
EBTAX: The Conversion of Ethane to Aromatics via Catalytic ...
Upload
independentCategory
view
3download
0
Fungal Genetics and Biology 74 (2015) 32–44
Contents lists available at ScienceDirect
Fungal Genetics and Biology
journal homepage: www.elsevier .com/ locate/yfgbi
The old 3-oxoadipate pathway revisited: New insights in the catabolismof aromatics in the saprophytic fungus Aspergillus nidulans
http://dx.doi.org/10.1016/j.fgb.2014.11.0021087-1845/� 2014 Elsevier Inc. All rights reserved.
⇑ Corresponding author at: Instituto de Tecnologia Química e Biológica, Univer-sidade Nova de Lisboa, Av. da República, EAN, 2781-901 Oeiras, Portugal. Fax: +351214411277.
E-mail address: [email protected] (C. Silva Pereira).
Tiago M. Martins a, Diego O. Hartmann a, Sébastien Planchon b, Isabel Martins a, Jenny Renaut b,Cristina Silva Pereira a,⇑a Instituto de Tecnologia Química e Biológica, Universidade Nova de Lisboa, Av. da República, 2780-157 Oeiras, Portugalb Proteomics Platform, Centre de Recherche Public – Gabriel Lippmann, Belvaux, Luxembourg
a r t i c l e i n f o
Article history:Received 19 June 2014Accepted 23 November 2014Available online 3 December 2014
Keywords:Aromatic compounds catabolismAspergillus nidulans3-Oxoadipate pathwayProtocatechuate/catechol branchesSaprophytic Ascomycota fungi
a b s t r a c t
Aspergilli play major roles in the natural turnover of elements, especially through the decomposition ofplant litter, but the end catabolism of lignin aromatic hydrocarbons remains largely unresolved. The 3-oxoadipate pathway of their degradation combines the catechol and the protocatechuate branches, eachusing a set of specific genes. However, annotation for most of these genes is lacking or attributed topoorly- or un-characterised families. Aspergillus nidulans can utilise as sole carbon/energy source eitherbenzoate or salicylate (upstream aromatic metabolites of the protocatechuate and the catechol branches,respectively). Using this cultivation strategy and combined analyses of comparative proteomics, genemining, gene expression and characterisation of particular gene-replacement mutants, we preciselyassigned most of the steps of the 3-oxoadipate pathway to specific genes in this fungus. Our findingsdisclose the genetically encoded potential of saprophytic Ascomycota fungi to utilise this pathway andprovide means to untie associated regulatory networks, which are vital to heightening their ecologicalsignificance.
� 2014 Elsevier Inc. All rights reserved.
1. Introduction
Aspergillus species are widespread in terrestrial habitats wherethey have an important role in the natural cycling of chemical ele-ments, particularly in the decomposition of plant litter. These fun-gal saprophytes can, in general, efficiently degrade complex plantpolymers including starches, hemicelluloses, celluloses, pectinsand other sugar polymers. Quinate and shikimate, comprisingmore than 10 wt% of decaying leaf litter, are other abundant carbonsources for aspergilli. The aromatic protocatechuate is the endintermediate of their utilisation pathways – well studied in bothAspergillus nidulans and Neurospora crassa (Hawkins et al., 1993;Tang et al., 2011). Plant lignin-related aromatic hydrocarbons,which constitute also an abundant carbon source for saprophyticfungi, are usually considered inhibitory compounds in the decom-position of leaf litter and fermentation of lignocellulosic hydroly-sates (Palmqvist and Hahn-Hägerdal, 2000).
The peripheral (upper) pathways of the aerobic catabolism ofaromatic hydrocarbons have been extensively investigated in fungi
but the subsequent central pathways are still poorly understood(Wright, 1993). They usually occur ultimately via one of five inter-mediates, either catechol, protocatechuate, homogentisate, genti-sate or hydroquinone (Harwood and Parales, 1996). Thehomogentisate degradation pathway (involved e.g. in the phenylal-anine metabolism) has been recently resolved in Penicillium chrys-ogenum (Veiga et al., 2012). On the contrary, the 3-oxoadipatepathway that combines the protocatechuate and the catecholbranches, remains largely overlooked, notwithstanding its signifi-cance in the end degradation of numerous exogenous aromatichydrocarbons (Harwood and Parales, 1996). It is generally acceptedthat the composing biochemical reactions of this pathway in bac-teria and fungi are rather well conserved, despite some distin-guishable features (Harwood and Parales, 1996). The twobranches of the pathway progress mostly through identical reac-tion steps that yield chemically analogous intermediates. Theyconverge with the formation of 3-oxoadipate (b-ketoadipate)which is channelled into the tricarboxylic acid (TCA) cycle. Estab-lished knowledge on aspergilli revealed that they are able to useboth branches of the 3-oxoadipate pathway using exclusivelybranch-specific genes (Boschloo et al., 1990; Kuswandi andRoberts, 1992). However, most reaction steps of this pathwayeither lack the assigned gene or are functionally assigned to multi-

T.M. Martins et al. / Fungal Genetics and Biology 74 (2015) 32–44 33
ple genes that encode poorly- or un-characterised protein familiesoften predicted based on ‘‘nebulous’’ homologies to their bacterialcounterparts. For example, the 3-carboxy-cis,cis-muconate inter-mediate (protocatechuate branch) undergoes specific cycloisomer-isation in bacteria and fungi (Harwood and Parales, 1996) but thegenes currently associated to the downstream reactions in fungidisregards this fact. Better understanding of the aromatic degrada-tion pathways in fungi can push the technological development ofnovel biorefinery and bioremediation strategies (Palmqvist andHahn-Hägerdal, 2000).
The present contribution aims to elucidate the 3-oxoadipatepathway in the model fungus A. nidulans. Combined comparativeproteomics, gene expression and functional studies were under-taken using either benzoate or salicylate as sole carbon/energysource, in comparison with growth on acetate (control). Benzoateand salicylate constitute, respectively, the upstream aromaticmetabolites of the protocatechuate and the catechol branches ofthe 3-oxoadipate pathway. This strategy allowed to assigning mostof the steps of this pathway to specific genes. Our findings spotteda few incorrect conceptions predicted from bacterial studies andstrengthened the significance of fundamental studies on modelfungi/yeast.
2. Materials and methods
2.1. Strain and growth conditions
A. nidulans FGSC A4 asexual spores were harvested and main-tained as frozen suspensions at �80 �C (Martins et al., 2014b). Cul-tures were initiated with 105 spores/mL and incubated with orbitalagitation (160 rpm) in the dark at 27 �C. Batch cultivations wereperformed in 250 mL screw thread flasks with a working volumeof 50 mL. A low nitrogen minimal medium was used containingper litre 1 g K2HPO4, 3 g NaNO3, 0.01 g ZnSO4�7H2O, 0.005 g CuSO4-
�5H2O, 0.5 g MgSO4�7H2O, 0.01 g FeSO4�7H2O, 0.5 g KCl. Filter ster-ilised salts were added to an autoclave sterilised phosphate (pH7.0) solution. The carbon sources were added either directly tothe phosphate solution for sterilisation (150 mM sodium acetate– control) or to the mineral media after filter sterilisation(50 mM sodium benzoate or sodium salicylate). A. nidulans FGSCA1145 and A1147 (as well as the gene replacement mutants) usedsimilar cultivation conditions but media were supplemented withthe appropriate nutrients requirements (Table 1).
2.2. Metabolites identification and quantification
Culture media were analysed by ultra-performance liquid chro-matography (UPLC) for the identification and/or quantification ofthe aromatic compounds and their degradation intermediates(Martins et al., 2014b). The chromatographic profiles wereacquired at 228 nm and 204 nm, respectively for the quantification
Table 1List of Aspergillus nidulans strains used in this study.
Strain Relevant phenotype Source
A4 Glasgow wild type; veA+ FGSCa
A1145 pyrG89; pyroA4; nkuA::argB; riboB2 FGSCa
A1147 pyrG89; argB2; pabaB22; nkuA::argB; riboB2 FGSCa
DOH4.7 DAN4531::pyrGAf; pyrG89; argB2; pabaB22; nku::argB;riboB2
This study
DOH9.1 DAN3895::pyrGAf; pyrG89; pyroA4; nkuA::argB; riboB2 This studyDOH11.2 DAN4061::pyrGAf; pyrG89; pyroA4; nkuA::argB; riboB2 This studyDOH12.2 DAN1151::pyrGAf; pyrG89; pyroA4; nkuA::argB; riboB2 This studyDOH13.5 DAN5232::pyrGAf; pyrG89; pyroA4; nkuA::argB; riboB2 This study
a Fungal Genetics Stock Center, Kansas City, MO, USA.
of benzoate (retention time, Rt: 3.20 min) and salicylate (Rt:3.44 min). Further identifications, usually at 212 nm or 260 nm,used commercial standards of protocatechuate, 4-hydroxybenzo-ate, catechol, 2,3-dihydroxybenzoate and cis,cis-muconate (Rt:1.81; 2.18; 2.40; 2.54 and 1.83, respectively), as well as standardsof cis,trans-muconate, trans,trans-muconate and muconolactoneprepared using standard synthesis methods (Elvidge et al., 1950)(Rt: 2.04; 1.71 and 1.40 min, respectively). 3-Carboxymuconolac-tone (Rt: 1.24) and 3-carboxy-cis,cis-muconate (Rt: 1.34) wereputatively identified based on their absorption spectra.
2.3. Protein samples preparation
Mycelial and extracellular proteins were obtained and quanti-fied as previously described (Carvalho et al., 2013; Martins et al.,2014a). Mycelial (100 mg powder mixed with 20 mgpoly(vinyl)polypyrrolidone) and extracellular (concentrated using10 kDa centrifugal devices) proteins were precipitated with coldacetone containing 10% w/v trichloroacetic acid and 60 mM dithi-othreitol (DTT). The washed pellets were dissolved in similar buf-fers that contain 7 M urea, 2 M thiourea, 4% w/v CHAPS, 60 mMDTT and 1.0% v/v of matching IPG buffer (GE Healthcare), withaddition of 1% (w/v) Triton X-100 in the case of extracellular pro-teins, and finally clarified by centrifugation.
2.4. Two-dimensional gel electrophoresis (2DE)
200 lg of mycelial and 50 lg of extracellular proteins wereloaded to 13 cm IPG strips pH 3–10 NL and pH 3–5.6 NL, respec-tively. Protein isoelectric focusing, electrophoresis, staining, imageacquisition and analysis followed previously described methods(Carvalho et al., 2013; Martins et al., 2014a). The gels were stainedwith flamingo dye (Bio-Rad) or colloidal coomassie blue for imageacquisition or spot excision, respectively. Average values of thenormalised volumes from the biological triplicates of each growthcondition determined the up-accumulation level of each proteinand were used for one-way ANOVA analysis with Progenesis Same-Spots software v2.0 (Nonlinear Dynamics, UK). The level of signif-icance was defined at p-value < 0.05.
