
Diagnostic assays and sampling protocols for the detection of Batrachochytrium dendrobatidis
Upload
independentCategory
view
1download
0
Experimental Exposures of Boreal Toads (Bufo boreas) to aPathogenic Chytrid Fungus (Batrachochytrium dendrobatidis)
Cynthia Carey,1 Judsen E. Bruzgul,1 Lauren J. Livo,1 Margie L. Walling,2 Kristin A. Kuehl,1
Brenner F. Dixon,1 Allan P. Pessier,3 Ross A. Alford,4 and Kevin B. Rogers5
1Department of Integrative Physiology, University of Colorado, Boulder, CO 80309-03542Department of Environmental and Radiological Health Sciences/Epidemiology, Colorado State University, Ft. Collins, CO 805213Department of Pathology, Zoological Society of San Diego, PO Box 120551, San Diego, CA 92112-05514School of Tropical Biology, James Cook University, Townsville, Queensland 4811, Australia5Colorado Division of Wildlife, PO Box 775777, Steamboat Springs, CO 80477
Abstract: One of the major causes of worldwide amphibian declines is a skin infection caused by a pathogenic
chytrid fungus (Batrachochytrium dendrobatidis). This study documents the interactions between this pathogen
and a susceptible amphibian host, the boreal toad (Bufo boreas). The amount of time following exposure until
death is influenced by the dosage of infectious zoospores, duration of exposure, and body size of the toad. The
significant relation between dosage and the number of days survived (dose-response curve) supports the
hypothesis that the degree of infection must reach a particular threshold of about 107–108 zoosporangia before
death results. Variation in air temperature between 12�C and 23�C had no significant effect on survival time.
The infection can be transmitted from infected to healthy animals by contact with water containing zoospores;
no physical contact between animals is required. These results are correlated with observations on the pop-
ulation biology of boreal toads in which mortalities associated with B. dendrobatidis have been identified.
Key words: Batrachochytrium dendrobatidis, amphibian pathogen, Bufo boreas, chytrid fungus, chytridiomy-
cosis
INTRODUCTION
Populations of many species of amphibians have experi-
enced serious declines over the last several decades (Alford
and Richards, 1999; Stuart et al., 2004). Although factors
such as habitat destruction and introduction of invasive
species have contributed to these declines, infectious dis-
ease has been identified as another significant cause (Berger
et al., 1998; Carey, 2000; Carey et al., 1999, 2003a, 2003b;
Daszak et al., 1999, 2003). A recently discovered chytrid
fungus (Batrachochytrium dendrobatidis), initially isolated
from captive amphibians suffering from a mycotic skin
disease (Longcore et al., 1999; Nichols et al., 1998; Pessier et
al., 1999), has now been linked to mass mortalities of wild
amphibian populations in many areas, including Europe,
South America, Central America, Australia, New Zealand,
and North America (Berger et al., 1998; Bishop, 2000;
Bosch et al., 2001; Bradley et al., 2002; Green et al., 2002;
Green and Kagarise Sherman, 2001; Lips et al., 2003; Muths
et al., 2003; Ron et al., 2003; Ron and Merino, 2000). For a
recent list of geographic localities and amphibian species onPublished online: January 18, 2006
Correspondence to: Cynthia Carey, e-mail: [email protected]
EcoHealth 3, 5–21, 2006DOI: 10.1007/s10393-005-0006-4
Original Contributions
� 2006 EcoHealth Journal Consortium

which this chytrid fungus has been found, see Carey et al.,
(2003a) or Speare and Berger (2000). This pathogen,
thought to have recently emerged (Daszak et al., 2003),
demonstrates little genetic diversity among isolates col-
lected in various locations around the world (Morehouse et
al., 2003). The closest chytrid relative from which this
pathogenic form evolved is currently not known (James et
al., 2000). Although the geographic origin of B. dendro-
batidis has not been proven, evidence from museum
specimens suggests it may have originated in Africa (Wel-
don et al., 2004).
A wide variety of bacterial, viral, and fungal agents are
normally found on the skin, in the digestive tract, and in
other tissues of amphibians (Granoff, 1969; Taylor, 2001;
Taylor et al., 2001). However, this chytrid fungus and a
group of ranaviruses are the only pathogens for which there
is a demonstrated correlation between the degree of
infection observed in laboratory animals and similar de-
grees of infection from animals captured in nature,
including those from populations experiencing mass mor-
tality events and population declines (Carey et al., 2003a,
2003b; Green et al., 2002; Jancovich et al., 1997; Nichols et
al., 2001). Although most members of the phylum Chy-
tridiomycota typically meet their nutritional requirements
by breaking down organic matter in aquatic systems, some
species are parasitic upon selected invertebrates, such as
insects (Longcore et al., 1999). B. dendrobatidis is the first
chytrid fungus known to be parasitic upon a vertebrate
host. Zoospores of this fungus preferentially attack kerati-
nocytes in the skin of metamorphosed amphibians
(Longcore et al., 1999; Pessier et al., 1999). Although
amphibian larvae appear to lack keratin in their epidermis,
this pathogen has been identified in the keratinized
mouthparts of tadpoles and toes of premetamorphic tad-
poles of a few species (Berger et al., 1998; Carey et al.,
2003a, 2003b; Fellers et al., 2001; Rachowicz and Vreden-
burg, 2004; Marantelli et al., 2004).
In the few years since its discovery, some progress has
been made in understanding the interactions of this fungal
pathogen with amphibians (see reviews by Berger et al.,
2004; Carey et al., 2003a, 2003b; Daszak et al., 2003;
Davidson et al., 2003; Longcore et al., 1999; McDonald et
al., 2005; Nichols et al., 2001; Pessier et al., 1999; Retallick
et al., 2004). Nichols et al., (2001) experimentally exposed
captive Dendrobates tinctorius and D. auratus to the type
isolate of B. dendrobatidis by dripping a solution containing
zoospores onto the back and legs of these amphibians once
a day for 30 days or 4–5 days per week for 4 weeks or by
immersing them in water containing chytrid zoospores in
their cages. All exposed animals died with skin infections,
whereas control animals did not develop infections. Some
amphibian species, such as bullfrogs (Rana catesbeiana)
and tiger salamanders (Ambystoma tigrinum), apparently
carry the pathogen on their epidermis without developing
lethal infections; therefore, they can serve as reservoirs for
the transmission of chytridiomycosis to susceptible species
(Daszak et al., 2003; Davidson et al., 2003; Hanselmann et
al., 2004; Weldon et al., 2004). Further, B. dendrobatidis can
persist in an endemic state in healthy frogs once an epi-
demic wave has passed through amphibian populations
(McDonald et al., 2005; Retallick et al., 2004). To date,
however, many questions remain about the interaction of
this pathogen with amphibians, including what constitutes
the minimal infective dose of zoospores necessary to cause
a lethal infection, how various environmental factors affect
the ability of zoospores to cause an infection, and how
various amphibian species differ in susceptibility to infec-
tion by this pathogen. Much more information about these
interactions is needed to develop effective methods for
preventing further amphibian mortalities and population
declines.
We here report on a series of experiments designed to
examine the interaction of B. dendrobatidis with the boreal
toad (Bufo boreas), a species that is known to be susceptible
to infection by this pathogen. Boreal toads, widely dis-
tributed throughout many parts of the western United
States, suffered severe population declines and extinctions
in the southeastern part of their range (Carey, 1993; Corn
et al., 1989) in the late 1970s to early 1980s. As a result, this
species is classified as ‘‘endangered’’ in Colorado (Goettl,
1997) and New Mexico, a ‘‘species of special concern’’ in
Wyoming (Keinath and Bennett, 2000), and a ‘‘sensitive
species’’ in Utah (Utah Division of Wildlife Resources,
1998); it is classified as ‘‘warranted but precluded’’ for
federal listing as a threatened or endangered species by the
U.S. Fish and Wildlife Service. Since 1996, B. dendrobatidis
has been associated with mortalities in some of the few
remaining populations of toads in Colorado (Muths et al.,
2003). Its presence in a few museum specimens collected in
Colorado during the 1970s and the similarities between the
patterns of historical declines and current mass mortalities
suggest, but do not conclusively prove, that the historical
die-offs of this species in Colorado in the 1970s were due to
this pathogen (Carey, 1993; Carey et al., 1999). The boreal
toad is an ideal model system for examining the interac-
tions between Batrachochytrium and amphibians because
6 Cynthia Carey et al.

