidues principally responsible for the Bohr effectand other heterotropic interactions in human Hbare known from structural and functional studies ofnative and modified Hb (Perutz, 1980), and many ofthese are found to be altered in trout Hb I (Bossaet al., 1978; Barra et al., 1983).
Unlike human Hb and trout Hb IV, trout Hb Ishows no change in circular dichroism on ligation,
Abbreviations used: Hb, haemoglobin; DPG,diphosphoglycerate; HbPb, Pagothenia bernachii Hb;PEG, polyethylene glycol.
0022–2836/96/240749–12 $18.00/0 7 1996 Academic Press Limited

Crystal Structures of Trout Hb I750
Table 1. Alignment of fish and human a globin sequences from the Nterminus to Pro37(C2) of the human proteinTrout I .SLTAKDKSVVKAFWGKISGKADVVGAEALGRDKMLTAYPHuman .VLSPADKTNVKAAWGKVGAHAGEYGAEALER..MFLSFPMarbled electric ray .VLSEGNKKAIKNLLQKIHSQTEVLGAEALAR..LFECHPSpiny dogfish .VLSAADKTAIKHLTGSLRTNAEAWGAESLAR..MFATTPG. acuticeps .SLSDKDKAAVRALWSTISKSSDAIGNDALSR..MIVVYPC. mawsoni .SLSDKDKAAVKALWTTISKSSDAIGNDALSR..MIVVYPBluefin tuna TTLSDKDKSTVKALWGKISKSADAIGADALGR..MLAVYPBlack rockcod .SLSTKDKETVKAFWSKVSGKSEDIGNDALSR..MLVVYPAtlantic salmon MSLTARDKSVVNAFWGKIKGKADVVGAEALGR..MLTAYPGoldfish .SLSDKDKAVVKALWAKIGSRADEIGAEALGR..MLTVYPElectric eel .SLTAKSKSIVKAFWGKIGSRADDIGAEAFGR..MLTVYPCarp .SLSDKDKAAVKIAWAKISPKADDIGAEALGR..MLTVYPTrout IV .SLSAKDKANVKAIWGKILPKSDEIGEQALSR..MLVVYPCoelocanth .GLTAADKTLIKSIWGKVEKETEAIGVEALVR..LFKCFPSouth American lungfish MRFSQDDEVLIKEAWGLLHQIPNA.GGEALAR..MFSCYP
It can be seen that the Asp-Lys insertion (between Arg31 and Met32 of human aglobin) in the trout Hb I sequence (Pir1:HATR1) is not found in other sequences.
and the CD spectra have been taken to indicate thattrout Hb I has a smaller proportion of amino acidresidues in a helical conformation than human Hb(Brunori, 1975). We have determined the structureof trout Hb I in both the deoxy and carbonmonoxyforms to examine the structure and allosterictransition of the protein. Despite the low level ofsequence identity between human Hb and trout HbI the structures of the two proteins are very similarin both the T and R states, and suggest possibleintramolecular interactions for stabilising the Tstate of trout Hb I which do not require heterotropicligands.
Results
Structure description
Trout Hb I, like human Hb, contains 146 residuesin each b subunit (Barra et al., 1983). According tothe published sequence, trout Hb I has three more,residues, 144 in all, in the a subunit (Bossa et al.,1978). Two of these, an aspartic acid residue and alysine residue, are inserted between the conservedArg31 and Met32 in the middle of the B helix(Table 1). No insertions are seen at this point inother fish Hbs, and we see no evidence for theseresidues in the electron density maps. There is clear
evidence for the extra alanine residue at position 47in the trout I a chain. The Asp-Lys insertiontherefore appears to be a sequencing error and wehave omitted these residues from our models. (BothArg31 and Met32 are clearly visible in the maps; thearginine residue forms hydrogen bonds withGlu27a and Gln127b and the methionine residuesits in an apolar pocket.) The a subunits of the troutHb I models described here therefore contain 142amino acid residues. The Ca trace of the protein canbe closely superimposed on that of human Hb overthe whole structure (Figure 1). This shows that theextra residue creates a kink in the main chain atposition 47 in the a CE corner although residues 147and 48 to 141 of human a globin overlap closelyresidues 1 to 47 and 49 to 142 of trout I a globin inboth the deoxy and carbonmonoxy forms. Likehuman Hb, trout Hb I has seven a-helices in the asubunit and eight in the b subunit. In the a subunitthe C helix is distorted, and the E helix isinterrupted at residue 71. The b F helix is bent inthe same way as in human Hb. This high degree ofsimilarity between human Hb and trout Hb I isfound in both the deoxy and carbonmonoxy formsof the protein, despite the low sequence identitybetween the two proteins and slight rigid-bodyshifts of various a-helices in the structure. Tables 2and 3 show the rms deviations of main chain atoms
Figure 1. A cross-sectional slabthrough the Ca trace of deoxy troutHb I (thick lines) also showing theCa trace of deoxy human Hb (thinlines; Fermi et al., 1984) superim-posed by least-squares fit of theBGH frame (Baldwin & Chothia,1979). The asterisk indicates theposition of Ala a47 which appearsto be an insertion in the fish a chainwhen aligned with the humanprotein. The extra amino acidresidue at the CE corner extends theloop in the chain at this point butotherwise the backbone atoms alignvery closely.

Crystal Structures of Trout Hb I 751
Table 2. The rms deviations between deoxy trout Hb Iand deoxy human Hb
Trout Hb I
Dimer Tetramer
Deoxy BGH All BGH All
Human 0.627 1.151 0.780 1.260
The least-squares overlap was carried out using 91 Ca atomsof the BGH frame or 1148 atoms over the main chain of thedimer, and 182 Ca atoms or 2296 atoms in the tetramer.
Figure 2. A schematic diagram showing interactionsformed at the a1b2 interface in the T (deoxy) and R (oxy)states of human Hb.
between trout Hb I and human Hb, using Ca atomsfrom the BGH frame (Baldwin & Chothia, 1979) andall main chain atoms (omitting residue 48 of troutHb I). Both an ab dimer and the whole tetramer arecompared. While the carbonmonoxy form of thetrout protein (determined under low-salt con-ditions) closely matches the high-salt human HbCOstructure, the tetramer fits much less well over thelow-salt R2 structure of human HbCO (Silva et al.,1992).
