
Somatic hypermutation of human mitochondrial and nuclear DNA by APOBEC3 cytidine deaminases, a...
6
Somatic hypermutation of human mitochondrial and nuclear DNA by APOBEC3 cytidine deaminases, a pathway for DNA catabolism Rodolphe Suspène a,1 , Marie-Ming Aynaud a,1 , Denise Guétard a , Michel Henry a , Grace Eckhoff a , Agnès Marchio b , Pascal Pineau b , Anne Dejean b,c , Jean-Pierre Vartanian a , and Simon Wain-Hobson a,2 a Institut Pasteur, Molecular Retrovirology Unit, Centre National de la Recherche Scientifique URA3015 and b Nuclear Organization and Oncogenesis Unit, 75724 Paris cedex 15, France; and c Institut National de la Santé et de la Recherche Médicale, U579, 75724 Paris cedex 15, France Edited by Reuben S. Harris, University of Minnesota, Minneapolis, MN, and accepted by the Editorial Board February 3, 2011 (received for review July 7, 2010) The human APOBEC3 (A3A–A3H) locus encodes six cytidine deam- inases that edit single-stranded DNA, the result being DNA pep- pered with uridine. Although several cytidine deaminases are clearly restriction factors for retroviruses and hepadnaviruses, it is not known if APOBEC3 enzymes have roles outside of these set- tings. It is shown here that both human mitochondrial and nuclear DNA are vulnerable to somatic hypermutation by A3 deaminases, with APOBEC3A standing out among them. The degree of editing is much greater in patients lacking the uracil DNA-glycolyase gene, indicating that the observed levels of editing reflect a dynamic composed of A3 editing and DNA catabolism involving uracil DNA-glycolyase. Nonetheless, hyper- and lightly mutated sequen- ces went hand in hand, raising the hypothesis that recurrent low- level mutation by APOBEC3A could catalyze the transition from a healthy to a cancer genome. T he human APOBEC3 (A3) locus encodes seven expressed genes (A3A–A3H), of which six are demonstrably cytidine deaminases with specificity for single-stranded DNA (ssDNA) (1). The A3 locus arose in placental mammals by duplication of AICDA and subsequent expansion (2). Activation-induced cyti- dine deaminase (AICDA) is presently the only enzyme known to induce mutations in the human genome and initiates class-switch recombination and somatic hypermutation of rearranged Ig genes (3). Several human A3 genes are up-regulated in inflammatory settings by type I and II interferons, although activation depends to some degree on the cell type (4–8). Given this finding, it is not surprising that A3F and A3G are involved in the restriction of retroviruses and hepadnaviruses where nascent single-stranded cDNA remains vulnerable until double-strand synthesis is com- pleted (9–16). In late stages of hepatitis B virus- (HBV) associ- ated cirrhosis there was significant up-regulation of four to five A3 genes, with A3G being the major restriction factor able to edit up to 35% of HBV genomes (17). DNA viral genomes, such as human papillomavirus (HPV), can also undergo editing both in vivo and in tissue culture, presumably during transcription or replication (18). Three A3 deaminases, A3A, A3C, and A3H could hyperedit HPV genomes ex vivo, suggesting that the phenomenon was occurring in the nucleus because the strictly cytoplasmic deaminases, A3F and A3G, did not edit transfected HPV DNA (18). Along with a re- cent article showing that A3A could edit transfected EGFP plasmid DNA (7), they highlight the potential of A3 enzymes to edit foreign DNA. The presence of uridine in cellular DNA is read as a danger signal, the uracil base resulting from cytidine deamination being subject to excision, particularly by uracil DNA-glycosylase (UNG), the nuclear form (UNG2) being far more active than its mitochondrial counterpart (UNG1). Uracil excision followed by abasic endonuclease cleavage of the DNA strand can lead to repair or degradation. Small fragments of nu- clear and mitochondrial DNA (mtDNA) can be found in the cytoplasm, which trigger a number of cytoplasmic DNA sensor molecules and are associated with DNA degradation (19, 20). As there are six human A3 enzymes, all with ssDNA specificity, the question arises as to whether they can edit host-cell DNA. Given the high mtDNA copy number per cell, we explored the possibility that a fraction of mtDNA might be vulnerable to A3 editing, especially in inflamed tissues where there is demonstrable up-regulation of A3 genes (17). We show here that a small frac- tion of human mitochondrial genomes are phenomenally edited by APOBEC3 deaminases in the cytoplasm. It transpires that nuclear DNA (nuDNA), notably segments of MYC and TP53, can be hyperedited by APOBEC3A. For both mitochondrial and nuDNA, editing reflects a dynamic between A3 activity and DNA degradation involving UNG. Results APOBEC3 Hypermutation of mtDNA. A nested PCR/3DPCR ap- proach was taken to the amplification of mtDNA from 36 DNA samples spanning alcoholic, HBV + , HBV + HCV + , and HCV + cirrhosis (17, 21). A region corresponding to part of MT-COI at ∼6,200 bp was chosen, as it maps within the large single- stranded replication intermediate. Mitochondrial DNA could be recovered at temperatures as low as 82.2 °C from 17 of 36 (∼47%) samples tested, much lower than for a control liver or cloned MT- COI DNA (Td∼86 °C) (Fig. 1 A and B). Molecular cloning and sequencing of the PCR products proved that they represented bona fide cytidine-edited mtDNA, with both strands being vulnerable (Fig. 1C and Fig. S1A), although minus-strand hypermutants were generally more abundant (shown as G->A hypermutants with respect to the plus strand). Although minus-strand editing was generally in the 30 to 80% range, occasionally a sequence with up to 98% of targets could be edited (Fig. 1 C and D). As A3 and AICDA catalyzed cytidine deamination is partic- ularly sensitive to the 5′ base, a bulk dinucleotide analysis was performed for minus- and plus-strand hypermutants (Fig. 1 E and F). The strong biases for TpC and CpC among minus-strand hypermutants are hallmarks of APOBEC3 editing (11, 15, 22). For plus-strand editing, the clear preference for CpC is a hall- mark of A3G (Fig. 1F), no matter the template (11). These slightly different patterns probably reflect the greater C content of the plus strand generating more oligoC tracts, which are known hotspots for A3G. A clonal analysis of the dinucleotide Author contributions: R.S., J.-P.V., and S.W.-H. designed research; R.S., M.-M.A., D.G., M.H., G.E., and A.M. performed research; P.P. and A.D. contributed new reagents/analytic tools; R.S., M.-M.A., J.-P.V., and S.W.-H. analyzed data; and J.-P.V. and S.W.-H. wrote the paper. The authors declare no conflict of interest. This article is a PNAS Direct Submission. R.S.H. is a guest editor invited by the Editorial Board. 1 R.S. and M.-M.A. contributed equally to this work. 2 To whom correspondence should be addressed. E-mail: [email protected]. This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10. 1073/pnas.1009687108/-/DCSupplemental. 4858–4863 | PNAS | March 22, 2011 | vol. 108 | no. 12 www.pnas.org/cgi/doi/10.1073/pnas.1009687108
Transcript of Somatic hypermutation of human mitochondrial and nuclear DNA by APOBEC3 cytidine deaminases, a...

