
Subcortical connections of the superior colliculus in the mustache bat,Pteronotus parnellii
19
THE JOURNAL OF COMPARATIVE NEUROLOGY 263:179-197 (1987) Subcortical Connections of the Superior Colliculus in the Mustache Bat, Pteronotus parnellii E. COVEY, W.C. HALL, AND J.B. KOBLER Departments of Surgery (Otolaryngology) (E.C., J.B.K.) and Anatomy (W.C.H.), Duke University Medical Center, Durham, North Carolina 27710 ABSTRACT The mustache bat, Pteronotus parnellii, depends on echolocation to nav- igate and capture prey. This adaptation is reflected in the large size and elaboration of brainstem auditory structures and in the minimal develop- ment of visual structures. The superior colliculus, usually associated with orienting the eyes, is nevertheless large and well developed in Pteronotus. This observation raises the question of whether the superior colliculus in the echolocating bat has evolved to play a major role in auditory rather than visual orientation. The connections of the superior colliculus in Pteronotus were studied with the aid of anterograde and retrograde transport of wheat germ agglutinin conjugated to HRP. These results indicate that the superior colliculus of Pteronotus is composed almost entirely of the layers beneath stratum opticum. The retinal projection is restricted to a very thin zone just beneath the pial surface. Prominent afferent pathways originate in motor structures, particularly the substantia nigra and the deep nuclei of the cerebellum. Sensory input from the auditory system originates in three brainstem nuclei: the inferior colliculus, the anterolateral periolivary nu- cleus, and the dorsal nuclei of the lateral lemniscus. The projections from these auditory structures terminate mainly in the central tier of the deep layer. The most prominent efferent pathways are those to medial motor structures of the contralateral brainstem via the predorsal bundle and to the ipsilateral midbrain and pontine tegmentum via the lateral efferent bundle. Ascending projections to the diencephalon are mainly to the medial dorsal nucleus and zona incerta. Thus, the superior colliculus in Pteronotus possesses well-developed anatomical connections that could mediate reflexes for orienting its ears, head, or body toward objects detected by echolocation. Key words: tectum, auditory-motor pathways, sensorimotor integration The mustache bat, Pteronotus parnellii, like many other microchiropterans (e.g., Cotter, '85), has small, rudimen- tary eyes and an almost vestigial optic nerve but, at the same time, possesses a large, well-developed superior collic- ulus (see Fig. 1). The main function of the superior collicu- lus, or optic tectum, has traditionally been considered to be the orientation of the eyes toward the source of an external stimulus (Schneider, '69; Wurtz and Goldberg, '71; Sprague, '75; Raczkowski et al., '76; Sparks, '78; Wurtz and Albano, '80; Hall and May, '84). Because Pteronotus depends almost entirely on echolocation to navigate and capture prey, it seems likely that the large superior colliculus of this species is associated with guiding movements of its ears, head, and body on the basis of auditory cues. If this is true, the superior colliculus should have extensive connections with the highly developed auditory structures used for echoloca- tion (Zvorykin, '59; Henson, '70; Baron, '74; Zook and Casseday, '82a) and with motor structures that play a role in orientation. Therefore, this species may prove to be an excellent model to use in studying the representation of Accepted March 19, 1987. J.B. Kobler's current address is Eaton Peabody Laboratory, Mas- sachusetts Eye and Ear Infirmary, 243 Charles Street, Boston, MA 02114. 0 1987 ALAN R. LISS. INC.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Subcortical connections of the superior colliculus in the mustache bat,Pteronotus parnellii

THE JOURNAL OF COMPARATIVE NEUROLOGY 263:179-197 (1987)
Subcortical Connections of the Superior Colliculus in the Mustache Bat,
Pteronotus parnellii
E. COVEY, W.C. HALL, AND J.B. KOBLER Departments of Surgery (Otolaryngology) (E.C., J.B.K.) and Anatomy (W.C.H.),
Duke University Medical Center, Durham, North Carolina 27710
ABSTRACT The mustache bat, Pteronotus parnellii, depends on echolocation to nav-
igate and capture prey. This adaptation is reflected in the large size and elaboration of brainstem auditory structures and in the minimal develop- ment of visual structures. The superior colliculus, usually associated with orienting the eyes, is nevertheless large and well developed in Pteronotus. This observation raises the question of whether the superior colliculus in the echolocating bat has evolved to play a major role in auditory rather than visual orientation. The connections of the superior colliculus in Pteronotus were studied with the aid of anterograde and retrograde transport of wheat germ agglutinin conjugated to HRP. These results indicate that the superior colliculus of Pteronotus is composed almost entirely of the layers beneath stratum opticum. The retinal projection is restricted to a very thin zone just beneath the pial surface. Prominent afferent pathways originate in motor structures, particularly the substantia nigra and the deep nuclei of the cerebellum. Sensory input from the auditory system originates in three brainstem nuclei: the inferior colliculus, the anterolateral periolivary nu- cleus, and the dorsal nuclei of the lateral lemniscus. The projections from these auditory structures terminate mainly in the central tier of the deep layer. The most prominent efferent pathways are those to medial motor structures of the contralateral brainstem via the predorsal bundle and to the ipsilateral midbrain and pontine tegmentum via the lateral efferent bundle. Ascending projections to the diencephalon are mainly to the medial dorsal nucleus and zona incerta. Thus, the superior colliculus in Pteronotus possesses well-developed anatomical connections that could mediate reflexes for orienting its ears, head, or body toward objects detected by echolocation.
Key words: tectum, auditory-motor pathways, sensorimotor integration
The mustache bat, Pteronotus parnellii, like many other microchiropterans (e.g., Cotter, '85), has small, rudimen- tary eyes and an almost vestigial optic nerve but, at the same time, possesses a large, well-developed superior collic- ulus (see Fig. 1). The main function of the superior collicu- lus, or optic tectum, has traditionally been considered to be the orientation of the eyes toward the source of an external stimulus (Schneider, '69; Wurtz and Goldberg, '71; Sprague, '75; Raczkowski et al., '76; Sparks, '78; Wurtz and Albano, '80; Hall and May, '84). Because Pteronotus depends almost entirely on echolocation to navigate and capture prey, it seems likely that the large superior colliculus of this species is associated with guiding movements of its ears, head, and
body on the basis of auditory cues. If this is true, the superior colliculus should have extensive connections with the highly developed auditory structures used for echoloca- tion (Zvorykin, '59; Henson, '70; Baron, '74; Zook and Casseday, '82a) and with motor structures that play a role in orientation. Therefore, this species may prove to be an excellent model to use in studying the representation of
Accepted March 19, 1987. J.B. Kobler's current address is Eaton Peabody Laboratory, Mas-
sachusetts Eye and Ear Infirmary, 243 Charles Street, Boston, MA 02114.
0 1987 ALAN R. LISS. INC.

