
Euroregiunile de cooperare transfrontaliera din Bazinul inferior al Dunarii
Upload
independentCategory
view
0download
0
EXPERIMENTAL NEUROLOGY 37, 469-480 (1972)
Binaural Interaction in the Inferior Colliculus of the Frog
RAMON KAULEN, WLADIMIRO LIFSCHITZ. CARMEN PALAZZJ. AND
HUGO ADRIAN 1
Department of Physiology and Biophysics. Ulriwvsity of Chile, Santiago. Chile
Rcccivcd May 17, 1972
Neural activity in the torus semicircularis (colliculus) of the frog was studied. Tones of different frequency, intensity, and duration were used for stimulation of each ear or both ears simultaneously. Unit responses were classified as contra-
lateral units, ipsilateral units, and bilateral units. This last category was sub- divided into pure bilateral units, units with contralateral inhibition, and units with ipsilateral inhibition. Percentage of occurrence of these units are similar
to these found in the cat and rabbit. This may imply that a general pattern of auditory functional projection is common to vertebrates.
Introduction
Localization of sound sources implies determining the direction followed by the soundpath and distance of the source. Experimental studies have been related to the behavioral and neurophysiological aspects of sound lo- calization mainly in mammals, including man, cat, rabbit, monkey (1. 10. 17). These studies have found that accurate localization depends on the differences in intensity and time of arrival of the sound reaching the two ears. Frequency cues are also important.
Previous studies have shown that physical parameters of the stimulus are among the aspects involved in the problem of lateralization. Other as- pects are related to the functional properties of the auditory system, since neurophysiological studies have shown complex excitatory-inhibitory inter- actions at the different nuclei of the auditory pathway. The results of these physiological events will determine the amount of afferences to the highest levels which produce the significant discrimination related to sound direc- tion,
Neurophysiological studies in cat and rabbit (1) have shown that cell unit activity in the medial genicuiate body, superior olive, trapezoid nu-
1 This research was supported by Grant NB 03610 NIH, United States Public Health Service. The authors gratefully acknowledge the technical assistance of Mr. Sergio Ramirez, Mr. Juan Valencia. Mr. RenP Roi and Miss Micaela Perez.
469 Co yright 0 1972 by Academic Press, Inc. AlP’ht f ng s o reproduction in any form reserved.

470 KAULEN ET AL.
cleus and colliculus depends on the combined input of the two ears. Medial geniculate body units, both in cat and rabbit, show a very similar activity evoked by stimuli delivered to the ears. The percentage of units respond- ing to contralateral and ipsilateral stimulation is similar in both animals. The type of excitatory-inhibitory interactions provoked by stimultaneous or successive stimulation of the two ears produces similar discharge pat- terns as well.
The present work aims to study cell activity in regard to binaural stimu- lation in the inferior colliculus of the frog, to establish if the pattern pre- viously observed in the medial geniculate body of the cat and rabbit is also present here. If this is so, we may conclude that in the evolution of vertebrates there is a general pattern of functional characteristics of unit activity as determined by binaural interaction at different levels.
Methods
Experiments were carried out in 15 adult frogs of both sexes, each weighing about 300 g. Animals were anesthetized with 20% aqueous solu- tion of urethane. 0.8 ml/100 g wt, injected in the dorsal lymphatic sac; body temperature was kept at about 20-24 C. Wet cotton was wrapped around the body and the legs in order to maintain water balance.
Standard operating techniques were used to expose the optic tectum. An indium microelectrode was used for recording. The electrode was driven by a stereotaxic instrument which permitted displacements in laterolateral, anteroposterior and vertical directions. The neural activity from the mi- croelectrode was amplified by an FM 122 Tektronix preamplifier; and dis- played on the upper channel of a 502 Tektronix oscilloscope. The stimulus artifact appeared on the other oscilloscope channel. A C-4 Grass camera photographed the events displayed on the screen of a slave oscilloscope.
Pure tones, generated by a 200 CDR Hewlett Packard oscillator were used as stimuli. The tone was introduced to the input of a 829 C Gras- son Stadler electronic switch, producing gradual rise and decay times in order to avoid electric transients.
Stimulation rate and duration of the stimuli were determined by a pro- gramming unit composed of several 162 and 161 Tektronix stimulators. The sound was filtered to eliminate frequencies below 100 cycle/set. The intensity was controlled with a 350 D Hewlett Packard atenuators. It was possible to stimulate each ear singly or both ears. The close sound delivery system consisted of THD 39 earphones connected to plastic tubes and me- tallic hollow pieces fitted to the frog’s ear with rubber tubing. The inten- sity of the sound was measured in SPL db attenuation, corresponding to 90 db over a sound pressure of 0.0002 dyne/cm’ in a range from 0.2-4 kc.
Photographic records from each particular experiment were mounted on

