
The influence of vibrissal somatosensory processing in rat superior colliculus on prey capture
10
THE INFLUENCE OF VIBRISSAL SOMATOSENSORY PROCESSING IN RAT SUPERIOR COLLICULUS ON PREY CAPTURE P. D. N. FAVARO, a1 T. S. GOUVÊA, a1 S. R. DE OLIVEIRA, a N. VAUTRELLE, b P. REDGRAVE b AND E. COMOLI a * a Department of Physiology, School of Medicine of Ribeirão Preto, University of São Paulo, Avenida Bandeirantes, 3900, CEP: 14049- 900, Ribeirão Preto, São Paulo, Brazil b Department of Psychology, University of Sheffield, Sheffield, S10 2TP, UK Abstract—The lateral part of intermediate layer of superior colliculus (SCl) is a critical substrate for successful predation by rats. Hunting-evoked expression of the activity marker Fos is concentrated in SCl while prey capture in rats with NMDA lesions in SCl is impaired. Particularly affected are rapid orienting and stereotyped sequences of actions associated with predation of fast moving prey. Such deficits are consis- tent with the view that the deep layers of SC are important for sensory guidance of movement. Although much of the rele- vant evidence involves visual control of movement, less is known about movement guidance by somatosensory input from vibrissae. Indeed, our impression is that prey contact with whiskers is a likely stimulus to trigger predation. More- over, SCl receives whisker and orofacial somatosensory in- formation directly from trigeminal complex, and indirectly from zona incerta, parvicelular reticular formation and so- matosensory barrel cortex. To better understand sensory guidance of predation by vibrissal information we investi- gated prey capture by rats after whisker removal and the role of superior colliculus (SC) by comparing Fos expression after hunting with and without whiskers. Rats were allowed to hunt cockroaches, after which their whiskers were removed. Two days later they were allowed to hunt cockroaches again. Without whiskers the rats were less able to retain the cock- roaches after capture and less able to pursue them in the event of the cockroach escaping. The predatory behaviour of rats with re-grown whiskers returned to normal. In parallel, Fos expression in SCl induced by predation was significantly reduced in whiskerless animals. We conclude that whiskers contribute to the efficiency of rat prey capture and that the loss of vibrissal input to SCl, as reflected by reduced Fos expression, could play a critical role in predatory deficits of whiskerless rats. © 2011 IBRO. Published by Elsevier Ltd. All rights reserved. Key words: predatory behaviour, vibrissae, somatosensory information, sensorimotor integration. Insect predation is a fundamental motivated behaviour of many rodents, including rats. As such it is an interesting paradigm which has been used to study patterns of innate predatory hunting. Cockroaches have been considered a suitable prey because they are relatively innocuous, easily overcome, and appear very palatable with both hunting and subsequent consumption contributing to a high hedo- nic value associated with the activity (Comoli et al., 2005). Previous studies have shown that insect predation in rats is associated with the expression of Fos protein at the lateral part of intermediate layer of superior colliculus (SCl) (Comoli and Canteras, 1998). Moreover, rats with local bilateral NMDA lesions in SCl typically fail to orient towards and chase the cockroaches with the series of stereotyped movements commonly seen in the predatory hunting of intact controls (Comoli and Canteras, 2000; Furigo et al., 2010). The superior colliculus (SC) is known to be responsible for detecting and orienting the head and eyes toward visual stimuli (Grantyn, 1988; Munoz and Guitton, 1989; Wurtz, 1996; Sparks, 1999; Boehnke and Munoz, 2008). In the case of rodents it is also important for positioning the head in space during a variety of oral tasks, such as predation, foraging for food, and nocifensive responses directed to localized pain (Redgrave et al., 1990; Welzl et al., 1984; Steiner and Huston, 1992; Wang and Redgrave, 1997; Ewert et al., 1999). It is also involved in gaze-shifts di- rected towards auditory, somatosensory and even olfac- tory cues (Grobstein, 1988; Dean et al., 1989; King, 2004; Boehnke and Munoz, 2008; Felsen and Mainen, 2008). Moreover SC is critical for the detection and guidance of initial responses to unexpected objects in the visual field (Grantyn, 1988; Munoz and Guitton, 1989; Wurtz, 1996; Sparks, 1999; Coizet et al., 2003; Comoli et al., 2003), and orienting the head away from potentially threatening stimuli (Dean et al., 1989; Brandão et al., 1994, 1999). In the light of our previous behavioural observations, the movement of prey, in particular their contacts with the rats’ whiskers seem to be the critical stimulus responsible for evoking hunting and to the up-regulation of Fos in the SCl. Anatomical and electrophysiological data show that SCl receives whisker and orofacial-related somatosensory information from a range of sources including the spinal (SpV) and principal trigeminal nuclei (PrV) (Killackey and Erzurumlu, 1981; Huerta et al., 1983; Comoli and Can- teras, 2000; Hemelt and Keller, 2007), zona incerta (ZI) (Kolmac et al., 1998; Comoli and Canteras, 2000, parvice- lular reticular nucleus (PARN) and somatosensory cortex (SSp) (Comoli and Canteras, 2000; Cohen et al., 2008). In line with this view, the fast attacks associated with prey 1 PDNF and TSG had the same participation in the study. *Corresponding author. Tel: 55-16-3602-4525; fax: 55-16-3633-0017. E-mail address: [email protected] (E. Comoli). Abbreviations: AQ, cerebral aqueduct; FLI, Fos-like-immunoreactivity; PAG, periaqueductal gray; SAI, stratum album intermediale; SAP, stratum album profundum; SC, superior colliculus; SCl, superior col- liculus, lateral part; SCm, superior colliculus, medial part; SGI, stratum griseum intermediale; SGP, stratum griseum profundum; SGS, stra- tum griseum superficiale; SZ, stratum zonale. Neuroscience 176 (2011) 318 –327 0306-4522/11 $ - see front matter © 2011 IBRO. Published by Elsevier Ltd. All rights reserved. doi:10.1016/j.neuroscience.2010.12.009 318
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of The influence of vibrissal somatosensory processing in rat superior colliculus on prey capture

U9
2
*EAPslgt
Neuroscience 176 (2011) 318–327
THE INFLUENCE OF VIBRISSAL SOMATOSENSORY PROCESSING IN
RAT SUPERIOR COLLICULUS ON PREY CAPTUREP. D. N. FAVARO,a1 T. S. GOUVÊA,a1
S. R. DE OLIVEIRA,a N. VAUTRELLE,b
P. REDGRAVEb AND E. COMOLIa*aDepartment of Physiology, School of Medicine of Ribeirão Preto,
niversity of São Paulo, Avenida Bandeirantes, 3900, CEP: 14049-00, Ribeirão Preto, São Paulo, Brazil
bDepartment of Psychology, University of Sheffield, Sheffield, S10TP, UK
Abstract—The lateral part of intermediate layer of superiorcolliculus (SCl) is a critical substrate for successful predationby rats. Hunting-evoked expression of the activity marker Fosis concentrated in SCl while prey capture in rats with NMDAlesions in SCl is impaired. Particularly affected are rapidorienting and stereotyped sequences of actions associatedwith predation of fast moving prey. Such deficits are consis-tent with the view that the deep layers of SC are important forsensory guidance of movement. Although much of the rele-vant evidence involves visual control of movement, less isknown about movement guidance by somatosensory inputfrom vibrissae. Indeed, our impression is that prey contactwith whiskers is a likely stimulus to trigger predation. More-over, SCl receives whisker and orofacial somatosensory in-formation directly from trigeminal complex, and indirectlyfrom zona incerta, parvicelular reticular formation and so-matosensory barrel cortex. To better understand sensoryguidance of predation by vibrissal information we investi-gated prey capture by rats after whisker removal and the roleof superior colliculus (SC) by comparing Fos expression afterhunting with and without whiskers. Rats were allowed to huntcockroaches, after which their whiskers were removed. Twodays later they were allowed to hunt cockroaches again.Without whiskers the rats were less able to retain the cock-roaches after capture and less able to pursue them in theevent of the cockroach escaping. The predatory behaviour ofrats with re-grown whiskers returned to normal. In parallel,Fos expression in SCl induced by predation was significantlyreduced in whiskerless animals. We conclude that whiskerscontribute to the efficiency of rat prey capture and that theloss of vibrissal input to SCl, as reflected by reduced Fosexpression, could play a critical role in predatory deficits ofwhiskerless rats. © 2011 IBRO. Published by Elsevier Ltd. Allrights reserved.
