
Frequency discrimination and stimulus deviance in the inferior colliculus and cochlear nucleus
Upload
independentCategory
view
0download
0
P1: GXB
Cellular and Molecular Neurobiology [cemn] pp793-cemn-461951 March 13, 2003 12:35 Style file version Oct 23, 2000
Cellular and Molecular Neurobiology, Vol. 23, No. 2, April 2003 ( C© 2003)
Early Postnatal Sound Exposure Induces LastingNeuronal Changes in the Inferior Colliculus of SenescenceAccelerated Mice (SAMP8): A Morphometric Studyon GABAergic Neurons and NMDA Expression
Dietrich Ernst Lorke,1 Lai Yung Wong,2 Helen W. L. Lai,2 Paul W. F. Poon,3
Aiqun Zhang,4 Wood Yee Chan,2 and David Tai Wai Yew2,5
Received December 15, 2002; accepted December 18, 2002
SUMMARY
Senescence-acceleration-prone mice (SAMP8) provide a model to study the influence ofearly postnatal sound exposure upon the aging auditory midbrain. SAMP8 were exposed toa 9-kHz monotone of either 53- or 65-dB sound pressure level during the first 30 postnataldays, the neurons in the auditory midbrain responding selectively to 9 kHz were localizedby c-fos immunohistochemistry and the following parameters were compared to controlSAMP8 not exposed to sound: mortality after sound exposure, dendritic spine density, andquantitative neurochemical alterations in this 9-kHz isofrequency lamina. For morphomet-ric analysis, animals were examined at 1, 4, and 8 months of age. Serial sections of the inferiorcolliculus were Golgi impregnated or stained immunohistochemically for the expression ofε1 subunit of NMDA receptor or GABA. Mortality after exposure to 53 dB was the sameas in controls, but was markedly increased from 7 months of age onward after postnatalexposure to 65 dB. No gross morphological alterations were observed in the auditory mid-brain after sound exposure. However, sound exposure to 53 or 65 dB significantly reduceddendritic spine density by 11% at 4 months or by 11–17% both at 1 and 4 months of age,respectively. The effect of sound exposure upon neurons expressing the NMDAε1 subunitwas dose-dependent. Increasing with age until 4 months in control mice and remainingessentially stable thereafter, the percentage of NMDAε1-immunoreactive neurons was sig-nificantly elevated by 40–66% in 1- and 8-month-old SAMP8 exposed to 53 dB, whereas nosignificant effect of 65 dB was apparent. The proportion of GABAergic cells declined withage in controls. It was significantly decreased at 1 month after 53 and 65 dB sound exposure.In contrast, it was elevated at later stages, being significantly increased at 4 months afterexposure to 53 dB and at 8 months after exposure to 65 dB. The total cell number in the9-kHz isofrequency lamina of SAMP8 decreased with age, but was not affected by expo-sure to either 53 or 65 dB. The present results indicate that early postnatal exposure to a
1 Institute of Neuroanatomy, University Hospital Hamburg–Eppendorf, Hamburg, Germany.2 Department of Anatomy, The Chinese University of Hong Kong, Shatin, New Territories, Hong Kong
SAR, China.3 Department of Physiology, College of Medicine, Institute of Basic Medical Sciences, National Cheng
Kung University, Tainan, Taiwan.4 Institute of Geriatrics and Gerontology, Chinese PLA General Hospital, Beijing, People’s Republic of
China.5 To whom correspondence should be addressed at Department of Anatomy, Chinese University of Hong
Kong, Shatin, New Territories, Hong Kong; e-mail: [email protected].
143
0272-4340/03/0400-0143/0 C© 2003 Plenum Publishing Corporation

P1: GXB
Cellular and Molecular Neurobiology [cemn] pp793-cemn-461951 March 13, 2003 12:35 Style file version Oct 23, 2000
144 Lorke, Wong, Lai, Poon, Zhang, Chan, and Yew
monotone of mild intensity has long-term effects upon the aging auditory brain stem. Someof the changes induced by sound exposure, e.g., decline in spine density, are interpreted asaccelerations of the normal aging process, whereas other effects, e.g., increased NMDAε1expression after 53 dB and elevated GABA expression after both 53 and 65 dB, are notmerely explicable by accelerated aging.
KEY WORDS: aging; auditory midbrain; c-fos; dendritic spine; GABA; inferior colliculus;mouse; NMDA receptor; presbyacusis; SAMP8; senescence accelerated; sound exposure;spine density.
INTRODUCTION
Age-related hearing loss (presbyacusis) is a debilitating disorder affecting about50% of the United States’ population aged 75 years and older (Caspary et al., 1999;Seidman et al., 2002). Presbyacusis manifests itself by difficulties in speech compre-hension, particularly in noisy environment. Combined with the inability to processcomplex auditory signals, this slow deterioration of auditory function is a majorreason for the tendency of affected people to withdraw from active participation insociety (Caspary et al., 1999) and seriously impairs the social, emotional, and medicalwell-being of elderly individuals.
Age-related changes in the auditory system occur at various levels of both of theperipheral and the central nervous systems (CNSs) (for a review, see Seidman et al.,2002; Willot et al., 2001). One of the major and best-characterized auditory structuresin the CNS is the inferior colliculus, which is involved in complex acoustic signal pro-cessing (Caspary et al., 1999; Jiang et al., 1997), receiving input from almost all otherauditory nuclei (Caird, 1991; McAlphine et al., 1998; Oliver and Shneiderman, 1991;Palombi and Caspary, 1996). Age-related changes in the inferior colliculus seriouslyaffect the GABAergic system (Caspary et al., 1995, 1999; Helfert et al., 1999). Lossof the GABA-synthesizing enzyme GAD (glutamic acid decarboxylase), reductionin the number of GABAergic cells and in GABA content, decreased GABA releaseand loss of GABA(B) receptors in the aging rodent inferior colliculus have beenreported (Caspary et al., 1995, 1999). Moreover, a significant reduction in the den-sities of both GABA immunoreactive and GABA immunonegative synapses linkedto a corresponding loss of dendrites is observed, which has been related to dendriticdecline (Helfert et al., 1999). Dendritic spines are subject to age-related changes,too, and spine morphology is correlated with cognitive functions in the aging brain(Scheibel et al., 1975, 1976). The other major neurotransmitter in the inferior collicu-lus is glutamate. Excitatory responses in the rodent inferior colliculus are mediatedby glutamate via NMDA and AMPA receptors (Kelly and Zhang, 2002; Ma et al.,2002; Zhang and Kelly, 2001). Age-related changes in NMDA expression, i.e., re-duced expression of subunits ζ1 (NR1), ε1 (NR2A), and ε2 (NR2B), associated withfunctional deficits, have been reported in various areas of the brain (Clayton et al.,2002; Clayton and Browning, 2001; Magnusson et al., 2002).
Little is known about the causes underlying age-related degenerative changesin the auditory midbrain (see Willott et al., 2001, for a review). The inferior col-liculus is one of the most metabolically active areas of the brain (Gross et al., 1987;Zeller et al., 1997) and is therefore subject to a greater amount of oxidative stress

P1: GXB
Cellular and Molecular Neurobiology [cemn] pp793-cemn-461951 March 13, 2003 12:35 Style file version Oct 23, 2000
Auditory Midbrain in Senescence Accelerated Mice After Sound Exposure 145
during aging (Helfert et al., 2002). This increased sensitivity to stress agents and adecreased repairing capacity following stress may lead to an increased sensitivityto noise-induced damage of the auditory system (Miller et al., 1998). In mice, earlypresbyacusis is associated with an increased sensitivity to noise (Miller et al., 1998).On the other hand, animal studies indicate that the allocation of hearing loss in anelderly individual into a noise component and an aging component is more complexthan additivity of decibels (Mills et al., 1998). Although exposure to intense noiseis likely to have negative effects upon hearing by the time old age is reached, theinteraction of noise- and age-related hearing loss is difficult to determine (Willottet al., 2001).
Elucidation of the relationship between age-related and noise-induced hearinglosses is therefore important for understanding the pathogenetic mechanisms under-lying them and for minimizing or preventing hearing loss (McFadden et al., 1998). Todistinguish noise-induced changes from age-related alterations occurring indepen-dently from noise, a model system is required allowing the systematic investigationof a large number of individuals reaching senescence in a short time. The senes-cence accelerated mouse (SAM) mutant developed by Takeda et al. (1981, 1991)provides such a model. Eleven substrains have been described, eight SAM-prone(SAMP) and three SAM-resistant lines. Mice of each SAMP line exhibit age-relatedpathological characteristics, which include degenerative arthropathy, osteoporosis,cataract, thymoma, and a variety of neuropathological abnormalities, which are alsocommon features in human aging (for a review, see Kawamata et al., 1997; Okumaand Nomura, 1998). SAMP strain 8 is particularly susceptible to neuropathologicalchanges (for a review, see Morley, 2002), e.g., overproduction of amyloid precursorprotein andβ-amyloid (Morley et al., 2002; Takemura et al., 1993), defects of brain mi-crovessels (Ueno et al., 2001), dysfunction of the cholinergic system (Onozuka et al.,2002), and other neurotransmitter changes (Flood et al., 1998; Kondziella et al., 2002;Nomura et al., 1997), leading to deficits in learning and memory (Nomura and Okuma,1999; Spangler et al., 2002) with relatively low incidence of other aging phenotypes(Morley et al., 2002).
Recent electrophysiological studies of the inferior colliculus have shown thatold SAMP8 individuals exhibit reduced responses to sound and a loss in sensitivityof high frequencies and that this general loss in auditory sensitivity is aggravatedby sound exposure during the early postnatal development (Chiu et al., 2002). Todetermine the underlying morphological and neurochemical basis of this enhancednoise-induced deterioration of inferior colliculus function in accelerated senescence,we have undertaken a morphometric examination of the SAMP8 inferior colliculus.During the early postnatal development, we have subjected SAMP8 to a moderatetone of 65 dB sound pressure level (SPL), which has been shown to lead to clusteringin frequency tuning of normal rat inferior colliculus neurons (Poon and Chen, 1992),and to a lower sound intensity of 53 dB SPL, which induces plastic changes in theSAMP8 inferior colliculus (Chiu et al., 2002). Because NMDA receptors are essentialfor mediating acoustic responses in the inferior colliculus and because loss of den-dritic spines and decrease in GABAergic neurons are regular features of age-relatedhearing loss, we have visualized NMDA receptor expressing and GABA synthesiz-ing neurons by immunohistochemistry and examined dendritic spines by the Golgi

P1: GXB
Cellular and Molecular Neurobiology [cemn] pp793-cemn-461951 March 13, 2003 12:35 Style file version Oct 23, 2000
146 Lorke, Wong, Lai, Poon, Zhang, Chan, and Yew
technique. The aim of this study is to measure densities of dendritic spines, NMDA re-ceptor expressing neurons, and GABA synthesizing neurons in the inferior colliculusof SAMP8 after exposure to sound of different intensities.
