
Diseases of Salivary Glands: Review - Al-Rafidain Dental ...
Upload
independentCategory
view
4download
0
Ann. appl. Biol. (1982), 100, 553-558 With I plate Printed in Great Britain
553
A study of glands on cotton responsible for the high pH and cation concentration of the leaf surface
BY CAROLE J. ELLEMAN AND PHILIP F. ENTWISTLE Institute of Virology, Natural Environment Research Council, Mansjield Road,
Oxford OX1 3SR
(Accepted I 4 January 1982)
S U M M A R Y
Groups of hygroscopic microcrystalline particles were found on the leaf surface of cotton, Gossypium hirsutum and were shown to be produced by glands in the epidermis. The glands were hydathode-like, about 20 pm in diameter by 25 pm long, with swollen, rounded, apices. They secreted high concentrations of magnesium and potassium and some sulphur and calcium, as indicated by X-ray microanalysis, onto the leaf surface. The proportions of elements present in gland secretions could be altered by changing the root environment of the plant. When the microcrystalline particles were removed and made up in distilled water the resulting solution had a pH of 10.2. It was established that the glands are responsible for the high pH and cation concentrations on the cotton leaf surface. The morphology and structure of the glands was investigated by light and electron microscopy and their possible function is discussed.
I N T R O D U C T I O N
The leaf surface of cotton was first reported as being alkaline in nature by Smith (1923). Since then many authors have referred to the high pH of the cotton leaf surface, cotton leaf dew and leaf washes (McCleod, Yearian & Young, 1977; Young, Yearian & Kim, 1977; Andrews & Sikorowski, 1973; Harr, Guggenheim, Boller & Oertli, 1980). Reported measurements of the pH vary from 7-4 to 10.7 and the alkalinity has been attributed to the presence of the basic cations magnesium, calcium and potassium which probably occur as carbonates and bicarbonates (Andrews & Sikorowski, 1973; Young et al., 1977). Both Young et al. (1977) and Harr et al. (1980) report that there are particularly high levels of magnesium present on the cotton leaf surface. Our interest in the conditions prevailing on the cotton leaf surface arose from work on the nuclear polyhedrosis virus (NPV) of larval Spodoptera littoralis Boisd. (Lepidoptera: Noctuidae); the virus is rapidly inactivated on the surface of cotton (by factors in addition to ultra violet light), as are other NPVs, both under laboratory and field conditions (Young et al., 1977; McCleod et a1.,1977; Bullock, 1967; Young & Yearian, 1974; Yearian & Young, 1974; Ignoffo et al., 1972; Roome & Daoust, 1976).
A study was made of the cotton leaf surface during which glands were found. Further investigation of these structures demonstrated that they were responsible for the high concentration of cations on the leaf surface and the high pH.
M A T E R I A L S A N D M E T H O D S
Cotton seed, Gossypium hirsutum L. variety Delta Pine 16, was obtained from the Delta and Pine Land Company, Scott, Mississippi 387722. Plants were grown in John Innes potting compost number 2 mixed with an equal part of sand in growth cabinets at a constant 0 1982 Association of Applied Biologists

554 C . J . E L L E M A N A N D P . F . E N T W I S T L E
temperature of 25 OC on a regime of 18 h daylight. For root environment experiments seed was germinated in vermiculite and watered with Sachs solutions (Harris Biologicals Ltd). The plants (lid not grow very well under these conditions and were used at a smaller size than the control plants.
Scanning electron microscopy (SEM) was carried out on a Jeol T20 microscope. the column of which is small enough to maintain a vacuum when fresh or partially dried leaf material is placed on the stage. However. for the best results specimens were either critical point dried and then gold sputtered before examination or simply sputtered without prior preparation. Critical point drying was performed by first fixing the leaf tissue in glutaraldehyde then dehydrating in an acetone series before placing the specimens in acetone in the critical point drying apparatus. X-ray microanalysis was carried out on a Jeol SJ35 with an EDAX attachment for microanalysis. Specimens for microanalysis received no prior treatment apart from coating with carbon, This permits the elemental analysis of extremely small areas of tissue. The technique is limited by the atomic weight of the element: the lowest atomic number that would have been detected was 23. i.e. Sodium. They were mounted on carbon-nylon grids.