Allosteric mechanism
Vertebrate haemoglobins are characterised by anincrease in oxygen affinity with oxygen saturation,an effect necessary for efficient oxygen transport. Inthe case of human Hb, extensive structural study ofthe protein in different ligation states by Perutz andcollaborators has shown that it adopts twoprincipal structures, the allosteric equilibriumbetween them depending on ligand binding to thehaems (Perutz et al., 1987). The T (tense) statefavoured by deoxy Hb shows lower oxygen affinitythan the R (relaxed) state. Ligand binding to thehaem groups of deoxy Hb pulls on the proximalhistidine residue to which the iron atom iscovalently bonded, triggering the change inquaternary structure. The change from T to R stateinvolves relatively minor changes in the tertiarystructure of the individual subunits, but one abdimer rotates relative to another; thus the a1b1
subunit contact is largely preserved but the a1b2
contact is altered on ligand binding. Studies ofintermediate ligation states suggest that the b FGcorner and a C helix act as the principal switchregion, sensing ligation of the haem iron atom andmoving relative to each other (Figure 2). In contrast,
the a FG corner and b C helix behave as a flexiblejoint (Baldwin & Chothia, 1979).
Although refined to a lower resolution thanhuman Hb in the R and T states, our models stillpermit clear features of the allosteric mechanism tobe seen. The rms deviations between the BGHframes of the two trout Hb models suggestcoordinate errors of 0.20 to 0.25 A over thebetter-ordered parts of the structures, which agreeswith the rms errors indicated by the Luzzati plots(data not shown) for both models. Cruickshank haspointed out however that estimates of errors incrystal structures from Luzzati plots should betreated with caution (Dodson et al., 1995). Theaverage temperature factors of protein atoms in thetwo structures (23.3 A2 for deoxy and 29.0 A2 forcarbonmonoxy) imply an rms amplitude ofvibration of about 0.55 A to 0.60 A.
Most residues forming the a C helix-b FG cornercontact region of human Hb are preserved in troutHb I. The interactions across the a1b2 interface introut Hb I and human Hb in the deoxy andcarbonmonoxy forms are shown in Tables 4 to 7.The change from Phe to Tyr of 41b(C7) significantlyalters the contacts made by the residue in both
Table 3. The rms deviations between carbonmonoxytrout Hb I and carbonmonoxy human Hb in the R stateand R2 state
Trout Hb I
Dimer Tetramer
Carbonmonoxy BGH All BGH All
Human 0.632 1.314 0.807 1.447Human R2 0.636 1.500 1.429 2.251
The least-squares overlap was carried out using 91 Ca atomsof the BGH frame or 1148 atoms over the main chain of thedimer, and 182 Ca atoms or 2296 atoms in the tetramer.

Crystal Structures of Trout Hb I752
Table 4. Contacts <3.5 A between a1 and b2 in deoxytrout Hb I
Deoxy trout Hb Ia1 b2 Distance (A)
K 40 NZ F 146 O 3.5T 41 CB F 97 O 3.5T 41 CG2 Y 145 CE2 3.5Y 42 CE2 R 40 NH1 3.0Y 42 CZ D 99 OD2 3.5Y 42 OH D 99 CB 3.3Y 42 OH D 99 CG 3.4Y 42 OH D 99 OD2 2.6L 92 O R 40 CD 3.5R 93 CB R 40 CD 3.4R 93 CZ P 36 O 3.4R 93 NH2 P 36 O 3.2R 93 NH2 Q 39 OE1 2.9D 95 CB D 99 OD2 3.4D 95 OD1 W 37 CE2 3.4D 95 OD1 W 37 NE1 2.6N 98 ND2 D 99 OD2 2.8Y 141 CD2 W 37 CE3 3.3R 142 CD V 34 O 3.1R 142 NH1 V 34 CG1 3.3R 142 NH1 V 34 O 2.7
Table 6. Contacts <3.5 A between a1 and b2 incarbonmonoxy trout Hb I
Carbonmonoxy trout Hb Ia1 b2 Distance (A)
Q 38 OE1 D 99 OD2 3.3Q 38 NE2 D 99 OD2 3.1R 93 O R 40 CD 3.2R 93 O R 40 NE 3.4D 95 CB Y 41 OH 3.5D 95 CG Y 41 OH 3.3D 95 OD1 Y 41 CZ 3.4D 95 OD1 Y 41 OH 2.4D 95 OD1 D 99 CB 3.2D 95 OD1 N 102 ND2 3.0P 96 CD W 37 CG 3.5R 142 CD V 34 O 3.0R 142 NH1 V 34 CG1 3.2R 142 NH1 V 34 O 2.8R 142 NH1 Y 35 CE2 3.4
instead forms an Asn turn by bonding to thecarbonyl oxygen atom of Asp99b(G1). In human Hban Asn turn is formed in the T state, and in the Rstate Asn102b(G4) bonds to the Asp95a2(G1)carboxyl group. The mutation Phe41b(C7) to Tyrhas been detected clinically (Hb Mequon), andengineered into human Hb by site-directedmutagenesis. The mutant protein shows loweredoxygen affinity, which would be expected given thelarger number of hydrogen bonds the Tyrside-chain can form in the T state of the protein(Baudin et al., 1992).
