
Mechanical properties of bat wing membrane skin
22
J. Zool., Lond. (1996) 239, 357-378 Mechanical properties of bat wing membrane skin S. M. SWARTZ*, M. S. GROVES Department of Ecology & Evolutionary Biology, Brown University, Providence, RI, USA H. D. KIM AND W. R. WALSH~ Division of Engineering, Brown University, Providence, RI, USA (SMS, HDK, WRW) Orthopedic Biomechanics Laboratory, Rhode Island Hospital, Providence, RI, USA (HDK, WRW) (Accepted 27 July 1995) (With 1 plate and 9 figures in the text) The skin of the bat wing is functionally unique among mammals: it serves as a major locomotor organ in addition to its protective and regulatory functions. We used tensile testing to investigate the mechanical capabilities of wing membrane skin, and compared stiffness, strength, load at failure, and energy absorption among specific wing regions and among a variety of bat taxa. We related these characteristics to the highly architectural fibrous supporting network of the wing membrane. We found that all material properties showed a strong anisotropy. In particular, wing membrane skin shows maximum stiffness and strength parallel to the wing skeleton, and greatest extensibility parallel to the wing’s trailing edge. We also found significant variation among wing regions. The uropatagium (tail membrane) supported the greatest load at failure, and the plagiopatagium (proximal wing membrane between lateral body wall and hand skeleton) is the weakest and most extensible part of the wing. We believe that the increased load bearing ability of the uropatagium relates to its key role in capture of insect prey, and that the great extensibility of the plagiopatagium promotes development of camber near the wing’s centre of lift. In interspecificcomparisons, energy absorption and load to failure were greatest in Artibeus jumuicensis, the largest bat in our sample and the species with the highest wing loading, suggesting that wing loading may play a role in dictating the functional design of wing membranes. Introduction Skin serves a number of important functions in all vertebrates, primarily by providing a protective and regulatory barrier between the organism’s internal environment and the external world. However, at least twice in vertebrate history, in the evolution of bats and of pterosaurs, vertebrate skin has taken on an additional major function: formation of a wing membrane used in powered flight. In these taxa, skin must participate in generating lift and thrust and transmitting locomotor stresses during flight. The histological structure of wing skin is distinctive in both bats and pterosaurs, presumably in association with flight function. In pterosaurs, closely spaced parallel fibres, possibly homo- logous to the keratinous scales and feathers of other archosaurs, invested the ventral surface of the wing membrane, transmitting aerodynamic forces from the wing surface to the upper limb skeleton (Padian & Rayner, 1993). In bats, the wing membrane comprises two layers of unusually * Address for correspondence + Present address: Centre for Biomedical Engineering, University of New South Wales, Sydney, Australia 351 0 1996 The Zoological Society of London
Transcript of Mechanical properties of bat wing membrane skin

J . Zool., Lond. (1996) 239, 357-378
Mechanical properties of bat wing membrane skin
S . M. SWARTZ*, M. S . GROVES Department of Ecology & Evolutionary Biology, Brown University, Providence, RI, USA
H. D. KIM A N D W. R. W A L S H ~ Division of Engineering, Brown University, Providence, RI, USA (SMS, HDK, WRW)
Orthopedic Biomechanics Laboratory, Rhode Island Hospital, Providence, RI, USA (HDK, WRW)
(Accepted 27 July 1995)
(With 1 plate and 9 figures in the text)
The skin of the bat wing is functionally unique among mammals: it serves as a major locomotor organ in addition to its protective and regulatory functions. We used tensile testing to investigate the mechanical capabilities of wing membrane skin, and compared stiffness, strength, load at failure, and energy absorption among specific wing regions and among a variety of bat taxa. We related these characteristics to the highly architectural fibrous supporting network of the wing membrane. We found that all material properties showed a strong anisotropy. In particular, wing membrane skin shows maximum stiffness and strength parallel to the wing skeleton, and greatest extensibility parallel to the wing’s trailing edge. We also found significant variation among wing regions. The uropatagium (tail membrane) supported the greatest load at failure, and the plagiopatagium (proximal wing membrane between lateral body wall and hand skeleton) is the weakest and most extensible part of the wing. We believe that the increased load bearing ability of the uropatagium relates to its key role in capture of insect prey, and that the great extensibility of the plagiopatagium promotes development of camber near the wing’s centre of lift. In interspecific comparisons, energy absorption and load to failure were greatest in Artibeus jumuicensis, the largest bat in our sample and the species with the highest wing loading, suggesting that wing loading may play a role in dictating the functional design of wing membranes.