INFERIOR COLLICULUS 471
cardboard. These were compared to show the influence of the stimulation of the individual ears on the behavior of a particular unit.
Results
Results are based on the neural activity of 117 units in the torus semi- circularis (colliculus) of the frog. Very few of the recorded units showed spontaneous activity ; since the few cases where spontaneous activity oc- curred might have been attributable to the effect of the microelectrode pen- etration, these units were not considered in the results reported in this paper, even though they presented binaural effects.
Two general discharge patterns were observed in the torus semicircu- laris of the frog. Spike activity was observed in some units for the entire duration of the tone stimulus, while others discharged with one to five spikes at the onset of the stimulus, even when the duration of the tone was varied.
Three types of units were found in the frog’s torus semicircularis de- pending on the ear stimulated. Some units responded to the ipsilateral ear; others discharged only to the contralateral stimulation. The last group of units showed different kind of activities depending on the excitatory- inhibitory effect of the two ears.
Figure 1 shows a recording from a unit that responded only to stimula- tion of the ear contralateral to the torus semicircularis. This unit showed only one discharge when stimulated by a tone of high intensity through the contralateral ear.
The ipsilateral ear did not evoke any response even when the stimulus intensity was 20 db higher than the stimulus delivered through the contra- lateral ear. In order to find out if the ipsilateral stimulation had any influ- ence on the unit discharge, both ears were simultaneously stimulated. Even in this case there was no response to ipsilateral stimulation, as is shown by the fact that only one spike was present during the discharge. The unit was driven only by the contralateral ear. From now on this type of unit will be called contralateral unit.
Figure 2 shows another example of this type of unit. In this case the discharge continued for the duration of the tone, with six to eight spikes. There was no response to stimulation of the ipsilateral ear, although the intensity of the tone was 30 db higher than the tone delivered to the con- tralateral ear.
Figure 3 shows the variation of the discharge duration of this same type of unit when tones of different lengths were used. In this case, the dis- charge increased from three or four to ten spikes as the tone duration in- creased from 200 to 100 msec. From 20 msec on, a tonic discharge of lower frequency is observed, after an initial burst of high frequency spikes.

472 KAULEN ET AL.
C.E. 70 db C.E. 10 db I.E. 90 db I.E. 90 db
-+--+
+-+
-+--+
-+--+
-t-+
-e--r---
TONE DURATION 50 msec
loop 0.1 3 Kc L-
50 msec FIG. 1. Example of torus semicircularis units driven by the contralateral ear [Note
that there is only one spike as responses to the tone stimulation (see text)]. Abbrevi- ations : C.E., contralateral ear ; I.E., ipsilateral ear; C.E. and I.E., simultaneous stim- ulation of both ears. Vertical columns correspond to oscilloscope traces to consecutive stimulation. The former description will be the same for the following figures.
Figure 4 presents the activity of a collicular neuron driven hy the ipsilat- era1 ear. In this case stimulation of the contralateral ear showed no effect. Simultaneous stimulation of both ears produced the same effects as stimu- lation of the ipsilateral ear alone. This type will be called ipsilateral unit.
The unit shown in Fig. 5 corresponds to a bilateral unit. Stimulation of either ear or both ears simultaneously yields the same response. This type wilI be called a pure bilateral unit.
In Figure 6 another type of bilateral unit is shown. This type is elicited by contralateral stimulation and inhibited by ipsilateral stimulation. No re- sponse is observed when the ipsilateral ear is stimulated alone although the intensity is 30 db higher than the threshold intensity. When the tone is de- livered to both ears at the same time, no response is obtained ; but if the sound intensity is diminished in the inhibitory ipsilateral ear, the response

