
Probiotics, Prebiotics, and Phytogenic Substances for ... - MDPI
Upload
khangminh22Category
view
1download
0
SUBSTANCES STIMULATING GLUCOSE CATABOLISM BY THEOXIDATIVE REACTIONS OF THE PENTOSE PHOSPHATEPATHWAY IN HUMAN ERYTHROCYTES
Arieh Szeinberg, Paul A. Marks
J Clin Invest. 1961;40(6):914-924. https://doi.org/10.1172/JCI104330.
Research Article
Find the latest version:
https://jci.me/104330/pdf

SUBSTANCESSTIMULATING GLUCOSECATABOLISMBY THEOXIDATIVE REACTIONSOF THE PENTOSEPHOSPHATE
PATHWAYIN HUMANERYTHROCYTES*
By ARIEH SZEINBERGt AND PAUL A. MARKS
(From the Department of Medicine, College of Physicians and Surgeons, Columbia University,New York, N. Y.)
(Submitted for publication January 9, 1961; accepted February 3, 1961)
Mature human erythrocytes possess the enzymesnecessary to catabolize glucose to CO2 via pentosephosphate pathway or to lactic acid via the Emb-den-Meyerhof pathway. These cells are deficientin certain enzymes of the tricarboxylic acid cycle.In the non-nucleated red cells, the oxidative reac-tions of pentose phosphate pathway appear to bethe sole mechanism for glucose conversion to CO2and for the generation of TPNH (2, 3). Sev-eral investigators have suggested that a majorfactor in the control of the rate of glucose catabo-lism via the pentose phosphate pathway is the meta-bolic demand for TPNH (4-6). It has been dem-onstrated that addition of dyes capable of actingas electron acceptors from TPNH (i.e., methyleneblue, phenazine methosulphate, pyocyanin) in-creases the rate of activity of this pathway in eryth-rocytes, liver slices and mammary glands (7-9).Stimulation of the pentose phosphate pathway bypyruvate in bovine corneal epithelium, by insulinin rat mammary tissue, and by ammonium saltsin yeasts has been ascribed to supply of substratesrequiring TPNHfor their metabolism or to stim-ulation of enzyme systems which utilize TPNH(8, 10-12).
Among the systems requiring TPNH is thatinvolved in the maintenance of glutathione in thereduced state (13). It has been demonstratedthat human erythrocytes deficient in glucose-6-phosphate dehydrogenase (G6PD) are unable tomaintain their level of reduced glutathione uponincubation with a variety of agents, such asacetylphenylhydrazine, primaquine, ascorbic acid,alpha naphthol, nitrofurantoin, cysteine and fava
* This work was supported in part by Grants CY 2332and RG7368 of the United States Public Health Service.A preliminary report of a portion of these data has ap-peared in abstract form (1).
t Visiting Scholar in Medicine. Present address: Gov-ernment Hospital, Tel Hashomer, Israel.
bean extract ( 14-19). This finding suggests thatsuch substances might produce a relative insuffi-ciency in the supply of TPNHeither by inhibitingreactions in which TPNH is generated or byleading to an increase in the requirement for thiscoenzyme which exceeds the maximal capacity ofcells deficient in G6PD. It is possible to distin-guish between these mechanisms by determiningwhether these substances inhibit or stimulate therate of oxidation of glucose by the reactions ofthe pentose phosphate pathway.
In this investigation a study has been madeof the effect of the physiological substances-cysteine, ascorbic acid and pyruvate-and ofagents which can cause hemolytic anemia-acetyl-phenylhydrazine, nitrofurantoin, a- and- 3-naph-thol, primaquine and fava bean extract-on therate of glucose oxidation via the pentose phosphatepathway by red cells. Observations have beenmade with erythrocytes from normal subj ects andfrom persons with a genetically determined de-ficiency of G6PD. It has been found that thesesubstances increase the oxidation rate of glucosecarbon atom 1 to CO2 by red cells, but not therate of glucose conversion to lactic acid. Thestimulatory effect of these substances is muchgreater with erythrocytes with normal levels ofG6PDthan with cells from persons with a markeddeficiency of this enzyme. The increased rate ofglucose oxidation caused by these compounds isnot attributable to a direct action on G6PD.Rather, it was found that certain of these sub-stances oxidize TPNH in the presence of di-aphorase.
METHODS
Preparation of erythrocytes. Human venous blood,collected in siliconized tubes containing heparin, wasmixed with an equal volume of 3 per cent dextran(Larco, grade HH, mol wt ca. 225,000) and 3 per centglucose in 0.15 M sodium chloride, for separation of
914

PENTOSEPHOSPHATEPATHWAYINT RED CELLS
erythrocytes from leukocytes and platelets by a methoddescribed in detail elsewhere (20). The erythrocyteswere washed 3 times with 20 vol of cold 0.15 M sodiumchloride solution and resuspended in Krebs-Ringer phos-phate buffer, pH 7.4 (21), to a cell concentration ofabout 4 X 108 per mm3. This erythrocyte suspension wascontaminated with not more than 500 leukocytes per mm3and 10,000 platelets per mm3.
Hemolysates, when employed, were prepared by lysisof 1 vol of washed packed erythrocytes with 4 vol ofcold distilled water.
Isotope experiments. Two ml of the erythrocyte sus-pension in Krebs-Ringer phosphate buffer was placedin Warburg type flasks equipped with two side-arms.Glucose labeled in positions 1, 2 or 6 with C14 and non-labeled glucose were placed in a side-arm together withadditional substances as indicated in the tables. The finalvolume was made to 3 ml with Krebs-Ringer phosphatebuffer. After 10 minutes' equilibration with an ap-propriate gas, the contents of the side-arm was tippedinto the main compartment and incubation carried out at370 C for 2 hours. At the end of this period of incu-bation, 1.0 ml 50 per cent trichloroacetic acid was in-troduced into the previously used side-arm through thegas vent of the stopper and 0.2 ml hyamine hydroxideand 0.2 ml toluene (21) were placed in the second side-arm. The contents of the side-arm containing the solu-tion of acid was then tipped into the main compartment.Quantitative diffusion of released C1402 into the hyaminehydroxide mixture was achieved by shaking the flasksat 37° C for 1 hour. The hyamine-toluene mixture wasquantitatively transferred into bottles containing 1.0 mlof absolute ethanol and 5.0 ml of 0.3 per cent 2,5-diphenyl-oxazole and 0.03 per cent 2,2'-p-phenylene-bis-(5-phenyl-oxazole) in toluene. The assay of radioactivity was per-formed in a liquid scintillation counter with an efficiencyof 60 per cent and a background of 30 cpm (22).