In human Hb both 38a(C3) and 41a(C6) arethreonine residues. The Asp99(G1) residue of theadjacent b2 subunit lies between these twothreonine residues in deoxy Hb, but slides past C3as the protein moves to the R state. Perutz (1970)likened this fitting of the Asp into the gaps betweenresidues along the C helix to the knuckles of onehand passing over those of the other. The threonineC3 side-chain makes no hydrogen bonds except tothe carbonyl group of His97b2(FG4) in the R stateonly. In trout Hb I, residue 97b is Phe; 38a is Gln(as in the majority of fish Hb sequences) butappears to perform a similar role, making nohydrogen bonds to other protein atoms in the Tstate, but bonding to the side-chains of Asn98a(G4)and Asp99b2(G1) in the R state. These bonds may
quaternary states however. Tyr is found at thisposition in about 10% of vertebrate Hb sequences,especially fish and crocodile Hbs, although bothtrout Hb IV and P. bernacchii Hb (HbPb) whichexhibit the Root effect have His instead. In deoxytrout Hb I, Tyr41b makes three hydrogen bonds tothe side-chains of Arg40b(C6) and Asn102b(G4),and the nitrogen atom of Asp99b(G1). In the R statethese bonds are lost and the Tyr instead forms abond to Asp95(G1) of a2, preventing this residuefrom interacting with Trp37b(C3) as it does in theT state. In human Hb this highly conservedtryptophan residue forms hydrogen bonds withAsp95a2(G1) in the T state and with Asn102b(G4) inthe R state. This asparagine residue, absolutelyconserved among vertebrate Hbs, also has slightlydifferent interactions in human and trout I Hbs. Indeoxy trout Hb I the Ne atom forms a hydrogenbond with the Oh of Tyr41b(C7), but in the R state
Table 5. Contacts <3.5 A between a1 and b2 in deoxyhuman Hb
Deoxy human Hba1 b2 Distance (A)
K 40 CE H 146 OXT 3.4K 40 NZ H 146 C 3.4K 40 NZ H 146 OXT 2.5Y 42 CE2 D 99 OD1 3.7Y 42 CZ D 99 OD1 3.3Y 42 OH D 99 CB 3.4Y 42 OH D 99 CG 3.3Y 42 OH D 99 OD1 2.5L 91 O R 40 NE 2.8L 91 O R 40 CZ 3.5L 91 O R 40 NH2 3.1R 92 O R 40 CD 3.3R 92 O R 40 NE 3.3D 94 OD1 W 37 NE1 3.0D 94 OD2 E 101 CG 3.4N 97 ND2 D 99 OD1 2.8R 141 CD V 34 O 3.4R 141 NHl V 34 O 2.9
Table 7. Contacts <3.5 A between a and b2 incarbonmonoxy human Hb
Carbonmonoxy human Hba1 b2 Distance (A)
T 38 OG1 H 97 C 3.5T 38 OG1 H 97 O 2.9R 92 CB W 37 O 3.5R 92 CG W 37 CA 3.4R 92 CG W 37 O 3.3R 92 NH1 P 36 O 3.4R 92 NH2 Q 39 OE1 3.3D 94 OD2 N 102 OD1 2.8P 95 CD W 37 CD2 3.3P 95 CD W 37 CE3 3.5V 96 CG1 D 99 OD1 3.0Y 140 OH W 37 CB 3.3R 141 NH2 V 34 O 2.6

Crystal Structures of Trout Hb I 753
not be very strong, however, since the temperaturefactors of the Gln side-chain atoms are fairly high(045 A2). Despite conservation between humanand trout I Hbs, tyrosine residue 42a(C7) behavesdifferently in the two proteins. In the T state of bothhuman and trout Hb I this residue forms hydrogenbonds to the nitrogen atom of Asp95a(G1) in thesame chain and the side-chain of Asp99b2(G1). In Rstate human Hb both these bonds are lost, but in Rstate trout Hb I only the bond to Asp99b2 is lost.
It can also be seen in Tables 4 to 7 that the twohighly conserved C terminal residues (Tyr-Arg) ofthe a subunits have different contacts in R statehuman and trout I Hbs, even though the pocket intowhich the tyrosine residue fits is also very wellconserved, and these two residues adopt verysimilar conformations in the T state. Tyr141a ofcarbonmonoxy trout Hb I fits deeper into the pocketthan its equivalent in human Hb, apparently due todifferences in the position of Tyr37b2 which packsagainst the tyrosine residue. The penultimatetyrosine residue of the R state b subunits is alsoshifted relative to that in human Hb. Cys93b inhuman Hb keeps the tyrosine residue about 6 Aaway from the side-chain of the proximal histidineresidue (92b), but is changed to an alanine residuein trout Hb I, which allows the tyrosine residue tocome within 4 A of His92b.
Few mutations are found at the aFG corner-b Chelix contact region. In deoxy human Hb, Arg92a1(FG4) points towards Glu 43b2(CD2). Thisglutamate residue is mutated to a glycine residue introut Hb I, so the arginine residue (93a in the fishprotein) forms a hydrogen bond with Gln39b2(C5)instead. The same interaction is found in oxy T statehuman Hb, an intermediate on the pathwaybetween the deoxy T and oxy R states (Paoli et al.,1996). In the R state of human and trout I Hbs thisarginine residue side-chain has high temperaturefactors and makes no close contacts with otherprotein atoms.
Although the a1b1 interface appears structurallypassive it does play an important role inestablishing the allosteric equilibrium between theR and T states. It is known that the Hbs of certainspecies of high-flying geese have unusually highoxygen affinity due to mutations which weaken thesubunit interaction (Zhang et al., 1996). ReplacingPro119a(H2) with Ala, or Met55b(D6) with Ser,greatly increases the oxygen affinity of human Hb(Weber et al., 1993). Both 33b(B15) and 34b(B16) arevaline residues in human Hb, but in trout Hb I, 33bis an isoleucine residue. This residue makes contactwith the highly conserved proline residue a(H2)across the a1b1 contact close to a(H6), which is anisoleucine residue in trout Hb I but an alanineresidue in the great majority of vertebrate Hbs. Themuch larger side-chain of Ile124a H6) is accommo-dated to some extent by the neighbouring b chainmoving away, but it makes close contact withIle33b(B15) and Val34b(B16). In the fish protein,Ile33b(B15) seems to push Met55b(D6) away fromPro120a(H2); the methionine residue has a high
temperature factor and does not seem to interactwith the proline residue in either quaternary state.Thus the Pro-Met contact of human Hb appears tobe replaced in trout Hb I by two other contactsinvolving isoleucine residues 33b(B15) and124a(H6). These unusual amino acid residues atthese positions could play a role in lowering theoxygen affinity of the protein.
Haem pockets
In both a and b subunits the residues in the haempockets take up very similar positions in bothhuman and trout Hb I and there are no mutationswhich appear to alter the ligand environmentsignificantly. The main difference is in the a subunitwhere the distal valine residue (E11) of human Hbis replaced by an isoleucine residue in the fishprotein (Figure 3). In the deoxy trout Hb I structurethis isoleucine residue covers the haem andcompletely blocks ligand access to the iron atom,whereas in the carbonmonoxy structure theside-chain rotates away from the haem to makeroom for the ligand. There is no indication of awater molecule in the a haem pocket, in contrast tohuman deoxy Hb (Fermi et al., 1984). Mutagenicstudies in human Hb have shown that replacingValE11 of human Hb with Ile does not impose greatsteric hindrance on ligand binding and thismutation does not alter the oxygen equilibriumcurve of the protein (Mathews et al., 1989; Tameet al., 1991). There are only slight structural changesin the haem pocket of human a globin on ligation,the iron atom moving closer to the porphyrin planeand the haem flattening slightly (Perutz et al., 1987).In common with human Hb, ligation of trout Hb Icauses a translation of the haem by 0.6 A into theprotein and away from E11 to create space for theligand. The b subunits of human Hb and trout HbI also behave very similarly on ligation; the haemtranslates along and rotates around the F helix, andflattens slightly against residues at the b FG corner.The carbon monoxide ligand appears quite bent inboth a and b subunits (Figure 4). The two atomswere moved by hand to make the Fe-C and C-Obonds co-linear, but they quickly moved away fromthese positions on re-refinement. The ligand atomsare not well represented in the electron density,particularly in the b subunit, and their coordinateerrors are probably higher than those of most atomswithin the body of the protein.