INFERIOR COLLICULUS 473
reappears. This type of unit is called a bilateral unit with ipsilateral inhibi- tion. Units in which the contralateral ear gives the inhibitory effect have also been found. These units are called bilateral units with contralateral in- hibition.
Percentages for the occurrence of these units in the frog are presented in Table 1. Contralateral units form the main group, 57.20/o, while the ipsi- lateral units amount only to 2.5%. Table 1 includes the percentages of sub- division for the bilateral units as well.
Each unit was found and studied in relation to its best frequency re- sponse. Data obtained indicates that units respond to frequencies from 100 to 2,300 cycle/set. The distribution between these two frequencies, is shown in Fig. 7. Most of the units responded to a tone of 800 cycle/set with a second peak at 180 cycle/set. Two other minor peaks appear at 550 and 1,200 cycle/set.
C.E. 16 db C.E. 46 db I.E. 76 db
I.E.46 db
TONE DURATION 50 msec
1 Kc 200 p
50 msec FIG. 2. Shows another of contralateral unit. Note that there is a discharge that lasts
the length of the tone (see text).

474 KAULEN ET AL.
20 msec 25 msec 32 msec 50 msec 100 msec
STIMULUS INTENSITY 16 db 1 Kc
200 )lv L 100 msec
FIG. 3. Shows a unit that discharged during the whole length of tone. Tone was delivered to the contralateral ear with duration that are shown at the top of the col- umn. Note that this unit follows the different durations of the tone (see text).
TABLE 1
CLASSIFICATION OF UNITS ISOLATED IN THE TORUS SEMICIRCULARIS OF THE FROG
Contralateral Ipsilateral Bilateral units units units
Neurons recorded 67 3 47
% 57.2 2.5 40.1
Neurons recorded
y. of all neurons a
Bilateral units
Driven by Ipsilateral Contralateral both ears inhibition inhibition
-
24 21 2
20.5 17.9 1.7
a Total number of units recorded. 117.

INFERIOR COLLICULUS 475
I.E. 60 db C.E. 80 db I.E. 60 db
C.E. 80 db
T-l-----
TONE DURATION 50 mscc
0.55 Kc loop
63 msec FIG. 4. Example of unit driven by the ipsilateral ear (see text).
Discussion
It is well-known that the electrical stimulation of the temporal lobe in human patients determines sound localization referred mainly to the contralateral side (10). Patients with unilateral temporal lobe damage have a greater deficit in discrimination of distorted speech in the con- tralateral ear than in the ipsilateral (2, 5. 8. 9, 16). Animals unilat- erally deprived of the auditory cortex plus s1 and sZ show impairment in localizing direction of the sound. This deficit is more evident in the side opposite to the lesion (16). From comparison of deficits shown in patients and animals it is possible to infer that a similar mechanism are involved. Different explanations have been given in an effort to explain this effect of

476 KAULEN ET AL.
better localization or discrimination in the ear ipsilateral to the lesion. They point in general to a greater reduction in the auditory contralateral field than in the ipsilateral (16).
Electrophysiological evidences point to a greater projection from either ear to the contralateral cortex than to the ipsilateral cortex (3, 4, 7, 13-15). Deficits in human patients, experimental deficits in animals, and electro-physiological studies suggest the existence of a general pattern in the projection from one ear to the auditory cortex.
Work done in this laboratory on the medial geniculate body of the cat and rabbit (1) show that neurons can be classified according to the ear that influences them in contralateral units, ipsilateral units and bilateral units (Table 2). The percentages displayed in Table 2 show that there is a greater number of neurons of the medial geniculate body driven by the contralateral ear than by the ipsilateral ear. Since the medial geniculate
C.E. L6 db I.E. 10 db C.E. 46 db
I.E. LO db
-Y--V---
---
---
---
TdNE DURATION 50 msec
0.9 Kc 200 pv T 50
FIG. 5. Shows a unit discharge driven by both ears (see text).

