
The chemical biomarkers C2C, Coll2-1, and Coll2-1NO2 provide complementary information on type II...
11
ARTHRITIS & RHEUMATISM Vol. 56, No. 10, October 2007, pp 3336–3346 DOI 10.1002/art.22875 © 2007, American College of Rheumatology The Chemical Biomarkers C2C, Coll2-1, and Coll2-1NO 2 Provide Complementary Information on Type II Collagen Catabolism in Healthy and Osteoarthritic Mice L. G. Ameye, 1 M. Deberg, 2 M. Oliveira, 1 A. Labasse, 2 J. M. Aeschlimann, 1 and Y. Henrotin 2 Objective. Compared with wild-type (WT) mice, biglycan/fibromodulin double-deficient mice develop se- vere knee osteoarthritis. We undertook this study to compare type II collagen catabolism in the 2 geno- types and to compare the usefulness of 3 biomarkers of collagen degradation (C2C [also known as Col2- 3/4C long mono ] as well as the peptide Coll2-1 and its nitrated form, Coll2-1NO 2 ) for evaluating collagen ca- tabolism in vivo. Methods. In 15 WT mice and 15 biglycan/ fibromodulin double-deficient mice, we determined se- rum levels of C2C at ages 66 and 141 days, and we determined serum levels of Coll2-1 and Coll2-1NO 2 at ages 49, 81, 95, and 141 days. Expression of the biomar- kers in knee sections was examined using immuno- histochemistry. Results. The mean concentrations of C2C and Coll2-1 were higher in biglycan/fibromodulin double- deficient mice at all time points. For C2C and Coll2-1, the ratio of the serum concentration in biglycan/ fibromodulin double-deficient mice to that in WT mice (the double-deficient:WT ratio) was constant over time and was 1.63 and 1.15, respectively. In contrast, the double-deficient:WT ratio for Coll2-1NO 2 varied and, depending on age, was >1 or <1. No significant corre- lation was found between the expression of the different biomarkers, except for a weak, negative correlation between Coll2-1NO 2 and C2C. In both genotypes, anti- bodies to each biomarker labeled some fibroblasts in the tendons and menisci as well as chondrocytes above the tidemark in articular cartilage. Growth plates were unstained. For each biomarker, extracellular staining was limited to fibrocartilage areas in the tendons and menisci in all mice and was limited to some focal lesions of the cartilage in biglycan/fibromodulin double- deficient mice. Conclusion. The different double-deficient:WT ratios observed with C2C, Coll2-1, and Coll2-1NO 2 in the absence of any correlation between the expression of the 3 biomarkers indicate that these biomarkers give complementary, rather than redundant, information about in vivo type II collagen catabolism. Osteoarthritis (OA) is one of the leading causes of pain and disability in the elderly. Its high prevalence and its moderate-to-severe impact on daily life pose a significant public health problem (1). Despite intensive research, a cure remains elusive for this disease with important socioeconomic consequences. The intrinsic difficulty in finding a cure is compounded by the low sensitivity of diagnostic and monitoring tools. For exam- ple, the destruction of articular cartilage is an important feature of OA, and radiographic change in joint space width (JSW) is widely recognized as the gold standard end point for measuring destruction of articular carti- lage and OA progression. However, this end point is an indirect measure of cartilage integrity, since cartilage is invisible on radiographs and its integrity must be in- ferred from the spacing between bones. Furthermore, change in JSW has a low signal-to-noise ratio; annual changes are only 0.1–0.2 mm in OA patients (2), close to the precision error of repetitive measurements. Alterna- 1 L. G. Ameye, PhD, M. Oliveira, MSc, J. M. Aeschlimann, PhD: Nestle ´ Research Center, Lausanne, Switzerland; 2 M. Deberg, PhD, A. Labasse, MSc, Y. Henrotin, PhD: University of Lie `ge, Lie `ge, Belgium. Drs. Deberg and Henrotin hold patents for immunoassays for the chemical biomarkers Coll2-1 and Coll2-1NO 2 and receive royalties from these patents. Address correspondence and reprint requests to L. G. Ameye, PhD, Nutrition and Health Department, Nestle ´ Research Center, Vers-chez-les-Blanc, 1000 Lausanne 26, Switzerland. E-mail: laurent. [email protected]. Submitted for publication January 24, 2007; accepted in revised form June 12, 2007. 3336
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of The chemical biomarkers C2C, Coll2-1, and Coll2-1NO2 provide complementary information on type II...

ARTHRITIS & RHEUMATISMVol. 56, No. 10, October 2007, pp 3336–3346DOI 10.1002/art.22875© 2007, American College of Rheumatology
The Chemical Biomarkers C2C, Coll2-1, and Coll2-1NO2Provide Complementary Information on
Type II Collagen Catabolism inHealthy and Osteoarthritic Mice
L. G. Ameye,1 M. Deberg,2 M. Oliveira,1 A. Labasse,2 J. M. Aeschlimann,1 and Y. Henrotin2
Objective. Compared with wild-type (WT) mice,biglycan/fibromodulin double-deficient mice develop se-vere knee osteoarthritis. We undertook this study tocompare type II collagen catabolism in the 2 geno-types and to compare the usefulness of 3 biomarkersof collagen degradation (C2C [also known as Col2-3/4Clong mono] as well as the peptide Coll2-1 and itsnitrated form, Coll2-1NO2) for evaluating collagen ca-tabolism in vivo.
Methods. In 15 WT mice and 15 biglycan/fibromodulin double-deficient mice, we determined se-rum levels of C2C at ages 66 and 141 days, and wedetermined serum levels of Coll2-1 and Coll2-1NO2 atages 49, 81, 95, and 141 days. Expression of the biomar-kers in knee sections was examined using immuno-histochemistry.
Results. The mean concentrations of C2C andColl2-1 were higher in biglycan/fibromodulin double-deficient mice at all time points. For C2C and Coll2-1,the ratio of the serum concentration in biglycan/fibromodulin double-deficient mice to that in WT mice(the double-deficient:WT ratio) was constant over timeand was �1.63 and �1.15, respectively. In contrast, thedouble-deficient:WT ratio for Coll2-1NO2 varied and,depending on age, was >1 or <1. No significant corre-lation was found between the expression of the different
biomarkers, except for a weak, negative correlationbetween Coll2-1NO2 and C2C. In both genotypes, anti-bodies to each biomarker labeled some fibroblasts in thetendons and menisci as well as chondrocytes above thetidemark in articular cartilage. Growth plates wereunstained. For each biomarker, extracellular stainingwas limited to fibrocartilage areas in the tendons andmenisci in all mice and was limited to some focal lesionsof the cartilage in biglycan/fibromodulin double-deficient mice.
Conclusion. The different double-deficient:WTratios observed with C2C, Coll2-1, and Coll2-1NO2 inthe absence of any correlation between the expression ofthe 3 biomarkers indicate that these biomarkers givecomplementary, rather than redundant, informationabout in vivo type II collagen catabolism.