Residue 85b(Fl) is generally Phe in vertebrateHbs but is Tyr in many fish Hb sequences includingtrout I. This side-chain points towards the E helix,and in human Hb the Phe fits between Ala70b(E14)and Ala74b(E18) (Figure 5). The extra Oh group ofthe Tyr in this position is accommodated by thisturn of the E helix ‘‘stretching’’ so that the carbonyloxygen atom of 70b(E14) does not hydrogen bonddirectly to the nitrogen atom of Ala74b(E18); in thedeoxy form of the protein both of these atoms formbonds with the tyrosine residue. Ligation of thehaem causes a coordinated translation of the F

Crystal Structures of Trout Hb I754
Figure 3. The 2Fo − Fc electron density over the a haem of deoxy trout Hb I, contoured at 1s. The Cd atom of IleE11over the haem appears slightly displaced from the 2Fo − Fc density, but refines stably with a B-factor of 29.5 A2. TheCg atoms have B-factors close to 18 A2.
helix, prising it away from the E helix and pullingTyr85b(F1) away from 74b(E18) and so losing ahydrogen bond in the R state.
Trout Hb I is known to be considerably morethermostable than human Hb (Hofrichter et al.,1991), and to dissociate into dimers much lessreadily in the oxygenated form (Brunori et al., 1973;Brunori, 1975). The extra bonds formed by the twoPhe to Tyr mutations and the altered a1b1 interfacemay be partly responsible by tightening the fold ofthe individual subunits and the ab dimers, but the
larger buried surface area in the R state between abdimers must also contribute significantly to thiseffect (Table 8).
Bohr effect
Considerable controversy has surrounded theBohr effect of human Hb and whether it arises froma small set of functional groups or from a largenumber of weak interactions (Russu & Ho, 1986;Derewenda et al., 1990; Shih et al., 1993). Roughly
Figure 4. The 2Fo − Fc electron density map over the a haem of carbon-monoxy trout Hb I, contoured at 1s. Theligand appears strongly bent, but higher-resolution data are required to determine the ligand geometry accurately.Ligand geometry was not restrained in the refinement. Ligand density is weak at the b haem suggesting less than fulloccupancy and/or partial oxidation.
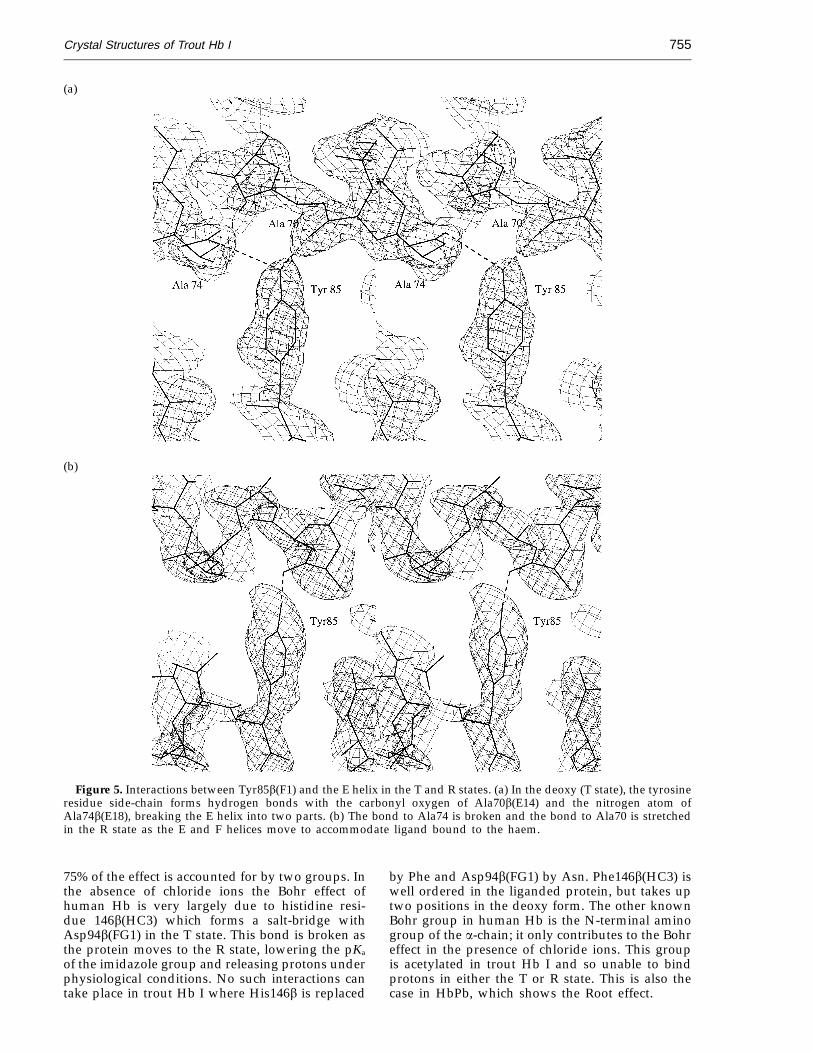
Crystal Structures of Trout Hb I 755
(a)
(b)
Figure 5. Interactions between Tyr85b(F1) and the E helix in the T and R states. (a) In the deoxy (T state), the tyrosineresidue side-chain forms hydrogen bonds with the carbonyl oxygen of Ala70b(E14) and the nitrogen atom ofAla74b(E18), breaking the E helix into two parts. (b) The bond to Ala74 is broken and the bond to Ala70 is stretchedin the R state as the E and F helices move to accommodate ligand bound to the haem.
75% of the effect is accounted for by two groups. Inthe absence of chloride ions the Bohr effect ofhuman Hb is very largely due to histidine resi-due 146b(HC3) which forms a salt-bridge withAsp94b(FG1) in the T state. This bond is broken asthe protein moves to the R state, lowering the pKa
of the imidazole group and releasing protons underphysiological conditions. No such interactions cantake place in trout Hb I where His146b is replaced
by Phe and Asp94b(FG1) by Asn. Phe146b(HC3) iswell ordered in the liganded protein, but takes uptwo positions in the deoxy form. The other knownBohr group in human Hb is the N-terminal aminogroup of the a-chain; it only contributes to the Bohreffect in the presence of chloride ions. This groupis acetylated in trout Hb I and so unable to bindprotons in either the T or R state. This is also thecase in HbPb, which shows the Root effect.