chytrid-free boreal toadlets can be obtained from a Colo-
rado Division of Wildlife hatchery and because an isolate of
this fungal pathogen from a boreal toad in a Colorado
population is available for experimentation.
These experiments were designed to investigate (1)
whether lethal infections due to chytridiomycosis can be
induced experimentally in boreal toadlets, (2) how vari-
ations in temperature and body mass affect survival of
boreal toadlets exposed to B. dendrobatidis zoospores, (3)
whether uninfected boreal toadlets can become infected by
exposure to water in which B. dendrobatidis-infected
toadlets have been housed, and (4) how the duration of
exposure to, and the dosage of B. dendrobatidis zoospores
affect the postexposure survival of boreal toadlets. An-
swers to these questions are important for identifying
factors that might contribute to the ability of B. dendro-
batidis to infect and kill boreal toads in nature. Although
the interactions between zoospores and amphibians may
differ in nature and in the laboratory, experimental
measurements of the number of zoospores necessary to
initiate a fatal infection and the amount of exposure time
necessary for infection to occur, as well as experimental
determination of whether or not amphibians can become
infected by contact with water containing zoospores in the
laboratory, are valuable steps in understanding these
phenomena in nature. In addition, it is important to
understand how environmental factors, such as tempera-
ture, affect the dynamics of infection of amphibians by
B. dendrobatidis.
MATERIALS AND METHODS
General Husbandry
Because boreal toads are designated as endangered or
threatened throughout the southeastern part of their geo-
graphic distribution, our experiments were necessarily re-
stricted to toadlets that were provided from the Colorado
Division of Wildlife’s John W. Mumma Native Aquatic
Species Restoration Facility (NASRF). The toadlets used in
these experiments were raised at NASRF from eggs or
tadpoles collected at five different localities in the moun-
tains of Colorado in late May through June. Because of the
relative synchronicity of egg laying at various boreal toad
breeding locations, the age of toadlets used in this study
probably differed by less than 6 weeks, regardless of the
locality of origin. Because the development of a polymerase
chain reaction (PCR) assay for B. dendrobatidis occurred
after the experiments described here were conducted, we
were unable to verify that all the toadlets were free of
chytrid infection at the start of each experiment. However,
the toadlets were raised by the hatchery in a manner that
minimized exposure to B. dendrobatidis and no outbreaks
of this pathogen have been reported for hatchery toads.
Subsequent PCR analysis of samples from the hatchery
indicates that the hatchery currently is free from contam-
ination with this pathogen. Therefore, we feel confident
that the toadlets had not been exposed to B. dendrobatidis
prior to their use in these experiments.
After transport from NASRF to the University of
Colorado in Boulder, young-of-the-year toadlets were
maintained at 23�C, the temperature shown to be optimal
for B. dendrobatidis growth (Piotrowski et al., 2004), on a
12-hour/12-hour light/dark cycle in troughs containing tap
water. Toadlets were fed Drosophila or pin-head crickets
(Fluker Farms, Port Allen, LA) three times a week. The
crickets were fed Fluker’s Calcium Fortified Cricket
Quencher in order to provide vitamins and minerals to the
toadlets. During experiments, toadlets were housed in
walk-in environmental chambers in which temperature,
light, and feeding conditions were controlled as described
above, unless specified otherwise in individual experiments.
In each experiment, however, toadlets were held in 20%
Holtfreter’s solution (pH 6.5, the optimal pH for B. dend-
robatidis growth [Piotrowski et al., 2004]), made with
double-distilled water to minimize exposure of chytrid
zoospores to contaminants in tap water.
B. dendrobatidis Cultures and Exposures
Isolate JEL#275 of B. dendrobatidis was used to expose
toadlets to this pathogen. This strain was originally isolated
from an infected boreal toad from Clear Creek County,
Colorado, by Joyce Longcore. Cultures were grown in H-
broth (10 g tryptone and 3.2 g glucose/liter distilled water)
at 23�C for 4–7 days prior to the beginning of an experi-
ment. On the day that toadlets were to be exposed to B.
dendrobatidis, zoospores were filtered through a sterile 20
lm nylon mesh filter (Spectra/Mesh�; Spectrum Labora-
tories, Rancho Dominguez, CA) to remove sporangia and
then an aliquot of the filtrate was counted with a hemacy-
tometer to determine the concentration of zoospores per
milliliter in the filtrate. A solution containing the desired
concentration of zoospores was made by diluting the filtered
H-broth culture with 20% Holtfreter’s solution (pH 6.5)
and sufficient penicillin/streptomycin (Sigma, St. Louis,
Exposure of Boreal Toads to B. dendrobatidis 7

MO) to comprise 1% of the final volume. Toads in the
exposure groups were placed in aliquots of this solution.
Because the toadlets were exposed to the chytrid zoospores
in a nonsterile environment, penicillin/streptomycin was
necessary to minimize bacterial growth on the nutrients in
the H-broth. Toads in control groups were exposed to
identical proportions of B. dendrobatidis-free H-broth and
penicillin/streptomycin in 20% Holtfreter’s solution.
Detection of B. dendrobatidis DNA in Toad Skin
These experiments were conducted before a PCR test for B.
dendrobatidis DNA was developed. In anticipation of the
development of this test, samples were collected during or
at the end of these experiments by scraping the ventral skin
with a sharpened wooden dowel, which was then placed in
1 ml of 0.25 M EDTA (pH 8) saturated with NaCl. These
samples were subsequently analyzed by Pisces Molecular
(Boulder, CO) for DNA specific to B. dendrobatidis using
PCR primers developed by Annis et al., (2004). Toad car-
casses were stored in 10% formalin. Sections of skin from
the ventral pelvic region (pelvic or drink patch) were
processed for histological examination and embedded in
paraffin. Sections (5 lm) were stained with hematoxylin
and eosin.
Experiment 1: Can Infections with
Batrachochytrium Be Experimentally Induced in
Boreal Toadlets, and How Do Temperature and
Body Mass Affect the Survival of These Toadlets
after Exposure?
Our first experiment was designed to test whether boreal
toadlets could be infected experimentally with the B.
dendrobatidis isolate JEL#275. Each of 160 toadlets was
weighed, placed in an individual 236 ml plastic Ziploc
container with a perforated lid, and randomly assigned to
one of four groups: control or exposed, each maintained at
either 23�C or 12�C. Mass ranged from 1.6–38.0 g (aver-
age ± standard error [SE] mass = 12.41 ± 0.56 g). Analysis
of variance (ANOVA) revealed no significant differences in
mean initial mass for toadlets among groups (F = 0.170,
P = 0.6806) or between the two temperature treatments
(F = 2.324, P = 0.1294). The group · temperature inter-
action was also nonsignificant (F = 44.701, degree of free-
dom [DF] = 1, P = 0.3430).
For toadlets in exposure groups, we used what we
believed to be a large dose (106 zoospores/toadlet daily) and
a relatively long exposure time (72 hours) to maximize the
chance of producing skin infections and mortality under
laboratory conditions. Toads in the exposure groups were
exposed individually to this dose of zoospores in 20 ml of
Holtfreter’s solution constituted as described above. This
volume was adequate to cover the floor of the Ziploc
container and to immerse the ventral side of the toadlet
throughout the exposure period. Control toadlets were
individually exposed to an identical volume of the sham
exposure solution. Fresh solutions containing zoospores
were made from B. dendrobatidis cultures each day. After
24 and 48 hours, the initial exposure solutions were dis-
carded from each container and replaced with the same
volume of identically constituted solutions. After 72 hours,
all toadlets were transferred from the small containers and
placed individually in 15 · 30 · 11 cm plastic containers
with 200 ml of B. dendrobatidis-free 20% Holtfreter’s
solution (pH 6.5). Holtfreter’s solutions in the cages were
replaced three times per week when the toadlets were fed.
The containers were held flat so that the ventral surfaces of
the toadlets in a normal sitting posture were in continuous
contact with the solution.
In order to examine the time course between exposure
and the appearance of detectable infection, five toadlets
from each group were sacrificed by cervical dislocation,
followed by spinal and cerebral pithing, on a random
sampling schedule on postexposure days 1, 3, 8, 14, and 21.
Skin scrapes were collected for PCR analysis, and carcasses
were preserved for histological examination.
Thirty-two of the 80 exposed toadlets died on or prior
to their assigned sampling date. On the assumption that
these toadlets died from B. dendrobatidis infections (as-
sessed after death with histology and, later, PCR), experi-
ment 1 afforded additional opportunities to analyze the
effects of two variables, temperature variation and toadlet
body mass, that could plausibly affect the number of days
to death. The relation of the number of days to death as a
function of body mass was evaluated with least-squares
regression. The average time of survival of toadlets that
died prior to their scheduled sampling date in two tem-
perature treatments, 23�C and 12�C, was evaluated using
Student’s t-test. Tissue samples from all toadlets that died
prior to their scheduled sampling date were analyzed by
PCR.
As the experiment progressed, we noted several
behavioral and physiological differences between control
and exposed toadlets at each temperature. Anecdotal
observations of behavioral differences were not statistically
8 Cynthia Carey et al.