Lactic acid determinations were performed by an enzy-matic procedure (23). An aliquot of the trichloroaceticacid extract from the incubation mixtures was centrifugedand the supernatant fluid recovered and adjusted to pH10.0 with 1.0 N NaOH. Assay mixtures were set up incuvets as follows: 2 ml glycine-NaOH buffer, 0.2 M,pH 10.5; 0.2 ml DPN, 1.5 X 10- M; 0.2 ml semicarbazideHCl, 1.0 M (adjusted to pH 10.5 with NaOH); 0.4 mlwater and 0.2 ml of the test solution. The optical densityat 340 mu of each assay mixture was determined, and0.050 ml lactic acid dehydrogenase (twice recrystallized,Nutritional Biochemicals Corp.) was added and thechange in optical density at 340 m/u was followed until nofurther increase was observed. As standards, solutionsof lithium lactate of known concentration were assayedwith each set of determinations. In preliminary ex-periments it was established that cysteine, ascorbic acid,acetylphenylhydrazine, nitrofurantoin, a-naphthol andprimaquine in the concentrations used in these studiesdid not interfere with the determination of lactic acid.
Glucose was estimated in barium hydroxide filtrates(24) by the 0-amino-diphenyl method of Timell, Glaude-mans and Currie (25). This procedure was chosen be-
cause none of the compounds studied, with the exceptionof acetylphenylhydrazine, interfered with the color reac-tion. Glucose could not be estimated in the presence ofacetylphenylhydrazine by this procedure or by theanthrone method (26), by the iodometric titration methodof Hagedorn-Jensen (27) or by an enzymatic procedureusing "Glucostat" reagents of Worthington BiochemicalCorporation (28).
G6PD and 6-phosphogluconic dehydrogenase activi-ties were assayed spectrophotometrically (29, 30). Aunit of enzyme activity is defined as the amount of en-zyme required to give an increase in optical density of0.001 per minute at 340 m/u (31). Erythrocytes fromnormal subjects assayed by this method had an activityof glucose-6-phosphate dehydrogenase of 10,000 to 20,000U per g hemoglobin, while those from subjects with afully expressed hereditary deficiency of this enzyme, 500to 4,000 U per g hemoglobin.
Materials. Erythrocyte G6PD was purified from nor-mal human erythrocytes by a method described in detailelsewhere (32). Spinach TPNH diaphorase was pre-pared according to the method of Avron and Jagendorf(33). An extract of fava bean was prepared by themethod of Walker and Bowman (18). G6PD partiallypurified from yeast and a preparation of diaphorase pre-pared from Clostridiun kluyveri were purchased fromSigma Chemical Corp. Primaquine phosphate waskindly provided to us by Winthrop Laboratories and ni-trofurantoin [N (5-nitro-2-furfurylidene) -1-aminohydan-toin] by Eaton Laboratories.
RESULTS
Stimulation of glucose oxidation to CO,Incubation of normal erythrocytes with cysteine,
ascorbic acid, acetylphenylhydrazine, primaquine,nitrofurantoin, a- or /3-naphthol is associated witha 3- to 18-fold increase in the rate of C140 forma-tion from glucose-i-C14 (Table I). A consistentlysmaller increase in the rate of C140. formation isobserved upon incubation of G6PD-deficient redcells under the same conditions. In the absence ofadded compounds, the rate of glucose oxidationto C140 is similar in normal and G6PD-deficientcells.
The increased rate of C1402 formation in thepresence of these compounds is not caused by anaction of these agents directly on glucose. Thisis indicated by the observation that when glucose-1-C14 is incubated with these compounds withoutadded erythrocytes or with boiled suspensions oferythrocytes no C140, is formed.
The increase in the rate of oxidation of glu-cose to C140. in the presence of these substancesinvolves glucose-6-phosphate (G6P) which is re-
915

ARIEH SZEINBERG AND PAUL A. MARKS
TABLE I
Effect of various compounds on the rate of C1402 formation from glucose-i-C'4 byerythrocytes of normal and G6PD-deficient subjects
Cells from normal subjectst Cells from mutant subjectst
Activity of C1402 Activity of C1402No. of No. of
Compound added* Concentration exp. Range Mean exp. Range Mean
M jtc X10-3/109 RBC ,Ic X10-3/10 RBCNone 9 0.28-0.75 0.41 8 0.16-0.52 0.35Methylene blue 1.6 X 10-4 8 7.9-25.1 16.7 6 1.8-5.2 4.0Cysteine 3.5 X 10-2 4 4.1-7.1 5.8 4 1.5-2.7 2.0Ascorbic acid 2.8 X 10-2 4 6.2-7.7 6.9 3 1.5-3.8 2.6Acetylphenylhydrazine$ 3.3 X 10-2 5 2.8-5.7 3.6 4 0.9-1.5 1.3a-Naphtholt 6.9 X 10-3 6 3.2-5.5 4.4 4 0.9-2.4 1.7Bl-Naphtholl 6.9 X 10-3 3 3.0-5.8 4.1 3 1.6-2.1 1.8Primaquine phosphate: 7.8 X 10-3 7 0.8-2.5 1.6 4 0.43-0.90 0.64Nitrofurantoint 4.2 X 10-3 6 2.4-3.3 2.7 3 0.87-2.1 1.3
* Ascorbic acid and cysteine were added as neutralized solutions in Krebs-Ringer phosphate buffer. The other com-pounds were added as dry powders. Final glucose concentration, 1 X 10-2 M; amount of glucose-i-C'4 added, 0.6 ,uc/flask; erythrocytes added, 6.9 to 9.0 X 109 flask; gas phase, 100% oxygen; incubation time, 2 hours.
t Normal subjects had red cell G6PDactivities between 10,000 and 20,000 U/g hemoglobin. Each compound wasassayed with erythrocytes of one mutant Caucasian female of Spanish extraction and a number of Negro mutants. TheG6PDactivity in erythrocytes of all these subjects was 1,000 to 4,000 U/g hemoglobin.
I These compounds are only slightly soluble in water. Their solubility in the incubation mixture used in these ex-periments is not known. Accordingly, the concentrations indicated represent calculations based on the amounts addedand not necessarily on the amounts actually in solution.
cycled via pentose phosphate, as well as G6Pformed from glucose. Addition of the variouscompounds to erythrocytes incubating with glu-cose-2-C14 is associated with an increased rate ofC140, formation (Table II). In fact, in the pres-
ence of cysteine, ascorbic acid and acetylphenyl-hydrazine, the relative increase in the rate of C140,formation from glucose-2-C14 is higher than thatfrom glucose-i-C14. When none of these com-
pounds is added, the ratio of the rate of formationof C1409 from glucose-i-C'4 to that from glucose-2-C14 is about 6, while in the presence of these com-
pounds it decreased to about 2. A similar de-
crease in this ratio is observed in the presence ofmethylene blue. This is in agreement with thereport of Brin and Yonemoto (7). The decreasein the ratio of the rate of C1402 formed from glu-cose-i-Cl4 to that from glucose-2-C14 suggeststhat the stimulation of the recycling of pentosephosphate to G6P is greater than the stimulationof glucose carbon 1 oxidation. The present dataprovide no explanation of the mechanism of thiseffect.