180 E. COVEY, W.C. HALL, AND J.B. KOBLER
auditory information in the mammalian 'tectum and in establishing connections between the auditory and motor systems in mammals.
With these ideas in mind, we initiated a series of experi- ments on the connections of the superior colliculus in Pter- onotus. The results show that the superior colliculus of Pteronotus is composed almost entirely of the layers be- neath stratum opticum. The pattern of efferent connections resembles that seen in all vertebrate species. The most prominent afferent connections originate in motor struc- tures; sensory input from auditory and somatosensory sources is relatively more prominent than visual input. Taken together, these results support the idea that the superior colliculus of Pteronotus is involved primarily in orienting to auditory stimuli.
MATERIALS AND METHODS Sixteen mustache bats, Pteronotus parnellii parnellii, were
used in this study. For comparison, an extensive collection of cases with various brainstem and cortical injections of anterograde and retrograde tracers was available. Before all procedures, animals were anesthetized with Metofane (Methoxyflurane). In the present study, wheat germ agglu- tinin conjugated to HRP (WGA-HRP) was injected electro- phoretically with micropipettes. Five animals had injections located in the superior colliculus, three in the inferior collic- ulus, two in the deep cerebellar nuclei, one in the anterola- teral periolivary nucleus, two in the dorsal nucleus of the lateral lemniscus, two in the retina, and one in the main sensory nucleus of the trigeminal nerve.
A 5% solution of WGA-HRP (from Dr. R.G. Carey, Barrow Neurobiological Institute, or Sigma) dissolved in physiolog- ical saline was placed in micropipettes with tip diameters ranging from 1 to 10 pm. The electrode was aimed at the target by using a combination of stereotaxic and visual techniques. In many cases, neural activity was recorded with the injection electrode to determine the auditory re- sponse properties of neurons at the injection site. The WGA- HRP was injected by applying pulses of 1-2-pamp current, positive at the electrode, for 10-20 minutes. The duration of each pulse was 7 seconds, with 7 seconds between pulses. After a survival period of 24 hours, the animals were per- fused with phosphate-buffered saline followed by 8% for- malin and 2% sucrose in phosphate buffer. These solutions were followed by 10% sucrose in phosphate buffer. The brains were sectioned at 40 pm in the frontal plane on a freezing microtome. The sections were generally divided into three sets: two sets were treated with tetramethylben- zidine (TMB) according to the method of Mesulam ('78) and one set was treated with diaminobenzidine (DAB) according to the method of LaVail ('72). One of the sets treated with TMB was counterstained with neutral red in order to ex- amine cytoarchitecture; the other set was left unstained to allow optimum visualization of the label. The set treated with DAB was counterstained with cresyl violet. Sections were examined microscopically and photographed with the aid of both lightfield and darkfield illumination. The loca- tions of labeled cells, axons, and terminals were mapped on drawings of the sections with a camera lucida.
ALP0 BC BIC BP Ce CG ~~
CN CP c u D DNLL F Ha HT I IC ICC ICp ICX INLL Int I 0 IP L LGv LL LR MD MG MNTB MRF NA NPC n .Vm n. Vsm
n. Vsp n. XI1 n. VII opt PC PG Po m PY RB RN RTP ~~~~
sc SCi SCP scs SN SOC Ve VIII VNLL VP X 21
Abbreviations
anterolateral periolivary nucleus brachium conjunctivum brachium of the inferior colliculus brachium pontis cerebellum central gray cochlear nucleus cerebral peduncle cuneiform nucleus dentate nucleus dorsal nucleus of the lateral lemniscus fastigial nucleus habenula habenulotegmental tract nucleus interpositus inferior colliculus central nucleus of the inferior colliculus pericentral nucleus of the inferior colliculus external nucleus of the inferior colliculus intermediate nucleus of the lateral lemniscus intralaminar nuclear group inferior olive interpeduncular nucleus lateral thalamic nuclear group ventral lateral geniculate body lateral lemniscus lateral reticular nucleus medial dorsal thalamic nucleus medial geniculate body medial nucleus of the trapezoid body mesencephalic reticular formation nucleus ambiguus nucleus of the posterior commissure motor nucleus of the trigeminal nerve main sensory nucleus of the trigeminal nerve (n.+t. = nucleus + tract) spinal nucleus of the trigeminal nerve nucleus of the hypoglossal nerve nucleus of the facial nerve stratum opticum posterior commissure pontine gray posterior thalamic nuclear group pretectum pyramidal body restiform body red nucleus nucleus reticularis tegmenti pontis superior colliculus intermediate layers of the superior colliculus deep layers of the superior colliculus superficial layers of the superior colliculus (s-tso = superficial gray layers + stratum opticum) substantia nigra superior olivary complex vestibular nucleus auditory nerve ventral nucleus of the lateral lemniscus ventral posterior thalamic nucleus vagus nerve zona incerta
RESULTS Organization of the superior colliculus -
The cytoarchitecture of the brainstem auditory nuclei in Figure 1 shows frontal sections through the superior col- Pteronotus has been described (Zook and Casseday, '82a); liculus of Pteronotus stained by three different methods. the nomenclature for other brainstem nuclei is based on Each method is useful because it reveals certain features of comparison with other species. the laminar organization better than do the others. Figure

CONNECTIONS OF THE BAT SUPERIOR COLLICULUS 181
Fig. 1. The layers of the superior colliculus in Pteronotus can be defined on the basis of their cytoarchitecture (A), myeloarchitecture (B), and histo- chemical reactivity (C). The left half of (A) is a lightfield photomicrograph of a Nissl-stained frontal section through the superior colliculus taken from a celloidin-embedded brain. B is a frontal section from a celloidin-embedded
brain stained to show fibers. C is a photomicrograph of a frontal section stained for cytochrome oxidase activity. This section is taken from a more rostra1 location than those in A and B. Calibration marks represent 500 pm. Shrinkage is greater in celloidinembedded material than in frozen, so the sections in A and B appear smaller than the one in C.

182 E. COVEY, W.C. HALL, AM) J.B. KOBLER

CONNECTIONS OF THE BAT SUPERIOR COLLICULUS 183
Fig. 3. Darkfield photomicrographs to show (A) the predorsal bundle as it leaves the superior colliculus and crosses the midline, (B) anterograde transport to the lateral pontine nuclei, and (C) anterograde transport to the inferior olive. To provide landmarks, the cerebral aqueduct is outlined in white in A, and the lower edge of the brainstem is outlined in C. The calibration mark is for all three photomicrographs and represents 500 pm
1A shows a Nissl-stained section accompanied by a drawing of the same section. Figure 1B shows a section stained for myelinated fibers with the Heidenhain method (Mallory, '381, and Figure 1C shows a section stained for cytochrome oxidase (Wong-Riley, '79). Our identification of the layers is based partly on their appearance in these stains and partly on their patterns of connections. In Pteronotus, the superfi- cial layers, strata zonale and griseum superficiale, together with stratum opticum extend less than 50 pm inward from the surface of the superior colliculus. This superficial region is very thin and we were unable to distinguish among its component layers. The superficial layers are most clearly seen in the Nissl-stained section in Figure 1A as a cell- sparse region on the surface of the superior colliculus. Fol- lowing injections of WGA-HRP in the retina, the superficial layers are clearly visible as a thin zone of labeled axons and terminals (see Fig. 71, corresponding to this cell-sparse region.
The intermediate layers are relatively large, with a total thickness of approximately 40 pm. They probably corre- spond to strata griseum and album intermediale in other mammals. In Nissl-stained sections it is possible to distin- guish an outer granular layer that contains small, densely packed cells and, beneath it, an inner layer of larger, more sparsely distributed cells. The outer granular layer is heav- ily labeled after an injection in the deep cerebellar nuclei
Fig. 2. Drawings to illustrate the distribution of anterograde transport after an injection of WGA-HRP in the superior colliculus. The sections are arranged from rostral to caudal starting at the upper left-hand corner. The injection site is shown in black; the stippled area indicates the region of dense label surrounding the injection site. Anterograde transport is shown as fine dots and labeled fibers are shown as stippled lines. The arrow in section 92 points to the crossing of the predorsal bundle. The boxes on sections 92, 86, and 20 indicate the approximate regions shown in the photomicrographs of Figure 3.
(see Fig. 7). In sections stained for cytochrome oxidase, three sublayers within the intermediate zone can easily be distinguished. The outermost layer (IVa) is homogeneously stained and corresponds to the granular layer seen in Nissl- stained material. In the middle layer (Nb), many fiber bundles running rostrocaudally appear as small, round, light-colored patches. In fiber-stained sections (Fig. lB), the bundles in layer IVb appear as round, dark patches. The inner layer (IVc) contains a few darkly stained multipolar cells and no light patches in sections processed for cyto- chrome oxidase. Sublayers of the intermediate zone with similar features have been described in other species (Wie- ner, '86).
The lower layers probably correspond to stratum album intermediale and to strata griseum and album profundum in other mammals. In Pteronotus they are approximately 500-600 pm thick and are unusually well differentiated. There are many fiber bundles running mediolaterally throughout the deep layer, giving it a striped appearance. The fiber bundles are clearly visible as light stripes in Nissl-stained sections and in sections processed for cyto- chrome oxidase. Conversely, they appear as black stripes in fiber-stained sections. The deep layers contain many large multipolar cells; these cells are segregated in a band run- ning through the middle of the deep layer and can be seen in Nissl-stained sections (Fig. 1A). This central tier, con- taining the large cells, is the main site of termination of the auditory structures (see below).
Connections of the superior colliculus Efferent projections. The overall distribution of efferent
projections from the superior colliculus in Pteronotus is illustrated in Figure 2. Because this injection is large and the labeling is heavy, anterograde and retrograde transport are shown separately to avoid confusion. The injection of WGA-HRP is confined mostly to the intermediate and deep layers of the rostral superior colliculus. Although the supe- rior colliculus in Pteronotus is the source of projections to both the diencephalon and brainstem, just as it is in other