analyzed. However, on the last sampling date, the number
of respirations per minute of all surviving exposed toadlets
(n12�C = 6, n23�C = 5) and randomly selected control
toadlets (n12�C = 6, n23�C = 4) were counted; the mean
number of respirations per minute of control and exposed
animals was compared with two-way ANOVA, with treat-
ment (control or exposed) and temperature (12�C or 23�C)
as the independent variables.
When the experiment was terminated 3 weeks after the
initial exposure, some control and all of the remaining ex-
posed animals were sacrificed. Holtfreter’s solution and the
containers in which exposed and control animals had been
sitting for 24 hours were immediately used in experiment 2.
Experiment 2: Can Uninfected Boreal Toadlets
Become Infected by Exposure to Liquid (i.e.,
Holtfreter’s Solution) in Which Infected Toadlets
Have Been Housed?
This experiment was designed to determine whether chy-
tridiomycosis could be transmitted to uninfected animals
through exposure to Holtfreter’s solutions used in the
previous experiment to house infected toadlets. Twenty-
one control animals remained at the end of experiment 1:
11 had been maintained at 12�C and 10 at 23�C. These
toadlets were assigned randomly to containers from
experiment 1 which had housed either control or exposed
toadlets in 200 ml Holtfreter’s solution that had been in the
container the preceding 24 hours. Toadlets for experiment
2 remained in the experiment 1 Holtfreter’s solution for 48
hours, after which the liquid was replaced with clean, B.
dendrobatidis-free, 20% Holtfreter’s solution (pH 6.5). This
solution was then changed three times per week during
feedings. Survival of toadlets was monitored daily for 34
days. When a toadlet died, the date of death was recorded
and skin scrapes were collected for PCR analysis. A log-
rank test on censored survival data (StatXact; Cytel Statis-
tical Software, Cambridge, MA) was used to assess whether
the patterns of survival of control and exposed groups
differed significantly.
Body masses of toadlets used this experiment ranged
between 6 and 30 g. Average body masses of control and
exposed toadlets, compared using two-way ANOVA, did
not differ significantly (P = 0.6782, DF = 1) and there was
no significant interaction term (P = 0.6636, DF = 1).
However, the average mass of the combined control and
exposed toadlets at 23�C was significantly heavier than the
control and exposed body masses at 12�C.
Experiments 3 and 4: How Do the Time of Exposure
and the Dosage of B. dendrobatidis Zoospores Affect
the Survival of Boreal Toadlets?
As noted below (see Results), PCR analysis and histology of
tissues from exposed toadlets in experiment 1 indicated
that exposure to 106 zoospores per day for 3 days was
sufficient to cause chytridiomycosis and death. Experiment
3 examined the effects of a variety of dosages and durations
of exposure on the number of days survived. The experi-
ment was designed as a 2 · 5 factorial, with one factor
being exposure duration (1 or 3 days) and the other being
dosage of zoospores/20 ml (0 [control], an estimated 1
zoospore, 100, 104, or 106 zoospores], resulting in a total of
10 experimental groups with 15 toadlets each. Toadlets
were weighed and randomly assigned to one of the 10
groups. Mean initial toadlet mass did not differ signifi-
cantly among groups (F = 0.576, DF = 9, P = 0.8154).
Individual toadlets from control and varying dosage
groups designated for 1-day exposure were placed in 236
ml Ziploc containers containing 20% Holtfreter’s solution
(pH 6.5) with either B. dendrobatidis-free broth (controls)
or broth containing various concentrations of zoospores
for 24 hours, after which the solutions were replaced daily
with 20 ml Holtfreter’s solution (pH 6.5). After 3 days,
toadlets were placed in individual 15 · 30 · 11 cm plastic
boxes for the duration of the experiment. Even though
these toadlets were exposed to B. dendrobatidis zoospores
for only 1 day, this procedure on days 2 and 3 ensured that
handling of the toadlets in the 1-day and 3-day exposures
was identical. Toadlets that were exposed for 3 days and
their controls were kept in the small Ziploc containers for 3
days and given fresh B. dendrobatidis-containing or B.
dendrobatidis-free doses of Holtfeter’s solution daily. At the
end of the 3-day exposure, these groups were also placed
individually in large plastic boxes at 23�C for the 42-day
duration of the experiment. Toads were fed and water was
changed three times per week; mortality was monitored
daily. Skin scrapes for PCR analysis were collected at death
or, for toadlets that survived the entire 42-day period, at
the end of the experiment.
In experiment 4, we examined the effect of 1-day
exposure to a different set of zoospore concentrations (0
[control], 20, 40, 60, 100, and 103 zoospores/20 ml) on
survivability with groups of 10 Bufo boreas toadlets each.
This experiment was also conducted at 23�C and moni-
tored for 42 days under the same housing and care con-
ditions as in experiment 3. No significant differences in
Exposure of Boreal Toads to B. dendrobatidis 9

initial toadlet mass existed among groups (F5,54 = 1.725,
P = 0.1445). At the end of the experiment, skin scrapes
were collected for PCR analysis.
Survival curves of control and exposure groups in
experiments 3 and 4 were evaluated by the StatXact log-
rank test for censured survival data. This test indicated
whether the rates of death and percentage of animals in
each group that died differed significantly among dosages
and (in experiment 3) the duration of exposure. Addi-
tionally, the known fate modeling procedure in program
MARK (White and Burnham, 1999) was used to isolate the
most parsimonious models from suites of candidate models
containing all possible first-order combinations of dose,
exposure duration (in experiment 3), and body mass to
determine their relative importance in determining the
length of survival. Program MARK uses second-order
Akaiki information criteria (AICc; Burnham and Anderson,
2002) to identify the model that best describes the data
without sample overparameterization. Using this approach
for model selection is superior to traditional hypothesis
testing for this data set because it allows simultaneous
comparison of multiple candidate models, balances preci-
sion and bias when selecting the appropriate model
(Burnham and Anderson, 2002), and is not restricted to
parametric data.
In addition, we examined whether the results of these
experiments were consistent with a simple model of the
disease process, based on the assumption that populations
of B. dendrobatidis on the host grow exponentially until a
threshold density is reached that produces host mortality.
Exponential population growth can be represented as a
linear increase of log(number of parasites) with time. A
linear function has two parameters, the slope and the
intercept. The intercept for these population growth lines is
the number of zoospores that settled on the first day of
exposure, day 1 of this experiment. For treatments in which
toadlets were exposed to an estimated single zoospore, this
must equal log10(1) for successful infections. For other
treatments, this number should be equal to log10(np), where
n is the number of zoospores to which the toadlets in each
treatment were exposed and p is the proportion of zoosp-
ores that colonize a toadlet in 1 day of exposure. For
experiment 3, we assumed that p was constant across all
treatments, estimated p using two methods, and used the
average of these estimates for our model. One estimate of p
was calculated as the proportion of toadlets that were ex-
posed to an estimated single zoospore for 1 day that became
infected. The second was calculated for the treatment in
which individuals were exposed to 100 zoospores as the
probability of colonization per zoospore that made the zero
term of the binomial distribution equal to the observed
proportion of failures to infect. We also assumed that all
population growth lines had the same slopes. If all the
growth lines had the same slopes, differing only in inter-
cepts, and mortality occurs when these growth lines cross a
threshold number, then the common slope of the growth
lines can be estimated by dividing 2 (the difference in ele-
vation in log units between the lines for initial exposures to
102, 104, and 106 zoospores) by the differences in time to
mortality averaged between all possible pairs of individuals
in the treatments exposed for 1 day to 102 or 104 and to 104
or 106 zoospores. We compared the average time to mor-
tality predicted by these four lines with the observed time to
mortality for toadlets exposed to zoospores for 1 day. We
cross-checked these results with those for individuals ex-
posed to zoospores for 3 days. We assumed that the slopes
and intercepts of the B. dendrobatidis population growth
lines remained the same as those for individuals exposed for
1 day. We calculated the total estimated B. dendrobatidis
population on individuals in each treatment as the sum of
the three population growth lines for the populations star-
ted on each day of exposure and determined estimated times
to mortality by inspecting the values of these sums.
We also examined how well our growth models fit the
data from experiment 4. We assumed that the slopes of
population growth lines remained the same as those esti-
mated for the third experiment since the conditions under
which toadlets were maintained following infection were
identical. We estimated the intercepts of the growth lines
for each exposure level in the same manner but used a value
of p derived from the observed rates of infection in this
experiment, by calculating the p values necessary to make
the zero term of the binomial distribution equal to the
observed proportion of failures to infect for the 20, 40, and
60 zoospore treatments and averaging these values.
RESULTS
Experiment 1: Can Infections with B. dendrobatidis
Be Experimentally Induced in Boreal Toadlets, and
How Do Temperature and Body Mass Affect the
Survival of These Toadlets after Exposure?
Both histological examination of ventral skin and PCR
analysis for B. dendrobatidis DNA in ventral skin confirmed
that exposure of boreal toadlets to 106 zoospores for 3 days
10 Cynthia Carey et al.