Sections for light microscopy were prepared by fixing in glutaraldehyde, dehydrating in acetone followed by embedding in Epikote as for an electron microscope (EM) preparation. Sections were then cut (1-2 ,pm thick) using an LKB sectioner and a glass knife. The cut sections were placed on microscope slides and stained with 1% methylene blue and 1% azure in 1% borax prior to examination.
pH was measured using a glass electrode and a Philips PW9418 meter. Electrical conductivity was measured with a small pair of metal electrodes set 1 mm apart attached to an appropriate circuit and an Avometer. Readings were in mA and a calibration of the electrode using sodium chloride is shown in Text-fig. 1 .
R E S U L T S
When plants were first brought into the laboratory from the growth cabinet at 8-12 weeks of age it was noticed that the leaf surface was covered in what appeared to be dust. A closer examination using the light microscope revealed that the dust consisted of discrete particles tending to cluster round the veins of the leaf (Plate. fig. 1). The particles of dust were only stable in a dry atmosphere and readily dissolved to form damp patches on the leaf surface when the humidity was raised. e.g. by placing a leaf in a container with a piece of damp tissue paper.
When fresh leaf material was examined in the SEM the particles of dust could be clearly seen to be composed of aggregates of crystals the form of which (Plate. fig. 2 ) varied from one specimen to another (Plate, fig. 3). In some specimens. as in Plate. fig. 3. a structure was present which suggested that a gland lying underneath the multicrystalline particle might be responsible fo r its production. When leaf specimens were critical point dried before examination all traces of ‘dust’ were lost and the glands were clearly visible (Plate. fig. 4). Glands and their products were observed on all parts of the plant. upper and lower leaf surfaces, petioles and sepals. They were present even on the very youngest leaves although here they were much closer together and frequently had no crystalline particles associated with them. In addition, the glands on the younger leaves did not collapse under the electron beam and sometimes appeared as intact structures on specimens that had not been critical point dried (Plate. fig. 5). All glands appeared to be situated above veins or veinlets. were hydathode-like, about 20 pm in diameter by 25 pm long. with swollen, rounded apices.
Light microscope sections of the glands showed them to be multicellular organs at the base of which. in the epidermis. were several specialised cells (Plate. fig. 6).
When the crystalline particles were removed from the cotton leaf by brushing with a stiff brush. and dissolved in distilled water (16 mg/ml) the resulting solution had a pH of 10.2. This
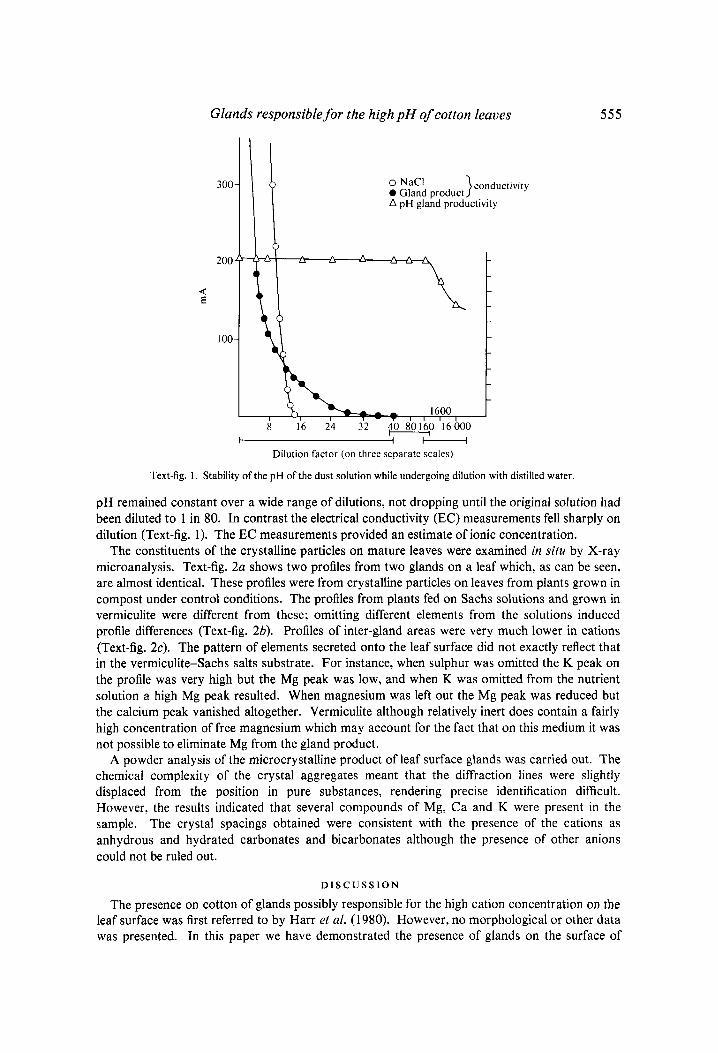
Glands responsible for the high pH of cotton leaves 5 5 5
300
200
d &
100
l b
o NaCl )conductivity 0 Gland product A pH gland productivity
I , , 16,001 8 16 24 32 40 80160 16000
t I t-----l Dilution factor (on three separate scales)
Text-fig. 1. Stability of the pH of the dust solution while undergoing dilution with distilled water.
pH remained constant over a wide range of dilutions, not dropping until the original solution had been diluted to 1 in 80. In contrast the electrical conductivity (EC) measurements fell sharply on dilution (Text-fig. 1). The EC measurements provided an estimate of ionic concentration.