Osteoarthritis (OA) is one of the leading causesof pain and disability in the elderly. Its high prevalenceand its moderate-to-severe impact on daily life pose asignificant public health problem (1). Despite intensiveresearch, a cure remains elusive for this disease withimportant socioeconomic consequences. The intrinsicdifficulty in finding a cure is compounded by the lowsensitivity of diagnostic and monitoring tools. For exam-ple, the destruction of articular cartilage is an importantfeature of OA, and radiographic change in joint spacewidth (JSW) is widely recognized as the gold standardend point for measuring destruction of articular carti-lage and OA progression. However, this end point is anindirect measure of cartilage integrity, since cartilage isinvisible on radiographs and its integrity must be in-ferred from the spacing between bones. Furthermore,change in JSW has a low signal-to-noise ratio; annualchanges are only 0.1–0.2 mm in OA patients (2), close tothe precision error of repetitive measurements. Alterna-
1L. G. Ameye, PhD, M. Oliveira, MSc, J. M. Aeschlimann,PhD: Nestle Research Center, Lausanne, Switzerland; 2M. Deberg,PhD, A. Labasse, MSc, Y. Henrotin, PhD: University of Liege, Liege,Belgium.
Drs. Deberg and Henrotin hold patents for immunoassays forthe chemical biomarkers Coll2-1 and Coll2-1NO2 and receive royaltiesfrom these patents.
Address correspondence and reprint requests to L. G. Ameye,PhD, Nutrition and Health Department, Nestle Research Center,Vers-chez-les-Blanc, 1000 Lausanne 26, Switzerland. E-mail: [email protected].
Submitted for publication January 24, 2007; accepted inrevised form June 12, 2007.
3336

tive, more powerful surrogate markers of OA are there-fore needed.
Magnetic resonance imaging (MRI) is a promis-ing technique, since it allows direct quantification ofcartilage volume and evaluation of noncartilaginoustissues, such as ligaments and menisci (3). Biochemicalbiomarkers are other interesting alternatives for thediagnosis, prognosis, and treatment monitoring of OA(for recent reviews, see refs. 4 and 5), since they aremore dynamic and often have a more suitable signal-to-noise ratio than imaging techniques (6).
Collagen molecules have been frequently studiedas potential biochemical markers for cartilage metabo-lism. These molecules, organized into fibrils, make up�50% of the organic dry weight of articular cartilage(7). In adult cartilage, 90% of collagen is type IIcollagen, which is mainly composed of 3 identicalpolypeptide (�1[II]) chains. These chains are synthe-sized as procollagen precursors whose carboxy-terminal,nonhelical propeptides are cleaved enzymatically andused as markers of collagen synthesis (8). Once thetrimmed type II collagen molecules are released extra-cellularly, they assemble into fibrils and form a network,which has high tensile strength and protects the cartilageagainst the swellings that result from hydration of neg-atively charged proteoglycan aggregates in the extracel-lular matrix. Extracellular collagen crosslinking furtherincreases the tensile strength of the collagen network.
The turnover of type II collagen in mature carti-lage is limited (9), and damage to the collagen network,a hallmark of many joint diseases including OA (10–13),is thought to result in the irreversible loss of cartilagestructure integrity. In OA articular cartilage in vivo,intact type II collagen molecules are initially cleavedinto three-quarter–length and one-quarter–length piecesby various matrix metalloproteinases (MMPs), includingMMP-13 (14–16). The initial cleavage unwinds thecollagen triple helix and is followed by further enzymaticcleavage. Biomarkers of collagen catabolism includeneoepitopes generated by the cleavage of the collagenmolecule (13,17), “denaturation” epitopes that resultfrom unwinding and subsequent cleavage of the triplehelix (10,18,19), and collagen fragments containingglucosyl-galactosyl pyridinoline crosslinks (20).
Antibodies to biomarkers of types I and II colla-gen degradation (C1,2C) and type II collagen degrada-tion (C2C) recognize a neoepitope located at thecarboxy-terminal end of the initial MMP-generatedthree-quarter–length fragment (13). The names C1,2Cand C2C denote the collagen specificity of the antibod-ies: C1,2C recognizes both type I and type II collagen,whereas C2C is specific to type II collagen alone. The
specificity of the 2 antibodies is linked to the length ofthe neoepitopes that are recognized. Indeed, the C1,2Cantibody is also called Col2-3/4Cshort because it recog-nizes a short, 7–amino acid epitope that is shared by typeI and type II collagen, while C2C is sometimes calledCol2-3/4Clong mono because it recognizes a longer epitopecomprising the 7–amino acid C1,2C epitope plus anadditional glutamate residue that restricts its specificityto type II collagen (21).
A correlation between the expression of thesemarkers and disease activity has been reported in anumber of studies. The synovial fluid level of C2C issignificantly increased following joint instability in a dogmodel of OA (22) and partial-thickness cartilage lesionin a sheep model (23). The serum levels of C2C/C-propeptide of type II collagen, which indicate the ratioof type II collagen degradation to synthesis, are alsoelevated in OA-susceptible compared with OA-resistantguinea pigs (24). In humans, calcitonin has been shownto reduce the serum levels of C2C and alleviate OAsymptoms (25). Furthermore, positive correlations havebeen found between the MRI T2 parameter and serumlevels of C2C in OA knees (26). However, no correlationhas been observed between baseline C2C or C1,2C levelsand subsequent joint space narrowing in 120 obesewomen with unilateral knee OA (27).
The peptide Coll2-1 (108HRGYPGLDG116) andits nitrated form, Coll2-1NO2 (108HRGY[NO2]PGLDG116), are located in the triple helix of the type IIcollagen molecule and are 2 of the most recently usedbiomarkers of collagen degradation (19). In healthysubjects, the serum concentration of Coll2-1 remainsstable throughout life, while the serum concentration ofColl2-1NO2 is higher in young adults (ages 26–30 years).Both Coll2-1 and Coll2-1NO2 concentrations are ele-vated in the serum of patients with primary knee OAand early rheumatoid arthritis (RA) compared with thatof healthy subjects (19). Moreover, the increase inurinary Coll2-1 and Coll2-1NO2 over a 1-year period ispredictive of the radiographic progression of OA (28).
There is increasing interest in the clinical rele-vance of chemical biomarkers for OA. Multiple case–control and cohort studies recently investigated theusefulness of such biomarkers for the clinical manage-ment of OA, and all recent structure-modifying clinicaltrials included biomarkers as secondary end points.However, the limited availability of biologic samples andthe high cost of analysis restrict the number of biomar-kers included in individual studies. Since collagen degra-dation is an important feature of OA, many studiesinclude at least 1 biomarker of type II collagen catabo-lism, but few (19,29) include multiple markers of this
BIOMARKERS OF TYPE II COLLAGEN CATABOLISM 3337
process. Instead, simultaneous monitoring of other bio-logic processes, such as collagen synthesis, bone resorp-tion, cytokine production, or turnover of other matrixmolecules, is often preferred.