92
8
RE
TRO
GR
AD
E
Figu
re 4

CONNECTIONS OF THE BAT SUPERIOR COLLICULUS 185
mammalian species, in Pteronotus the projections to struc- tures associated with the visual system are sparse while projections to certain brainstem motor structures are un- usually prominent.
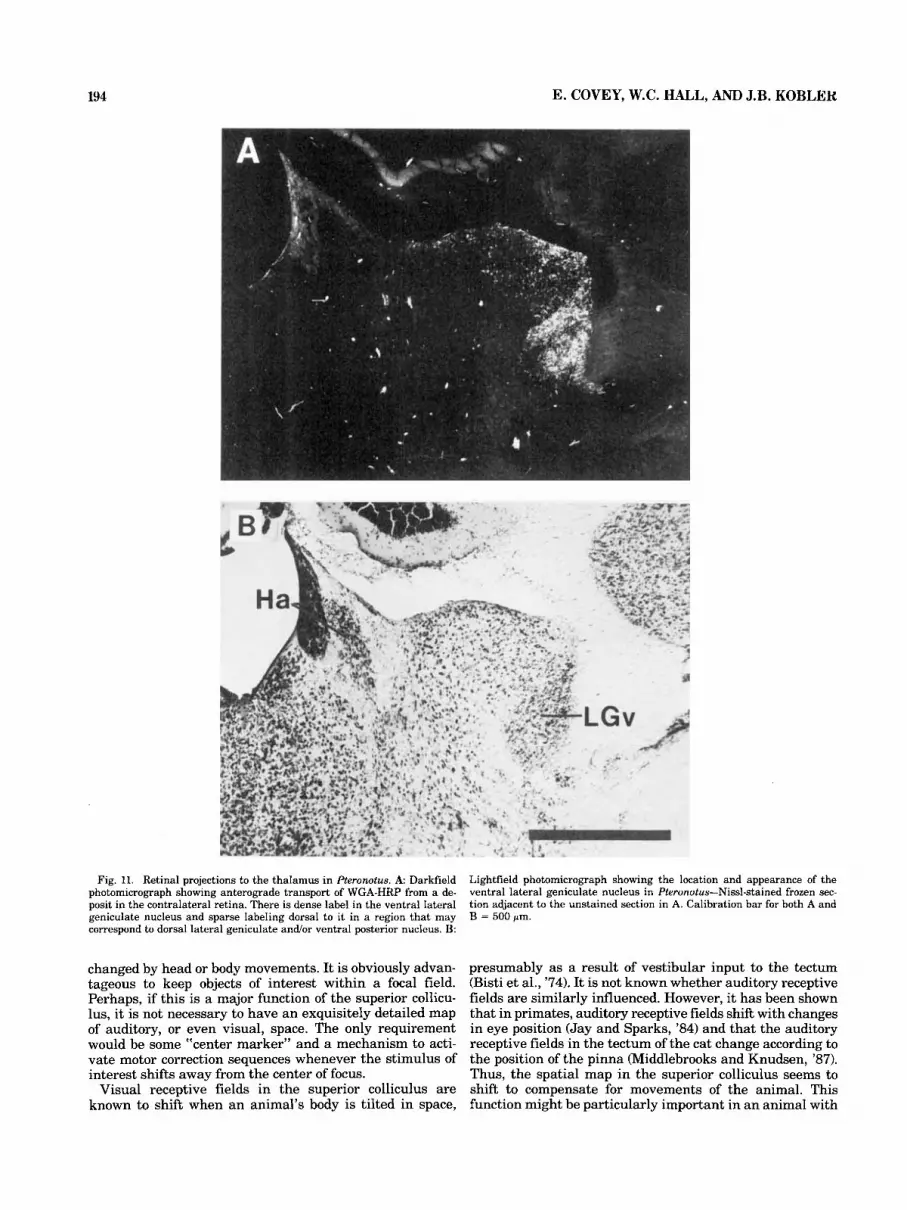
Ascending projections to the diencephalon. In the dorsal thalamus, a small amount of label is present just lateral to the intralaminar group (section 128, Fig. 2). Part of this tectal-recipient zone may correspond to the lateral poste- rior nucleus and pulvinar of other species. Some of the cells in this region of the thalamus receive sparse retinal projections (Fig. 11) and therefore may form a poorly differ- entiated dorsal lateral geniculate nucleus. In fact, Cotter (‘85) identifies this entire region as the dorsal lateral ge- niculate body in the bats Eptesicus and Artibeus. The pro- jections to certain other thalamic areas are prominent, especially those to the medial dorsal nucleus, to the intra- laminar areas (Fig. 2, section 1281, and to a region just medial to the medial geniculate body (Fig. 2, section 103). It is likely that this latter zone includes the supragenicu- late nucleus (Fig. 6) and posterior nuclear group (Figs. 2, 6) as defined in other species (e.g., Morest, ’65; Burton and Jones, ’76). In this experiment, the medial dorsal nucleus is labeled on both sides. In the ventral thalamus, a ventral lateral geniculate nucleus can be identified in Pteronotus on the basis of its position (Jones, ’85) and its reception of dense retinal projections (see Fig. 11). However, following the injection illustrated in Figure 2, as well as other injec- tions that included a large portion of the superficial layers (see Fig. 6), neither anterograde nor retrograde label can be detected in this area. Bilateral projections to zona in- certa are moderately dense (section 128, Fig. 2). The con- tralateral fibers in this pathway appear to cross the midline in the supraoptic decussation.
Ipsilateral to the injection site, shown in Figure 2, the label is most dense in the lateral part of the tegmentum. The main terminal zone extends from the lateral midbrain tegmentum (section 103) to the pontine tegmentum at the level of the superior olivary complex (section 62). Especially prominent ipsilat- era1 pathways can be traced to the ventral pons, including the nucleus reticularis tegmenti pontis (section 77), to the lateral pontine nuclei (section 86), and to the reticular for- mation medial to the superior olivary complex (section 62). This latter projection does not include any of the periolivary cell groups. In the lateral tegmentum there are labeled cells in the paralemniscal area and labeled fibers, probably en route to the reticular formation and pons (see Fig. 6, sec- tions 109 and 104). There is no cell group in Pteronotus that can be identified with certainty as the parabigeminal nu- cleus on the basis of cytoarchitecture or connections.
Contralateral to the injection site are pathways to the opposite superior colliculus, mainly the deep layers, and to the mesencephalic reticular formation (Fig. 2, section 103). Most of these projections appear t o cross the midline via
Descending projections to the brainstem.
Fig. 4. Drawings to illustrate the distribution of retrogradely labeled cells after an injection of WGA-HRP in the superior colliculus. This is the same case as that shown in Figure 2. The sections are arranged from rostral to caudal beginning at the upper left. The injection site is depicted as in Figure 2. The boxes on sections 115,95, 77, 34, and 20 indicate the approxi- mate regions shown in the photomicrographs of Figure 5.
the tectal commissure. The most prominent descending pathway to the contralateral brainstem crosses in the dor- sal tegmental decussation near the red nucleus (Fig. 2, section 92, and Fig. 3A) and descends toward the pons adjacent to the midline as the predorsal bundle. As in other mammals, fibers leave at right angles along the course of the predorsal bundle and terminate in the adjacent medial reticular formation of the pons (Fig. 2, sections 20-77). Other fibers descend ventrally and terminate in the nu- cleus reticularis tegmenti pontis (Fig. 2, section 77; Fig. 3B). One striking feature of this pathway in Pteronotus is the very dense projection to the inferior olive (Fig. 2, section 20; Fig. 3C).
Afferent projections. Figure 4 illustrates the locations of cells labeled by retrograde transport in the same case shown in Figure 2. Rostra1 to the superior colliculus, two structures contain labeled cells. The first (Fig. 4, section 128) lies in the subthalamus just beneath the ventral nu- clear group of the dorsal thalamus and probably corre- sponds to the zona incerta of other species. This is the only diencephalic area found to contain labeled cells. The ven- tral lateral geniculate nucleus, most surprisingly, is devoid of labeled cells. In the pretectum, a small region near the posterior commissure contains labeled cells (Fig. 4, section 115, and Fig. 5D); this group of cells is probably the nucleus of the posterior commissure.
Caudal to the superior colliculus large numbers of labeled cells are present in the inferior colliculus ipsilateral to the injection (Fig. 4, section 77; Fig. 5B). The labeled cells in the external nucleus of the inferior colliculus are multipo- lar cells and are among the largest neurons in the inferior colliculus. Another group of labeled cells is located ven- trally and medially, in the central nucleus. These include many small cells typical of this region as well as some large multipolar cells similar to those in the external nucleus. In some sections the labeled cells in the central nucleus to- gether with those in the external nucleus form a continuous semicircle. In other sections they form a vertical band in the ventromedial part of the inferior colliculus. Labeled cells are also found in the most rostral part of the inferior colliculus, in the area described by some authors (e.g., Meh- ler, ’69; RoBards et al., ’79) as the “intercollicular zone.” A few labeled cells are found contralateral to the injection in both the central and external nuclei of the inferior colliculus.
Two additional brainstem areas associated with the au- ditory system contain labeled cells. The dorsal nucleus of the lateral lemniscus contains labeled cells bilaterally. Most of the labeled cells are located in the most rostral and medial part of this nucleus (Fig. 4, section 77). The antero- lateral periolivary nucleus ipsilateral to the injection also contains many labeled cells (Fig. 4, section 77, and Fig. 5F). This cell group lies just anterior to the medial superior olive and posterior and ventromedial to the ventral nucleus of the lateral lemniscus.
One additional area that may have close associations with the auditory system projects to the superior colliculus. After an injection in the superior colliculus, many labeled cells are found in the lateral tegmental (or paralemniscal) region ventral to the inferior colliculus and medial and rostral to the dorsal nucleus of the lateral lemniscus (see Fig. 6, section 109).
In other parts of the midbrain, many labeled cells are present bilaterally. They are especially numerous in sub-