The constituents of the crystalline particles on mature leaves were examined in situ by X-ray microanalysis. Text-fig. 2a shows two profiles from two glands on a leaf which, as can be seen, are almost identical. These profiles were from crystalline particles on leaves from plants grown in compost under control conditions. The profiles from plants fed on Sachs solutions and grown in vermiculite were different from these; omitting different elements from the solutions induced profile differences (Text-fig. 2b). Profiles of inter-gland areas were very much lower in cations (Text-fig. 2c). The pattern of elements secreted onto the leaf surface did not exactly reflect that in the vermiculite-Sachs salts substrate. For instance, when sulphur was omitted the K peak on the profile was very high but the Mg peak was low, and when K was omitted from the nutrient solution a high Mg peak resulted. When magnesium was left out the Mg peak was reduced but the calcium peak vanished altogether. Vermiculite although relatively inert does contain a fairly high concentration of free magnesium which may account for the fact that on this medium it was not possible to eliminate Mg from the gland product.
A powder analysis of the microcrystalline product of leaf surface glands was carried out. The chemical complexity of the crystal aggregates meant that the diffraction lines were slightly displaced from the position in pure substances, rendering precise identification difficult. However, the results indicated that several compounds of Mg, Ca and K were present in the sample. The crystal spacings obtained were consistent with the presence of the cations as anhydrous and hydrated carbonates and bicarbonates although the presence of other anions could not be ruled out.
D I S C U S S I O N
The presence on cotton of glands possibly responsible for the high cation concentration on the leaf surface was first referred to by Harr et al. (1 980). However, no morphological or other data was presented. In this paper we have demonstrated the presence of glands on the surface of

556
3 0 -
C . J . E L L E M A N A N D P . F . E N T W I S T L E
Mg SCI K C a
Gland I Gland 2
I M p S CI KCa
1 2 3 4 Electron \ 011s
Text-fig. 2. (a) An EDAX X-ray microanalysis profile of two separate dust particles on the surface of a mature cotton leaf from a plant grown in compost. (h) Three profiles of dust particles. each from plants grown in water culture in media deficient in S. Mg or K . (c) Two profiles from a leaf taken from a plant grown in water culture: one is from a dust particle. the other the background leaf material. The nutrient solution was complete.
cotton by light and electron microscopy and have shown their secretions to be rich in several cations. Sections of the glands revealed that they are multicellular organs originating from specialised cells in the epidermal layer of the leaf. The glands were found on all parts of the plant examined. on both old and young leaves although. except under conditions of extreme water stress, crystals were produced less readily by the young leaves. Some evidence was obtained that electrical conductivity (used as an indicator of ionic concentration) on the very young leaves was particularly high, seeming to indicate that these glands were active. Such higher conductivity may be due to the greater density of glands on leaves not fully expanded. The microcrystalline groups were hygroscopic. readily absorbing water and dissolving to form damp patches on the leaves when the atmosphere was humid.