Somatic hypermutation of human mitochondrial andnuclear DNA by APOBEC3 cytidine deaminases,a pathway for DNA catabolismRodolphe Suspènea,1, Marie-Ming Aynauda,1, Denise Guétarda, Michel Henrya, Grace Eckhoffa, Agnès Marchiob,Pascal Pineaub, Anne Dejeanb,c, Jean-Pierre Vartaniana, and Simon Wain-Hobsona,2
aInstitut Pasteur, Molecular Retrovirology Unit, Centre National de la Recherche Scientifique URA3015 and bNuclear Organization and Oncogenesis Unit,75724 Paris cedex 15, France; and cInstitut National de la Santé et de la Recherche Médicale, U579, 75724 Paris cedex 15, France
Edited by Reuben S. Harris, University of Minnesota, Minneapolis, MN, and accepted by the Editorial Board February 3, 2011 (received for review July 7, 2010)
The human APOBEC3 (A3A–A3H) locus encodes six cytidine deam-inases that edit single-stranded DNA, the result being DNA pep-pered with uridine. Although several cytidine deaminases areclearly restriction factors for retroviruses and hepadnaviruses, itis not known if APOBEC3 enzymes have roles outside of these set-tings. It is shown here that both human mitochondrial and nuclearDNA are vulnerable to somatic hypermutation by A3 deaminases,with APOBEC3A standing out among them. The degree of editing ismuch greater in patients lacking the uracil DNA-glycolyase gene,indicating that the observed levels of editing reflect a dynamiccomposed of A3 editing and DNA catabolism involving uracilDNA-glycolyase. Nonetheless, hyper- and lightly mutated sequen-ces went hand in hand, raising the hypothesis that recurrent low-level mutation by APOBEC3A could catalyze the transition froma healthy to a cancer genome.
The human APOBEC3 (A3) locus encodes seven expressedgenes (A3A–A3H), of which six are demonstrably cytidine
deaminases with specificity for single-stranded DNA (ssDNA)(1). The A3 locus arose in placental mammals by duplication ofAICDA and subsequent expansion (2). Activation-induced cyti-dine deaminase (AICDA) is presently the only enzyme known toinduce mutations in the human genome and initiates class-switchrecombination and somatic hypermutation of rearranged Ig genes(3). Several human A3 genes are up-regulated in inflammatorysettings by type I and II interferons, although activation dependsto some degree on the cell type (4–8). Given this finding, it is notsurprising that A3F and A3G are involved in the restriction ofretroviruses and hepadnaviruses where nascent single-strandedcDNA remains vulnerable until double-strand synthesis is com-pleted (9–16). In late stages of hepatitis B virus- (HBV) associ-ated cirrhosis there was significant up-regulation of four to fiveA3genes, with A3G being the major restriction factor able to editup to 35% of HBV genomes (17).DNA viral genomes, such as human papillomavirus (HPV),
can also undergo editing both in vivo and in tissue culture,presumably during transcription or replication (18). Three A3deaminases, A3A, A3C, and A3H could hyperedit HPV genomesex vivo, suggesting that the phenomenon was occurring in thenucleus because the strictly cytoplasmic deaminases, A3F andA3G, did not edit transfected HPV DNA (18). Along with a re-cent article showing that A3A could edit transfected EGFPplasmid DNA (7), they highlight the potential of A3 enzymes toedit foreign DNA. The presence of uridine in cellular DNA isread as a danger signal, the uracil base resulting from cytidinedeamination being subject to excision, particularly by uracilDNA-glycosylase (UNG), the nuclear form (UNG2) being farmore active than its mitochondrial counterpart (UNG1). Uracilexcision followed by abasic endonuclease cleavage of the DNAstrand can lead to repair or degradation. Small fragments of nu-clear and mitochondrial DNA (mtDNA) can be found in thecytoplasm, which trigger a number of cytoplasmic DNA sensormolecules and are associated with DNA degradation (19, 20).
As there are six human A3 enzymes, all with ssDNA specificity,the question arises as to whether they can edit host-cell DNA.Given the high mtDNA copy number per cell, we explored thepossibility that a fraction of mtDNA might be vulnerable to A3editing, especially in inflamed tissues where there is demonstrableup-regulation of A3 genes (17). We show here that a small frac-tion of human mitochondrial genomes are phenomenally editedby APOBEC3 deaminases in the cytoplasm. It transpires thatnuclear DNA (nuDNA), notably segments ofMYC and TP53, canbe hyperedited by APOBEC3A. For both mitochondrial andnuDNA, editing reflects a dynamic between A3 activity andDNA degradation involving UNG.