Crystal Structures of Trout Hb I756
Table 8. Surface area (A2) buried between ab dimersDeoxy Carbonmonoxy
Protein (A2) (A2)
Human Hb 3276 2974Trout Hb I 3263 3349
The contact areas between the a1 b1 and a2b2 dimers of humanHb and trout Hb I were calculated from the difference in solventaccessibility between the tetramer and the separated dimers. Thecalculation was carried out in QUANTA using a probe radius of1.4 A. In contrast to human Hb, trout Hb I shows an increase inthe buried surface area on switching to the R state. The closefit of the R state dimer pair explains the unusually highdimer-tetramer association constant of trout Hb I in the oxyform, which is 12 to 40 times greater than that of human oxy-Hb(Brunori, 1975).
Table 9. Data collection and refinement statisticsDataset Deoxy Carbonmonoxy
Resolution limits (A) 8.0 to 2.3 8.0 to 2.5R = S(=Fo= − =Fc=)/S=Fo= 0.162 0.162Rfree 0.247 0.286Intensity >3s (%) 95.4 75.2Rmerge (%) 4.9 6.5Completeness (%) 85.3 74.1No. of unique reflections 14,572 16,338Average multiplicity 4.1 1.4rms D (bond distances; A)
Bonded 0.014 (0.025) 0.014 (0.025)Angle related 0.030 (0.035) 0.033 (0.035)Interplanar 0.035 (0.050) 0.038 (0.050)
As Ito et al. (1995) have noted however, the effectdoes not seem to correlate with the knownsequences and functional properties of other fishHbs. Furthermore, this interaction could onlyaccount at best for half the Root effect whichinvolves the binding of four to five more protons inthe T state. Surprisingly His146b of Pagothenia Hbdoes not form a salt-bridge in the T state. It has arelatively high B-factor and points into the solventin both structures of the protein so its pK cannotchange substantially on ligand binding or release.
The aspartic acid residues identified by Ito et al.(1995) are conserved in trout Hb I, and acomparison of our structures with those ofPagothenia Hb shows that their positions are similarin both proteins. In the deoxy trout Hb I structurethe two Asp Ca atoms are about 0.5 A further apartthan in HbPb and the side-chains both hydrogenbond to the same water molecule instead of to eachother. The main difference between the two fishHbs around this site is residue 104(G6)b. InPagothenia Hb this is a Lys which points into thesolvent and away from Asp101b. In trout Hb I,however, there is an Arg at this position which inthe T state is well ordered and forms salt-bridgeswith Asp101b. This will prevent the pK of eitherAsp95a or 101b rising in the T state. In the R statethe Arg becomes disordered and these bonds break.Trout Hb IV also has an Arg at position 104b whichpresumably interacts with Asp101b in the sameway and prevents it from contributing to the Rooteffect. The cause of the strong stabilisation of the Tstate by protons remains to be explained in thisprotein. Parkhurst et al. (1983) found that removingthe C-terminal His from the b chains of carp Hbweakened the Root effect, even though this residuedoes not form a salt-bridge in Pagothenia Hb. Itoet al. (1995) have suggested that it may contributeto the Bohr effect through long-distance electro-static interactions. Even so, it appears that ‘‘the’’Root effect is brought about by several differentgroups of residues in different species of fish, someof which remain to be identified.
Discussion
From the two structures described above it can beseen that the allosteric mechanism of human Hbdetermined by Perutz and co-workers closely
Since only histidine residues have a side-chainwith a normal pK value in the physiological range,close attention has been paid to these residues inhuman and fish Hbs to find the cause of the‘‘missing’’ Bohr effect and the Root effect. Thereare, in fact, no histidine residues in trout Hb I whichare not also present in human Hb, and only eightof the 19 in the human sequence are common toboth. These include the proximal and distalhistidine residues of both subunits, which remark-ably are the only histidine residues found in the bsubunit of trout Hb I. The remaining four preservedhistidine residues (all in the a subunits) form thesame interactions as they do in human Hb. His45forms a hydrogen bond to a haem propionategroup in both R and T states, as in human Hb, butthe high temperature factors of the side-chainatoms indicate some disorder. His104 in the middleof the G helix is a key residue, in forming the a1b1
contact, sitting in a pocket between Tyr36a,Val108a, Asp108b and Ile112b. The environment isunchanged between R and T states in both humanHb and trout Hb I. His113 caps the G helix andforms a salt-link with Glu27a which is preserved ontransition between allosteric states. His123 alsoforms an invariant a1b1 contact, lying againstVal34b, but forms no hydrogen bonds. Severalhistidines ‘‘lost’’ from trout Hb I are replaced byapolar residues. However, in most cases the sameapolar residues also appear at these positions in fishHbs with the Root effect, so it seems unlikely thatthey contribute strongly to the Bohr effect.
The search for the residues responsible for theRoot effect of fish Hbs has led Camardella et al.(1992) and Ito et al. (1995) to determine the structureof Pagothenia Hb in the carbonmonoxy and deoxyforms (Brookhaven Data Bank entries 1pbx and1hbh). Unfortunately, the comparison of these Rand T state models has failed, even in the light ofextensive sequence information, to provide acomplete explanation of the effect. The onlyshort-range interaction found in the deoxy form ofthe protein which appeared to contribute signifi-cantly to the Root effect was between two aspartateresidues, 95(G1)a and 101(G3)b (Asp99b is alsonearby). The carboxyl groups of these residuesapproach each other very closely in the T state andappear to share a proton which is lost in the R state.

Crystal Structures of Trout Hb I 757
Figure 6. The Ramachandran plotfor the deoxy structure of troutHb I, drawn using PROCHECK(Laskowski et al., 1993). Glycineresidues are shown as triangles.
Figure 7. The Ramachandran plotfor the carbonmonoxy structure oftrout Hb I indicating the changes inf and c angles between the two abdimers in the asymmetric unit. Thepoints plotted for each residue andits equivalent in the two ab dimersare connected by a line. Overallthe two dimers overlap closelywith an rms deviation of 0.29 Aover all main chain atoms. Thelargest deviations from non-crystal-lographic symmetry occur aroundGly46b which in one dimer forms acrystal contact with a neighbouringHb tetramer. This Figure was pre-pared using SQUID (Oldfield, 1992).
matches that of trout Hb I, despite the limitedsequence identity between the two proteins. Boththe tertiary and quaternary structures of the fishand human proteins can be closely superimposed.Many interactions between the subunits arepreserved, but not always by conserved residues;
conversely, some conserved residues show slightlydifferent behaviour in the two proteins. Trout HbI is characterised by an almost complete loss ofinteraction with allosteric effectors which bind toother vertebrate haemoglobins. In many instancesthis can be explained from the protein sequence

Crystal Structures of Trout Hb I758
alone; for example DPG is known to bind humanHb by interacting with several positively chargedgroups including Lys82b (EF6; Arnone, 1972;Richards et al., 1993), and this well-conservedresidue is changed to leucine in trout Hb I. The lossof the Bohr effect can similarly be attributed to thereplacement of eight histidine residues found inhuman Hb by apolar residues in trout Hb I. Not allthe histidine residues ‘‘missing’’ from trout Hb Ihave been previously implicated in the Bohr effect,and one of those which is retained (His122a) hasbeen suggested as a possible Bohr group(Nishikura, 1978), although subsequent exper-iments showed no evidence for this (Ohe & Kajita,1980). From the structures of human Hb and troutHb I in the T and R states, however, it appears thatall the histidine residues found in trout I, except theproximal and distal histidine residues, have verysimilar environments in both quaternary structures.Several are highly conserved throughout vertebratehaemoglobins and form important interactionsacross the a1b1 contact.