The microcrystals produced by the glands when made up in solution had a pH of 10.2, which is slightly lower than values of leaf pH given by Harr et al. (1980) but higher than most other

Glands responsible for the high pH of cotton leaves 557
reported pH values for cotton dew or leaf washings. Measurements of pH and EC readings as the microcrystal solution was progressively diluted indicated that, on Delta Pine 16 grown under the conditions, the pH will remain constant and be the same on all leaves. This is in contrast to the results of Harr et al. (1980) who found that on some varieties of Gossypium barbadense L. the surface pH of the younger leaves was lower than the more mature leaves. This difference may be due to differing gland products. As demonstrated by use of Sachs solutions the root environment profoundly affects the elemental content of the gland secretions and this could account for (a) the variability in pH measurements for cotton leaf surfaces from different studies and (b) differences between leaves of different ages if the pH is more dependent on concentration when a different mixture of cations is present.
The purpose of the glands is not obvious. A high pH is inhibitory to fungal growth but may stimulate the growth of bacteria. Morphologically the glands bear a strong resemblance to the salt glands of Avicennia marina and the chalk secreting glands of Plumbago capensis (Shimony, Fahn & Reinhold, 1973; Sakai, 1970). On the salt marsh plants glands of this nature are responsible for maintaining the internal osmotic pressure of the plant at a physiological level against the concentration gradient caused by the salt water of the environment. This function is accomplished by pumping salts out of the plant onto the surface. It is possible that on cotton the function is similar, protecting the plant from internal high concentrations of cations when water is lost rapidly by transpiration in the semi-arid conditions which are its natural habitat. However, there is an alternative suggestion arising from the work of Mooney, Gulmon, Ehleringer & Rundel (1980). These authors found glands on Nolana mollis, a succulent leaved shrub of desert areas. The secretions of these glands were high in sodium, calcium and magnesium and were strongly hygroscopic. The deposits were able to absorb water from a non-saturated atmosphere which was then, at the expense of metabolic energy, available to the plant. The hygroscopic nature of the cotton gland product and the native habitat of cotton strongly suggest that their function may be similar to that of the glands of Nolana mollis, though it is also possible this is combined with an osmo-regulatory function.
Our many thanks are due to the Delta Pine Land Co. for the supply of cotton seed, to Jeol (UK) Ltd for assistance with X-ray crystallography, to Dr A. K. Cheetham, of the X-ray Crystallography Department, University of Oxford for help with powder analysis and to Mr M. Heslop of this Institute for constructing the circuit employed in measuring the electrical conductivity of cotton leaf surface fluids.
REFERENCES
ANDREWS, G. L. & SIKOROWSKI, P. P. ( 1 9 7 3 ) . Effects of cotton leaf surfaces on the polyhedrosis virus of Heliothis zea and Heliothis virescens (Lepidoptera: Noctuidae). Journal of Invertebrate Pathology 22,
BULLOCK, H. R. ( 1 9 6 7 ) . Persistence of Heliothis nuclear polyhedrosis virus on cotton foliage. Journal of 290-29 1.
Invertebrate Pathology 9 , 4 3 4 - 6 .
commercial cotton varieties. Coton et Fibres Tropicales 3 5 , 3 7 9 - 3 8 4 .
R. L. (1972). Field studies on the stability of the Heliothis nuclear polyhedrosis virus at various sites throughout the cotton belt. Environmental Entomology 1 , 3 8 8 - 3 9 0 .