Key words: predatory behaviour, vibrissae, somatosensoryinformation, sensorimotor integration.
1 PDNF and TSG had the same participation in the study.Corresponding author. Tel: �55-16-3602-4525; fax: �55-16-3633-0017.-mail address: [email protected] (E. Comoli).bbreviations: AQ, cerebral aqueduct; FLI, Fos-like-immunoreactivity;AG, periaqueductal gray; SAI, stratum album intermediale; SAP,tratum album profundum; SC, superior colliculus; SCl, superior col-iculus, lateral part; SCm, superior colliculus, medial part; SGI, stratum
riseum intermediale; SGP, stratum griseum profundum; SGS, stra-um griseum superficiale; SZ, stratum zonale.
0306-4522/11 $ - see front matter © 2011 IBRO. Published by Elsevier Ltd. All righdoi:10.1016/j.neuroscience.2010.12.009
318
Insect predation is a fundamental motivated behaviour ofmany rodents, including rats. As such it is an interestingparadigm which has been used to study patterns of innatepredatory hunting. Cockroaches have been considered asuitable prey because they are relatively innocuous, easilyovercome, and appear very palatable with both huntingand subsequent consumption contributing to a high hedo-nic value associated with the activity (Comoli et al., 2005).
Previous studies have shown that insect predation inrats is associated with the expression of Fos protein at thelateral part of intermediate layer of superior colliculus (SCl)(Comoli and Canteras, 1998). Moreover, rats with localbilateral NMDA lesions in SCl typically fail to orient towardsand chase the cockroaches with the series of stereotypedmovements commonly seen in the predatory hunting ofintact controls (Comoli and Canteras, 2000; Furigo et al.,2010).
The superior colliculus (SC) is known to be responsiblefor detecting and orienting the head and eyes toward visualstimuli (Grantyn, 1988; Munoz and Guitton, 1989; Wurtz,1996; Sparks, 1999; Boehnke and Munoz, 2008). In thecase of rodents it is also important for positioning the headin space during a variety of oral tasks, such as predation,foraging for food, and nocifensive responses directed tolocalized pain (Redgrave et al., 1990; Welzl et al., 1984;Steiner and Huston, 1992; Wang and Redgrave, 1997;Ewert et al., 1999). It is also involved in gaze-shifts di-rected towards auditory, somatosensory and even olfac-tory cues (Grobstein, 1988; Dean et al., 1989; King, 2004;Boehnke and Munoz, 2008; Felsen and Mainen, 2008).Moreover SC is critical for the detection and guidance ofinitial responses to unexpected objects in the visual field(Grantyn, 1988; Munoz and Guitton, 1989; Wurtz, 1996;Sparks, 1999; Coizet et al., 2003; Comoli et al., 2003), andorienting the head away from potentially threatening stimuli(Dean et al., 1989; Brandão et al., 1994, 1999).
In the light of our previous behavioural observations,the movement of prey, in particular their contacts with therats’ whiskers seem to be the critical stimulus responsiblefor evoking hunting and to the up-regulation of Fos in theSCl. Anatomical and electrophysiological data show thatSCl receives whisker and orofacial-related somatosensoryinformation from a range of sources including the spinal(SpV) and principal trigeminal nuclei (PrV) (Killackey andErzurumlu, 1981; Huerta et al., 1983; Comoli and Can-teras, 2000; Hemelt and Keller, 2007), zona incerta (ZI)(Kolmac et al., 1998; Comoli and Canteras, 2000, parvice-lular reticular nucleus (PARN) and somatosensory cortex(SSp) (Comoli and Canteras, 2000; Cohen et al., 2008). In
line with this view, the fast attacks associated with preyts reserved.
scbEi
P. D. N. Favaro et al. / Neuroscience 176 (2011) 318–327 319
capture by wild shrews depend on information from thewhiskers rather than visual and olfactory cues (Anjum etal., 2006).
Although SCl has an important role in predatory hunt-ing in rats (Comoli and Canteras, 2000; Furigo et al.,2010), the link to somatosensory input to the SCl from thevibrissae is still missing. The purpose of the present studywas, therefore, to determine the effects of removing thewhiskers both on the behavioural performance of preda-tory hunting and on immediate early gene (c-Fos) activa-tion in the SCl which has shown to be elicited by hunting(Comoli and Canteras, 1998).
EXPERIMENTAL PROCEDURES
The basic design of the present study involved behavioural ob-servations of predatory hunting together with measurements ofassociated Fos expression in the SCl in rats with intact whiskers,rats that had their whiskers subsequently removed, and ratswhose whiskers had been allowed to re-grow to their original size(35 days after removal).
Animal housing
All experimental procedures were performed with adults maleWistar rats (n�22) each weighting aproximately 230 g obtainedfrom local Ribeirão Preto breeding facilities. The rats were keptunder controlled temperature (23�2 °C) and 12-h light/dark cycle(lights on at 7 AM), and had free access to water and standardlaboratory diet. Experiments followed the National Institutes ofHealth Guidelines for the Care and Use of Laboratory Animals(NIH Publications No. 80-23, 1996), and were approved by theCommittee on Care and Use of Laboratory Animals of the Schoolof Medicine of Ribeirão Preto—University of São Paulo, Brazil(Protocol number 133/2007). Effort was made to minimize thenumber of animals used and their suffering.
Experimental procedure
Seven days before the experiments began, rats were handledrepeatedly by the same investigator who conducted the behav-ioural tests. Rats were individually placed into an acrylic cage(40�30�16 cm3) for 2 h each day in which free access to rat chowand water was available.
Subsequently, hunting sessions were carried out between09:00 and 12:00 h, during the light phase of the cycle. Rats wereinduced to hunt by simultaneously introducing five mature intactcockroaches (Leurolestes circunvagans), raised for this purposein our laboratory, into the experimental cage. The rapid movementof the prey was sufficient to elicit hunting and no additional in-ducement was needed. A hunting trial for each rat lasted until ithad caught all five cockroaches or 20 min had elapsed at whichpoint the trial was ended by the experimenter. Hunting was vid-eotaped using an overhead camera and the resulting record re-tained for additional behavioural analysis.
For the analysis of predatory behaviour seven animals wereallowed to hunt cockroaches during three successive sessions: (i)An initial baseline session (Vib) in which rats were allowed to huntwith their whiskers intact. (ii) Rats then had their whiskers re-moved while under anaesthesia (ketamine, xylazine andacepromazine; 45:5:0.5 mg/kg, i.p.) (União Farmacêutica Nacio-nal S/A, Embu-Guaçu, SP, Brazil; Laboratórios Calier S/A Barce-lona, Espanha; Rhobifarma Indústria Farmacêutica Ltda, SãoPaulo, SP, Brazil). Forty-eight hours later they were allowed tohunt for a second time (VibX). (iii) A period of 35 days was thenallowed during which the rats’ whiskers re-grew to their originalsize, at which point they were allowed to hunt for a third and final
time (VibR).For the histological analysis of c-Fos induced by hunting withand without whiskers we used 20 animals. These included: agroup of five animals who experienced the same Vib experimentalcondition of hunting with whiskers; a second group of five whoexperienced the VibX experimental condition of hunting withoutwhiskers; five animals from the final behavioural VibR session inwhich hunting was conducted with re-grown whiskers; and a con-trol group (n�5) who were similarly housed and handled, but werenever allowed to hunt nor had their whiskers removed (they wereplaced undisturbed in the experimental cage during the test pe-riod). After behavioural testing the brain tissue of all animals wastaken for immunohistochemical analysis (see below).