METHODS
Animals
Senescence accelerated mice (SAM, strain P8), both male and female, were keptunder defined environmental conditions (21◦C; 50% relative humidity; 12-h light–dark cycle) in the Laboratory Animal Services Centre of the Chinese Universityof Hong Kong. Standard diet (PicoLab Rodent Diet 20, PMI Nutrition Int., Inc.)and water were given ad libitum. The SAM colony had initially been establishedin 1999 from a seed colony of 20 SAMP8 mice originating from the Institute ofFrontier Medical Sciences, Kyoto University. In the course of the study, SAMP8controls were randomly selected and sent to the Evolutionary Biology Unit, SouthAustralian Museum, Adelaide, Australia, for genetic tests. Genetic authenticity wasconfirmed by differences at three loci characterizing the strain (Idh-1, Mod-1, andCar-2) using standard electrophoretic screening procedures (Higuchi, 1997). Duringthe entire experiment, the “Guiding Principles in the Care of and Use of LaboratoryAnimals” were observed.
Prolonged Acoustic Stimulation
Litters of newborn mice were reared by their mothers and kept in a sound-shielded chamber equipped with a quiet ventilation system built specifically for thispurpose. One hundred and eight SAMP8 mice were taken for histological and mor-phometric examination. They were divided into three groups (n = 36, each): Thefirst group (mild sound intensity) was exposed to a 9 kHz (optimal auditory sensi-tivity range in rodents (Clerici and Coleman, 1987)) continuous tone at an intensityof 53 dB SPL for a period of 10 h/day (8:00 P.M. to 6:00 A.M.). The sound was gen-erated from an integrated audio amplifier (Nynaco, Japan, model # SCA-200), afunction generator (American Reliance Inc., U.S.A., model # FG-513), and an audiomixer; it was broadcasted from a free field speaker (Pioneer SP77) located in thecenter of the chamber’s ceiling. Sound intensity was measured close to the animalsusing a calibration microphone (B&K 4191). The second group (higher sound in-tensity) was exposed in the same manner to a 9 kHz monotone of 65 dB SPL. Alltreatment began on postnatal day 1 and continued for 30 days. The control groupwas raised under the same conditions as the experimental animals were, but wasnot exposed to any sound. In addition, survival characteristics after sound expo-sure were determined. Animals underwent the same treatment (53, 65 dB soundexposure and controls not exposed to sound), but were not examined histolog-ically. Unpaired Student’s t-test was utilized to determine statistically significantdifferences with a p value set at 0.05. At the end of the treatment, all animalswere returned to the regular housing facilities. To ascertain the localization of the9 kHz isofrequency lamina in the SAMP8 inferior colliculus, six SAMP8 aged 2 and

P1: GXB
Cellular and Molecular Neurobiology [cemn] pp793-cemn-461951 March 13, 2003 12:35 Style file version Oct 23, 2000
Auditory Midbrain in Senescence Accelerated Mice After Sound Exposure 147
8 months were exposed to sound and processed for c-fos immunohistochemistry (seebelow).
Histology
Animals were sacrificed at 1, 4, and 8 months of age. They were first anesthetizedwith an i.p. injection of 100 mg/kg ketamine and 10 mg/kg xylazine, then the thoraciccavity was opened and, after incision of the jugular veins and the right atrium, an-imals were perfused transcardially with isotonic saline solution followed by a mix-ture of 4% paraformaldehyde and 0.35% glutaraldehyde in 0.1 M phosphate buffer(pH 7.4; 18 min). Thereafter, the skull was partially opened and the brain was care-fully removed. The whole brain was postfixed at 4◦C in the same fixative for another16 h. The inferior colliculus was then dissected and further processed for histologyat room temperature unless stated otherwise.
NMDAε1 Receptor and GABA Immunohistochemistry
Tissue specimens (n = 6 per age group) were rinsed in 0.1 M phosphate buffer(pH 7.4, 12 h, 4◦C), dehydrated in ascending series of ethanol and embedded inHistosec (Merck, Darmstadt, Germany) using xylene as an intermedium. Six-micrometer coronal serial sections were mounted, rehydrated in descending series ofethanol, and then predigested with 0.1% trypsin (BDH Laboratory Supplies, Poole,U.K.) in 0.05 M Tris buffered saline containing 0.1% CaCl2 (pH 7.6, 37◦C, 20 min)followed by two rinses in 0.01-M phosphate buffered saline (PBS; pH 7.4, 5 min).The endogenous peroxidase activity was blocked with 0.3% hydrogen peroxide inabsolute methanol for 30 min, followed by another three rinses in PBS (5 min each).To suppress nonspecific binding, sections were then incubated for 1 h in 2% nor-mal blocking serum (Vectastain ABC Kit, Vector Laboratories, Burlingame, CA)in 0.3% triton/PBS. Thereafter, sections were incubated overnight with the primaryantibody: polyclonal rabbit anti-GABA (1:1000, Chemicon, Ternecula, CA) or poly-clonal goat anti-NMDAε1(C-17) (1:400, Santa Cruz Biotechnology, Inc., Santa. Cruz,CA) for 24 h at 4◦C. The sections were then washed with three rinses of PBS con-taining 0.05% Tween 20 (5 min each) and incubated with the respective biotiny-lated secondary antibody in blocking solution for 30 min (1:200, Vectastain ABCKit, Vector Laboratories, Burlingame, CA). Sections were washed three times inPBS again (5 min) and incubated with ABC Reagent (Vectastain ABC Kit, VectorLaboratories, Burlingame, CA) for 30 min. The immunocytochemical staining sig-nals were visualized by incubating the sections in 0.05% of the substrate 3′3′ -diaminobenzidine tetrahydrochloride (DAB) in PBS containing 0.01% H2O2. In-cubation times for color development were 4 min for GABA immunoreaction and 2min for NMDAε1 immunoreaction. All staining procedures included negative con-trols with omission of the primary antibody, which did not show any immunoreaction.The sections were washed with distilled water and then counterstained with 0.5%cresyl fast violet in 0.1 M sodium acetate (pH 3.5, 5 min). They were differentiatedwith 95% ethanol, dehydrated, cleared, and covered in Permount (Fisher Scientific,Hampton, VA).

P1: GXB
Cellular and Molecular Neurobiology [cemn] pp793-cemn-461951 March 13, 2003 12:35 Style file version Oct 23, 2000
148 Lorke, Wong, Lai, Poon, Zhang, Chan, and Yew
Rapid Golgi Method
Tissue blocks of the inferior colliculus of six mice per experimental group wereexamined (total number: n = 54). After postfixation, excess fixative was blotted offfrom tissue specimens with a piece of tissue paper. Tissue blocks were then incubatedfor 7 days in 0.2% osmium tetroxide/3% potassium dichromate at 37.9◦C. Thereafter,salt deposits were removed by brushing the tissue blocks with a fine brush. Tis-sue was washed in distilled water and embedded in Histosec (Merck, Darmstadt,Germany) as described above. Ten-micrometer sections were cut, mounted on 3%gelatine-coated slides, and cover-slipped using Permount . The density of dendriticspines from 200 cells per animal located within the 9-kHz isofrequency lamina wasdetermined (see below).
c-fos Immunohistochemistry
To ascertain the exact location of the 9 kHz isofrequency laminae in the SAMP8inferior colliculus, two additional groups of SAMP8 (age: 2 months, n = 3; 8 months,n = 3) were examined. The mice were placed in the silent soundproof chamber for6 h. They were then exposed to a 9 kHz monotone of 65 dB SPL for 30 min. There-after, the sound was switched off and the animals were left in the silent soundproofchamber for another 30 min. They were then anesthetized with ketamine/xylazineand perfused with isotonic saline followed by 4% buffered paraformaldehyde as de-scribed above. The brain was removed from the skull and postfixed overnight in thesame fixative. The inferior colliculus was dissected, rinsed in 0.1 M phosphate buffer,and placed in cold 20% sucrose in 0.1-M phosphate buffer for 2 days followed by em-bedding in Tissue-Tek O.C.T. compound (Sakura Finetek, Torrance, CA). A totalof 40-µm-thick cryostat serial sections were prepared, mounted onto SuperFrost
Plus slides (Menzel, Burgdorf, Germany), and stored at −20◦C until further use.Sections were rehydrated in PBS and incubated with rabbit anti-c-fos antibody (SC-253, Santa Cruz Biotechnology, Inc., Santa Cruz, CA) diluted in PBS containing 2%normal goat serum and 10% Triton X-100 (48 h, 4◦C). They were then rinsed threetimes in PBS for 15 min and subjected to the Vectastain ABC detection method(Vector Laboratories, Burlingame, CA) as described above. DAB was used as chro-mogen (15 min). Sections were rinsed with running tap water, dehydrated throughgraded ethanol, cleared with xylene, and covered in Permount (Fisher Scientific,Hampton, VA).
Image Analysis
All images were aquired with an Axiophot 2 photomicroscope (Zeiss, Jena,Germany) equipped with an Optronics color video camera (Thales Optem, Fairport,NY) at a magnification of 400×. Six sections of each inferior colliculus sampled atregular intervals of 80µm were taken for analysis. Three-dimensional reconstructionof the 9 kHz isofrequency laminae was performed on the sections immunostainedfor c-fos using a Neurolucida (MicroBright Field, Inc., Colchester, VT) computerizedcamera lucida system. Total number of neurons was assessed using the same systemby counting all nuclei within a counting frame representing the 9 kHz isofrequency

P1: GXB
Cellular and Molecular Neurobiology [cemn] pp793-cemn-461951 March 13, 2003 12:35 Style file version Oct 23, 2000
Auditory Midbrain in Senescence Accelerated Mice After Sound Exposure 149
lamina. The total cell number was determined by correcting the crude number countaccording to Abercrombie’s formula for double counting (Abercrombie, 1946). Per-centages of GABA positive cells and NMDAε1 positive cells as well as their respec-tive cell sizes were measured within the 9 kHz isofrequency lamina and within thelamina immediately adjacent to it ventromedially (to assess specificity of 9 kHz re-action) using the Neurolucida imaging software, too. The density of dendritic spineswas determined on Golgi-stained sections using the Stereo Investigator imagingsoftware (MicroBright Field, Inc., Colchester, VT), which allows quantitative three-dimensional analysis of complete dendritic trees (Bock and Braun, 1999). Imageswere taken from the 9 kHz isofrequency lamina only, at a magnification of 400×.All counting was done using the same counting frames. All impregnated dendrites of200 cells per animal were analyzed. The number of dendritic spines was determinedafter measuring the entire length of the dendritic tree by tracing the whole dendritewhile counting all dendritic spines from the dendrite. Spine density was calculatedas number per 1 µm dendritic length. The unpaired Student’s t-test was used forstatistical analysis with p value set at 0.05.