Introduction
Skin serves a number of important functions in all vertebrates, primarily by providing a protective and regulatory barrier between the organism’s internal environment and the external world. However, at least twice in vertebrate history, in the evolution of bats and of pterosaurs, vertebrate skin has taken on an additional major function: formation of a wing membrane used in powered flight. In these taxa, skin must participate in generating lift and thrust and transmitting locomotor stresses during flight.
The histological structure of wing skin is distinctive in both bats and pterosaurs, presumably in association with flight function. In pterosaurs, closely spaced parallel fibres, possibly homo- logous to the keratinous scales and feathers of other archosaurs, invested the ventral surface of the wing membrane, transmitting aerodynamic forces from the wing surface to the upper limb skeleton (Padian & Rayner, 1993). In bats, the wing membrane comprises two layers of unusually
* Address for correspondence + Present address: Centre for Biomedical Engineering, University of New South Wales, Sydney, Australia
3 5 1
0 1996 The Zoological Society of London

358 S . M . SWARTZ ET A L .
thin epidermis on either side of a relatively thin dermis/hypodermis containing nerves, muscle fibres, blood vessels. and an intricate network of connective tissue fibre bundles (Quay, 1970; Sokolov, 1982). In contrast, body skin is single-layered, relatively thick, and in general, not distinctly different from that of non-flying mammals (Sokolov, 1982).
Although flying vertebrates appear to have structurally unique skin, the relationship between skin structure and its mechanical performance capabilities remains poorly understood. Indeed, physical properties of wing skin have received scant attention. In the only study conducted to date, Studier (1972) found a weak relationship between wing skin thickness and body weight in seven species of North American Microchiroptera. His data show no correlation between thickness and puncture strength, but he found some support for the hypothesis that taxa that feed on the ground or in areas of thick or thorny vegetation show increased puncture resistance. The variation in mechanical characteristics among various wing regions, relationships among mechanical parameters, and directional effects in skin properties were not addressed.
Here, we propose that the specialized function of bat wing membrane skin will subject this tissue to forces unique among mammals, and we hypothesize that this function dictates performance requirements for wing skin. To address this issue, we ask: 1) do skin mechanical properties in bat wings differ from the condition seen in skin from other mammalian taxa?; 2) do mechanical characteristics of wing skin vary among bats?; 3 ) do these differences, if any, relate directly to bats’ specialized skin architecture?; 4) does the skin in different parts of the bat wing vary in mechanical characteristics?; and 5) can these differences contribute to improved flight performance?
Bats are a diverse order, with close to a thousand living species, ranging in body size from a few to over 1200 g. They display a variety of dietary preferences and flight modes, varying in relative importance of hovering, gliding, manoeuvrability, flight speed, etc., and comprise at least 14 distinct family lineages. Because powered flight is a mechanically demanding activity, we predict that selection will operate to minimize skin mass and hence the mass of wing tissue to be supported and oscillated. We therefore expect that wing membrane skin will be thinner than predicted by standard allometric equations relating skin thickness to body mass in mammals (Calder, 1984). The mechanical requirements imposed on wings will depend on characteristics such as body size, flight mode, and dietary type (e.g. insect capture vs. fruit eating), particularly since wing loading (weight supported per unit area of wing surface) will change with mass and food load. We predict that characteristics relating to load bearing ability will be enhanced in species characterized by large body size and/or regular carrying of large food loads, particularly fruits. Adaptations for these biological roles can, however, only occur in the context of phylogenetic history, so in the absence of perfect optimization of structural design to functional requirements, we also predict that skin of closely related taxa will be more similar than that of more distant relatives.