Figure 5

CONNECTIONS OF THE BAT SUPERIOR COLLICULUS 187
stantia nigra pars reticulata on both sides, although the density is highest ipsilateral to the injection (Fig. 4, sec- tions 103 and 95, and Fig. 5E). Labeled cells are found bilaterally in the reticular formation and contralaterally in the deep layers of the superior colliculus opposite to the injection (Fig. 4, section 95). In the brainstem, mainly con- tralateral to the injection site, small neurons in the main sensory nucleus of the trigeminal nerve are labeled (section 56, Fig. 4). Many densely labeled cells are present in the fastigial and dentate nuclei of the cerebellum (Fig. 4, sec- tion 34, and Fig. 5A). The labeled cells in the dentate nucleus are mostly contralateral to the injection. In the fastigial nucleus, labeled cells are somewhat more numer- ous on the ipsilateral side. The most caudal brainstem area that contains a large number of labeled cells is the contra- lateral perihypoglossal region (Fig. 4, section 20; Fig. 6, section 44; Fig. 5C). Most of these labeled cells probably belong to the nucleus prepositus hypoglossi.
Figure 6 illustrates a second case with both retrograde and anterograde transport from an injection of WGA-HRP located in the caudal superior colliculus. We present it to supplement the case just illustrated because it includes parts of the superficial, intermediate, and deep layers but is smaller than the previous injection and farther removed from the midline. Overall, the patterns of both anterograde and retrograde transport are very similar to those already shown. For this reason, only the most important differences will be addressed. One main difference is that the projec- tions to the thalamus are virtually all ipsilateral. In the previous case there are bilateral projections to the medial dorsal nucleus that we attribute to the slight intrusion of the injection to the opposite side Wig. 4, section 95). The only other difference is the absence of labeled cells in the sensory nucleus of the trigeminal nerve and in the contra- lateral superior colliculus. Since we have found evidence for such projections in other cases, we attribute their ab- sence in this case to the more restricted injection site.
Laminar distribution of inputs to the superior colliculus
The laminar distribution of the inputs to the superior colliculus provides a basis for establishing relationships between the layers in Pteronotus and those in other mam- mals. Figure 7A shows a thin band of anterograde label
Fig. 5. Darkfield photomicrographs to illustrate labeled cells after an injection of WGA-HRP in the superior colliculus. A Labeled cells in the deep cerebellar nuclei. The dentate nucleus contralateral to the injection is illustrated, as are the fastigial nuclei ofboth sides. B: Cells in the ipsilateral inferior colliculus. Laterally, there is a band of large labeled cells in the external nucleus; ventrally and medially these merge with a group of smaller cells to form a continuous semicircle that may extend into the central nucleus. The ventral border and the lateral edge of the inferior colliculus are outlined. C: Labeled cells in the contralateral perihypoglossal area. The outline indicates the border of the fourth ventricle. D: Labeled cells in the nuclei of the posterior commissure bilaterally. The cerebral aqueduct is outlined to provide a landmark. E: Labeled cells in the substan- tia nigra ipsilateral to the injection. There are large labeled cells laterally and smaller labeled cells medially. The outlines show the substantia nigra and the cerebral peduncle below it. F: Labeled cells in the anterolateral periolivary nucleus ipsilateral to the injection (outlined). Dorsomedial to the labeled cells there is an area in the pontine gray that contains antero- grade label. The calibration mark in A is for all the photomicrographs and represents 500 pm.
near the surface of the superior colliculus after an injection of WGA-HRP in the eye. This band marks the limits of the superficial retinorecipient layers in Pteronotw. In contrast, Figure 7B illustrates anterograde label in the superior col- liculus after a large injection of WGA-HRP in the contralat- era1 dentate and interpositus nuclei of the cerebellum. The label occupies a broad band just ventral to the zone that receives visual projections. This dense band of label coin- cides almost exactly with the outer granular sublayer (IVa) of the intermediate gray layer. The region of sparse projec- tions below it corresponds to the inner sublayers (IVb and IVc) of the intermediate gray layer. Below these sublayers are moderately dense projections throughout the deep layer and into the central gray. After a much smaller injection in the dentate nucleus, label is confined to the granular tier of the intermediate layer in the superior colliculus contra- lateral to the injection site.
The three drawings in Figure 8 show the patterns of label that result from injections in the brainstem auditory struc- tures that project to the superior colliculus in Pteronotus. Figure 8A shows the distribution of label following an injec- tion of WGA-HRP in the anterolateral periolivary nucleus. The label in the tectum is ipsilateral to the injection and is sparse in all parts of the intermediate layers. In the deep layers, there are large heavily labeled axons surrounding cells, especially in the sublamina containing the large mul- tipolar cells. Figure 8B illustrates a case with a large injec- tion of WGA-HRP that involves both the external and central nuclei of the caudal inferior colliculus. The deep tectal layers contain light, uniformly distributed label. Very little label is present in the intermediate layers, and vir- tually none is present in the granular sublayer ma) . There is no label contralateral to the injection site. Figure 8C shows the label resulting from an injection of WGA-HRP in the dorsal nucleus of the lateral lemniscus. The label is present bilaterally, is of approximately equal density on the two sides, and is most dense in the deep layers. There is some scattered label in the intermediate layers. Large, heavily labeled axons can be seen swirling around the somata of large multipolar cells in the deep layer (Fig. 9). Thus, the primary target of all the auditory structures that project to the superior colliculus appears to be the deep layer, although there are sparse projections to the interme- diate layer as well.
To illustrate projections from one source of somatosensory input to the superior colliculus, Figure 10 shows the results of an injection of WGA-HRP centered in the main sensory nucleus of the spinal trigeminal nerve. There is patchy label in the contralateral superior colliculus, mostly in the intermediate layer, and it is most dense in the lateral part. In addition, there are a few labeled cells in the deep layer. Ipsilateral to the injection there is sparse label in the most lateral part of the intermediate layer and along the ventral border of the deep layer.
DISCUSSION The main purpose of this discussion is to compare the
pattern of tectal connections in Pteronotus with those in other species and to discuss the significance of this pattern for understanding the organization and functions of the superior colliculus. In the sections that follow, we shall consider first the efferent pathways, emphasizing those that are either absent or especially well developed in Pteronotus. We shall then consider the sources of input to the superior colliculus, their patterns of termination, if known, and the

188 E. COVEY, W.C. HALL, AND J.B. KOBLER

CONNECTIONS OF THE BAT SUPERIOR COLLICULUS 189
role of connections in establishing homologies between the layers in different species. In a final section, we shall dis- cuss the comparative anatomy of the superior colliculus as it relates to function.
Efferent pathways in Pteronotus Ascending pathways. The superior colliculus is the
source of ascending pathways to both the dorsal and the ventral thalamus; however, in Pteronotus these pathways are restricted primarily to those known to arise in the layers beneath stratum opticum (e.g., Harting et al., '73; Kawamura and Kobayashi, '75; Huerta and Harting, '84). In the dorsal thalamus the major projections are to the intralaminar nuclei, the medial dorsal nucleus, and the posterior nuclear group, including the suprageniculate nu- cleus. In other species the medial dorsal nucleus and intra- laminar group are known to receive projections from the deep tectal layers. These nuclei contain cells with responses temporally linked to the onset of eye movements, and they project to the frontal eye fields (Schlag and Schlag-Rey, '84; Schlag-Rey and Schlag, '84). It seems unlikely that Ptere notus would have well-developed frontal eye fields. How- ever, the frontal cortex in Pteronotus receives a rich audi- tory input as well as projections from the medial dorsal nucleus, intralaminar group, and suprageniculate nucleus (Kobler et al., '87). Thus, in the bat these pathways from the tectum to the frontal cortex via the dorsal thalamus may provide a link between auditory and motor functions involving the head and ears, analogous to the visuomotor functions of the frontal cortex in other mammals.
In most species the lateral thalamus receives extensive input from the superior colliculus, but in Pteronotus this projection is very sparse. The lateral posterior nucleus-if it is represented at all in Pteronotus-must receive very little tectal input. This conclusion is not surprising since these projections arise in the superficial layers (Harting et al., '73; Graham, '77; Hall and May, '84)-layers that are poorly developed in Pteronotus.
In the ventral thalamus, zona incerta receives a modest input while the area we define as the ventral lateral genic- ulate nucleus, on the basis of retinal projections and posi- tion, appears to receive no projections from the superior colliculus. The connections of the ventral lateral geniculate body are a particular puzzle in Pteronotus. In most mam- mals, the ventral lateral geniculate nucleus is regarded as part of the visual system. It receives projections from the retina (Giolli and Guthrie, '62; Laties and Sprague, '66; Mathers and Mascetti, '75), the superficial layers of the superior colliculus (Altman and Carpenter, '61; Harting et al., '73; Benevento and Fallon, '75; Albano et al., '79), and the visual cortex (Garey et al., '68; Hollander, '70; Graybiel, '74). It projects in turn to the deep tectal layers (Edwards et al., '74; Graybiel, '74; Swanson et al., '74; Kawamura et al., '78). In Pteronotus, a group of cells can be identified in a
Fig. 6. Drawings to illustrate anterograde and retrograde transport of WGA-HRP after an injection in the caudal superior colliculus. The stippling indicates anterograde transport, the large dots labeled cells. Sections are arranged from rostra1 to caudal beginning at the upper left.
position corresponding to that of the ventral lateral genic- ulate in other species (Jones, '85), and this region does receive retinal projections (Fig. 11). Moreover, our case with an injection of WGA-HRP in the dentate nucleus of the cerebellum (case 874) indicates that, as in other mammals (Graybiel, '74), the ventral lateral geniculate nucleus of Pteronotus receives input from the cerebellum. However, our evidence suggests that in Pteronotus the ventral lateral geniculate neither receives projections from nor sends fi- bers to the tectum. Even in cases such as the one illustrated in Figure 2, with large numbers of labeled cells throughout the brainstem, the ventral lateral geniculate nucleus is devoid of labeled cells. This result is surprising since the deep tectal layers are a major target of the ventral lateral geniculate nucleus in other species (Edwards et al., '74; Graybiel, '74; Swanson et al., '74; Kawamura et al., '78). Because of this apparent species difference, determining the further connections of the ventral lateral geniculate nucleus in Pteronotus may be of particular value for under- standing its functions.
The results on the ascending pathways suggest a general conclusion: the diencephalic nuclei traditionally associated with vision have little or no connection with the superior colliculus in Pteronotus, while nuclei that are not specifi- cally visual but are associated with the deeper, premotor layers of the tectum have prominent connections.
Descending pathways. In Pteronotus, both ipsilateral and contralateral descending pathways are prominent, and both have courses and distributions similar to those of other species. Once again, the main difference appears to be the sparseness or absence of pathways associated with the vi- sual system. For example, in most mammals, a prominent pathway to the parabigeminal nucleus arises primarily from the superficial layers (e.g., Graham, '77; Holcombe and Hall, '81). This pathway could not be identified in Pterone tus. In contrast, other pathways, such as those to the nu- cleus reticularis tegmenti pontis and to the inferior olive, are at least as prominent in Pteronotus as they are in most other species that have been described (e.g., Harting, '77; Weber and Harting, '78). The pathway to the lateral pon- tine nuclei is of particular interest because in most mam- mals this projection originates mainly in the superficial layers (Graham, '77; Holcombe and Hall, '81), but neverthe- less is quite prominent in Pteronotus. If this pathway is concerned with relaying sensory information to the cerebel- lum, the shift in the laminar origin of the projections in Pteronotus may be related both to the reduction of visual input to the superficial layers and to the increase in audi- tory input to the deep layers. The inferior olive, the lateral pontine nuclei, and the nucleus reticularis tegmenti pontis all project to the cerebellum (Hoddevik et al., '76; Frank- furter et al., '77; Weber and Harting, '78; Brodal and Bro- dal, '81) and all are very prominent in Pteronotus. If we are correct in surmising that the superior colliculus in Ptere notus plays a role in orienting the bat's body, head, or ears during echolocation, these pathways to the cerebellum must make an important contribution, perhaps reflecting the special demands of detecting and correcting errors in flight by using auditory feedback.
A projection to the lateral tegmentum, or paralemniscal area, is prominent in Pteronotus. It has been suggested that this area plays a role in the control of pinna movements in the cat (Henkel and Edwards, '78), and this may be true in the bat as well.

