
The lectin BJcuL induces apoptosis through TRAIL expression, caspase cascade activation and...
10
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/authorsrights
Transcript of The lectin BJcuL induces apoptosis through TRAIL expression, caspase cascade activation and...

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/authorsrights

Author's personal copy
The lectin BJcuL induces apoptosis through TRAIL expression,caspase cascade activation and mitochondrial membranepermeability in a human colon adenocarcinoma cell line
Danusa de Castro Damasio a, Stefanie Nolte a, Leonardo Puchetti Polak a,Anna Paula Brandt b, Nat�alia Borges Bonan a, Luciana Zischler a,Patrícia M. Stuelp-Campelo a, Silvia Maria S.C. Cadena b, Lúcia de Noronha a,Selene L. Elífio-Esposito a, Andr�ea Novais Moreno-Amaral a, *
a P�os-graduaç~ao em Ciencias da Saúde, Escola de Medicina, Pontifícia Universidade Cat�olica do Paran�a, 80215-901, Curitiba, Paran�a,Brazilb Departamento de Bioquímica e Biologia Molecular, Universidade Federal do Paran�a, Curitiba, Paran�a, Brazil
a r t i c l e i n f o
Article history:Received 7 April 2014Received in revised form 12 August 2014Accepted 14 August 2014Available online 3 September 2014
Keywords:Lectin BJcuLTRAIL expressionColon adenocarcinomaApoptosisMitochondria activityCaspase cascade
a b s t r a c t
It has been demonstrated that the cytotoxic effect of BJcuL, the lectin isolated fromBothrops jararacussu venom, on human gastric carcinoma is accompanied by the inhibitionof extracellular matrix adhesion, cytoskeleton disassembly and apoptosis induction. Thepresent study aimed to evaluate the apoptosis mechanisms triggered by the BJcuL inter-action with specific glycans on the surface of HT29 human colon adenocarcinoma cells. Theresults demonstrated that BJcuL interacts with glycoligands targets on the cell, which wereinhibited in the presence of D-galactose. It shows a dose-dependently cytotoxic effect thatis inhibited in the presence of D-galactose. A dose-dependent cell aggregation decrease wasalso observed for the HT29 cells. Analysis of cell proliferation inhibition was assessed byanti-PCNA and demonstrated that lectin diminishes PCNA expression when compared withuntreated cells. Differences in apoptotic marker expression estimated by immunohisto-chemistry revealed that the lectin promotes an increase in TRAIL expression, leading to anincrease in the expression of FADD, caspase-8 and Bax. Besides the increased expression ofapoptosis-related proteins, our results revealed that the lectin promotes a mitochondrialrespiration decrease and a 75% increase in the amount of cytochrome c released. Togetherthese results suggest that the cytotoxicity of BJcuL can sensitize pro-apoptotic proteins inthe cytoplasm and mitochondria, leading to the apoptotic cascade.
© 2014 Elsevier Ltd. All rights reserved.
1. Introduction
Lectins are molecules that are considered to be of greatscientific and pharmacological importance because oftheir role in many biological phenomena such as the in-duction of platelet aggregation (Ozeki et al., 1994),
hemagglutination (Mastro et al., 1986), the activation oflymphocyte proliferation, cell growth regulation (Knibbset al., 1994), cell adhesion (Barondes, 1984; Sharon andLis, 1989) and apoptosis activation (Perillo et al., 1998).These multifunctional lectin abilities are due to its char-acteristic recognition of carbohydrates in solution and onthe cell surface (Sharon and Lis, 2004). The lectin purifiedfrom Bothrops jararacussu snake venom (BJcuL) comprisestwo subunits of approximately 15 kDa and demonstrates a
* Corresponding author. Tel./fax: þ55 41 3271 2285.E-mail address: [email protected] (A.N. Moreno-Amaral).
Contents lists available at ScienceDirect
Toxicon
journal homepage: www.elsevier .com/locate/ toxicon
http://dx.doi.org/10.1016/j.toxicon.2014.08.0620041-0101/© 2014 Elsevier Ltd. All rights reserved.
Toxicon 90 (2014) 299e307

Author's personal copy
high affinity for glycoconjugates containing b-D-galacto-sides (de Carvalho et al., 1998). BJcuL is classified as a C-type lectin that requires calcium to bind to specific sugarsthrough a conserved carbohydrate recognition domain(CRD) (Powell and Varki, 1995; Drickamer and Taylor,1998). Recently, we described the ability of this lectin toinduce potent activation of neutrophils in a non-cytotoxicway (Elifio-Esposito et al., 2011), revealing that BJcuL maystimulate a wide variety of immune functions includingthe first-line defense against pathogens, cell traffickingand the induction of the innate immune response. Amongthe biological activities previously described for this lectin,its effectiveness in inhibiting tumor cell proliferationshould be emphasized. BJcuL was able to inhibit thegrowth of human renal cell carcinoma and pancreaticcancer cells (Pereira-Bittencourt et al., 1999), and it wasalso described to promote the same effect on glioma andendothelial cell progression in vitro (de Carvalho et al.,2001). These results imply that lectin serves as a poten-tial tool for decreasing tumor progression; however, themechanisms involved in the biological activities proposedfor BJcuL are unclear. Our previous results demonstratedthat BJcuL was able to promote apoptosis in the gastriccarcinoma cell lines MKN45 and AGS, which was visual-ized by DNA fragmentation. This effect manifested as adecrease in the cell viability, disorganization of the actinfilaments and inhibition of extracellular matrix adhesion(Nolte et al., 2012). In this study, we investigated theintracellular pathways that result in the programmed celldeath activation induced by a specific interaction with theBJcuL and glycoproteins on the surface of a colon adeno-carcinoma cell line.
2. Materials and methods
BJcuL: B. jararacussu venomwas kindly provided by theButantan Institute (S~ao Paulo, Brazil), and the lectin BJcuLwas purified from crude venom by affinity chromatographywith an agarose-D-galactose column as previouslydescribed (Elifio-Esposito et al., 2007). Biotinylation usingsulfo-NHS-LC-biotin (Pierce) was performed according tothe manufacturer's recommendations. To all assays, lectinconcentrations were 0.6e10 mg/ml according previouslydetermined by Nolte et al. (2012).
