
Sequencing the genome of Marssonina brunnea reveals fungus-poplar co-evolution

Research in Microbiology 161 (2010) 720e728www.elsevier.com/locate/resmic
b-Galactofuranose-containing structures present in the cell wall of thesaprophytic fungus Cladosporium (Hormoconis) resinae
Renata Calixto a, Bianca Mattos a, Vera Bittencourt b, Lıvia Lopes a, Lauro Souza c,Guilherme Sassaki c, Thales Cipriani c, Maria Silva a, Eliana Barreto-Bergter a,*
a Instituto de Microbiologia, Centro de Ciencias da Saude, Universidade Federal do Rio de Janeiro (UFRJ),Bloco I, Ilha do Fundao, 21941-970, Rio de Janeiro,
RJ, Brazilb Instituto Biomedico, UNIRIO, Rio de Janeiro, RJ, Brazil
cDepartamento de Bioquımica e Biologia Molecular, Universidade Federal do Parana (UFPR), 81531-980, Curitiba, PR, Brazil
Received 12 March 2010; accepted 18 July 2010
Available online 7 August 2010
Abstract
A peptidogalactomannan was isolated from mycelia of Cladosporium (Hormoconis) resinae and characterized using methylationefrag-mentation analysis, partial acid hydrolysis and 1H and 13C-NMR spectroscopy. The galactomannan component was a branched structure andconsisted of a main chain containing (1/6)-linked a-D-Manp residues substituted at O-2 by side chains containing (1/2)-linked a-D-Manpresidues. b-D-Galf residues were present as side chains of 3e4 units that are (1/5)-interlinked. This structure is very similar to a pGM isolatedfrom Aspergillus fumigatus and differs from that of Cladosporium werneckii (currently named Hortaea werneckii), with this pGM and otherfungal galactomannans having single terminal (1/6)-linked b-Galf residues. The importance of the carbohydrate moiety of Cladosporiumresinae pGM in immunoassays was also demonstrated. On FACS examination, a decrease (60%) in rabbit serum anti- C. resinae binding toC. resinae conidia occurred when this serum had been previously incubated with pGMs from C. resinae and A. fumigatus or with mannoproteinfrom Candida parapsilosis, suggesting the presence of cross-reactive determinants in these fungi.� 2010 Elsevier Masson SAS. All rights reserved.
Keywords: Cladosporium resinae; Hormoconis resinae; Peptidogalactomannan; NMR spectroscopy; Carbohydrate epitopes
1. Introduction
Species of the genus Cladosporium are relevant chromo-blastomycosis agents (Cladophialophora carrionii, currentlynamed C. carrionii) (Yegres et al., 1991; Vitale et al., 2009),allergens (Cladosporium cladosporioides, C. herbarum)(Latge et al., 1988), and plant pathogens (Cladosporium ful-vum, C. herbarum) (van Kan et al., 1992). Cladosporiumresinae (Hormoconis resinae) is a saprophytic species,commonly found as a contaminant in fuel oil storage tanks(Lindley and Heyderman, 1985; Seifert et al., 2007). Aprevious report on analysis of the cell wall from two strains of
* Corresponding author. Tel.: þ55 21 25626741; fax: þ55 21 25608344.
E-mail address: [email protected] (E. Barreto-Bergter).
0923-2508/$ - see front matter � 2010 Elsevier Masson SAS. All rights reserved.
doi:10.1016/j.resmic.2010.07.005
C. carrionii, a chromoblastomycosis agent, and one strain ofC. resinae, suggested that the cell walls were composedmainly of hexoses as a (1/3)-linked b-glucan, with somegalactose and mannose residues, and melanin mostly presenton cytoplasmic melanosomes (San-Blas et al., 1996). Thepresence of melanin in this fungus has been related to viru-lence (Dixon et al., 1992), resistance to antifungal agents(Corrado et al., 1982) and survival in hostile environments(Potgieter and Alexander, 1966). The cell-wall compositionwas very similar in both strains (pathogenic and saprophytic),suggesting that in the process of infecting humans, the cellwall undergoes significant structural modifications. We nowreport a detailed investigation of the major glycoconjugatespresent on the C. resinae cell surface, carrying out gas-liquidchromatography (GCeMS), methylation analysis, 1H and 13CNMR spectroscopy, and ELISA and FACS techniques. Their

721R. Calixto et al. / Research in Microbiology 161 (2010) 720e728
structures were compared with other molecules found in thepathogenic fungi Aspergillus fumigatus and Candida para-psilosis that synthesize antigenic galactomannans andmannans (Barreto-Bergter et al., 1981; Shibata et al., 1995).
2. Materials and methods
2.1. Strain and culture conditions
Cladosporium resinae (Hormoconis resinae) ATCC 22712was maintained on slants of potato dextrose agar (PDA). Cellswere inoculated in Erlenmeyer flasks containing potatodextrose broth, which was incubated for 7 days at 25 �C withshaking. The mycelium was obtained via paper filtration,washed with distilled water, and stored at �20 �C.
2.2. Extraction of C. resinae mycelium and fractionationof glycoprotein with Cetavlon
The crude glycoprotein (2.3 g) was extracted frommycelium(134 g dry weight) with 0.05 M phosphate buffer, pH 7.2, at100 �C for 2 h, the filtrate was dialyzed, evaporated to a smallvolume, and then fractionated by Cetavlon precipitationaccording to Lloyd (1970). The mother liquor from the firstCetavlon precipitation was adjusted to pH 8.8 with 1% sodiumtetraborate hydrate and the resulting precipitate recovered bycentrifugation to give a major fraction (Fraction B). This, afteruncoupling Cetavlon from the complex with acetic acid, wasdialyzed and freeze-dried. It was further purified on a Superdex200 column (300 � 10 mm diameter) eluted with a continuousgradient of 0.15 M NaCl in 0.01 M sodium phosphate buffer.Eluted fractions were monitored by A280 for protein andcolorimetrically (A490) for carbohydrate (Dubois et al., 1956).Fractions containing the pGM were pooled, dialyzed againstdistilled water and lyophilized.
2.3. Extraction and fractionation of mannoproteins fromC. parapsilosis
Extraction of crude mannoproteins from C. parapsilosiswas carried out by suspending yeast cells in 0.05 M phosphatebuffer, pH 7.2, for further incubation at 100 �C for 2 h. Thesuspension was filtered to remove cells and the filtrate wasextensively dialyzed against distilled water. Mannoproteinswere finally precipitated by reaction with Fehling solution,resulting in the formation of a water-insoluble Cu2þ complex.Mannoproteins (pGM) were recovered by elimination of Cu2þ
by treatment of the precipitates with the Amberlite IR-120 (Hþ
form) resin, followed by neutralization, dialysis againstdistilled water and freeze-drying.