Incubation of erythrocytes with glucose-6-C14 isassociated with negligible amounts of C140, forma-tion. Addition of methylene blue, cysteine, as-
TABLE II
Influence of various compounds on the rates of C"402 formation from glucose-i-C'4 andglucose-2-C'4 by normal erythrocytes *
Ratio of C'402Activity of C1402 recovered from from glucose-1-C'4
to C'402 fromCompounds added Glucose-1-C14 Glucose-2-C'4 glucose-2-C"4
pC X10-3/1O RBC
None 0.54 0.09 6.0Methylene blue 10.2 5.0 2.1Cysteine 6.4 3.0 2.1Ascorbic acid 7.0 3.5 2.0Acetylphenylhydrazine 3.1 1.4 2.2ca-Naphthol 4.0 0.6 6.6Primaquine phosphate 1.5 0.37 4.0Nitrofurantoin 2.8 0.4 .8
* Conditions of experiment are as in Table I. Glucose-i-Cl' or glucose-2-C'4, 0.6 ,tc, was added to each flask. Theresults represent mean values of 3 experiments.
916

PENTOSEPHOSPHATEPATHWAYIN RED CELLS
corbic acid, acetylphenylhydrazine, primaquine,nitrofurantoin or a- or f8-naphthol to such incuba-tion mixtures causes no detectable increase in therate of glucose-6-C14 oxidation to C1402. Theseobservations are consistent with the fact that ma-
ture erythrocytes are lacking in the activity ofcertain enzymes of the tricarboxylic acid cycle(34). In addition, these data suggest that in ma-
ture red cells glyceraldehyde-3-phosphate whichmay be formed in the transketolase reaction isconverted to a very limited extent, if at all, tocarbon atoms 1 through 3 of G6P.
Oxygen dependence of the stimulatory effect
Stimulation of the C140, formation by the vari-ous compounds is less striking upon incubation ofred cells in an atmosphere of air or nitrogen thanin 100 per cent oxygen. The stimulatory effectwas least marked when the cells were pretreatedwith carbon monoxide and then incubated in an at-mosphere of 100 per cent nitrogen (Table III).The response to such treatment varied with dif-ferent compounds, the stimulation by acetylphenyl-hydrazine is almost completely abolished, whilethat by nitrofurantoin is only little affected.
Effect of concentration of the stimulating com-
pounds
The magnitude of stimulation in the rate of oxi-dation of glucose-1-C'4 to C1402 is related to the
TABLE III
Effect of treatment with carbon monoxide followed by incuba-tion in atmosphere of nitrogen on the rate of C'402formation
from glucose-l-C'4 by normal erythrocytes *
Activity of C1402recovered from
Control CO-treatedcells cells
(gas phase, (gas phase,Compound added 100% 02) 100% N2)
Ac XIO/109 RBCNone 0.51 0.11Methylene blue 15.4 5.4Cysteine 5.6 1.3Ascorbic acid 7.0 1.4Acetylphenylhydrazine 3.1 0.3a-Naphthol 4.3 1.5Primaquine phosphate 1.7 0.66Nitrofurantoin 2.8 1.9
* Conditions of experiment are as in Table I. TheCO-treated cells were obtained by passing carbon monoxidethrough the suspension of erythrocytes for 10 minutes be-fore distribution into the flasks. The results represent meanvalues of 3 experiments.
TABLE IV
Effect of plasma and of pyruvate on the rate of C1402 forma-tion from glucose-i- C'4 by normal cells *
Activity ofAddition Concentration C14O2
M' IL X10>-/109 RBC
None 0.47Plasmat 1.15Sodium pyruvate 1 X 10-3 0.95
1 X 10-2 1.4
* Conditions of experiment were described in Table I.The results represent mean values of 3 experiments.
t In the experiment with plasma, the Krebs-Ringer phos-phate buffer was replaced by 2 ml homologous cell-freeplasma. Control flasks containing plasma alone did notshow any C1402 formation.
concentration of the added compound. The con-centration of the compounds at which maximalstimulation is observed is indicated in Table I.Increasing the concentration beyond the optimalpoint is associated with a decrease in the degreeof stimulation. The maximal effect obtained withcysteine and ascorbic acid is observed at concen-trations of these substances 500 times that in nor-mal blood (which is 0.6 to 1.0x 10-4 M) (35).However, a twofold increase in the rate of C1402formation is observed upon addition of these com-pounds in concentrations of 3 x 10-4 M.
The low rate of oxidation of glucose-i-C'4 toC1402 which is observed when washed red bloodcells are incubated in a Krebs-Ringer phosphatebuffer might in part reflect a depletion of certainsubstances during the preliminary washing of thecells. Washed erythrocytes resuspended in plasmahave a higher rate of oxidation of glucose-i-C'4to C140, than do the same cells suspended in buffersolution (Table IV). This observation is in ac-cord with that of Murphy (2). It is possible thatthis effect of plasma may be attributed to its con-tent of substances such as ascorbic acid or cysteine,which stimulate the rate of glucose catabolismvia the pentose phosphate pathway. There are, ofcourse, other possible explanations for this effectof plasma and the problem requires further study.
Glucose utilization and lactic acid formation inpresence of the stimulating compoundsThe increased rate of glucose oxidation via the
pentose phosphate pathway in the presence of thevarious compounds is not associated with an in-crease in glucose catabolism via the Embden-
917

ARIEH SZEINBERG AND PAUL A. MARKS
TABLE V
Effect of various compounds on glucose utilization and lacticacid formation by normal erythrocytes *
Lactic acid GlucoseCompound added formed utilized
Aiimoles/109 RBC/2 hrsNone 1.15 0.61Methylene blue 0.92 0.72Ascorbic acid 0.93 0.69Cysteine 0.97 0.59Acetylphenylhydrazine 0.62 ta-Naphthol 0.50 0.60Primaquine phosphate 0.25 0.63Nitrofurantoin 1.00 0.60
* Conditions of experiment are as in Table I. The re-sults represent mean values of 4 experiments.
t Glucose utilization was not measured owing to lack ofa method free from interference by acetylphenylhydrazine.
Meyerhof pathway. In the presence of the stimu-lating compounds, no comparable increase in therate of glucose disappearance or lactic acid forma-tion is observed (Table V). On the contrary, un-der these conditions, a decrease in the rate of lac-tic acid formation is generally found. Primaquine,acetylphenylhydrazine and a-naphthol cause aparticularly significant decrease in the rate of lac-tic acid formation without a similar change in therate of glucose utilization. This finding suggeststhat these compounds may inhibit some step in theconversion of glucose to lactic acid beyond G6P.