Thus, although several biomarkers of type IIcollagen catabolism are now available, there are fewstudies that directly compare the relevance of each ofthese biomarkers for OA research or monitoring. Inparticular, it is currently unknown whether these bio-markers provide redundant information or whether theyprovide complementary, independent knowledge aboutcollagen catabolism. Without such comparative studies,the choice of a biomarker is random, dictated by eco-nomics or personal preferences, or is based on a limitednumber of reported studies. Longitudinal comparison ofmultiple biomarkers in well-characterized animal mod-els of OA (for a recent review, see ref. 30) could providemuch needed insight into the pros and cons of eachbiomarker for specific uses in OA research. Such animalstudies would allow a scientific, evidence-based selectionthat would ultimately improve the design and success ofstudies in humans.
To this end, the current study compared theusefulness of C2C, Coll2-1, and Coll2-1NO2 in theevaluation of collagen catabolism in healthy and OAmice. The evolution of these 3 biomarkers was comparedin wild-type (WT) mice and biglycan/fibromodulindouble-deficient mice between the ages of 7 weeks and 5months. The latter not only develop severe prematureknee OA (31), but they also develop a mild form of OAin their temporomandibular joints (32). Knee articularcartilage from these deficient mice does not display anyvisible structural damage at 1 month of age, but by 2months of age, small clefts, horizontal cartilage fibrilla-tion, and cracks are observed (31), and by 5 months ofage, the destruction of the articular cartilage reaches thetidemark throughout a large proportion of the joint. Insome areas, the destruction is so pronounced that itleads to eburnation and underlying subchondral sclerosis(Ameye LG: unpublished observations).
Other joint tissues besides articular cartilage arealso affected by the absence of biglycan and fibromodu-lin (31,33). Tendons of biglycan/fibromodulin double-deficient mice are less stiff, are composed of thinnercollagen fibrils, and develop an ectopic ossification thatresults in severe gait impairment. Taken together, thephenotype of biglycan/fibromodulin double-deficientmice is similar to that of STR/Ort mice, which alsodevelop OA. As in guinea pig OA (34) and human OA(for a discussion, see ref. 31), the structural weaknessesof tendons and ligaments are thought to be the origin ofOA in these deficient mice.
MATERIALS AND METHODS
Generation of biglycan and fibromodulin double-deficient mice. Mice deficient in biglycan and fibromodulinwere generated by gene targeting in embryonic stem cells, asdescribed previously (31,35,36). Since early observations sug-gested that the OA phenotype was more severe in males thanin females, only male mice were analyzed in this study. Micewere housed under specific pathogen–free conditions. Theexperiments were approved by the Swiss Cantonal InstitutionalAnimal Care and Use Committee and were in accordance withthe Swiss Law of Animal Protection. We studied 7-week-old
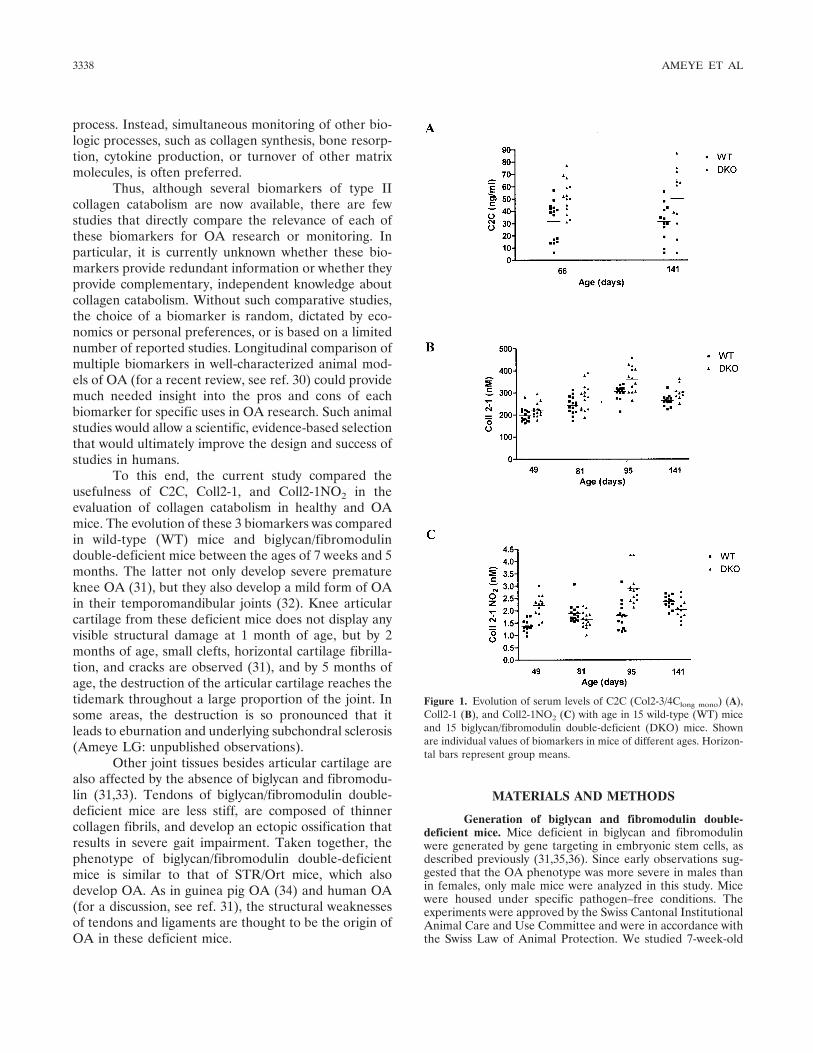
Figure 1. Evolution of serum levels of C2C (Col2-3/4Clong mono) (A),Coll2-1 (B), and Coll2-1NO2 (C) with age in 15 wild-type (WT) miceand 15 biglycan/fibromodulin double-deficient (DKO) mice. Shownare individual values of biomarkers in mice of different ages. Horizon-tal bars represent group means.
3338 AMEYE ET AL
C57BL/6 WT and biglycan/fibromodulin double-deficient mice(n � 15 per group) that were fed an AIN-93M diet.
Measurement of type II collagen biomarkers in serum.Blood was collected serially from 15 WT and 15 biglycan/fibromodulin double-deficient mice at ages 49, 66, 81, 95, and141 days. To facilitate blood sampling and induce vasodilation,mice were placed for a few minutes under a heating lamp, anda few drops of blood were collected in polypropylene tubesafter a small tail incision was made. Blood was allowed tocoagulate at room temperature for 2 hours and was thencentrifuged at 10,000g for 2 minutes. Sera were collected andstored frozen at �80°C until analysis. Quantification of C2Clevels in serum was performed directly by Ibex (Montreal,Quebec, Canada), the commercial manufacturer of the C2Ckit. Competitive enzyme-linked immunosorbent assays specificfor Coll2-1 and Coll2-1NO2, with limits of detection of 17mmoles/liter and 25 pmoles/liter, respectively, were used aspreviously described (19). The intra- and interassay coeffi-cients of variation were �10%, and the dilution curves wereparallel to the standard curve for both assays. The analyticrecoveries were, on average, 104.7% and 121.9% for theColl2-1 and Coll2-1NO2 assays, respectively.