2.5. Protein identification
The differential protein spots were excised from gels manually,and processed using the Ettan Digester robot of the Ettan Spot Han-dling Workstation (GE Healthcare) (Carvalho et al., 2013; Martinset al., 2014a). Samples (0.7 lL) were then spotted on MALDI-TOFtarget plates (Applied Biosystems), before the deposit of 0.7 lLCHCA (7 mg mL�1 in ACN 50%, TFA 0.1%). Peptide mass determina-tions were carried out using the AB SCIEX TOF/TOF™ 5800 System(AB SCIEX). Both peptide mass fingerprinting and tandem MS inreflectron mode analyses were performed. The ten most intensepeaks were selected automatically and used for MS/MS. Calibrationwas carried out with the peptide mass calibration kit for 4700(Applied Biosystems). Proteins were identified, with ProteinPilot,by searching against the NCBI nr database downloaded on the18/01/2013 (restricted to fungi taxa, 1635212 sequences) withMascot v2.3 (Matrix Science). All searches were executed allowingfor a mass window of 100 ppm for the precursor mass and 0.5 Dafor fragment ion masses. Search parameters allowed for carboxy-amidomethylation of cysteine as fixed modification. Methylationof aspartate or glutamate, and oxidation of methionine and trypto-phan (single oxidation, double oxidation and kynurenin) were setas variable modifications. Homology identification was retainedwith probability set at 95%. All identifications were confirmedmanually.

Tabl
e2
Gen
esas
sign
edin
the
3-ox
oadi
pate
path
way
and
uppe
rde
grad
atio
npa
thw
ays
inA
sper
gillu
sni
dula
ns.M
ycel
ialp
rote
ins
iden
tifi
edin
the
2DE
diff
eren
tial
lyac
cum
ulat
edpr
otei
nsp
ots
(fol
dch
ange
,FC)
inBe
nzoa
te(B
z)or
Salic
ylat
e(S
c)an
dex
pres
sion
leve
lsof
sele
cted
tran
scri
pts
mea
sure
dby
qRT-
PCR.
Prev
ious
prot
ein
anno
tati
onna
mes
init
alic
s.
Gen
eco
dePr
otei
n(E
Cn
um
ber)
Nam
ein
the
data
base
Myc
elia
lpr
otei
nsp
ots
Tran
scri
pt
Ass
ign
edn
ame
FCpI
/MW
(kD
a)p
Val
ue
Nos
.FC
Bz
ScTh
eo.
Exp.
Bz
Sc
Benz
oate
degr
adat
ion
and
prot
ocat
echu
ate
bran
chof
the
3-ox
oadi
pate
path
way
AN
1095
0B
zuA
,ben
zoat
e4-
mon
ooxy
gen
ase
(1.1
4.13
.12)
––
––
––
48.0
3.3
AN
1095
24-
hyd
roxy
ben
zoat
e-3-
mon
ooxy
gen
ase
(1.1
4.13
.2)
FAD
mon
ooxy
gena
se,p
utat
ive
––
––
––
79.8
3.3
AN
0876
NA
DPH
flav
inre
duct
ase
(1.5
.1.3
0)co
nser
ved
hypo
thet
ical
prot
ein
8.8
–8.
3/34
5.5/
337.
07e�
06
3121
25.4
1.5
AN
8566
prot
ocat
ech
uat
e-3,
4-di
oxyg
enas
e(1
.13.
11.3
)ca
tech
oldi
oxyg
enas
e,pu
tati
ve34
.5–
5.4/
355.
4/35
7.86
e�0
529
73a
90.0
1.8
AN
1151
carb
oxy-
cis,
cis-
mu
con
ate
cycl
ase
(5.5
.1.5
)ca
rbox
y-ci
s,ci
s-m
ucon
ate
cycl
ase,
puta
tive
4.3
–5.
8/34
6.1/
433.
00e�
02
2304
a26
.52.
6A
N52
323-
carb
oxym
uco
nol
acto
ne
hyd
rola
se(d
ecar
boxy
lati
ng)
(3.1
.1.-
)4-
carb
oxym
ucon
olac
tone
deca
rbox
ylas
efa
mily
prot
ein
4.1
–7.
8/26
5.4/
261.
60e�
04
4068
9.1
1.4
AN
1049
5su
ccin
yl-C
oA:3
-oxo
adip
ate
CoA
-tra
nsf
eras
e(2
.8.3
.6)
coen
zym
eA
tran
sfer
ase,
puta
tive
2.0
–6.
2/55
5.5/
563.
80e�
02
1489
12.1
5.6
Salic
ylat
ede
grad
atio
nan
dca
tech
olbr
anch
ofth
e3-
oxoa
dipa
tepa
thw
ayA
N21
14sa
licy
late
1-m
onoo
xyge
nas
e(d
ecar
boxy
lati
ng)
(1.1
4.13
.1)
salic
ylat
ehy
drox
ylas
e,pu
tati
ve–
5.5
6.4/
496.
5/54
1.10
e�0
216
181.
611
.9A
N74
18ph
enol
2-m
onoo
xyge
nas
e(1
.14.
13.7
)ph
enol
2-m
onoo
xyge
nase
,put
ativ
e–
9.2
6.1/
786.
4/22
8.55
e�0
446
291.
113
3.4
AN
6723
2,3-
dih
ydro
xybe
nzo
ate
carb
oxy-
lyas
e2,
3-di
hydr
oxyb
enzo
ate
carb
oxyl
yase
,put
ativ
e–
––
––
–2.
110
.9A
N45
32ca
tech
ol1,
2-di
oxyg
enas
e(1
.13.
11.1
)ca
tech
oldi
oxyg
enas
e,pu
tati
ve–
11.2
5.3/
335.
3/31
7.14
e�0
534
20a
1.0
43.9
AN
1895
Mai
A,m
aley
lace
toac
etat
eis
omer
ase
––
––
–�
1.0
2.1
AN
3895
mu
con
ate
isom
eras
e(C
oAtr
ansf
eras
efa
mil
yII
I)(5
.-.-
.-)
CAIB
/BA
IFfa
mily
enzy
me
––
––
––
�1.
23.
1A
N40
61m
uco
nol
acto
ne
isom
eras
e(5
.3.3
.4)
cons
erve
dhy
poth
etic
alpr
otei
n–
5.0
6.4/
166.
2/14
4.00
e�0
356
962.
438
.6A
N45
313-
oxoa
dipa
teen
ol-l
acto
ne
hyd
rola
se(3
.1.1
.24)
alph
a/be
tahy
drol
ase,
puta
tive
–1.
65.
0/56
6.2/
273.
60e�
02
6987
b1.
119
.3
aSa
me
iden
tifi
cati
onin
mu
ltip
lesp
ots:
[297
3,29
44,3
318,
4512
,286
9,29
15,3
182]
;[2
304,
2380
];[3
420,
3483
,463
7,32
33,3
163,
3104
,325
7,30
94,4
218,
5201
].b
Spot
sth
atre
trie
ved
mor
eth
anon
epr
otei
nid
enti
fica
tion
.
34 T.M. Martins et al. / Fungal Genetics and Biology 74 (2015) 32–44
2.6. Quantitative real-time PCR analysis (qRT-PCR)
Based on the AspGD sequences of A. nidulans genes (Arnaudet al., 2012) (www.aspergillusgenome.org), oligonucleotide pairswere designed using the GeneFisher2 web tool (http://bibi-serv.techfak.uni-bielefeld.de/genefisher2) (SupplementalTable S1). The qRT-PCR analysis was performed in a CFX96 ThermalCycler (Bio-Rad), using the SsoFast EvaGreen Supermix (Bio-Rad),250 nM of each oligonucleotide and the cDNA template equivalentto 5 ng of total RNA, at a final volume of 10 lL per well, in threetechnical and four biological replicates. The PCR conditions were:enzyme activation at 95 �C for 30 s; 40 cycles of denaturation at95 �C for 10 s and annealing/extension at 60 �C (59 �C forAN1151, AN10950 and AN10952) for 15 s; and melting curveobtained from 65 �C to 95 �C, consisting of 0.5 �C increments for5 s. Data analyses were performed using the CFX Manager software(Bio-Rad). The expression of each gene was taken as the relativeexpression in pair-wise comparisons of each condition relative tothe acetate control. The expression of all target genes was norma-lised by the expression of the c-actin gene, AN6542, selected asinternal control due to its constant levels in all conditions.
2.7. Creation of gene-replacement mutants
The following genes: AN1151, AN3895, AN4061, AN4531 orAN5232, were selected for gene replacement with Aspergillusfumigatus pyrG (pyrGAf) in A. nidulans pyrG89 strains (Table 1). Genereplacement was performed according to previously establishedmethods (Szewczyk et al., 2006). Briefly, the �1 kb flaking regionsof each gene coding sequence were amplified with primers P1 andP3, for 50-flank, or P4 and P6, for 30-flank, specifically designed foreach gene target (primers list in Supplemental Table S2). The pyrGAf
cassette was produced by amplification of the sequence from plas-mid pCDS60 (FGSC, Kansas City, MO, USA) with primers CDS164and CDS165. Fusion PCR of the flanking regions with the pyrGAf cas-sette were performed using nested primers P2 and P5 designed foreach gene target. PCR reactions for the 50 and 30 flanking regions ofeach gene target and the pyrGAf cassette were performed with aNZYProof DNA polymerase, whereas fusion PCR used NZYLongDNA polymerase (NZYTech) instead (MyCycler thermal cycler,Bio-Rad). The purified PCR fusion products (excised from the aga-rose gel electrophoresis and purified with NZYGelPure-NZYTech)were used to transform protoplasts of A. nidulans strain A1145(for AN1151, AN3895, AN4061 and AN5232) or strain A1147 (forAN4531) as previously described (Szewczyk et al., 2006). GenomicDNA purification (by UltraClean Microbial DNA Isolation Kit,MoBio) and additional PCR reactions were done to further confirmthe success of each transformation.
2.8. Gene-replacement mutants’ functional studies
The gene-replacement mutants used in this study (Table 1)were grown onto solid media (minimal mineral media jellifiedwith 15 g/L agar) supplemented with acetate (150 mM), benzoateor salicylate (50 mM), during 6 days. Corresponding batch culturesin six well plates (working volume of 4 mL per well, 90 rpm) weredone as previously described (Section 2.1), supplementing theminimal mineral media with acetate (control) or either aromaticcompound alone or in combination with acetate. In addition,mycelia (wild type, DAN1151, DAN5232), previously grown in ace-tate liquid media for 4 days, were exposed to benzoate (10 mM) for3 days. At the end of the cultivation time metabolites produced inany of the liquid cultures were analysed as described in Section 2.2.