ResultsAPOBEC3 Hypermutation of mtDNA. A nested PCR/3DPCR ap-proach was taken to the amplification of mtDNA from 36 DNAsamples spanning alcoholic, HBV+, HBV+HCV+, and HCV+
cirrhosis (17, 21). A region corresponding to part of MT-COIat ∼6,200 bp was chosen, as it maps within the large single-stranded replication intermediate. Mitochondrial DNA could berecovered at temperatures as low as 82.2 °C from 17 of 36 (∼47%)samples tested, much lower than for a control liver or clonedMT-COI DNA (Td∼86 °C) (Fig. 1 A and B). Molecular cloning andsequencing of the PCRproducts proved that they represented bonafide cytidine-edited mtDNA, with both strands being vulnerable(Fig. 1C and Fig. S1A), although minus-strand hypermutantswere generally more abundant (shown asG->Ahypermutants withrespect to the plus strand). Although minus-strand editing wasgenerally in the 30 to 80%range, occasionally a sequencewith up to98% of targets could be edited (Fig. 1 C and D).As A3 and AICDA catalyzed cytidine deamination is partic-
ularly sensitive to the 5′ base, a bulk dinucleotide analysis wasperformed for minus- and plus-strand hypermutants (Fig. 1 Eand F). The strong biases for TpC and CpC among minus-strandhypermutants are hallmarks of APOBEC3 editing (11, 15, 22).For plus-strand editing, the clear preference for CpC is a hall-mark of A3G (Fig. 1F), no matter the template (11). Theseslightly different patterns probably reflect the greater C contentof the plus strand generating more oligoC tracts, which areknown hotspots for A3G. A clonal analysis of the dinucleotide
Author contributions: R.S., J.-P.V., and S.W.-H. designed research; R.S., M.-M.A., D.G.,M.H., G.E., and A.M. performed research; P.P. and A.D. contributed new reagents/analytictools; R.S., M.-M.A., J.-P.V., and S.W.-H. analyzed data; and J.-P.V. and S.W.-H. wrote thepaper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission. R.S.H. is a guest editor invited by the EditorialBoard.1R.S. and M.-M.A. contributed equally to this work.2To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1009687108/-/DCSupplemental.
4858–4863 | PNAS | March 22, 2011 | vol. 108 | no. 12 www.pnas.org/cgi/doi/10.1073/pnas.1009687108
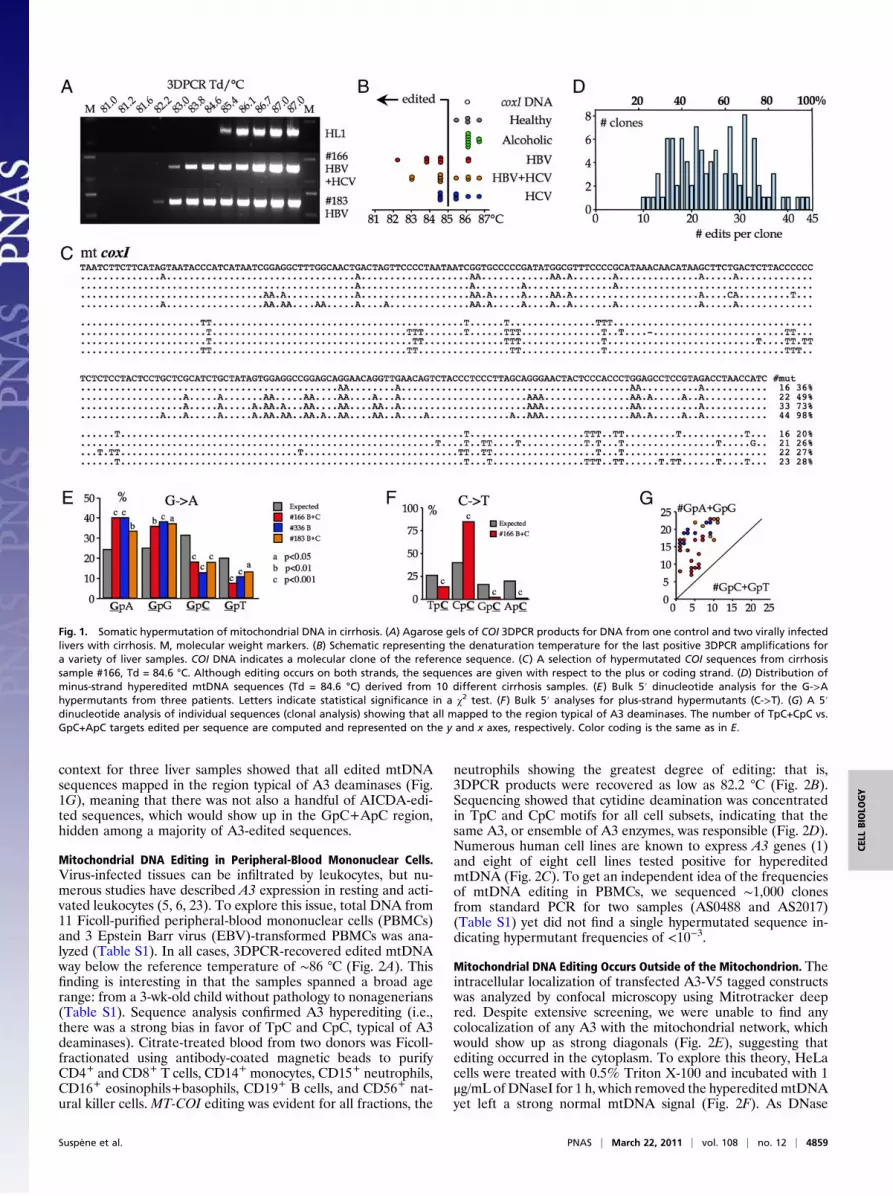
context for three liver samples showed that all edited mtDNAsequences mapped in the region typical of A3 deaminases (Fig.1G), meaning that there was not also a handful of AICDA-edi-ted sequences, which would show up in the GpC+ApC region,hidden among a majority of A3-edited sequences.
Mitochondrial DNA Editing in Peripheral-Blood Mononuclear Cells.Virus-infected tissues can be infiltrated by leukocytes, but nu-merous studies have described A3 expression in resting and acti-vated leukocytes (5, 6, 23). To explore this issue, total DNA from11 Ficoll-purified peripheral-blood mononuclear cells (PBMCs)and 3 Epstein Barr virus (EBV)-transformed PBMCs was ana-lyzed (Table S1). In all cases, 3DPCR-recovered edited mtDNAway below the reference temperature of ∼86 °C (Fig. 2A). Thisfinding is interesting in that the samples spanned a broad agerange: from a 3-wk-old child without pathology to nonagenerians(Table S1). Sequence analysis confirmed A3 hyperediting (i.e.,there was a strong bias in favor of TpC and CpC, typical of A3deaminases). Citrate-treated blood from two donors was Ficoll-fractionated using antibody-coated magnetic beads to purifyCD4+ and CD8+ T cells, CD14+ monocytes, CD15+ neutrophils,CD16+ eosinophils+basophils, CD19+ B cells, and CD56+ nat-ural killer cells. MT-COI editing was evident for all fractions, the
neutrophils showing the greatest degree of editing: that is,3DPCR products were recovered as low as 82.2 °C (Fig. 2B).Sequencing showed that cytidine deamination was concentratedin TpC and CpC motifs for all cell subsets, indicating that thesame A3, or ensemble of A3 enzymes, was responsible (Fig. 2D).Numerous human cell lines are known to express A3 genes (1)and eight of eight cell lines tested positive for hypereditedmtDNA (Fig. 2C). To get an independent idea of the frequenciesof mtDNA editing in PBMCs, we sequenced ∼1,000 clonesfrom standard PCR for two samples (AS0488 and AS2017)(Table S1) yet did not find a single hypermutated sequence in-dicating hypermutant frequencies of <10−3.