INFERIOR COLLICULUS 477
C.E. 50 I.E. 80 db C.E. 60 db C.E. 60 db
db I.E. 60 db I.E. 50 db
L
I @---
L
I ---,
* l
_-- I--
. A--II’ c
I
----
TONE DURATION T 50 msec 2oo)rv
FIG. 6. Shows a unit that is driven by the contralateral ear and it is inhibited by the ipsilateral ear. I f the intensity is diminished in the inhibitory ear the unit discharge reappears (see text).
body is the last relay station to the cortex, it seems reasonable to think that the same type of projection would exist on the level of the auditory cortex. These results could help to explain greater deficits in the ear con- tralateral to the lesion in human and animal subjects.
In consequence, an experiment was designed in lower vertebrates, to find out if the type of projection from the ear to the higher levels was the same that obtained in cat and rabbit. Similarity of percentages (Tables 1 and 2) suggest that there is a general basic pattern of functional projection in the auditory system of vertebrates.
As it is well-known, unit responses of neurons of auditory nuclei located below the inferior colliculus of the cat and rabbit (1) follow the entire du- ration of the tone. But at the level of the inferior colliculus two patterns of discharges can be classified: one common to the lower levels, i.e., that fol- lows the direction of the tone, and another that consists of a short burst of
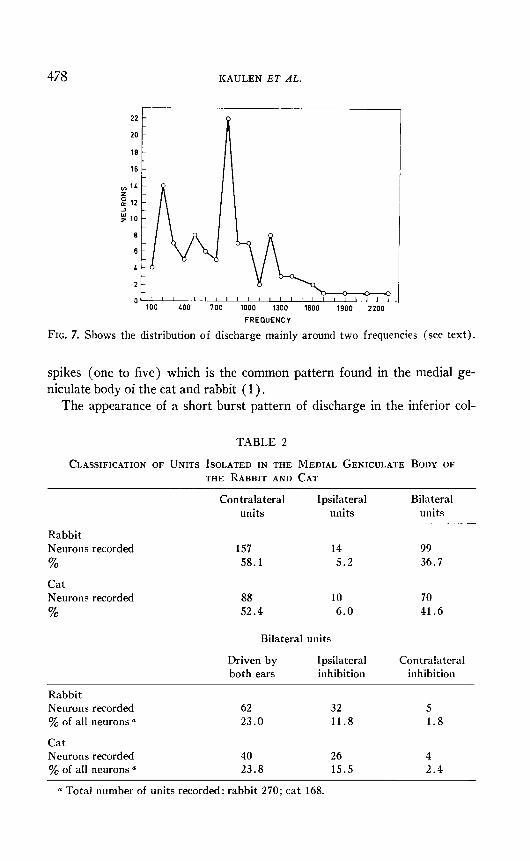
478 KAULEN ET AL.
FREWENCV
FIG. 7. Shows the distribution of discharge mainly around two frequencies (see text).
spikes (one to five) which is the common pattern found in the medial ge- niculate body of the cat and rabbit ( 1) .
The appearance of a short burst pattern of discharge in the inferior col-
TABLE 2
CLASSIFICATION OF UNITS ISOLATED IN THE MEDIAL GENICULATE BODY OF
THE RABBIT AND CAT
Contralateral units
Ipsilateral units
Bilateral units
Rabbit Neurons recorded 157 14 99 % 58.1 5.2 36.7
Cat Neurons recorded 88 10 70 % 52.4 6.0 41.6
Bilateral units
Driven by Ipsilateral both ears inhibition
Contralateral inhibition
Rabbit Neurons recorded 62 32 5 $& of all neurons 0 23.0 11.8 1.8
Cat Neurons recorded 40 26 4 yO of all neurons a 23.8 15.5 2.4
a Total number of units recorded: rabbit 270; cat 168.