2.4. Isolation and purification ofpeptidogalactomannans from Aspergillus fumigatus
Peptidogalactomannans were isolated from A. fumigatusmycelium according to Haido et al. (1998).
2.5. Analytical procedures
Neutral sugars were determined by the phenol/sulfuric acidmethod (Dubois et al., 1956) and protein by the Lowry method(Lowry et al. 1951).
2.6. Monosaccharide analysis
pGM was hydrolyzed in 3 M trifluoroacetic acid at 100 �Cfor 3 h. The resulting monosaccharides were converted to theiralditol acetates which were examined by GCeMS usinga capillary column of DB-225 (30 m � 0.25 mm i.d.), operatedat 50 �C (1 min), then stepwise at 40 �C min�1 to 230 �C, thenon hold. Helium was the carrier gas. The alditol acetates wereidentified by their typical retention times and electron impactspectra (Sassaki et al., 2005a).
2.7. Amino acid acetate methyl esters (aaAME) analysis
pGM was hydrolyzed with aqueous methanolic solution(200 ml, 1.5:8.5, v/v) containing 0.6 M HCl and 0.1% ofphenol and heated at 100 �C for 15 min, followed by evapo-ration under a N2 stream. The resulting methyl-amino esterswere then acetylated with pyridineeMeOHeAc2O (300 ml;1:1:4, v/v) at 100 �C for 60 min, giving rise to aaAMEs. In thisparticular case, the samples could not be cleaned or evapo-rated, being directly injected into the GCeMS to avoid aza-lactone formation as well as decomposition of the derivative.GCeMS analyses were performed using a SATURN 2000Rgas chromatograph equipped with a 30 m � 0.25 mm lowbleed/MS capillary columns. The temperature program foraaAME separation on CP-Sil-43CB was: injector 250 �C, ovenstart at 100e210 �C (20 �C min�1, then hold for 1 min),225 �C (10 �C min�1, then on hold for 75 min) and to 230 �C(10 �C min�1, then on hold for 5 min). Post-run analyses wereperformed after optimized separation, with the chromatogramsthen being analyzed using a 5.1 Saturn Workstation (Sassakiet al., 2008).
2.8. Methylation analysis
pGM was per-O-methylated using a procedure of Tischeret al. (2002), a modification of that of Ciucanu and Kerek(1984). The product was converted to the partially O-meth-ylated alditol acetates by successive acid hydrolysis, NaBD4
reduction, and acetylation, and was analyzed by GCeMSusing a capillary column of DB-225 (30 m � 0.25 mm i.d.)under the same conditions as those for preparation of alditolacetates, except that the constant temperature was 185 �C. Theproducts were identified by their retention times and typicalelectron impact spectra, by comparison with standard mixtures(Ciucanu and Kerek, 1984).
2.9. ESI-MS
Positive ESI-MS was carried out at atmospheric pressureionization (API) with a tandem triple quadrupole Quattro LC

722 R. Calixto et al. / Research in Microbiology 161 (2010) 720e728
spectrometer (Waters), using nitrogen as nebulizer and des-olvation gas. The sample (e500 mg ml�1) was prepared in LiCl(5 mM) and directly injected into its ESI source with a syringepump (KDScientific) at a constant flow rate of 10 ml min�1.The samples were examined in the total ion current (TIC)mode using the energy setup: cone 70 V and the capillary3.1 kV. Tandem-MS was performed by collision-induceddissociation mass spectrometry (CID-MS) using helium ascollision gas. The energies used in CID-MS ranged from 56 to60 V in positive ion detection. Energies used in CID-MSranged from 56 to 60 V in positive ion detection.
2.10. NMR spectroscopy
NMR spectra were obtained with a Bruker 400 MHzAVANCE III NMR spectrometer with a 5 mm inverse Zgradient probe. Signal assignments in the 1D 1H-(zgpr) and13C-NMR (zgpg) from the pGM were obtained after dissolu-tion in D2O and the chemical shifts of signals are expressed ind ppm, relative to an external standard of acetone (d 30.2 andd 2.224 for 13C and 1H signals, respectively). 2D NMRexperiments were carried out using HSQC (hsqcetgpsi).Coupled HSQC (pl 12 CPD decoupler pulse ¼ 120 dB) wasused to determine JCeH and the anomeric configuration.COSY (cosygpqf), TOCSY (mlevphpr.2), and HMBC(hmbcgplpndqf) were carried out according to the Brukermanual. The 2D experiments were recorded for quadraturedetection in the indirect dimension, COSY, TOCSY, andHSQC spectra were acquired using 16 scans per series of2 K � 256 data points. The HMBC spectrum was acquiredusing 32 scans with zero filling in F1 (4 K) prior to Fouriertransformation (Sassaki et al., 2001, 2005b).
2.11. Partial hydrolysis of pGM
C. resinae pGM (2 mg) was treated with 0.1 M HCl andheated at 100 �C for 20 min. The degraded glycoprotein wasrecovered on dialysis and freeze-drying of the retainedsolution.
2.12. Periodate oxidation of pGM
C. resinae pGM (4 mg) was treated with increasingconcentrations (10e100 mM) of sodium m-periodate accord-ing to Haido et al., (1998).
2.13. Trypsin treatment of pGM
C. resinae pGM (1 mg) was reduced by addition of 40 ml of2.5 mM dithiothreitol (DTT) at 56 �C for 45 min. Digestionwith trypsin was performed in 100 mM Tris/HCl buffer pH 8.5using a ratio of 1:20 enzyme/pGM at 37 �C for 24 h. Digestionwas stopped by boiling the treated sample.
2.14. b-Elimination of pGM
C. resinae pGM (5 mg) was chemically de-O-glycosylatedby mild reductive alkaline treatment under reducing conditions(Leitao et al. 2003). The liberated oligosaccharide alditolfraction was recovered as eluted material on dialysis, freeze-dried and examined by TLC (Leitao et al., 2003), ESI-MS, andMSeMS.