Mitochondrial DNA Editing Occurs Outside of the Mitochondrion. Theintracellular localization of transfected A3-V5 tagged constructswas analyzed by confocal microscopy using Mitrotracker deepred. Despite extensive screening, we were unable to find anycolocalization of any A3 with the mitochondrial network, whichwould show up as strong diagonals (Fig. 2E), suggesting thatediting occurred in the cytoplasm. To explore this theory, HeLacells were treated with 0.5% Triton X-100 and incubated with 1μg/mL of DNaseI for 1 h, which removed the hyperedited mtDNAyet left a strong normal mtDNA signal (Fig. 2F). As DNase
Fig. 1. Somatic hypermutation of mitochondrial DNA in cirrhosis. (A) Agarose gels of COI 3DPCR products for DNA from one control and two virally infectedlivers with cirrhosis. M, molecular weight markers. (B) Schematic representing the denaturation temperature for the last positive 3DPCR amplifications fora variety of liver samples. COI DNA indicates a molecular clone of the reference sequence. (C) A selection of hypermutated COI sequences from cirrhosissample #166, Td = 84.6 °C. Although editing occurs on both strands, the sequences are given with respect to the plus or coding strand. (D) Distribution ofminus-strand hyperedited mtDNA sequences (Td = 84.6 °C) derived from 10 different cirrhosis samples. (E) Bulk 5′ dinucleotide analysis for the G->Ahypermutants from three patients. Letters indicate statistical significance in a χ2 test. (F) Bulk 5′ analyses for plus-strand hypermutants (C->T). (G) A 5′dinucleotide analysis of individual sequences (clonal analysis) showing that all mapped to the region typical of A3 deaminases. The number of TpC+CpC vs.GpC+ApC targets edited per sequence are computed and represented on the y and x axes, respectively. Color coding is the same as in E.
Suspène et al. PNAS | March 22, 2011 | vol. 108 | no. 12 | 4859
CELL
BIOLO
GY

treatment alone did not reduce the edited DNA signal, thisrules out extracellular mtDNA as a source for the signal (Fig. 2F).If hyperediting occurred outside of the mitochondrion, it shouldnot be subject to DNA repair. Like all archaeabacterial poly-merases, Pfu is unable to amplify DNA harboring dU (24). Weperformed 3DPCR on first round PCRmtDNA amplified at 95 °Cusing Pfu. This strategy failed to recover any genomes beyondthe 85.4 to 86 °C benchmark temperature for unedited mtDNA(Fig. 2F). These findings show that A3 editing of mtDNA occursoutside of the mitochondrion and goes uncorrected.The high levels of hyperedited mtDNA in established cell lines
has thwarted detailed analysis of the phenomenon by trans-fection experiments. Only for A3A transfections, and to a lesserextent A3G, could a clear signal over background be found (Fig.2F). Uracil-DNA glycosylase inhibitor (UGI) is a small proteinfrom a Bacillus subtilis bacteriophage that can inhibit mammalianUNG, a crucial enzyme involved in excising uracil from DNA(25). When A3A or A3G were cotransfected with the UGI plas-mid, mtDNA recovered at temperatures as low as 81 °C (Fig. 2F).
Given this finding, we next turned to EBV transformed B-celllines from three ung−/− individuals [P1, P2, and P3 (26)].Hyperedited mtDNA was detected for all three samples down to81 °C, confirming the results had with UGI (Fig. 2F). To de-termine A3 expression levels, real-time quantitative PCR wasperformed for P1, P2, and P3, as well as two ung+ control EBV-transformed cell lines, Z (AICDA−/−) and T (Fig. S2A). Datawere normalized to the expression levels of four invariable ref-erence genes (CLK2, GUSB, HMBS, TBP). Although A3 ex-pression was in general similar, A3A showed some variationamong the samples (Fig. S2A). Analysis of total PBMC DNAfrom one of these individuals (P1) yielded the same result as thecell line, ruling out any influence from EBV on the observation.For P2 DNA, the frequency of mtDNA hypermutants in stan-dard PCR products was assessed by deep sequencing. Approxi-mately 0.5% were edited, greater than for PBMC DNA fromnormal individuals (vide supra). This finding translates to ∼25 percell, assuming ∼5 K mtDNA genomes per cell.
Fig. 2. Mitochondrial DNA editing occurs among PBMCs and in the cytoplasm. (A) Schematic representation for 3DPCR amplification for human PBMCs, EBV-transformed PBMCs, and purified sperm. (B) Comparable representation for PBMC subsets from donors ABD2 and -4. (C) Comparable representation forestablished human cell lines. The A3C1 and WTI lines are derivatives of Jurkat. (D) Clonal analysis of edited mtDNA sequences for PBMC cell subsets fromdonor ABD4. Color coding corresponds to that in B. (E) Confocal microscopic analysis of double positive A3-V5 transfected HeLa cells plus Mitotracker deepred. The absence of a diagonal in any plot indicates a lack of colocalization. (F) 3DPCR temperature-gradient agarose gels revealing recovery of editedmtDNA. The legend to the right describes the treatment, transfection, and sample; ± DNaseI-treated HeLa cells lyzed by Triton X-100 indicating that editingoccurs in the cytoplasm; Taq/Taq or Pfu/Taq indicates first-round amplification using Taq or Pfu DNA polymerase followed by Taq for 3DPCR. Pfu is unable tocopy DNA containing dU; pv, empty plasmid; UGI refers to the phage UNG inhibitor; P1-3 are human ung−/− cell lines and P1 PBMCs indicates total PBMC DNAfrom patient P1. (G) Schematic of last positive 3DPCR amplification temperature for the quail QT6 cell line transfected by a series of APOBEC plasmids induplicate. The prefixes “h” and “m” denote human and mouse, respectively. Both hA3Gn and hA3Gc denote the N and C domains alone of hA3G; the C288Sand C291S mutants of hA3G are inactive.
4860 | www.pnas.org/cgi/doi/10.1073/pnas.1009687108 Suspène et al.