190 E. COVEY, W.C. HALL, AND J.B. KOBLER
Fig. 7. Drawings and photomicrographs to illustrate the patterns of projections to the superior colliculus from (A) the contralateral retina; (B) an injection in the contralateral dentate nucleus of the cerebellum. C and D are darkfield photomicrographs of the sections illustrated in A and B. Calibration marks are 500 pm.
Afferent pathways in Pteronotus The superior colliculus in Pteronotus receives inputs from
various sensory modalities as well as from structures that probably modulate the premotor responses of deep tectal cells. These latter include prominent pathways from the deep cerebellar nuclei, the substantia nigra pars reticulata, and the nucleus prepositus hypoglossi. In most mammals these pathways are important for oculomotor reflexes, but our evidence suggests that their function may be different in the bat. The responses of cells in the substantia nigra pars reticulata of primates are linked to saccadic eye move- ments toward the source of an auditory stimulus; these cells fire when a sound stimulus is presented (Hikosaka and Wurtz, '83). Cells in the nucleus prepositus hypoglossi not only fire before visual saccades but also in response to neck movements (Gresty and Baker, '76). This cell group receives projections from the vestibular nuclei (Takagi, '25; Baker and Berthoz, '75) and cerebellum (Walberg, '61; Angaut and Brodal, '68) and may represent an important stage in ves- tibular, somatosensory, and motor integration.
The sources of sensory input to the tectum in Pteronotus include the retina, the sensory nucleus of the trigeminal nerve, and several brainstem auditory structures. Because the retinal input is limited to an extremely thin superficial zone we conclude that the visually mediated component of tectal function in Pteronotus must be of relatively minor importance. The nucleus of the trigeminal nerve is at least
one source of somatosensory input. This projection appears to be comparable in its magnitude and distribution to that in the rat (Killackey and Erzurumlu, '81; Huerta et al., '83) and cat (Baleydier and Mauguiere, '78; Nagata and Kruger, '79; Huerta et al., '81). The external nucleus of the inferior colliculus is another potential source of somatosensory pro- jections (e.g., Blomqvist et al., '78; RoBards et al., 1979; Bjorklund and Boivie, '84).
Auditory input to the superior colliculus in Pteronotus and other species
In most mammals the auditory projections to the tectum appear to be rather sparse. Possibly as a consequence of this sparseness, conflicting results have been reported re- garding their origins (e.g., Moore and Goldberg, '63, '66; Tarlov and Moore, '63; Casseday et al., '76; Edwards et al., '79; Kudo and Niimi, '80; Kudo, '81; Henkel, '83). Our results indicate that at least three distinct areas in the brainstem of Pteronotus contribute auditory input to the superior colliculus: the inferior colliculus, the dorsal nu- cleus of the lateral lemniscus, and the anterolateral perioli- vary nucleus.
Fibers projecting from the inferior to the superior collicu- lus of the cat were described long ago by Held (1893) and Cajal ('11). Later studies based on anterograde degenera- tion produced a variety of results. Some failed to demon-

CONNECTIONS OF THE BAT SUPERIOR COLLICULUS 191
strate such projections at all (e.g., Moore and Goldberg, '63; Casseday et al., '76); others indicated that the projection is at most a sparse one (Tarlov and Moore, '63; Moore and Goldberg, '66); and one (Powell and Hatton, '69) reported that, in the cat, lesions in all regions of the inferior collicu- lus, including those confined to the central nucleus, re- sulted in f 'strong topographically organized projections" to the superior colliculus. More recent experiments using re- trograde transport of HRP and anterograde transport of 3H- leucine have indicated that in the cat there are projections from the inferior to the superior colliculus but that they originate in the external and pericentral regions, not in the central nucleus (Edwards et al., '79; Kudo and Niimi, '80).
Our results indicate that in the bat the central and exter- nal nuclei of the inferior colliculus as well as the intercollic- ular zone project to the superior colliculus. Zook and Casse- day ('82a) have described an area in the ventromedial part of the inferior colliculus of Pteronotus that is sparsely pop- ulated with large multipolar cells and separates the medial from the anterolateral division of the central nucleus. This area receives input directly from the cochlear nucleus as well as from other brainstem auditory structures (Zook and Casseday, '85, '87). It appears to be the main, though not exclusive, source of projections from the central nucleus to the superior colliculus of Pteronotus. Because the central nucleus, external nucleus, and intercollicular zone each receive projections from a different subset of auditory nuclei (e.g., Aitkin et al., '78; RoBards et al., '79; Casseday and Covey, '87), these subdivisions may represent three very different sources of auditory input to the superior colliculus, with each source transmitting information that has under- gone a different type of processing.
Previous studies have reported projections from the dor- sal nucleus of the lateral lemniscus (DNLL) to the superior colliculus of the cat (Kudo, '81; Henkel, '831, tree shrew (Casseday et al., '79), and rat (Tanaka et al., '85). In con- trast, Edwards et al. ('79) reported that in the cat there was no such projection. One possible explanation for this dis- crepancy is that the region Edwards et al. ('79) called the paralemniscal area is equivalent to a region that other authors have included within the rostral region of DNLL. In Pteronotus we found labeled cells extending from approx- imately the middle to the most rostral and medial parts of DNLL after WGA-HRP injections in the superior colliculus. Tanaka et al. ('85) reported that in the rat the dorsal nu- cleus of the lateral lemniscus contains two separate popu- lations of cells-a caudal group that projects to the inferior colliculus and a rostral group that projects to the superior colliculus. Our results show that this is also true in PtercF notus. Most previous studies indicate that although the pathway from DNLL is bilateral, more cells project contra- laterally than ipsilaterally. This is also the case in Pteronotus.
There is very little evidence regarding auditory projec- tions to the superior colliculus from sources caudal to the inferior colliculus and DNLL. Our evidence identifies a distinct cell group, the anterolateral periolivary nucleus (ALPO), as a source of auditory projections to the superior colliculus in Pteronotus. ALPO consists of very large mul- tipolar cells that are easily distinguished from the small, densely packed multipolar cells in the ventral nucleus of the lateral lemmscus (VNLL) (Zook and Casseday, '82b). ALPO receives projections directly from the contralateral cochlear nucleus (see Zook and Casseday, '82b; Covey et a1 '84).
ALPO in the bat is almost certainly homologous to "pos- teromedial VNLL," a group of large cells reported to project
to the superior colliculus in the cat (Edwards et al., '79; Henkel, '83). Further support for this homology comes from the finding that in both the cat and bat this cell group projects to the suprageniculate nucleus of the dorsal thala- mus (Henkel, '83; Kobler et al., '87) as well as to the tectum.
The pathway from ALPO to the tectum has the capacity to provide relatively short-latency auditory input to the superior colliculus. In fact, latencies for auditory responses in the superior colliculus range from 3.6 msec up to 29 msec (e.g., Wise and Irvine, '83; Jen et al., '84; Wong, '84; Middle- brooks and Knudsen, '84). This wide range in latencies may reflect the diverse sources of auditory input, ranging fro=, multisynaptic pathways via the cortex to the pathway via ALPO which may involve only a single synapse between the cochlear nucleus and the tectum.
Edwards et al. ('79) reported projections to the superior colliculus from the dorsomedial periolivary nucleus and the ventral nucleus of the trapezoid body in the cat. We saw no evidence in the bat for projections from either of these sources.
The laminar organization of the superior colliculus in Pteronotus
The morphological features revealed by cytoarchitecture and the cytochrome oxidase stain indicate that the parts we call the intermediate and deep layers in Pteronotus correspond to strata griseum and album intermediale and profundum in other species (Wiener, '86). In Pteronotus as in other species the cerebellum projects to the intermediate and deep layers (Kawamura et al., '82; Uchida et al., '83; May and Hall, '86). In Pteronotus the cerebellotectal fibers terminate most densely in the intermediate layer and less densely in the deeper layers. In all species that have been studied, including Pteronotus, the auditory projections ter- minate mainly in the deep layers and sparsely in the inter- mediate layers. Pteronotus further resembles other species in that projections from the sensory nucleus of the trigemi- nal nerve terminate most densely in the lateral part of the intermediate layer. Thus, to the extent that we know the laminar distribution of projections in Pteronotus, it seems to be the same as in other species.
The terminal distributions from other sources-for exam- ple, the substantia nigra pars reticulata-remain to be es- tablished in Pteronotus. Until this information is available it is impossible to make more detailed comparisons. Since Pteronotus has unusually well-developed deep layers, one contribution of future studies in the bat might be to draw attention to laminar differences in connections that are not easily recognized in the intermediate and deep layers of other species.
The possible role of the superior colliculus in echolocating bats
One might expect that sound would be the most effective sensory stimulus for neurons in the deep layers of the superior colliculus in the bat. Indeed, in the few electro- physiological studies that have been conducted on bats, the authors apparently had no difficulty eliciting responses from tectal units by using pure tones or frequency-modulated sweeps (Jen et al., '84; Wong, '84). However, they do not report what percentage of cells responded to auditory stim- uli or whether "auditory" cells also responded to other stimulus modalities as they do in other species (e.g., Mere- dith and Stein, '86). Such information would be very useful for comparative purposes.
There is evidence that the intermediate and deep layers of the superior colliculus may be a stage in an ascending