2.15. Rabbit immune sera
White male rabbits were inoculated with freeze-driedmycelium of C. resinae (2 mg ml�1 dry weight) emulsified inan equal volume of complete Freund’s adjuvant and 1 ml ofemulsion was injected intradermally at weekly intervals for 3weeks (Haido et al., 1998). Then, throughout a one-weekperiod, the same concentration was used in three intravenousinjections at 2-day intervals. The hyperimmune serumobtained was used in ELISA experiments. Pre-immune serumwas taken as a control.
2.16. Flow cytometry
C. resinae conidia (1 � 106 cells) were harvested bycentrifugation at 7000 � g for 3 min, washed twice in phos-phate-buffered saline (PBS), pH 7.2, and incubated with anti-whole cell (1:400) antibody of C. resinae for 1 h at 37 �C. Theconidia were washed three times in PBS, pH 7.2 and incubatedwith fluorescein isothiocyanate (FITC)-labelled anti-rabbitIgG (1:100 dilution) for 1 h at 37 �C. Cells were then washedin PBS, pH 7.2, containing 0.1% sodium azide (FACs buffer),resuspended in the same buffer and analyzed using a FACScaninstrument (Becton Dickinson). On the basis of forward andside light scattering, individual conidia were gated in and cellclusters gated out. Positivity for each probe was defined asexceeding fluorescence, in relation to fluorescence of thesingle-control cells. For flow cytometric analysis, data from10 000 cells were analyzed using “Windows Multiple Docu-ment Interface Flow Cytometry Application (WinMDI) 2.8version software”. Conidia that were incubated only in thepresence of the secondary antibody were used as negativecontrols.
2.17. SDS-PAGE
Samples (C. resinae total extract, pGM) were solubilized at100 �C for 5 min in disrupting buffer containing 0.06 MTriseHCl, pH 6.8, 2% (w/v) SDS, 25% (v/v) glycerol and0.1% (w/v) bromophenol blue, with reducing conditions beingobtained by adding 5% (w/v) 2-mercaptoethanol immediatelybefore use (Laemmli, 1970) . Electrophoresis was performedin a 10% polyacrylamide separating system and the gelsstained for carbohydrate with the periodic acid-Schiff (PAS)reagent and for protein with Coomassie Blue.

723R. Calixto et al. / Research in Microbiology 161 (2010) 720e728
2.18. Western blot analysis
Following electrophoresis, samples were electrotransferredonto PVDF membranes, as previously described (Bahia et al.,1997) . After transfer, these were blocked with 3% BSA inPBS (10 mM phosphate buffer/0.15 M NaCl) at roomtemperature and incubated for 1 h at 37 �C with hyperimmuneserum against whole cells of C. resinae diluted 1:100. Themembranes were washed 3 � with PBS Tween, incubated withgoat anti-rabbit IgG (1:1000) in the same buffer, washed asabove, and developed with diaminobenzidine (DAB) in 0.1 MPBS buffer and H2O2.
2.19. ELISA
Solutions of 5 mg ml�1 of C. resinae pGM, intact and afterchemical treatment (partial hydrolysis, periodate oxidation,and b-elimination) were added onto wells of flat-bottomedpolyvinyl microtitre plates (Falcon-Becton & Dickinson,Franklin Lakes, NJ), which were maintained for 1 h at 37 �Cand overnight at 4 �C. After washing the plates with 0.05%phosphate buffered saline-Tween 20, the non-specific siteswere blocked by addition of 5% skimmed milk in 0.1%phosphate buffered saline-Tween 20. Rabbit immune seraagainst C. resinae (1/400) in blocking buffer (100 ml) wereadded to the wells and antibody binding was measured usinggoat anti-rabbit IgG antibody conjugated to horseradishperoxidase (Sigma) at 1/3000 (100 ml). Each step was fol-lowed by 1 h/37 �C incubation and washings with 0.1%phosphate buffered saline-Tween 20. The substrate used was0.4 mg mL�1 o-phenylenediamine and 0.4 mL mL�1 H2O2 in0.01 M sodium citrate buffer, pH 5.0. The enzyme reactionwas followed by measuring absorbance at 490 nm using anautomated microplate reader (BioRad ELISA Reader) 20 minafter terminating the reaction with 50 ml 1.5 M H2SO4. Inorder to verify antigenic similarities between the pGM of C.resinae, that of A. fumigatus, and the mannoprotein of C.parapsilosis, the cross-reactivity of these glycoproteins (5 mg/mL) with rabbit immune serum against C. resinae (1/400) wastested by ELISA, as described above.
2.20. Hapten inhibition tests
Solutions containing the oligosaccharide mixture, rangingfrom 0.5 to 500 ng/well, were separately added to the samevolume of rabbit immune serum against C. resinae (1/400) andpreincubated for 1 h at 37 �C. The immunoreactivity of themixture of reduced O-linked oligosaccharidic chains wasevaluated by ELISA, as described above.
2.21. Statistical analysis
Fig. 1. Elution profile of pGM from C. resinae on a Superdex 200 column
(10 � 300 mm). Fractions were monitored for protein (A280) (-) and
carbohydrates (A490) (o).
Statistical analyses were performed using GraphPad Prismversion 5.00 for Windows (GraphPad Software, San DiegoCA). One-way analysis of variance was used to compare thedifferences between groups, and individual comparisons of
groups were done using a Bonferroni posttest. A 90e95%confidence interval was determined in all experiments.
3. Results
3.1. Isolation and purification of C. resinaepeptidopolysaccharide (pGM)
The crude glycopeptides (53% carbohydrate; 31% protein)obtained from C. resinae mycelium by hot buffer extractionwere purified by selective precipitation with Cetavlon/sodiumtetraborate at pH 8.8 as previously described (Haido et al.1998; Pinto et al., 2001). The Cetavlon-precipitated material(Fraction B) was further fractionated by Superdex 200 columnchromatography giving rise to a homogeneous pGM fraction(Fig. 1). Further analysis was carried out on this fraction,which contained 77% carbohydrate and 23% protein.
pGM, on electrophoresis with SDS-PAGE, showed a bandwith an apparent molecular mass ranging from 45 to 42 kDawhen stained for protein and carbohydrate (Fig. 2, lanes 5 and6) and recognized by polyclonal rabbit antiserum against C.resinae mycelium (Fig. 2, lane 4). Trypsin treatment causedthe molecular mass of the pGM to decrease, confirming thepresence of the protein moiety in the pGM (Fig. 2, lane 7).