Histology and immunohistochemistry. Knee sectionswere prepared for histology as described previously (31). Forimmunohistochemistry, paraffin-embedded sections weredeparaffinized with xylene and then rehydrated in a series ofgraded ethanol solutions. Endogenous peroxidase activity wasblocked by incubating the sections in a freshly preparedsolution of 0.5% (volume/volume) H2O2 in absolute ethanolfor 10 minutes at room temperature. Glycosaminoglycans wereremoved using 0.4 units/liter proteinase-free chondroitinaseABC (Sigma, Bornem, Belgium) in 0.1M Tris HCl, pH 8.0, for30 minutes at 37°C. Nonspecific binding of antibodies wasblocked by incubating with 100 �l of 1% normal goat serum(Jackson ImmunoResearch, Lucron Bioproducts, De Pinte,Belgium) in 50 mM Tris, 138 mM NaCl (Tris buffered saline[TBS]), pH 7.6, for 30 minutes. All subsequent incubationswere performed at room temperature in a humidified cham-ber.
First, sections were incubated for 2 hours with 100 �lof the specific primary antibodies D3 (1:200), D37 (1:100), orC1,2C (1:200) diluted in TBS containing 1% normal goatserum. Sections incubated with normal rabbit serum instead ofthe primary antibody served as negative controls. Of note, anantibody against C1,2C was used instead of an anti-C2Cantibody because the latter is not suitable for immunohisto-chemistry. After rinsing, sections were incubated with 100 �lhorseradish peroxidase–conjugated goat anti-rabbit IgG
(Dako, Heverlee, Belgium) for 30 minutes, and peroxidase wasvisualized using 100 �l of 3,3�-diaminobenzidine (Sigma) for15 minutes. Finally, the sections were counterstained withhematoxylin for 1 minute.
The histologic data presented in this report are from 3different animals of the same genotype. Serial sections ofwhole joints were obtained for each animal, and staining wasperformed on nonconsecutive serial sections in order to exam-ine expression throughout the joint and to ensure that theobservations were genuine and not due to artifacts. To mini-mize experimental variability, tissues from each animal groupwere processed in parallel.
Statistical analysis. Differences between genotypesand ages were assessed using a two-way analysis of variance(ANOVA). Student’s unpaired t-test was used to compareexpression in the 2 animal groups at each age. Based on theindividual values from both genotypes, 2 � 2 correlationsbetween biomarkers were calculated. P values less than 0.05were considered significant.
RESULTS
Variation of serum levels of C2C, Coll2-1, andColl2-1NO2 with age and genotype. The serum concen-trations of 3 biomarkers of type II collagen catabolism(C2C, Coll2-1, and Coll2-1NO2) were examined in WTand biglycan/fibromodulin double-deficient mice be-tween the ages of 7 weeks and 5 months (Figure 1 andTable 1). A two-way ANOVA revealed an effect ofgenotype on all 3 biomarkers (P � 10�4 for C2C, P �10�4 for Coll2-1, and P � 10�4 for Coll2-1NO2) and aneffect of age on the levels of Coll2-1 and Coll2-1NO2(P � 10�4). A significant interaction between age andgenotype was observed for Coll2-1NO2 (P � 10�4).
The mean serum concentrations of C2C andColl2-1 were higher in the biglycan/fibromodulindouble-deficient mice than in the WT mice at all timepoints (see Table 1). For these 2 biomarkers, the ratio ofthe serum concentration in biglycan/fibromodulindouble-deficient mice to that in WT mice (the double-deficient:WT ratio), which was calculated from the meanvalues in Table 1, remained relatively constant at thedifferent ages examined (see Figure 1) and were �1.63
Table 1. Concentrations of C2C, Coll2-1, and Coll2-1NO2 in serum from wild-type (WT) mice and biglycan/fibromodulin double-deficient mice(n � 15 per group)*
Age ofmice, days
C2C, ng/ml Coll2-1, nM Coll2-1NO2, nM
WT mice Double-deficient mice P WT mice Double-deficient mice P WT mice Double-deficient mice P
49 – – – 200 � 30 222 � 31 0.06 1.38 � 0.22 2.22 � 0.42 0.00066 32 � 16 52 � 14 0.001 – – – – – –81 – – – 245 � 40 281 � 61 0.06 1.90 � 0.38 1.63 � 0.32 0.05395 – – – 305 � 32 362 � 60 0.005 1.81 � 0.54 2.89 � 0.67 0.000141 30 � 16 50 � 26 0.03 264 � 24 303 � 35 0.008 2.37 � 0.24 2.04 � 0.41 0.027
* Values are the mean � SD. C2C � Col2-3/4Clong mono.
BIOMARKERS OF TYPE II COLLAGEN CATABOLISM 3339

and �1.15 for C2C and Coll2-1, respectively. Whengroup medians were used to calculate the double-deficient:WT ratio instead of group means, the ratio forColl2-1 remained constant at 1.15, but the ratio for C2Cvaried with age and was 1.30 at age 66 days and 2.03 atage 141 days. In both genotypes, the mean concentra-tions of C2C were unchanged at ages 66 days and 141days; however, the levels of Coll2-1 increased with age,peaking at 95 days and declining thereafter (Figure 1).
Mean serum concentrations of Coll2-1NO2 in-creased progressively with age in WT mice but varied inbiglycan/fibromodulin double-deficient mice, withhigher values being observed at ages 49 days and 95 daysthan at ages 81 days and 141 days. Consequently, and incontrast to the constant double-deficient:WT ratios forC2C and Coll2-1, the double-deficient:WT ratio forColl2-1NO2 showed a “pseudo-cyclic” variation as themice aged. More specifically, the double-deficient:WTratio for Coll2-1NO2 was �0.86 at ages 81 days and 141days and �1.60 at ages 49 days and 95 days (see Table1). Similar results were obtained when group medianswere used to calculate the Coll2-1NO2 double-deficient:WT ratio. Correlations between any 2 of the 3biomarkers were poor and not significant, although aweak negative correlation at the limit of statisticalsignificance was observed between the levels of Coll2-1NO2 and C2C (r2 � 0.096, P � 0.049) (Figure 2).
C1,2C, Coll2-1, and Coll2-1NO2 expression inthe knees of healthy, WT mice and biglycan/fibromodulin double-deficient mice. To gain furtherinsight into the generation of these biomarkers of colla-gen catabolism, immunohistochemical staining of kneejoint sections was performed. Since the antibody to C2Cis not suitable for immunohistochemistry, an antibody toC1,2C was used. Both of these antibodies recognize thesame primary catabolic neoepitope, but while anti-C2Cis specific for type II collagen, anti-C1,2C recognizesboth type I and type II collagen epitopes.