Limiting-dilution of PBMCs and purified CD4 cells from pa-tient ABD2 were performed in triplicate, the hyperedited3DPCR signal being lost between 1,000 and 300 cells (f ≥ 10−3).A similar triplicate titration of P2 cells resulted in a loss of signalbetween 50 and 10 cells (f ≥ 2 × 10−2). Given 3DPCR as readout,these frequencies are underestimates. For P2 cells where thefrequencies of cells harboring edited mtDNA and bulk hyper-edited mtDNA are known, it can be concluded that a sizeablefraction of cells harbors a small fraction of hyperedited mtDNAsequences.All of the above analyses concerned editing of the theMT-COI
gene. To be sure that there was no bias, part of the MT-CYB andMT-ND2 genes from the cell lines P1, P2, and P3 were analyzedby 3DPCR. Like MT-COI, the former maps to the ssDNA rep-lication intermediate but MT-ND2 mapped to the dsDNA re-gion. Both DNA strands of MT-CYB and MT-ND2 were heavilyedited; a selection from P2 cells is shown in Fig. S1B.Although A3A and A3G can edit mtDNA, this doesn’t exclude
other A3 enzymes that could be individually, or together, re-sponsible for the high background. To address this question weused the QT6 quail cell line that has never shown an endogenouscytidine-editing background for retroviruses, presumably becausethe avian lineage does not encode A1 or A3 othologs (11). QT6cells were transfected by numerous APOBEC plasmids. ThePCR/3DPCR approach recovered hyperedited quail mtDNA for5 of 11 human cytidine deaminases, notably A3A, A3C, A3F,A3G, and A3H; the murine A1 andA3 enzymes were also capableof editing QT6 mtDNA (Fig. 2G).Given this result, it is possible that mtDNA editing may occur
beyond the primate world, with its seven-gene A3 locus. For
other mammals, the A3 gene number presently ranges from one(mice, rats, pigs), to three (cats) and six (horses) (27). In con-trast, birds, reptiles, and fish do not encode an A3 gene. EditedmtDNA was recovered from tree shrew hepatocyte cultures(Tupaia belangeri belongs to Scandentia, the closest order toPrimates) as well as horse and goat PBMCs (Table S2). In allcases, 5′ dinucleotide analysis revealed a penchant for editingwithin TpC and CpC, typical of A3 enzymes. Mouse DNA fromnumerous tissues was systematically negative for mtDNA editing(Table S2). These findings show that the phenomenon ofmtDNA editing is not uniform across placental mammals.
A3 Editing of the Human nuDNA. As the A3A, A3C, and A3Henzymes in particular can locate to the nucleus, we tested thepossibility that, occasionally, nuDNA might be vulnerable to A3editing. MYC is particularly mutated in leukemias and lympho-mas, and the tumor-suppressor gene TP53 is mutated in ∼50% ofcancers, with exon 8 encoding many hotspots (http://p53.free.fr).Several of the human cirrhosis and PBMC samples wheremtDNA editing was in evidence were screened using primers forMYC exon 2 and TP53 exon 8/intron 9. We were unable to re-cover massively hypermutated MYC or TP53 sequences.Given the impact of UNG on mtDNA editing (Fig. 2F), we
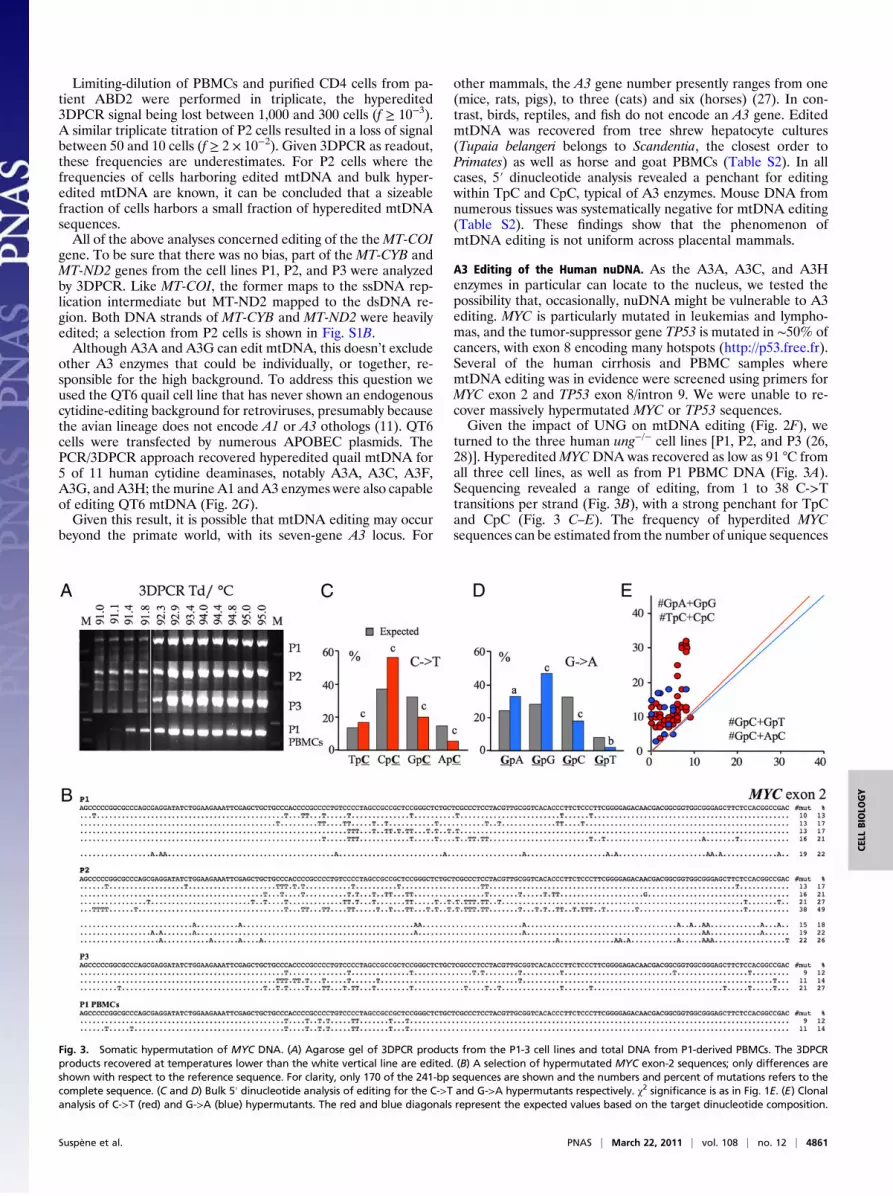
turned to the three human ung−/− cell lines [P1, P2, and P3 (26,28)]. HypereditedMYCDNA was recovered as low as 91 °C fromall three cell lines, as well as from P1 PBMC DNA (Fig. 3A).Sequencing revealed a range of editing, from 1 to 38 C->Ttransitions per strand (Fig. 3B), with a strong penchant for TpCand CpC (Fig. 3 C–E). The frequency of hyperdited MYCsequences can be estimated from the number of unique sequences
Fig. 3. Somatic hypermutation of MYC DNA. (A) Agarose gel of 3DPCR products from the P1-3 cell lines and total DNA from P1-derived PBMCs. The 3DPCRproducts recovered at temperatures lower than the white vertical line are edited. (B) A selection of hypermutated MYC exon-2 sequences; only differences areshown with respect to the reference sequence. For clarity, only 170 of the 241-bp sequences are shown and the numbers and percent of mutations refers to thecomplete sequence. (C and D) Bulk 5′ dinucleotide analysis of editing for the C->T and G->A hypermutants respectively. χ2 significance is as in Fig. 1E. (E) Clonalanalysis of C->T (red) and G->A (blue) hypermutants. The red and blue diagonals represent the expected values based on the target dinucleotide composition.