Behavioural analysis
A trained observer scored behavioural characteristics using theethological analysis software “The Observer XT®”, (version XT 7;Noldus Information Technology, Wagenigen, The Netherlands).The following events were measured: (i) Latency for the firstattack. (ii) Total time spent hunting—forward head and bodymovements directed towards a cockroach. (iii) The duration ofeach occasion the rat caught and held one of the prey—time fromwhen the prey disappeared below the rat from the extension of itsshoulder to its snout. This measure included the time spent han-dling and eating prey. (iv) Frequency of occasions when the ratimmediately followed cockroach after it had escaped (been re-leased)—the nose of the rat moving in the same direction as thecockroach in successive video frames. (v) Frequency of occasionswhen the rat failed to follow the cockroach after an escape—therat’s nose either remained stationary or moved in a differentdirection to that of the cockroach in successive video frames. (vi)The total number of cockroaches killed and eaten. (vii) Otherstates—including grooming, drinking and resting.
Immunohistochemical procedures
After all behavioural procedures had been completed, rats whosebrain tissue was to be analysed for c-Fos were deeply anesthe-tized (by ketamine, xylazine and acepromazine; 100:20:3 mg/kg,i.p.) (União Farmacêutica Nacional S/A, Embu-Guaçu, SP, Brazil;Laboratórios Calier S/A Barcelona, Espanha; Rhobifarma In-dústria Farmacêutica Ltda, São Paulo, SP, Brazil) 90 min aftertheir final hunting session. They were perfused transcardially witha brief saline rinse followed by an ice-cold solution of 4% parafor-maldehyde in a 0.1 M phosphate buffer at pH 7.4. Two hours laterthe brains were removed from the skull and cryoprotected with20% sucrose in 0.1 M phosphate buffer at 4 °C. The brains werethen frozen and sectioned coronally on a sliding microtome (LeicaMicrosystems, Germany) (30 �m) and sorted into four series. Oneeries was processed (rabbit anti-Fos, dilution 1:20,000; Calbio-hem, San Diego, CA, USA, lot # D00007099) with the avidin–iotin-peroxidase immunohistochemical technique (Vectastainlite ABC kit, Vector Laboratories, Burlingame, CA, USA) accord-
ng to the method of Gerfen and Sawchenko (1984) to revealFos-like immunoreactivity. Briefly, sections were washed in potas-sium phosphate-buffered saline (KPBS; pH 7.4) and incubated for90 min at room temperature in a solution of biotinylated goatanti-rabbit IgG (Vector Laboratories, Burlingame, CA, USA). Theywere then exposed to a mixed avidin–biotin horseradish peroxi-dase (HRP) complex solution (ABC Elite Kit, Vector Laboratories,Burlingame, CA, USA) for the same period. The peroxidase reac-tion product was visualized using the glucose oxidase procedure(Itoh et al., 1979) and the metal free 3,3’ diaminobenzidine tetra-hydrochloride (DAB, Sigma, St Louis, MO, USA) as a chromogen.Sections were exposed for 10 min to a 0.02% chromogen solutioncontaining 0.3% nickel ammonium sulfate in 0.05 M phosphatebuffer (pH 7.4) and glucose oxidase, followed by the addition of�-D-glucose to produce a blue-black product. The reaction was
stopped by extensive washing in KPBS, pH 7.4. The sections
sat
(pAc
P. D. N. Favaro et al. / Neuroscience 176 (2011) 318–327320
were mounted on gelatinized slides, and then dehydrated andcoverslipped with DPX (Sigma, St. Louis, MO, USA). An adjacentseries was stained with Thionin to serve as a cytoarchitectonicreference for the distribution of Fos labeling.
Fos-immunoreactive neurons were evaluated by an observerignorant of the rat’s experimental status. Sections were viewedunder a 10�objective of a Leika DM 2500 microscope equippedwith a camera lucida, and a graticule (0.3�0.3 mm2 area�0.09mm2) was used to manually quantify the number of Fos-positiveneurons in the lateral and medial sectors of the SC at mid rostro-caudal levels, AP � 6.0 mm. For a cell to be considered asexpressing Fos-like immunoreactivity, the nucleus of the neuronshad to be of appropriate size (ranging approximately, from 8 to 15�m) and shape (oval or round), show the characteristic blue-blacktaining of oxidized DAB-Ni, and be distinct from the backgroundt magnification of �10. Both regions of interest were counted in
hree sections from each rat.The figures were prepared using the Adobe Photoshop CS3
version 10.0.1; Adobe Systems, Mountain View, CA, USA) forhotomicrographs and Adobe Illustrator CS3 (Version 13.0.1;dobe System, Mountain View, CA, USA) for line drawings. Onlyontrast and brightness were adjusted.
Statistical analysis
For behavioural measurements we used a one-way or two-wayanalysis of variance (ANOVA) with repeated measures. However,because the hold-duration data were not normally distributed com-parisons were made with the non-parametric Kolmogorov–Smir-nov test. Data reflecting the density of Fos labeled cells wereanalysed using the one-way ANOVA followed by the Fisher’sPLSD multiple comparisons test for independent samples for lat-eral or medial SC. In each case the significance level was set at��5%. All the values are expressed as means�SEM.
RESULTS
Behavioural analysis
The purpose of the behavioural analysis was to determinethe effects of removing rats’ whiskers on their ability to huntcockroaches. We will first describe the normal hunting ofrats with intact whiskers, before outlining the effects whis-ker removal had on this behaviour.