Similar intracellular staining was obtained withantibodies to C1,2C (Figure 3), Coll2-1 (Figure 4), andColl2-1NO2 (Figure 5). In both animal groups, the 3antibodies labeled cells in the menisci, some tenocytes,and a fair proportion of the chondrocytes above thetidemark in the articular cartilage. The intracellularstaining of the chondrocytes was more prominent in theWT mice. Indeed, almost all the superficial chondro-cytes in dense rows that bordered the intact, articularcartilage surface were positive (Figures 3G, 4G, and5G). In biglycan/fibromodulin double-deficient mice,articular cartilage was already eroded in significant partsof the joint below the tidemark. In the areas where thearticular cartilage remained above the tidemark, intra-
cellular staining was observed in a significant proportionof the chondrocytes (see Figures 3C and E, 4B and D,and 5C and E). Chondrocytes below the tidemark werenot labeled in the knees from either animal group.
Extracellular expression of the biomarkers wassimilar, although the intensity of Coll2-1NO2 stainingwas lower than that of Coll2-1 or C1,2C staining. Dif-ferences in the intensity of intracellular staining werenot observed. In both genotypes, extracellular staining
Figure 2. Correlation between serum levels of C2C, Coll2-1, andColl2-1NO2 in wild-type mice and biglycan/fibromodulin double-deficient mice at age 141 days.
3340 AMEYE ET AL

was observed in some areas of the tendon and menisci,but staining was more developed in the hypertrophicmenisci and tendons of biglycan/fibromodulin double-deficient mice. Indeed, in the knees from WT animals,extracellular staining in tendons was mainly restricted to
their extremities and the tendon insertion site, while inbiglycan/fibromodulin double-deficient mice, a morewidespread staining was observed (compare Figures 3Fand 4F with Figures 3D and 4E). In addition, some,but not all, articular cartilage lesions in biglycan/
Figure 3. Immunostaining of C1,2C (Col2-3/4Cshort) in coronal knee sections from 5-month-old DKO mice (A–E) and WT mice (F–I). Arrowheadsindicate staining of the extracellular matrix. Note the extracellular staining of cartilage lesions in DKO mice (B and C) as well as in the insertionsites of the tendons in WT mice (F) and in the hypertrophic meniscus and tendons of DKO mice (D and E). Vertical arrows indicate intracellularstaining of chondrocytes, tenocytes, and cells in the meniscus (B, D, E, G, and H). The growth plates were negative (diagonal arrows) (A and G).White arrows indicate nonspecific labeling in the bone marrow due to the high endogenous peroxidase activity of granulocytes and red blood cells.As a negative control, normal rabbit serum was used instead of the primary anti-C1,2C antibody (I). Bars in A, D, and I � 800 �m; bars in B andH � 200 �m; bar in C � 100 �m; bar in E � 400 �m; bar in F � 600 �m; bar in G � 300 �m. See Figure 1 for definitions.
BIOMARKERS OF TYPE II COLLAGEN CATABOLISM 3341

fibromodulin double-deficient mice were stained extra-cellularly, while those in WT animals had no extracellu-lar staining. No extra- or intracellular staining was
observed in either genotype in the bone growth plate(Figures 3A, 4A, and 5A).
Staining was specific for all tissues except bone
Figure 4. Immunostaining of Coll2-1 in coronal knee sections from 5-month-old DKO mice (A–E) and WT mice (F–J). Arrowheads indicatestaining of the extracellular matrix. Extracellular staining was observed in the cartilage lesions of DKO mice (A, C, and E) as well as in the insertionsites of the tendons in WT mice (F, H, and I) and in the hypertrophic meniscus and tendons of DKO mice (D and E). B is an enlargement of a partof A and depicts the extracellular staining in the cartilage lesions of the DKO mice. G is an enlargement of a part of F and depicts the intracellularstaining of superficial chondrocytes in intact articular cartilage. Horizontal and vertical arrows indicate intracellular staining of chondrocytes,tenocytes, and cells in the meniscus. The growth plate and some chondrocytes were negative (diagonal arrows). Small white or black arrows indicatenonspecific labeling in the bone marrow due to the high endogenous peroxidase activity of granulocytes and red blood cells. As a negative control,normal rabbit serum was used instead of the primary anti–Coll2-1 antibody (J). Bars � 400 �m in A, E, F, and H; bars � 150 �m in B, G, and J;bars � 300 �m in C and D; bar � 200 �m in I. See Figure 1 for definitions.
3342 AMEYE ET AL
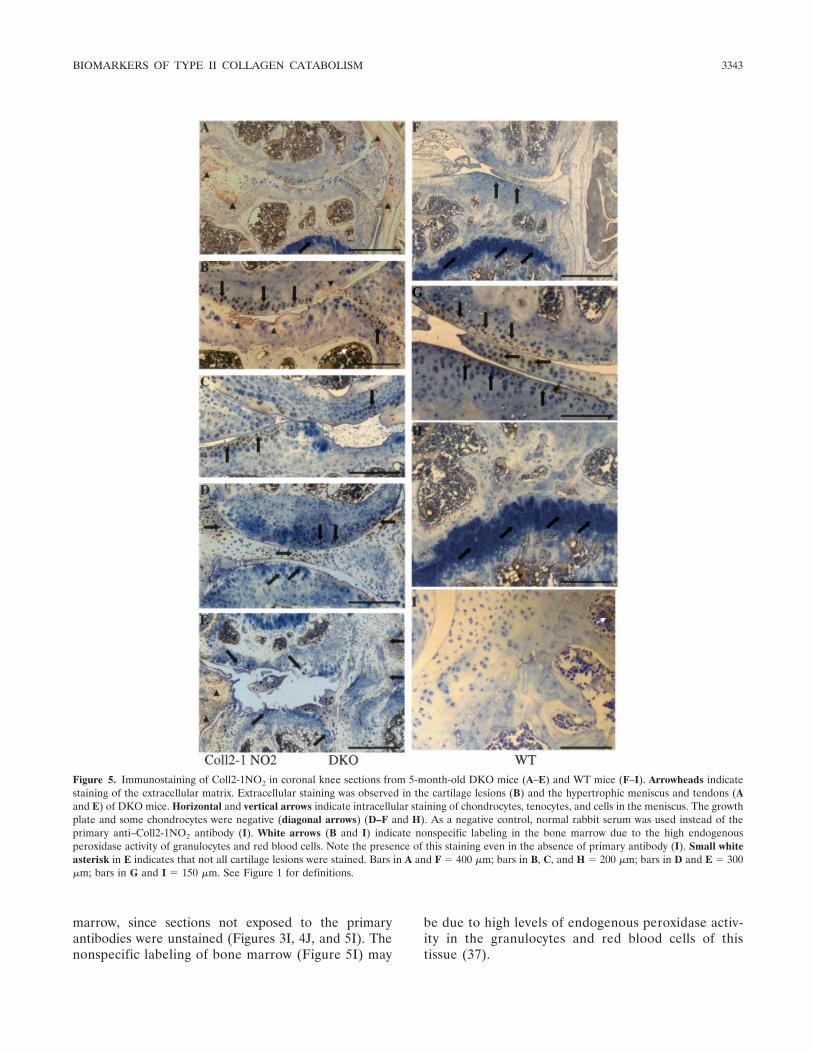
marrow, since sections not exposed to the primaryantibodies were unstained (Figures 3I, 4J, and 5I). Thenonspecific labeling of bone marrow (Figure 5I) may
be due to high levels of endogenous peroxidase activ-ity in the granulocytes and red blood cells of thistissue (37).