A
ICX
Fig. 8. Drawings to show projections to the superior colliculus from auditory structures. A. Trans- port from the ipsilateral anterolateral periolivary nucleus (ALPO). B: Transport from an injection in the ipsilateral inferior colliculus. C: Bilateral transport from the dorsal nucleus of the lateral lemnis- cus. Calibration marks represent 500 prn.

CONNECTIONS OF THE BAT SUPERIOR COLLICULUS 193
pathway that contains a map of auditory space (e.g., Gor- don, '73; Drager and Hubel, '75; Chalupa and Rhoades, '77; Harris et al., '80; Middlebrooks and Knudsen, '84; King and Palmer, '85). However, the precision of the auditory spatial map in most mammals does not match that of the barn owl (Knudsen, '82). Since both the sources and termi- nal distribution of auditory input to the superior colliculus in Pteronotus are prominent and well defined relative to other mammals that have been described (e.g., Powell and Hatton, '69; Kudo and Niimi, '801, one might expect echo- locating bats to have a precise map of auditory space, com- parable to that in the owl. Certainly, the deep layers in Pteronotus receive input from a variety of sources that could provide precise information about the location of an auditory stimulus. Nevertheless, in the other species of echolocating bats that have been examined, it appears that the auditory spatial map is no more precise than it is in species that do not echolocate; moreover, it appears to rep- resent a relatively small region of auditory space that ex- tends outward only about 40" from the point where the vertical and horizontal axes intersect. This field represents the region directly in front of the bat's head and corre- sponds roughly to the field of the emitted pulse (Simmons, '69; Shimozawa et al., 74; Jen et al., '84; Wong, '84). In a general way, this auditory focal area might be compared to the visual field seen by the fovea of a primate. In the bat the relation Of the focal area to the animal's body can be changed by pinna movements, comparable, perhaps, to changing the direction of gaze by eye movements in a primate. Naturally, as with direction of gaze, it can also be
Fig. 9. Darkfield photomicrograph to show details of endings on large cells in the deep layer of the superior colliculus after an injection of WGA- HRP in the dorsal nucleus of the lateral lemniscus. Arrows point to two of these large endings. Calibration bar is 100 pm.
Fig. 10. Projections to the superior colliculus from the main sensory nucleus of the trigeminal nerve. A: Anterograde transport is mostly confined to the intermediate layer of the contralateral superior colliculus. It has a patchy distribution and is most dense in the lateral part. There are a few labeled cells in the deep layer of the superior colliculus. B: Injection site. Calibration mark is for both panels and represents 500 pm.
194 E. COVEY, W.C. HALL, AND J.B. KOBLEK
Fig. 11. Retinal projections to the thalamus in Pteronotus. A Darkfield photomicrograph showing anterograde transport of WGA-HRP from a de- posit in the contralateral retina. There is dense label in the ventral lateral geniculate nucleus and sparse labeling dorsal to i t in a region that may correspond to dorsal lateral geniculate and/or ventral posterior nucleus. B:
Lightfield photomicrograph showing the location and appearance of the ventral lateral geniculate nucleus in Pteronotus-Nissl-stained frozen sec- tion adjacent to the unstained section in A. Calibration bar for both A and B = 500 pm.
changed by head or body movements. It is obviously advan- tageous to keep objects of interest within a focal field. Perhaps, if this is a major function of the superior collicu- lus, it is not necessary to have an exquisitely detailed map of auditory, or even visual, space. The only requirement would be some "center marker" and a mechanism to acti- vate motor correction sequences whenever the stimulus of interest shifts away from the center of focus.
Visual receptive fields in the superior colliculus are known to shift when an animal's body is tilted in space,
presumably as a result of vestibular input to the tectum (Bisti et al., '74). It is not known whether auditory receptive fields are similarly influenced. However, it has been shown that in primates, auditory receptive fields shift with changes in eye position (Jay and Sparks, '84) and that the auditory receptive fields in the tectum of the cat change according to the position of the pinna (Middlebrooks and Knudsen, '87). Thus, the spatial map in the superior colliculus seems to shift to compensate for movements of the animal. This function might be particularly important in an animal with