3.2. Monosaccharide composition of pGM
Acid hydrolysis of pGM, followed by successive sodiumborohydride reduction and acetylation, gave alditol acetates ofrhamnitol (Rha), ribitol (Rib), mannitol (Man), galactitol (Gal)and glucitol (Glc) at a molar ratio of 4:2.5:33:30:3 (GCeMS).
3.3. Amino acid composition of pGM
Results of analysis of the amino acid composition of pGMare shown in Table 1. The protein contained relatively highamounts of serine, threonine and isoleucine/leucine. Proline andphenylalanine were also present in relatively high quantities,

Table 2
Molar percentages of O-methyl alditol acetates obtained on methylation
analysis of peptidopolysaccharide from C. resinae.
O-Methylalditol
acetateaRetention
time (min)
Glycosidic
linkage
Mole %
2,3,4,6-Me4-Man 8.70 Manp-(1/ 6
2,3,5,6-Me4-Gal 9.02 Galf-(1/ 6
3,4,6-Me3-Man 11.10 /2)-Manp-(1/ 18
2,3,6-Me3-Gal 12.05 /5)-Galf-(1/ 41
2,3,4-Me3-Man 12.18 /6)-Manp-(1/ 12
2,3,6-Me3-Glc 12.28 /4)-Glcp-(1/ 13
2,3,5-Me3-Gal 13.45 /6)-Galf-(1/ tr.
3,4-Me2-Man 17.44 /2,6)-Manp-(1/ 11
2,4-Me2-Man 17.69 /3,6)-Manp-(1/ 2
2,3-Me2-Glc 18.09 /4,6)-Glcp-(1/ 1
a O-Methylalditol acetates obtained on successive methylation, hydrolysis,
reduction, and acetylation, analyzed by GCeMS (DB-225 column).
Fig. 2. SDS-PAGE gels and corresponding immunoblots of C. resinae total cell
extract, untreated and trypsin-degraded pGM. The figure shows differently
stained SDS-PAGE gels (lanes 1, 2, 5, 6 and 7) and corresponding immuno-
blots (lanes 3 and 4). Lane 1: Coomassie stained molecular weight marker;
lane 2: Coomassie-stained total cell extract; lane 3: immunoblot of lane 2
reacted with polyclonal rabbit serum against mycelium of C.resinae; lane 4:
immunoblot of purified pGM reacted with polyclonal rabbit serum against
mycelium of C.resinae; lane 5: purified pGM stained for sugars; lane 6: silver
stain of purified pGM; lane 7: silver stain of purified pGM treated with trypsin.
The arrow indicates undegraded pGM, ** indicates GM portion, * indicates
trypsin.
724 R. Calixto et al. / Research in Microbiology 161 (2010) 720e728
whereas alanine, glutamic acid, and glycine were present in lowquantities.
3.4. Methylation analysis of pGM
Methylation analysis (Table 2) of pGM showed it to havea branched structure giving rise to partially O-methylatedalditol acetates (GCeMS), corresponding to non-reducingend- (6%), 5-O- (41%) and 6-O-subst. Galf (tr.), non-reducingend-(6%), 2-O- (18%), 6-O-(12%), 2,6-di-O-(11%) and 3,6-di-O-subst. Manp (2%), and 4-O- (3%) and 4,6-di-O-subst. unitsof Glcp (1%).
e
Table 1
Retention times and EI-MS reporter fragments obtained by mass spectrometry
from the aaAME derivatives.
aaMe Rt % Reporter ions (m/z)
Ala 5.516 3.7 146e114e102e86
Gly 5.825 4.4 132e114e100e99e89
Val 5.878 tr 174e142e114e72Ile/Leu 6.255 29.3 188e156e128e99e86
Thr 7.448 14.2 218e158e131e113e98e89
Pro 7.883 11.5 172e140e128e112e70
Ser 8.105 25.4 204e144e102e84Glu 10.651 1.2 218e158e116e98e84
Phe 12.651 10.2 222e162e131e120e103e91e88
Tyr 44.547 tr 280e248e238e220e178e147e107e91e88Lys 65.608 tr 245e213e201e186e170e142e126e114e84
GCeMS analysis of aaAME on CP-Sil-43CB capillary column. tr traces
(relative quantities below 0.1%).
3.5. NMR spectroscopy of pGM
The 13C NMR spectrum of pGMwas complex and containedseveral signals in the C-1 region (Fig. 3A insert). Depending onthe shifts in a related galactofuranomannopyranan witha (1/6)-linked a-Manp main chain, obtained from myceliumof A. fumigatus (Barreto-Bergter et al. 1981), the three largestb-Galf signals were from consecutive exterior (1/5)-linkedside-chain units and were at d 108.24, 108.40 and at 107.70. Thelast signal would be from b-Galf units linked (1/2) to themannan core. This is based on our methylation data and thevalue of d 107.7 for the shielded C-1 of b-D-galactofuranosyl-(1/2)-methyl a-D-mannopyranoside (Gorin and Barreto-Bergter, 1981) . Other prominent signals may be assigned to5-O-substituted b-D-Galf units, such as those of C-2 and C-4(d 82.2 and 83.1), C-3 (d 77.6) and C-5 (d 76.7) (Gimenes-Abianet al. 2007). The other principal signal was at d 100.69 from non-substituted units of a (1/6)-linked a-mannopyranan mainchain. Smaller signals were present at d 103.60 and 102.2 fromrespective terminal and internal