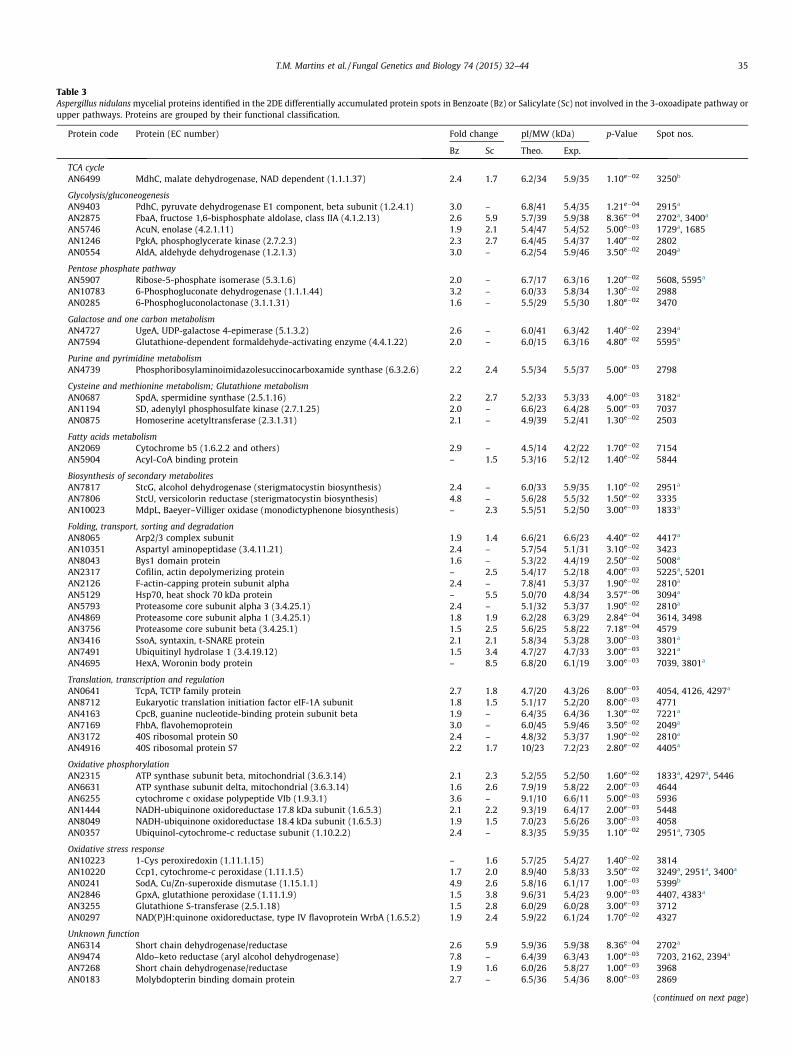
Table 3Aspergillus nidulans mycelial proteins identified in the 2DE differentially accumulated protein spots in Benzoate (Bz) or Salicylate (Sc) not involved in the 3-oxoadipate pathway orupper pathways. Proteins are grouped by their functional classification.
Protein code Protein (EC number) Fold change pI/MW (kDa) p-Value Spot nos.
Bz Sc Theo. Exp.
TCA cycleAN6499 MdhC, malate dehydrogenase, NAD dependent (1.1.1.37) 2.4 1.7 6.2/34 5.9/35 1.10e�02 3250b
Glycolysis/gluconeogenesisAN9403 PdhC, pyruvate dehydrogenase E1 component, beta subunit (1.2.4.1) 3.0 – 6.8/41 5.4/35 1.21e�04 2915a
AN2875 FbaA, fructose 1,6-bisphosphate aldolase, class IIA (4.1.2.13) 2.6 5.9 5.7/39 5.9/38 8.36e�04 2702a, 3400a
AN5746 AcuN, enolase (4.2.1.11) 1.9 2.1 5.4/47 5.4/52 5.00e�03 1729a, 1685AN1246 PgkA, phosphoglycerate kinase (2.7.2.3) 2.3 2.7 6.4/45 5.4/37 1.40e�02 2802AN0554 AldA, aldehyde dehydrogenase (1.2.1.3) 3.0 – 6.2/54 5.9/46 3.50e�02 2049a
Pentose phosphate pathwayAN5907 Ribose-5-phosphate isomerase (5.3.1.6) 2.0 – 6.7/17 6.3/16 1.20e�02 5608, 5595a
AN10783 6-Phosphogluconate dehydrogenase (1.1.1.44) 3.2 – 6.0/33 5.8/34 1.30e�02 2988AN0285 6-Phosphogluconolactonase (3.1.1.31) 1.6 – 5.5/29 5.5/30 1.80e�02 3470
Galactose and one carbon metabolismAN4727 UgeA, UDP-galactose 4-epimerase (5.1.3.2) 2.6 – 6.0/41 6.3/42 1.40e�02 2394a
AN7594 Glutathione-dependent formaldehyde-activating enzyme (4.4.1.22) 2.0 – 6.0/15 6.3/16 4.80e�02 5595a
Purine and pyrimidine metabolismAN4739 Phosphoribosylaminoimidazolesuccinocarboxamide synthase (6.3.2.6) 2.2 2.4 5.5/34 5.5/37 5.00e�03 2798
Cysteine and methionine metabolism; Glutathione metabolismAN0687 SpdA, spermidine synthase (2.5.1.16) 2.2 2.7 5.2/33 5.3/33 4.00e�03 3182a
AN1194 SD, adenylyl phosphosulfate kinase (2.7.1.25) 2.0 – 6.6/23 6.4/28 5.00e�03 7037AN0875 Homoserine acetyltransferase (2.3.1.31) 2.1 – 4.9/39 5.2/41 1.30e�02 2503
Fatty acids metabolismAN2069 Cytochrome b5 (1.6.2.2 and others) 2.9 – 4.5/14 4.2/22 1.70e�02 7154AN5904 Acyl-CoA binding protein – 1.5 5.3/16 5.2/12 1.40e�02 5844
Biosynthesis of secondary metabolitesAN7817 StcG, alcohol dehydrogenase (sterigmatocystin biosynthesis) 2.4 – 6.0/33 5.9/35 1.10e�02 2951a
AN7806 StcU, versicolorin reductase (sterigmatocystin biosynthesis) 4.8 – 5.6/28 5.5/32 1.50e�02 3335AN10023 MdpL, Baeyer–Villiger oxidase (monodictyphenone biosynthesis) – 2.3 5.5/51 5.2/50 3.00e�03 1833a
Folding, transport, sorting and degradationAN8065 Arp2/3 complex subunit 1.9 1.4 6.6/21 6.6/23 4.40e�02 4417a
AN10351 Aspartyl aminopeptidase (3.4.11.21) 2.4 – 5.7/54 5.1/31 3.10e�02 3423AN8043 Bys1 domain protein 1.6 – 5.3/22 4.4/19 2.50e�02 5008a
AN2317 Cofilin, actin depolymerizing protein – 2.5 5.4/17 5.2/18 4.00e�03 5225a, 5201AN2126 F-actin-capping protein subunit alpha 2.4 – 7.8/41 5.3/37 1.90e�02 2810a
AN5129 Hsp70, heat shock 70 kDa protein – 5.5 5.0/70 4.8/34 3.57e�06 3094a
AN5793 Proteasome core subunit alpha 3 (3.4.25.1) 2.4 – 5.1/32 5.3/37 1.90e�02 2810a
AN4869 Proteasome core subunit alpha 1 (3.4.25.1) 1.8 1.9 6.2/28 6.3/29 2.84e�04 3614, 3498AN3756 Proteasome core subunit beta (3.4.25.1) 1.5 2.5 5.6/25 5.8/22 7.18e�04 4579AN3416 SsoA, syntaxin, t-SNARE protein 2.1 2.1 5.8/34 5.3/28 3.00e�03 3801a
AN7491 Ubiquitinyl hydrolase 1 (3.4.19.12) 1.5 3.4 4.7/27 4.7/33 3.00e�03 3221a
AN4695 HexA, Woronin body protein – 8.5 6.8/20 6.1/19 3.00e�03 7039, 3801a
Translation, transcription and regulationAN0641 TcpA, TCTP family protein 2.7 1.8 4.7/20 4.3/26 8.00e�03 4054, 4126, 4297a
AN8712 Eukaryotic translation initiation factor eIF-1A subunit 1.8 1.5 5.1/17 5.2/20 8.00e�03 4771AN4163 CpcB, guanine nucleotide-binding protein subunit beta 1.9 – 6.4/35 6.4/36 1.30e�02 7221a
AN7169 FhbA, flavohemoprotein 3.0 – 6.0/45 5.9/46 3.50e�02 2049a
AN3172 40S ribosomal protein S0 2.4 – 4.8/32 5.3/37 1.90e�02 2810a
AN4916 40S ribosomal protein S7 2.2 1.7 10/23 7.2/23 2.80e�02 4405a
Oxidative phosphorylationAN2315 ATP synthase subunit beta, mitochondrial (3.6.3.14) 2.1 2.3 5.2/55 5.2/50 1.60e�02 1833a, 4297a, 5446AN6631 ATP synthase subunit delta, mitochondrial (3.6.3.14) 1.6 2.6 7.9/19 5.8/22 2.00e�03 4644AN6255 cytochrome c oxidase polypeptide VIb (1.9.3.1) 3.6 – 9.1/10 6.6/11 5.00e�03 5936AN1444 NADH-ubiquinone oxidoreductase 17.8 kDa subunit (1.6.5.3) 2.1 2.2 9.3/19 6.4/17 2.00e�03 5448AN8049 NADH-ubiquinone oxidoreductase 18.4 kDa subunit (1.6.5.3) 1.9 1.5 7.0/23 5.6/26 3.00e�03 4058AN0357 Ubiquinol-cytochrome-c reductase subunit (1.10.2.2) 2.4 – 8.3/35 5.9/35 1.10e�02 2951a, 7305
Oxidative stress responseAN10223 1-Cys peroxiredoxin (1.11.1.15) – 1.6 5.7/25 5.4/27 1.40e�02 3814AN10220 Ccp1, cytochrome-c peroxidase (1.11.1.5) 1.7 2.0 8.9/40 5.8/33 3.50e�02 3249a, 2951a, 3400a
AN0241 SodA, Cu/Zn-superoxide dismutase (1.15.1.1) 4.9 2.6 5.8/16 6.1/17 1.00e�03 5399b
AN2846 GpxA, glutathione peroxidase (1.11.1.9) 1.5 3.8 9.6/31 5.4/23 9.00e�03 4407, 4383a
AN3255 Glutathione S-transferase (2.5.1.18) 1.5 2.8 6.0/29 6.0/28 3.00e�03 3712AN0297 NAD(P)H:quinone oxidoreductase, type IV flavoprotein WrbA (1.6.5.2) 1.9 2.4 5.9/22 6.1/24 1.70e�02 4327
Unknown functionAN6314 Short chain dehydrogenase/reductase 2.6 5.9 5.9/36 5.9/38 8.36e�04 2702a
AN9474 Aldo–keto reductase (aryl alcohol dehydrogenase) 7.8 – 6.4/39 6.3/43 1.00e�03 7203, 2162, 2394a
AN7268 Short chain dehydrogenase/reductase 1.9 1.6 6.0/26 5.8/27 1.00e�03 3968AN0183 Molybdopterin binding domain protein 2.7 – 6.5/36 5.4/36 8.00e�03 2869
(continued on next page)
T.M. Martins et al. / Fungal Genetics and Biology 74 (2015) 32–44 35
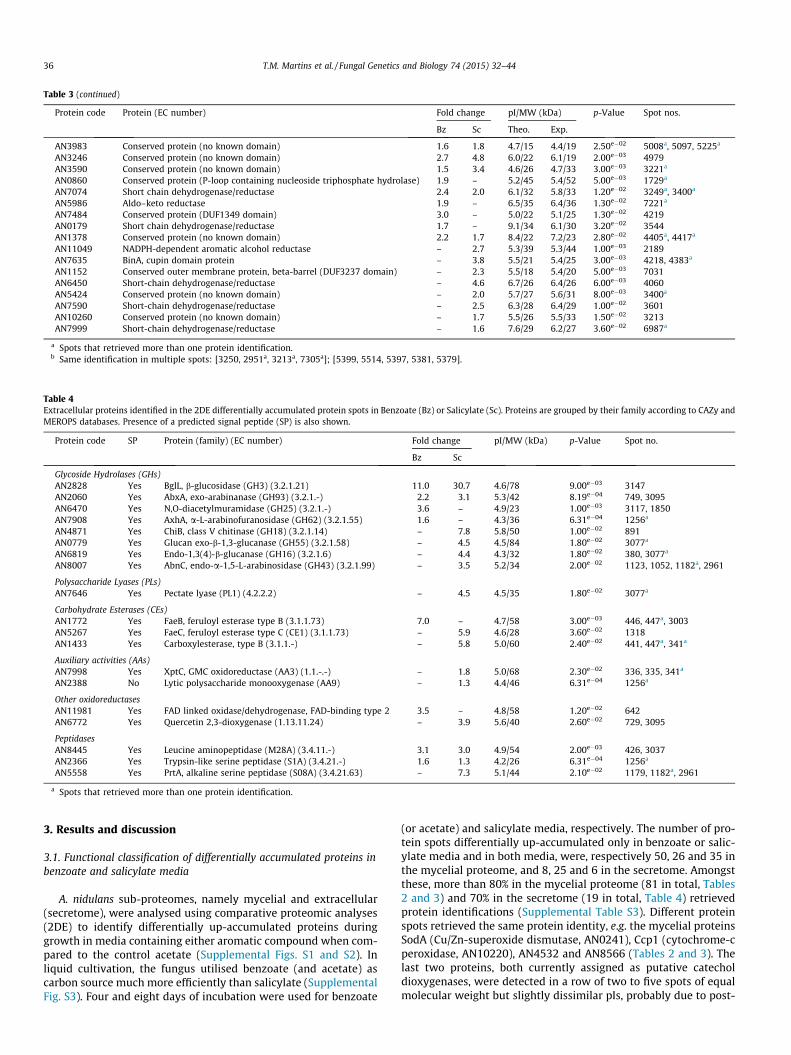