CONNECTIONS OF THE BAT SUPERIOR COLLICULUS 195
a high degree of three-dimensional mobility, such as the bat.
CONCLUSIONS Comparative anatomy suggests that the efferent tectal
pathways, particularly those that descend to motor areas of the brainstem, are very constant not only among mammals but throughout vertebrates (Rubinson, '68; Foster and Hall, '75; Ebbeson and Vanegas, '76; Smeets, '81; Hall and May, '84). On the other hand, the tectum may vary dramatically both in terms of the sensory modalities that project to it and the sources of these projections. Among the modalities that project to the tectum are the visual, somatosensory, in- frared, electroreceptive, and auditory (Drager and Hubel, '75; Kass et al., '78; Stein and Gaither, '81). Furthermore, in some species, the major source of these inputs may be the cerebral cortex rather than the brainstem (Casseday et al., '79; Berson and McIlwain, '83; Stein et al., '83). This suggests that the main function of the superior colliculus, as evidenced by the relative constancy of its premotor effer- ent pathways, has remained similar throughout verte- brates but that the specific sensory modalities that influence or initiate activity in these premotor pathways can vary. In other words, orienting responses may remain the constant function of the superior colliculus, but the stimuli that elicit these responses vary. Our studies suggest that in Pterone tus the superior colliculus is concerned primarily with ori- entation behavior directed through the auditory feedback process that occurs during echolocation.
ACKNOWLEDGMENTS E.C. was supported by NIH grant NS 21748 and NSF
grants BNS 82-17357 and BNS 85-20441. W.C.H. was sup- ported by Public Health Service grant EY-04060 and NSF grants BNS 81-09794 and BNS-04060. We thank J.H. Casseday for advice and support in this study and Karen Gates and Melanie Colson for technical assistance. We also thank Drs. J.H. Casseday, T. Hughes, P.J. May, and L. Schweitzer for their helpful suggestions on earlier drafts of this manuscript.
and thalamic projections of dorsal column and lateral cervical nuclei: A quantitative study in the cat. Brain Res. 141.355-341,
Brodal, P., and A. Brodal (1981) The olivocerebellar projection in the mon- key. Experimental studies with the method of retrograde tracing of horseradish peroxidase. J. Comp. Neurol. 201:375-393.
Burton, H., and E.G. Jones (1976) The posterior thalamic region and its cortical projection in new world and old world monkeys. J. Comp. Neu- rol. 168.249-302.
Cajal, S. Ramon y. (1911) Histologie du Systeme Nerveux de l'Homme et des Vertebres. Paris: Maloine.
Casseday, J.H., and E. Covey (1987) Central auditory pathways in direc- tional hearing. In W.A. Yost and G. Gourevitch (eds): Directional Hear- ing. New York Springer Verlag (in press).
Casseday, J.H., I.T. Diamond, and J.K. Harting (1976) Auditory pathways to the cortex in Tupaia glis. J. Comp. Neurol. 166:303-340.
Casseday, J.H., D.R. Jones, and I.T. Diamond (1979) Projections from the cortex to the tectum in the tree shrew, Tupaia glis. J. Comp. Neurol. I85:253-292.
Chalupa, I.M., and R.W. Rhoades (1977) Responses of visual, somatosensory and auditory neurons in the golden hamster's superior colliculus. J. Physiol. nond.)270:595-626.
Cotter, J.R. (1985) Retinofugal projections of the big brown bat, Eptesicus fuscus and the neotropical fruit bat, Artibeus jamaicensis. Am. J. Anat. 172.105-124.
Covey, E., D.R. Jones, and J.H. Casseday (19841 Projections from the supe- rior olivary complex to the cochlear nucleus in the tree shrew. J. Comp. Neurol. 226.289-305.
Drager, U.C., and D.H. Hubel (1975) Responses to visual stimulation and relationship between visual, auditory and somatosensory inputs in mouse superior colliculus. J. Neurophysiol. 38;690-713.
Ebbeson, S.O., and H. Vanegas (1976) Projections of the optic tectum in two teleost species. J. Comp. Neurol. 165:161-180.
Edwards, S.B., A.C. Rosenquist, and L.A. Palmer (1974) An autoradio- graphic study of ventral lateral geniculate projections in the cat. Brain Res. 72.282-287.
Edwards, S.B., C.L. Ginsburg, C.K. Henkel, and B.E. Stein (1979) Sources of subcortical projections to the superior colliculus in the cat. J. Comp. Neurol. 184:309-330.
LITERATURE CITED Aitkin, L.M., H. Dickhaus, W. Schult, and M. Zimmerman (1978) External
nucleus of inferior colliculus: Auditory and spinal somatosensory affer- ents and their interactions. J. Neurophysiol. 41.83'7-847.
Albano, J.E., T.T. Norton, and W.C. Hall (1979) Laminar origin of projections from the superficial layers of the superior colliculus in the tree shrew, Tupaia glis. Brain Res. 175.1-11.
Altman, J., and M.B. Carpenter (1961) Fiber projections of the superior colliculus in the cat. J. Comp. Neurol. 116:157-178.
Angaut, P., and A. Brodal (1968) The projection of the vestibulocerebellum onto the vestibular nuclei in the cat. Arch. Ital. Biol. 105:441-479.
Baker, R., and A. Berthoz (1975) Is the prepositus hypoglossi nucleus the source of another vestibular ocular pathway? Brain Res. 86:121-127.
Baleydier, C., and Mauguiere (1978) Projections of the ascending somes- thetic pathways to the cat superior colliculus visualized by the horserad- ish peroxidase technique. Exp. Brain Res. 31:43-50.
Baron, G. (1974) Differential phylogenetic development of the acoustic nu- clei among Chiroptera. Brain Behav. Evol. 9.7-40.
Benevento, L.A., and J.H. Fallon (1975) The ascending projections of the superior colliculus in the rhesus monkey (Macaca mulatta). J. Comp. Neurol. 199:339-362.
Berson, D.M., and J.T. McIlwain (1983) Visual cortical inputs to the deep layers of the cat's superior colliculus. J. Neurophysiol. 50:1143-1155.
Bisti, S., L. Maffei, and M. Piccolino (1974) Visuovestibular interactions in the cat superior colliculus. J. Neurophysiol. 37;146-155.
Bjorklund, M., and J. Boivie (1984) An anatomical study of the projections from the dorsal column nuclei to the midbrain in cat. Anat. Embryol. 170:29-43.
Blomqvist, A,, R. Flink, D. Bowsher, S. Griph, and J. Westman (1978) Tectal 17.201-248,
Foster, R.E., and W.C. Hall (1975) The connections and laminar organiza- tion of the optic tectum in a reptile (Iguana iguana). J. Comp. Neurol. 163.39'7-426.
Frankfurter, A., J.T. Weber, and J.K. Harting (1977) Brainstem projection to lobule VII of the posterior vermis in the squirrel monkey: As demon- strated by the retrograde axonal transport of tritiated horseradish per- oxidase. Brain Res. 124:135-139.
Garey, L.J., E.G. Jones, and T.P.S. Powell (1968) Interrelationships of striate and extrastriate cortex with the primary relay sites of the visual path- way. J. Neurol. Neurosurg. Psychiatry 31:135-157.
Giolli, R.A., and M.D. Guthrie (1962) The primary optic projections in the rabbit: An experimental degeneration study. J. Comp. Neurol. 13699- 126.
Gordon, B. (1973) Receptive fields of deep layers of the cat superior collicu- lus. J. Neurophysiol. 36:157-178.
Graham, J. (1977) An autoradiographic study of the efferent connections of the superior colliculus in the cat. J. Comp. Neurol. 173:629-654.
Graybiel, A.M. (1974) Visuocerebellar and cerebello-visual connections in- volving the ventral lateral geniculate nucleus. Exp. Brain Res. 20.303- 306.
Gresty, M., and R. Baker (1976) Neurons with visual receptive field, eye movement and neck displacement sensitivity within and around the nucleus prepositus hypoglossi in the alert cat. Exp. Brain Res. 24.429- 433.
Hall, W.C., and P.J. May (1984) The anatomical basis for sensorimotor transformations in the superior colliculus. In W.D. Neff (ed): Contribu- tions to Sensory Physiology, Vol. 8. Orlando: Academic Press, pp. 1-40.
Harris, L.R., C. Blakemore, and M. Donaghy (1980) Integration of visual and auditory space in the mammalian superior colliculus. Nature 288:56-59.
Harting, J.K. (1977) Descending pathways from the inferior colliculus: An autoradiographic analysis in the rhesus monkey (Macaca mulattd. J. Comp. Neurol. 173.583-612.
Harting, J.K., W.C. Hall, I.T. Diamond, and G.F. Martin (1973) Anterograde degeneration study of the superior colliculus in Tupaia glis: Evidence for a subdivision between superficial and deep layers. J. Comp. Neurol. 148.361-386.
Held, H. (1893) Die centrale Gehorleitung. Arch. Anat. Physiol. Anat. Abt.