Suspène et al. PNAS | March 22, 2011 | vol. 108 | no. 12 | 4861
CELL
BIOLO
GY

derived from 0.5-μg input DNA (∼150 K copies). As ∼30 to 50%of sequences were identical, the number of distinct sequences is inthe range of the number of unique sequences. Hence, thehypereditedMYC frequencies are in the range of 20/150 K ∼10−4.At such low frequencies, correlations with macroscopic, such asbulk transciption, levels are going to be fraught with problems.To explore nuDNA editing experimentally, we transfected
HeLa cells with individual A3-V5 constructs ±UGI and soughthyperedited MYC DNA at 72 h posttransfection. Only for theA3A+UGI combination (three of three transfections) was hy-pereditedMYCDNA recovered (Fig. S3 A and E). When a lowertemperature range was explored, DNA was recovered down to88 °C. Phenomenally edited sequences with up to ∼70% targetresidues edited were recovered (Fig. S3B). Although both strandswere edited, slightly fewer plus-strand hypermutants were re-covered. Therewas considerable sequence heterogeneity, indicatingmultiple independent deamination events. Not surprisingly, thedegree of editing was less for 3DPCR products recovered at 92.3than at 91 °C (Fig. S3C). Indeed, sequences with one to three C->Ttransitions were recovered, suggesting that there is a very largespectrum from editing, essentially from a few to ∼70%. The 5′ di-nucleotide context was strongly in favor of TpC+CpC and biasedagainst GpC+ApC (Fig. S3D).Not surprisingly, A3A transfection of a 293T cells stably
expressing UGI resulted in hyperedited MYC DNA (Fig. S3B).Nuclear and cytoplasmic fractions were made from P1, P2, P3,and A3A ± UGI transfected cells according to a variety of dif-ferent protocols. However, the fractions were insufficiently pureto draw conclusions as to the site of nuDNA editing. As UNGimpacted the detectable levels of MYC editing, we attempted theinverse, transfection by both A3A and the nuclear form of hu-man UNG, UNG2. As P3 gave the highest A3A genes expression(Fig. S2A), it and 293T-UGI cells were transfected with andwithout UNG2 (29). For both P3 and 293T-UGI cells, less editedMYC DNA was recovered compared with the A2-negative con-trol, reinforcing the notion that observed levels of editing reflecta dynamic involving UNG (Fig. S2 B and C).In an analogous fashion, the PCR/3DPCR approach was ap-
plied to a region spanning most of exon 8 and part of intron 9 ofTP53. Like MYC, hypermutated DNA was obtained only fromthe A3A+UGI cotransfection (Fig. S4A). The data are highlycomparable with massively edited sequences harboring up to59% of mutated cytidine residues, with the exception that onlyminus-strand hypermutants were found (Fig. S4B). A3A editingspanned both exon 8 and intron 9 without discrimination. Editingwas once again biased in favor of TpC+CpC (Fig. S4C).
DiscussionThe antiviral roles hitherto ascribed to A3 cytidine deaminasesfit well with the observation that numerous human A3 geneswere up-regulated by interferons (4, 5, 7, 8). It turns out that thisactivity represents just one aspect, the present data revealing anunsuspected pathway for DNA catabolism of both humangenomes initiated by cytidine editing, with A3A being particu-larly important for nuDNA editing. UNG impacts the observedlevels of editing, but DNases may also be part of the equation,and follows from the greater editing frequencies in ung−/− cells.Accordingly, there is more A3 editing than detected in normalsamples. Given the mutation load, A3 hyperedited nuDNA issynonymous with cell death, the phenomenal loss of informationmaking the process irreversible.Any hypothesis concerning mtDNA or nuDNA editing has to
address a sequence with >50% of edited cytidines. To date,editing of this sort is found within the close confines of retroviralcapsid structures, whereA3 concentrations are in the 20- to 200-μMrange, phenomenal enzyme concentrations by anymeasure, whichare essentially the result of capsid volumes in the zeptolitre range(30, 31). In contrast, A3G hyperediting of ssDNA in vitro pla-
teaued at∼20%of cytidines deaminated at∼2 μMof enzyme (32),meaning that the hyperediting observed cannot be simply thechance encounter of free enzyme and target ssDNA in a per-meablized or collapsed cell. The Triton-dependent DNase sensi-tivity of edited mtDNA supports this deduction. Hypermutatedhuman DNA therefore implies compartmentalization or cofactorswhere ssDNA and A3 remain together, rather than permitted todrift apart, and so contributing to the efficiency of the catabolicpathway. With this finding in mind, it must be noted that there isa hierarchy to A3 editing: HBV genomes are vulnerable to editingby six hA3 enzymes (17) and mtDNA could be edited by five (Fig.2E). HPV DNA was hyperedited by the three Zinc-finger mono-domaindeaminasesA3A,A3C, andA3H,but nuDNA is vulnerableto A3A only. Hence, nuDNA editing cannot result from a non-physiological encounter of nuDNA with A3A, say in a lysed cell,because the other active A3 enzymes would be involved.Although A3 enzymes are involved in catabolizing DNA, there
was a broad continuum with lightly edited sequences occurringalongside the hyperedited sequences (Fig. S3C). As 3DPCRselects highly edited over lightly edited DNA, the fraction of thelatter is certainly underestimated (17). Obviously, cells with A3-edited genomes will be subject to the winnowing power of pu-rifying selection, with few survivors. The crucial question is, de-spite the highly deleterious mutant spectrum resulting from A3editing, can minute numbers of such cells escape death?The answer may well be positive for several reasons. First, A1
and AICDA transgenic mice generally develop cancers dictated bythe tg promoter (33–35). Second, the explosion in cancer genomicsis showing that thousands of GC->AT transitions characterizea cancer genome (36–38). Third, although attention is focused onthe massively edited mitochondrial and nuDNA sequences, chro-mosomal DNA from a majority of P1 to P3 cells (ung−/− cell lines)contains uracil, as shown by the comet assay (28). As A3 genes areup-regulated by inflammatory environments (4, 5, 7, 8, 17), it seemsthat A3 deaminases will provide a recurrent source of genomicstress, particularly for persistent infections like viral hepatitis.In conclusion, there is far more cytidine deamination of hu-
man genomes than hitherto suspected. Indeed, the notion ofspontaneous cytidine deamination may well need revisiting. Themutation load generated by hypermutation is incompatible withreplication, indicating an unsuspected mechanism of DNA ca-tabolism. The findings reveal a physiological role for the A3enzymes outside of the antiviral paradigm. Clearly, several A3enzymes can function as human DNA mutators, and APO-BEC3A, like AICDA, becomes a serious candidate for nudginggenomes down the long and winding road to cancer.