Table 3 (continued)
Protein code Protein (EC number) Fold change pI/MW (kDa) p-Value Spot nos.
Bz Sc Theo. Exp.
AN3983 Conserved protein (no known domain) 1.6 1.8 4.7/15 4.4/19 2.50e�02 5008a, 5097, 5225a
AN3246 Conserved protein (no known domain) 2.7 4.8 6.0/22 6.1/19 2.00e�03 4979AN3590 Conserved protein (no known domain) 1.5 3.4 4.6/26 4.7/33 3.00e�03 3221a
AN0860 Conserved protein (P-loop containing nucleoside triphosphate hydrolase) 1.9 – 5.2/45 5.4/52 5.00e�03 1729a
AN7074 Short chain dehydrogenase/reductase 2.4 2.0 6.1/32 5.8/33 1.20e�02 3249a, 3400a
AN5986 Aldo–keto reductase 1.9 – 6.5/35 6.4/36 1.30e�02 7221a
AN7484 Conserved protein (DUF1349 domain) 3.0 – 5.0/22 5.1/25 1.30e�02 4219AN0179 Short chain dehydrogenase/reductase 1.7 – 9.1/34 6.1/30 3.20e�02 3544AN1378 Conserved protein (no known domain) 2.2 1.7 8.4/22 7.2/23 2.80e�02 4405a, 4417a
AN11049 NADPH-dependent aromatic alcohol reductase – 2.7 5.3/39 5.3/44 1.00e�03 2189AN7635 BinA, cupin domain protein – 3.8 5.5/21 5.4/25 3.00e�03 4218, 4383a
AN1152 Conserved outer membrane protein, beta-barrel (DUF3237 domain) – 2.3 5.5/18 5.4/20 5.00e�03 7031AN6450 Short-chain dehydrogenase/reductase – 4.6 6.7/26 6.4/26 6.00e�03 4060AN5424 Conserved protein (no known domain) – 2.0 5.7/27 5.6/31 8.00e�03 3400a
AN7590 Short-chain dehydrogenase/reductase – 2.5 6.3/28 6.4/29 1.00e�02 3601AN10260 Conserved protein (no known domain) – 1.7 5.5/26 5.5/33 1.50e�02 3213AN7999 Short-chain dehydrogenase/reductase – 1.6 7.6/29 6.2/27 3.60e�02 6987a
a Spots that retrieved more than one protein identification.b Same identification in multiple spots: [3250, 2951a, 3213a, 7305a]; [5399, 5514, 5397, 5381, 5379].
Table 4Extracellular proteins identified in the 2DE differentially accumulated protein spots in Benzoate (Bz) or Salicylate (Sc). Proteins are grouped by their family according to CAZy andMEROPS databases. Presence of a predicted signal peptide (SP) is also shown.
Protein code SP Protein (family) (EC number) Fold change pI/MW (kDa) p-Value Spot no.
Bz Sc
Glycoside Hydrolases (GHs)AN2828 Yes BglL, b-glucosidase (GH3) (3.2.1.21) 11.0 30.7 4.6/78 9.00e�03 3147AN2060 Yes AbxA, exo-arabinanase (GH93) (3.2.1.-) 2.2 3.1 5.3/42 8.19e�04 749, 3095AN6470 Yes N,O-diacetylmuramidase (GH25) (3.2.1.-) 3.6 – 4.9/23 1.00e�03 3117, 1850AN7908 Yes AxhA, a-L-arabinofuranosidase (GH62) (3.2.1.55) 1.6 – 4.3/36 6.31e�04 1256a
AN4871 Yes ChiB, class V chitinase (GH18) (3.2.1.14) – 7.8 5.8/50 1.00e�02 891AN0779 Yes Glucan exo-b-1,3-glucanase (GH55) (3.2.1.58) – 4.5 4.5/84 1.80e�02 3077a
AN6819 Yes Endo-1,3(4)-b-glucanase (GH16) (3.2.1.6) – 4.4 4.3/32 1.80e�02 380, 3077a
AN8007 Yes AbnC, endo-a-1,5-L-arabinosidase (GH43) (3.2.1.99) – 3.5 5.2/34 2.00e�02 1123, 1052, 1182a, 2961
Polysaccharide Lyases (PLs)AN7646 Yes Pectate lyase (PL1) (4.2.2.2) – 4.5 4.5/35 1.80e�02 3077a
Carbohydrate Esterases (CEs)AN1772 Yes FaeB, feruloyl esterase type B (3.1.1.73) 7.0 – 4.7/58 3.00e�03 446, 447a, 3003AN5267 Yes FaeC, feruloyl esterase type C (CE1) (3.1.1.73) – 5.9 4.6/28 3.60e�02 1318AN1433 Yes Carboxylesterase, type B (3.1.1.-) – 5.8 5.0/60 2.40e�02 441, 447a, 341a
Auxiliary activities (AAs)AN7998 Yes XptC, GMC oxidoreductase (AA3) (1.1.-.-) – 1.8 5.0/68 2.30e�02 336, 335, 341a
AN2388 No Lytic polysaccharide monooxygenase (AA9) – 1.3 4.4/46 6.31e�04 1256a
Other oxidoreductasesAN11981 Yes FAD linked oxidase/dehydrogenase, FAD-binding type 2 3.5 – 4.8/58 1.20e�02 642AN6772 Yes Quercetin 2,3-dioxygenase (1.13.11.24) – 3.9 5.6/40 2.60e�02 729, 3095
PeptidasesAN8445 Yes Leucine aminopeptidase (M28A) (3.4.11.-) 3.1 3.0 4.9/54 2.00e�03 426, 3037AN2366 Yes Trypsin-like serine peptidase (S1A) (3.4.21.-) 1.6 1.3 4.2/26 6.31e�04 1256a
AN5558 Yes PrtA, alkaline serine peptidase (S08A) (3.4.21.63) – 7.3 5.1/44 2.10e�02 1179, 1182a, 2961
a Spots that retrieved more than one protein identification.
36 T.M. Martins et al. / Fungal Genetics and Biology 74 (2015) 32–44
3. Results and discussion
3.1. Functional classification of differentially accumulated proteins inbenzoate and salicylate media
A. nidulans sub-proteomes, namely mycelial and extracellular(secretome), were analysed using comparative proteomic analyses(2DE) to identify differentially up-accumulated proteins duringgrowth in media containing either aromatic compound when com-pared to the control acetate (Supplemental Figs. S1 and S2). Inliquid cultivation, the fungus utilised benzoate (and acetate) ascarbon source much more efficiently than salicylate (SupplementalFig. S3). Four and eight days of incubation were used for benzoate
(or acetate) and salicylate media, respectively. The number of pro-tein spots differentially up-accumulated only in benzoate or salic-ylate media and in both media, were, respectively 50, 26 and 35 inthe mycelial proteome, and 8, 25 and 6 in the secretome. Amongstthese, more than 80% in the mycelial proteome (81 in total, Tables2 and 3) and 70% in the secretome (19 in total, Table 4) retrievedprotein identifications (Supplemental Table S3). Different proteinspots retrieved the same protein identity, e.g. the mycelial proteinsSodA (Cu/Zn-superoxide dismutase, AN0241), Ccp1 (cytochrome-cperoxidase, AN10220), AN4532 and AN8566 (Tables 2 and 3). Thelast two proteins, both currently assigned as putative catecholdioxygenases, were detected in a row of two to five spots of equalmolecular weight but slightly dissimilar pIs, probably due to post-

Fig. 1. Classification of differentially up-accumulated mycelial proteins in thepresence of either aromatic compound: by protein class (A) and by functionalcharacterisation (B).