A number of mammalian haemoglobins areknown which do not interact with organicphosphates such as DPG, and these proteins arefound to have large apolar residues replacinghistidine 2b(NA2); Perutz & Imai, 1980). Theseresidues stabilise the T state since they are moreexposed to solvent water in the R state. No suchresidues are found in trout Hb I, yet the proteinshows a much lower oxygen affinity than humanHb under stripped conditions (R. E. Weber & A.Fago, unpublished results). Other interactions mustcompensate for the loss of the Bohr groups andheterotropic effector binding sites which stabilisethe T state. Since the haem pockets of the fishprotein show no obvious reason for the decreasedligand affinity the interactions responsible must befound elsewhere in the protein. Four residues havebeen identified which may lead to a relativestabilisation of the T state, two in the a1b1 contact,one in the a1b2 contact and one at the beginning ofthe b F helix. Of these, Tyr41b(C7) and Tyr85b(F1)are not unique to trout Hb I and probably form thesame interactions in other Hbs. The evidence for theproposed stabilisation of the T state due to Ile33band Ile124a is less compelling, and this may be apeculiar feature of trout Hb I. These extra bondsmay partly account for the known thermostability ofthe protein (relative to human Hb) as well as the lowoxygen affinity. These mutations could thereforeprove useful in the design of genetically engineeredHbs intended for use as artificial blood substitutes.
The refined crystal structures contrast with theconclusion reached by Brunori (1975) on the basisof circular dichroism experiments that the helicalcontent of the protein was significantly less than inhuman Hb. The apparent conflict with the X-raydata is however reconciled by more recent workwhich shows that the mean residue ellipticity of aprotein at 222 nm is not directly proportional tohelical content (as has been assumed previously),but is also dependent on helix length (Hirst &
Brooks, 1994). The fragmentation of long helices,without a reduction in the number of helicalresidues, significantly reduces the mean residueellipticity at 222 nm. As described above, the bsubunit E helix in trout Hb I is indeed fragmentedby the mutation Phe85b(F1) to Tyr, whicheffectively breaks it into two shorter helices. The Chelix of the a subunits is also very distorted, butbeing short perhaps contributes rather less to theCD spectrum. We suggest that Tyr85b is respon-sible for the altered CD profile of trout Hb. Othervertebrate Hbs with this mutation will presumablyalso show reduced CD at 222 nm.
Experimental MethodsBlood samples were drawn from the caudal blood
vessels of trout into heparinised syringes and bubbledwith carbon monoxide. The red cells were washed withsaline before being lysed with cold water. Membranousmaterial was removed by saturating the lysate withsodium chloride and spinning at 12,000 g for one hour.The supernatant was then ultracentrifuged at 230,000 gfor one hour. The protein was dialysed against 50 mMTris-HCl (pH 8.5). Separation of the different Hbcomponents was achieved using a HiLoad monoQcolumn (Pharmacia) equilibrated with the same buffer.Trout Hb I passes through the column immediately;components II, III and IV can be eluted with salt. Hb Iwas then dialysed against 5 mM Tris-HCl (pH 8.0) undercarbon monoxide, concentrated to 10 mg/ml and storedunder CO at 4°C in a rubber-stoppered flask.
Crystals of the carbonmonoxy protein were grownusing hanging drops. Thin, plate-like crystals grew from15% (w/v) polyethylene glycol (PEG) 4000, 50 mMTris-HCl (pH 8.0) mixed with 10 mg/ml protein. Crystalswere mounted under air. Precession photographsshowed the crystal to be orthorhombic, and an initialdataset collected using a Xentronics multi-wire chamberX-ray detector showed the crystals to be in space groupP212121 with cell dimensions a = 65.2 A b = 79.8 A andc = 122.9 A. The crystals diffracted only weakly using arotating anode X-ray generator, but data could becollected to 2.8 A in York. Using a second crystal, datawere collected to 2.1 A on the Weissenberg camera at thePhoton Factory in Tsukuba, Japan, but this dataset has alarge blind region. Limited data collection time pre-vented a more complete dataset being collected. TheXentronics data were processed with XDS (Kabsch, 1988),and Weissenberg data with DENZO (Z. Otwinowski,Yale University). The two datasets were merged and aresolution cut-off of 2.5 A applied to give a reasonablycomplete dataset.
Deoxy trout Hb I batch crystallisations were set up ina glove-box filled with argon using oxy-Hb which wasdeoxygenated in situ. Oxy-Hb was prepared byilluminating a sample of carbonmonoxy protein under astream of air. The ligation state of the protein waschecked after 30 minutes by examining its visiblespectrum. Deoxygenation of the protein was achievedusing the method of Perutz (1968), in which iron(II)citrate is prepared in an anaerobic environment andadded to the protein, buffer and precipitant in the tube.Crystals were grown in 50 mM bisTris-HCl (pH 6.0), 15%PEG 8000. Crystals of two different habits were grown,thin needles from the first set of crystallisations and thick,tabular crystals from the second. Both forms were

Crystal Structures of Trout Hb I 759
mounted under argon. X-ray data were collected in Yorkusing a Rigaku Raxis-II image plate system, but a verylong axis (0306 A) permitted only low-resolution (to3.3 A) data to be collected. The space group appeared tobe P61 or P65, and subsequent molecular replacementshowed it to be P6522 with cell dimensions a = b = 63.3 A,c = 312.7 A and an ab dimer in the asymmetric unit.A tabular crystal, mounted on a small face to orient thelong axis along the capillary, was used to collect data at PX9.6 at the Synchrotron Radiation Source, Daresbury, usinga Mar image plate system with a plate diameter of300 mm. Data were collected in two passes, firstly 20images of 1.5° each at 360 mm (2.3 A at the plate edge) andthen ten images of 3° each at 540 mm. Data were processedwith MOSFLM. The final dataset has an Rmerge of 4.9% withaverage redundancy 4.1. Strong diffraction was observedto at least 2.0 A, but data could not be measured to thisresolution due to the closeness of the spots. The tabularcrystal form has a slightly longer 6-fold axis than theneedle form (312 A), but the same space group.