a-Manp-(1/2)-(a-Manp)nside-chain groups linked to the mannan core, and that at d 98.97should arise from 2,6-di-O-substituted units in the main chain.On examination of its C-6 signals, the signal at d 66.9, whencompared with the unsubstituted resonance of the a-shift of5.4 ppm found for the mannan of baker’s yeast and which arisesfrom Manp-(1/6)-a-Manp groups, and the signal at d 63.3(C-60), should have arisen from b-Galf-(1/2)-a-Manp groups,since methyl b-D-Galf- (1/2)-a-D-Manp has signals at d 63.8(C-60) and 61.9 (C-6), respectively (Gorin and Barreto-Bergter,1981). An additional C-6 signal at d 61.0 was detected in theNMRspectrum and corresponds to units ofa-hexosaminewhichshould have a C-6 resonance at higher field than that ofa-mannopyranose (Bock and Peterson, 1983; Tischer et al.,2002). The anomeric configuration was obtained by coupledHSQC. All JCeH values from the mannopyranosyl units weretypical of a configurationvarying from 169 to 171Hz, consistentwith Perlin and Casu (1969), who reported 169 Hz for a-Glcpand 160Hz forb-Glcpmoieties. The only possible conformationand configuration of the galactosyl units from peptidogalacto-mannan gave rise to characteristic hemiketalic b C-1 signals of
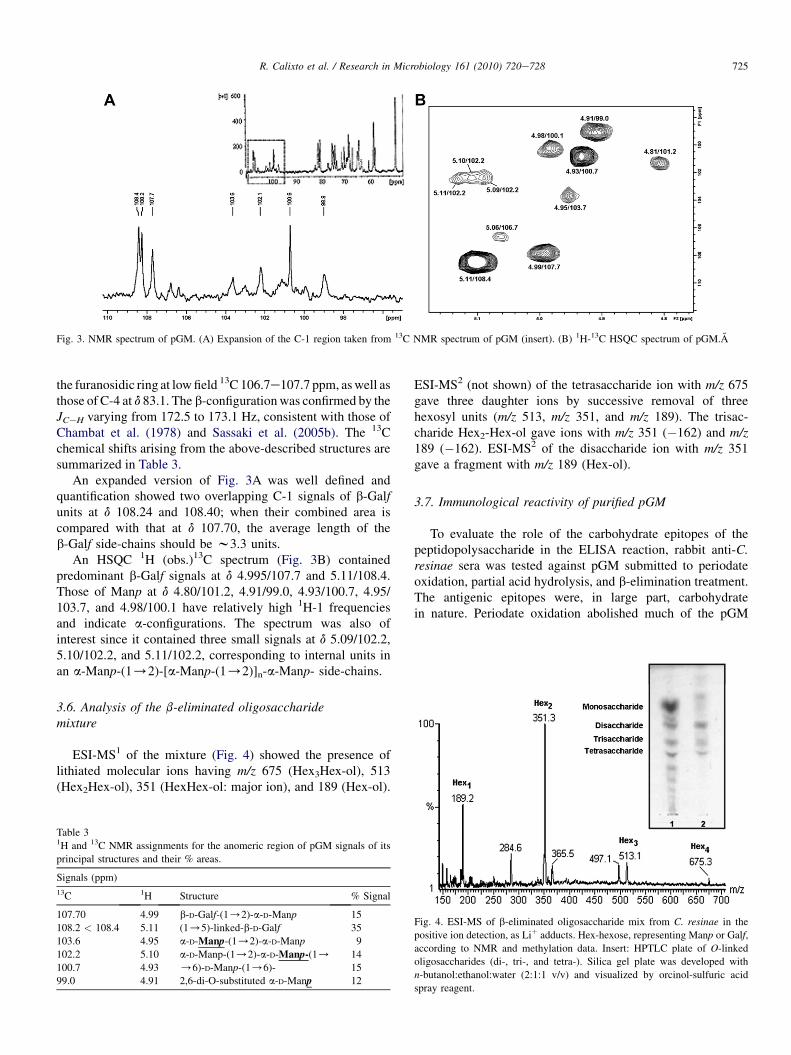
Fig. 3. NMR spectrum of pGM. (A) Expansion of the C-1 region taken from 13C NMR spectrum of pGM (insert). (B) 1H-13C HSQC spectrum of pGM.A
725R. Calixto et al. / Research in Microbiology 161 (2010) 720e728
the furanosidic ring at lowfield 13C 106.7e107.7 ppm, aswell asthose of C-4 at d 83.1. The b-configurationwas confirmed by theJCeH varying from 172.5 to 173.1 Hz, consistent with those ofChambat et al. (1978) and Sassaki et al. (2005b). The 13Cchemical shifts arising from the above-described structures aresummarized in Table 3.
An expanded version of Fig. 3A was well defined andquantification showed two overlapping C-1 signals of b-Galfunits at d 108.24 and 108.40; when their combined area iscompared with that at d 107.70, the average length of theb-Galf side-chains should be w3.3 units.
An HSQC 1H (obs.)13C spectrum (Fig. 3B) containedpredominant b-Galf signals at d 4.995/107.7 and 5.11/108.4.Those of Manp at d 4.80/101.2, 4.91/99.0, 4.93/100.7, 4.95/103.7, and 4.98/100.1 have relatively high 1H-1 frequenciesand indicate a-configurations. The spectrum was also ofinterest since it contained three small signals at d 5.09/102.2,5.10/102.2, and 5.11/102.2, corresponding to internal units inan a-Manp-(1/2)-[a-Manp-(1/2)]n-a-Manp- side-chains.
3.6. Analysis of the b-eliminated oligosaccharidemixture
ESI-MS1 of the mixture (Fig. 4) showed the presence oflithiated molecular ions having m/z 675 (Hex3Hex-ol), 513(Hex2Hex-ol), 351 (HexHex-ol: major ion), and 189 (Hex-ol).
Table 31H and 13C NMR assignments for the anomeric region of pGM signals of its
principal structures and their % areas.
Signals (ppm)
13C 1H Structure % Signal
107.70 4.99 b-D-Galf-(1/2)-a-D-Manp 15
108.2 < 108.4 5.11 (1/5)-linked-b-D-Galf 35
103.6 4.95 a-D-Manp-(1/2)-a-D-Manp 9
102.2 5.10 a-D-Manp-(1/2)-a-D-Manp-(1/ 14
100.7 4.93 /6)-D-Manp-(1/6)- 15
99.0 4.91 2,6-di-O-substituted a-D-Manp 12
ESI-MS2 (not shown) of the tetrasaccharide ion with m/z 675gave three daughter ions by successive removal of threehexosyl units (m/z 513, m/z 351, and m/z 189). The trisac-charide Hex2-Hex-ol gave ions with m/z 351 (�162) and m/z189 (�162). ESI-MS2 of the disaccharide ion with m/z 351gave a fragment with m/z 189 (Hex-ol).
3.7. Immunological reactivity of purified pGM
To evaluate the role of the carbohydrate epitopes of thepeptidopolysaccharide in the ELISA reaction, rabbit anti-C.resinae sera was tested against pGM submitted to periodateoxidation, partial acid hydrolysis, and b-elimination treatment.The antigenic epitopes were, in large part, carbohydratein nature. Periodate oxidation abolished much of the pGM
Fig. 4. ESI-MS of b-eliminated oligosaccharide mix from C. resinae in the
positive ion detection, as Liþ adducts. Hex-hexose, representing Manp or Galf,
according to NMR and methylation data. Insert: HPTLC plate of O-linked
oligosaccharides (di-, tri-, and tetra-). Silica gel plate was developed with
n-butanol:ethanol:water (2:1:1 v/v) and visualized by orcinol-sulfuric acid
spray reagent.