Cell culture: Human colon adenocarcinoma HT29 cells(ATCC) were obtained from IPATIMUP (Porto, Portugal).Cells were cultured in RPMI-1640 medium (Gibco Labora-tories, Grand Island, NY, USA) supplemented with 10%inactivated fetal calf serum (FCS) (ICN Biomedicals, Aurora,OH, USA), 100 U/ml penicillin and 100 mg/ml streptomycin(Gibco Laboratories) in a humidified atmosphere contain-ing 5% CO2 at 37 �C. Prior to each experimental procedure,cells were incubated with different concentrations of BJcuLfor 24 h at 37 �C or untreated tumor cells incubated onlywith 5% RPMI, washed with PBS, detached from the platewith PBS/EDTA (2 mM) and collected by centrifugation for5 min at 200 � g at 4 �C.
Localization of the lectin glycoligands on the sur-face of carcinoma cells: HT29 cells were seeded oncoverslips coated with Biobond (EM Science, Fort Wash-ington, PA, USA), fixed with 2% formaldehyde at room
temperature (RT) for 20 min and incubated with biotinyl-BJcuL (10 mg/ml) or PBS for 1 h at RT. After washing inPBS, the cells were incubated with streptavidin-FITC(GibcoBRL) for 30 min. The coverslips with cells weremounted on histological slides using Fluormount-G(EMScience, Gibbstown, NJ), and images were analyzedby fluorescence microscopy (Leica DMBL with a cameraattached). For the inhibition assay, biotinyl-BJcuL waspreincubated for 45 min at 37� C with 2 mM D-galactose(D-Gal) or N-acetyl-D-glucosamine (GlcNAc) (Sigma, St.Louis, MO) (Nolte et al., 2012).
MTT cell viability assay: The BJcuL cytotoxic effect wasevaluated using a conventional MTT assay. After incubationwith BJcuL (0.6, 1.25, 1.8 or 2.5 mg/ml) for 24 h at 37� C, thecells were incubated with 5 mg/ml MTT (10 ml) for 2 h.Untreated tumor cells were incubated with PBS alone orBJcuL that was preincubated for 45 min with 2 mM Gal orGlcNAc. After aspiration of the medium, DMSO was added,and the optical density (OD) wasmeasured at 550 nm usinga multiwell scanner (Tecan Sunrise ™).
Cell adhesion assay: Two different cell adhesion assayprotocols were used. In the first protocol, a tumor cellssuspension (1 � 104 cells/well in 5% RPMI) containingBJcuL (2.5 mg/ml) or 5% RPMI alone was distributed in aMatrigel-coated (BD e Biosystems) 96-well microplateand incubated for 24 h at 37 �C. In the second protocol, thetumor cells (1 � 104 cells/well in 5% RPMI) were distrib-uted in the matrigel-coated wells and incubated for 1 h at37 �C to promote adherence. Afterward, BJcuL (2.5 mg/ml)or 5% RPMI was added to the specific wells, and the cellswere incubated for 24 h at 37 �C. To determine if lectinalone promoted cell detachment and evaluate whetherthese cells maintained viability in both protocols, the cellsin the supernatant were carefully transferred to a freshplate and incubated for an additional 24 h at 37 �C for cellviability analysis. Cells that remained adherent to thematrigel-coated wells were analyzed by an MTT assay asdescribed above.
Cell aggregation assay: Adenocarcinoma cells (1 �104)were incubated with BJcuL (0.6 or 2.5 mg/ml in 5% RPMI) ina 96-well plate coated with semi-solid agar (0.66% in PBS)for 24 h at 37 �C. Untreated tumor cells were incubatedonly with 5% RPMI, and aggregation was evaluated usingan inverted microscope. For inhibition assays, BJcuL waspre-incubated for 45 min at 37� C with 2 mM Gal orGlcNAc.
Evaluation of marker of cellular proliferation andapoptosis: After incubation with BJcuL (2.5 mg/ml) or 5%RPMI alone for 24 h at 37 �C, HT29 adenocarcinoma cells(5 � 104 cells/ml) were mounted on slides by cytospincentrifugation and fixed in 4% paraformaldehyde for20 min, and the slides were then subjected to an immu-nocytochemical reaction. For this reaction, they wereincubated with 5% hydrogen peroxide in methanol for15 min, washed with distilled water and moistened withTris-phosphate buffer (TPB). An immunoperoxidase assaywas performed as reported by Chong and colleagues (2009)(Chong et al., 2009) with modifications. Briefly, specificmonoclonal antibodies for the detection of PCNA (Dako-Cytomation®) (1:400), Bax (DakoCytomation®) (1:50),TRAIL (Novocastra®) (1:50), FADD (Abcam®) (1:30) and
D.C. Damasio et al. / Toxicon 90 (2014) 299e307300

Author's personal copy
Caspase-8 (ABR Affinity Bio Reagents®) (1:100) werediluted in TPB and used as primary antibodies. After anovernight incubation with the primary antibodies, the cellswerewashed and coveredwith TPB for 15min. The reactionwas developed using the peroxidase conjugated secondaryantibody solution Advance (DakoCytomation®) and visu-alized by DAB (DakoCytomation®) addition. Up to 10 highpower field (400�) images were captured with a lightmicroscope coupled to a digital camera, and immuno-positive areas (mm2) were counted using Image Pro-Plussoftware (Media Cybernetics, MD, USA). Significant differ-ences in the immunopositive areas among the untreatedand BJcuL-stimulated cells were determined by statisticalanalysis.