Stimulation of the oxidation of glucose to CO2 bypyruvateThe addition of pyruvate to the incubation me-
dium causes an increase in the rate of oxidation ofglucose-i-C14 to C140.2 by red cells. Several in-
vestigators working with other tissues, have re-
ported that pyruvate may stimulate glucose catabo-lism via the pentose phosphate pathway (5, 10,11). It has been suggested that this effect may
result from pyruvate acting as an electron acceptorfrom TPNH in the presence of lactic dehydroge-nase. In the present study, no evidence has beenobtained that lactic acid dehydrogenase of eryth-rocytes can reduce pyruvate in the presence ofTPNH. Lactic acid dehydrogenase activity hasbeen assayed by the method of Wroblewski andLaDue (36). No activity of this enzyme is dem-onstrated in hemolysates when TPNHis used as
the hydrogen donor over the pH range of 6.0 to7.4. Similarly, no evidence has been obtainedthat the pyruvate stimulation of the rate of oxi-dation of glucose to CO, might be mediated by a
pyridine nucleotide transhydrogenase. Thus, in-cubation of a hemolysate with TPNH, DPNandpyruvate does not result in a decrease of opticaldensity at 340 mu. This finding suggests thathemolysates are lacking a transhydrogenating en-
zyme which could lead to DPNHformation andpermit the subsequent reduction of pyruvate underthese conditions.
Stimulation of the oxidation of glucose to CO2 byextract of Vicia fava
An increase in the rate of oxidation of glucose-1-C14 to C1409 is observed upon incubation oferythrocytes with an extract of fava bean (TableVI). In the main, this effect is due to a thermo-stable constituent of the extract, as a definite in-
TABLE VI
Effect of extract of Vicia fava on the rate of formation ofC1402 from glucose-1-C14 by normal erythrocytes *
Unboiled extract Boiled extractt
Cells added Extract added Activity of C1402 Extract added Activity of C402
ml Ac X10J3/vessel ml jic XIO 3/vesselErythrocytest 0 3.1 0 3.1Erythrocytest 0.25 6.9 0.25 3.1Erythrocytest 0.50 8.1 0.50 6.9Erythrocytest 1.0 9.0 1.0 7.2Boiled erythrocytes§ 0 0Boiled erythrocytes§ 1.0 0.25 1.0 0None 0.25 0.12 0.25 0None 1.0 0.30 1.0 0
* Conditions of experiment are as in Table I.t The extract of Vicia fava was heated at 1000 C for 10 min prior to addition.t 5 X 109 Erythrocytes were added.§ The erythrocytes were heated at 1000 C for 10 min prior to addition.
918

PENTOSEPHOSPHATEPATHWAYIN RED CELLS 919
TABLE VII
Influence of various compounds on the activity of G6P and 6-phosphogluconicdehydrogenases in hemolysates and of purified G6PD*
Activity of hemolysates
Activity of 6-Phospho-purified gluconic
Compound added Concentration G6PD G6PD dehydrogenase
U/min U/min U/minNone 11.0 21.1 12.0Cysteine 1 X 10-2 M 11.3 20.2 11.8Ascorbic acid 1 X 10-2 M 10.9 21.0 12.1Acetylphenylhydrazine 1.0 X 10-2 M 9.5 18.0 11.8a-Naphthol 33%1c saturated solutiont 6.5 14.0 11.9Primaquine 1.3 X 10-3 M 9.4 19.1 11.0Nitrofurantoin 33c%0 saturated solution 5.4 9.7 7.3
* The assay mixture contained 0.5 ml 0.1 MMgC12, 0.1 ml of 2.5 X 10-3 MTPN, 0.5 ml of 0.01 MG6P, 0.005 mlG6PDpurified from normal human erythrocytes or 0.050 ml hemolysate from a normal subject, 1.0 ml of a solution of therespective test substance in Krebs-Ringer phosphate buffer and water to the final volume of 3.0 ml. For 6-phospho-gluconic dehydrogenase assay, 0.5 ml of 0.01 M 6-phosphogluconate was added instead of G6P. Saturated solutionsof nitrofurantoin and a-naphthol were prepared by suspending 10 mg of the respective compounds in 10 ml of Krebs-Ringer phosphate buffer, shaking the suspensions for 10 minutes at room temperature and then centrifuging them.
t 33% Saturated solution indicates that the saturated solution of these compounds was in a threefold dilution in thefinal reaction mixture.
crease in the rate of C140, formation is observedin the presence of a boiled preparation. Thehigher rate of C1402 formation upon incubationwith an unboiled extract requires further elucida-tion, since a small quantity of C140 was alsoformed from glucose-1-C14 in the absence of eryth-rocytes, suggesting that the extract itself con-tained a system capable of oxidizing glucose toCO2.
Studies of the mechanism of stimulation of glucosecatabolism via the pentose phosphate pathway
1) Effect of the stimulatory compounds oni theactivity of G6PD. The possibility has been in-vestigated that the increase in the rate of glucoseoxidation to C1402 caused by the various agentsmay be a consequence of a direct action on the ac-tivity of G6PD. No increase in the activity ofpreparations of this enzyme purified from eryth-rocytes or of the enzyme in crude hemolysates isobserved upon addition of any of the compoundsstudied (Table VII). In fact, a-napththol andnitrofurantoin inhibit G6PD activity, a findingsimilar to that previously reported by Desforges,Kalaw and Gilchrist (37). Nitrofurantoin alsoinhibits 6-phosphogluconic dehydrogenase activity.
2) Effect of the stimnulatory compounds onTPNH oxidation. The following experimentswere performed to determine if the compoundsstimulating glucose oxidation via the pentose phos-
phate pathway cause a direct or enzyme-mediatedoxidation of TPNH.
a) TPNHwas added to solutions of cysteine,ascorbic acid, acetylphenylhydrazine, a-naphthol,primaquine and nitrofurantoin buffered at pH7.4 and the change of optical density at 340 mpufollowed with time. Of these several compounds,only cysteine causes a non-enzymatic oxidation ofTPNH. The rate of oxidation increases (butnot by a linear relationship) with rising cysteineconcentration to 1.0 x 10-2 M. Further increasein cysteine concentration is associated with a fallin the rate of TPNHoxidation.