726 R. Calixto et al. / Research in Microbiology 161 (2010) 720e728
antigenic activity (Fig. 5A). Treatment with dilute acid, whichremoved labile galactofuranosyl side-chain residues,decreased the antigenicity of this molecule (Fig. 5A). A strongdecrease in reactivity was also observed with de-O-glycosy-lated pGM (lacking O-linked oligosaccharide chains)(Fig. 5A).
To evaluate the immunodominance of the O-linked oligo-saccharides, they were tested in an ELISA hapten system asinhibitors of the reactivity between pGM and hyperimmunerabbit antisera against whole cells of C. resinae. Inhibitions ashigh as 40% were obtained (Fig. 5B).
3.8. Antigenic cross-reactivity between C. resinae pGMand other fungal glycoproteins
Galactofuranose- and mannose-containing polysaccharidesare present in many fungi. We therefore tested antigenic simi-larity of the galactomannan of C. resinae to other gal-actomannan and mannan using rabbit antisera against wholecells of C. resinae. It was found that the reactivity of C. resinaerabbit antiserawithC. resinae pGMwas similar to that observedwith A. fumigatus pGM, as well as mannoprotein from C. par-apsilosis (Fig. 6A). By FACS, a decrease in rabbit sera anti-C. resinae binding to C. resinae conidia (e60%) could bedemonstrated when this sera was previously incubated withpeptidogalatomannans fromC. resinae andA. fumigatus, orwithmannoprotein from C. parapsilosis, suggesting the presence of
Fig. 5. Antigenicity of the pGM fraction of C. resinae. (A) ELISA of
untreated, de-O-glycosylated, partially hydrolyzed and periodate-treated pGM
probed with rabbit anti-C. resinae serum. *p < 0.001. (B) Inhibition of pGM
reactivity to rabbit anti-C. resinae serum (1:400) by O-linked oligosaccharide
fraction from pGM (hapten).
Fig. 6. Cross-reactivity of the rabbit anti-resinae serum with glycoproteins of
C. resinae A. fumigatus and C. parapsilosis. (A) Reactivity of rabbit anti-C.
resinae serum with pGM from C. resinae, pGM from A. fumigatus and
mannoprotein from C. parapsilosis. All glycoproteins were used at 5 mg/mL.
(B) FACS analysis showing inhibition of C. resinae conidia reactivity to rabbit
anti-C. resinae serum (1:400) by pGM from C. resinae, pGM from A. fumi-
gatus and mannoprotein from C. parapsilosis. Positive controls indicate the
reactivity of each glycoprotein with sera anti-C. resinae. An irrelevant
immunoglobulin was used in negative controls.*p < 0.001.
cross-reactive determinants in these fungi (Fig. 6B). Since theimmunodominant epitopes present in pGM of A.fumigatuscontain a b-Galf-(1/5)-b-Galf side chain, it is possible thatsuch structures are also present in C. resinae. The C. resinaeantisera is also reactive for branchingmoieties ofC. parapsilosismannoprotein, which contained side chains of (1/2)-linked a-D-mannopyranose residues. The presence of side chains of(1/2)-linked a-D-Manp units in the C. resinae galactomannanwas confirmed by methylation and NMR analysis (Table 2,Fig. 3B).
4. Discussion
Glycoconjugates isolated from fungi comprise a wide rangeof components with different structures and immunologicalreactivity. These can be associated with the composition ofculture media as well as temperature of growth (Calera et al.,1994; Bahia et al., 1997). We now report a partial structuralcharacterization of a pGM from a saprophytic strain of C.resinae, which was performed in order to correlate its structure

727R. Calixto et al. / Research in Microbiology 161 (2010) 720e728
with a pGM from mycelium of a pathogenic strain of A.fumigatus, previously studied by our group (Haido et al., 1998;Leitao et al., 2003).
The pGM from C. resinae consists of a main chain con-taining (1/6)-linked a-D-mannopyranosyl residuessubstituted at O-2 by side chains containing consecutive(1/2)-linked a-D-mannopyranose residues. b-D-galactofur-anosyl units are present as side chains of w3.3 units that are(1/5)-interlinked. Methylation analysis showed the presenceof non-reducing end- and 5-O-substituted galactofuranosylunits (Table 2), although the formation of 2,3,6Me3galactitolacetate is consistent with 4-O-linked galactopyranosyl and5-O-linked galactofuranosyl units. However, NMR resultsshow only the latter to be present. Such a complex D-galacto-D-mannan is very similar to a pGM isolated from A. fumigatusAF 2109 (Barreto-Bergter et al., 1981). It differs from thegalactomannans (pGM) isolated from the dimorphic fungusCladosporium werneckii, the causative agent of tinea nigrahaving single terminal b-galactofuranosyl-(1/6)-units(Lloyd, 1970) and from (1/6)-linked linear b-galactofur-anosyl polymers linked to a small amount of mannan, isolatedfrom Malassezia furfur and Malassezia pachydermatis(Shibata et al., 1995). The galactofuranosyl unit is essential forsurvival or virulence of some pathogenic microorganisms andis a dominant immunogen (Bennet et al. 1985). In fungi,galactofuranosyl units have been found in polymers fromAspergillus, Penicillium, Histoplasma, Paracoccidioides, Tri-chophyton and Fonsecae species (Shibata et al. 2009; Prietoet al. 2004).
The mannan moiety of the C. resinae galactomannan con-sisted of a main chain of (1/6)-linked a-D-mannopyranoseresidues substituted at HO-2 by a-D-mannopyranosyl units thatare (1/2)-interlinked, similar to that of several species ofAspergillus (Barreto-Bergter et al., 1980) and the mannan ofC. parapsilosis (Shibata et al., 1995).
The importance of the carbohydrate moiety of C. resinaepGM in immunoassays has also been demonstrated in thiswork. Removal of side chains of (1/5)-linked b-D-gal-actofuranosyl units, by partial acid hydrolysis with 0.1 M HClat 100 �C decreased the reactivity with C. resinae serum.Previous reports demonstrated that the carbohydrate portion ofpGM molecules contained b-Galf-bearing chains, regarded tobe the immunodominant epitopes (Bennet et al., 1985), espe-cially when they are (1/5)-linked (Nothermans et al., 1988).More recent results from our group showed that the immu-nodominant epitopes in A. fumigatus pGM were present in thetetra- and hexasaccharides, which contain a b-Galf-(1/5)-b-Galf- terminal groups (Leitao et al. 2003).