RESULTS
Mortality After Sound Exposure
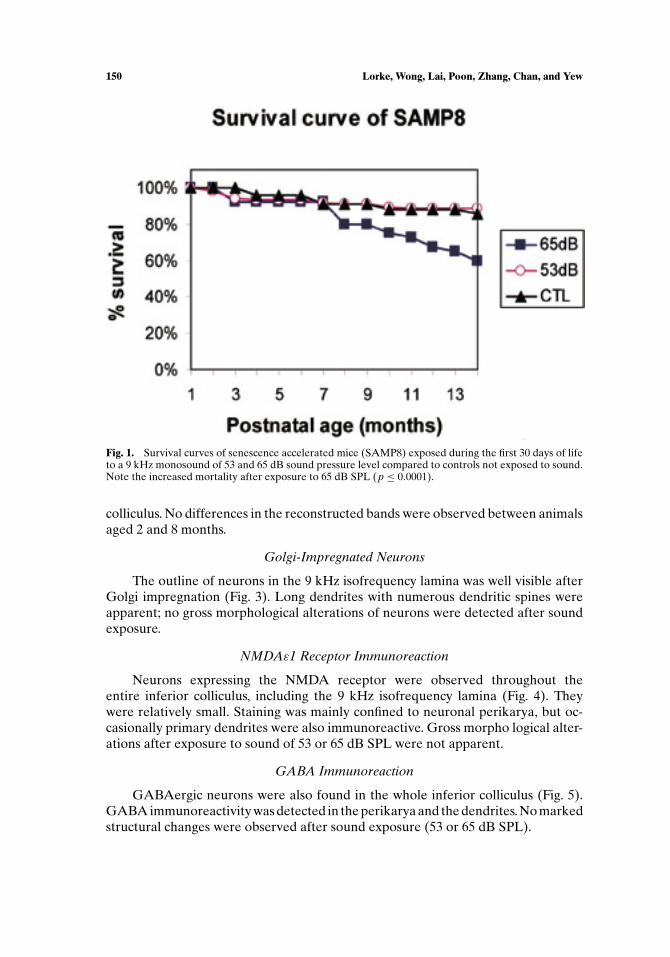
SAMP8 controls not exposed to sound started to die at the age of about 3 months.Thereafter, mortality continued steadily at an approximate rate of 1.0% per monthreaching a cumulative mortality of 14% at the end of 14 months (Fig. 1). Deathof the animals, though occasionally occurring without prodromal signs, was oftenpreceded by a rapid loss in body weight, yellow coloring of the fur, and a markedreduction in motor activity. Pneumonia, tumors of the liver, splenomegaly, and eyelesions were regular findings at autopsy. Mortality was essentially the same afterearly postnatal exposure to low- (53 dB SPL) sound levels, the monthly mortalityrate being 0.795% and the cumulative mortality reaching 11% after 14 months. Incontrast, exposure to the higher-sound levels (65 dB SPL) was accompanied by asignificant increase in mortality (p ≤ 0.0001). Although survival until 6 months wasnot altered after exposure to 65 dB SPL, mortality was markedly higher thereafter,reaching a monthly mortality rate of 2.68% and a cumulative mortality of 40% atthe end of 14 months.
Morphology
c-fos Immunoreaction
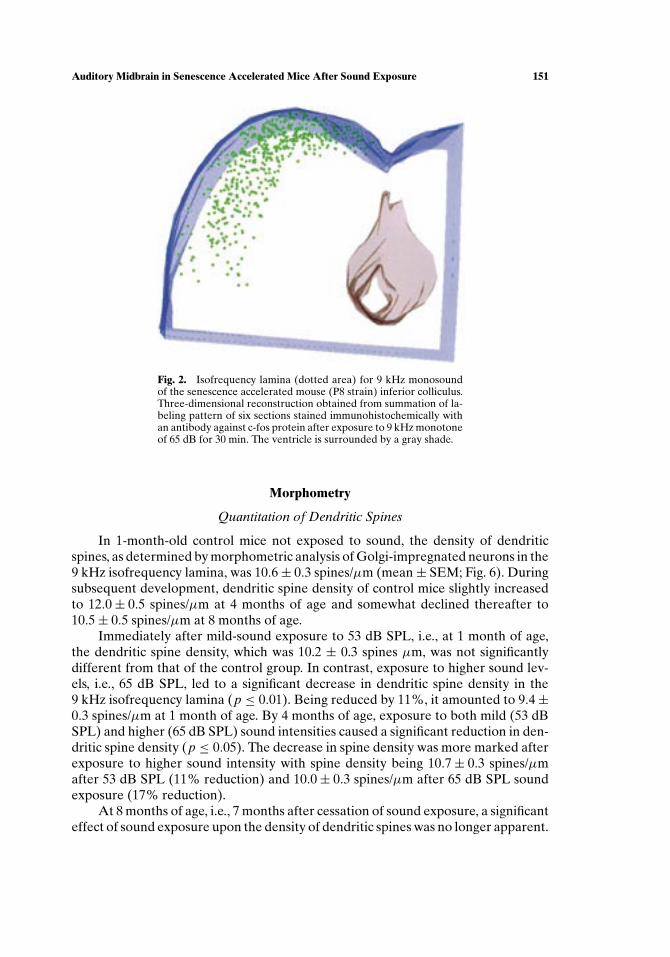
Neurons in the SAMP8 inferior colliculus responding to acoustic stimulation bya 9 kHz monotone were visualized by c-fos immunohistochemistry (Fig. 2). Stainingrevealed a c-shaped band of labeled cells, which was situated close to the meningealsurface, extended ventrolaterally from the extreme dorsomedial edge of the inferiorcolliculus, and observed throughout the entire rostrocaudal extent of the inferior
P1: GXB
Cellular and Molecular Neurobiology [cemn] pp793-cemn-461951 March 13, 2003 12:35 Style file version Oct 23, 2000
150 Lorke, Wong, Lai, Poon, Zhang, Chan, and Yew
Fig. 1. Survival curves of senescence accelerated mice (SAMP8) exposed during the first 30 days of lifeto a 9 kHz monosound of 53 and 65 dB sound pressure level compared to controls not exposed to sound.Note the increased mortality after exposure to 65 dB SPL (p ≤ 0.0001).
colliculus. No differences in the reconstructed bands were observed between animalsaged 2 and 8 months.
Golgi-Impregnated Neurons
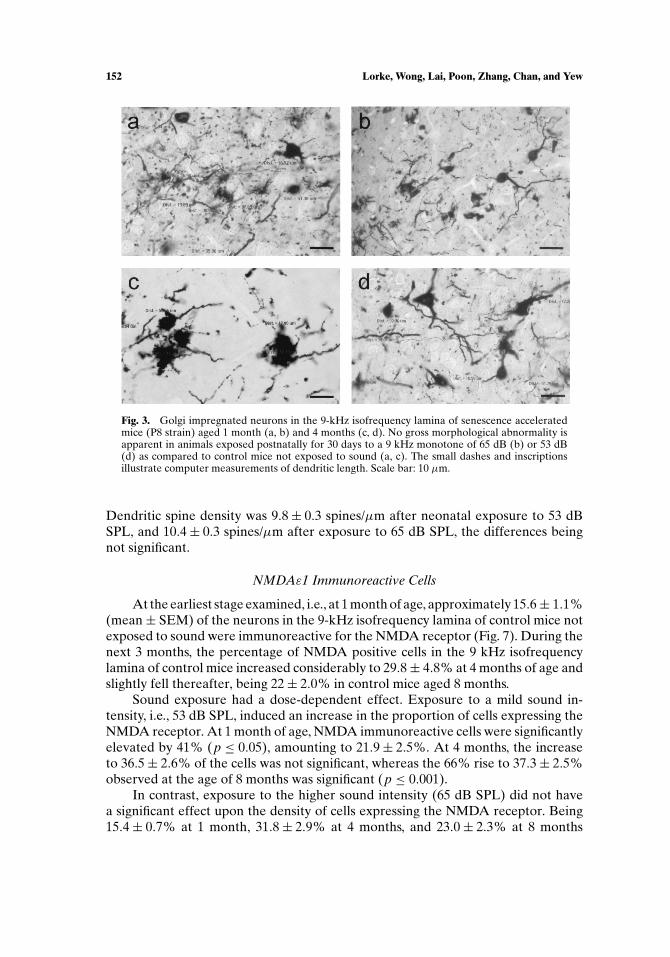
The outline of neurons in the 9 kHz isofrequency lamina was well visible afterGolgi impregnation (Fig. 3). Long dendrites with numerous dendritic spines wereapparent; no gross morphological alterations of neurons were detected after soundexposure.