Structure determination and refinement
Both structures were solved by molecular replacementusing the program AMORE (Navaza, 1990) implementedin the CCP4 protein crystallography suite of programs(CCP4, 1994). Data between 8.0 A and 4.0 A resolutionwere used in both cases. The initial model for the deoxyform was constructed from the coordinates of humandeoxy-haemoglobin (Fermi et al., 1984). The side-chainswere mutated to the sequence of trout Hb I usingQUANTA (Biosym-MSI, San Diego, CA). The structurewas first refined using the simulated annealing methodof X-PLOR (Brunger, 1992) during which the R-factordropped from 49% to 30%. Further stages of refinementwere carried out by the restrained least-squaresprocedure of Konnert & Hendrickson (1980) usingPROLSQ (CCP4, 1994). Water molecules were addedusing ARP (Lamzin & Wilson, 1993). The Fe-Ne2 distancewas restrained to 2.1 A. Manual model adjustment wascarried out with the program O (Jones & Kjeldgaard,1991). The final R-factor is 16.2% with a free R-factor of24.7% for data between 8.0 A and 2.3 A. The refinedmodel, which includes 173 water molecules, has goodstereochemistry with root-mean-square deviations of0.014 A and 2.70° from ideal bond lengths and bondangles, respectively.
The refined deoxy structure was used as the molecularreplacement model for carbonmonoxy trout Hb I. Therefinement proceeded as described above, althoughinitial manual fitting was done using 2Fo − Fc maps inwhich the electron density was averaged between thetwo ab dimers in the asymmetric unit with DM (Cowtan,1994). Non-crystallographic restraints were appliedthroughout refinement. The final model has 163 watermolecules. Some oxidation appears to have occurred,leading to poor electron density over the ligand. Datacollection and refinement statistics for both structures aregiven in Table 9. The Ramachandran plot for the deoxystructure is shown in Figure 6, and for the carbonmonoxystructure in Figure 7.
AcknowledgementsWe thank Dr Angela Fago for helpful comments on the
manuscript and Dr Madeleine Moore for collecting dataat the Photon Factory, Tsukuba. J.R.H.T. is a Royal
Society University Research Fellow and J.C.W. has aTraining Fellowship from the Medical Research Council.This work was supported in part by the BBSRC (grantnumber GR /H68864) and the Danish Natural ScienceResearch Council.
ReferencesArnone, A. (1972). X-ray diffraction study of binding
of 2,3 diphosphoglycerate to human deoxy-haemoglobin. Nature, 237, 146–149.
Baldwin, J. & Chothia, C. (1979). Haemoglobin: thestructural changes related to ligand binding and itsallosteric mechanism. J. Mol. Biol. 129, 175–220.
Barra, D., Petruzzelli, R., Bossa, F. & Brunori, M. (1983).Primary structure of haemoglobin from trout. Aminoacid sequence of the b chain of trout Hb I. Biochim.Biophys. Acta, 742, 72–77.
Baudin, V., Pagnier, J., Lacaze, N., Bihoreau, M. T.,Kister, J., Marden, M., Kiger, L. & Poyart, C. (1992).Allosteric properties of hemoglobin b-41 (C7) Phe toTyr—a stable low-oxygen-affinity variant syn-thesized in Escherichia coli. Biochim. Biophys. Acta,1159, 223–226.
Binotti, I., Giovenco, S., Giardina, B., Antonini, E.,Brunori, M. & Wyman, J. (1971). Studies on thefunctional properties of fish hemoglobins. Theoxygen equilibrium of the isolated hemoglobincomponents from trout blood. Arch. Biochem. Biophys.142, 274–280.
Bossa, F., Barra, D., Petruzzelli, R., Martini, F. & Brunori,M. (1978). Primary structure of haemoglobin fromtrout (Salmo irideus). Amino acid sequence of the achain from trout Hb I. Biochim. Biophys. Acta, 536,298–305.
Brunger, A. (1992). X-PLOR Version 3.1, Yale University,CT, USA.
Brunori, M. (1975) Molecular adaptation to physiologicalrequirements: the hemoglobin system of trout. Mol.Cell. Biochem. 9, 1–39.
Brunori, M., Bonaventura, J., Bonaventura, C., Giardina,B., Bossa, F. & Antonini, E. (1973). Hemoglobinsfrom trout: structural and functional properties. Mol.Cell. Biochem. 1, 189–196.
Camardella, L., Caruso, C., Davino, R., di Prisco, G.,Rutigliano, B., Tamburrini, M., Fermi, G. & Perutz,M. F. (1992). Hemoglobin of the Antarctic fishPagothenia bernachii. Amino acid sequence, oxygenequilibria and crystal structure of its carbonmonoxyderivative. J. Mol. Biol. 224, 449–460.
CCP4. (1994). The CCP4 suite: programs for proteincrystallography. Acta Crystallog. sect. D, 50, 760–763.
Cowtan, K. (1994). DM: an automated procedure forphase improvement by density modification. JointCCP4 and ESF-EACBM Newsletter on Protein Crystal-lography, 31, 34–38.
Derewenda, Z., Dodson, G., Emsley, P., Harris, D., Nagai,K., Perutz, M. & Renaud, J.-P. (1990). Stereochem-istry of carbon monoxide binding to normal humanadult and Cowtown haemoglobins. J. Mol. Biol. 211,515–519.
di Prisco, G. & Tamburrini, M. (1992). The hemoglobinsof marine and fresh water fish: the search forcorrelations with physiological adaptation. Comp.Biochem. Physiol. 102B, 661–671.
Dodson, E., Kleywegt, G. J. & Wilson, K. (1995). Reportof a workshop on the use of statistical validators inprotein X-ray crystallography. Acta Crystallog. sect.D, 52, 228–234.

Crystal Structures of Trout Hb I760
Fermi, G., Perutz, M., Shaanan, B. & Fourme, R. (1984).The crystal structure of human deoxy haemoglobinat 1.74 A resolution. J. Mol. Biol. 175, 159–174.
Hirst, J. & Brooks, C. (1994). Helicity, circular dichroismand molecular dynamics of proteins. J. Mol. Biol. 243,173–179.
Hofrichter, J., Henry, E. R., Szabo, A., Murray, L. P.,Ansari, A., Jones, C. M., Coletta, M., Falcioni, G.,Brunori, M. & Eaton, W. A. (1991). Dynamics of thequaternary conformational change in trout hemo-globin. Biochemistry, 30, 6583–6598.