Figure 5. Immunostaining of Coll2-1NO2 in coronal knee sections from 5-month-old DKO mice (A–E) and WT mice (F–I). Arrowheads indicatestaining of the extracellular matrix. Extracellular staining was observed in the cartilage lesions (B) and the hypertrophic meniscus and tendons (Aand E) of DKO mice. Horizontal and vertical arrows indicate intracellular staining of chondrocytes, tenocytes, and cells in the meniscus. The growthplate and some chondrocytes were negative (diagonal arrows) (D–F and H). As a negative control, normal rabbit serum was used instead of theprimary anti–Coll2-1NO2 antibody (I). White arrows (B and I) indicate nonspecific labeling in the bone marrow due to the high endogenousperoxidase activity of granulocytes and red blood cells. Note the presence of this staining even in the absence of primary antibody (I). Small whiteasterisk in E indicates that not all cartilage lesions were stained. Bars in A and F � 400 �m; bars in B, C, and H � 200 �m; bars in D and E � 300�m; bars in G and I � 150 �m. See Figure 1 for definitions.
BIOMARKERS OF TYPE II COLLAGEN CATABOLISM 3343

DISCUSSION
The expression of 3 biomarkers of type II colla-gen catabolism, C2C, Coll2-1, and Coll2-1NO2, wasexamined longitudinally in the serum and in knee sec-tions of 15 WT mice and 15 OA biglycan/fibromodulindouble-deficient mice. This is the first study to comparethese 3 biomarkers directly. Blood was collected frommice 5 times between the ages of 7 weeks and 5 months,but the limited amount of blood available from live miceprevented the measurement of all 3 biomarkers at eachtime point except 141 days, when mice were killed. Assuch, serum levels of C2C were measured in 66- and141-day-old mice, and serum levels of Coll2-1 andColl2-1NO2 were measured in 49-, 81-, 95-, and 141-day-old mice. Since our goal was to monitor collagen catab-olism in articular cartilage, blood was not collectedbefore the age of 7 weeks, since younger animals arerapidly growing and since growth plate cartilage metab-olism contributes importantly to the overall type IIcollagen metabolism. The last blood sample was col-lected at age 5 months, when biglycan/fibromodulindouble-deficient mice already display severe knee OAwith important articular cartilage destruction (31). Atthis age, the knee growth plate is unlikely to contributeto the serum levels of the biomarkers, since it does notstain positively with any of the 3 antibodies.
Serum concentrations of the 3 molecules suggestthat C2C and Coll2-1, but not Coll2-1NO2, are usefulbiomarkers of collagen degradation in OA mice. Indeed,double-deficient:WT ratios �1 for C2C and Coll2-1reflect the expected increase in type II collagen catabo-lism in OA biglycan/fibromodulin double-deficient mice.In contrast, this ratio for Coll2-1NO2 varied and, de-pending on age, was �1 or �1. The distinct evolution ofserum concentrations of Coll2-1 and Coll2-1NO2 withage suggests that the production of these 2 epitopesrepresents different biologic processes. Moreover, al-though Coll2-1NO2 is the nitrated form of Coll2-1, itsconcentration is not simply an indication of the totalamount of Coll2-1 epitopes present. Thus, our dataindicate that while a study of Coll2-1 expression allowsthe extent of type II collagen catabolism to be moni-tored, examination of Coll2-1NO2 levels measures adistinct aspect of this catabolism that is not necessarilydirectly correlated with the total amount of type IIcollagen catabolism in the organism.
Since nitration of Coll2-1 requires oxidativestress, the concentration of Coll2-1NO2 may indicate theextent of oxidative stress in articular cartilage and thelevel of inflammation in the synovium. In support of thishypothesis, Coll2-1NO2 concentrations are higher in RA
than in OA and correlate with C-reactive protein levels(19). Therefore, the cyclic variations in Coll2-1NO2levels that we observed in biglycan/fibromodulin double-deficient mice may reflect transient increases in synovialinflammation or bursts of oxidative stress related tocartilage degradation. However, further research is re-quired to confirm this hypothesis.
In contrast to healthy human subjects, in whomconcentrations of Coll2-1NO2 are higher in young adults(ages 26–30 years) than in older subjects (ages 36–65years) (19), in WT mice, serum levels of Coll2-1NO2increased with age. The young age of the mice in ourstudy may explain this discrepancy. The serum levels ofColl2-1 increased up to age 95 days in both WT andbiglycan/fibromodulin double-deficient mice and de-clined thereafter. This age-related effect is not related tothe development of OA, since it is observed both inbiglycan/fibromodulin double-deficient mice, which de-velop severe and premature knee OA, and in healthyWT mice. Instead, this effect probably reflects changesin the intensity of the overall type II collagen turnover asthe mice age. If true, this suggests that turnover of typeII collagen peaks in mice at �3 months of age, later thanthe long bone growth spurt (38).
The double-deficient:WT ratio for C2C washigher than that for Coll2-1 (�1.63 versus �1.15),suggesting that C2C is a more sensitive biomarker thanColl2-1 for monitoring increased type II collagen catab-olism. This difference in magnitude and the poor andnonsignificant correlation between the expression of the2 biomarkers suggest that C2C and Coll2-1 levels reflectdifferent biologic processes and/or stages in type IIcollagen degradation and that they provide complemen-tary rather than redundant information on type IIcollagen catabolism.
Antibodies to C2C recognize the first catabolicneoepitope generated during the degradation of thetriple helix. The C2C epitope is located three-quarters ofthe total length of the collagen molecule away from theN-telopeptide (13). Conversely, epitope recognized bythe anti–Coll2-1 antibody is located in the triple helix,relatively close to the N-telopeptide (19). Unmasking ofthe Coll2-1 epitope may require unwinding of the triplehelix and/or multiple enzymatic digestion of the collagenmolecule after the generation of the C2C neoepitope bythe first MMP. Thus, C2C concentrations may representearly stages of type II collagen degradation, whileColl2-1 may be a late degradation product. Kineticstudies of collagen degradation are required to elucidatethis process.
In any event, the independent behavior of C2Cand Coll2-1 expression suggests that these 2 biomarkers
3344 AMEYE ET AL

could both be examined in clinical investigations of OA,as has recently been done for the 622–632 peptidederived from the �1 chain of human type II collagen(HELIX-II) and C-terminal crosslinking telopeptide oftype II collagen (CTX-II) (29). A small case studyshowed that, independently of each other, HELIX-IIand CTX-II levels are associated with rapidly destructivehip OA and that measurement of both molecules im-proves diagnosis and the monitoring of disease progres-sion in patients.