De-O-glycosylation (removal of O-linked oligosaccharideschains) of pGM by reductive alkaline b-elimination gave riseto a w70% decrease in its antigenic reactivity, which waspractically eliminated by m-periodate treatment. Antibodiesrecognizing Galf-containing structures may therefore recog-nize both N-linked high molecular weight polysaccharides (de-O-pGM) and the O-linked oligosaccharides in the pGMmolecules. To verify this, immunodominance of the O-linkedoligosaccharide chains present in the C. resinae pGM was
evaluated in an ELISA hapten system, as inhibitors of reac-tivity between the pGM and the C. resinae sera. Inhibition ashigh as 40% was obtained, suggesting that O-linked oligo-saccharides linked to pGM are important determinants ofreactivity, as already described in peptidopolysaccharides fromA. fumigatus (Leitao et al., 2003), Pseudallescheria boydii(Pinto et al., 2001) and Sporothrix schenckii (Bernardes-Engelmann et al., 2009). Our results show that the antige-nicity of the intact pGM could be partly associated withO-glycans, in addition to galactomannan (supposedly N-linkedglycan). The peptidogalactomannan (pGM) from A. fumigatusand the peptidorhamnomann (PRM) from P. boydii, thatcontain both N- and O-glycans, have the same behavior whentested against either anti- A.fumigatus or anti-P. boydii rabbitantiserum, respectively (Leitao et al., 2003; Pinto et al., 2001).
Cross-reactivity between pGM from C. resinae and mannanfrom C. parapsilosis was also observed by flow cytometry andsuggests that another significant structural feature of the pGMis a high amount of a-D-mannopyranose residues substituted atHO-2 by a-D-mannopyranosyl units that are (1/2)-inter-linked, as present in C. resinae pGM and C. parapsilosismannan (Shibata et al., 1995).
Acknowledgements
We thank Maria de Fatima F. Soares for technical assis-tance and Profs. P.A.J. Gorin and Walter Oelemann for advice,encouragement and critical reading of the manuscript. Thiswork was supported by grants from the Conselho Nacional deDesenvolvimento Cientifico e Tecnologico (CNPq (CNPq)),Fundacao de Amparo a Pesquisa do Estado do Riode Janeiro (FAPERJ (FAPERJ)), Programa de Nucleos deExcelencia (PRONEX (PRONEX)) and Coordenacao deAperfeicoamento de Pessoal de Ensino Superior (CAPES(CAPES)).
References
Bahia,M.C.F.R.,Vieira,R.P.,Mulloy,B.,Hartmann,R., Barreto-Bergter, E., 1997.
The structures of polysaccharides and glycolipids of Aspergillus fumigatus
grown in the presence of human serum. Mycopathologia 137, 17e25.
Barreto-Bergter, E., Gorin, P.A.J., Travassos, L.R., 1981. Cell constituents of
mycelia and conidia of Aspergillus fumigatus. Carbohydr. Res. 95,
205e218.
Barreto-Bergter, E., Travassos, L.R., Gorin, P.A.J., 1980. Chemical structure of
the D-galacto-D-mannan component from hyphae of Aspergillus niger and
other Aspergillus spp. Carbohydr. Res. 86, 273e285.Bennet, J.E., Bhattacharjee, A.K., Glaudemans, C.P.J., 1985. Galactofuranosyl
groups are immunodominant in Aspergillus fumigatus galactomannan. Mol.
Immunol. 23, 251e254.
Bernardes-Engelmann,A.R., Loureiro y Penha, C.V., Benvenuto, F., Braga, J.U.,
Barros, M.L., Orofino-Costa, R., Lopes-Bezerra, L., 2009. A comparative
serological study of the SsCBF antigenic fraction isolated from three
Sporothrix schenckii strains. Med. Mycol. 47, 874e878.
Bock, K., Peterson, C., 1983. Carbon-13 nuclear magnetic resonance spectros-
copy of monosaccharides. Adv. Carbohydr. Chem. Biochem. 41, 27e66.
Calera, J.A., Lopez Medrano, R., Overejo, M.C., Perente, P., Leal, F., 1994.
Variability of Aspergillus nidulans antigens with media and time and
temperature of growth. Infect. Immun. 62, 2322e2333.

728 R. Calixto et al. / Research in Microbiology 161 (2010) 720e728
Chambat, G., Joseleau, J.-P., Lapeyre, M., Lefebvre, A., 1978. Presence of
D-galactofuranose in the capsular polysaccharide of Klebsiella serotype
K-41: synthesis of 5,6-di- O-methyl-D-galactofuranose. Carbohydr. Res.
63, 323e326.
Ciucanu, J., Kerek, F., 1984. A simple and rapid method for the permethylation
of carbohydrates. Carbohydr. Res. 131, 209e217.
Corrado, M.L., Kramer, M., Cummings, M., Eng, R.H., 1982. Susceptibility of
dematiaceous fungi to amphotericin B, miconazole, ketoconazole, flu-
cyosine and rifampin alone and in combination. Sabouradia 20, 109e113.
Dixon, D.M., Migliozzi, J., Cooper Jr., C.R., Solis, O., Breslin, B.,
Szaniszlo, P.J., 1992. Melanized and non-melanized multicellular form
mutants of Wangiella derantitidis in mice: mortality and histopathology
studies. Mycoses 35, 17e21.
Dubois, M., Gilles, K.A., Hamilton, J.K., Rebers, P.A., Smith, E., 1956.
Colorimetric method for determination of sugar and related substance.
Anal. Chem. 28, 350e356.Gimenes-Abian, M.I., Bernabe, M., Leal, J.A., Jimenez-Barbero, J., Prieto, A.,
2007. Structure of a galactomannan isolated from the cell wall of the
fungus Lineolata rhizophorae. Carbohydr. Res. 342, 2599e2603.Gorin, P.A.J., Barreto-Bergter, E.M., da Cruz, F.S., 1981. The chemical
structure of the D-galacto-D-mannan component of Trypanosoma cruzi:13C-NMR dependence on structure of D-galactose to D-mannose linkage.
Carbohydr. Res. 88, 177e188.Haido, R.M.T., Silva,M.H., Ejzemberg, R., Leitao, E.A., Hearn, V.M., Evans, E.
G.V., Barreto-Bergter, E., 1998. Analysis of peptidogalactomannans from
the mycelial surface of Aspergillus fumigatus. Med. Mycol. 36, 313e321.