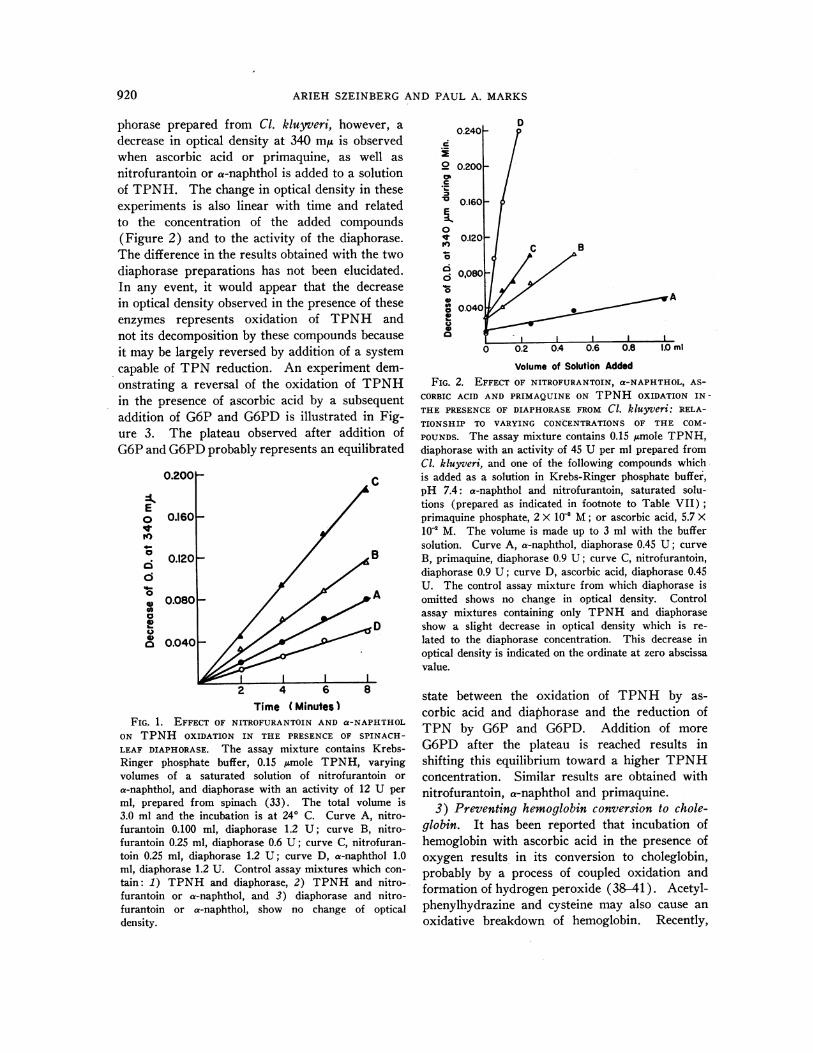
b) The experiments indicated above were thenperformed with the addition of a preparation of aTPNH-specific diaphorase purified from spinachleaves or a diaphorase prepared from Cl. kluyveriwhich had activity with either TPNHor DPNH.The rate of oxidation of TPNHin the presence ofcysteine was not influenced by the addition of eitherof these diaphorase preparations. In the presenceof the diaphorase from spinach leaves, addition ofnitrofurantoin or a-naphthol to a solution ofTPNHcauses a decrease in optical density at 340m,u. This decrease is linear with time and its mag-nitude is related to the concentration of thesecompounds and to the diaphorase activity added(Figure 1). The addition of ascorbic acid, pri-maquine or acetylphenylhydrazine to such a mix-ture is not associated with a change in opticaldensity at 340 mt. In the presence of the dia-
ARIEH SZEINBERGAND PAUL A. MARKS
phorase prepared from Cl. kluyveri, however, adecrease in optical density at 340 m/A is observedwhen ascorbic acid or primaquine, as well asnitrofurantoin or a-naphthol is added to a solutionof TPNH. The change in optical density in theseexperiments is also linear with time and relatedto the concentration of the added compounds(Figure 2) and to the activity of the diaphorase.The difference in the results obtained with the twodiaphorase preparations has not been elucidated.In any event, it would appear that the decreasein optical density observed in the presence of theseenzymes represents oxidation of TPNH andnot its decomposition by these compounds becauseit may be largely reversed by addition of a systemcapable of TPN reduction. An experiment dem-onstrating a reversal of the oxidation of TPNHin the presence of ascorbic acid by a subsequentaddition of G6P and G6PD is illustrated in Fig-ure 3. The plateau observed after addition ofG6Pand G6PDprobably represents an equilibrated
0.200 C
Eo 0.1601W)
a~~~~~~~~~0.120 B
0 A
2 4 6 8Time (Minutes)
FIG. 1. EFFECT OF NITROFURANTOIN AND a-NAPHTHOLON TPNH OXIDATION IN THE PRESENCE OF SPINACH-LEAF DIAPHORASE. The assay mixture contains Krebs-Ringer phosphate buffer, 0.15 /Amole TPNH, varyingvolumes of a saturated solution of nitrofurantoin ora-naphthol, and diaphorase with an activity of 12 U perml, prepared from spinach (33). The total volume is3.0 ml and the incubation is at 240 C. Curve A, nitro-furantoin 0.100 ml, diaphorase 1.2 U; curve B, nitro-furantoin 0.25 ml, diaphorase 0.6 U; curve C, nitrofuran-toin 0.25 ml, diaphorase 1.2 U; curve D, a-naphthol 1.0ml, diaphorase 1.2 U. Control assay mixtures which con-tain: 1) TPNH and diaphorase, 2) TPNH and nitro-furantoin or a-naphthol, and 3) diaphorase and nitro-furantoin or ax-naphthol, show no change of opticaldensity.
o 0.200_
0.160 -
0
- 0, 120 -; Se
o0,08060
004
0 0.2 0.4 0.6 0.8 1.0 ml
Volume of Solution AddedFIG. 2. EFFECT OF NITROFURANTOIN, a-NAPHTHOL, AS-
CORBIC ACID AND PRIMAQUINE ON TPNH OXIDATION IN-THE PRESENCEOF DIAPHORASE FROM Cl. kluyveri: RELA-TIONSHIP TO VARYING CONCENTRATIONSOF THE COM-POUNDS. The assay mixture contains 0.15 imole TPNH,diaphorase with an activity of 45 U per ml prepared fromCl. kluyveri, and one of the following compounds whichis added as a solution in Krebs-Ringer phosphate buffer,pH 7.4: a-naphthol and nitrofurantoin, saturated solu-tions (prepared as indicated in footnote to Table VII);primaquine phosphate, 2 X 10' M; or ascorbic acid, 5.7 X102 M. The volume is made up to 3 ml with the buffersolution. Curve A, a-naphthol, diaphorase 0.45 U; curveB, primaquine, diaphorase 0.9 U; curve C, nitrofurantoin,diaphorase 0.9 U; curve D, ascorbic acid, diaphorase 0.45U. The control assay mixture from which diaphorase isomitted shows no change in optical density. Controlassay mixtures containing only TPNH and diaphoraseshow a slight decrease in optical density which is re-lated to the diaphorase concentration. This decrease inoptical density is indicated on the ordinate at zero abscissavalue.
state between the oxidation of TPNH by as-corbic acid and diaphorase and the reduction ofTPN by G6P and G6PD. Addition of moreG6PD after the plateau is reached results inshifting this equilibrium toward a higher TPNHconcentration. Similar results are obtained withnitrofurantoin, a-naphthol and primaquine.