Background
Bat wings comprise several distinct regions which vary in function during flight and in prey capture. The proximal region of the wing, the plagiopatagium, includes within it the centre of lift and hence has greater involvement in the production of lift, while the manoeuvrable, relatively prehensile hand wing, the rluctj~lopatagitmz, is functionally more important in generating thrust (Vaughan, 1970; Altenbach, 1979) (Fig. 1 ) . The tail membrane or uropatugium also generates some lift and thrust during flight, but additionally plays an important role in prey capture in

BAT WING SKIN MECHANICAL PROPERTIES 359
Dactylopatagi
9
n
FIG. 1. Diagram of ventral view of a bat wing showing the specific wing regions investigated in our study: the dactylopatagium, the plagiopatagium, and the uropatagium, as well as the propatagium, not included in these analyses. Test sampling regions are indicated with asterisks.
insectivorous species, functioning as a scoop to propel prey items toward the hunting bat's open mouth. We hypothesize that functional differentiation is reflected in the mechanical characteristics of the wing skin, and that such differences, if they exist, can be understood in reference to the functional differentiation of the wing, particularly along a proximal to distal gradient.
The bat wing membrane is supported by a combination of muscular and connective elements, many of which are grossly apparent in a transilluminated wing (Plate I). The primary connective tissue framework of the wing is a system of collagen and elastin fibre bundles organized into a regular, mesh-like scaffolding (Holbrook & Odland, 1978). The fibre bundles of this soft tissue skeleton are readily visible to the naked eye when the wing is illuminated; they range from 130-300 pm in diameter in a diverse sample of mega- and microchiropteran bats (Swartz & KO, unpubl. data). It has been suggested that the fibrous net permits flexibility while providing tensile strength and limiting wing extensibility (Holbrook & Odland, 1978). The architecture of this fibrous skeleton has been described only for Tadarida brasiliensis; the following is taken from this account, and certainly applies to other species to at least some degree. The bundles extend in a network among the bones of the arm, forearm, and hand, or from these bones to the muscle bundles in the leading or trailing edge of the wing (Plate I, Fig. 2). Fibres traverse the wing in straight, curved, or branching paths. Within the plagiopatagium, and the dactylopatagium to a lesser extent, fibre bundles form an orthogonal net, with each bundle crossed by a number of others at approximately right angles to its primary orientation. The fibres appear to approach the bones at 90" to the bones' long axes. Within this net, nerves, skeletal muscle fibre bundles, and blood vessels traverse the wing. In T. brasiliensis, few cross-hatching fibres appear in the more distal portions of the wing.
SEM of fibre bundle cross-sections and amino acid analysis of wing membrane proteins show

360 S. M . SWARTZ ET A L .
P L A T E I . Photograph of ventral surface of the left elbow region of a bat wing (Peropteryx kuppleri) illuminated from the dorsal aspect. The propatagium and part of the plagiopatagium are shown. The gross architecture of the fibre network is readily apparent. with the orthogonal net pattern displayed in the region between the radius and the fifth digit. Fine lines indicate corrugations running roughly parallel to radius and fifth digit and at varying angles to the predominant fibre bundles. h, humerus: r, radius; mcv, fifth metacarpal; ppv. fifth proximal phalanx; f, representative fibres; c, representative corrugations.
that each bundle comprises a composite of collagen and elastin fibres (Holbrook & Odland, 1978). The collagen and elastin fibres run in a largely parallel fashion, although the elastin fibres appear to branch within each bundle. Each bundle has a core composed primarily of elastin, with a 'sheath' of collagen fibres around its circumference. Elastic fibres appear to dominate the bundles, accounting for around 90% of the fibres in a typical cross-section (Holbrook & Odland, 1978). This is probably of critical importance in determining the mechanical characteristics of wing skin. Elastic fibres from other mammalian tissues typically show a unique ability to extend reversibly to about twice their resting length (Harkness, 1968), primarily owing to the behaviour of the protein elastin, while extensibility of collagen is much more limited (Harkness, 1968; Silver, 1987).