NMDAε1 Receptor Immunoreaction
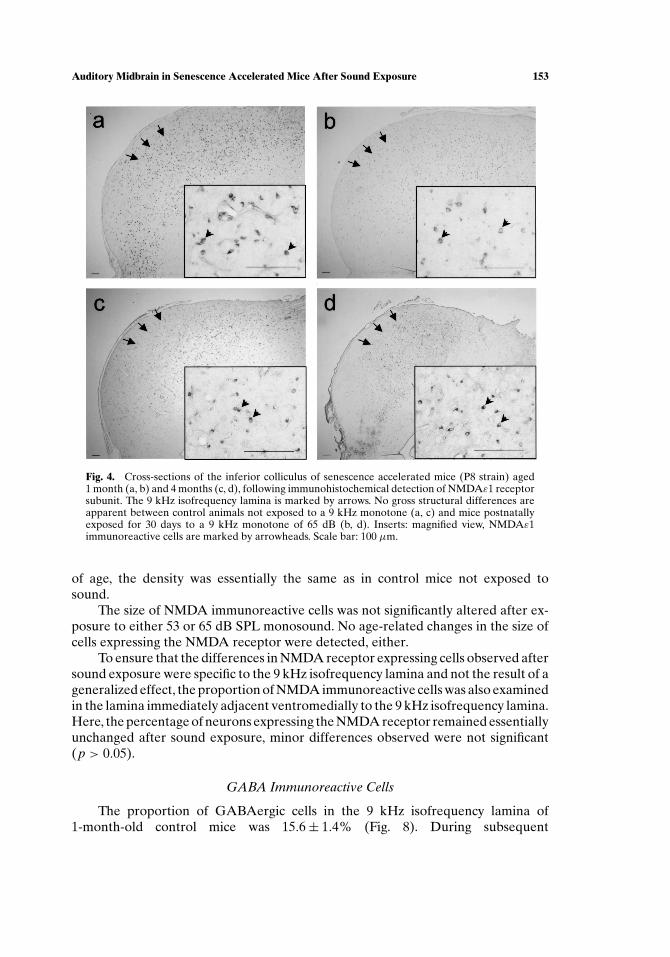
Neurons expressing the NMDA receptor were observed throughout theentire inferior colliculus, including the 9 kHz isofrequency lamina (Fig. 4). Theywere relatively small. Staining was mainly confined to neuronal perikarya, but oc-casionally primary dendrites were also immunoreactive. Gross morpho logical alter-ations after exposure to sound of 53 or 65 dB SPL were not apparent.
GABA Immunoreaction
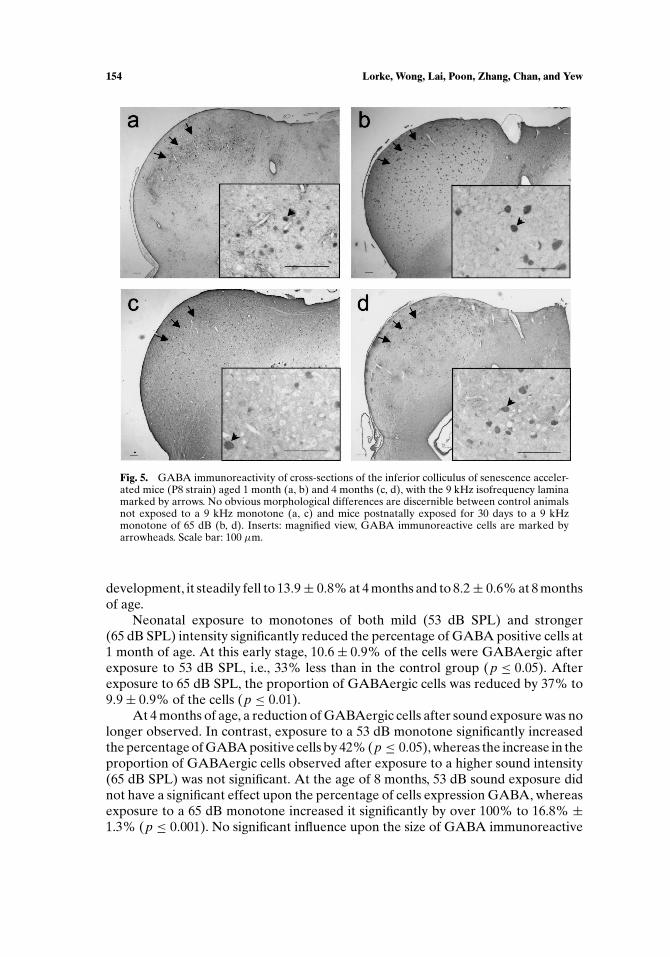
GABAergic neurons were also found in the whole inferior colliculus (Fig. 5).GABA immunoreactivity was detected in the perikarya and the dendrites. No markedstructural changes were observed after sound exposure (53 or 65 dB SPL).
P1: GXB
Cellular and Molecular Neurobiology [cemn] pp793-cemn-461951 March 13, 2003 12:35 Style file version Oct 23, 2000
Auditory Midbrain in Senescence Accelerated Mice After Sound Exposure 151
Fig. 2. Isofrequency lamina (dotted area) for 9 kHz monosoundof the senescence accelerated mouse (P8 strain) inferior colliculus.Three-dimensional reconstruction obtained from summation of la-beling pattern of six sections stained immunohistochemically withan antibody against c-fos protein after exposure to 9 kHz monotoneof 65 dB for 30 min. The ventricle is surrounded by a gray shade.
Morphometry
Quantitation of Dendritic Spines
In 1-month-old control mice not exposed to sound, the density of dendriticspines, as determined by morphometric analysis of Golgi-impregnated neurons in the9 kHz isofrequency lamina, was 10.6± 0.3 spines/µm (mean± SEM; Fig. 6). Duringsubsequent development, dendritic spine density of control mice slightly increasedto 12.0± 0.5 spines/µm at 4 months of age and somewhat declined thereafter to10.5± 0.5 spines/µm at 8 months of age.
Immediately after mild-sound exposure to 53 dB SPL, i.e., at 1 month of age,the dendritic spine density, which was 10.2 ± 0.3 spines µm, was not significantlydifferent from that of the control group. In contrast, exposure to higher sound lev-els, i.e., 65 dB SPL, led to a significant decrease in dendritic spine density in the9 kHz isofrequency lamina (p ≤ 0.01). Being reduced by 11%, it amounted to 9.4±0.3 spines/µm at 1 month of age. By 4 months of age, exposure to both mild (53 dBSPL) and higher (65 dB SPL) sound intensities caused a significant reduction in den-dritic spine density (p ≤ 0.05). The decrease in spine density was more marked afterexposure to higher sound intensity with spine density being 10.7± 0.3 spines/µmafter 53 dB SPL (11% reduction) and 10.0± 0.3 spines/µm after 65 dB SPL soundexposure (17% reduction).
At 8 months of age, i.e., 7 months after cessation of sound exposure, a significanteffect of sound exposure upon the density of dendritic spines was no longer apparent.
P1: GXB
Cellular and Molecular Neurobiology [cemn] pp793-cemn-461951 March 13, 2003 12:35 Style file version Oct 23, 2000
152 Lorke, Wong, Lai, Poon, Zhang, Chan, and Yew
Fig. 3. Golgi impregnated neurons in the 9-kHz isofrequency lamina of senescence acceleratedmice (P8 strain) aged 1 month (a, b) and 4 months (c, d). No gross morphological abnormality isapparent in animals exposed postnatally for 30 days to a 9 kHz monotone of 65 dB (b) or 53 dB(d) as compared to control mice not exposed to sound (a, c). The small dashes and inscriptionsillustrate computer measurements of dendritic length. Scale bar: 10 µm.
Dendritic spine density was 9.8± 0.3 spines/µm after neonatal exposure to 53 dBSPL, and 10.4± 0.3 spines/µm after exposure to 65 dB SPL, the differences beingnot significant.
NMDAε1 Immunoreactive Cells
At the earliest stage examined, i.e., at 1 month of age, approximately 15.6± 1.1%(mean± SEM) of the neurons in the 9-kHz isofrequency lamina of control mice notexposed to sound were immunoreactive for the NMDA receptor (Fig. 7). During thenext 3 months, the percentage of NMDA positive cells in the 9 kHz isofrequencylamina of control mice increased considerably to 29.8± 4.8% at 4 months of age andslightly fell thereafter, being 22± 2.0% in control mice aged 8 months.
Sound exposure had a dose-dependent effect. Exposure to a mild sound in-tensity, i.e., 53 dB SPL, induced an increase in the proportion of cells expressing theNMDA receptor. At 1 month of age, NMDA immunoreactive cells were significantlyelevated by 41% (p ≤ 0.05), amounting to 21.9± 2.5%. At 4 months, the increaseto 36.5± 2.6% of the cells was not significant, whereas the 66% rise to 37.3± 2.5%observed at the age of 8 months was significant (p ≤ 0.001).
In contrast, exposure to the higher sound intensity (65 dB SPL) did not havea significant effect upon the density of cells expressing the NMDA receptor. Being15.4± 0.7% at 1 month, 31.8± 2.9% at 4 months, and 23.0± 2.3% at 8 months
P1: GXB
Cellular and Molecular Neurobiology [cemn] pp793-cemn-461951 March 13, 2003 12:35 Style file version Oct 23, 2000
Auditory Midbrain in Senescence Accelerated Mice After Sound Exposure 153
Fig. 4. Cross-sections of the inferior colliculus of senescence accelerated mice (P8 strain) aged1 month (a, b) and 4 months (c, d), following immunohistochemical detection of NMDAε1 receptorsubunit. The 9 kHz isofrequency lamina is marked by arrows. No gross structural differences areapparent between control animals not exposed to a 9 kHz monotone (a, c) and mice postnatallyexposed for 30 days to a 9 kHz monotone of 65 dB (b, d). Inserts: magnified view, NMDAε1immunoreactive cells are marked by arrowheads. Scale bar: 100 µm.
of age, the density was essentially the same as in control mice not exposed tosound.
The size of NMDA immunoreactive cells was not significantly altered after ex-posure to either 53 or 65 dB SPL monosound. No age-related changes in the size ofcells expressing the NMDA receptor were detected, either.
To ensure that the differences in NMDA receptor expressing cells observed aftersound exposure were specific to the 9 kHz isofrequency lamina and not the result of ageneralized effect, the proportion of NMDA immunoreactive cells was also examinedin the lamina immediately adjacent ventromedially to the 9 kHz isofrequency lamina.Here, the percentage of neurons expressing the NMDA receptor remained essentiallyunchanged after sound exposure, minor differences observed were not significant(p > 0.05).