Rats were placed individually in the experimental cageafter which five intact mature cockroaches were intro-duced. Initially, a rat sniffed around the cage until a preywas located, often by the whiskers, at which point the ratrushed towards the cockroach and tried to capture it, eitherwith the forepaws or immediately with the mouth. Whilehunting the rats were able to orient themselves optimallywith respect to the prey and immediately after capture,rapidly manipulated the cockroach into a position where akilling bite could be directed towards the prey’s head. Moreoften than not the cockroach was then eaten. On someoccasions, rats with intact whiskers were so motivated theywould leave the cockroach they had just caught to catchanother that happened to distract it. Alternatively, the ratswith intact whiskers would sometimes appear to play withthe prey in much the same way as a cat would play with amouse—letting it escape only to re-capture it again. Re-sults of the quantitative analysis of hunting by rats withintact whiskers are presented in first data column of Table
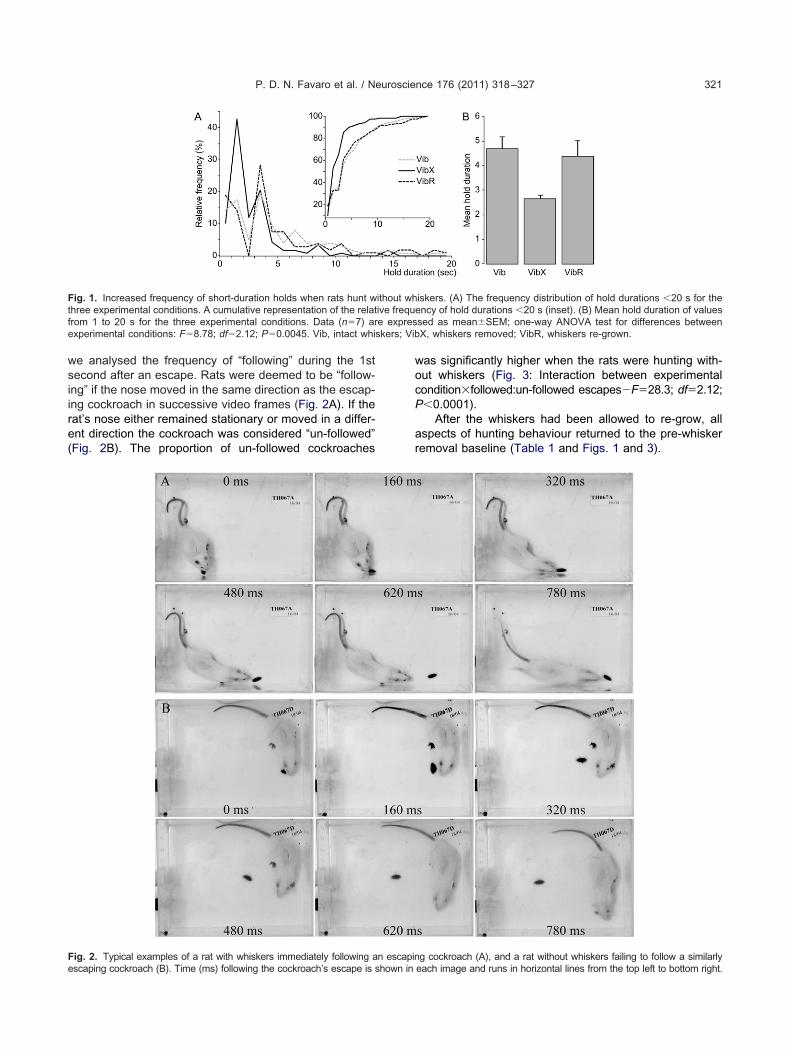
1 and Figs. 1 and 3.Despite already having had the experience of hunting,several aspects of the predatory behaviour of rats werequite different when their whiskers were removed. How-ever, we will describe first the similarities. Rats withoutwhiskers appeared equally well motivated to catch cock-roaches. Thus, in terms of latency to first attack, timeduring the session spent hunting, total time in contact withthe prey, together with the number of cockroaches caught,killed and consumed, rats without whiskers did not differsignificantly from the previous session when their whiskerswere intact (data column 2 of Table 1). However, despitebeing well motivated, rats without whiskers were clearlyless proficient in the sensorimotor aspects of hunting andprey capture. First, they appeared to have difficulty orient-ing themselves correctly towards the prey to facilitate anefficient strike, either with the mouth or the paws. Second,having caught or made contact with a cockroach they haddifficulty manipulating it into a position where it could beeasily killed. Thus, on many occasions the cockroach wasable to escape at which point the whiskerless rats oftenfailed to follow the escaping prey. To provide quantitativemeasures of these behavioural problems we measuredfirst, the duration prey were as held after each attack, andsecondly, in the event of the cockroach escaping, whetheror not it was immediately followed. Hold durations �20 swere excluded from the analysis, because by this time acaught cockroach was most often dead or injured, andtherefore unlikely to escape. The distribution of hold dura-tions across the three experimental sessions are illustratedin Fig. 1A. Very short-hold durations (1–2 s) were morefrequent when rats were hunting without whiskers, thuscompared with the initial control condition, the distributionof hold-durations �20 s was significantly different for ratswithout whiskers (Chi-square�28.035, P�0.0001), butnot after the whiskers were allowed to re-grow (Chi-square�1.626, P�0.8869). This was confirmed with ananalysis of the mean duration of holds �20 s which wassignificantly shorter for rats hunting without whiskers (Fig.1B; F�8.78; df�2.12; P�0.0045). These data are consis-tent with the failure of rats without whiskers being able toorient themselves appropriately during an attack and sub-sequently manipulating the prey to prevent its escape.However, the short-hold duration data confounded whis-kerless incompetence with “cat-and-mouse” play wherethe cockroach was “deliberately” released by rats with
Table 1. Behavioural measurements during experimental session
Behaviouralmeasurements
Experimental conditions (n�7)
Vib VibX VibR
Latency to attack (s) 18.9�4.6 25.7�6.9 11.3�6.0Hunt-duration (s) 235.3�71.2 262.0�67.1 234.3�66.5Total contact time (s) 467.6�33.9 429.0�64.7 338.9�41.0Total # of cockroaches-
killed/consumed1.2�0.1 1.3�0.1 1.1�0.1
Mean-attacks/kill 4.5�0.8 4.7�0.4 3.8�0.7
Values are mean�SEM. Differences between conditions were as-sessed by one-way repeated measures ANOVA.
whiskers, then only to be recaptured. To disambiguate this
P
ar
skers; Vib
e
P. D. N. Favaro et al. / Neuroscience 176 (2011) 318–327 321
we analysed the frequency of “following” during the 1stsecond after an escape. Rats were deemed to be “follow-ing” if the nose moved in the same direction as the escap-ing cockroach in successive video frames (Fig. 2A). If therat’s nose either remained stationary or moved in a differ-ent direction the cockroach was considered “un-followed”(Fig. 2B). The proportion of un-followed cockroaches
Fig. 1. Increased frequency of short-duration holds when rats hunt wthree experimental conditions. A cumulative representation of the relatfrom 1 to 20 s for the three experimental conditions. Data (n�7) arexperimental conditions: F�8.78; df�2.12; P�0.0045. Vib, intact whi
Fig. 2. Typical examples of a rat with whiskers immediately following a
scaping cockroach (B). Time (ms) following the cockroach’s escape is shown inwas significantly higher when the rats were hunting with-out whiskers (Fig. 3: Interaction between experimentalcondition�followed:un-followed escapes�F�28.3; df�2.12;
�0.0001).After the whiskers had been allowed to re-grow, all
spects of hunting behaviour returned to the pre-whiskeremoval baseline (Table 1 and Figs. 1 and 3).
iskers. (A) The frequency distribution of hold durations �20 s for theency of hold durations �20 s (inset). (B) Mean hold duration of valuessed as mean�SEM; one-way ANOVA test for differences betweenX, whiskers removed; VibR, whiskers re-grown.
g cockroach (A), and a rat without whiskers failing to follow a similarly
ithout whive freque expres
n escapin
each image and runs in horizontal lines from the top left to bottom right.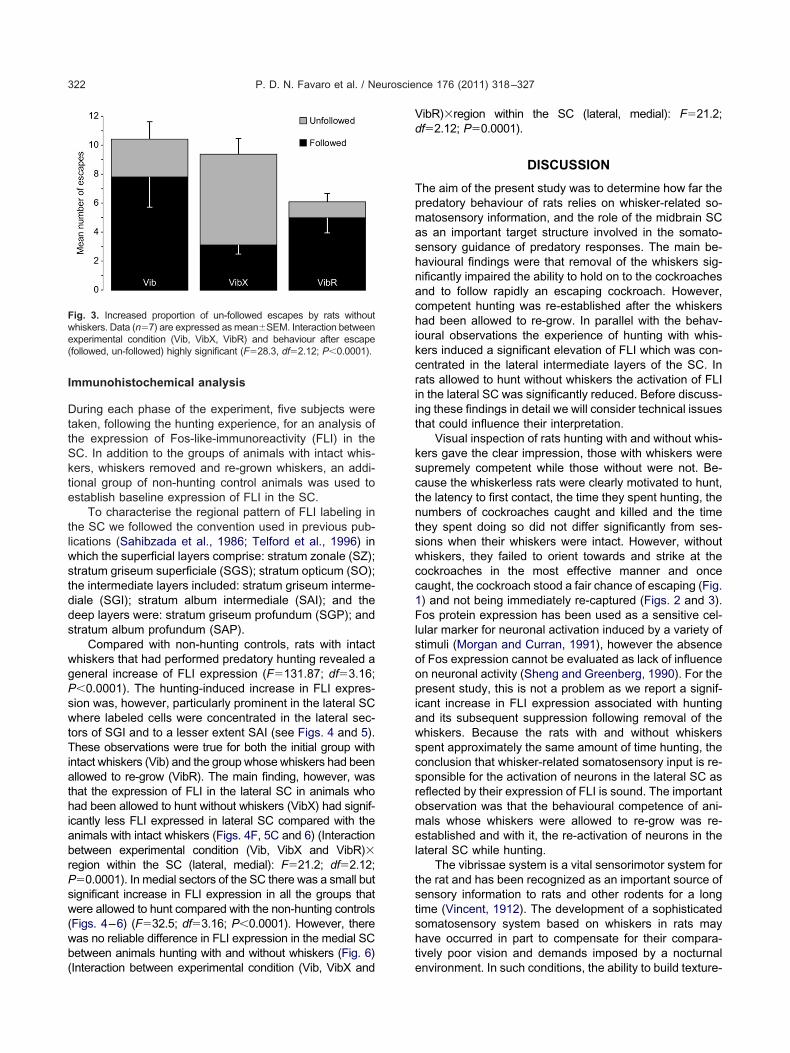
TiathiabrPsw(wb(
Vd
P. D. N. Favaro et al. / Neuroscience 176 (2011) 318–327322
Immunohistochemical analysis
During each phase of the experiment, five subjects weretaken, following the hunting experience, for an analysis ofthe expression of Fos-like-immunoreactivity (FLI) in theSC. In addition to the groups of animals with intact whis-kers, whiskers removed and re-grown whiskers, an addi-tional group of non-hunting control animals was used toestablish baseline expression of FLI in the SC.