Latge, J.P., Bouziane, H., Diaquin, M., 1988. Ultrastructure and composition
of the conidial wall of Cladosporium cladosporioides. Can. J. Microbiol.
34, 1325e1329.
Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of
the head of bacteriophage T4. Nature 227, 680e685.
Leitao,E.A.,Bittencourt,V.C.B.,Haido,R.M.T.,Valente,A.P., Peter-Katalinik, J.
, Letzel, J., de Souza, L.M., Barreto-Bergter, E., 2003. b-Galactofuranose-
containing O-linked oligosaccharides present in the cell wall peptidoga-
lactomannan of Aspergillus fumigatus contain immunodominant epitopes.
Glycobiology 13, 681e692.
Lindley, N.D., Heyderman, M.T., 1985. Alkane utilization by Cladosporium
resinae: the importance of extended lag phases when assessing substrate
optima. FEMS Microbiol. Ecol. 31, 307e310.
Lloyd, K.O., 1970. Isolation, characterization and partial structure of pepti-
dogalactomannan from the yeast form of Cladosporium werneckii.
Biochemistry 9, 3446e3453.Lowry, O.H., Rosebrough, N.J., Fan, A.L., Randall, R.J., 1951. Protein
measurement with the Folin phenol reagent. J. Biol. Chem. 193, 265e275.
Nothermans, S., Veeneman, G.H., van Zuylen, C.W.E., Hoogerhout, P.,
van Boon, J.H., 1988. (1/5)-Linked b-D-galactofuranosides are immu-
nodominant in extracellular polysaccharides of Penicillium and Aspergillus
species. Mol. Immunol. 25, 975e979.
Perlin, A.S., Casu, B., 1969. Carbon-13 and proton magnetic resonance spectra
of D-glucose-13C. Tetrahedron Lett., 2921e2924.
Pinto, M.R., Mulloy, B., Haido, R.M.T., Travassos, L.R., Barreto-Bergter, E.,
2001. A peptidorhamnomannan from the mycelium of Pseudallescheria
boydii is a potential diagnostic antigen of this emerging human pathogen.
Microbiology 147, 1499e1506.
Potgieter, H.J., Alexander, M., 1966. Susceptibility and resistance of several
fungi to microbial lysis. J. Bacteriol. 91, 1526e1532.
Prieto, A., Ahrazem, O., Bernabe, M., Leal, J.A., 2004. Polysaccharides F1SS:
taxonomic and evolutionary characters for ascomycetes. In: San-Blas, G.,
Calderone, R.A. (Eds.), Pathogenic Fungi: Structural Biology and
Taxonomy. Caister Academic Press, England, pp. 319e360.
San-Blas, G., Guanipa, O., Moreno, B., Pekerar, S., San-Blas, F., 1996.
Cladosporium carrionii and Hormoconis resinae (C. resinae): cell wall
and melanin studies. Curr. Microbiol. 332, 11e16.
Sassaki, G.L., Iacomini, M., Gorin, P.A.J., 2005a. Methylation-GCeMS
analysis of arabinofuranose- and galactofuranose-containing structures:
rapid synthesis of partially O-methylated alditol acetate standards. Acad.
Bras. Cienc. 77, 223e234.
Sassaki, G.L., Gorin, P.A.J., Tischer, C.A., Iacomini, M., 2001. Sulfonogly-
colipids from the lichenised basidiomycete Dictyonema glabratum: isola-
tion, NMR and ESI-MS approaches. Glycobiology 11, 345e351.
Sassaki, G.L., Gorin, P.A., Reis, R.A., Serrato, R.V., Elifio, S.L., Iacomini, M.,
2005b. Carbohydrate, glycolipid, and lipid components from the photo-
biont (Scytonema sp.) of the lichen Dictyomema glabratum. Carbohydr.
Res. 340, 1808e1817.
Sassaki,G.L., Souza, L.M., Serrato,R.V., Thales, R.,Cipriani, T.R., Gorin, P.A.J.,
Iacomini, M., 2008. Application of acetate derivatives for gas chromatogra-
phyemass spectrometry: novel approaches on carbohydrates, lipids and
amino acids analysis. J. Chromatogr. A 1208, 215e222.
Seifert, K.A., Hughes, S.J., Boulay, H., Louis-Seize, G., 2007. Taxonomy,
nomenclature and phylogeny of three cladosporium-like hyphomycetes,
Sorocybe resinae, Seifertia azaleae and the Hormoconis anamorph of
Amorphotheca resinae. Stud. Mycol. 58, 235e245.Shibata, N., Ikuta, K., Imai, T., Satoh, Y., Satoh, R., Suzuki, A., Kojima, C.,
Kobayashi, H., Hisamichi, K., Suzuki, S., 1995. Existence of branched side
chains in the cell wall mannan of pathogenic yeast, Candida albicans.
Structure-antigenicity relationship between the cell wall mannans ofCandida
albicans and Candida parapsilosis. J. Biol. Chem. 270, 1113e1122.
Shibata, N., Saitoh, T., Tadokoro, Y., Okawa, Y., 2009. The cell wall gal-
actomannan antigen fromMalassezia furfur andMalassezia pachydermatis
contains b-1,6-linked linear galactofuranosyl residues and its detection has
diagnostic potential. Microbiology 155, 3420e3429.
Tischer, C.R., Gorin, P.A.J., Iacomini, M., 2002. The free reducing oligosac-
charides of gum arabic: aids for structural assignments in the poly-
saccharide. Carbohydr. Polym. 47, 151e158.van Kan, J.A., Joosten, M.H., Wagemakers, C.A., Van Den Berg-Velthuis, G.C.,
De Wit, P.J., 1992. Differential accumulation of mRNAs encoding
extracellular and intracellular PR proteins in tomato induced by virulent
and avirulent races of Cladosporium fulvum. Plant Mol. Biol. 20, 513e527.
Vitale, R.G., Perez-Blanco, M., De Hoog, G.S., 2009. In vitro activity of
antifungal drugs against Cladophialophora species associated with human
chromoblastomycosis. Med. Mycol. 47, 35e40.Yegres, F., Richard-Yegres, N., Nishimura, K., Miyaji, M., 1991. Virulence
and pathogenicity of human and environmental isolates of Cladosporium
carrionii in newborn ddY mice. Mycopathologia 114, 71e76.
Copyright © 2022 FDOKUMEN




