caused chytridiomycosis. Histological lesions observed in
exposed toadlets were moderate epidermal hyperplasia
characterized by increased epidermal thickness and disor-
ganization of keratinocytes and mild to moderate ortho-
keratotic hyperkeratosis (Fig. 1). Within keratinocytes in
the stratum corneum, there were moderate numbers of
characteristic chytrid thalli, including zoosporangia with
developed zoospores and septate (colonial) thalli charac-
teristic of B. dendrobatidis. Chytrid thalli were evident in
histological sections as early as 9 days following infection.
Tissues of only 3 of the 80 exposed animals tested negative
for B. dendrobatidis DNA; these were sampled either on day
1 or day 3 following exposure. Additionally, 32 exposed
animals died on or prior to their scheduled sampling date.
Because both histological analysis and PCR indicated the
presence of B. dendrobatidis in the skin of these animals, we
concluded that 3-day exposure to 106 zoospores was suf-
ficient to cause death in boreal toadlets.
One control toadlet died on the first day of the 21-
day experiment and was found to be free of B. dendro-
batidis infection by PCR analysis. Tissues of 6 of the 20
control toadlets submitted for PCR analysis were weakly
positive for B. dendrobatidis DNA, probably due to con-
tamination during the sampling procedure at the end of
the study. Although instruments were cleaned with etha-
nol between samples, this cleaning procedure may not
have been sufficient to remove all B. dendrobatidis DNA,
resulting in contamination. No other control animals,
including those toadlets nominally positive for B. dend-
robatidis DNA, died prematurely before their assigned
sampling date.
The mean number of days survived by toadlets that
died before their scheduled sampling date was 14.0 ± 0.8
days (n = 14) and 13.8 ± 0.8 days (n = 18) at 12�C and
23�C, respectively. These values did not differ significantly
(t = 0.188, P = 0.8525, DF = 30), indicating that tem-
perature variation over this range had no significant effect
on the survival time of these boreal toadlets.
Body mass of all toadlets in experiment 1 ranged be-
tween 1–38 g, but the masses of those that died on or before
their scheduled sampling date ranged only between 4–19 g.
Our regression analysis indicated that toadlet body mass
within this range had a significant effect on the number of
days survived. Because temperature had no significant ef-
fect on survival time, the days of survival of toadlets held at
12�C and 23�C were pooled and analyzed; a highly signif-
icant correlation existed between the number of days sur-
vived following exposure to B. dendrobatidis zoospores and
toadlet body mass (r = 0.795, n = 30, P < 0.0001; Fig. 2).
The least-squares regression equation best describing this
relationship is as follows: Days survived = 8.79 + 0.545 ·Mass (g) (F = 49.798, P < 0.0001). An SAS t-test (Satt-
erthwaite method) showed a highly significant relationship
between mass and survival time, with shorter survival times
associated with smaller masses (t = 3.62, P = 0.0018).
Control and exposed toadlets exhibited several
behavioral differences as the experiment progressed. Spe-
cifically, exposed toadlets held their bodies out of water as
Figure 1. Histopathology of Bufo boreas experi-
mentally infected with B. dendrobatidis. A: Skin
from an uninfected control, magnified ·40. B:
Skin from a toad exposed to B. dendrobatidis 12
days previously. The epidermis is thickened with
disorganized keratinocytes (hyperplasia), and
there is a cluster of chytrid thalli within the
superficial keratinized layers (stratum corneum,
center), magnified ·40. C: Detail of chytrid thalli
within the stratum corneum. Numerous devel-
opmental stages are present including a flask-
shaped zoosporangium containing numerous
discrete zoospores.
Exposure of Boreal Toads to B. dendrobatidis 11

much as possible by climbing on the walls of their con-
tainer or by adopting a four-legged posture that raised their
ventral surface above the water on the bottom of the cage.
Sometimes exposed toadlets elevated their toes out of the
water. In comparison, control toadlets usually sat in the
water and were rarely observed with ventral body surfaces
out of the water.
ANOVA indicated that control toadlets had a signifi-
cantly higher respiration rate than exposed toadlets by the
end of the 3-week experiment (F = 10.6, DF = 1, P =
0.0047; Table 1). Temperature had no significant effect on
respiration rates of exposed and control toadlets held at
12�C and 23�C (F = 3.592, DF = 1, P = 0.0752), nor did the
interaction between treatment and temperature (F = 2.770,
DF = 1, P = 0.1144).
Experiment 2: Can Uninfected Boreal Toadlets
Become Infected by Exposure to Water in Which
Infected Toadlets Have Been Housed?
Exposure to water in which toadlets infected with B.
dendrobatidis had been living for 24 hours caused signifi-
cant chytridiomycosis and death of boreal toadlets (Fig. 3).
A log-rank test for censored survival data indicated a sig-
nificant difference in the pattern of survival of control and
Figure 2. Number of days survived by boreal toadlets (Bufo boreas) as a function of body mass following exposure to 106 zoospores of
B. dendrobatidis for 3 days. Each point represents data for one toad.
Table 1. Mean Breaths per Minute of Control (Nonexposed)
Boreal Toadlets (Bufo boreas) or Toads Exposed to Chytrid Fungal
Zoospores (Batrachochytrium dendrobatidis) at Two Constant
Temperatures
Treatment and temperature Mean ± SE breaths/min n
Control (12�C) 147 ± 5.4 6
Exposed (12�C) 125 ± 9.1 6
Control (23�C) 195 ± 9.0 4
Exposed (23�C) 128 ± 24.5 5
Figure 3. Changes in the percentage survival of groups of boreal
toadlets (Bufo boreas) as a function of time (days) during a 34-day
experiment following placement in water in which uninfected toads
(control group) or toads infected with B. dendrobatidis (exposed
group) had been held. n = 10 for controls (5 at 23�C, 5 at 12�C), n =
11 for exposed (5 at 23�C, 6 at 12�C).
12 Cynthia Carey et al.

exposed toadlets (two-sided exact inference P = 0.0001).
Mortalities of exposed toadlets began on day 16. Exposed
toadlets held at 12�C and 23�C survived an average of
23.8 ± 2.42 (n = 6) and 25.45 ± 2.56 (n = 4) days,
respectively, during the 34-day experiment. Only one of the
exposed toadlets (at 23�C) survived the full 34 days of the
experiment. All of the exposed toadlets tested positive for
B. dendrobatidis DNA by PCR analysis. In contrast, 9 of the
10 toadlets (5 at 12�C and 5 at 23�C) transferred to water
previously used by control (nonexposed) toadlets survived
the entire 34 days of the experiment. One control toadlet
(at 23�C) died on day 34. None of the control toadlets,
including the one that died on day 34, tested positive for B.
dendrobatidis DNA.
During the course of this experiment, toadlets placed
in water previously occupied by B. dendrobatidis-exposed
toadlets exhibited avoidance of water and decreased rates of
respiration as noted in exposed toadlets from experiment 1.
Experiments 3 and 4: How Do the Time of Exposure
and the Dosage of B. dendrobatidis Zoospores Affect
the Survival of Boreal Toadlets?
Both the number of zoospores to which a toadlet was ex-
posed (dosage) and the number of days of exposure
strongly affected the duration of survival of exposed boreal
toadlets (log-rank test two-sided exact inference P =
0.0001). The average number of days survived by toadlets
exposed to 106 zoospores (15.5 ± 1.9 and 16.4 ± 3.9 days
for the 1- and 3-day exposure groups, respectively) did not
differ significantly (Fig. 4). At lower dosages (an estimated
1, 100, or 10,000 zoospores), both dosage and duration of
exposure to B. dendrobatidis zoospores significantly affected
the mean length of survival, percent mortality, and per-
centage of toadlets actually infected by the exposure treat-
ment (Table 2).
The duration of exposure had no effect on the per-
centage of toadlets surviving at high dosages: all toadlets in
groups exposed to 10,000 and 106 zoospores died by the
end of the test (Table 2). At lower dosages, only 13% of
toadlets exposed for 3 days to an estimated single zoospore
survived, whereas 93% of those exposed to that dosage for
only 1 day survived the 42-day experiment (Table 2).
Finally, both the time of exposure and the dosage af-
fected the percentage of toadlets in each group infected by
B. dendrobatidis. For example, only 38% of the toadlets
became infected when exposed to an estimated single
zoospore for 24 hours, yet 100% were infected by a 3-day
exposure (Table 2).
Duration of exposure, dose, and toadlet mass were
factors available for analysis for their effects on survival
time in experiment 3. These factors were evaluated by
ranking a suite of candidate models in program MARK
using the logit link function (Table 3). The model that
included exposure duration and dose (model A) had
nearly three times the support (AICc weight = 0.572) than
the one that also included mass (model B, AICc weight =
0.225). Of the top four models (models A-D), those that
included exposure duration as a variable (A and B) were
almost four times more strongly supported than those
without (C and D). Dose was selected in all four of the
top models, demonstrating its importance in predicting
survival of toadlets. The most parsimonious model was as
follows:
Figure 4. Experiment 3: Changes in the per-
centage survival of groups of boreal toadlets (Bufo
boreas) exposed to different dosages of B.
dendrobatidis zoospores for 1 or 3 days in a
42-day experiment. n = 15 for each group.
Exposure of Boreal Toads to B. dendrobatidis 13