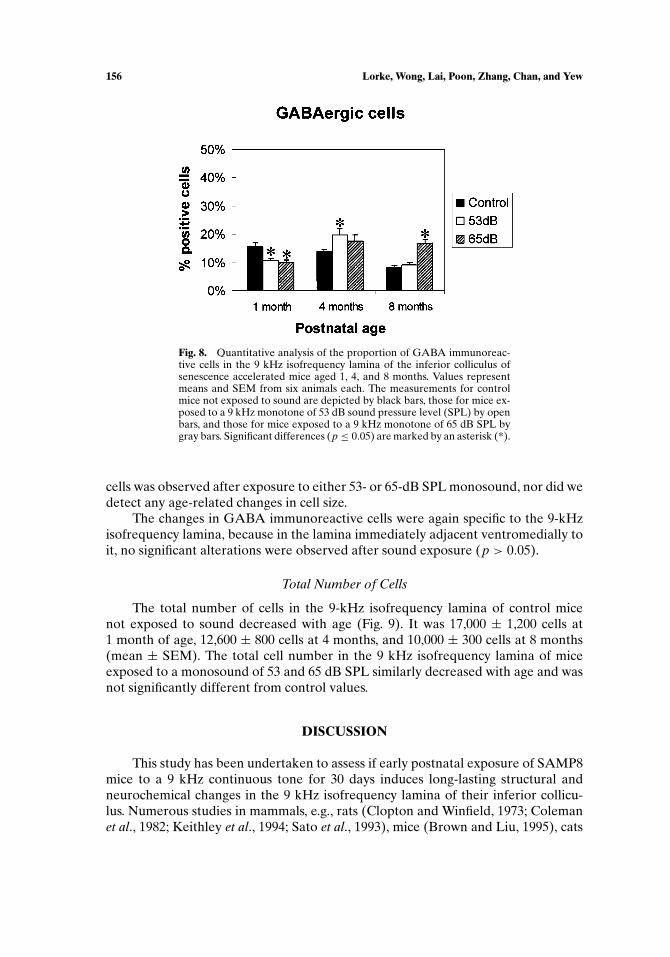
GABA Immunoreactive Cells
The proportion of GABAergic cells in the 9 kHz isofrequency lamina of1-month-old control mice was 15.6± 1.4% (Fig. 8). During subsequent
P1: GXB
Cellular and Molecular Neurobiology [cemn] pp793-cemn-461951 March 13, 2003 12:35 Style file version Oct 23, 2000
154 Lorke, Wong, Lai, Poon, Zhang, Chan, and Yew
Fig. 5. GABA immunoreactivity of cross-sections of the inferior colliculus of senescence acceler-ated mice (P8 strain) aged 1 month (a, b) and 4 months (c, d), with the 9 kHz isofrequency laminamarked by arrows. No obvious morphological differences are discernible between control animalsnot exposed to a 9 kHz monotone (a, c) and mice postnatally exposed for 30 days to a 9 kHzmonotone of 65 dB (b, d). Inserts: magnified view, GABA immunoreactive cells are marked byarrowheads. Scale bar: 100 µm.
development, it steadily fell to 13.9± 0.8% at 4 months and to 8.2± 0.6% at 8 monthsof age.
Neonatal exposure to monotones of both mild (53 dB SPL) and stronger(65 dB SPL) intensity significantly reduced the percentage of GABA positive cells at1 month of age. At this early stage, 10.6± 0.9% of the cells were GABAergic afterexposure to 53 dB SPL, i.e., 33% less than in the control group (p ≤ 0.05). Afterexposure to 65 dB SPL, the proportion of GABAergic cells was reduced by 37% to9.9± 0.9% of the cells (p ≤ 0.01).
At 4 months of age, a reduction of GABAergic cells after sound exposure was nolonger observed. In contrast, exposure to a 53 dB monotone significantly increasedthe percentage of GABA positive cells by 42% (p ≤ 0.05), whereas the increase in theproportion of GABAergic cells observed after exposure to a higher sound intensity(65 dB SPL) was not significant. At the age of 8 months, 53 dB sound exposure didnot have a significant effect upon the percentage of cells expression GABA, whereasexposure to a 65 dB monotone increased it significantly by over 100% to 16.8% ±1.3% (p ≤ 0.001). No significant influence upon the size of GABA immunoreactive

P1: GXB
Cellular and Molecular Neurobiology [cemn] pp793-cemn-461951 March 13, 2003 12:35 Style file version Oct 23, 2000
Auditory Midbrain in Senescence Accelerated Mice After Sound Exposure 155
Fig. 6. Quantitative analysis of spine density, expressed as the numberof spines per 1 µm dendritic length, in the 9 kHz isofrequency laminaof the inferior colliculus of senescence accelerated mice aged 1, 4, and8 months. Values represent means and SEM from six animals each. Themeasurements for control mice not exposed to sound are depicted byblack bars, those for mice exposed to a 9 kHz monotone of 53 dB soundpressure level (SPL) by open bars, and those for mice exposed to a 9 kHzmonotone of 65 dB SPL by gray bars. Significant differences (p ≤ 0.05)are marked by an asterisk (*).
Fig. 7. Quantitative analysis of the proportion of cells immunoreactive forthe ε1 subunit of the NMDA receptor in the 9 kHz isofrequency laminaof the inferior colliculus of senescence accelerated mice aged 1, 4, and8 months. Values represent means and SEM from six animals each. Themeasurements for control mice not exposed to sound are depicted by blackbars, those for mice exposed to a 9 kHz monotone of 53 dB sound pressurelevel (SPL) by open bars, and those for mice exposed to a 9 kHz monotoneof 65 dB SPL by gray bars. Significant differences (p ≤ 0.05) are markedby an asterisk (∗).
P1: GXB
Cellular and Molecular Neurobiology [cemn] pp793-cemn-461951 March 13, 2003 12:35 Style file version Oct 23, 2000
156 Lorke, Wong, Lai, Poon, Zhang, Chan, and Yew
Fig. 8. Quantitative analysis of the proportion of GABA immunoreac-tive cells in the 9 kHz isofrequency lamina of the inferior colliculus ofsenescence accelerated mice aged 1, 4, and 8 months. Values representmeans and SEM from six animals each. The measurements for controlmice not exposed to sound are depicted by black bars, those for mice ex-posed to a 9 kHz monotone of 53 dB sound pressure level (SPL) by openbars, and those for mice exposed to a 9 kHz monotone of 65 dB SPL bygray bars. Significant differences (p ≤ 0.05) are marked by an asterisk (*).
cells was observed after exposure to either 53- or 65-dB SPL monosound, nor did wedetect any age-related changes in cell size.
The changes in GABA immunoreactive cells were again specific to the 9-kHzisofrequency lamina, because in the lamina immediately adjacent ventromedially toit, no significant alterations were observed after sound exposure (p > 0.05).
Total Number of Cells
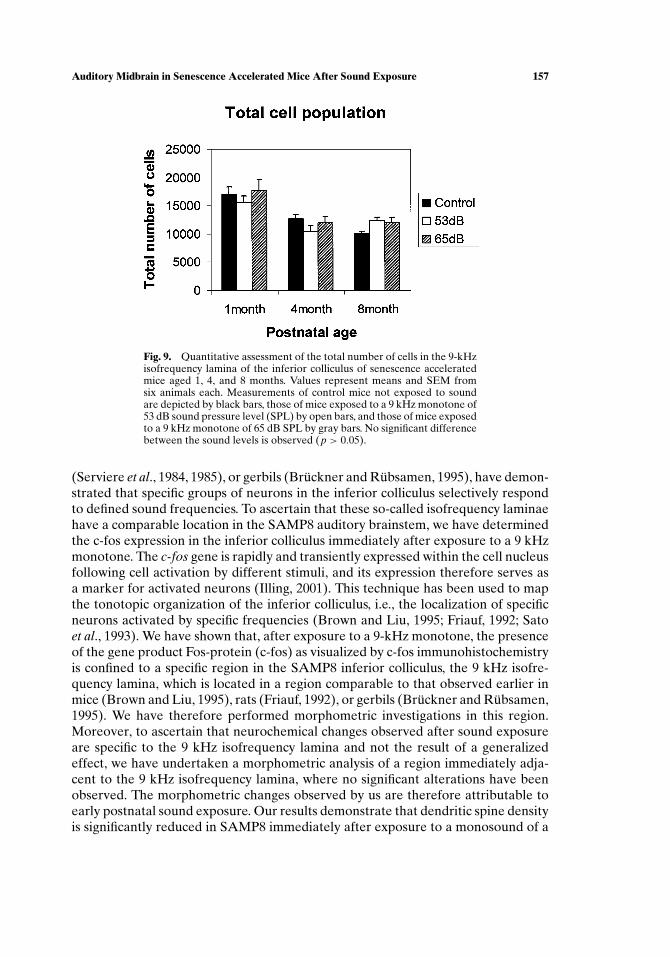
The total number of cells in the 9-kHz isofrequency lamina of control micenot exposed to sound decreased with age (Fig. 9). It was 17,000 ± 1,200 cells at1 month of age, 12,600 ± 800 cells at 4 months, and 10,000 ± 300 cells at 8 months(mean ± SEM). The total cell number in the 9 kHz isofrequency lamina of miceexposed to a monosound of 53 and 65 dB SPL similarly decreased with age and wasnot significantly different from control values.
DISCUSSION
This study has been undertaken to assess if early postnatal exposure of SAMP8mice to a 9 kHz continuous tone for 30 days induces long-lasting structural andneurochemical changes in the 9 kHz isofrequency lamina of their inferior collicu-lus. Numerous studies in mammals, e.g., rats (Clopton and Winfield, 1973; Colemanet al., 1982; Keithley et al., 1994; Sato et al., 1993), mice (Brown and Liu, 1995), cats
P1: GXB
Cellular and Molecular Neurobiology [cemn] pp793-cemn-461951 March 13, 2003 12:35 Style file version Oct 23, 2000
Auditory Midbrain in Senescence Accelerated Mice After Sound Exposure 157
Fig. 9. Quantitative assessment of the total number of cells in the 9-kHzisofrequency lamina of the inferior colliculus of senescence acceleratedmice aged 1, 4, and 8 months. Values represent means and SEM fromsix animals each. Measurements of control mice not exposed to soundare depicted by black bars, those of mice exposed to a 9 kHz monotone of53 dB sound pressure level (SPL) by open bars, and those of mice exposedto a 9 kHz monotone of 65 dB SPL by gray bars. No significant differencebetween the sound levels is observed (p > 0.05).