Oxygen uptake: After cells incubation in the presence ornot of BJcuL (2.5 mg/ml) for 24 h at 37 �C, HT29 cells werewashed with PBS, detached from the plates using PBS/EDTA(2 mM) and centrifuged for 5 min at 200 � g at 4 �C. Theoxygen uptake in cell suspensions (1 � 106 cells) wasmonitored inRPMImediumbyhigh-resolution respirometry(Oxygraph-2k; Oroboros Instruments, Innsbruck, Austria).The measurements were made in two chambers at 37 �Cunder gentle agitation. The oxygen solubility factor wasdetermined to be 0.92 for the RPMI medium at local baro-metric pressure (91 kPa), and the concentration of oxygen atair saturation was 172.42 mM. Oxygen consumption wasmeasured in different states of respiration as previouslydescribed (Gnaiger, 2003, 2009), and it was defined as fol-lows: Routine: oxygen consumption in the absence of in-hibitors or uncouplers; Leak: respiration in the presence ofoligomycin (2 mg/ml); and Uncoupled: oxygen consumptionin the presence of the FCCP uncoupler (0.5 mM). The oxygenflow in these states was corrected by subtracting the non-mitochondrial respiration, which was obtained after theaddition of rotenone (0.5 mM) and antimycin (3 mg/ml). Theresults were expressed as pmol of oxygen flow per secondmultiplied by the total cells [pmols/(s * 106 cells)] and as themean ± S.D.
Quantification of cytochrome c release: The tech-nique described by (Appaix et al., 2000) was modified toquantify the cytochrome c released in HT29 adenocarci-noma cells after BJcuL incubation. For this purpose,HT29 cells were incubated with BJcuL (2.5 mg/ml) or onlywith 5% RPMI for 24 h at 37 �C, and after this time, theywere detached from the plates with PBS/EDTA (2 mM)and centrifuged for 5 min at 200 � g at 4 �C. The cellswere then resuspended in PBS for determination of theprotein concentration by the Bradford method (Bradford,1976) and pelleted again for incubation with 1 ml reac-tion buffer (200 mM sucrose, 10 mM TriseHCl, 10 mMEGTA, pH 7.2) supplemented with 10 mM digitonin for30 min at 4 �C. The samples were then centrifuged(10.000 � g; 30 min), and the supernatant was filtered(Millipore 0.2 mm) for an OD measurement at 405 nm.The values were compared to a cytochrome c (Sigma, St.Louis, MO) calibration curve.
Statistical analysis: Each experiment was performed atleast three times in duplicate, and the results are presentedas the mean ± S.D. Differences of means were comparedusing Student's t test and were considered significant whenp � 0.05.
3. Results
3.1. BJcuL interacts with glycoligands targets on the HT29 cellsurface and diminishes the viability of these cells
BJcuL was biotinylated to evaluate its interaction withcarbohydrates on the surface of colon adenocarcinomacells (HT29). The results demonstrated that the cells werepositively labeled when incubated with biotinyl-BJcuLalone (Fig. 1A) as well as when BJcuL was previouslyincubated with the non-specific carbohydrate (GlcNAc)(data not shown). In contrast, when lectin was pre-incubated with its specific carbohydrate (i.e., Gal), label-ing was not observed (Fig. 1B), which was similar to thecells lacking biotinyl-BJcuL incubation (data not shown).The lectin-mediated cytotoxicity of the tumor cells wasevaluated by the MTT method, and it was observedthat incubation with different BJcuL concentrationsresulted in decreased cell viability in a dose-dependentmanner (Fig. 1C) up to ~ 65% for the two highest doses(Fig. 1C), and the lowest dose provided a ~50% decrease incell viability. The cytotoxic profile was altered whenBJcuL was pre-incubated with its specific carbohydrate(i.e., Gal), which restored the cell viability to the samelevel observed in untreated cells to the lowest concen-trations used, but about 60% of viability was restoredwith the highest concentrations (Fig. 1C). When it waspre-incubated with non-specific carbohydrates (i.e.,GlcNAc), the cytotoxic effect was again evident (data notshown).
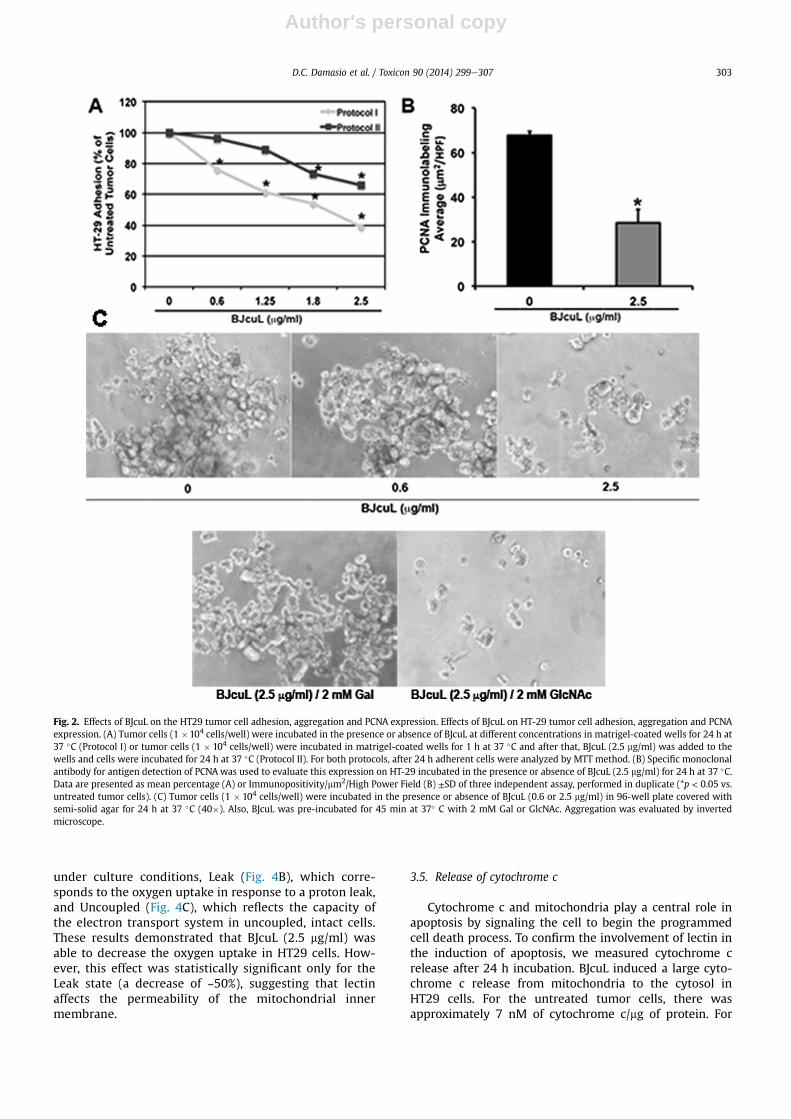
3.2. BJcuL inhibits cell adhesion, PCNA expression andprevents HT29 cell aggregation
Adhesion to the matrigel-coated wells was high whenHT29 cells were incubated with 5% RPMI (Fig. 2Ae ProtocolI), and it was dose-dependently reduced when the cellswere incubated with BJcuL, even at the lowest concentra-tion (0.6 mg/ml). The highest dose (2.5 mg/ml) promoted60% inhibition when the cells and BJcuL were simulta-neously added to the matrigel-coated wells (Fig. 2A e