To characterise the regional pattern of FLI labeling inthe SC we followed the convention used in previous pub-lications (Sahibzada et al., 1986; Telford et al., 1996) inwhich the superficial layers comprise: stratum zonale (SZ);stratum griseum superficiale (SGS); stratum opticum (SO);the intermediate layers included: stratum griseum interme-diale (SGI); stratum album intermediale (SAI); and thedeep layers were: stratum griseum profundum (SGP); andstratum album profundum (SAP).
Compared with non-hunting controls, rats with intactwhiskers that had performed predatory hunting revealed ageneral increase of FLI expression (F�131.87; df�3.16;P�0.0001). The hunting-induced increase in FLI expres-sion was, however, particularly prominent in the lateral SCwhere labeled cells were concentrated in the lateral sec-tors of SGI and to a lesser extent SAI (see Figs. 4 and 5).
hese observations were true for both the initial group withntact whiskers (Vib) and the group whose whiskers had beenllowed to re-grow (VibR). The main finding, however, washat the expression of FLI in the lateral SC in animals whoad been allowed to hunt without whiskers (VibX) had signif-
cantly less FLI expressed in lateral SC compared with thenimals with intact whiskers (Figs. 4F, 5C and 6) (Interactionetween experimental condition (Vib, VibX and VibR)�egion within the SC (lateral, medial): F�21.2; df�2.12;�0.0001). In medial sectors of the SC there was a small butignificant increase in FLI expression in all the groups thatere allowed to hunt compared with the non-hunting controls
Figs. 4–6) (F�32.5; df�3.16; P�0.0001). However, thereas no reliable difference in FLI expression in the medial SCetween animals hunting with and without whiskers (Fig. 6)
Fig. 3. Increased proportion of un-followed escapes by rats withoutwhiskers. Data (n�7) are expressed as mean�SEM. Interaction betweenexperimental condition (Vib, VibX, VibR) and behaviour after escape(followed, un-followed) highly significant (F�28.3, df�2.12; P�0.0001).
Interaction between experimental condition (Vib, VibX and
ibR)�region within the SC (lateral, medial): F�21.2;f�2.12; P�0.0001).
DISCUSSION
The aim of the present study was to determine how far thepredatory behaviour of rats relies on whisker-related so-matosensory information, and the role of the midbrain SCas an important target structure involved in the somato-sensory guidance of predatory responses. The main be-havioural findings were that removal of the whiskers sig-nificantly impaired the ability to hold on to the cockroachesand to follow rapidly an escaping cockroach. However,competent hunting was re-established after the whiskershad been allowed to re-grow. In parallel with the behav-ioural observations the experience of hunting with whis-kers induced a significant elevation of FLI which was con-centrated in the lateral intermediate layers of the SC. Inrats allowed to hunt without whiskers the activation of FLIin the lateral SC was significantly reduced. Before discuss-ing these findings in detail we will consider technical issuesthat could influence their interpretation.
Visual inspection of rats hunting with and without whis-kers gave the clear impression, those with whiskers weresupremely competent while those without were not. Be-cause the whiskerless rats were clearly motivated to hunt,the latency to first contact, the time they spent hunting, thenumbers of cockroaches caught and killed and the timethey spent doing so did not differ significantly from ses-sions when their whiskers were intact. However, withoutwhiskers, they failed to orient towards and strike at thecockroaches in the most effective manner and oncecaught, the cockroach stood a fair chance of escaping (Fig.1) and not being immediately re-captured (Figs. 2 and 3).Fos protein expression has been used as a sensitive cel-lular marker for neuronal activation induced by a variety ofstimuli (Morgan and Curran, 1991), however the absenceof Fos expression cannot be evaluated as lack of influenceon neuronal activity (Sheng and Greenberg, 1990). For thepresent study, this is not a problem as we report a signif-icant increase in FLI expression associated with huntingand its subsequent suppression following removal of thewhiskers. Because the rats with and without whiskersspent approximately the same amount of time hunting, theconclusion that whisker-related somatosensory input is re-sponsible for the activation of neurons in the lateral SC asreflected by their expression of FLI is sound. The importantobservation was that the behavioural competence of ani-mals whose whiskers were allowed to re-grow was re-established and with it, the re-activation of neurons in thelateral SC while hunting.
The vibrissae system is a vital sensorimotor system forthe rat and has been recognized as an important source ofsensory information to rats and other rodents for a longtime (Vincent, 1912). The development of a sophisticatedsomatosensory system based on whiskers in rats mayhave occurred in part to compensate for their compara-tively poor vision and demands imposed by a nocturnal
environment. In such conditions, the ability to build texture-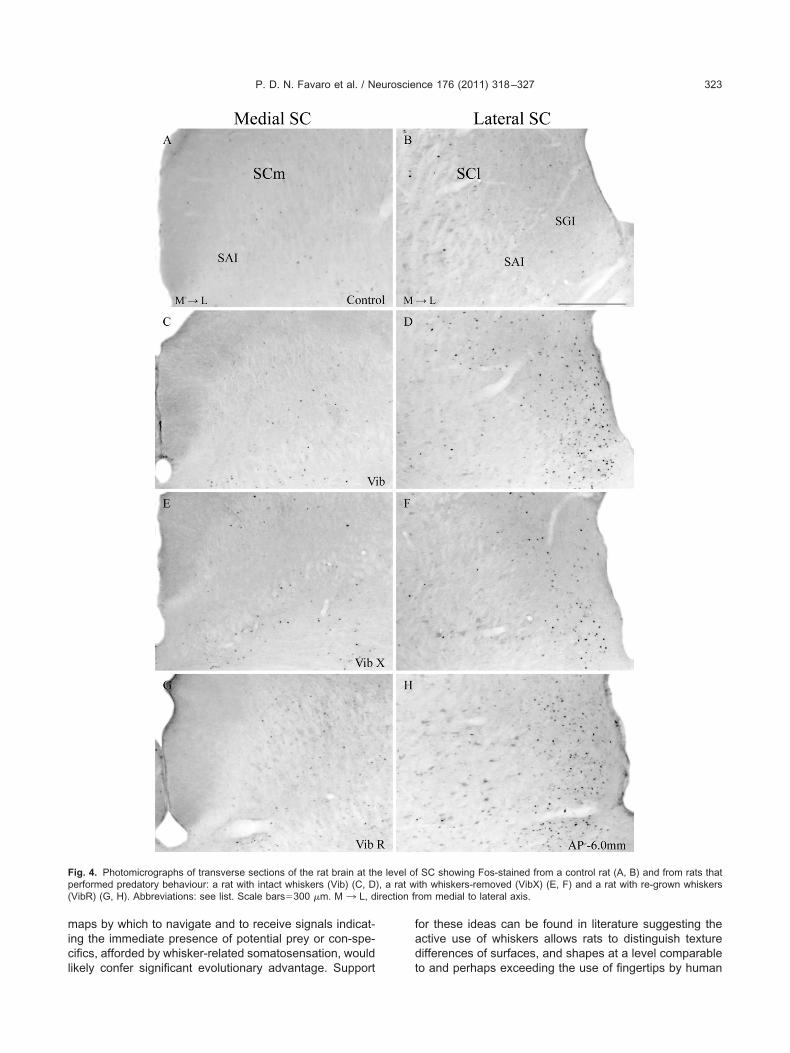
irection fr
P. D. N. Favaro et al. / Neuroscience 176 (2011) 318–327 323
maps by which to navigate and to receive signals indicat-ing the immediate presence of potential prey or con-spe-cifics, afforded by whisker-related somatosensation, would
Fig. 4. Photomicrographs of transverse sections of the rat brain at thperformed predatory behaviour: a rat with intact whiskers (Vib) (C, D(VibR) (G, H). Abbreviations: see list. Scale bars�300 �m. M ¡ L, d
likely confer significant evolutionary advantage. Support
for these ideas can be found in literature suggesting theactive use of whiskers allows rats to distinguish texturedifferences of surfaces, and shapes at a level comparable
f SC showing Fos-stained from a control rat (A, B) and from rats thatith whiskers-removed (VibX) (E, F) and a rat with re-grown whiskersom medial to lateral axis.
e level o), a rat w
to and perhaps exceeding the use of fingertips by human
xactly to s
P. D. N. Favaro et al. / Neuroscience 176 (2011) 318–327324
and non-human primates (Guic-Robles et al., 1989, 1992;Carvell and Simons, 1990; Prigg et al., 2002; Arabzadeh etal., 2005; Kleinfeld et al., 2006; Wolfe et al., 2008). More-over the sensitivity of the whisker system to direction andspeed of whisker deflections would be essential for guidingthe fore-paws or the mouth to a moving target (Brecht etal., 1997).