(Serviere et al., 1984, 1985), or gerbils (Bruckner and Rubsamen, 1995), have demon-strated that specific groups of neurons in the inferior colliculus selectively respondto defined sound frequencies. To ascertain that these so-called isofrequency laminaehave a comparable location in the SAMP8 auditory brainstem, we have determinedthe c-fos expression in the inferior colliculus immediately after exposure to a 9 kHzmonotone. The c-fos gene is rapidly and transiently expressed within the cell nucleusfollowing cell activation by different stimuli, and its expression therefore serves asa marker for activated neurons (Illing, 2001). This technique has been used to mapthe tonotopic organization of the inferior colliculus, i.e., the localization of specificneurons activated by specific frequencies (Brown and Liu, 1995; Friauf, 1992; Satoet al., 1993). We have shown that, after exposure to a 9-kHz monotone, the presenceof the gene product Fos-protein (c-fos) as visualized by c-fos immunohistochemistryis confined to a specific region in the SAMP8 inferior colliculus, the 9 kHz isofre-quency lamina, which is located in a region comparable to that observed earlier inmice (Brown and Liu, 1995), rats (Friauf, 1992), or gerbils (Bruckner and Rubsamen,1995). We have therefore performed morphometric investigations in this region.Moreover, to ascertain that neurochemical changes observed after sound exposureare specific to the 9 kHz isofrequency lamina and not the result of a generalizedeffect, we have undertaken a morphometric analysis of a region immediately adja-cent to the 9 kHz isofrequency lamina, where no significant alterations have beenobserved. The morphometric changes observed by us are therefore attributable toearly postnatal sound exposure. Our results demonstrate that dendritic spine densityis significantly reduced in SAMP8 immediately after exposure to a monosound of a

P1: GXB
Cellular and Molecular Neurobiology [cemn] pp793-cemn-461951 March 13, 2003 12:35 Style file version Oct 23, 2000
158 Lorke, Wong, Lai, Poon, Zhang, Chan, and Yew
higher intensity (65 dB SPL), i.e., at 1 month of age, and 3 months after exposure to amonosound of both mild (53 dB SPL) and higher intensities (65 dB SPL). Dendriticspines are tiny membranous compartments consisting of a head connected to thedendrite by a thin neck. Being the contact sites for most excitatory synapses in thebrain (for a review, see Matus, 2000; Sabatini et al., 2001; Segal et al., 2000), theyare highly dynamic structures that compartmentalize [Ca2+] (Reid, 2002) and showactivity- and experience-dependent morphological plasticity (Lendvai et al., 2000;Segal and Andersen, 2000). The current hypothesis of spine function proposes thatthey constitute the site of neuronal plasticity and long-term, stable memory in CNS(Sabatini et al., 2001; Segal et al., 2000; Segal and Anderson, 2000). This is achievedeither by forming novel spines or by eliminating existing ones (pruning).
Neurons maintain an optimal level of total synaptic input by increasing synapticstrength when activation is low and decreasing it when activation is high (Turrigianoet al., 1998). This can be realized through activity-dependent redistribution of re-ceptors (Craig, 1998) and through regulation of the total number of active synapses(Harris, 1999). Stronger activation results in spine loss as shown in vitro in slice cul-tures and in vivo after epileptic seizures (Harris, 1999). In chicken, auditory imprint-ing, involving exposure to an acoustic stimulus, results in spine loss (Wallhausser andScheich, 1987). Our observation that acoustic stimulation induces reduction in den-dritic spine density is in good agreement with these results. Our results indicate thatthis effect is dose-dependent. The stronger stimulus (65 dB SPL) is intense enoughto already cause a decrease in dendritic spine density immediately after sound ex-posure, i.e., at 1 month of age. In contrast, the effect of the weaker stimulus (53 dBSPL) only becomes apparent 3 months later, i.e., at 4 months of age, when both 53and 65 dB SPL induce a significant reduction in spine density. At a later stage, i.e.,at 8 months of age, when spine density has declined in control animals, this pruningeffect of sound exposure is no longer detectable. The accelerated loss of dendriticspines can be related to electrophysiological measurements in the SAMP8 inferiorcolliculus revealing a general loss in responsive auditory units after sound exposure(Chiu et al., 2002). The decrease in synaptic density observed in the old control infe-rior colliculus supports a similar observation in the C57BL/6 model of presbyacusis(Kazee et al., 1995).
There is ample evidence that activity-dependent and learning-induced selectivereduction of spine synapses requires NMDA receptor activation (for a review, seeConstantine-Paton and Cline, 1998; Jontes and Smith, 2000; Matus, 2000; Segal et al.,2000). NMDA receptors have been implicated in intiating the relevant structuralchanges that result in the refinement of synaptic connection (Constantine-Paton andCline, 1998). Inhibition of the NMDA receptor results in a sixfold increase in spinesin the developing lateral geniculate nucleus (Harris, 1999). Loss of dendritic spines,induced by auditory filial imprinting, can be blocked by NMDA receptor antagonists(Bock and Braun, 1999). Given the effect of acoustic stimulation upon spine density,we have therefore investigated NMDA receptor expression in the inferior colliculus.
Our data show that, in controls not exposed to sound, the percentage of cellswith NMDA-receptor immunoreactivity increases until 4 months of age and slightlydeclines thereafter. The developmental pattern of NMDAε1 receptor expression inthe cerebral cortex and hippocampus of normal aging mice similarly shows an initial

P1: GXB
Cellular and Molecular Neurobiology [cemn] pp793-cemn-461951 March 13, 2003 12:35 Style file version Oct 23, 2000
Auditory Midbrain in Senescence Accelerated Mice After Sound Exposure 159
rise and a subsequent decline (Magnusson et al., 2002). However, peak NMDAε1 ex-pression is only reached at 10 months of age, which is considerably later than in ourstudies. This difference is, however, not surprising considering that the premature ag-ing process of the SAMP8 brain is likely to cause an acceleration of the physiologicalage-related decline in NMDAε1 receptor expression.
The most striking finding is a differential effect of different sound intensitiesupon the proportion of neurons expressing the NMDA receptor. Exposure to soundlevels of mild intensity (53 dB SPL) significantly increases the numerical density ofneurons expressing the NMDA receptor. This effect has been detectable in all agegroups examined, the difference being significant at 1 and 8 months of age. Ourobservation that the number of neurons expressing the NMDA receptors is elevatedafter sound exposure to 53 dB is in good agreement with numerous studies on therelationship between sound exposure and NMDA receptor expression. The NMDAreceptor mediates and regulates excitatory neuronal responses in the adult inferiorcolliculus (Kelly and Zhang, 2002; Ma et al., 2002; Zhang and Kelly, 2001). NMDAreceptors are involved in remodeling dendritic spines, which we have found to bedecreased in density immediately and 3 months after sound exposure. Expression ofNMDA receptor subunit mRNAs in the rat auditory brain stem, i.e., cochlear nucleiand central nucleus of inferior colliculus, is increased after intracochlear electricalstimulation of auditory pathways (Liao et al., 2000). Sensory deprivation decreasesexpression of NMDA receptor subunits ζ1 (NR1), ε1 (NR2A), and ε2 (NR2B) in therat inferior colliculus (Marianowski et al., 2000). The elevated NMDA expression isalso consistent with electrophysiological data showing a rise in spontaneously activeunits in the SAMP8 inferior colliculus after exposure to 53-dB SPL (Chiu et al., 2002).
Although the elevated percentage of NMDAε1 receptor expressing neuronsafter neonatal exposure to 53 dB SPL thus fits well into this picture, our observationthat NMDAε1 receptor expression remains unchanged after exposure to 65 dB SPLis more difficult to interpret. A differential effect of mild and higher sound intensitieshas also been reported in an electrophysiological study of the 9 kHz isofrequencylamina of SAMP8 exposed to a monotone (Chiu et al., 2002). In this study, a weakertone (53 dB SPL) had a stronger effect than the louder one (65 dB SPL). Thereare several possible explanations for this paradox influence. Cochlear damage maybe elicited by the stronger stimulus, but not by the weaker one. The 65 dB soundlevel is, however, some 45 dB weaker than the one usually applied to cause cochlearlesions in mice (Davis et al., 2001; Ohlemiller et al., 2000). It is possible, though,that SAMP8 are more susceptible to sound-induced cochlear damage. Support forthis assumption comes from a study on SAMP1 showing that age-related hearingimpairment and loss of spiral ganglion neurons progress more rapidly in SAMP1than in the senescence-resistant strain (Saitoh et al., 1994). An alternative hypothesisexplaining the absence of changes in NMDR receptor expression may be stress-related changes induced by exposure to a stronger sound intensity. Support for thishypothesis comes from our observation that mortality is markedly increased afterexposure to 65 dB SPL, but not to 53 dB SPL. A thorough histological examinationon the cochlea and the spiral ganglion of SAMP8 after sound exposure as well asan analysis of stress-related hormone levels is therefore envisaged. In summary, itappears that the results of the 65 dB group may be contaminated by effects other than

P1: GXB
Cellular and Molecular Neurobiology [cemn] pp793-cemn-461951 March 13, 2003 12:35 Style file version Oct 23, 2000
160 Lorke, Wong, Lai, Poon, Zhang, Chan, and Yew
a simple rise in neuronal activity during early life. The findings in the 53 dB groupare probably more physiological, an assumption which is supported by a clearer bestfrequency clustering to the exposing tone of 9 kHz after exposure to 53 dB SPL (Chiuet al., personal communication).
In addition, our study shows an effect of sound exposure upon GABAergic cellsin the 9 kHz isofrequency lamina of SAMP8. The slight age-related decline in thepercentage of GABAergic cells observed in control mice not exposed to sound isin good agreement with previous studies showing that the number of GABAergiccells, the GABA content, GABA release, and the activity of the rate-limiting GABAsynthesizing enzyme glutamic acid decarboxylase (GAD) are all reduced in the agingrodent inferior colliculus (Caspary et al., 1995, 1999). Moreover, we have detecteda significant reduction in GABA immunoreactive neurons immediately after expo-sure to a monotone of both 53 and 65 dB SPL. GABA shapes responses to ampli-tude modulated stimuli in the inferior colliculus (Caspary et al., 2002). A decreasedGAD activity has been observed after exposure to traumatic noise levels (Milbrandtet al., 2000), and sensory deprivation increases expression of the GABA(a) subunit(Marianowski et al., 2000). These observations are consistent with our finding ofdecreased GABA immunoreactivity immediately after sound exposure.