Protocol I). However, when cells were incubated onmatrigel-coated wells (1 h) prior to the lectin addition theinhibitory effect on the HT29 cell adhesionwas diminished.Under this condition, only the higher BJcuL concentrations(1.8 and 2.5 mg/ml) were able to reduce cell adhesion, whilelower concentrations had no significant effect (Fig. 2A e
Protocol II).Analysis of the proliferating cell nuclear antigen
(PCNA) expression that is involved in cell proliferationsignaling revealed that HT29 cell incubation with BJcuL(2.5 mg/ml) induced an inhibition of the PCNA expression,which is highly expressed on untreated tumor cells. PCNAexpression decreased approximately 60% in HT29 cellsincubated with the lectin compared with untreated cells(Fig. 2B). To verify the action of the lectin BJcuL in pro-moted cell detached, HT29 cells were incubated with thelowest and highest concentrations of BJcuL (i.e., 0.6 and2.5 mg/ml), and cellular aggregation was evaluated with alight microscope. We observed a typical aggregationprofile for the untreated tumor cells and a dose-
D.C. Damasio et al. / Toxicon 90 (2014) 299e307 301

Author's personal copy
dependent detached after incubation with both BJcuLconcentrations (Fig. 2C). This effect was abrogated whenlectin was pre-incubated with its specific carbohydrate(D-Gal) and otherwise was not affected by GlcNAc(Fig. 2C). We also observed that detached cells by BJcuLincubation, which were collected from the supernatantsof the cultures, were not viable as revealed by the MTTassay (not shown).
3.3. BJcuL can alter the expression of apoptosis markersexpressed on HT29 cells
Immunocytochemical analysis of the obtained HT29 cellimages were evaluated for differences in apoptosis markerexpression after incubation for 24 h at 37 �C in the presenceor absence of BJcuL (2.5 mg/ml). Our results demonstratedthat TRAIL expression increased approximately 130%(Fig. 3) on HT29 cells incubated with BJcuL compared withuntreated tumor cells. Though the observed TRAILexpression was increased by incubation with BJcuL, weinvestigated the intracellular mechanisms involved in theapoptosis pathways that are induced by lectin in HT29 cells.
Thus, we evaluated FADD expression, which subsequentlyconnects with caspase-8. We observed increased FADD andcaspase-8 expression (Fig. 3), which were associated withthe apoptosis pathway in HT29 cells after incubation withBJcuL. Compared with untreated tumor cells, FADDexpression increased approximately 1.5-fold, and caspase-8expression increased approximately 3-fold in BJcuLinduced cells (Fig. 3).
Apoptotic stimuli can trigger the interaction of apoptosis-inducing proteins such as Baxwithmitochondria and lead tocytochrome c release, resulting in the activation of caspases.Our results reveal that HT29 tumor cells incubated withBJcuL demonstrate a significant increase in Bax expression(Fig. 3) compared with untreated tumor cells.
3.4. Oxygen uptake
Considering the involvement of mitochondria in celldeath processes, this study also evaluated the effect oflectin on different states of mitochondrial respiration.The respiration states were defined as Routine (Fig. 4A),which demonstrates metabolic aerobic respiration
Fig. 1. Effect of the BJcuL interaction with HT29 tumor cells. (A) Biotinyl-BJcuL recognizes a specific glycan on the tumor cell surface. Cells adherent to coverslipswere fixed and incubated with biotinyl-BJcuL (10 mg/ml) for 1 h at RT. The BJcuL interaction was observed after the cells incubation with streptavidin-FITC for30 min. (B) For the inhibition assay, biotinyl-BJcuL was preincubated with 2 mM Gal for 45 min at 37 �C. Images were analyzed by fluorescence microscopy (LeicaDMBL). (C) The BJcuL cytotoxic effect on HT29 cells. Cells were incubated with different lectin concentrations for 24 h at 37 �C or only with 5% RPMI (0). Af-terward, the cells were subjected to the MTT assay. The data are presented as the mean percentage ± SD for three independent assays performed in duplicate(*p < 0.05 vs. untreated tumor cells and **p < 0.01 vs. untreated tumor cells).
D.C. Damasio et al. / Toxicon 90 (2014) 299e307302
Author's personal copy
under culture conditions, Leak (Fig. 4B), which corre-sponds to the oxygen uptake in response to a proton leak,and Uncoupled (Fig. 4C), which reflects the capacity ofthe electron transport system in uncoupled, intact cells.These results demonstrated that BJcuL (2.5 mg/ml) wasable to decrease the oxygen uptake in HT29 cells. How-ever, this effect was statistically significant only for theLeak state (a decrease of ~50%), suggesting that lectinaffects the permeability of the mitochondrial innermembrane.