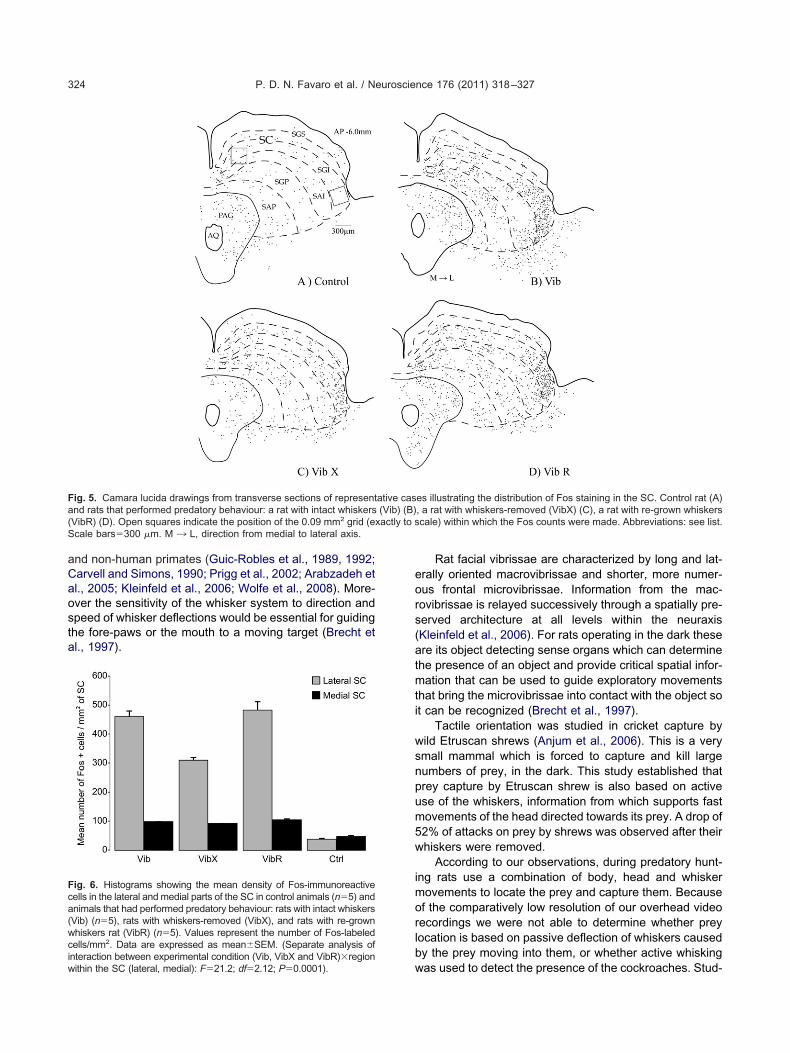
Fig. 5. Camara lucida drawings from transverse sections of representand rats that performed predatory behaviour: a rat with intact whiskers(VibR) (D). Open squares indicate the position of the 0.09 mm2 grid (eScale bars�300 �m. M ¡ L, direction from medial to lateral axis.
Fig. 6. Histograms showing the mean density of Fos-immunoreactivecells in the lateral and medial parts of the SC in control animals (n�5) andanimals that had performed predatory behaviour: rats with intact whiskers(Vib) (n�5), rats with whiskers-removed (VibX), and rats with re-grownwhiskers rat (VibR) (n�5). Values represent the number of Fos-labeledcells/mm2. Data are expressed as mean�SEM. (Separate analysis of
interaction between experimental condition (Vib, VibX and VibR)�regionwithin the SC (lateral, medial): F�21.2; df�2.12; P�0.0001).Rat facial vibrissae are characterized by long and lat-erally oriented macrovibrissae and shorter, more numer-ous frontal microvibrissae. Information from the mac-rovibrissae is relayed successively through a spatially pre-served architecture at all levels within the neuraxis(Kleinfeld et al., 2006). For rats operating in the dark theseare its object detecting sense organs which can determinethe presence of an object and provide critical spatial infor-mation that can be used to guide exploratory movementsthat bring the microvibrissae into contact with the object soit can be recognized (Brecht et al., 1997).
Tactile orientation was studied in cricket capture bywild Etruscan shrews (Anjum et al., 2006). This is a verysmall mammal which is forced to capture and kill largenumbers of prey, in the dark. This study established thatprey capture by Etruscan shrew is also based on activeuse of the whiskers, information from which supports fastmovements of the head directed towards its prey. A drop of52% of attacks on prey by shrews was observed after theirwhiskers were removed.
According to our observations, during predatory hunt-ing rats use a combination of body, head and whiskermovements to locate the prey and capture them. Becauseof the comparatively low resolution of our overhead videorecordings we were not able to determine whether preylocation is based on passive deflection of whiskers causedby the prey moving into them, or whether active whisking
es illustrating the distribution of Fos staining in the SC. Control rat (A), a rat with whiskers-removed (VibX) (C), a rat with re-grown whiskerscale) within which the Fos counts were made. Abbreviations: see list.
ative cas(Vib) (B)
was used to detect the presence of the cockroaches. Stud-

P. D. N. Favaro et al. / Neuroscience 176 (2011) 318–327 325
ies based on high speed video recordings showed thatobject localization by rats can certainly be achieved byactive touch (Knutsen et al., 2005; Towal and Hartmann,2006; Mitchinson et al., 2007; Ahissar and Knutsen, 2008;Grant et al., 2009) as well as for prey capture by theEtruscan shrew (Anjum et al., 2006). However, in ourexperimental situation it is likely that both active and pas-sive deflection of whiskers was used in prey capture; activewhen the animals were moving forward hunting and pas-sive when a cockroach had the misfortune to enter thewhisker field while the rat was either pausing or consuminga cockroach it had captured.
In our study we observed that VibX group often failedto initiate pursuit after a cockroach had escaped. Perhapsthis was because, in the absence of whiskers informationthe rat was unable to initiate the fast translational head andbody movements required for immediate re-capture (Fig.2B). In contrast, the groups with intact whiskers wouldoften immediately re-orient themselves in order to facilitatethe cockroache’s re-capture (Fig. 2A). Although our datademonstrate that whiskers provide important informationnecessary for effective prey detection and capture, furtherstudies with high resolution video capability are necessaryto clarify the extent to which active and passive deflectionof the whiskers may elicit different components of thepredatory response.