Materials and MethodsSamples. All of the liver materials have been previously described (17). Thestudy was approved by an institutional human research review board (RBM2005–019). Informed consent was obtained for each patient. The PBMCsamples were either from healthy anonymous donors or obtained by writteninformed consent from sperm donors, following approval by local ethicscommittees (Comités Consultatifs de Protection des Personnes dans laRecherche Biomédicale N°98353, and Comité de protection des personnes,number AC-2009–886 Germéthèque) and the institutional review board(Centre de Recherche de Biochimie 2003.6).
Cells. One million HeLa cells were seeded in six-well plates and transfected 24h later by 3 μg of total plasmid using Fugene6 (Roche). Three days later, cellswere washed in cold PBS, covered by 1 mL of 10 mM Tris.HCl pH7.4, 2 mMMgCl2, and put on ice for 5 min, after which they were incubated with 0.5%(final) Triton X-100 for 5 min on ice. Lysis was confirmed by microscopy. Cellswere subsequently incubated with 1 U/μL DNaseI (Roche) for 1 h at 37 °C,after which DNA was extracted as per usual.
Confocal Microscopy. Approximately 105 HeLa cells were transfected in 24-well plates with C-terminal V5-tagged A3 constructs. At 72 h post-transfection, cells were incubated with 400 nM Mitotracker Deep Red FM(Invitrogen) for 40 min at 37 °C. After PBS washing, cells were fixed in 50/50
4862 | www.pnas.org/cgi/doi/10.1073/pnas.1009687108 Suspène et al.

methanol/acetone for 20 min at −20 °C. The anti-V5 antibody (Invitrogen)was incubated at 1/200 for 1 h at room temperature, followed by incubationwith a fluoresceine-conjugated secondary antibody for 1 h at room tem-perature. Slides were stained with DAPI and mounted with Vectashield.Confocal Imaging was performed using a Zeiss AxioImagerZ2 LSM700.
PCR. All DNAs were extracted using the Epicentre kit. All amplifications wereperformed using first-round standard PCR followed by nested 3DPCR (21)(Table S3). PCR was performed with 2.5 U Taq (Bioline) or 2.5 U Pfu (Stra-tagene) DNA polymerase per reaction. PCR products were cloned using theTOPO vector and sequencing was outsourced to Cogenics. Detailed in-formation on PCR, 3D-PCR and real time quantitative PCR are provided in SIMaterials and Methods.
ACKNOWLEDGMENTS. We thank Drs. Carlo Battiston, Vincenzo Mazzaferro,Kenneth McElreavey, Marie-Noëlle Ungeheuer, and Vesna Mellon for humanDNA samples, Prof. Wolfram Gerlich for tupaia liver samples, Dr. PhilippeBlancou for horse and goat blood, Dr. Francina Langa Vives for the purifiedmouse oocytes, Dr. Anne Durandy for the five Epstein Barr virus-transformedB-cell lines and P1 DNA, Dr. Vincenzo Di Bartolo for the WTI and A3C1 celllines, Pascal Roux for help with confocal microscopy, and Christophe Rusniokfor providing invaluable help with bioinformatics. This work is supported inpart by a postdoctoral fellowship from l’Association pour la Recherche sur leCancer (to R.S.); a graduate fellowship from La Ligue contre le Cancer (toM.-M.A.); the Pasteur Foundation through its Zuccaire Internship Program(G.E.); and Grants from the Institut Pasteur, Agence Nationale de Recherchessur le SIDA, Agence Nationale de Recherches, Institut National de la Santé etde la Recherche Médicale, and the Centre National de la Recherche Scientifi-que. The Unit is “Equipe labelisée LIGUE 2010.”