T.M. Martins et al. / Fungal Genetics and Biology 74 (2015) 32–44 37
translational modifications (Supplemental Fig. S1B), and also inprotein spots with less than 10 kDa of their theoretical molecularweight, indicating that proteolysis might modulate their activity.
Amongst the identified mycelial proteins in differentially up-accumulated protein spots, the most representative classes sortedusing the gene information in AspGD (Arnaud et al., 2012) wereoxidoreductases (42%), hydrolases (13%), transferases (9%) andlyases (6%) (Fig. 1A). Function is still lacking for 44% of these pro-teins (KEGG, AspGD and Aspergillus Comparative Database), andmost of the annotated ones play roles in metabolism (29%), energymetabolism (7%) or genetic information processing (15%) (Fig. 1B).Amongst the genes presently annotated in the degradation of aro-matic compounds KEGG functional category (a total of 22 genes, 14of them associated to the conversion of salicylate to catechol), onlysalicylate 1-monooxygenase (AN2114) and catechol 1,2-dioxygen-ase (AN4532) were here found up-accumulated.
3.2. Benzoate and salicylate general impact on metabolism
Filamentous fungi are capable to utilise as carbon sources mul-tiple compounds that are directly channelled into the TCA cyclethus requiring gluconeogenesis. They are used as sources of eitheronly acetyl-CoA (acetate, ethanol and even-numbered fatty acids)or acetyl-CoA and succinate (aromatic acids and odd-numberedfatty acids), therefore depending or not on the glyoxylate cycle,respectively. Benzoate and salicylate general impact on primarymetabolism is schematically represented in Fig. 2. Up-accumula-tion of enzymes in gluconeogenesis (AcuN, PgkA and FbaA) andTCA cycle (MdhC) occurred in either aromatic media (Table 3). Util-isation of benzoate as carbon source up-accumulated enzymes of
the oxidative phase of the pentose phosphate pathway (Table 3and Fig. 2), including 6-phosphogluconate dehydrogenase(AN10783) that is believed to be the main source of NADPH regen-eration in the cell (Poulsen et al., 2005). This pathway may be usedfor anabolic purposes, consistent with the up-accumulation ofenzymes participating in the biosynthesis of nucleotides andamino acids (Table 3).
The use of succinate-producing carbon sources also affected theoxidative phosphorylation pathway, leading to up-accumulation ofproteins in complexes I, III, IV and V but not II (succinate dehydro-genase) (Table 3). This likely increased the flux of reactive oxygenspecies, hence causing up-accumulation of oxidative stressresponse associated proteins, e.g. Ccp1, SodA and glutathione per-oxidase GpxA (AN2846). Oxidative stress can in turn generatedamaged and misfolded proteins that are mainly degraded by theubiquitin proteasome system (Aiken et al., 2011), here consistentwith the up-accumulation of proteasome core subunits (AN3756,AN4869, AN5793) and ubiquitinyl hydrolase 1 (AN7491) (Table 3).
The identified mycelial proteins of unknown function (21 out of81) are mainly conserved proteins with no known domains orshort chain dehydrogenase/reductase family proteins (seven intotal). Similar up-accumulation of several dehydrogenase/reduc-tase family proteins was observed in Phanerochaete chrysosporiumin the presence of benzoate (Matsuzaki et al., 2008). Particulararylaldehyde and arylalcohol dehydrogenases are thought to medi-ate the conversion of benzoate to benzyl alcohol via intermediateformation of benzaldehyde in this fungus (Matsuzaki et al.,2008), none of which shows high homology to those found up-accumulated here. Similar reactions occur during degradation of4-methylphenol by A. fumigatus (Jones et al., 1993) but the identi-fication of the enzymes is still lacking. Their function might betherefore associated with the degradation of complex aromaticcompounds – reactions usually induced by small aromatics suchas benzoate, catechol, 4-hydroxybenzoate or protocatechuate (deVries et al., 2002). Such induction mechanism was also noticedby the up-accumulation in the secretome of feruloyl esterase typeB (FaeB, AN1772) and type C (FaeC, AN5267) in benzoate and salic-ylate media, respectively (Table 4). The encoding genes are possi-bly regulated by different aromatic compounds (de Vries et al.,2002). The remaining up-accumulated proteins in the secretome,mainly carbohydrate-active enzymes and associated proteins(Lombard et al., 2014), reveal similar responses in either aromaticcompound medium (Table 4).
3.3. The protocatechuate branch of the 3-oxoadipate pathway
Benzoate 4-monooxygenase (BzuA, AN10950), together withthe cytochrome P450 reductase partner (AN0595), mediate theconversion of benzoate to 4-hydroxybenzoate in a single reactionstep in A. nidulans (Fraser et al., 2002) (Fig. 3). BzuA orthologousprotein of Cochliobolus lunatus (CYP53A15) mediates not only thissingle step reaction but also the conversion of benzoate to protoc-atechuate in a two-step reaction depending on the cytochromeP450 reductase partner involved, respectively CPR1 and CPR2(Lah et al., 2011). Both benzoate 4-monooxygenase (PcCYP1f) andthe cytochrome P450 (CYP63A1) were induced by benzoate duringP. chrysosporium growth in a glucose medium (Matsuzaki et al.,2008). In the benzoate medium BzuA was not identified amongstthe up-accumulated proteins (neither AN0595) but bzuA was herefound exceptionally up-regulated (48-fold, Table 2). BzuA locatesin the membrane (Fraser et al., 2002) which possibly hindered itsextraction. In addition, the cytochrome b5 (AN2069) found up-accumulated in the benzoate medium (2.9-fold, Table 3) can alsoserve as electron donor to cytochrome P450 enzymes (Lah et al.,2011). In P. chrysosporium CYP63A1 was proposed as a 4-hydroxy-benzoate-3-monooxygenase (yielding protocatechuate), which are
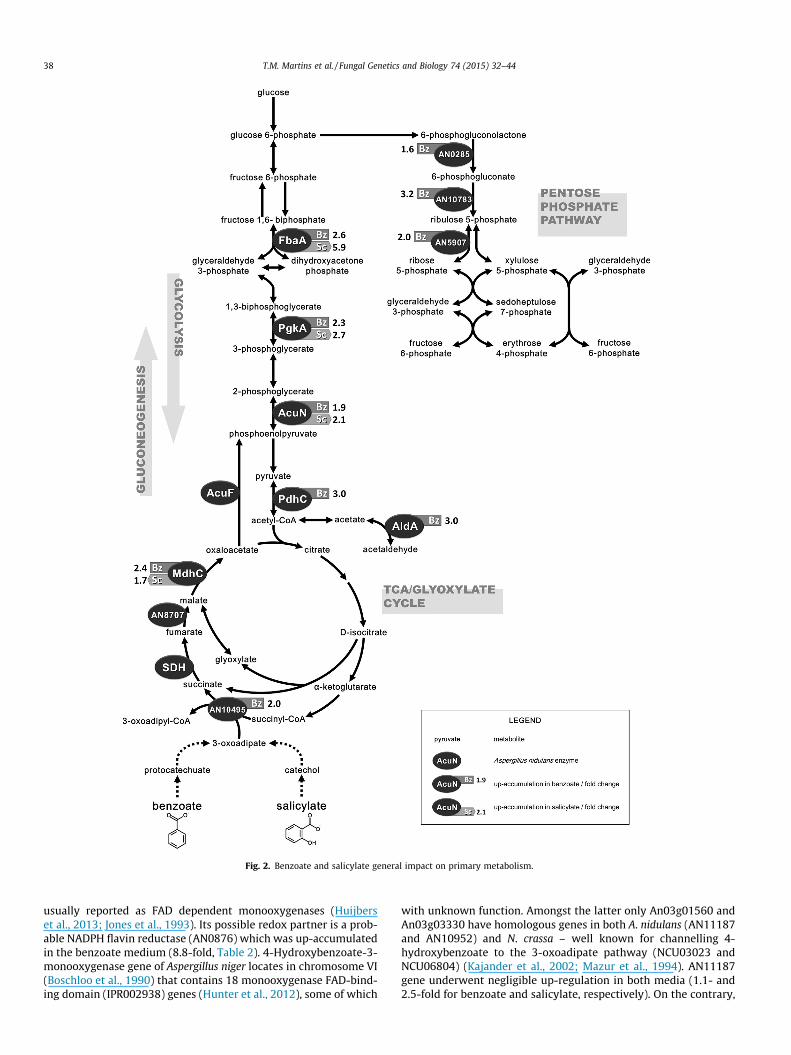
Fig. 2. Benzoate and salicylate general impact on primary metabolism.
38 T.M. Martins et al. / Fungal Genetics and Biology 74 (2015) 32–44
usually reported as FAD dependent monooxygenases (Huijberset al., 2013; Jones et al., 1993). Its possible redox partner is a prob-able NADPH flavin reductase (AN0876) which was up-accumulatedin the benzoate medium (8.8-fold, Table 2). 4-Hydroxybenzoate-3-monooxygenase gene of Aspergillus niger locates in chromosome VI(Boschloo et al., 1990) that contains 18 monooxygenase FAD-bind-ing domain (IPR002938) genes (Hunter et al., 2012), some of which
with unknown function. Amongst the latter only An03g01560 andAn03g03330 have homologous genes in both A. nidulans (AN11187and AN10952) and N. crassa – well known for channelling 4-hydroxybenzoate to the 3-oxoadipate pathway (NCU03023 andNCU06804) (Kajander et al., 2002; Mazur et al., 1994). AN11187gene underwent negligible up-regulation in both media (1.1- and2.5-fold for benzoate and salicylate, respectively). On the contrary,
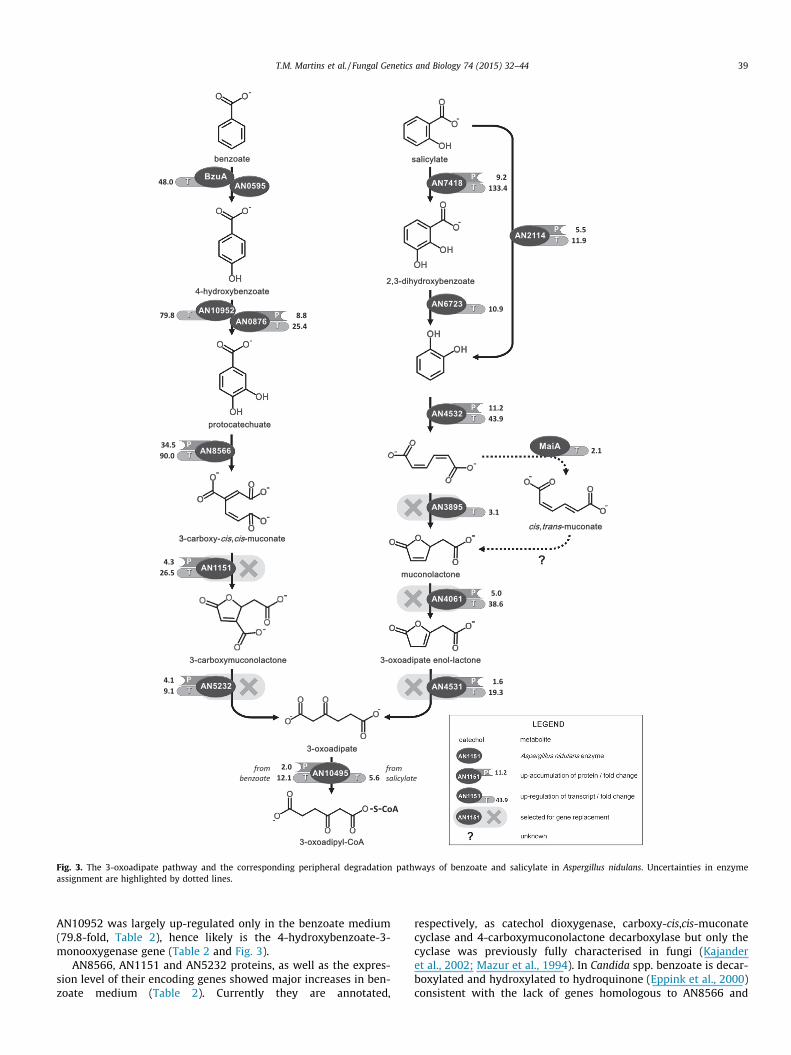
33-carboxy-cis,cis-muconate
2,3-dihydroxybenzoate
3-oxoadipate
3-oxoadipyl-CoA
-S-CoA
cis,trans-muconate
3-oxoadipate enol-lactone
salicylate
muconolactone
4-hydroxybenzoate
benzoate
protocatechuate
AN0876P T
8.8 25.4 T
BzuAT 48.0 T AN0595
AN10952T 79.8 T
T 5.6 T
AN10495
P T
2.0 12.1 T
from benzoate
from salicylate
MaiA T 2.1 T
?