The SC is an ancient midbrain structure considered themammalian homologue of the optic tectum. It is widelyacknowledged that the SC is responsible for the sensori-motor transformations required to direct gaze towards oraway from unexpected, biologically salient events (Wurtzand Albano, 1980; Sparks, 1986; Dean et al., 1989; Steinand Meredith, 1993). The SC contains a variable numberof alternating cell and fiber rich layers (Kanaseki andSprague, 1974; Reiner, 1994; May, 2006), although a func-tional distinction is often made (Casagrande et al., 1972)between the superficial layers, which receive spatially or-ganized retinotopic input giving rise to a retinotopicallycoded map, and deep layers, which receive numerousmultisensory (including somatosensory input from thewhiskers) and non-sensory modulatory inputs (Stein andMeredith, 1993; Sefton and Dreher, 1995; Boehnke andMunoz, 2008). In rodents a critical distinction between themedial and lateral sectors of SC has been made on func-tional grounds (Dean et al., 1989). Thus, the ecology of therat is such that most of its predators (e.g. larger mammals,hawks, etc.), are detected initially as a movement in theupper visual field. Since in the retinotopic representationsit is the medial SC that such movement will be detected,appropriate stimulation of this region can elicit a fully inte-grated defensive reactions (Dean et al., 1988, 1991). Onthe other hand, appetitive stimuli for the rodent (prey,offspring, etc.) generally appear or can be found in thelower visual field which is represented in the lateral SC.Consequently, appropriate stimulation of this region typi-cally elicits approach and appetitive behaviours. Of partic-ular relevance to the present study, are reports demon-strating that integrity of the lateral SC is essential for
effective predatory hunting (Comoli and Canteras, 2000;Furigo et al., 2010). This work is extended by the presentinvestigation by showing that signals from the whiskersrelayed to this region are likely to be a critical componentof the circuitry responsible for efficient predation. In partic-ular, our behavioural and immunohistochemical resultshighlight the importance of whisker input to the lateral SCas a source of information necessary to target a prey andgenerate the movements required for a successful attack.
Confirming electrophysiological experiments in rodentshave shown that neurons in the lateral SC are responsiveto somatosensory stimuli applied to face and whiskers(Tiao and Blackemore, 1976; Dräger and Hubel, 1976).Further studies have demonstrated that the lateral SC isdifferently responsive to perioral somatosensory stimulithat can be used to guide objects towards the mouth in theform of a “bitting reflex” (Redgrave et al., 1990; Keay et al.,1990). There are several potential sources of whisker-related somatosensory input to the lateral SC. The mostobvious are the direct afferent projections from the brain-stem trigeminal system (Hemelt and Keller, 2007). Thusthe lateral SC receives whisker and orofacial-related so-matosensory information from both the SpV and PrV (Kil-lackey and Erzurumlu, 1981; Huerta et al., 1983; Rhoadeset al., 1989; Comoli and Canteras, 2000; Hemelt andKeller, 2007). Other indirect inputs originate from the zonaincerta (Kolmac et al., 1998; Comoli and Canteras, 2000),parvicelular reticular nucleus and somatosensory barrelcortex (Comoli and Canteras, 2000; Cohen et al., 2008).An important recent paper by Felsen and Mainen (2008)reported that the lateral SC is also critical for orienting toolfactory stimuli. Olfactory cues are likely to play a criticalmotivational role in predatory behavioural responses (Co-moli et al., 2005). Consequently it was not surprising toobserve that VibX group was highly motivated to hunt,showing no delays to start chasing the cockroaches andspending similar amounts of time hunting and engagingwith the prey. Their problem was one of competence. It istherefore likely that whiskers removal resulted in a loss ofeffective spatial localization of fast moving prey and con-sequent lack of specific sensory information required todirect orienting movements of head, body and mouth to-wards a moving target. Thus, our data are consistent withthe large body of work showing that phasic whisker-relatedinput to the lateral SC is specialised for the initial detectionof biologically salient stimuli that require immediate appet-itive action (Cohen et al., 2007, 2008; Hemelt and Keller,2007), including the detection and capture of cockroaches.
Acknowledgments—This research was supported by grants fromFundação de Amparo à Pesquisa do Estado de São Paulo (FA-PESP, n°. 07/03655-4) and Fundação Apoio ao Ensino e Pes-quisa e Assistência Hospital das Clínicas da Faculdade de Me-dicina de Ribeirão Preto awarded to E.C.. P.N.F., T.S.G., andS.R.O. were supported by FAPESP fellowships. P.R. was in re-ceipt of support from the Wellcome Trust.
REFERENCES
Ahissar E, Knutsen PM (2008) Object localization with whiskers. Biol
Cybern 98:449–458.
F
G
G
G
G
G
G
H
H
I
K
K
K
KK
K
K
M
M
M
M
P. D. N. Favaro et al. / Neuroscience 176 (2011) 318–327326
Anjum F, Turni H, Mulder PG, van der Burg J, Brecht M (2006) Tactileguidance of prey capture in Etruscan shrews. Proc Natl Acad SciU S A 103:16544–16549.
Arabzadeh E, Zorzin E, Diamond ME (2005) Neuronal encoding oftexture in the whisker sensory pathway. PLoS Biol 3:e17.
Boehnke SE, Munoz DP (2008) On the importance of the transientvisual response in the superior colliculus. Curr Opin Neurobiol18:1–8.
Brandão ML, Cardoso SH, Melo LL, Motta V, Coimbra NC (1994)Neural substrate of defensive behavior in the midbrain tectum.Neurosci Biobehav Rev 18:339–346.
Brandão ML, Anseloni VZ, Pandóssio JE, De Araújo JE, Castilho VM(1999) Neurochemical mechanisms of the defensive behavior inthe dorsal midbrain. Neurosci Biobehav Rev 23:863–875.
Brecht M, Preilowski B, Merzenich MM (1997) Functional architectureof the mystacial vibrissae. Behav Brain Res 84:81–97.
Carvell GE, Simons DJ (1990) Biometric analyses of vibrissal tactilediscrimination in the rat. J Neurosci 10:2638–2648.
Casagrande VA, Harting JK, Hall WC, Diamond IT, Martin GF (1972)Superior colliculus of the tree shrew: a structural and functionalsubdivision into superficial and deep layers. Science 177:444–447.
Cohen JD, Hirata A, Castro-Alamancos MA (2007) Early sensorypathways for detection of fearful conditioned stimuli: tectal andthalamic relays. J Neurosci 27:7762–7776.
Cohen JD, Hirata A, Castro-Alamancos MA (2008) Vibrissa sensationin superior colliculus: wide-field sensitivity and state-dependentcortical feedback. J Neurosci 28:11205–11220.
Coizet V, Comoli E, Westby GW, Redgrave P (2003) Phasic activationof substantia nigra and the ventral tegmental area by chemicalstimulation of the superior colliculus: an electrophysiological inves-tigation in the rat. Eur J Neurosci 17:28–40.
Comoli E, Canteras NS (1998) Characterization of the neural systemsmobilized during predatory behavior in rats. In: Proceedings of the28th Annual Meeting of the Society for Neuroscience, vol. 24, p678; November 1998. Los Angeles, CA. Abstract 265.17.
Comoli E, Canteras NS (2000) Lateral region of intermediate layer ofsuperior colliculus: a key site involved in the motor control ofpredatory hunting. In: Proceedings of the 30th Annual Meetingof the Society for Neuroscience, vol. 26, p 2257; November 2000.New Orleans, LA. Abstract 847.5.
Comoli E, Coizet V, Boyes J, Bolam JP, Canteras NS, Quirk RH,Overton PG, Redgrave P (2003) A direct projection from superiorcolliculus to substantia nigra for detecting salient visual events. NatNeurosci 6:974–980.
Comoli E, Ribeiro-Barbosa ER, Negrão N, Goto M, Canteras NS(2005) Functional mapping of the prosencephalic systems involvedin organizing predatory behavior in rats. Neuroscience 130:1055–1067.
Dean P, Mitchell IJ, Redgrave P (1988) Contralateral head movementsproduced by microinjection of glutamate into superior colliculus ofrats: evidence for mediation by multiple output pathways. Neuro-science 24:491–500.
Dean P, Redgrave P, Westby GWM (1989) Event or emergency? Tworesponse systems in the mammalian superior colliculus. TrendsNeurosci 12:137–147.
Dean P, Simkins M, Hetherington L, Mitchell IJ, Redgrave P (1991)Tectal induction of cortical arousal: evidence implicating multipleoutput pathways. Brain Res Bull 26:1–10.
Dräger UC, Hubel DH (1976) Topography of visual and somatosen-sory projections to mouse superior colliculus. J Neurophysiol39(1):91–101.