3) Preventing hemoglobin conversion to chole-globin. It has been reported that incubation ofhemoglobin with ascorbic acid in the presence ofoxygen results in its conversion to choleglobin,probably by a process of coupled oxidation andformation of hydrogen peroxide (38-41). Acetyl-phenylhydrazine and cysteine may also cause anoxidative breakdown of hemoglobin. Recently,
920

PENTOSEPHOSPHATEPATHWAYIN RED CELLS
Mills and Randall have reported purifying fromred cells of rat and of cow a GSH peroxidase,which catalyzes the reduction of H202 by reducedglutathione (GSH) leading to the formation ofoxidized glutathione (GSSG) (40, 42, 43). Ad-dition of GSHor GSSGplus a system generatingTPNH to hemolysates of rat erythrocytes pre-vented the conversion of hemoglobin to choleglo-bin by ascorbic acid. However, in the presentstudy this mechanism could not be demonstratedin hemolysates prepared from human erythrocytes.Addition of GSH does not decrease the rate ofcholeglobin formation in the presence of ascorbicacid (Figure 4). Indeed, a slight increase in therate of choleglobin formation was observed uponaddition of GSH.1 TPNHdoes afford protectionagainst choleglobin formation, but a similar ac-tivity was exerted by TPN. Thus, no specific re-quirement for the reduced form of TPN to preventthe conversion of hemoglobin to choleglobin couldbe demonstrated in hemolysates of human redcells.
0400k
1i.E0
a4
0.300k
0.200[-
0.100
Addition of | |Addition ofG6PD G6P I IG6PD
\ ~~~~~Control without\I DIAPHORASE
-Addition \ )[of T PNH %
'1'5 0 5 10 15 20 25 30 35 40 45TIME (Minutes)
FIG. 3. REVERSAL OF THE DECREASE IN OPTICAL DEN-
SITY AT 340 MA OF A MIXTURE OF TPNH, ASCORBIC ACID
AND DIAPHORASEBY ADDITION OF G6P AND G6PD. Theinitial assay mixture (from -5 to 0 minutes) contains12 jmoles of ascorbic acid and 0.45 U of diaphorase pre-
pared from Cl. kluyveri. The control mixture containsno diaphorase (broken line). At zero time, 0.15 4mole
of TPNH is added. At 20 minutes 0.5 ml of 0.1 MMgCl2, 0.5 ml of 0.01 M G6P and 0.050 ml of G6PD(containing 30 U of enzyme activity) are added. Thisaddition is associated with a decrease in optical densityin both mixtures which probably reflects the dilution ofthe assay mixtures. At 36 minutes, an additional 60 Uof G6PD is added to both test and control mixtures.
1 Formation of oxidation products of hemoglobin upon
incubation of crystalline hemoglobin with GSHhas alsobeen observed by Mills and Randall, but it was preventedby addition of purified GSHperoxidase (41).
zm 20
0
lo 5
5-
4 ~~~~~~~ConW (without AA)
0 30 60 90 120 150 180Incubation Time (Minutes)
FIG. 4. THE EFFECT OF PYRIDINE NUCLEOTIDES AND OFGSHON THE RATE OF CHOLEGLOBIN FORMATION IN THEPRESENCE OF ASCORBIC ACID. The incubation mixturescontain Krebs-Ringer phosphate solution, hemolysatesprepared from normal human erythrocytes, and additionsas indicated adjacent to each curve. The final concen-trations of the reagents are: hemoglobin, 1.0 g per 100ml; ascorbic acid (AA), 2.8 X 10' M; TPN and TPNH,3.0 X 10' M; and GSH, 1.0 X 10' M. The final volumeof the solution is 3 ml. The incubation is at 37° C. Asimilar pattern of results, i.e., a decrease in the rate ofcholeglobin formation in the presence of TPNand TPNH,but not of GSH, is obtained with two additional sets ofconditions. 1) Hemoglobin, 0.1 g per 100 ml; ascorbicacid, 7.2 X 10' M; TPN and TPNH, 3.0 x 10' M; andGSH, 8.0 x 10- M. 2) Hemoglobin, 1.0 g per 100 ml; as-corbkc acid, 3.6 X 10' M; TPN and TPNH, 3.0 x 10' M;and GSH, 8.0 X 10' M. The method of Lemberg, Leggeand Lockwood (44) is used for choleglobin determination.
DISCUSSION
The present study indicates that the rate of oxi-dation of glucose via the pentose phosphate path-way may be selectively stimulated by cysteine, as-corbic acid or pyruvate, as well as by methyleneblue, acetylphenylhydrazine, a- and 83-naphthol,primaquine phosphate and nitrofurantoin. Theseobservations suggest that among the determinantsof the activity of the oxidative reactions of thepentose phosphate pathway in human erythrocytesin vivo may be the concentration of certain physio-logical substances, such as ascorbic acid and cys-teine as well as a variety of agents or their meta-bolic products, to which the cells may becomeexposed.
921
go

ARIEH SZEINBERG AND PAUL A. MARKS
The mode of action of the various compoundsin stimulating the rate of glucose catabolism via thepentose phosphate pathway may differ. This issuggested by the observation that conversion ofhemoglobin to carboxyhemoglobin followed by in-cubation in an atmosphere of nitrogen affects to adifferent extent the stimulatory activity of thevarious compounds.
It is probable that the increase in the rate ofoxidation of glucose to CO2 results from a more-rapid rate of reoxidation of TPNHformed in theoxidative steps of the pentose phosphate pathway.It was found that cysteine may lead to a non-en-zymatic oxidation of TPNH, while nitrofurantoin,ascorbic acid, primaquine and a-naphthol maycause TPNHoxidation in the presence of a suita-ble diaphorase. Scott and Griffith have purifieda diaphorase from human erythrocytes, whichcatalyzes the reduction of methemoglobin byTPNH (although at a much slower rate than byDPNH) (45). The activity of nitrofurantoin,ascorbic acid, a-naphthol and primaquine in thepresence of such a diaphorase from human eryth-rocytes has not yet been investigated. Such stud-ies are necessary to determine the applicability ofthe present findings with plant or bacterial di-aphorase in explaining the metabolic events inhuman erythrocytes. It is of interest thata-naphthol and nitrofurantoin inhibit G6PD ac-tivity yet increase the rate of oxidation of glucose-1-C14 to C1402. This suggests that the inhibitionof the activity of the enzyme by these agents is in-sufficient to make the enzyme activity rather thanthe concentration of TPN the factor limiting therate of glucose oxidation by erythrocytes.
A number of investigators have concluded thatthe reoxidation of TPNHis the limiting factor inthe rate of oxidation of glucose by the pentosephosphate pathway in a variety of tissues (4-12).This conclusion is based primarily on the observa-tions that substances which may serve as electronacceptors from TPNH cause a marked increasein the rate of oxidation of glucose carbon atom1 to CO2. The present findings are consistentwith this hypothesis. In both normal cells andcells with a deficiency of G6PD, addition of com-pounds which can increase the rate of TPNHre-oxidation is associated with an increase in the rateof oxidation of glucose to CO2. However, in thecells with low G6PD levels, the increment in the
rate of glucose oxidation which may be achievedupon addition of the various stimulatory com-pounds appears to be limited by the activity ofthis enzyme.