AN5232
P T
4.1 9.1 T
AN4061P T
5.0 38.6 T
AN4531 P T
1.6 19.3 T
3-carboxymuconolactone
AN1151
P T
4.3 26.5 T
AN8566
P T
34.5 90.0 T
AN3895 T 3.1 T
AN4532P T
11.2 43.9 T
AN6723 T 10.9 T
AN2114P T
5.5 11.9 T
AN7418P T
9.2 133.4 T
Fig. 3. The 3-oxoadipate pathway and the corresponding peripheral degradation pathways of benzoate and salicylate in Aspergillus nidulans. Uncertainties in enzymeassignment are highlighted by dotted lines.
T.M. Martins et al. / Fungal Genetics and Biology 74 (2015) 32–44 39
AN10952 was largely up-regulated only in the benzoate medium(79.8-fold, Table 2), hence likely is the 4-hydroxybenzoate-3-monooxygenase gene (Table 2 and Fig. 3).
AN8566, AN1151 and AN5232 proteins, as well as the expres-sion level of their encoding genes showed major increases in ben-zoate medium (Table 2). Currently they are annotated,
respectively, as catechol dioxygenase, carboxy-cis,cis-muconatecyclase and 4-carboxymuconolactone decarboxylase but only thecyclase was previously fully characterised in fungi (Kajanderet al., 2002; Mazur et al., 1994). In Candida spp. benzoate is decar-boxylated and hydroxylated to hydroquinone (Eppink et al., 2000)consistent with the lack of genes homologous to AN8566 and

Fig. 4. Aspergillus nidulans gene-replacement mutants grown in acetate liquid medium (control) or in benzoate or salicylate liquid medium with or without acetate co-supplementation (8 days). Genes of the protocatechuate branch were essential for benzoate utilisation as carbon source (AN1151 and AN5232). Deletion of AN3895, AN4061and AN4531 genes in the catechol branch showed phenotypes with increasing levels of growth impairment.
40 T.M. Martins et al. / Fungal Genetics and Biology 74 (2015) 32–44
AN5232. In the present study, DAN1151 (Table 1) was unable togrow in the presence of benzoate, regardless of acetate co-supple-mentation (Figs. 4 and 5).
In addition to AN8566 there are another four intradiol ring-cleavage catechol dioxygenase genes in A. nidulans (N-terminal:IPR007535 and C-terminal: IPR000627 domains (Hunter et al.,2012)). These genes, namely AN4532, AN8998, AN0764 andAN9363, may be classed in three sub-groups of a single evolution-ary lineage family, namely catechol 1,2-dioxygenase, hydroxyqui-nol 1,2-dioxygenase and protocatechuate 3,4-dioxygenase. Amember of the first sub-group, previously characterised and iden-tified in Candida albicans (Tsai and Li, 2007), is orthologous toAN4532 and AN8998 (paralogous genes). Candida parapsilosis uti-lises benzoate (and derivatives) through a hydroxyquinol variantof the 3-oxoadipate pathway, hence its single catechol dioxygen-ase-like gene belongs to the second sub-group (Holesova et al.,2011), which is orthologous to AN0764 and AN9363. These find-ings support AN8566 as the protocatechuate 3,4-dioxygenase gene(Table 2 and Fig. 3).
Fungi are devoid of 4-carboxymuconolactone decarboxylasegenes because they form 3-carboxymucolactone and not 4-carbo-xymuconolactone as reported for bacteria (Harwood and Parales,1996); regardless AN1252, AN4023 and AN5232 have been anno-tated as such. In fungi, conversion of 3-carboxymuconolactone to3-oxoadipate is mediated by a single enzyme with both hydrolase
and decarboxylase activities (Thatcher and Cain, 1970). AN5232gene was induced in benzoate medium (noticed at both proteinand gene level) but not in the salicylate medium (Table 2). Thisgene is highly conserved among aspergilli, on opposition to theother two genes sharing the same domain (IPR003779) (Arnaudet al., 2012), and shows the highest sequence identity to the onlyorthologous gene of N. crassa (NCU04594). The corresponding dele-tion mutant showed similar phenotype to DAN1151 (Figs. 4 and 5).When exposed to benzoate (cultures pre-grown in acetate med-ium) both mutants accumulated protocatechuate and 4-hydroxy-benzoate, similar to that observed in the wild type. In addition,in DAN1151 and DAN5232 cultures, respectively 3-carboxy-cis-cis-muconate and 3-carboxymuconalactone were putatively iden-tified as well (Fig. 6A). Based on these findings, AN5232 shouldbe functionally assigned as 3-carboxymuconolactone hydrolase(decarboxylating) (Fig. 3).
Metabolic convergence of the two branches of the 3-oxoadipatepathway occurs with formation of 3-oxoadipate (Fig. 3). The down-stream reaction is mediated by a 3-oxoadipate-CoA transferase.Aspergillus nidulans contains two putative succinyl-CoA:3-oxo-acid-CoA transferase family genes, namely AN5669 and AN10495.The former has been putatively identified as succinyl-CoA:2-oxo-acid-CoA transferase gene because of its differentially expressionin glycerol medium in three aspergilli (Salazar et al., 2009).AN10495 protein was up-accumulated only in the benzoate med-

Fig. 5. Aspergillus nidulans gene-replacement mutants grown onto solid mediacontaining acetate, benzoate or salicylate (6 days). Genes of the protocatechuatebranch were essential for benzoate utilisation as carbon source (AN1151 andAN5232). Genes in the catechol branch were not essential for salicylate utilisationas carbon source (AN3895, AN4061 and AN4531), probably because the fungusutilised the hydroxyquinol variant of the 3-oxoadipate pathway during growth onsolid media.
T.M. Martins et al. / Fungal Genetics and Biology 74 (2015) 32–44 41
ium but the encoding gene underwent up-regulation in both media(Table 2), being presumably a succinyl-CoA:3-oxoadipate-CoAtransferase (Fig. 3).
3.4. The catechol branch of the 3-oxoadipate pathway
Salicylate has been shown to be transformed to catechol eitherdirectly or with phenol or 2,3-dihydroxybenzoate as intermediates(Kirimura et al., 2010; Wright, 1993). Fungi in general, and A. nidu-lans in particular, are capable of hydroxylating salicylate to yield2,3-dihydroxybenzoate (Wright, 1993). The latter undergoes anon-oxidative decarboxylation by 2,3-dihydroxybenzoate car-boxy-lyase (AN6723, EC 4.1.1.46) which has been extensively stud-ied in aspergilli. The conversion through phenol, which wouldrequire a reversible salicylate decarboxylase, is unlikely to occurin aspergilli. This reaction, previously characterised in Trichosporonmoniliiforme (Kirimura et al., 2010), involves an orthologue ofAN6723 but the gene products showed different substrate specific-ity (Kamath and Vaidyanathan, 1990). Two hydroxylating enzymes
were found to up-accumulate only in salicylate medium, namelyAN2114 and AN7418, currently annotated as salicylate 1-monoox-ygenase (decarboxylating) and phenol 2-monooxygenase, respec-tively. Both the corresponding genes, as well as AN6723,underwent major up-regulation only in the salicylate medium(Table 2). These observations are consistent with catechol forma-tion from salicylate either directly or with 2,3-dihydroxybenzoateas intermediate (Fig. 3).
Catechol conversion to cis,cis-muconate in C. albicans involves acatechol 1,2-dioxygenase (Tsai and Li, 2007), of which the encodinggene shows the highest sequence identity with AN4532. Asexpected, the last underwent major induction in the salicylatemedium, at both protein (11 spots, see Supplemental Fig. S1) andgene levels (Table 2).
AN4531, presently recognised as either uncharacterised oralpha/beta-hydrolase, underwent major up-regulation only in thesalicylate medium and the encoded protein was also up-accumu-lated (Table 2). The bacterial 3-oxoadipic enol-lactone hydrolasecontains ca. 260 amino acids and proteins of similar length areencoded by several fungal homologous genes (two exons)(Arnaud et al., 2012). AN4531 is currently annotated as codifyinga protein with 510 amino acid residues (three exons) but therelated protein spot retrieved an experimental molecular weightmuch lower than the theoretical one (27 and 56 kDa, respectively,Table 2). The protein codifying sequence of AN4531 is probablyshorter (likely containing only two exons), consequently consistentwith a 3-oxoadipic enol-lactone hydrolase gene (Fig. 3). In linewith this hypothesis, growth of the corresponding gene-replace-ment mutant in liquid media in the presence of salicylate wasextremely impaired, regardless of acetate co-supplementation(Fig. 4). The very scarce mycelia produced a dark brown metabo-lite. The corresponding chromatogram shows accumulation of apeak with retention time similar to muconolactone (Fig. 6B). Its fullspectrum shows overlapping of muconolactone and an uncharac-terised compound (kmax at 300 nm), likely 3-oxoadipate enol-lactone.