Ewert JP, Buxbaum-Conradi H, Glagow M, Röttgen A, Schürg-PfeifferE, Schwippert WW (1999) Forebrain and midbrain structures in-volved in prey-catching behaviour of toads: stimulus-response me-diating circuits and their modulating loops. Eur J Morphol 37(2–3):
172–176.Felsen G, Mainen ZF (2008) Neural substrates of sensory-guidedlocomotor decisions in the rat superior colliculus. Neuron60:137–148.
urigo IC, de Oliveira WF, de Oliveira AR, Comoli E, Baldo MVC,Mota-Ortiz SR, Canteras NS (2010) The role of supeior colliculusin predatory behavior. Neuroscience 165:1–15.
erfen CR, Sawchenko PE (1984) An anterograde neuroanatomicaltracing method that shows the detailed morphology of neurons,their axons and terminals: immunohistochemical localization of anaxonally transported plant lectin, Phaseolus vulgaris leucoag-glutinin (PHA-L). Brain Res 290(2):219–238.
rant RA, Mitchinson B, Fox CW, Prescott TJ (2009) Active touchsensing in the rat: anticipatory and regulatory control of whiskersmovements during surface exploration. J Neurophysiol 101:862–874.
rantyn R (1988) Gaze control through superior colliculus: structureand function. In: Neuroanatomy of the oculomotor system (Buttner-Ennever JA, ed), pp 273–333. Amsterdam: Elsevier.
robstein P (1988) Between the retinotectal projection and directedmovement: topography of a sensorimotor interface. Brain BehavEvol 31:34–48.
uic-Robles E, Valdivieso C, Guajardo G (1989) Rats can learn aroughness discrimination using only their vibrissal system. BehavBrain Res 31(3):285–289.
uic-Robles E, Jenkins WM, Bravo H (1992) Vibrissal roughnessdiscrimination is barrelcortex-dependent. Behav Brain Res 48(2):145–152.
emelt ME, Keller A (2007) Superior sensation: superior colliculusparticipation in rat vibrissa system. BMC Neurosci 8:12.
uerta MF, Frankfurter A, Harting JK (1983) Studies of the principalsensory and spinal trigeminal nuclei of the rat: projections to thesuperior colliculus, inferior olive, and cerebellum. J Comp Neurol220(2):147–167.
toh K, Konishi A, Nomura S, Mizuno N, Nakamura Y, Sugimoto T(1979) Application of coupled oxidation reaction to electron micro-scopic demonstration of horseradish peroxidase: cobalt-glucoseoxidase method. Brain Res 175(2):341–346.
anaseki T, Sprague JM (1974) Anatomical organization of pretectaland tectal laminae in the cat. J Comp Neurol 158:319–337.
eay K, Westby GW, Frankland P, Dean P, Redgrave P (1990)Organization of the crossed tecto-reticulo-spinal projection in rat–II. Electrophysiological evidence for separate output channels tothe periabducens area and caudal medulla. Neuroscience 37(3):585–601.
illackey HP, Erzurumlu RS (1981) Trigeminal projections to the su-perior colliculus of the rat. J Comp Neurol 201(2):221–242.
ing AJ (2004) The superior colliculus. Curr Biol 14(9):R335–R338.leinfeld D, Ahissar E, Diamond ME (2006) Active sensation: insights
from the rodent vibrissa sensorimotor system. Curr Opin Neurobiol16:435–444.
olmac CI, Power BD, Mitrofanis J (1998) Patterns of connectionsbetween zona incerta and brainstem in rats. J Comp Neurol396(4):544–555.
nutsen PM, Derdikman D, Ahissar E (2005) Tracking whisker andhead movements in unrestrained behaving rodents. J Neuro-physiol 93(4):2294–2301.
ay PJ (2006) The mammalian superior colliculus: laminar structureand connections. Prog Brain Res 151:321–378.
itchinson B, Martin CJ, Grant RA, Prescott TJ (2007) Feedbackcontrol in active sensing: rat exploratory whisking is modulated byenvironmental contact. Proc R Soc B 274:1035–1041.
organ JI, Curran T (1991) Stimulus-transcription coupling in thenervous system: involvement of the inducible proto-oncogenes fosand jun. Annu Rev Neurosci 14:421–451.
unoz DP, Guitton D (1989) Fixation and orientation control by thetecto-reticulo-spinal system in the cat whose head is unrestrained.
Rev Neurol (Paris) 145:567–579.
P. D. N. Favaro et al. / Neuroscience 176 (2011) 318–327 327
Prigg T, Goldreich D, Carvell GE, Simons DJ (2002) Texture discrim-ination and unit recordings in the rat whiskers/barrel system.Physiol Behav 77:671–675.
Redgrave P, Dean P, Westby GW (1990) Organization of the crossedtecto-reticulo-spinal projection in rat–I. Anatomical evidence forseparate output channels to the periabducens area and caudalmedulla. Neuroscience 37(3):571–584.
Reiner A (1994) Laminar distribution of the cells of origin of ascendingand descending tectofugal pathways in turtles: implications for theevolution of tectal lamination. Brain Behav Evol 43(4–5):254–292.
Rhoades RW, Fish SE, Chiaia NL, Bennett-Clarke C, Mooney RD(1989) Organization of the projections from the trigeminal brains-tem complex to the superior colliculus in the rat and hamster:anterograde tracing with Phaseolus vulgaris leucoagglutinin andintra-axonal injection. J Comp Neurol 289(4):641–656.
Sahibzada N, Dean P, Redgrave P (1986) Movements resemblingorientation or avoidance elicited by electrical stimulation of thesuperior colliculus in rats. J Neurosci 6:723–733.
Sheng M, Greenberg ME (1990) The regulation and function of c-fosand other immediate early genes in the nervous system. Neuron4(4):477–485.
Sparks DL (1986) Translation of sensory signals into commands forcontrol of saccadic eye movements: role of the primate superiorcolliculus. Physiol Rev 66:118–171.
Sparks DL (1999) Conceptual issues related to the role of the superiorcolliculus in the control of gaze. Curr Opin Neurobiol 9:698–707.
Sefton AJ, Dreher B (1995) Visual system. In: The rat nervous system
(Paxinos G, ed), pp 833–898. Academic Press Inc.Stein BE, Meredith MA (1993) The merging of the senses. The MITPress.
Steiner H, Huston JP (1992) Control of behavior under apomorphineby sensory input from the face. Psychopharmacol (Berl) 109(4):390–394.
Telford S, Wang S, Redgrave P (1996) Analysis of nociceptive neu-rons in the rat superior colliculus using c-fos immunohistochemis-try. J Comp Neurol 375:601–617.
Tiao YC, Blackemore C (1976) Functional organization in the superiorcolliculus of the golden hamster. J Comp Neurol 168(4):483–503.
Towal RB, Hartmann MJ (2006) Right-left asymmetries in the whiskingbehavior of rats anticipate head movements. J Neurosci 26(34):8838–8846. Erratum in: J Neurosci 26(40):10079.
Vincent SB (1912) The function of vibrissae in the behavior of the whiterat. Behav Monogr 1:1–85.
Wang S, Redgrave P (1997) Microinjections of muscimol into lateralsuperior colliculus disrupt orienting and oral movements in theformalin model of pain. Neuroscience 81(4):967–988.
Welzl H, Schwarting R, Kulajta J, Huston JP (1984) Perioral bitting andturning after intranigral injection of a GABA- or metenkephalin-agonist: role of the thalamus and superior colliculus. Exp Brain Res55:438–444.
Wolfe J, Hill DN, Pahlavan S, Drew PJ, Kleinfeld D, Feldman DE(2008) Texture coding in the rat whisker system: slip-stick versusdifferential resonance. PLoS Biol 6(8):e215.
Wurtz RH, Albano JE (1980) Visual-motor function of the primatesuperior colliculus. Annu Rev Neurosci 3:189–226.
Wurtz RH (1996) Vision for the control of movement. The FriedenwaldLecture. Invest Ophthalmol Vis Sci 11:2130–2145.
(Accepted 9 December 2010)(Available online 14 December 2010)




