There is another aspect of this stimulation ofglucose catabolism via the pentose phosphate path-way which is of possible significance. A selectivedecrease in the activity of G6PD occurs as redcells age in vivo (46). Old erythrocytes, as wellas cells with a genetically determined deficiencyin G6PD (47) have an increased susceptibilityto destruction under normal conditions and, par-ticularly, upon exposure to a variety of agents in-cluding cysteine, acetylphenylhydrazine, nitro-furantoin, a- and 8-naphthol, primaquine and thefava bean. This has suggested that the dehy-drogenase reactions of the pentose phosphate path-way play an important role in the maintenance ofthe integrity of the red cell (14, 46, 47). Tt is gen-erally presumed that this is so because these reac-tions provide TPNH whose reductive potential,either directly or possibly through the generationof reduced glutathione, provides protection againstthe damaging effects of oxidative agents (14, 17,40-43, 48-54). The present study suggests thatone mechanism by which these substances could leadto a decreased survival time of red cells is by re-oxidizing TPNH. In the cells with low levels ofG6PD the capacity to generate TPNH may beinadequate to provide the necessary reductive po-tential to protect against the oxidative effects ofthese compounds. The finding that TPN orTPNH, but not GSH, could protect hemoglobinagainst denaturation to choleglobin in the pres-ence of ascorbic acid suggests that the protectionafforded by pyridine nucleotides need not be medi-ated through GSH. It has been demonstratedthat TPN and TPNH, but not GSH, are impor-tant to the stability of proteins other than hemo-globin, e.g., G6PD (32, 55, 56).
SUMMARY
The rate of oxidation of glucose-i-C'4 and ofglucose-2-C'4 to C1402 by erythrocytes is stimulatedby certain physiological substances such as cysteine,ascorbic acid and pyruvate, as well as by agentswhich can cause acute hemolytic anemias, namely,primaquine, nitrofurantoin, acetylphenylhydrazine,a- and f8-naphthol and an extract of fava bean.
922

PENTOSEPHOSPHATEPATHWAYIN RED CELLS
This stimulation is more marked with cells withnormal levels of glucose-6-phosphate dehydro-genase (G6PD) than with cells deficient in thisenzyme. These compounds cause no increase inthe rate of glucose utilization or in the rate oflactic acid formation.
The increased rate of catabolism of glucose viathe pentose phosphate pathway caused by theseagents is not a result of a direct effect on G6PDactivity. Indeed, a-naphthol and nitrofurantoininhibit G6PD activity. Evidence has been ob-tained to suggest that the stimulation of the path-way is caused by an increased rate of reoxidationof TPNH in the presence of these compounds.A diaphorase-mediated oxidation of TPNH bynitrofurantoin, a-naphthol, ascorbic acid and pri-maquine has been demonstrated. Cysteine canreoxidize TPNH even in the absence of a di-aphorase. The increased rate of reoxidation ofTPNHmay be a factor in the mechanism by whichthese compounds lead to the destruction of agedred cells and erythrocytes with a genetically de-termined deficiency in G6PD.
ACKNOWLEDGMENT
The skillful help of Miss Theresa Fiorino is gratefullyacknowledged.
REFERENCES
1. Szeinberg, A., and Marks, P. A. Study of the roleof the pentose phosphate pathway in red cell me-tabolism. Clin. Res. 1960, 8, 248.
2. Murphy, J. R. Erythrocyte metabolism. II. Glu-cose metabolism and pathways. J. Lab clin. Med.1960, 55, 286.
3. Johnson, A. B., and Marks, P. A. Glucose metabo-lism and oxygen consumption in normal and glu-cose-6-phosphate dehydrogenase deficient humanerythrocytes. Clin. Res. 1958, 6, 187.
4. Dickens, F. Part II. Enzymes in red blood cells.Recent advances in knowledge of the hexosemonophosphate shunt. Ann. N. Y. Acad. Sci.1958, 75, 71.
5. Holzer, H. Carbohydrate metabolism. Ann. Rev.Biochem. 1959, 28, 171.
6. deDuve, C., and Hers, H. G. Carbohydrate metabo-lism. Ann. Rev. Biochem. 1957, 26, 149.
7. Brin, M., and Yonemoto, R. H. Stimulation of theglucose oxidative pathway in human erythrocytesby methylene blue. J. biol. Chem. 1958, 230, 307.
8. McLean, P. Carbohydrate metabolism of mammarytissue. III. Factors in the regulation of pathwaysof glucose catabolism in the mammary gland ofthe rat. Biochim. biophys. Acta 1960, 37, 296.
9. Cahill, G. F., Jr., Hastings, A. B., Ashmore, J., andZottu, S. Studies on carbohydrate metabolism inrat liver slices. X. Factors in the regulation ofpathways of glucose metabolism. J. biol. Chem.1958, 230, 125.
10. Kinoshita, J. H. The stimulation of the phospho-gluconate oxidation pathway by pyruvate in bo-vine corneal epithelium. J. biol. Chem. 1957, 228,247.
11. Wenner, C. E. Oxidation of reduced triphosphopyri-dine nucleotide by ascites tumor cells. J. biol.Chem. 1959, 234, 2472.
12. Holzer, H., and Witt, I. Beschleunigung des oxy-dativen Pentosephosphatcyclus in Hefezellen durchAmmoniumsalze. Biochim. biophys. Acta 1960, 38,163.
13. Rall, T. W., and Lehninger, A. L. Glutathione re-ductase of animal tissues. J. biol. Chem. 1952,194, 119.
14. Beutler, E. The hemolytic effect of primaquine andrelated compounds: A review. Blood 1959, 14, 103.
15. Zinkham, W. H., and Childs, B. A defect of glu-tathione metabolism in erythrocytes from patientswith a naphthalene-induced hemolytic anemia. Pe-diatrics 1958, 22, 461.
16. Kimbro, E. L., Jr., Sachs, M. V., and Torbert, J. V.Mechanism of the hemolytic anemia induced bynitrofurantoin (Furadantin). Further observationson the incidence and significance of "primaquine-sensitive" red cells. Bull. Johns Hopk. Hosp. 1957,101, 245.
17. Szeinberg, A., Adam, A., Ramot, B., Sheba, C., andMyers, F. The incorporation of isotopically la-belled glycine into glutathione of erythrocytes withglucose-6-phosphate dehydrogenase deficiency. Bio-chim. biophys. Acta 1959, 36, 65.
18. Walker, D. B., and Bowman, J. E. In vitro effectof Vicia faba extracts upon reduced glutathioneof erythrocytes. Proc. Soc. exp. Biol. (N. Y.)1960, 103, 476.
19. Mela, C., and Perona, G. P. Sulla resistenza invitro del glutatione ridotto eritrocitario al succo difave fresche; nota preventiva. Boll. Soc. ital.Biol. sper. 1959, 35, 146.
20. Marks, P. A., Gellhorn, A., and Kidson, C. Lipidsynthesis in human leukocytes, platelets, and eryth-rocytes. J. biol. Chem. 1960, 235, 2579.
21. Cohen, P. P. in Umbreit, W. W., Burris, R. H., andStauffer, J. F. Manometric Techniques and Re-lated Methods for Study of Tissue Metabolism.Minneapolis, Burgess Publishing, 1957, p. 149.