Catechol 1,2-dioxygenase (AN4532) and AN4531 genes areclustered in the genomes of A. nidulans and other aspergilli(Arnaud et al., 2012). Homologous genes are enclosed in a genomicinsert of five highly conserved genes (Maguire et al., 2013) (Supple-mental Table S4 and Fig. S5) that is highly correlated to phenol andcatechol degradation capacity in Ascomycota yeasts (Holesovaet al., 2011; Middelhoven, 1993; Tsai and Li, 2007). One of theinsert genes is homologous to AN4061 that in salicylate mediumwas up-regulated and its encoded protein also increased (Table 2).Further BLAST gene analyses showed that the same insert is mostlyconserved in the Basidomycota yeast Trichosporon asahii, as well asin some Ascomycota fungi including Fusarium spp. (four of the fivegenes rearranged in order) and N. crassa (Supplemental Table S4).In addition, the uncharacterised genes AN4061 and AN3895 areclustered in most aspergilli even if not in A. nidulans. AN3895showed similar behaviour to AN4061, notwithstanding at a muchlower expression level (Table 2), suggesting that they mightencode the enzymes required for this branch of the 3-oxoadipatepathway. In particular, they should be involved either in the muc-onates isomerisation or lactonisation or in the subsequent isomeri-sation of muconolactone. In a previous study we demonstratedthat 2-chloro-cis,cis-muconate was converted to the cis,trans iso-mer (Martins et al., 2014b). This reaction is likely mediated bymaleylacetoacetate isomerase (MaiA, AN1895) (Seltzer and Hane,1988), of which the encoding gene underwent minor up-regulationin the salicylate medium (Table 2). maiA low expression level in thesalicylate media is consistent with the accumulation of both iso-mers in the culture media (Supplemental Fig. S4). Conversion ofeither muconate isomer to muconolactone must be mediated bya muconate cycloisomerase since at the neutral pH used (not
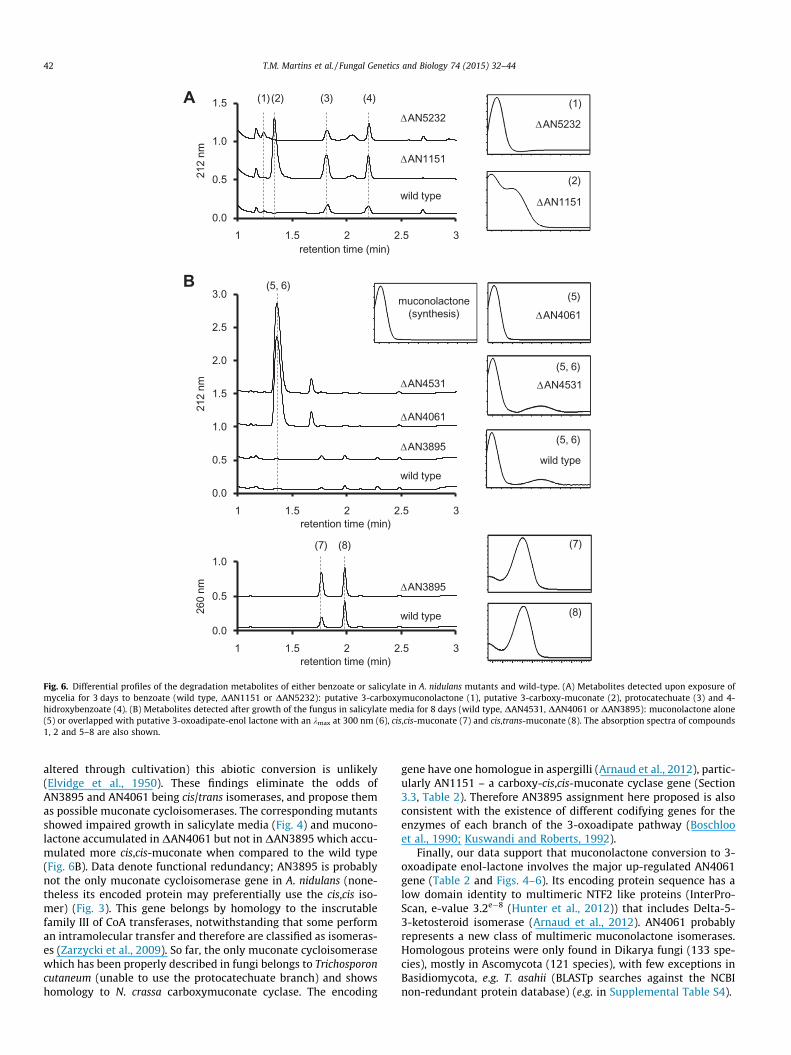
0.0
0.5
1.0
1.5
2.0
2.5
3.0
1 1.5 2 2.5 3
212
nm
retention time (min)
0.0
0.5
1.0
1 1.5 2 2.5 3
260
nm
retention time (min)
0.0
0.5
1.0
1.5
1 1.5 2 2.5 3
212
nm
retention time (min)
(1)(2) (3) (4)A
wild type
ΔAN5232
(5, 6)B
wild type
ΔAN3895
ΔAN1151
ΔAN4061
ΔAN4531
wild type
ΔAN3895
ΔAN1151
ΔAN5232
(1)
(2)
muconolactone(synthesis) ΔAN4061
(5)
(5, 6)
(5, 6)
wild type
ΔAN4531
(7)
(8)
(7) (8)
Fig. 6. Differential profiles of the degradation metabolites of either benzoate or salicylate in A. nidulans mutants and wild-type. (A) Metabolites detected upon exposure ofmycelia for 3 days to benzoate (wild type, DAN1151 or DAN5232): putative 3-carboxymuconolactone (1), putative 3-carboxy-muconate (2), protocatechuate (3) and 4-hidroxybenzoate (4). (B) Metabolites detected after growth of the fungus in salicylate media for 8 days (wild type, DAN4531, DAN4061 or DAN3895): muconolactone alone(5) or overlapped with putative 3-oxoadipate-enol lactone with an kmax at 300 nm (6), cis,cis-muconate (7) and cis,trans-muconate (8). The absorption spectra of compounds1, 2 and 5–8 are also shown.
42 T.M. Martins et al. / Fungal Genetics and Biology 74 (2015) 32–44
altered through cultivation) this abiotic conversion is unlikely(Elvidge et al., 1950). These findings eliminate the odds ofAN3895 and AN4061 being cis/trans isomerases, and propose themas possible muconate cycloisomerases. The corresponding mutantsshowed impaired growth in salicylate media (Fig. 4) and mucono-lactone accumulated in DAN4061 but not in DAN3895 which accu-mulated more cis,cis-muconate when compared to the wild type(Fig. 6B). Data denote functional redundancy; AN3895 is probablynot the only muconate cycloisomerase gene in A. nidulans (none-theless its encoded protein may preferentially use the cis,cis iso-mer) (Fig. 3). This gene belongs by homology to the inscrutablefamily III of CoA transferases, notwithstanding that some performan intramolecular transfer and therefore are classified as isomeras-es (Zarzycki et al., 2009). So far, the only muconate cycloisomerasewhich has been properly described in fungi belongs to Trichosporoncutaneum (unable to use the protocatechuate branch) and showshomology to N. crassa carboxymuconate cyclase. The encoding
gene have one homologue in aspergilli (Arnaud et al., 2012), partic-ularly AN1151 – a carboxy-cis,cis-muconate cyclase gene (Section3.3, Table 2). Therefore AN3895 assignment here proposed is alsoconsistent with the existence of different codifying genes for theenzymes of each branch of the 3-oxoadipate pathway (Boschlooet al., 1990; Kuswandi and Roberts, 1992).
Finally, our data support that muconolactone conversion to 3-oxoadipate enol-lactone involves the major up-regulated AN4061gene (Table 2 and Figs. 4–6). Its encoding protein sequence has alow domain identity to multimeric NTF2 like proteins (InterPro-Scan, e-value 3.2e�8 (Hunter et al., 2012)) that includes Delta-5-3-ketosteroid isomerase (Arnaud et al., 2012). AN4061 probablyrepresents a new class of multimeric muconolactone isomerases.Homologous proteins were only found in Dikarya fungi (133 spe-cies), mostly in Ascomycota (121 species), with few exceptions inBasidiomycota, e.g. T. asahii (BLASTp searches against the NCBInon-redundant protein database) (e.g. in Supplemental Table S4).

T.M. Martins et al. / Fungal Genetics and Biology 74 (2015) 32–44 43
A. nidulans capacity to utilise very efficiently salicylate as car-bon source in solid media (Fig. 5) is dramatically hindered duringliquid cultivation (Supplemental Fig. S3). Deletion of genes in thecatechol branch (AN3895, AN4061 and AN4531) showed pheno-types able to utilise rather efficiently salicylate as carbon sourceduring growth in solid media (Fig. 5). However, both DAN4061and DAN4531 barely grow in the salicylate liquid medium.DAN3895 divergent phenotype emphasises that this is not the onlylikely muconate cycloisomerase gene. Likely the activation of thehydroxyquinol variant of the 3-oxoadipate pathway was activatedonly during solid cultivation, i.e. under stringent regulation by theenvironmental conditions.
Finally, the growth impairment of all these mutant phenotypesin the salicylate liquid media was partially resolved in media co-supplemented with acetate, except for DAN4531 (Fig. 4). Possibly,growth inhibitory metabolites accumulating in media peakedwhen the last step of the catechol branch was interrupted. The cor-responding mutant could not bypass the media toxicity even whenco-supplemented with acetate.
4. Conclusions
This contribution resolved fundamentals of the degradation ofaromatic compounds in A. nidulans. In particular, it assigns previ-ously uncharacterised genes to specific reaction steps of the 3-oxoadipate pathway and also corrected some misconceptions pre-dicted from bacterial studies. The gene families here identified dis-close the genetically encoded potential to undertake just one orboth branches of the pathway in fungi. Deletion of essential genesin the catechol branch showed phenotypes able to utilise salicylateas carbon source during growth in solid media. This was likelyensured through the activation of the hydroxyquinol variant ofthe 3-oxoadipate pathway that was, however, largely hinderedduring growth in liquid media. On the contrary, genes of the prot-ocatechuate branch were essential for benzoate utilisation as car-bon source in either solid or liquid media. The last was true evenin media co-supplemented with acetate, in line with the recogni-sed toxicity of benzoate and/or its degradation intermediates.Novel studies focussing the regulatory networks of this pathwayshould be encouraged. Fungi catabolic potential remains still lar-gely untapped but our data inspire the development of engineeredstrains for efficient degradation of disparate aromatic compoundsin biorefinery and/or bioremediation applications.
Acknowledgments
T.M. and D.O.H. are grateful to Fundação para a Ciência e a Tecn-ologia (FCT) – Portugal for the fellowships SFRH/BPD/70064/2010and SFRH/BD/66396/2009. The work was partially supported byFCT Grants: PEst-OE/EQB/LA0004/2013 and PTDC/AAC-CLI/119100/2010.