INFERIOR COLLICULUS 479
liculus seems to indicate that the discharges coming from lower levels of the auditory pathways suffer a process of integration, ‘since an onset pat- tern is found. Results obtained in this experimental work show the two types of patterns are present in the auditory neurons of the torus semicir- cular& of frog. The percentage burst pattern is about 60% of the recorded units. The remaining percentages are in accordance with those found in the cat and rabbit. It is of interest to consider the presence of short burst and long duration discharges in different vertebrates and possibly to all of them.
It has been described that the spectrum of audible frequencies to the bullfrog ranges between 100 cycle/set to 3000-4000 cycle/set (6). Refer- ence results obtained in the Chilean frog show that its audible frequencies varies from 100 cycle/set to an upper limit of around 2300 cycle/set. The comparison of the cited results, suggests that the audible frequency range for the frog depends on the particular species.
The distribution of auditory neurons of the torus semicircularis accord- ing to their best frequencies is shown in Fig. 7. Four peaks are shown but only two are prominent. This last distribution corresponds to that de- scribed by other authors (6, 11, 12). The meaning of these four peaks could be related to the four frog calls : mating, warning, distress, or release (6).
Another aspect that seems to have some interest is the influence of the duration of the tone on the number of discharges that the neurons of the torus semicircularis. A large spike count is obtained for a given stimulus duration. This counts for shorter or longer stimulus durations. Even for different attenuations, if these findings are true, they point to the possibil- ity of a best duration for a given neuron located in the midbrain. These re- sults agree with the findings of Potter ( 11, 12).
References
1. ADRIAX, H. 0.. W. M. LIFSCHITZ, R. J. TAVITAS, and F. P. GALLI. 1966. Activ- ity of neural units in medial geniculate body of cat and rabbit. J. Nezlroplz~lsiol. 29 : 1046-1060.
2. BOCCA. E. 1955. Binaural Hearing: Another approach. Laryngoscopc 65 : 1164- 1171.
3. BREJER. F., and R. S. Dow. 1939. The acoustic area of cerebral cortex in the cat. .4 combined oscillographic and cytoarchitectonic study. J. Neuropkysiol. 2 : 308-31s.
4. BKEXEIL F. 1952. Analyze oscillographiques des responses sensorielles des Ccorses cerebral et cerebelleuse. Reef. Scurol. 67 : 65-92.
5. CALEARO, C. 1957. Binaural summation in lesions of the temporal lobe. Acfa Ofo- rltinolaryrrgol. 45 : 392-395.
6. CAl’RAiXIc.4, R. R. Characteristics of the calls in the Bullfrog’s vocal repertoire.

480 KAULEN ET AL.
1967. The 74th meeting of the acoustical society of America, Miami Beach (13-17 November).
7. GALAMBOS, R. J., E. ROSE, R. B. BROMILEY, and J. R. HUGHES. 1952. Micro- electrode studies on the Medial Geniculate body of cat. II Responses to clicks. J. Neurophysiol. 15 : 359-380.
8. GOLDSTEIN, R. A. GOODMAN, and R. B. KING. 1956. Hearing and speech in in- fantile hemiplegia before and after left hemipherectomy. Ncnrologg 6 : 869-875.
9. JERGER, J. 1960. Observations on auditory behavior in lesions of the central audi- tory pathways. Arch. Otolaryngol. 71: 797-W.
10. PENFIELD, W., and T. RASMUSSEN. 1950. “The Cerebral of Man: A Clinical Study of Localization of Functions.” Macmillan, New York.
11. POTTER, H. D. 1965. Mesencephalic auditory region of the bullfrog. J. Newo- fdzysiol. 28 : 1132-1154.
12. POTTER, H. D. 1965. Patterns of acoustically evoked discharges of neurons in the mesencephalon of the bullfrog. J. Nrz~rophgsiol. 28 : 1155-1184.
13. ROSENZWEIG, M. R. 1951. Representation of the two ears at the auditory cortex.
J. Cowp. Physiol. 67 : 147-158. 14. ROSENZWEIG, M. R. 1954. Cortical correlations of auditory localization and of
related perceptual phenomena. J. Camp. Physiol. Psychol. 47: 269-276. 15. ROSENZWEIG, M. R. 1961. Development of research on physiological mechanisms
of auditory localization. Psychol. Bull. 58 : 376-889. 16. SANCHEZ-L• NGO, L. P., and F. M. FORSTER. 1958. Clinical significances of im-
pairments of sound localization. Nc~rvolog~~ 8 : 119-125. 17. STROMINGER, N. L. 1969. Localization of sound in space after unilateral and bi-
lateral ablation of auditory cortex. E.rp. Nczrrol. 25 : 521-533. 18. TUNTURI, A. R. 1960. Anatomy and physiology of the auditory cortex, pp. 181-
200. In “Neural Mechanisms of the Auditory and Vestibular System.” G. L. Rasmunssen and W. F. Windle [Eds.]. Thomas, Springfield, Illinois.
Copyright © 2022 FDOKUMEN




