S ¼ expð4:987 þ 0:459DUR � 0:684DOSEÞ1 � expð4:987 þ 0:459DUR � 0:684DOSEÞ
where S is daily survival, DUR is the exposure duration, and
DOSE is the dose used.
Toadlets were exposed to 100 zoospores for 1 day in
both experiments 3 and 4. The percentage of toadlets sur-
viving the 42-day experiments was similar (27% in exper-
iment 3 and 30% in experiment 4). A t-test showed no
significant difference in mean survival time for toadlets at
this common dosage level (mean survival(experiment 3) =
34.5 ± 1.6 days, mean survival(experiment 4) = 33.0 ± 2.3
days; t = 0.562, P = 0.5796, DF = 23). These results give us
confidence that the conditions under which the experi-
ments were run, although offset in time, were sufficiently
similar that the results can be compared between the two
experiments.
Except for the 100-zoospore exposure replicate, toadlets
were exposed to a different set of dosages in experiment 4.
All toadlets exposed to 1,000 zoospores died within 42 days.
At dosages lower than 1,000 zoospores, the percentage of
toadlets surviving the 42-day experiment and the percentage
infected with B. dendrobatidis varied among groups (Ta-
ble 2, Fig. 5); some toadlets exposed to lower dosages lived
the full 42 days (Fig. 5, Table 2). One control animal also
tested weakly positive for B. dendrobatidis DNA (Table 2).
A similar suite of candidate models were ranked by
program Mark for experiment 4 (Table 4). Because
experiment 4 had only a single exposure duration and the
doses were within an order of magnitude, the contribution
Table 3. Models Considered by Program MARK to Predict Survival (S) in Toadlets from Experiment 3 and Their Ranking by AICc:
Parameters Considered Include Exposure Duration (Dur), Dose, and Mass
Model Parameters AICc
Delta
AICc
AICc
Weight
Model
Likelihood Par Deviance
A S(DurDose) 835.965 0.00 0.572 1.0000 3 829.960
B S(DurDoseMass) 837.832 1.87 0.225 0.3933 4 829.823
C S(Dose) 838.727 2.76 0.144 0.2514 2 834.724
D S(DoseMass) 840.476 4.51 0.060 0.1048 3 834.470
E S(Dur) 913.565 77.60 0.000 0.0000 2 909.562
F S(DurMass) 915.209 79.24 0.000 0.0000 3 909.203
G S(.) 917.353 81.39 0.000 0.0000 1 915.352
Par, number of parameters; S(.), daily survival, was the only parameter used in the model.
Table 2. Proportion (%) of Toadlets Surviving 42 Days following Exposure to Varying Dosages of Chytrid Fungi Zoospores and
Proportion That Tested Positive for B. dendrobatidis by PCR: Control Groups Were Not Exposed to the Fungus
Experiment 3 dosages (n = 15) Control 1 100 10,000 1,000,000
3-Day exposure
Surviving 42 days 100% 13% 0% 0% 0%
Positive 0% 100% n/a n/a n/a
1-Day exposure
Surviving 42 days 100% 93% 27% 0% 0%
Positive 0% 38% n/a n/a n/a
Experiment 4 dosages (n = 10) Control 20 40 60 100 1000
1-Day exposure
Surviving 42 days 100% 50% 60% 40% 30% 0%
Positive 10% 100% 60% 90% 90% 100%
n/a, not applicable.
14 Cynthia Carey et al.

of mass was more than three times that for dose in terms of
toadlet survival (model I AICc weight = 0.375 vs. model J
AICc weight = 0.118). Even though a small range of masses
was used in this experiment, a size-related effect on survival
was clearly demonstrated, with increasing mass resulting in
increased survival time. This mass effect can be summa-
rized as follows:
S ¼ expð2:568 þ 0:155MASSÞ1 þ expð2:568 þ 0:155MASSÞ
For model I, S is daily survival and MASS is given in grams.
We used the data from experiments 3 and 4 to address
the question of whether death occurs when the number of B.
dendrobatidis sporangia infecting a toadlet exceeds a
threshold. Our models of the growth of B. dendrobatidis
populations on individuals fit the data closely (Table 5) and
predict that the threshold for death is about 107–108 zoo-
sporangia per toadlet. Using the models fitted for 1-day
exposures in experiment 3 produced predicted times to
mortality that were close to the actual times experienced by
individuals exposed to zoospores over 3 days. The intercepts
estimated for experiment 4 using the proportion of animals
successfully infected are low, suggesting that rates of suc-
cessful establishment of zoospores on hosts were lower in
this experiment than in experiment 3. However, the models
also fit the data for this experiment reasonably well. The
relationship between observed and predicted median dates
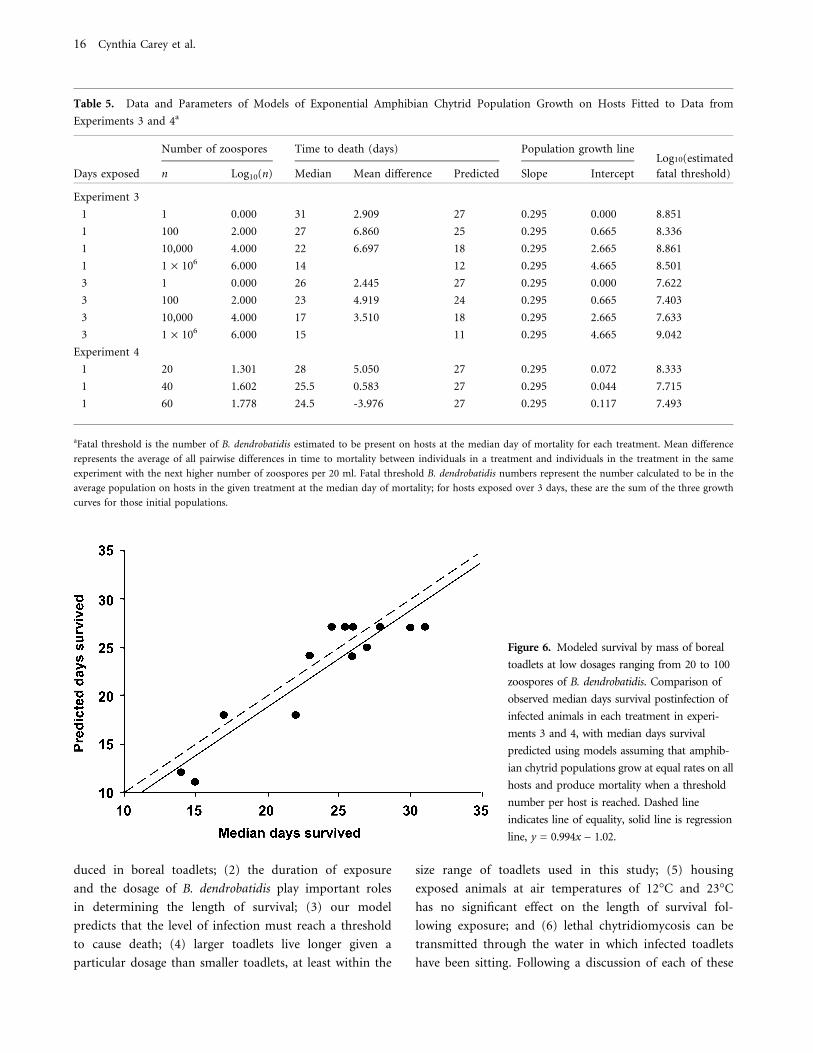
of mortality for experiments 3 and 4 is shown in Figure 6.
The fact that the regression accounts for most of the vari-
ation in the data and has a slope almost identical to 1
suggests that our models are a good reflection of the actual
disease process in boreal toads.
DISCUSSION
We believe that this study has a number of significant
findings: (1) chytridiomycosis can be experimentally in-
Figure 5. Experiment 4: changes in the per-
centage survival of groups of boreal toadlets
(Bufo boreas) exposed to different dosages of
B. dendrobatidis zoospores for 1 day in a 42-day
experiment. n = 10 for each group.
Table 4. Models Considered by Program MARK to Predict Survival (S) in Toadlets from Experiment 4 and Their Ranking by AICc:
Parameters Considered Include Dose and Mass
Model Parameters AICc
Delta
AICc
AICc
Weight
Model
Likelihood Par Deviance
H S(DoseMass) 304.472 0.00 0.441 1.0000 3 298.455
I S(Mass) 304.797 0.33 0.375 0.8497 2 300.789
J S(Dose) 307.117 2.65 0.118 0.2664 2 303.108
K S(.) 308.280 3.81 0.066 0.1489 1 306.278
Par, number of parameters; S(.), daily survival, was the only parameter used in the model.
Exposure of Boreal Toads to B. dendrobatidis 15
duced in boreal toadlets; (2) the duration of exposure
and the dosage of B. dendrobatidis play important roles
in determining the length of survival; (3) our model
predicts that the level of infection must reach a threshold
to cause death; (4) larger toadlets live longer given a
particular dosage than smaller toadlets, at least within the
size range of toadlets used in this study; (5) housing
exposed animals at air temperatures of 12�C and 23�C
has no significant effect on the length of survival fol-
lowing exposure; and (6) lethal chytridiomycosis can be
transmitted through the water in which infected toadlets
have been sitting. Following a discussion of each of these
Table 5. Data and Parameters of Models of Exponential Amphibian Chytrid Population Growth on Hosts Fitted to Data from
Experiments 3 and 4a
Number of zoospores Time to death (days) Population growth lineLog10(estimated
Days exposed n Log10(n) Median Mean difference Predicted Slope Intercept fatal threshold)
Experiment 3
1 1 0.000 31 2.909 27 0.295 0.000 8.851
1 100 2.000 27 6.860 25 0.295 0.665 8.336
1 10,000 4.000 22 6.697 18 0.295 2.665 8.861
1 1 · 106 6.000 14 12 0.295 4.665 8.501
3 1 0.000 26 2.445 27 0.295 0.000 7.622
3 100 2.000 23 4.919 24 0.295 0.665 7.403
3 10,000 4.000 17 3.510 18 0.295 2.665 7.633
3 1 · 106 6.000 15 11 0.295 4.665 9.042
Experiment 4
1 20 1.301 28 5.050 27 0.295 0.072 8.333
1 40 1.602 25.5 0.583 27 0.295 0.044 7.715
1 60 1.778 24.5 -3.976 27 0.295 0.117 7.493
aFatal threshold is the number of B. dendrobatidis estimated to be present on hosts at the median day of mortality for each treatment. Mean difference
represents the average of all pairwise differences in time to mortality between individuals in a treatment and individuals in the treatment in the same
experiment with the next higher number of zoospores per 20 ml. Fatal threshold B. dendrobatidis numbers represent the number calculated to be in the
average population on hosts in the given treatment at the median day of mortality; for hosts exposed over 3 days, these are the sum of the three growth
curves for those initial populations.
Figure 6. Modeled survival by mass of boreal
toadlets at low dosages ranging from 20 to 100
zoospores of B. dendrobatidis. Comparison of
observed median days survival postinfection of
infected animals in each treatment in experi-
ments 3 and 4, with median days survival
predicted using models assuming that amphib-
ian chytrid populations grow at equal rates on all
hosts and produce mortality when a threshold
number per host is reached. Dashed line
indicates line of equality, solid line is regression
line, y = 0.994x – 1.02.
16 Cynthia Carey et al.