After 4 and 8 months, however, we have observed an increase in the percentageof GABAergic immunoreactive neurons in the 9 kHz isofrequency lamina of theinferior colliculus, which is significant at 4 months for a sound level of 53 dB andat 8 months for 65 dB SPL. This long-term rise in the proportion of GABAergicneurons contrasts with the decrease seen as short-term effect of sound exposure. In-creased GABA immunoreaction at older ages is, however, in good agreement withphysiological data collected on 15 month old SAMP8 mice demonstrating a generalloss in auditory sensitivity after the same early postnatal exposure to a 9-kHz mono-tone of 53 or 65 dB SPL (Chiu et al., personal communication). Several underlyingmechanisms for this increased density in GABAergic neurons are conceivable. For-mation of new GABAergic neurons is very unlikely because neurons of the rodentinferior colliculus are generated in the prenatal and early postnatal period (Altmanand Bayer, 1981). Death of non-GABAergic neurons, which may produce a relativeincrease in the proportion of GABAergic neurons, is also not a probable explanation,because we have found the total number of neurons to decrease with age in parallelin all three experimental groups with no significant difference between them. Wetherefore believe that the increased density of GABAergic neurons after exposureto sounds reflects a subset of cells, whose initial level of GABA synthesis has beentoo low to be detected and which have increased GABA synthesis as a result of stim-ulation. It is well known that GABA plays an important role in long-term synapticplasticity (Gaiarsa et al., 2002). Elevated GABA synthesis in the inferior colliculusafter sound exposure is probably a compensatory mechanism to suppress overactivitydue to elevated NMDA expression by an increase in inhibitory GABAergic neurons.A similar rise in GABAergic neurons in the entire inferior colliculus, possibly relatedto elevated neuronal activity, has been found in a study on salicylate-induced tinnitus(Poon et al., unpublished results). Comparable changes have also been reported inother brain regions, e.g., the murine neocortex, where vibrissa stimulation inducesincreased density in GABAergic cells (Siucinska et al., 1999).

P1: GXB
Cellular and Molecular Neurobiology [cemn] pp793-cemn-461951 March 13, 2003 12:35 Style file version Oct 23, 2000
Auditory Midbrain in Senescence Accelerated Mice After Sound Exposure 161
Whatever the underlying cytokinetic basis of this increased density ofGABAergic neurons is, our data both on dendritic spine density, NMDA recep-tor expression, and GABAergic neurons demonstrate that early postnatal exposureto sound results in long-term changes in the 9 kHz isofrequency lamina of SAMP8mice. These observations corroborate parallel electrophysiological findings showingthat exposure of SAMP8 to mild sound levels leads to plastic changes in the inferiorcolliculus lasting into old age and that mild and stronger sound intensities have a dif-ferential effect upon the auditory midbrain (Chiu et al., personal communication). Itis tempting to speculate that early postnatal exposure to a mild sound level has a ben-eficial effect preventing physiological damage, as demonstrated by numerous studies(see Niu and Canlon, 2002, for a review; Willott and Turner, 1999). In contrast, earlyexposure to higher sound intensities, even way below the threshold causing cochlearlesions, possibly has a devastating influence upon the auditory system, which mayonly become apparent at old age.
Our study demonstrates that early postnatal exposure to sound in some regardsaccelerates the normal aging process, as demonstrated by the reduced density ofdendritic spines. Other alterations induced by early sound exposure are not simplyexplicable by accelerated aging, e.g., increased NMDAε1 expression after 53 dB andelevated GABA expression after exposure to both 53 and 65 dB SPL.
ACKNOWLEDGMENTS
The work described in this paper was fully supported by a grant from the Re-search Grants Council of the Hong Kong Special Administrative Region (Project no.CUHK4260/99M). The authors thank Samuel Wong for skillful technical assistancein preparing the photographs.
REFERENCES
Abercrombie, M. (1946). Estimation of nuclear population from microtome sections. Anat. Rec. 94:239–246.
Altman, J., and Bayer, S. A. (1981). Time of origin of neurons of the rat inferior colliculus and the relationsbetween cytogenesis and tonotopic order in the auditory pathway. Exp. Brain Res. 42:411–423.
Bock, J., and Braun, K. (1999). Blockade of N-methyl-D-aspartate receptor activation suppresses learning-induced synaptic elimination. Proc. Natl. Acad. Sci. U.S.A. 96:2485–2490.
Brown, M. C., and Liu, T. S. (1995). Fos-like immunoreactivity in central auditory neurons of the mouse.J. Comp. Neurol. 357:85–97.
Bruckner, S., and Rubsamen, R. (1995). Binaural response characteristics in isofrequency sheets of thegerbil inferior colliculus. Hear. Res. 86:1–14.
Caird, D. (1991). Processing in the colliculi. In Altschuler, R. A., Bobbin, R. P., Clopton B. M., andHoffman, D. W. (eds.), Neurobiology of Hearing: The Central Auditory System, Raven Press, NewYork, pp. 253–292.
Caspary, D. M., Holder, T. M., Hughes, L. F., Milbrandt, J. C., Mckernan, R. M., and Naritoku, D. K.(1999). Age-related changes in GABAA receptor subunit composition and function in rat auditorysystem. Neuroscience 93:307–312.
Caspary, D. M., Milbrandt, J. C., and Helfert, R. H. (1995). Central auditory aging: GABA changes in theinferior colliculus. Exp. Gerontol. 30:349–360.
Caspary, D. M., Palombi, P. S., and Hughes, L. F. (2002). GABAergic inputs shape responses to amplitudemodulated stimuli in the inferior colliculus. Hear. Res. 168:163–173.

P1: GXB
Cellular and Molecular Neurobiology [cemn] pp793-cemn-461951 March 13, 2003 12:35 Style file version Oct 23, 2000
162 Lorke, Wong, Lai, Poon, Zhang, Chan, and Yew
Chiu, T. W., Poon, P. W. F., Yew, D. T. W., and Chan, W. Y. (manuscript submitted for publication). Long-term changes of response in the auditory midbrain of senescence accelerated mice after early soundexposure. J. Neurol. Sci.
Clayton, D. A., and Browning, M. D. (2001). Deficits in the expression of the NR2B subunit in thehippocampus of aged Fischer 344 rats. Neurobiol. Aging 22:165–168.
Clayton, D. A., Grosshans, D. R., and Browning, M. D. (2002). Aging and surface expression of hippocam-pal NMDA receptors. J. Biol. Chem. 277:14367–14369.
Clerici, W. J., and Coleman, J. R. (1987). Resting and pure tone evoked metabolic responses in the inferiorcolliculus of young adult and senescent rats. Neurobiol. Aging 8:171–178.
Clopton, B. M., and Winfield, J. A. (1973). Tonotopic organization in the inferior colliculus of the rat.Brain Res. 29:355–358.
Coleman, J. R., Campbell, M., and Clerici, W. J. (1982). Observations on tonotopic organization withinthe rat inferior colliculus using 2-deoxy-D[1-3H] glucose. Otolaryngol. Head Neck Surg. 90:795–800.
Constantine-Paton, M., and Cline, H. T. (1998). LTP and activity-dependent synaptogenesis: The morealike they are, the more different they become. Curr. Opin. Neurobiol. 8:139–148.
Craig, A. M. (1998). Activity and synaptic receptor targeting: The long view. Neuron 21:459–462.Davis, R. R., Newlander, J. K., Ling, X.-B., Cortopassi, G. A., Krieg, E. F., and Erway, L. C. (2001). Genetic
basis for susceptibility to noise-induced hearing loss in mice. Hear. Res. 155:82–90.Flood, J. F., Farr, S. A., Uezu, K., and Morley, J. E. (1998). Age-related changes in septal serotonergic,
GABAergic and glutamatergic facilitation of retention in SAMP8 mice. Mech. Ageing Dev. 105:173–188.
Friauf, E. (1992). Tonotopic order in the adult and developing auditory system of the rat as shown by c-fosimmunocytochemistry. Eur. J. Neurosci. 4:798–812.
Gaiarsa, J.-L., Caillard, O., and Ben-Ari, Y. (2002). Long-term plasticity at GABAergic and glycinergicsynapses: Mechanisms and functional significance. Trends Neurosci. 25:564–570.
Gross, P. M., Sposito, N. M., Pettersen, S. E., Panton, D. G., and Fenstermacher, J. D. (1987). Topographyof capillary density, glucose metabolism, and microvascular function within the rat inferior colliculus.J. Cereb. Blood Flow Metab. 7:154–160.
Harris, K. M. (1999). Structure, development, and plasticity of dendritic spines. Curr. Opin. Neurobiol.9:343–348.
Helfert, R. H., Galtz, F. R., III, Wilson, T. S., Ramkumar, V., and Hughes, L. F. (2002). Hsp70 in the inferiorcolliculus of Fischer-344 rats: Effects of age and acoustic stress. Hear. Res. 170:155–165.
Helfert, R. H., Sommer, T. J., Meeks, J., Hofstetter, P., and Hughes, L. F. (1999). Age-related synapticchanges in the central nucleus of the inferior colliculus of Fischer-344 rats. J. Comp. Neurol. 406:285–298.
Higuchi K. (1997). Genetic characterization of senescence-accelerated mouse (SAM). Exp. Gerontol.32:129–138.
Illing, R.-B. (2001). Activity-dependent plasticity in the adult auditory brainstem. Audiol. Neurootol.6:319–345.
Jiang, D., McAlpine, D., and Palmer, A. R. (1997). Detectability index measures of binaural masking leveldifference across populations of inferior colliculus neurons. J. Neurosci. 17:9331–9339.
Jontes, J. D., and Smith, S. J. (2000). Filopodia, spines, and the generation of synaptic diversity. Neuron27:11–14.
Kawamata, T., Akiguchi, I., Yagi, H., Irino, M., Sugiyama, H., Akiyama, H., Shimada, A., Takemura, M.,Ueno, M., Kitabayashi, T., Ohnishi, K., Seriu, N., Higuchi, K., Hosokawa, M., and Takeda, T. (1997).Neuropathological studies on strains of senescence-accelerated mice (SAM) with age-related deficitsin learning and memory. Exp. Gerontol. 32:161–169.
Kazee, A. M., Han, L. Y., Spongr, V. P., Walton, J. P., Salvi, R. J., and Flood, D. G. (1995). Synaptic loss inthe central nucleus of the inferior colliculus correlates with sensorineural hearing loss in the C57BL/6mouse model of presbycusis. Hear. Res. 89:109–120.
Keithley, E. M., Lo, J., and Ryan, A. F. (1994). 2-Deoxyglucose uptake patterns in response to pure tonestimuli in the aged rat inferior colliculus. Hear. Res. 80:79–85.
Kelly, J. B., and Zhang, H. (2002). Contribution of AMPA and NMDA receptors to excitatory responsesin the inferior colliculus. Hear. Res. 168:35–42.
Kondziella, D., Bidar, A., Urfjell, B., Sletvold, O., and Sonnewald, U. (2002). The pentylenetetrazole-kindling model of epilepsy in SAMP8 mice: Behavior and metabolism. Neurochem. Int. 40:413–418.
Lendvai, B., Stern, E. A., Chen, B., and Svoboda, K. (2000). Experience-dependent plasticity of dendriticspines in the developing rat barrel cortex in vivo. Nature 404:876–881.