1. Jarmuz A, et al. (2002) An anthropoid-specific locus of orphan C to U RNA-editing
enzymes on chromosome 22. Genomics 79:285–296.2. Conticello SG, Thomas CJ, Petersen-Mahrt SK, NeubergerMS (2005) Evolution of the AID/
APOBEC family of polynucleotide (deoxy)cytidine deaminases.Mol Biol Evol 22:367–377.3. Di Noia JM, Neuberger MS (2007) Molecular mechanisms of antibody somatic
hypermutation. Annu Rev Biochem 76:1–22.4. Bonvin M, et al. (2006) Interferon-inducible expression of APOBEC3 editing enzymes
in human hepatocytes and inhibition of hepatitis B virus replication. Hepatology 43:
1364–1374.5. Koning FA, et al. (2009) Defining APOBEC3 expression patterns in human tissues and
hematopoietic cell subsets. J Virol 83:9474–9485.6. Refsland EW, et al. (2010) Quantitative profiling of the full APOBEC3mRNA repertoire in
lymphocytes andtissues: Implications forHIV-1 restriction.NucleicAcidsRes38:4274–4284.7. Stenglein MD, Burns MB, Li M, Lengyel J, Harris RS (2010) APOBEC3 proteins mediate
the clearance of foreign DNA from human cells. Nat Struct Mol Biol 17:222–229.8. Wang FX, Huang J, Zhang H, Ma X, Zhang H (2008) APOBEC3G upregulation by alpha
interferon restricts human immunodeficiency virus type 1 infection in human
peripheral plasmacytoid dendritic cells. J Gen Virol 89:722–730.9. Chelico L, Pham P, Calabrese P, Goodman MF (2006) APOBEC3G DNA deaminase acts
processively 3′ —> 5′ on single-stranded DNA. Nat Struct Mol Biol 13:392–399.10. Harris RS, et al. (2003) DNA deamination mediates innate immunity to retroviral
infection. Cell 113:803–809.11. Henry M, et al. (2009) Genetic editing of HBV DNA by monodomain human APOBEC3
cytidine deaminases and the recombinant nature of APOBEC3G. PLoS ONE 4:e4277.12. Lecossier D, Bouchonnet F, Clavel F, Hance AJ (2003) Hypermutation of HIV-1 DNA in
the absence of the Vif protein. Science 300:1112.13. Mangeat B, et al. (2003) Broad antiretroviral defence by human APOBEC3G through
lethal editing of nascent reverse transcripts. Nature 424:99–103.14. Mariani R, et al. (2003) Species-specific exclusion of APOBEC3G from HIV-1 virions by
Vif. Cell 114:21–31.15. Suspène R, et al. (2005) Extensive editing of both hepatitis B virus DNA strands by
APOBEC3 cytidine deaminases in vitro and in vivo. ProcNatl Acad Sci USA 102:8321–8326.16. Wiegand HL, Doehle BP, Bogerd HP, Cullen BR (2004) A second human antiretroviral
factor, APOBEC3F, is suppressed by the HIV-1 and HIV-2 Vif proteins. EMBO J 23:
2451–2458.17. Vartanian JP, et al. (2010) Massive APOBEC3 editing of hepatitis B viral DNA in
cirrhosis. PLoS Pathog 6:e1000928.18. Vartanian JP, Guétard D, Henry M, Wain-Hobson S (2008) Evidence for editing of
human papillomavirus DNA by APOBEC3 in benign and precancerous lesions. Science
320:230–233.19. Fernandes-Alnemri T, Yu JW, Datta P, Wu J, Alnemri ES (2009) AIM2 activates the
inflammasome and cell death in response to cytoplasmic DNA. Nature 458:509–513.20. Stetson DB, Ko JS, Heidmann T, Medzhitov R (2008) Trex1 prevents cell-intrinsic
initiation of autoimmunity. Cell 134:587–598.
21. Suspène R, Henry M, Guillot S, Wain-Hobson S, Vartanian JP (2005) Recovery ofAPOBEC3-edited human immunodeficiency virus G->A hypermutants by differentialDNA denaturation PCR. J Gen Virol 86:125–129.
22. Beale RC, et al. (2004) Comparison of the differential context-dependence of DNAdeamination by APOBEC enzymes: Correlation with mutation spectra in vivo. J MolBiol 337:585–596.
23. Chen K, et al. (2006) Alpha interferon potently enhances the anti-humanimmunodeficiency virus type 1 activity of APOBEC3G in resting primary CD4 T cells. JVirol 80:7645–7657.
24. Greagg MA, et al. (1999) A read-ahead function in archaeal DNA polymerases detectspromutagenic template-strand uracil. Proc Natl Acad Sci USA 96:9045–9050.
25. Wang Z, Mosbaugh DW (1988) Uracil-DNA glycosylase inhibitor of bacteriophagePBS2: Cloning and effects of expression of the inhibitor gene in Escherichia coli. JBacteriol 170:1082–1091.
26. Imai K, et al. (2003) Human uracil-DNA glycosylase deficiency associated withprofoundly impaired immunoglobulin class-switch recombination. Nat Immunol 4:1023–1028.
27. LaRue RS, et al. (2008) The artiodactyl APOBEC3 innate immune repertoire showsevidence for a multi-functional domain organization that existed in the ancestor ofplacental mammals. BMC Mol Biol 9:104.
28. Kavli B, et al. (2005) B cells from hyper-IgM patients carrying UNG mutations lackability to remove uracil from ssDNA and have elevated genomic uracil. J Exp Med 201:2011–2021.
29. Chen R, Le Rouzic E, Kearney JA, Mansky LM, Benichou S (2004) Vpr-mediatedincorporation of UNG2 into HIV-1 particles is required to modulate the virus mutationrate and for replication in macrophages. J Biol Chem 279:28419–28425.
30. Benjamin J, Ganser-Pornillos BK, Tivol WF, Sundquist WI, Jensen GJ (2005) Three-dimensional structure of HIV-1 virus-like particles by electron cryotomography. J MolBiol 346:577–588.
31. Watts NR, et al. (2002) The morphogenic linker peptide of HBV capsid protein formsa mobile array on the interior surface. EMBO J 21:876–884.
32. Suspène R, et al. (2004) APOBEC3G is a single-stranded DNA cytidine deaminase andfunctions independently of HIV reverse transcriptase. Nucleic Acids Res 32:2421–2429.
33. Okazaki IM, et al. (2003) Constitutive expression of AID leads to tumorigenesis. J ExpMed 197:1173–1181.
34. Takai A, et al. (2009) A novel mouse model of hepatocarcinogenesis triggered by AIDcausing deleterious p53 mutations. Oncogene 28:469–478.
35. Yamanaka S, et al. (1995) Apolipoprotein B mRNA-editing protein induceshepatocellular carcinoma and dysplasia in transgenic animals. Proc Natl Acad Sci USA92:8483–8487.
36. Greenman C, et al. (2007) Patterns of somatic mutation in human cancer genomes.Nature 446:153–158.
37. Pleasance ED, et al. (2010) A small-cell lung cancer genome with complex signaturesof tobacco exposure. Nature 463:184–190.
38. Sjöblom T, et al. (2006) The consensus coding sequences of human breast andcolorectal cancers. Science 314:268–274.
Suspène et al. PNAS | March 22, 2011 | vol. 108 | no. 12 | 4863
CELL
BIOLO
GY




