findings, the findings of this study will be correlated with
observations of the population biology of boreal toad
populations experiencing mass mortalities associated with
B. dendrobatidis.
Chytridiomycosis Can Be Experimentally Induced
in Boreal Toadlets
This study showed that lethal chytridiomycosis can be
experimentally induced in the laboratory in boreal toadlets
by exposure to B. dendrobatidis zoospores and, in most
instances, death followed exposure within 5–7 weeks. In
our first experiment, exposure to 106 zoospores for 72
hours caused infections in 96% of boreal toadlets in that
group. These results coupled with those of Nichols et al.,
(2001) indicate that chytridiomycosis can be induced
readily in susceptible amphibian species.
Behavioral and physiological changes in the toadlets
were noted as the experiment progressed. First, infected
toadlets tried to avoid contact with water in the bottom of
their cages. The avoidance of water, regardless of the
motivational cause underlying the behavior, should pro-
long survival following infection because the rate of rein-
fection from zoospores released from the skin surface
would be low whenever the ventral surface was dry. Second,
the rate of respirations decreased as the severity of the
infections increased. These results may reflect a gradual
inhibition of metabolism by pathological changes in the
skin caused by B. dendrobatidis. These observations do not
support the hypothesis that B. dendrobatidis kills amphib-
ians by blocking oxygen uptake through the skin. If this
hypothesis were true, lung ventilation should increase if
cutaneous respiration is curtailed by B. dendrobatidis
infection. However, our data show the opposite trend.
Duration of Exposure and Dosage of B. dendrobatidis
Play Important Roles in the Length of Survival
The results of experiments 3 and 4 indicate that the initial
dosage of zoospores to which toadlets were exposed and the
length of exposure have significant effects on the number of
toadlets infected and the number of days survived (Figs. 4
and 5). Low dosages and a 1-day exposure fostered longer
average survival times than high dosages and a 3-day
exposure duration. However, average mortality of groups
exposed to 1 or 3 days of the highest dosage level of 106
zoospores did not differ significantly. The significance of
these data will be discussed in the next section.
Although toadlets were exposed for 24 hours to their
individual dose of zoospores in a small (20 ml) amount of
solution, not all individuals became infected at low expo-
sures. Our estimates of initial sizes of B. dendrobatidis
populations on toadlets (Table 5) suggest that approxi-
mately 4% of the zoospores present in the solution used for
exposures became established on the host during 24 hours
in experiment 3, while only 1% became established during
24 hours of exposure in experiment 4. Therefore, it appears
that a relatively high proportion (96–99%) of the zoospores
present in the immediate vicinity of toadlets fail to colonize
them successfully during a 24-hour period.
Reinfection by Zoospores Released by an Individual
Is Necessary for the Level of Infection to Reach a
Threshold Necessary to Cause Death
In experiment 1, both PCR and histological analysis of
toadlets killed at intervals throughout the experiment
showed increases in the amount of B. dendrobatidis
present and indicated that death occurred with moderate
to heavy infections. In this study, even a nominal dose of
1 zoospore in the water may be sufficient to result
eventually in a lethal infection. Our data show an inverse
correlation between dosage and length of survival: the
higher the number of zoospores in the initial exposure,
the shorter the time to death. The finding that the
number of days survived by boreal toadlets following
exposure to B. dendrobatidis zoospores is directly related
to dosage supports the contention that heavy infections
are needed to cause mortality. Exposure to a few zoosp-
ores causes infection, but time is needed for multiple
reinfections to reach the threshold required to kill the
animal. Exposure to a large number of zoospores causes
death more rapidly than a small number because a large
number of zoospores can relatively quickly produce the
critical degree of infection necessary to cause death.
Similarly, the length of exposure is also important for
reaching the critical level of infection to cause death. With
the exception of toadlets infected with 106 zoospores, the
average days of survival was lower in toadlets exposed for
3 days than for 1 day for a given dosage.
Our growth model (Fig. 6) indicates that the approxi-
mate threshold for infection to cause death ranges between
107 and 108 zoosporangia (thalli). When a zoospore invades
amphibian skin, it invades a keratinocyte and transforms into
a sporangium, which produces zoospores. Mature zoospores
exit from the sporangium through a discharge tube into the
Exposure of Boreal Toads to B. dendrobatidis 17

water adjacent to the epidermis. The zoospores then can
reinfect the same host or find other hosts. The mechanism by
which B. dendrobatidis kills amphibians is as yet unknown,
but two hypotheses have been advanced (Berger et al., 1998;
Carey et al., 2003a) that are consistent with the idea that a
certain level of infection must be reached to cause death.
First, the fungus might produce a toxin that diffuses from the
epidermis into the body and causes lethal tissue damage. A
heavy concentration of sporangia would produce more toxin
than a light one. Second, the presence of this chytrid in the
epidermis causes hyperplasia and hyperkeratosis, or excess
production of skin layers. Thickening of the skin layers in the
ventral skin of the pelvic area is hypothesized to cause lethal
disruptions in water and ion balance. A heavy infection
would result in a greater degree of ionic and/or water
imbalance than a light one.
Larger Toadlets Will Live Longer Given a Particular
Dosage than Smaller Toadlets, at Least within the
Size Range of Toadlets Used in This Study
Body size was a significant factor in determining the length
of survival of an infected toadlet in experiments 1, 3, and 4.
The size range of toadlets used in these experiments does
not represent the total variation in body mass observed in
field populations because these toadlets were young of the
year. Some boreal toads, particularly females, can grow to
over 80 g (Carey et al., 2005), but the endangered status of
this toad precludes use of large boreal toads in laboratory
experiments. Therefore, the significant association between
the duration of survival of boreal toadlets and body mass
may not necessarily hold for larger toads.
It is unknown why smaller animals died more rapidly
than larger ones. We have demonstrated that exposing
toadlets of similar sizes to different doses of zoospores leads
to differences in time to death consistent with the existence
of a threshold number of thalli necessary to cause death. It
seems likely that the size of this threshold is directly related
to the surface area of the animal. The threshold of small
animals, therefore, is both smaller and reached more
quickly than the threshold for larger animals. The effect
may relate to the fact that small toadlets have a relatively
small surface area of ventral skin. Alternatively, the skin of
smaller toadlets might be easier to penetrate than that of
larger ones. Also, there might be size-, and therefore age-,
related differences in maturation of skin immune defenses,
such as the ability to synthesize and secrete antimicrobial
peptides.
Housing Exposed Animals at Air Temperatures of
12�C and 23�C Has No Significant Effect on Length
of Survival Following Exposure
Numerous anecdotal accounts suggest that temperature can
be an important cofactor contributing to the success of B.
dendrobatidis in causing mortality of amphibians. Specifi-
cally, a number of amphibian deaths linked to B. dendro-
batidis coincide with the onset of seasonally cold
temperatures (Berger et al., 2004; Carey, 2000; Carey et al.,
1999; McDonald et al., 2005; Retallick et al., 2004). Further,
studies of a tropical frog indicate that B. dendrobatidis
infection may be cured by exposure to a temperature of
37�C (Woodhams et al., 2003).
These previous studies document that very high and
very low temperatures affect the interaction between B.
dendrobatidis and its hosts. This study, conducted at mod-
erate temperatures, found that housing exposed animals at
12�C or 23�C had no significant effect on survival in both
experiments 1 and 2. Most physiological and cellular rate
processes increase two- to threefold for every 10�C rise in
temperature (the Q10 rule) (Rome et al., 1992). Several
important factors could be temperature-dependent,
including the growth rate of sporangia, the rate of zoospore
release, and the swimming speed, penetration rate, and
keratinocyte invasion rate by zoospores. However, this study
shows that the overall length of time survived by B. dend-
robatidis-infected boreal toadlets does not obey the Q10 rule.
Additional research on the effect of temperature on the
interactions between B. dendrobatidis and its hosts is needed.
Lethal Chytridiomycosis Can Be Transmitted
through the Water in Which Infected Boreal
Toadlets Have Been Sitting
Experiment 2 confirms the findings of Marantelli et al.,
(2004) that direct body contact is not necessary for chy-
tridiomycosis to be transmitted. Within 21 days after
infection with 106 zoospores per day for 3 days, toadlets
shed sufficient zoospores into Holtfreter’s solution to kill
previously uninfected toadlets. A recent study indicates that
water can remain infective for up to 7 weeks after the
introduction of B. dendrobatidis (Johnson and Speare,
2003). Therefore, this pathogen, shed by an infected indi-
vidual in a breeding pond in June, could be transmitted to
an uninfected individual coming into contact with the
water at this location for much of the summer activity
period, even if no other infected amphibians were present.
18 Cynthia Carey et al.