22. Passmann, J. M., Radin, N. S., and Cooper, J. A. D.Liquid scintillation technique for measuring car-bon-14-dioxide activity. Analyt. Chem. 1956, 28,484.
23. Horn, H. D., and Bruns, F. H. Quantitative Bestim-mung von L(+) - Milchsaure mit Milchsaurede-hydrogenase. Biochim. biophys. Acta 1956, 21,378.
923

ARIEH SZEINBERGAND PAUL A. MARKS
24. Somogyi, M. A method for the preparation of bloodfiltrates for the determination of sugar. J. biol.Chem. 1930, 86, 655.
25. Timell, T. E., Glaudemans, C. P. J., and Currie, A. L.Spectrophotometric method for determination ofsugars. Analyt. Chem. 1956, 28, 1916.
26. Scott, T. A., Jr., and Melvin, E. H. Determ-nationof dextran with anthrone. Analyt. Chem. 1953,25, 1656.
27. Rappaport, F. Rapid Microchemical Methods forBlood and C.S.F. Examinations. New York,Grune and Stratton, 1942, p 242.
28. Saifer, A., and Gerstenfeld, S. The photometric mi-crodetermination of blood glucose with glucoseoxidase. J. Lab. clin. Med. 1958, 51, 448.
29. Kornberg, A., and Horecker, B. L. Glucose-6- phos-phate dehydrogenase in Methods in Enzymology,S. P. Colowick and N. 0. Kaplan, Eds. NewYork, Academic Press, 1955, vol. 1, p. 323.
30. Horecker, B. L., and Smyrniotis, P. Z. Phospho-gluconic acid dehydrogenase from yeast. J. biol.Chem. 1951, 193, 371.
31. Marks, P. A., and Gross, R. T. Erythrocyte glu-cose-6-phosphate dehydrogenase deficiency: Evi-dence of differences between Negroes and Cauca-sians with respect to this genetically determinedtrait. J. clin. Invest. 1959, 38, 2253.
32. Marks, P. A., Szeinberg, A., and Banks, J. Eryth-rocyte glucose-6-phosphate dehydrogenase of nor-
mal and mutant human subjects. J. biol. Chem.1961, 236, 10.
33. Avron, M., and Jagendorf, A. T. A TPNH diaph-orase from chloroplasts. Arch. Biochem. 1956,65, 475.
34. Rubinstein, D., Ottolenghi, P., and Denstedt, 0. F.The metabolism of the erythrocyte. XIII. Enzymeactivity in the reticulocyte. Canad. J. Biochem.1956, 34, 222.
35. Stein, W. H., and Moore, S. The free amino acidsof human blood plasma. J. biol. Chem. 1954, 211,915.
36. Wroblewski, F., and LaDue, J. S. Lactic dehydro-genase activity in blood. Proc. Soc. exp. Biol.(N. Y.) 1955, 90, 210.
37. Desforges, J. F., Kalaw, E., and Gilchrist, P. In-hibition of glucose-6-phosphate dehydrogenase byhemolysis inducing drugs. J. Lab. clin. Med. 1960,55, 757.
38. Lemberg, R., and Legge, J. W. Hematin Compoundsand Bile Pigments. New York, Interscience Pub-lishers, 1949, p. 476.
39. Kaziro, K., Kikuchi, G., and Kawai, M. Reaction ofverdohemoglobin formation in vitro, with specialreference to its quantitative treatment. J. Bio-chem. (Tokyo) 1956, 43, 161.
40. Mills, G. C. Hemoglobin catabolism: I. Glutathioneperoxidase, an erythrocyte enzyme which protectshemoglobin from oxidative breakdown. J. biol.Chem. 1957, 229, 189.
41. Mills, G. C., and Randall, H. P. Hemoglobin catabo-lism: The protection of hemoglobin from oxida-tive breakdown in the intact erythrocyte. J. biol.Chem. 1958, 232, 589.
42. Mills, G. C. The purification and properties of glu-tathione peroxidase of erythrocytes. J. biol. Chem.1959, 234, 502.
43. Mills, G. C. Glutathione peroxidase and the destruc-tion of hydrogen peroxide in animal tissues. Arch.Biochem. 1960, 86, 1.
44. Lemberg, R., Legge, J. W., and Lockwood, W. H.Coupled oxidation of ascorbic acid and haemoglo-bin. 3. Quantitative studies on choleglobin forma-tion. Estimation of haemoglobin and ascorbic acidoxidation. Biochem. J. 1941, 35, 339.
45 Scottt, E. M., and Griffith, I. V. The enzymic de-fect of hereditary methemoglobinemia: Diaphorase.Biochim. biophys. Acta 1959, 34, 584.
46. Marks, P. A., Johnson, A. B., and Hirschberg, E.Effect of age on the enzyme activity in erythro-cytes. Proc. nat. Acad. Sci. (Wash.) 1958, 44,529.
47. Carson, P. E., Flanagan, C. L., Ickes, C. E., andAlving, A. S. Enzymatic deficiency in prima-quine-sensitive erythrocytes. Science 1956, 124,434.
48. Keilin, D., and Hartree, E. F. Spectroscopic studyof permeability and lysis of red blood corpuscles.Nature (Lond.) 1946, 157, 210.
49. Fegler, G. Relationship between reduced glutathionecontent and spontaneous haemolysis in shed blood.Nature (Lond.) 1952, 170, 624.
50. Benesch, R. E., and Benesch, R. Relation betweenerythrocyte integrity and sulfhydryl groups. Arch.Biochem. 1954, 48, 38.
51. Beutler, E., Robson, M., and Buttenwieser, E. Themechanism of glutathione destruction and pro-
tection in drug-sensitive and nonsensitive erythro-cytes. In vitro studies. J. clin. Invest. 1957, 36,617.
52. Horecker, B. L., and Hiatt, H. H. Pathways ofcarbohydrate metabolism in normal and neoplasticcells. New Engl. J. Med. 1958, 258, 177 and 225.
53. Marks, P. A. Red cell glucose-6-phosphate and6-phosphogluconic dehydrogenases and nucleosidephosphorylase. Science 1958, 127, 1338.
54. Jandl, J. H., Engle, L. K., and Allen, D. W. Oxi-dative hemolysis and precipitation of hemoglobin.I. Heinz body anemias as an acceleration of redcell aging. J. clin. Invest. 1960, 39, 1818.
55. Marks, P. A., Banks, J., and Gross, R. T. Glucose-6-phosphate dehydrogenase thermostability in leuko-cytes of Negroes and Caucasians with erythrocytedeficiency of this enzyme. Biochem. biophys. Res.Com. 1959, 1, 199.
56. Kirkman, H. N. Characteristics of glucose-6-phos-phate dehydrogenase from normal and primaquine-sensitive erythrocytes. Nature (Lond.) 1959, 184,1291.
924
Copyright © 2022 FDOKUMEN




