3.5. Release of cytochrome c
Cytochrome c and mitochondria play a central role inapoptosis by signaling the cell to begin the programmedcell death process. To confirm the involvement of lectin inthe induction of apoptosis, we measured cytochrome crelease after 24 h incubation. BJcuL induced a large cyto-chrome c release from mitochondria to the cytosol inHT29 cells. For the untreated tumor cells, there wasapproximately 7 nM of cytochrome c/mg of protein. For
Fig. 2. Effects of BJcuL on the HT29 tumor cell adhesion, aggregation and PCNA expression. Effects of BJcuL on HT-29 tumor cell adhesion, aggregation and PCNAexpression. (A) Tumor cells (1 �104 cells/well) were incubated in the presence or absence of BJcuL at different concentrations in matrigel-coated wells for 24 h at37 �C (Protocol I) or tumor cells (1 � 104 cells/well) were incubated in matrigel-coated wells for 1 h at 37 �C and after that, BJcuL (2.5 mg/ml) was added to thewells and cells were incubated for 24 h at 37 �C (Protocol II). For both protocols, after 24 h adherent cells were analyzed by MTT method. (B) Specific monoclonalantibody for antigen detection of PCNA was used to evaluate this expression on HT-29 incubated in the presence or absence of BJcuL (2.5 mg/ml) for 24 h at 37 �C.Data are presented as mean percentage (A) or Immunopositivity/mm2/High Power Field (B) ±SD of three independent assay, performed in duplicate (*p < 0.05 vs.untreated tumor cells). (C) Tumor cells (1 � 104 cells/well) were incubated in the presence or absence of BJcuL (0.6 or 2.5 mg/ml) in 96-well plate covered withsemi-solid agar for 24 h at 37 �C (40�). Also, BJcuL was pre-incubated for 45 min at 37� C with 2 mM Gal or GlcNAc. Aggregation was evaluated by invertedmicroscope.
D.C. Damasio et al. / Toxicon 90 (2014) 299e307 303

Author's personal copy
BJcuL-treated cells, there was a 75% increase in the amountof cytochrome c released (Fig. 5).
4. Discussion
Several lectins have been shown to have immunomo-dulating and antitumor activities (Haas et al., 1999; Kiss
et al., 1997; Pryme et al., 1996). Our study demonstratedthat the BJcuL exerted cytotoxic effects on human colonadenocarcinoma cells (HT29) that were mediated by itsinteraction with the cell surface, and this was demon-strated to be dependent on carbohydrate recognition. Weobserved that different BJcuL concentrations decreased cellviability and that the presence of Gal abolished the lectin
Fig. 3. The relationship between BJcuL and apoptosis marker expression on HT29 cells. HT29 cells (5 � 104 cells/ml) incubated in the absence (0) or presence ofBJcuL (2.5 mg/ml) for 24 h at 37 �C were mounted on slides by cytospin centrifugation and fixed with 4% paraformaldehyde for 20 min. An immunocytochemicalreaction using specific monoclonal antibodies for the detection of TRAIL, FADD, Caspase-8 and Bax was performed. The reaction was developed using theperoxidase conjugated secondary antibody solution Advance (DakoCytomation®) and visualized by DAB (DakoCytomation®) addition. Data are presented as themean immunopositivity/mm2/High Power Field ±SD for three independent assays performed in duplicate (*p < 0.05 and **p < 0.01 vs. untreated tumor cells).
Fig. 4. Effects of BJcuL on oxygen uptake by HT29 cells. Cells were incubated in the absence (0) or presence of BJcuL (2.5 mg/ml) for 24 h at 37 �C. Oxygen uptakewas monitored in RPMI medium by high-resolution respirometry (Oxygraph-2k; Oroboros Instruments, Innsbruck, Austria). The oxygen flow in the (A) Routine,(B) Leak and (C) Uncoupled states was corrected by subtracting the non-mitochondrial respiration, which was obtained after the addition of rotenone (0.5 mM)and antimycin (3 mg/ml). The results are expressed as oxygen flow per cell [pmols/(s * 106 cells)] as the mean ± SD of three independent assays. (**p < 0.01 vs.untreated tumor cells).
D.C. Damasio et al. / Toxicon 90 (2014) 299e307304

Author's personal copy
cytotoxic effects. In contrast, pre-incubation with 2 mMGlcNAc had no effect on lectin binding and tumor cellcytotoxicity. These results suggest that the binding andcytotoxic effects of lectin are due the carbohydrate recog-nition domain (CRD). Many reports have shown that lectinsinduce apoptosis in tumor cells through their carbohydratebinding capability. However, our cytotoxic assay showedthat even in the presence of its specific carbohydrate, BJcuLwas able to maintain about 40% of cytotoxic effect in thehighest concentrations on HT29 tumor cells. It is possiblethat, the galactose or galactose-containing sugars couldblock partially the BJcuL activity, not hindering completelyits antitumor activity in the highest concentrations. This isinteresting since the highest concentration of BJcuL wasshowed to promote the antitumor activity, observedthrough the increased apoptosis markers expression. Theglycan-target on the tumor cell recognized by BJcuL is notknown yet, but we can suggest that in the presence ofhighest concentration of BJcuL the cytotoxic activity isstrong and irreversible as showed by decrease the oxygenuptake and intense cytochrome-c release in HT-29 cells.Thus, similar to other lectins, BJcuL could decode the in-formation contained in the oligosaccharide units conju-gated to membrane proteins or lipids (Kennedy et al., 1995;Loris et al., 1998) and trigger a signaling cascade, inducing acellular response.
The antitumoral actions of lectins appear to occurthrough diverse mechanisms. Lebectin, a snake venom C-type lectin from Macrovipera lebectina inhibited prolifera-tion and adhesion mediated by integrin in several tumorcell lines and completely blocked cell migration towardfibronectin in a haptotaxis test, also preventing the inva-sion of tumor cells to fibrin gels (Sarray et al., 2009). Ourgroup demonstrated that BJcuL causes the disruption ofactin filaments in two gastric cancer cell lines, AGS and
MKN45, and it interferes with ECM adhesion (Nolte et al.,2012). In the present study we observed that BJcuL alsocaused HT29 cell disaggregation and changes in cellecelland cell-extracellular matrix adhesion. It has been sug-gested that these activities may be interconnected becausedisruption of the actin cytoskeleton may disable the ECM-cell adhesion. These phenomena should be carefullyanalyzed because they are factors that are central to theconversion of a premalignant lesion into an invasive car-cinoma, or conversely, they may contribute to a reductionin tumor growth (Sarray et al., 2009).