Appendix A. Supplementary material
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.fgb.2014.11.002.
References
Aiken, C.T., Kaake, R.M., Wang, X., Huang, L., 2011. Oxidative stress-mediatedregulation of proteasome complexes. Mol. Cell. Proteomics 10 (5), R110.006924.
Arnaud, M.B., Cerqueira, G.C., Inglis, D.O., Skrzypek, M.S., Binkley, J., Chibucos, M.C.,Crabtree, J., Howarth, C., Orvis, J., Shah, P., 2012. The Aspergillus GenomeDatabase (AspGD): recent developments in comprehensive multispeciescuration, comparative genomics and community resources. Nucleic Acids Res.40 (D1), D653–D659.
Boschloo, J., Paffen, A., Koot, T., Tweel, W., Gorcom, R.v., Cordewener, J., Bos, C., 1990.Genetic analysis of benzoate metabolism in Aspergillus niger. Appl. Microbiol.Biotechnol. 34 (2), 225–228.
Carvalho, M.B., Martins, I., Medeiros, J., Tavares, S., Planchon, S., Renaut, J., Núñez, O.,Gallart-Ayala, H., Galceran, M.T., Hursthouse, A., Silva Pereira, C., 2013. Theresponse of Mucor plumbeus to pentachlorophenol: a toxicoproteomics study. J.Proteomics 78, 159–171.
de Vries, R., van Kuyk, P., Kester, H., Visser, J., 2002. The Aspergillus niger faeB geneencodes a second feruloyl esterase involved in pectin and xylan degradationand is specifically induced in the presence of aromatic compounds. Biochem. J.363, 377–386.
Elvidge, J., Linstead, R., Sims, P., Orkin, B., 1950. The third isomeric (cis-trans-)muconic acid. J. Chem. Soc., 2235–2241.
Eppink, M., Cammaart, E., Van Wassenaar, D., Middelhoven, W., van Berkel, W.,2000. Purification and properties of hydroquinone hydroxylase, a FAD-dependent monooxygenase involved in the catabolism of 4-hydroxybenzoatein Candida parapsilosis CBS604. Eur. J. Biochem. 267 (23), 6832.
Fraser, J.A., Davis, M.A., Hynes, M.J., 2002. The genes gmdA, encoding an amidase,and bzuA, encoding a cytochrome P450, are required for benzamide utilizationin Aspergillus nidulans. Fungal Genet. Biol. 35 (2), 135–146.
Harwood, C.S., Parales, R.E., 1996. The b-ketoadipate pathway and the biology ofself-identity. Annu. Rev. Microbiol. 50 (1), 553–590.
Hawkins, A.R., Lamb, H.K., Moore, J.D., Charles, I.G., Roberts, C.F., 1993. ReviewArticle: The pre-chorismate (shikimate) and quinate pathways in filamentousfungi: theoretical and practical aspects. J. Gen. Microbiol. 139 (12), 2891–2899.
Holesova, Z., Jakubkova, M., Zavadiakova, I., Zeman, I., Tomaska, L., Nosek, J., 2011.Gentisate and 3-oxoadipate pathways in the yeast Candida parapsilosis:identification and functional analysis of the genes coding for 3-hydroxybenzoate 6-hydroxylase and 4-hydroxybenzoate 1-hydroxylase.Microbiology 157 (7), 2152–2163.
Huijbers, M.M., Montersino, S., Westphal, A.H., Tischler, D., van Berkel, W.J., 2013.Flavin dependent monooxygenases. Arch. Biochem. Biophys. 544, 2–17.
Hunter, S., Jones, P., Mitchell, A., Apweiler, R., Attwood, T.K., Bateman, A., Bernard, T.,Binns, D., Bork, P., Burge, S., 2012. InterPro in 2011: new developments in thefamily and domain prediction database. Nucleic Acids Res. 40 (D1), D306–D312.
Jones, K.H., Trudgill, P.W., Hopper, D.J., 1993. Metabolism of p-cresol by the fungusAspergillus fumigatus. Appl. Environ. Microbiol. 59 (4), 1125–1130.
Kajander, T., Merckel, M.C., Thompson, A., Deacon, A.M., Mazur, P., Kozarich, J.W.,Goldman, A., 2002. The structure of Neurospora crassa 3-carboxy-cis, cis-muconate lactonizing enzyme, a b propeller cycloisomerase. Structure 10 (4),483–492.
Kamath, A.V., Vaidyanathan, C., 1990. Structural requirements for the induction andinhibition of 2,3-dihydroxybenzoic acid decarboxylase of Aspergillus niger. Curr.Microbiol. 20 (2), 111–114.
Kirimura, K., Gunji, H., Wakayama, R., Hattori, T., Ishii, Y., 2010. Enzymatic Kolbe–Schmitt reaction to form salicylic acid from phenol: enzymatic characterizationand gene identification of a novel enzyme, Trichosporon moniliiforme salicylicacid decarboxylase. Biochem. Biophys. Res. Commun. 394 (2), 279–284.
Kuswandi, K., Roberts, C., 1992. Genetic control of the protocatechuic acid pathwayin Aspergillus nidulans. J. Gen. Microbiol. 138 (4), 817–823.
Lah, L., Podobnik, B., Novak, M., Korošec, B., Berne, S., Vogelsang, M., Kraševec, N.,Zupanec, N., Stojan, J., Bohlmann, J., 2011. The versatility of the fungalcytochrome P450 monooxygenase system is instrumental in xenobioticdetoxification. Mol. Microbiol. 81 (5), 1374–1389.
Lombard, V., Ramulu, H.G., Drula, E., Coutinho, P.M., Henrissat, B., 2014. Thecarbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 42(D1), D490–D495.
Maguire, S.L., ÓhÉigeartaigh, S.S., Byrne, K.P., Schröder, M.S., O’Gaora, P., Wolfe, K.H.,Butler, G., 2013. Comparative genome analysis and gene finding in Candidaspecies using CGOB. Mol. Biol. Evol. 30 (6), 1281–1291.
Martins, I., Garcia, H., Varela, A., Núñez, O., Planchon, S., Galceran, M.T., Renaut, J.,Rebelo, L.P., Silva Pereira, C., 2014a. Investigating Aspergillus nidulans secretomeduring colonisation of cork cell walls. J. Proteomics 98, 175–188.
Martins, T.M., Núñez, O., Gallart-Ayala, H., Leitão, M.C., Galceran, M.T., Silva Pereira,C., 2014b. New branches in the degradation pathway of monochlorocatecholsby Aspergillus nidulans: a metabolomics analysis. J. Hazard. Mater. 268, 264–272.
Matsuzaki, F., Shimizu, M., Wariishi, H., 2008. Proteomic and metabolomic analysesof the white-rot fungus Phanerochaete chrysosporium exposed to exogenousbenzoic acid. J. Proteome Res. 7 (6), 2342–2350.
Mazur, P., Henzel, W.J., Mattoo, S., Kozarich, J.W., 1994. 3-Carboxy-cis, cis-muconatelactonizing enzyme from Neurospora crassa: an alternate cycloisomerase motif.J. Bacteriol. 176 (6), 1718–1728.
Middelhoven, W.J., 1993. Catabolism of benzene compounds by ascomycetous andbasidiomycetous yeasts and yeast-like fungi – a literature-review and anexperimental approach. Anton. Leeuw. Int. J. G. 63 (2), 125–144.
Palmqvist, E., Hahn-Hägerdal, B., 2000. Fermentation of lignocellulosichydrolysates. II: inhibitors and mechanisms of inhibition. Bioresour. Technol.74 (1), 25–33.
Poulsen, B.R., Nøhr, J., Douthwaite, S., Hansen, L.V., Iversen, J.J., Visser, J., Ruijter, G.J.,2005. Increased NADPH concentration obtained by metabolic engineering of thepentose phosphate pathway in Aspergillus niger. FEBS J. 272 (6), 1313–1325.
Salazar, M., Vongsangnak, W., Panagiotou, G., Andersen, M.R., Nielsen, J., 2009.Uncovering transcriptional regulation of glycerol metabolism in Aspergillithrough genome-wide gene expression data analysis. Mol. Genet. Genomics 282(6), 571–586.

44 T.M. Martins et al. / Fungal Genetics and Biology 74 (2015) 32–44
Seltzer, S., Hane, J., 1988. Maleylacetoacetate cis–trans isomerase: one-step doublecis–trans isomerization of monomethyl muconate and the enzyme’s probablerole in benzene metabolism. Bioorg. Chem. 16 (4), 394–407.
Szewczyk, E., Nayak, T., Oakley, C.E., Edgerton, H., Xiong, Y., Taheri-Talesh, N.,Osmani, S.A., Oakley, B.R., 2006. Fusion PCR and gene targeting in Aspergillusnidulans. Nat. Protoc. 1 (6), 3111–3120.
Tang, X., Dong, W., Griffith, J., Nilsen, R., Matthes, A., Cheng, K.B., Reeves, J., Schuttler,H.B., Case, M.E., Arnold, J., Logan, D.A., 2011. Systems biology of the qa genecluster in Neurospora crassa. PLoS ONE 6 (6), e20671.
Thatcher, D., Cain, R., 1970. Metabolism of aromatic compounds by fungi:conversion of b-carboxymuconolactone into 3-oxoadipate in Aspergillus niger.Biochem. J. 120 (4), 28–29.
Tsai, S.-C., Li, Y.-K., 2007. Purification and characterization of a catechol 1, 2-dioxygenase from a phenol degrading Candida albicans TL3. Arch. Microbiol. 187(3), 199–206.
Veiga, T., Solis-Escalante, D., Romagnoli, G., ten Pierick, A., Hanemaaijer, M.,Deshmuhk, A., Wahl, A., Pronk, J.T., Daran, J.M., 2012. Resolving phenylalaninemetabolism sheds light on natural synthesis of Penicillin G in Penicilliumchrysogenum. Eukaryot. Cell 11 (2), 238–249.
Wright, J., 1993. Fungal degradation of benzoic acid and related compounds. WorldJ. Microbiol. Biotechnol. 9 (1), 9–16.
Zarzycki, J., Brecht, V., Müller, M., Fuchs, G., 2009. Identifying the missing steps ofthe autotrophic 3-hydroxypropionate CO2 fixation cycle in Chloroflexusaurantiacus. Proc. Natl. Acad. Sci. U.S.A. 106 (50), 21317–21322.
Copyright © 2022 FDOKUMEN






![Regulation of Lysine Catabolism through Lysine[mdash]Ketoglutarate Reductase and Saccharopine Dehydrogenase in Arabidopsis](https://static.fdokumen.com/doc/165x107/631cc83693f371de19019c93/regulation-of-lysine-catabolism-through-lysinemdashketoglutarate-reductase-and.jpg)