Tissue expression of the 3 biomarkers was verysimilar. As expected, all 3 molecules were expressed inthe extracellular matrix, around superficial lesions in thearticular cartilage of biglycan/fibromodulin double-deficient OA mice, but were absent from the matrix ofhealthy, intact articular cartilage. Extracellular stainingwas also observed in fibrocartilaginous soft tissues, suchas the menisci and the tendon insertion sites that areknown to secrete type II collagen. This observationindicates that such markers not only measure the artic-ular cartilage degradation but also type II collagencatabolism in the other soft tissues of the joint. Theextracellular staining of the menisci and tendons wasmore conspicuous in the tissues from biglycan/fibromodulin double-deficient mice than in those fromWT mice. This result was expected, since these tissues inbiglycan/fibromodulin double-deficient animals are hy-pertrophic and prone to ectopic, endochondral ossifica-tion (31). In addition to this extracellular staining, all celltypes known to synthesize type II collagen (chondro-cytes, cells in menisci, and some tenocytes) expressed all3 epitopes intracellularly. While such expression mightbe expected for biomarkers of collagen synthesis, it issomewhat more surprising for biomarkers of collagendegradation.
There are 2 possible explanations for this obser-vation. First, the antibodies may recognize epitopes onnascent collagen fibrils. However, the lack of cross-reactivity between antibodies to C1,2C, Coll2-1, andColl2-1NO2 and denatured collagen molecules (13,19) isnot consistent with this hypothesis. A second, moreprobable, explanation is that intracellular staining re-flects cellular uptake of degraded collagen molecules, aspreviously shown for hyaluronan and aggrecan frag-ments (39,40). If this is indeed the case, labeling ofhealthy chondrocytes not only in WT mice, but also inbiglycan/fibromodulin double-deficient mice, reflectsphysiologic type II collagen turnover. Certainly, suchturnover in healthy mice is not negligible, since C2C,Coll2-1, and Coll2-1NO2 are readily detectable in theserum of WT mice.
We recognize that our study has some limitations.
Since C2C measurements were only made at 2 of the 5time points, a more complete comparison of its expres-sion with that of Coll2-1 and Coll2-1NO2 is lacking.Differences between group mean and median values forC2C might result from the small group size (n � 15) andmake it difficult to draw any conclusion about C2Cconcentrations as a function of age. Finally, the studyexamined 3 biomarkers of type II collagen catabolismwhen several others exist.
In conclusion, a longitudinal comparison of C2C,Coll2-1, and Coll2-1NO2 expression with age in WT andOA mice indicated that these 3 biomarkers of type IIcollagen catabolism are independent of each other andprovide complementary, rather than redundant, infor-mation on in vivo type II collagen degradation. Sincethese biomarkers reflect different aspects and/or stagesof type II collagen degradation, their combined use mayincrease their usefulness for the diagnosis, prognosis,and treatment monitoring of OA in humans.
ACKNOWLEDGMENT
We thank Dr. Anne Donnet for her careful review ofthe English.
AUTHOR CONTRIBUTIONS
Dr. Ameye had full access to all of the data in the study andtakes responsibility for the integrity of the data and the accuracy of thedata analysis.Study design. Ameye.Acquisition of data. Deberg, Oliveira, Labasse, Henrotin.Analysis and interpretation of data. Ameye, Deberg, Henrotin.Manuscript preparation. Ameye, Deberg, Henrotin.Statistical analysis. Aeschlimann.
REFERENCES
1. Yelin E. The economics of osteoarthritis. In: Brandt KD, DohertyM, Lohmander LS, editors. Osteoarthritis. 2nd ed. Oxford: OxfordUniversity Press; 2003. p. 17–21.
2. Brandt KD, Mazzuca SA. Lessons learned from nine clinical trialsof disease-modifying osteoarthritis drugs [review]. ArthritisRheum 2005;52:3349–59.
3. Conaghan PG, Felson D, Gold G, Lohmander S, Totterman S,Altman R. MRI and non-cartilaginous structures in knee osteoar-thritis. Osteoarthritis Cartilage 2006;14 Suppl A:A87–94.
4. Garnero P, Rousseau JC, Delmas PD. Molecular basis and clinicaluse of biochemical markers of bone, cartilage, and synovium injoint diseases [review]. Arthritis Rheum 2000;43:953–68.
5. Kraus VB. Biomarkers in osteoarthritis. Curr Opin Rheumatol2005;17:641–6.
6. Schaller S, Henriksen K, Hoegh-Andersen P, Sondergaard BC,Sumer EU, Tanko LB, et al. In vitro, ex vivo, and in vivomethodological approaches for studying therapeutic targets ofosteoporosis and degenerative joint diseases: how biomarkers canassist? Assay Drug Dev Technol 2005;3:553–80.
7. Thonar EJ, Masuda K, Manicourt DH, Kuettner KE. Structureand function of normal human adult articular cartilage. In: Regin-ster JY, Pelletier JP, Pelletier J, Henrotin Y, editors. Osteoarthri-
BIOMARKERS OF TYPE II COLLAGEN CATABOLISM 3345

tis: clinical and experimental aspects. Berlin: Springer-Verlag;1999. p. 1–19.
8. Sugiyama S, Itokazu M, Suzuki Y, Shimizu K. Procollagen II Cpropeptide level in the synovial fluid as a predictor of radiographicprogression in early knee osteoarthritis. Ann Rheum Dis 2003;62:27–32.
9. Nelson F, Dahlberg L, Laverty S, Reiner A, Pidoux I, Ionescu M,et al. Evidence for altered synthesis of type II collagen in patientswith osteoarthritis. J Clin Invest 1998;102:2115–25.
10. Hollander AP, Pidoux I, Reiner A, Rorabeck C, Bourne R, PooleAR. Damage to type II collagen in aging and osteoarthritis startsat the articular surface, originates around chondrocytes, andextends into the cartilage with progressive degeneration. J ClinInvest 1995;96:2859–69.
11. Hollander AP, Heathfield TF, Webber C, Iwata Y, Bourne R,Rorabeck C, et al. Increased damage to type II collagen inosteoarthritic articular cartilage detected by a new immunoassay.J Clin Invest 1994;93:1722–32.
12. Hollander AP, Heathfield TF, Liu JJ, Pidoux I, Roughley PJ, MortJS, et al. Enhanced denaturation of the � (II) chains of type-IIcollagen in normal adult human intervertebral discs comparedwith femoral articular cartilage. J Orthop Res 1996;14:61–6.
13. Billinghurst RC, Dahlberg L, Ionescu M, Reiner A, Bourne R,Rorabeck C, et al. Enhanced cleavage of type II collagen bycollagenases in osteoarthritic articular cartilage. J Clin Invest1997;99:1534–45.
14. Knauper V, Lopez-Otin C, Smith B, Knight G, Murphy G.Biochemical characterization of human collagenase-3. J BiolChem 1996;271:1544–50.