Correlations of the Findings of This Study with
Observations on Populations of Boreal Toads in
Colorado Experiencing Mass Mortalities Associated
with B. dendrobatidis
These studies were conducted in environmental conditions
that differed considerably from those in the habitat of
boreal toads, and as a result, they cannot be used to predict
the survival times of boreal toads infected with B. dendro-
batidis in nature. The conditions in these experiments were
optimal for B. dendrobatidis: 23�C and pH 6.5 are within
the optimal ranges for growth of this fungus in laboratory
culture (Piotrowski et al., 2004). Furthermore, in this
study, toads were held in continuous contact with water.
Since toads can reinfect themselves with zoospores pre-
sumably only when they are in water or on a moist sub-
strate, constant contact with water undoubtedly promoted
reinfection and shortened the survival time relative to the
probable survival time if the toads had been periodically
allowed to dry their skin. In contrast, boreal toad body
temperatures, the pH of the water with which they come
into contact, and the amount of daily contact with water
vary considerably in their native habitat in ways that should
promote longer survival times of infected toads in the field
than in these experimental conditions. Body temperatures
of boreal toads fluctuate between near freezing and almost
30�C daily during their 3- to 4-month summer activity
period (Carey, 1978). Toads spend the other 7–8 months of
each year in hibernation under snow and ice, where their
body temperatures remain near freezing. The pH of the
water in which these toads come into contact varies widely
from location to location and throughout the year (Jones et
al., 2001). Finally, nonbreeding individuals of this species
spend at least part of each day during their summer active
period away from contact with water.
Despite these differences, the results of this study can
contribute to our understanding of some of the obser-
vations that we have made on boreal toad populations
experiencing mass mortalities associated B. dendrobatidis.
During these events, population sizes decrease either to
extinction or to very low levels (Carey, 1993; Muths et
al., 2003). We have noticed that larger toads (adults)
tended to persist in the population longer than smaller
ones (postmetamorphic juveniles) and that adult females
tended to survive more years than adult males following
the initial documentation of the presence of B. dendro-
batidis and subsequent mortality of most toads in the
population.
The range of body sizes used in this study was small
compared to the full range (up to more than 80 g) of body
masses found in the field (Carey et al., 2005). Because
approximately 4–6 years are required to grow to adult body
size, typical boreal toad populations have a broad spectrum
of body sizes. Although further study is necessary to verify
that body size has an effect on survival in large boreal toads,
our data are consistent with observations that juveniles and
smaller males died out before larger females in infected
populations (Carey, personal observation).
We presume that most boreal toads become infected
with B. dendrobatidis in aquatic environments, such as
breeding ponds, hibernacula, or moist substrate on which
adults sit and bask in the sun during the summer. This
pathogen, shed by an infected individual breeding in June,
could be transmitted to an uninfected individual coming
into contact with the water in this location for much of the
summer active period, even if no other infected amphibians
were present. The number of zoospores released in the field
from an infected boreal toad is not known, but we antici-
pate that a lightly infected individual would shed fewer
zoospores than a heavily infected one. Therefore, the dos-
age to which an uninfected animal would be exposed in the
field would vary with the number of infected individuals
present, the severity of their infections, the proximity of the
healthy individual to the infected one, the life span of
zoospores, and probably other factors.
Adult females exhibit several behaviors that could
minimize their risk of infection by B. dendrobatidis
zoospores and that support our observations that adult
females in infected populations may live longer generally
than adult males during an outbreak of B. dendrobatidis.
Adult males are likely to spend several weeks during
breeding each spring in continuous contact with water and
to have frequent skin-to-skin contact with other adult
males. The risk of infection in breeding ponds is minimized
for females because they spend less than 1 day at a breeding
site during egg laying and because they do not breed every
year (Carey et al., 2005). Females may also minimize the
risk of infection by hibernating more frequently as solitary
individuals than males, some of whom have been observed
to hibernate communally (Carey, personal observation).
Most boreal toad populations in Colorado became ex-
tinct in the late 1970s through early 1980s (Carey, 1993;
Corn et al., 1989). One hypothesis to explain why the few
relict populations survived these mass extinction events was
that they might have possessed some sort of immune resis-
tance against whatever pathogen might have caused the mass
Exposure of Boreal Toads to B. dendrobatidis 19

mortalities, thought at the time to be Aeromonas hydrophila
(Carey, 1993). Although the exact causes of the 1970s
mortalities will never be proven conclusively, the presence of
chytridiomycosis in museum specimens collected in Colo-
rado during the mid-1970s and the similarities between the
pattern of mass mortalities in the 1970s and 1990s (the latter
known to be linked to B. dendrobatidis; Muths et al., 2003)
suggest that this pathogen was the infectious agent (Carey et
al., 1999). Toadlets in this study originated from five pop-
ulations located in different mountain ranges in the Colo-
rado Rockies. Because individuals from each of these
populations proved to be susceptible to this pathogen under
the conditions of these experiments, we conclude that the
persistence of these relict populations was not likely related
to immunity to this pathogen.
Acknowledgments
This study was supported by a National Science Founda-
tion grant (Integrated Research Challenges in Environ-
mental Biology, DEB-0213851). We thank the Colorado
Division of Wildlife, especially Mark Jones, Chuck Loeffler,
and Craig Fetkavich, for facilitating use of animals from the
John W. Mumma Native Aquatic Species Restoration
Facility for this research. Joyce E. Longcore generously
provided B. dendrobatidis cultures and advice throughout
the course of these experiments. John Wood of Pisces
Molecular, Inc. made several recommendations concerning
PCR sampling, for which we are grateful.
REFERENCES
Alford RA, Richards SJ (1999) Global amphibian declines: aproblem in applied ecology. Annual Review of Ecology andSystematics 30:133–165
Annis SL, Dastoor F, Ziel H, Daszak P, Longcore JE (2004) ADNA-based assay identifies Batrachochytrium dendrobatidis inamphibians. Journal of Wildlife Diseases 40:420–428
Berger L, Speare R, Daszak P, Green DE, Cunningham AA, GogginCL, et al. (1998) Chytridiomycosis causes amphibian mortalityassociated with population declines in the rain forests of Aus-tralia and Central America. Proceedings of the National Academyof Sciences USA 95:9031–9036
Berger L, Speare R, Hines HB, Marantelli G, Hyatt AD, McDonaldKR, et al. (2004) Effect of season and temperature on mortalityin amphibians due to chytridiomycosis). Australian VeterinaryJournal 82:434–439
Bishop P (2000) Chytrid fungi identified from dying frogs in NewZealand. FrogLog 39:1–2
Bosch J, Martınez-Solano I, Garcıa-Parıs M (2001) Evidence of achytrid fungus infection involved in the decline of the commonmidwife toad (Alytes obstetricans) in protected areas of centralSpain. Biological Conservation 97:331–337
Bradley GA, Rosen PC, Sredl MJ, Jones TR, Longcore JE (2002)Chytridiomycosis in native Arizona frogs. Journal of WildlifeDiseases 38:206–212
Burnham KP, Anderson DR (2002) Model Selection and Multi-model Inference: A Practical Information-Theoretic Approach,New York: Springer
Carey C (1978) Factors affecting body temperatures of toads.Oecologia 35:197–219
Carey C (1993) Hypothesis concerning the causes of the disap-pearance of boreal toads from the mountains of Colorado.Conservation Biology 7:355–362
Carey C (2000) Infectious disease and worldwide declines ofamphibian populations, with comments on emerging diseases incoral reef organisms and in humans. Environmental HealthPerspectives 108(Suppl 1):1–8
Carey C, Bradford DF, Brunner JL, Collins JP, Davidson EW,Longcore JE, et al. (2003a) Biotic factors in amphibian popu-lation declines. In: Amphibian Decline: An Integrated Analysis ofMultiple Stressor Effects, Linder G, Krest SK, Sparling DW(editors), Pensacola, FL: Society of Environmental Toxicologyand Chemistry, pp 153–208
Carey C, Cohen N, Rollins-Smith L (1999) Amphibian declines:an immunological perspective. Developmental and ComparativeImmunology 23:459–472
Carey C, Corn PS, Jones MS, Livo LJ, Muths E, Loeffler CW (2005)Environmental and life history factors that limit recovery insouthern Rocky Mountain populations of boreal toads (Bufo boreaboreas). In: Status and Conservation of US Amphibians, Lannoo MJ(editor), Berkeley: University of California Press, pp 222–236
Carey C, Pessier AP, Peace AD (2003b) Pathogens, infectiousdisease, and immune defenses. In: Amphibian Conservation,Semlitch RD (editor), Washington DC: Smithsonian Institu-tion, pp 125–136
Corn PS, Stolzenburg W, Bury RB (1989) Acid precipitationstudies in Colorado and Wyoming: interim report of surveys ofmontane amphibians and water chemistry. US Fish and WildlifeService Biological Report 80:1–56
Daszak P, Berger L, Cunningham AA, Hyatt AD, Green DE,Speare R (1999) Emerging infectious diseases and amphibianpopulation declines. Emerging Infectious Diseases 5:735–748
Daszak P, Cunningham AA, Hyatt AD (2003) Infectious diseaseand amphibian population declines. Diversity and Distributions9:141–150
Davidson EW, Parris M, Collins JP, Longcore JE, Pessier AP,Brunner J (2003) Pathogenicity and transmission of chytridi-omycosis in tiger salamanders (Ambystoma tigrinum). Copeia2003:601–607
Fellers GM, Green DE, Longcore JE (2001) Oral chytridiomycosisin the mountain yellow-legged frog (Rana muscosa). Copeia2001:945–953
Goettl JP Jr (1997) Boreal toad (Bufo boreas) recovery plan(southern Rocky Mountain population), Denver: ColoradoDivision of Wildlife
Granoff A (1969) Viruses of amphibia. Current Topics in Micro-biology and Immunology 50:107–137
Green DE, Converse KA, Schrader AK (2002) Epizootiology ofsixty-four amphibian morbidity and mortality events in the
20 Cynthia Carey et al.