Incubation of the HT29 cells with BJcuL cause an alteredbalance between proliferation and apoptosis with adecreased expression in the proliferation markers and anincrease expression in apoptosis-related proteins and canresult in a dramatic perturbation of tumor cell homeostasis.We observed decreased proliferating cell nuclear antigen(PCNA) expression, which is highly expressed in the un-treated tumor cell, reinforcing the hypothesis that theexacerbated proliferation, which is typical for tumor cells,is reduced by lectin. Finding a molecule capable of inhib-iting an active pathway could be easier than recovering aninactive one, which is the case for proteins of tumor sup-pressor pathways that have impaired their function. Cellproliferation and apoptosis are essential for healthy tissueshomeostasis, but cancer can be triggered by action ofdifferent carcinogens that cause a loss of balance betweenthese two processes (Gaur and Aggarwal, 2003; Fulda andDebatin, 2004).
Lectins such as WGA, SFL (Shophora flavescens lectin),POL (Polygonatum odoratum lectin), and PCL (Phaselouscoccineus lectin) demonstrated cytotoxic effects on cancercell lines due to the induction of apoptosis through caspaseactivation (Gastman et al., 2004; Liu and Rabinovich, 2005;Liu et al., 2009; Chen et al., 2009). In this study, usingimmunocytochemistry, we confirm the potential of BJcuL-mediated apoptosis induction that was previously re-ported (Nolte et al., 2012), since HT29 cells demonstratedan increased expression of TRAIL, which is a member of theTNF protein family. Members of the TNF receptor (TNFR)family are type I transmembrane proteins. In this family,sequence homology is mainly found within the extracel-lular regions that mediate ligand-receptor binding. TRAILbinds to the cell surface death receptors DR4 and DR5,which are O-glycosylated proteins, and their glycosylationdomains form clusters and increase receptor sensitivity(Ashkenazi, 2008).
TRAIL-R1 (DR4 or APO-2) and TRAIL-R2 (DR5) are type Itransmembrane proteins containing death domain (Reddyand Standiford, 2010) that mediate apoptosis. When acti-vated, TRAIL receptors undergo trimerization and recruitthe Fas-associated death domain (FADD) adapter protein totheir DD (Kischkel et al., 2000). FADD, subsequently, in-teracts with pro-caspase-8 to form a complex at the re-ceptor level called the death inducing signaling complex(DISC). As the DISC is formed, it induces the activation ofcaspase-8, which activates downstream effector caspases(extrinsic apoptotic pathway) or cleaves BH3-only proteins(e.g., Bid), which translocate into mitochondria to activatethe intrinsic apoptotic pathway (Fulda and Debatin, 2006;Luo et al., 1998) and induce cytochrome c release through
Fig. 5. Effect of BJcuL (2.5 mg/ml) on the cytochrome c release in HT29adenocarcinoma cells. After incubation, cells were resuspended in PBS forprotein determination by the Bradford method and incubated with 1 ml ofreaction buffer (200 mM sucrose, 10 mM TriseHCl, 10 mM EGTA, pH 7.2)supplemented with 10 mM digitonin for 30 min, at 4 �C. The samples werethen centrifuged, and the supernatant was filtered for OD determination at405 nm. The values were compared to a cytochrome c calibration curve. Dataare presented as the mean percentage ±SD of three independent assaysperformed in duplicate (*p < 0.05 vs. untreated tumor cells).
D.C. Damasio et al. / Toxicon 90 (2014) 299e307 305

Author's personal copy
Bax activation. The results described in this study demon-strated that the lectin binds to a glycosylated target presenton the membrane, and this interaction could trigger acascade of kinases via the activation of TRAIL receptors,which consequently activate the apoptosis pathway. Thishypothesis can be supported by findings that demonstratedincreased FADD, caspase-8 and Bax expression levels afterincubating HT29 cells with BJcuL.
Mitochondria play a central role in the cell deathmechanism (Adams and Cory, 2007; Strasser et al., 2011)and Bax deficiency has been related to acquisition ofresistance to TRAIL (LeBlanc et al., 2002; Zhu et al., 2012).In this study, the increased release of cytochrome c incells treated with BJcuL suggests that lectin was capableof promoting an increase in the permeability of theexternal mitochondrial membrane through the increasein Bax expression. The cytochrome c release may besubsequently responsible for the respiration inhibitionobserved in BJcuL-treated cells. However, an inhibitoryeffect on the respiratory chain complexes cannot be dis-counted, because the effects of the uncoupled state pro-moted by BJcuL (unbound electron transport to ATPsynthesis) was less pronounced compared with otherrespiration states (Routine and Leak). Of note, the Leakrespiration state in which a decrease in oxygen con-sumption occurs in response to the re-entry of protonsinto the mitochondrial matrix was greatly affected byBJcuL. This result suggests that in addition to impairingthe outer membrane permeability, lectin BJcuL was ableto cause alterations in inner mitochondrial membranepermeability.
The resistance to TRAIL-mediated apoptosis of cancercells in vivo remains a challenging issue for the successfulapplication of TRAIL for cancer therapy. One effectivestrategy to overcome this obstacle is to combine recombi-nant TRAIL or monoclonal antibodies which target DR5with other anti-cancer agents (Bernardi et al., 2012), (Zhangand Fang, 2005). Lacour e cols (2001) (Lacour et al., 2001),using a combinatory treatment with TRAIL and 5-FU,cisplatin or doxorubicin overcame the resistance ofHT29 cells to TRAIL by increased FADD and caspase-8 ex-pressions, followed by release of cytochrome c from mito-chondria. In a very similar way, BJcuL induced an increasein TRAIL expression, recruited FADD and consequentlyinduced higher caspase-8 expression. In addition, increasedBax expression in HT29 cells incubated with BJcuL can leadto cytochrome c release and altered cellular respiration.However, even though we have not identified the BJcuL-specific glycan target on the HT29 cell surface, we caninfer that BJcuL has multiple mechanisms of action thatinvolve proliferation inhibition and the apoptotic processactivation.