Ito, N., Komiyama, N. H. & Fermi, G. (1995). Structure ofdeoxyhaemoglobin of the Antarctic fish Pagotheniabernachii with an analysis of the structural basis ofthe Root effect by comparison of the liganded andunliganded haemoglobin structures. J. Mol. Biol. 250,648–658.
Jones, T. A. & Kjeldgaard, M. (1991). O—The Manual,Uppsala University, Sweden.
Kabsch, W. (1988). Evaluation of single-crystal X-raydiffraction data from a position-sensitive detector.J. Appl. Crystallog. 21, 916–924.
Kilmartin, J. V., Breen, J. J., Roberts, G. C. K. & Ho, C.(1973). Direct measurement of the pK values of analkaline Bohr group in human hemoglobin. Proc.Natl Acad. Sci. USA, 70, 1246–1249.
Konnert J. H. & Hendrickson, W. A. (1980). A restrainedparameter thermal factor refinement procedure. ActaCrystallog. sect A, 36, 344–350.
Lamzin, V. S. & Wilson, K. S. (1993). Automatedrefinement of protein models. Acta Crystallog. sect. D,49, 129–147.
Laskowski, R. A., MacArthur, M. W., Moss, D. S. &Thornton, J. M. (1993). PROCHECK: a program tocheck the stereochemical quality of protein struc-tures. J. Appl. Crystallog. 26, 283–291.
Mathews, A. J., Rohlfs, R. J., Olson, J. S., Tame, J., Renaud,J.-P. & Nagai, K. (1989). The effects of E7 and E11mutations on the kinetics of ligand binding to R statehuman hemoglobin. J. Biol. Chem. 264, 16573–16583.
Navaza, J. (1990). Accurate computation of the rotationmatrices. Acta Crystallog. sect. A, 46, 619–620.
Nishikura, K. (1978). Identification of histidine-122a inhuman haemoglobin as one of the unknown alkalineBohr effect groups by hydrogen-tritium exchange.Biochem. J. 173, 671–677.
Ohe, M. & Kajita, A. (1980). Changes in pKa values ofindividual residues of human hemoglobin uponreaction with carbon monoxide. Biochemistry, 19,4443–4450.
Oldfield, T. J. (1992). SQUID: a program for theanalysis and display of data from crystallographyand molecular dynamics. J. Mol. Graph. 10, 247–252.
Paoli, M., Liddington, R., Tame, J., Wilkinson, A. &Dodson, G. (1996). Crystal structure of T statehaemoglobin with oxygen bound at all four haems.J. Mol. Biol. 256, 775–792.
Parkhurst, L. J., Goss, D. J. & Perutz, M. F. (1983). Kineticand equilibrium studies on the role of the b-147histidine in the Root effect and cooperativity of carphemoglobin. Biochemistry, 22, 88–118.
Perutz, M. (1968). Preparation of haemoglobin crystals.J. Crystal Growth, 2, 54–56.
Perutz, M. (1970). Stereochemistry of cooperative effectsin haemoglobin. Nature, 228, 726–739.
Perutz, M. & Imai, K. (1980). Regulation of oxygenaffinity of mammalian haemoglobins. J. Mol. Biol.136, 669–678.
Perutz, M. F., Fermi, G. & Shih, T.-B. (1984). Structure ofdeoxyhemoglobin Cowtown [His HC3(146b) to Leu]:origin of the alkaline Bohr effect and electrostaticinteractions in hemoglobin. Proc. Natl Acad. Sci. USA,81, 4781–4784.
Perutz, M., Fermi, G., Luisi, B., Shaanan, B. & Liddington,R. (1987). Stereochemistry of cooperative mechan-isms in hemoglobin. Acc. Chem. Res. 20, 309–321.
Richard, V., Dodson, G. G. & Mauguen, Y. (1993). Humandeoxyhaemoglobin-2,3-diphosphoglycerate complexlow salt structure at 2.5 A resolution. J. Mol. Biol. 233,270–274.
Riggs, A. (1979). Studies of the Amazonian fishes: anoverview. Comp. Biochem. Physiol. 62A, 257–272.
Russu, I. & Ho, C. (1986) Assessment of roles ofb146-histidyl and other histidyl residues in the Bohreffect af human normal adult hemoglobin. Biochem-istry, 25, 1706–1716.
Shih, D. T., Luisi, B. F., Miyazaki, G., Perutz, M. F. &Nagai, K. (1993). A mutagenic study of the allostericlinkage of His(HC3)146b in haemoglobin. J. Mol. Biol.230, 1291–1296.
Silva, M. M., Rogers, P. H. & Arnone, A. (1992). A thirdquaternary structure of human hemoglobin A at1.7 A resolution. J. Mol. Biol. 267, 17248–17256.
Tame, J., Shih, D. T.-B., Pagnier, J., Fermi, G. & Nagai, K.(1991). The functional role of the distal valine (E11)residue of the a subunits in human haemoglobin.J. Mol. Biol. 218, 761–767.
Tan, A., De Young, A. & Noble, R. (1972). The pHdependence of the affinity, kinetics and cooperativityof ligand binding to carp hemoglobin. J. Biol. Chem.247, 2493–2498.
Weber, R. E. (1982). Intraspecific adaptation of hemo-globin function to oxygen availability. In Exogenousand Endogenous Influences on Metabolic and NeuralControl (Addink, A. D. F. & Spronk, N., eds), pp.87–102, Pergamon Press, Oxford and New York.
Weber, R. E. (1990). Functional significance and struc-tural basis of multiple hemoglobins with specialreference to ectothermic vertebrates. Comp. Physiol. 6,58–75.
Weber, R. E., Wood, S. & Lomholt, J. (1976). Temperatureacclimation and oxygen-binding properties of bloodand multiple haemoglobins of rainbow trout. J Expt.Biol. 65, 333–354.
Weber, R. E., Jessen, T.-H., Malte, H. & Tame, J. (1993).Mutant hemoglobins (a119-Ala and b55-Ser): functionsrelated to high altitude adaptation in geese. J. Appl.Physiol. 75, 2646–2655.
Zhang, J., Hua, Z., Tame, J. R. H., Lu, G., Zhang, R. & Gu,X. (1996). The crystal structure of a high affinityspecies of haemoglobin (bar-headed goose haemo-globin in the oxy form). J. Mol. Biol. 255, 484–493.
Edited by R. Huber
(Received 14 December 1995; received in revised form 25 March 1996; accepted 11 April 1996)
Note added in proof: The crystal structure of a Root effect Hb was recently solved and its functionalimplications discussed (Mylvaganam et al., 1996, Nature Struct. Biol. 3, 275–283).




