USA, 1996–2001. Annals of the New York Academy of Sciences969:323–339
Green DE, Kagarise Sherman C (2001) Diagnostic histologicalfindings in Yosemite toads (Bufo canorus) from a die-off in the1970s. Journal of Herpetology 35:92–103
Hanselmann R, Rodrıguez A, Lampo M, Fajardo-Ramos L,Aguirre AA, Kilpatrick AM, et al. (2004) Presence of anemerging pathogen of amphibians in introduced bullfrogsRana catesbeiana in Venezuela. Biological Conservation 120:115–119
James TY, Porter D, Leander CA, Vilgalys R, Longcore JE (2000)Molecular phylogenetics of the Chytridiomycota supports theutility of ultrastructural data in chytrid systematics. CanadianJournal of Botany 78:336–350
Jancovich JK, Davidson EW, Morado JF, Jacobs BL, Collins JP(1997) Isolation of a lethal virus from the endangered tigersalamander Ambystoma tigrinum stebbinsi. Diseases of AquaticOrganisms 31:161–167
Johnson ML, Speare R (2003) Survival of Batrachochytriumdendrobatidis in water: quarantine and disease control impli-cations. Emerging Infectious Diseases 9:922–925
Jones MS, Livo LJ, Holland AA (2001) Boreal Toad ResearchProgress Report 2000, Denver: Colorado Division of Wildlife
Keinath D, Bennett J (2000) Distribution and Status of the BorealToad (Bufo boreas boreas) in Wyoming, Cheyenne: U.S. Fish andWildlife Service
Lips KR, Green DE, Papendick R (2003) Chytridiomycosis in wildfrogs from southern Costa Rica. Journal of Herpetology 37:215–218
Longcore JE, Pessier AP, Nichols DK (1999) Batrachochytriumdendrobatidis gen. et sp. nov., a chytrid pathogenic toamphibians. Mycologia 91:219–227
Marantelli G, Berger L, Speare R, Keegan L (2004) Distribution ofthe amphibian chytrid Batrachochytrium dendrobatidis andkeratin during tadpole development. Pacific Conservation Biol-ogy 10:173–179
McDonald KR, Mendez D, Muller R, Freeman AB, Speare R(2005) Decline in the prevalence of chytridiomycosis in frogpopulations in North Queensland, Australia. Pacific Conserva-tion Biology 11:114–120
Morehouse EA, James TY, Ganley ARD, Vilgalys R, Berger L,Murphy P, et al. (2003) Multilocus sequence typing suggests thechytrid pathogen of amphibians is a recently emerged clone.Molecular Ecology 12:395–403
Muths E, Corn PS, Pessier AP, Green DE (2003) Evidence fordisease-related amphibian decline in Colorado. Biological Con-servation 110:357–365
Nichols DK, Lamirande EW, Pessier AP, Longcore JE (2001)Experimental transmission of cutaneous chytridiomycosis indendrobatid frogs. Journal of Wildlife Diseases 37:1–11
Nichols DK, Pessier AP, Longcore JE (1998) Cutaneous chy-tridiomycosis in amphibians: an emerging disease? In: Pro-ceedings of the American Association of Zoo Veterinarians/
American Association of Wildlife Veterinarians, Joint Conference,Media, Pennsylvania: American Association of Zoo Veteri-narians, pp 269–271
Pessier AP, Nichols DK, Longcore JE, Fuller MS (1999) Cutaneouschytridiomycosis in poison dart frogs (Dendrobates spp.) andWhite’s tree frogs (Litoria caerulea). Journal of VeterinaryDiagnostic Investigation 11:194–199
Piotrowski JS, Annis SL, Longcore JE (2004) Physiology of Ba-trachochytrium dendrobatidis, a chytrid pathogen of amphibians.Mycologia 96:9–15
Rachowicz LJ, Vredenburg VT (2004) Transmission of Batracho-chytrium dendrobatidis within and between amphibian lifestages. Diseases of Aquatic Organisms 61:75–82
Retallick RWR, McCallum H, Speare R (2004) Endemic infectionof the amphibian chytrid fungus in a frog community post-decline. Public Library of Science Biology 2:e351
Rome LC, Stevens ED, John-Alder HB (1992) The influence oftemperature and thermal acclimation on physiological function.In: Environmental Physiology of the Amphibians, Feder ME,Burggren WW (editors), Chicago: University of Chicago Press,pp 183–205
Ron SR, Duellman WE, Coloma LA, Bustamante MR (2003)Population decline of the Jambato toad Atelopus ignescens(Anura: Bufonidae) in the Andes of Ecuador. Journal of Her-petology 37:116–126
Ron SR, Merino A (2000) Amphibian declines in Ecuador: over-view and first report of chytridiomycosis from South America.FrogLog 42:2–3
Speare R, Berger L (2000) Global distribution of chytridiomycosisin amphibians. Available: http://www.jcu.edu.au/school/phtm/PHTM/frogs/chyglob.htm [accessed June 31, 2005]
Stuart SN, Chanson JS, Cox NA, Young BE, Rodrigues ASL, Fis-chman DL, et al. (2004) Status and trends of amphibian declinesand extinctions worldwide. Science 306:1783–1786
Taylor SK (2001) Mycoses. In: Amphibian Medicine and CaptiveHusbandry, Wright KM, Whitaker BR (editors), Malabar, FL:Krieger, pp 181–191
Taylor SK, Green DE, Wright KM, Whitaker BR (2001) Bacterialdiseases. In: Amphibian Medicine and Captive Husbandry, WrightKM, Whitaker BR (editors), Malabar, FL: Krieger, pp 159–179
Utah Division of Wildlife Resources (1998) Western toad. In:Inventory of Sensitive Vertebrate and Invertebrate Species: A Pro-gress Report, pp 159–161. Available: http://dwredc.nr.utah.gov/ucdc/ViewReports/vertrpt.pdf [accessed December 15, 2005]
Weldon C, du Preez LH, Hyatt AD, Muller R, Speare R (2004)Origin of the amphibian chytrid fungus. Emerging InfectiousDiseases 10:2100–2105
White GC, Burnham KP (1999) Program MARK: survival esti-mation from populations of marked animals. Bird Study46(Suppl):120–138
Woodhams D, Alford RA, Marantelli G (2003) Emerging diseaseof amphibians cured by elevated body temperature. Diseases ofAquatic Organisms 55:65–67
Exposure of Boreal Toads to B. dendrobatidis 21
Copyright © 2022 FDOKUMEN