Acknowledgments
The authors thank Ms Ana Paula Camargo Martins andMarina Azevedo for skillful technical assistance and theButantan Institute/Brazil for providing B. jararacussuvenom. We also thank BD Bioscience for supplying thematrigel. This research was supported by grants from CNPq(80964/2007-6).
Conflict of interest
All authors certify that there is no potential conflict ofinterests with any financial organization or entity withinterest in the subject matter or materials discussed inmanuscript entitled The lectin BJcuL induces apoptosisthrough TRAIL expression, caspase cascade activation andmitochondrial membrane permeability in a human colonadenocarcinoma cell line.
Transparency document
Transparency document related to this article can befound online at http://dx.doi.org/10.1016/j.toxicon.2014.08.062.
References
Adams, J.M., Cory, S., 2007. The Bcl-2 apoptotic switch in cancer devel-opment and therapy. Oncogene 26, 1324e1337.
Appaix, F., Minatchy, M., Riva-Lavieille, C., Olivares, J., Antonsson, B.,Saks, V.A., 2000. Rapid spectrophotometric method for quantitationof cytochrome c release from isolated mitochondria or permeabilizedcells revisited. Biochim. Biophys. Acta 1457, 175e181.
Ashkenazi, A., 2008. Targeting the extrinsic apoptosis pathway in cancer.Cytokine Growth Factor Rev. 19, 325e331.
Barondes, S.H., 1984. Soluble lectins: a new class of extracellular proteins.Science 223, 1259e1264.
Bernardi, S., Secchiero, P., Zauli, G., 2012. State of art and recent de-velopments of anti-cancer strategies based on TRAIL. Recent Pat.Anticancer Drug. Discov. 7, 207e217.
Bradford, M.M., 1976. A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein-dyebinding. Anal. Biochem. 72, 248e254.
Chen, J., Liu, B., Ji, N., Zhou, J., Bian, H.J., Li, C.Y., Chen, F., Bao, J.K., 2009. Anovel sialic acid-specific lectin from Phaseolus coccineus seeds withpotent antineoplastic and antifungal activities. Phytomedicine 16,352e360.
Chong, H., Cheah, S.H., Ragavan, M., Johgalingam, V.T., 2009. Developmentof an indirect enzyme immunoassay using monoclonal antibodies forthe measurement of 17alpha-hydroxyprogesterone in human serum.J. Immunoass. Immunochem. 30, 166e179.
de Carvalho, D.D., Marangoni, S., Oliveira, B., Novello, J.C., 1998. Purifica-tion and characterization of a new lectin from the venom of snakeBothrops jararacussu. Biochem. Biol. Mol. Inter 44, 933e938.
de Carvalho, D.D., Schmitmeier, S., Novello, J.C., Markland, F.S., 2001. Effectof BJcuL (a lectin from the venom of the snake Bothrops jararacussu)on adhesion and growth of tumor and endothelial cells. Toxicon 39,1471e1476.
Drickamer, K., Taylor, M.E., 1998. Evolving views of protein glycosylation.Trends Biochem. Sci. 23, 321e324.
Elifio-Esposito, S.L., Hess, P.L., Moreno, A.N., Lopes-Ferreira, M.,Ricart, C.A.O., Souza, M.V., Hasselmann-Zielinski, F., Becker, J.A.,Pereira, L.F., 2007. A C-Type lectin from the venom of Bothrops jar-aracussu can adhere to extracellular matrix proteins and induce therolling of leukocytes. J. Venom. Anim. Toxins Incl. Trop. Dis. 13,782e799.
Elifio-Esposito, S., Tomazeli, L., Schwartz, C., Gimenez, A.P., Fugii, G.M.,Fernandes, L.C., Zishler, L.F., Stuelp-Campelo, P.M., Moreno, A.N., 2011.Human neutrophil migration and activation by BJcuL, a galactosebinding lectin purified from Bothrops jararacussu venom. BMCImmunol. 12, 10.
Fulda, S., Debatin, K.M., 2004. Targeting apoptosis pathways in cancertherapy. Curr. Cancer Drug. Targets 4, 569e576.
Fulda, S., Debatin, K.M., 2006. Extrinsic versus intrinsic apoptosis path-ways in anticancer chemotherapy. Oncogene 25, 4798e4811.
Gastman, B., Wang, K., Han, J., Zhu, Z.Y., Huang, X., Wang, G.Q.,Rabinowich, H., Gorelik, E., 2004. A novel apoptotic pathway asdefined by lectin cellular initiation. Biochem. Biophys. Res. Commun.316, 263e271.
Gaur, U., Aggarwal, B.B., 2003. Regulation of proliferation, survival andapoptosis by members of the TNF superfamily. Biochem. Pharmacol.66, 1403e1408.
D.C. Damasio et al. / Toxicon 90 (2014) 299e307306

Author's personal copy
Gnaiger, E., 2003. Oxygen conformance of cellular respiration. Aperspective of mitochondrial physiology. Adv. Exp. Med. Biol. 543,39e55.
Gnaiger, E., 2009. Capacity of oxidative phosphorylation in human skel-etal muscle: new perspectives of mitochondrial physiology. Int. J.Biochem. Cell. Biol. 41, 1837e1845.
Haas, H., Falcone, F.H., Schramm, G., Haisch, K., Gibbs, B.F., Klaucke, J.,Poppelmann, M., Becker, W.M., Gabius, H.J., Schlaak, M., 1999. Dietarylectins can induce in vitro release of IL-4 and IL-13 from human ba-sophils. Eur. J. Immunol. 29, 918e927.