15. Neuhold LA, Killar L, Zhao W, Sung ML, Warner L, Kulik J, et al.Postnatal expression in hyaline cartilage of constitutively activehuman collagenase-3 (MMP-13) induces osteoarthritis in mice.J Clin Invest 2001;107:35–44.
16. Wu W, Billinghurst RC, Pidoux I, Antoniou J, Zukor D, TanzerM, et al. Sites of collagenase cleavage and denaturation of type IIcollagen in aging and osteoarthritic articular cartilage and theirrelationship to the distribution of matrix metalloproteinase 1 andmatrix metalloproteinase 13. Arthritis Rheum 2002;46:2087–94.
17. Christgau S, Garnero P, Fledelius C, Moniz C, Ensig M, Gineyts E,et al. Collagen type II C-telopeptide fragments as an index ofcartilage degradation. Bone 2001;29:209–15.
18. Charni N, Juillet F, Garnero P. Urinary type II collagen helicalpeptide (HELIX-II) as a new biochemical marker of cartilagedegradation in patients with osteoarthritis and rheumatoid arthri-tis. Arthritis Rheum 2005;52:1081–90.
19. Deberg M, Labasse A, Christgau S, Cloos P, Bang HD, ChapelleJP, et al. New serum biochemical markers (Coll 2-1 and Coll 2-1NO2) for studying oxidative-related type II collagen networkdegradation in patients with osteoarthritis and rheumatoid arthri-tis. Osteoarthritis Cartilage 2005;13:258–65.
20. Jordan KM, Syddall HE, Garnero P, Gineyts E, Dennison EM,Sayer AA, et al. Urinary CTX-II and glucosyl-galactosyl-pyridino-line are associated with the presence and severity of radiographicknee osteoarthritis in men. Ann Rheum Dis 2006;65:871–7.
21. Poole AR, Ionescu M, Fitzcharles MA, Billinghurst RC. Theassessment of cartilage degradation in vivo: development of animmunoassay for the measurement in body fluids of type IIcollagen cleaved by collagenases. J Immunol Methods 2004;294:145–53.
22. Chu Q, Lopez M, Hayashi K, Ionescu M, Billinghurst RC, JohnsonKA, et al. Elevation of a collagenase generated type II collagenneoepitope and proteoglycan epitopes in synovial fluid followinginduction of joint instability in the dog. Osteoarthritis Cartilage2002;10:662–9.
23. Lu Y, Markel MD, Swain C, Kaplan LD. Development of partialthickness articular cartilage injury in an ovine model. J Orthop Res2006;24:1974–82.
24. Huebner JL, Kraus VB. Assessment of the utility of biomarkers ofosteoarthritis in the guinea pig. Osteoarthritis Cartilage 2006;14:923–30.
25. Manicourt DH, Azria M, Mindeholm L, Thonar EJ, DevogelaerJP. Oral salmon calcitonin reduces Lequesne’s algofunctionalindex scores and decreases urinary and serum levels of biomarkersof joint metabolism in knee osteoarthritis. Arthritis Rheum 2006;54:3205–11.
26. King KB, Lindsey CT, Dunn TC, Ries MD, Steinbach LS, Majum-dar S. A study of the relationship between molecular biomarkersof joint degeneration and the magnetic resonance-measured char-acteristics of cartilage in 16 symptomatic knees. Magn ResonImaging 2004;22:1117–23.
27. Mazzuca SA, Poole AR, Brandt KD, Katz BP, Lane KA, LobanokT. Associations between joint space narrowing and molecularmarkers of collagen and proteoglycan turnover in patients withknee osteoarthritis. J Rheumatol 2006;33:1147–51.
28. Deberg MA, Labasse AH, Collette J, Seidel L, Reginster JY,Henrotin YE. One-year increase of Coll 2-1, a new marker of typeII collagen degradation, in urine is highly predictive of radiologicalOA progression. Osteoarthritis Cartilage 2005;13:1059–65.
29. Garnero P, Charni N, Juillet F, Conrozier T, Vignon E. Increasedurinary type II collagen helical and C telopeptide levels areindependently associated with a rapidly destructive hip osteoar-thritis. Ann Rheum Dis 2006;65:1639–44.
30. Ameye LG, Young MF. Animal models of osteoarthritis: lessonslearned while seeking the “Holy Grail.” Curr Opin Rheumatol2006;18:537–47.
31. Ameye L, Aria D, Jepsen K, Oldberg A, Xu T, Young MF.Abnormal collagen fibrils in tendons of biglycan/fibromodulin-deficient mice lead to gait impairment, ectopic ossification, andosteoarthritis. FASEB J 2002;16:673–80.
32. Wadhwa S, Embree MC, Kilts T, Young MF, Ameye LG. Accel-erated osteoarthritis in the temporomandibular joint of biglycan/fibromodulin double-deficient mice. Osteoarthritis Cartilage 2005;13:817–27.
33. Ameye L, Young MF. Mice deficient in small leucine-rich proteo-glycans: novel in vivo models for osteoporosis, osteoarthritis,Ehlers-Danlos syndrome, muscular dystrophy, and corneal dis-eases. Glycobiology 2002;12:107–16R.
34. Quasnichka HL, Anderson-MacKenzie JM, Tarlton JF, Sims TJ,Billingham ME, Bailey AJ. Cruciate ligament laxity and femoralintercondylar notch narrowing in early-stage knee osteoarthritis.Arthritis Rheum 2005;52:3100–9.
35. Xu T, Bianco P, Fisher L, Longenecker G, Smith E, Goldstein S,et al. Targeted disruption of the biglycan gene leads to anosteoporosis-like phenotype in mice. Nat Genet 1998;20:78–82.
36. Svensson L, Aszodi A, Reinholt F, Fassler R, Heinegard D,Oldberg A. Fibromodulin-null mice have abnormal collagenfibrils, tissue organization, and altered lumican deposition intendon. J Biol Chem 1999;274:9636–47.
37. Li CY, Ziesmer SC, Lazcano-Villareal O. Use of azide andhydrogen peroxide as an inhibitor for endogenous peroxidase inthe immunoperoxidase method. J Histochem Cytochem 1987;35:1457–60.
38. Halloran BP, Ferguson VL, Simske SJ, Burghardt A, Venton LL,Majumdar S. Changes in bone structure and mass with advancingage in the male C57BL/6J mouse. J Bone Miner Res 2002;17:1044–50.
39. Embry JJ, Knudson W. G1 domain of aggrecan cointernalizes withhyaluronan via a CD44-mediated mechanism in bovine articularchondrocytes. Arthritis Rheum 2003;48:3431–41.
40. Embry Flory JJ, Fosang AJ, Knudson W. The accumulation ofintracellular ITEGE and DIPEN neoepitopes in bovine articularchondrocytes is mediated by CD44 internalization of hyaluronan.Arthritis Rheum 2006;54:443–54.
3346 AMEYE ET AL




