Liao, W.-H., van den Abbeele, T., Herman, P., Frachet, B., Huy, P. T. B., Lecain, E., and Marianowski,R. (2000). Expression of NMDA, AMPA and GABAA receptor subunit mRNAs in the rat auditorybrainstem. II: Influence of intracochlear electrical stimulation. Hear. Res. 150:12–26.

P1: GXB
Cellular and Molecular Neurobiology [cemn] pp793-cemn-461951 March 13, 2003 12:35 Style file version Oct 23, 2000
Auditory Midbrain in Senescence Accelerated Mice After Sound Exposure 163
Ma, C. L., Kelly, J. B., and Wu, S. H. (2002). AMPA and NMDA receptors mediate synaptic excitation inthe rat’s inferior colliculus. Hear. Res. 168:25–34.
Magnusson, K. R., Nelson, S. E., and Young, A. B. (2002). Age-related changes in the protein expressionof subunits of the NMDA receptor. Mol. Brain Res. 99:40–45.
Marianowski, R., Liao, W.-H., van den Abbeele, T., Fillit, P., Herman, P., Frachet, B., and Huy, P. T. B.(2000). Expression of NMDA, AMPA and GABAA receptor subunit mRNAs in the rat auditorybrainstem. I: Influence of early auditory deprivation. Hear. Res. 150:1–11.
Matus, A. (2000). Actin-based plasticity in dendritic spines. Science 290:754–758.McAlpine, D., Jiang, D., Shackleton, T. M., and Palmer, A. R. (1998). Convergent input from brainstem
coincidence detectors onto delay-sensitive neurons in the inferior colliculus. J. Neurosci. 18:6026–6039.
McFadden, S. L., Campo, P., Ding, D., and Quaranta, N. (1998). Effects of noise on inferior colliculusevoked potentials and cochlear anatomy in young and aged chinchillas. Hear. Res. 117:81–96.
Milbrandt, J. C., Holder, T. M., Wilson, M. C., Salvi, R. J., and Caspary, D. M. (2000). GAD levels andmuscimol binding in rat inferior colliculus following acoustic trauma. Hear. Res. 147:251–260.
Miller, J. M., Dolan, D. F., Raphael, Y., and Altschuler, R. A. (1998). Interactive effects of aging with noiseinduced hearing loss. Scand. Audiol. Suppl. 48:53–61.
Mills, J. H., Dubno, J. R., and Boettcher, F. A. (1998). Interaction of noise-induced hearing loss andpresbyacusis. Scand. Audiol. Suppl. 48:117–122.
Morley, J. E. (2002). The SAMP8 mouse: A model of Alzheimer disease? Biogerontology 3:57–60.Morley, J. E., Farr, S. A., and Flood, J. F. (2002). Antibody to amyloid β protein alleviates impaired
acquisition, retention, and memory processing in SAMP8 mice. Neurobiol. Learn. Mem. 78:125–138.
Niu, X., and Canlon, B. (2002). Protecting against noise trauma by sound conditioning. J. Sound Vibrat.250:115–118.
Nomura, Y., Kitamura, Y., Ohnuki, T., Arima, T., Yamanaka, Y., Sasaki, K., and Oomura, Y. (1997).Alterations in acetylcholine, NMDA, benzodiazepine receptors and protein kinase C in the brainof the senescence-accelerated mouse: An animal model useful for studies on cognitive enhancers.Behav. Brain Res. 83:51–55.
Nomura, Y., and Okuma, Y. (1999). Age-related defects in lifespan and learning ability in SAMP8 mice.Neurobiol. Aging 20:111–115.
Ohlemiller, K. K., Wright, J. S., and Heidbreder, A. F. (2000). Vulnerability to noise-induced hearingloss in ‘middle-aged’ and young adult mice: A dose-response approach in CBA, C57BL, and BALBinbred strains. Hear. Res. 149:239–247.
Okuma, Y., and Nomura, Y. (1998). Senescence-accelerated mouse (SAM) as an animal model of seniledementia: Pharmacological, neurochemical and molecular biological approach. Jpn. J. Pharmacol.78:399–404.
Oliver, D. L., and Shneiderman, A. (1991). The anatomy of the inferior colliculus: A cellular basis forintegration of monaural and binaural information. In Altschuler, R. A., Bobbin, R. P., Clopton,B. M., and Hoffman, D. W. (eds.), Neurobiology of Hearing: The Central Auditory System, RavenPress, New York, pp. 195–222.
Onozuka, M., Watanabe, K., Fujita, M., Tomida, M., and Ozono, S. (2002). Changes in the septohippocam-pal cholinergic system following removal of molar teeth in the aged SAMP8 mouse. Behav. BrainRes. 18:197–204.
Palombi, P. S., and Caspary, D. M. (1996). Responses of young and aged Fischer 344 rat inferior colliculusneurons to binaural tonal stimuli. Hear. Res. 100:59–67.
Poon, P. W., and Chen, X. (1992). Postnatal exposure to tones alters the tuning characteristics of inferiorcollicular neurons in the rat. Brain Res. 585:391–394.
Reid, C. A. (2002). The role of dendritic spines: Comparing the complex with the simple. Eur. J. Pharmacol.447:173–176.
Sabatini, B. L., Maravall, M., and Svoboda, K. (2001). Ca2+ signaling in dendritic spines. Curr. Opin.Neurobiol. 11:349–356.
Saitoh, Y., Hosokawa, M., Shimada, A., Watanabe, Y., Yasuda, N., Takeda, T., and Murakami, Y. (1994).Age-related hearing impairment in senescence-accelerated mouse (SAM). Hear. Res. 75:27–37.
Sato, K., Houtani, T., Ueyama, T., Ikeda, M., Yamashita, T., Kumazawa, T., and Sugimoto, T. (1993).Identification of rat brainstem sites with neuronal Fos protein induced by acoustic stimulation withpure tones. Acta Otolaryngol. Suppl. 500:18–22.
Scheibel, M. E., Lindsay, R. D., Tomiyasu, U., and Scheibel, A. B. (1975). Progressive dendritic changesin aging human cortex. Exp. Neurol. 47:392–403.
Scheibel, M. E., Lindsay, R. D., Tomiyasu, U., and Scheibel, A. B. (1976). Progressive dendritic changesin the aging human limbic system. Exp. Neurol. 53:420–430.

P1: GXB
Cellular and Molecular Neurobiology [cemn] pp793-cemn-461951 March 13, 2003 12:35 Style file version Oct 23, 2000
164 Lorke, Wong, Lai, Poon, Zhang, Chan, and Yew
Segal, M., and Andersen, P. (2000). Dendritic spines shaped by synaptic activity. Curr. Opin. Neurobiol.10:582–586.
Segal, M., Korkotian, E., and Murphy, D. D. (2000). Dendritic spine formation and pruning: Commoncellular mechanisms? Trends Neurosci. 23:53–57.
Seidman, M. D., Ahmad, N., and Bai, U. (2002). Molecular mechanisms of age-related hearing loss. AgeingRes. Rev. 1:331–343.
Serviere, J., Webster, W. R., Brown, M., and Kohout, V. (1985). Both [1-14C] glucose and 2-[1-14C]deoxyglucose produce selective iso-frequency labelling in the inferior colliculus of the cat with shortstimulation periods. Neurosci. Lett. 16:127–134.
Serviere, J., Webster, W. R., and Calford, M. B. (1984). Isofrequency labelling revealed by a combined[14C]-2-deoxyglucose, electrophysiological, and horseradish peroxidase study of the inferior collicu-lus of the cat. J. Comp. Neurol. 228:463–477.
Siucinska, E., Kossut, M., and Stewart, M. G. (1999). GABA immunoreactivity in mouse barrel field afteraversive and appetitive classical conditioning training involving facial vibrissae. Brain Res. 843:62–70.
Spangler, E. L., Patel, N., Speer, D., Hyman, M., Hengemihle, J., Markowska, A., and Ingram, D. K. (2002).Passive avoidance and complex maze learning in the senescence accelerated mouse (SAM): Age andstrain comparisons of SAMP8 and R1. J. Gerontol. A. Biol. Sci. Med. Sci. 57:B61–B68.
Takeda, T., Hosokawa, M., and Higuchi, K. (1991). Senescence-accelerated mouse (SAM): A novel murinemodel of accelerated senescence. J. Am. Geriatr. Soc. 39:911–919.
Takeda, T., Hosokawa, M., Takeshita, S., Irino, M., Higuchi, K., Matsushita, T., Tomita, Y., Yasuhira, K.,Hamamoto, H., Shimizu, K., Ishii, M., and Yamamuro, T. (1981). A new murine model of acceleratedsenescence. Mech. Ageing Dev. 17:183–194.
Takemura, M., Nakamura, S., Akiguchi, I., Ueno, M., Oka, N., Ishikawa, S., Shimada, A., Kimura, J.,and Takeda, T. (1993). Beta/A4 proteinlike immunoreactive granular structures in the brain ofsenescence-accelerated mouse. Am. J. Pathol. 142:1887–1897.
Turrigiano, G. G., Leslie, K. R., Desai, N. S., Rutherford, L. C., and Nelson, S. B. (1998). Activity-dependentscaling of quantal amplitude in neocortical neurons. Nature 391:892–896.
Ueno, M., Sakamoto, H., Kanenishi, K., Onodera, M., Akiguchi, I., and Hosokawa, M. (2001). Ultra-structural and permeability features of microvessels in the hippocampus, cerebellum and pons ofsenescence-accelerated mice (SAM). Neurobiol. Aging 22:469–478.
Wallhausser, E., and Scheich, H. (1987). Auditory imprinting leads to differential 2-deoxyglucose uptakeand dendritic spine loss in the chick rostral forebrain. Brain Res. 428:29–44.
Willott, J. F., Chisolm, T. H., and Lister, J. J. (2001). Modulation of presbycusis: Current status and futuredirections. Audiol. Neurootol. 6:231–249.
Willott, J. F., and Turner, J. G. (1999). Prolonged exposure to an augmented acoustic environment ame-liorates age-related auditory changes in C57BL/6J and DBA/2J mice. Hear. Res. 135:78–88.
Zeller, K., Rahner-Welsch, S., and Kuschinsky, W. (1997). Distribution of Glut1 glucose transporters indifferent brain structures compared to glucose utilization and capillary density of adult rat brains.J. Cereb. Blood Flow Metab. 17:204–209.
Zhang, H., and Kelly, J. B. (2001). AMPA and NMDA receptors regulate responses of neurons in the rat’sinferior colliculus. J. Neurophysiol. 86:871–880.
Copyright © 2022 FDOKUMEN




