Kennedy, J.F., Palva, G., Corella, M.T.S., Cavalcant, M.S.M., Coelho, L.C.B.B.,1995. Lectins, versatile proteins of recognition: a review. Carbohydr.Polym. 8627, 219e230.
Kischkel, F.C., Lawrence, D.A., Chuntharapai, A., Schow, P., Kim, K.J.,Ashkenazi, A., 2000. Apo2L/TRAIL-dependent recruitment of endog-enous FADD and caspase-8 to death receptors 4 and 5. Immunity 12,611e620.
Kiss, R., Camby, I., Duckworth, C., De Decker, R., Salmon, I., Pasteels, J.L.,Danguy, A., Yeaton, P., 1997. In vitro influence of Phaseolus vulgaris,Griffonia simplicifolia, concanavalin A, wheat germ, and peanut ag-glutinins on HCT-15, LoVo, and SW837 human colorectal cancer cellgrowth. Gut 40, 253e261.
Knibbs, R.N., MacCallum, D.K., Lillie, J.H., Goldstein, I.J., 1994. Wild-typeand cultured Ehrlich ascites tumour cells differ in tumorigenicity,lectin binding patterns and binding to basement membranes. Gly-cobiology 4, 419e428.
Lacour, S., Hammann, A., Wotawa, A., Corcos, L., Solary, E., Dimanche-Boitrel, M.T., 2001. Anticancer agents sensitize tumor cells to tumornecrosis factor-related apoptosis-inducing ligand-mediated caspase-8activation and apoptosis. Cancer Res. 61, 1645e1651.
LeBlanc, H., Lawrence, D., Varfolomeev, E., Totpal, K., Morlan, J., Schow, P.,Fong, S., Schwall, R., Sinicropi, D., Ashkenazi, A., 2002. Tumor-cellresistance to death receptoreinduced apoptosis through mutationalinactivation of the proapoptotic Bcl-2 homolog Bax. Nat. Med. 8,274e281.
Liu, F.T., Rabinovich, G.A., 2005. Galectins as modulators of tumour pro-gression. Nat. Rev. Cancer 5, 29e41.
Liu, B., Min, M.W., Bao, J.K., 2009. Induction of apoptosis by ConcanavalinA and its molecular mechanisms in cancer cells. Autophagy 5,432e433.
Loris, R., Hamelryck, T., Bouckaert, J., Wyns, L., 1998. Legume lectinstructure. Biochim. Biophys. Acta 1383, 9e36.
Luo, X., Budihardjo, I., Zou, H., Slaughter, C., Wang, X., 1998. Bid, a Bcl2interacting protein, mediates cytochrome c release from
mitochondria in response to activation of cell surface death receptors.Cell 94, 481e490.
Mastro, A.M., Hurley, D.J., Winning, R.K., Filipowski, R., Ogilvie, M.L.,Gartner, T.K., 1986. Mitogenic activity of snake venom lectins. Cell.Tissue Kinet. 19, 557e566.
Nolte, S., de Castro Damasio, D., Barea, A.C., Gomes, J., Magalhaes, A.,Mello Zischler, L.F., Stuelp-Campelo, P.M., Elifio-Esposito, S.L., Roque-Barreira, M.C., Reis, C.A., Moreno-Amaral, A.N., 2012. BJcuL, a lectinpurified from Bothrops jararacussu venom, induces apoptosis inhuman gastric carcinoma cells accompanied by inhibition of celladhesion and actin cytoskeleton disassembly. Toxicon 59, 81e85.
Ozeki, Y., Matsui, T., Hamako, J., Suzuki, M., Fujimura, Y., Yoshida, E.,Nishida, S., Titani, K., 1994. C-type galactoside-binding lectin fromBothrops jararaca venom: comparison of its structure and functionwith those of botrocetin. Arch. Biochem. Biophys. 308, 306e310.
Pereira-Bittencourt, M., Carvalho, D.D., Gagliardi, A.R., Collins, D.C.,1999. The effect of a lectin from the venom of the snake, Bothropsjararacussu, on tumor cell proliferation. Anticancer Res. 19,4023e4025.
Perillo, N.L., Marcus, M.E., Baum, L.G., 1998. Galectins: versatile modula-tors of cell adhesion, cell proliferation, and cell death. J. Mol. Med.Berl. 76, 402e412.
Powell, L.D., Varki, A., 1995. I-type lectins. J. Biol. Chem. 270,14243e14246.
Pryme, I.F., Pusztai, A.J., Grant, G., Bardocz, S., 1996. Phytohemagglutinin-induced gut hyperplasia and the growth of a mouse lymphosarcomatumor. J. Exp. Ther. Oncol. 1, 171e176.
Reddy, R.C., Standiford, T.J., 2010. Effects of sepsis on neutrophil chemo-taxis. Curr. Opin. Hematol. 17, 18e24.
Sarray, S., Siret, C., Lehmann, M., Marrakchi, N., Luis, J., El Ayeb, M.,Andre, F., 2009. Lebectin increases N-cadherin-mediated adhesionthrough PI3K/AKT pathway. Cancer Lett. 285, 174e181.
Sharon, N., Lis, H., 1989. Lectins as cell recognition molecules. Science 246,227e234.
Sharon, N., Lis, H., 2004. History of lectins: from hemagglutinins to bio-logical recognition molecules. Glycobiology 14, 53Re62R.
Strasser, A., Cory, S., Adams, J.M., 2011. Deciphering the rules of pro-grammed cell death to improve therapy of cancer and other diseases.EMBO J. 30, 3667e3683.
Zhang, L., Fang, B., 2005. Mechanisms of resistance to TRAIL-inducedapoptosis in cancer. Cancer Gene Ther. 12, 228e237.
Zhu, S., Li, T., Tan, J., Yan, X., Zhang, D., Zheng, C., Chen, Y., Xiang, Z., Cui, H.,2012. Bax is essential for death receptor-mediated apoptosis inhuman colon cancer cells. Cancer Biother. Radiopharm. 27, 577e581.
D.C. Damasio et al. / Toxicon 90 (2014) 299e307 307




