196
Henkel, C.K. (1983) Evidence of sub-collicular projections to medial genicu- late nucleus in the cat: An autoradiographic and horseradish peroxidase study. Brain Res. 259:21-30.
Henkel, C.K., and S.B. Edwards (1978) The superior colliculus control of pinna movements in the cat: Possible anatomical connections. J. Comp. Neurol. 182763-776.
Henson, O.W. (1970) The central nervous system. In W.A. Wimsatt (ed): Biology of Bats, Vol. 4. New York: Academic Press, pp. 95-142.
Hikosaka, 0.. and R.H. Wurtz (1983) Visual and oculomotor functions of monkey substantia nigra pars reticulata. I. Relation of visual and audi- tory responses to saccades. J. Neurophysiol. 49:1230-1252.
Hoddevik, G.H., A. Brodal, and F. Walberg (1976) The olivocerebellar projec- tion in the cat studied with the method of retrograde axonal transport of horseradish peroxidase. 111. The projection of the visual vermal area. J. Comp. Neural. 169:155-170.
Holcombe, V., and W.C. Hall (1981) Laminar origin of ipsilateral tectopon- tine pathways. Neuroscience 6:255-260.
Hollander, H. (1970) The projection from the visual cortex to the lateral geniculate body (LGB): An experimental study with silver impregnation methods in the cat. Exp. Brain Res. lOr219-235.
Huerta, M.F., A.J. Frankfurter, and J.K. Harting (1981) The trigeminocol- licular projection in the cat: Patch-like endings within the intermediate gray. Brain Res. 211:l-13.
Huerta, M.F., A. Frankfurter, and J.K. Harting (1983) Studies of the prin- cipal sensory and spinal trigeminal nuclei of the r a t Projections to the superior colliculus, inferior olive, and cerebellum. J. Comp. Neurol. 220: 147-167.
Huerta, M.F., and J.K. Harting (1984) The mammalian superior colliculus: Studies of its morphology and connections. In H. Vanegas (ed): Compar- ative Neurology of the Optic Tectum. New York: Plenum, pp. 687-773.
Jay, M.F., and D.L. Sparks (1984) Auditory receptive fields in primate superior colliculus shift with changes in eye position. Nature 309:345- 347.
Jen, P.H., X. Sun, T. Kamada, S. Zhang, and T. Shimozawa (1984) Auditory response properties and spatial response areas of superior collicular neurons of the FM bat, Eptesicus fuscus. J. Comp. Physiol. 154:407-413.
Jones, E.G. (1985) The Thalamus, New York: Plenum Press. Kass, L., M.S. Loop, and P.H. Hartline (1978) Anatomical and physiological
localization of visual and infrared cell layers in tectum of pit vipers J. Comp. Neurol. 182:811-820.
Kawamura, S., and E. Kobayashi (1975) Identification of laminar origin of some tecto-thalamic fibers in the cat. Brain Res. 91:281-285.
Kawamura, S., N. Fukushima, S. Hattori, and T. Tashior (1978) A ventral lateral geniculate nucleus projection to the dorsal thalamus and mid- brain in the cat. Exp. Brain Res. 3lt95-106.
Kawamura, S., S. Hattori, S. Higo, and T. Matsuyama (1982) The cerebellar projections to the superior colliculus and pretectum in the cat: An autoradiographic and horseradish peroxidase study. Neuroscience 7: 1673-1689.
Killackey, H.P., and R.S. Erzurumlu (1981) Trigeminal projections to the superior colliculus of the rat. J. Comp. Neurol. 201:221-242.
King, A.J., and A.R. Palmer (1985) Integration of visual and auditory information in bimodal neurones in the guinea pig superior colliculus. Exp. Brain Res. 60:492-500.
Knudsen, E.I. (1982) Auditory and visual maps of space in the optic tectum of the owl. J. Neurosci. 21177-1194.
Kobler, J.M., S.F. Isbey, and J.H. Casseday (1987) Auditory pathways to the frontal cortex of the mustache bat, Pteronotus parnellii. Science 236r824- 826.
Kudo, M. (1981) Projections of the lateral lemniscus in the cat: An autora diographic study. Brain Res. 22157-69.
Kudo, M., and K. Niimi (1980) Ascending projections of the inferior collicu- lus in the c a t An autoradiographic study. J. Comp. Neurol. 191r545- 556.
Laties, A.M., and J.M. Sprague (1966) The projection of optic fibers to visual centers in the cat. J. Comp. Neurol. 127r35-70.
LaVail, J.H., and M.M. LaVail (1972) Retrograde axonal transport in the central nervous system. Science 176:1416-1417.
Mallory, F.B. (1938) Pathological Technique: A Practical Manual for Work- ers in Pathological Histology Including Directions for the Performance of Autopsies and Microphotography. Philadelphia: W.B. Saunders.
E. COVEY, W.C. HALL, AND J.B. KOBLER
Ann. NY Acad. Sci. 169r424-468. Meredith, M.A., and B.E. Stein (1986) Visual, auditory, and somatosensory
convergence on cells in superior colliculus results in multisensory inte- gration. J. Neurophysiol. 56:640-662.
Mesulam, M.-M. (1978) Tetramethylbenzidine for horseradish peroxidase neurohistochemistry: A noncarcinogenic blue reaction product with su- perior sensitivity for visualizing afferents and efferents. J. Histochem. Cytochem. 26:106-117.
Middlebrooks, J.C., and E.I. Knudsen (1984) A neural code for auditory space in the cat's superior colliculus. J. Neurosci. 4r2621-2634.
Middlebrooks, J.C., and E.I. Knudsen (1987) Changes in ear position modify the spatial tuning of auditory units in the cat's superior colliculus. J. Neurophysiol. 57r672-687.
Moore, R.Y., and J.M. Goldberg (1963) Ascending projections of the inferior colliculus in the cat. J. Comp. Neurol. 121t109-136.
Moore, R.Y., and J.M. Goldberg (1966) Projections of the inferior colliculus in the monkey. Exp. Neurol. 14t429-438.
Morest, D.K. (1965) The lateral tegmental system of the midbrain and the medial geniculate body: Study with Golgi and Nauta methods in cat. J. Anat. 99:611-634.
Nagata, T., and L. Kruger (1979) Tactile neurons of the superior colliculus of the c a t Input and physiological properties. Brain Res. 174t19-37.
Powell, E.W., and J.B. Hatton (1969) Projections of the inferior colliculus in the cat. J. Comp. Neurol. 136:183-192.
Raczkowski, D., V.A. Casagrande, and I.T. Diamond (1976) Visual neglect in the tree shrew after interruption of the descending projections of the deep superior colliculus. Exp. Neurol. 5Ot14-29.
RoBards, M.J., D.W. Watkins, and R.B. Masterton (1979) An anatomical study of some somesthetic afferents to the intercollicular terminal zone of the midbrain of the opossum. J. Comp. Neurol. 17Ot499-524.
Rubinson, K. (1968) Projections of the tectum opticum of the frog. Brain Behav. Evol. 1x529-561.
Schlag, J., and M. Schlag-Rey (1984) Visuomotor functions of central thala- mus in monkey. 11. Unit activity related to visual events, targeting, and fixation. J. Neurophysiol. 51: 1175-1195.
Schlag-Rey, M., and J. Schlag (1984) Visuomotor functions of central thala- mus in monkey. I. Unit activity related to spontaneous eye movements. J. Neurophysiol. 51:1149-1174.
Schneider, G.E. (1969) Two visual systems. Brain mechanisms for localiza- tion and discrimination are dissociated by tectal and cortical lesions. Science 163t895-902.
Simmons, J.A. (1969) Acoustic radiation patterns for the echolocating bats Chibnycteris rubiginosa and Eptesicus fuscus. J. Acoust. SOC. Am. 46:1054-1056.
Shimozawa, T., N. Suga, P. Hendler, and S. Schuetze (1974) Directional sensitivity of echolocation system in bats producing frequency-modu- lated signals. J. Exp. Biol. 6053-69.
Smeets, W. (1981) Efferent tectal pathway in the chondrichthyan, Schlior- hinus conicula, and the ray, Raja clauata. J. Comp. Neural. 195:13-24.
Sparks, D.L. (1978) Functional properties of neurons in the monkey superior colliculus: Coupling of neuronal activity and saccade onset. Brain Res. 156:l-16.
Sprague, J.M. (1975) Mammalian tectum: Intrinsic organization, afferent inputs and integrative mechanisms. Anatomical substrate. Neurosci. Res. Prog. Bull. 13:204-213.
Stein, B.E., and N.S. Gaither (1981) Sensory representation in reptilian optic tectum: Some comparisons with mammals. J. Comp. Neurol. 202t69-87.
Stein, B.E., R.F. Spencer, and S.B. Edwards (1983) Corticotectal and corti- cothalamic efferent projections of SN somatosensory cortex in cat, J. Neurophysiol. 5Or896-909.
Swanson, L.W., W.M. Cowan, and E.G. Jones (1974) An autoradiographic study of the efferent connections of the ventral lateral geniculate nu- cleus in the albino rat and the cat. J. Comp. Neurol. 156:143-164.
Takagi, J. (1925) Studien zur vergleichenden Anatomie des Nucleus vestib- ularis triangularis. I. Der Nucleus intercalatus und der Nucleus prae- positus hypoglossi. Arb. Neurol. Inst. Univ. Wien. 27t157-188.
Tanaka, K., 0. Katsumi, A. Tokunaga, and S. Sugita (1985) The organiza- tion of neurons in the nucleus of the lateral lemniscus projecting to the superior and inferior colliculi in the rat. Brain Res. 341252-260.
Tarlov, E.C., and R.Y. Moore (1963) The tecto-thalamic connections in the brain of the rabbit. J. Comp. Neurol. 126r403-422.
logical properties of neurons in the ventral lateral geniculate nucleus of Uchida, K., N. ~ i ~ ~ ~ ~ , T, Sugimoto, K, Itoh, and M, Kudo (1983) Direct projections from the cerebellar nuclei to the superior colliculus in the the rabbit. Exp. Neurol. 46t506-520. rabbit An HRP study. J. Comp. Neurol. 216:319-326.
squirrel. Exp. Brain Res. 65:200-212. Walberg, F. (1961) Fastigiofugal fibers to the perihypoglossal nuclei in the cat. Exp. Neurol. 3:525-541.
Mathers, L.H., and G.G. Mascetti (1975) Electrophysiological and morpho-
May, P.J., and W.C. Hall (1986) The cerebellotectal pathway in the grey
Mehler, W.R. (1969) Some neurological species differences: A posteriori.

CONNECTIONS OF THE BAT SUPERIOR COLLICULUS 197
Weber, J.T., and J.K. Harting (1978) Parallel pathways connecting the primate superior colliculus with the posterior vermis. In C.R. Noback (ed): Sensory Systems of Primates. New York Plenum Press, pp. 135- 149.
Wiener, S.I. (1986) Laminar distribution and patchiness of cytochrome oxi- dase in mouse superior colliculus. J. Comp. Neurol. 244t137-148.
Wise, L.Z., and D.R.F. Irvine (1983) Auditory response properties of neurons in deep layers of cat superior colliculus. J. Neurophysiol. 4936'74-685.
Wong, D. (1984) Spatial tuning of auditory neurons in the superior colliculus of the echolocating bat, Myotis lucifugus. Hear. Res. I6:261-270.
Wong-Riley, M. (1979) Changes in the visual system of monocularly sutured or enucleated cats demonstrable with cytochrome oxidase histochemis- try. Brain Res. 17Itll-28.
Wurtz, R.H., and J.E. Albano (1980) Visual motor function of the primate superior colliculus. Annu. Rev. Neurosci. 3t189-226.
Wurtz, R.H., and M.F. Goldberg (1971) Superior colliculus cell responses related to eye movements in awake monkeys. Science 17It82-84.
Zook, J.M., and J.H. Casseday (1982a) Cytoarchitecture of auditory system in lower brainstem of the mustache bat, Pteronotus pamelli i . J. Comp. Neurol. 207:l-13.
Zook, J.M., and J.H. Casseday (1982b) Origin of ascending projections to inferior colliculus in the mustache bat, Pteronotus pamellii. J. Comp. Neurol. 207t14-28.
Zook, J.M., and J.H. Casseday (1985) Projections from the cochlear nuclei in the mustache bat, Pteronotus pamellii. J. Comp. Neurol. 237:307-324.
Zook, J.M., and J.H. Casseday (1987) Convergence of ascending pathways at the inferior colliculus of the mustache bat, Pteronotus pamellii. J. Comp. Neurol. (in press).
Zvorykin, V.P. (1959) Morphological basis of locative and supersonic abili- ties in bat. Arkh. Anat. Gistol. Embriol. 36319-31.




