MCCLEOD, P. J., YEARIAN, w. c . & YOUNG, s. Y. ( 1 9 7 7 ) . Inactivation ofBaculovirus heliothis by ultra violet irradiation, dew and temperature. Journal of Invertebrate Pathology 30,234-24 1.
MOONEY, H. A., GULMON, s. L., EHLERINGER, J. RUNDEL, P. w. ( 1 9 8 0 ) . Atmospheric water uptake by an Atacama Desert shrub. Science 2 0 9 , 6 9 3 - 6 9 4 .
ROOME, R. E. & DAOUST, R. A. ( 1 9 7 6 ) . Survival of the nuclear polyhedrosis of Heliothis armigera on crops and in soil in Botswana. Journal of Invertebrate Pathology 2 1 , 7 - 1 2 .
HARR, J., GUGGENHEIM, R., BOLLER, TH. & OERTLI, J. J. ( 1 9 8 0 ) . High pH-values on the leaf surface Of
IGNOFFO, C. M., BRADLEY, J . R. JR., GUILLARD, F. R., HARRIS, F. A., FALCON, L. A., LARSON, L. V. & MCGARR,

558 C . J . E L L E M A N A N D P . F . E N T W I S T L E
SAKAI, w. s. (1970). Scanning electron microscopy and energy dispersive X-ray analysis of chalk secreting leaf glands of Plumbago capensis. American Journal of Botany 61,94-99.
SHIMONY, c., FAHN, A. & REINHOLD, L. (1973). Ultrastructure and ion gradients in the salt glands of Aricennia marina (Forssk) Vierh. New Phytology 72, 27-36.
SMITH. c. M. (1923). Journd of Agricultural Research 26, 192-194.
YEARIAN, w. c. & YOUNG, s. Y . (1974). Persistence of Heliothis nuclear polyhedrosis virus on cotton plant parts. Entiironmental Entomology 3, 1035-1036.
YOUIVUC. s. Y. & YEARIAN, w. c. (1974). Persistence of Heliothis NPV on foliage of cotton, soybean and tomato. Enoironmental Entomology 3.253-5.
YOUNG, S. Y., YEARIAN. W. C. & KIM, K. s. (1977). Effect of dew from cotton and soybean foliage on activity of Heliothis NPV. Journal of Invertebrate Pathology 29, 105-1 11.
Excretion from leaves as a factor in arsenical injury to plants.
(Received 3 Nocember 1981)
EXPLANATION OF PLATE
Fig. 1. A photomicrograph of the upper leaf surface of cotton taken through a light microscope. (Magnification =
Fig. 2. An SEM micrograph of the upper surface of a mature cotton leaf. (Specimen preparation: gold sputtered, magnification = 4 9 0 0 ~ . ~ = 2 Fig. 3. An SEM micrograph of the upper surface of a mature cotton leaf. (Specimen preparation: none. magnification :- 1 5 0 0 ~ . - - = 5 p 1 2 . 5 k V . ) G = G l a n d .
Fig. 4. An SEM micrograph of the upper surface of a cotton leaf showing an intact gland. (Specimen preparation: critical point dried and gold sputtered, magnification = 1687 x . ~ = 5 p 2 0 kV.)
Fig. 5 . An SEM micrograph of a young cotton leaf showing several intact glands placed quite close together and many leaf hairs. (Specimen preparation: gold sputtered. magnification = 7 5 0 x . ~ = lop.)
Fig. 6. A light microscope section through a gland on a young cotton leaf showing the multicellular structure. (Specimen preparation: fixed in glutaraldehyde, dehydrated in acetone. embedded in epikote, stained with methylene blue. azure and borax. magnification = 2 4 0 0 ~ . ~ = 5 p.)
2 4 ~ , - = 5 0 0 p . )
12.5 kV.)

Annals of Applied Biology, Vol. 100, No. 3
CAROLE J. ELLEMAN AND PHILIP F. ENTWISTLE
Plate I
(Fucingp. 558)
Copyright © 2022 FDOKUMEN




