Muscles of two types insert into the connective tissue network. Some relatively large muscle bundles originate on the wing skeleton and insert into the membrane network; they anchor the membrane to the bone and are beiieved to control membrane tension and camber. In many taxa, there is a discrete coracocutaneous muscle originating from the coracoid process of the scapula and inserting into the fibre network of the plagiopatagium, and a humeropatagialis muscle, originating from the distal humeral shaft and extending distally into the plagiopatagium (Strickler, 1978). Intrinsic muscles, originating and inserting within the membrane itself, are

BAT WING SKIN MECHANICAL PROPERTIES
(4 361
FIG. 2. (a) The layout of fibre bundles in a typical bat wing; craniocaudally running plagiopatagiales muscles are also illustrated. The details of this pattern vary among bat species. (b) Schematic of a cross-section through a typical wing membrane network fibre bundle; white fibres are elastin; black fibres are collagen (adapted from Holbrook & Odland, 1978).
also a common feature of bat wings. Microchiropteran bats lack the smooth muscles of the arrectores pilorum, and it has been suggested that the abundant and complex skeletal muscles of wing membranes have secondarily taken over this smooth muscle function (Quay, 1968). The intrinsic muscles are typically restricted to the plagiopatagium, running craniocaudally in parallel to digit V, and vary among bats in cross-sectional area, length, and number. There is no obvious correlation between muscular development and wing structure or flight mode (Gupta, 1967).
Based on structural design, there are several possible functions for the fibrous skeleton. Small muscle bundles within the membrane can insert or originate from the fibres (Holbrook & Odland, 1978), allowing precise control of wing shape and tension. The fibre bundles themselves insert on to the wing bones perpendicular to their long axes, providing a means for modulating bone loading. The elastin components may facilitate passive contraction when the wing joints are flexed, and help keep the wing tightly folded to prevent tearing during climbing, roosting, etc. Most likely, the fibre network determines the mechanical properties of the wing, and given the large elastin contribution, one might expect that wing skin may have properties comparable to those of mammalian elastic ligaments.
Methods
To assess wing membrane properties under tensile loading, we excised test samples of uniform size and shape from the plagiopatagium (the proximal area of the wing membrane bordered by the body, the arm, the fifth digit, the trailing edge, and the hindlimb), dactylopatagium (the distal area of the membrane between the digits) and uropatagium (connecting the hindlimbs and the body, including the tail when present) (Fig. 1) of previously frozen cadavers of a number of bat taxa (Table I). The taxonomic sampling includes both

362 S . M. SWARTZ E T A L .
TABLE I Summurj, of charuc~trristic~ of ta.w included in this studv. Sample si:e for each species folloivs species name in parentheses, with the ,first number giring the numher of individuals sampled and the second giving the number of test specimens. Wing loading. iispect ratio. and dietar! categorization values are front Norberg & Rayner, 1987: additional characteristics are from
the same source and N o ~ a k , 1991 ~
Wing Body mass loading
Taxonomic assignment (9) (g/m2) Aspect ratio Diet Other characteristics
Vespertilionidae M.voti.7 lucifugzrs (9. 54) A4,wtis velifer (2. 14) Epresicus .fuscus ( 5 . 30) Lasiiirus cinereiis ( 1.7)
Pteronotus parnellii ( 2 . 16) Pteronotus simapirrensi~ ( 1.8)
'4rtibeus jamaicensi.c ( I , 7 ) Uroderma hilobutum (1, 7 )
Taduridu hrasiliensis (2 . 9 )
Mormoopidae
Ph yllostomidae
Molossidae
7.1 9.1
15.9 33.0
22.9 8.1
47.0 15.4
12.5
1.5 6.3 9.4
16.5
8.8
16.6 10.1
11.5
low low average high
average average
low low
high
insectivorous insectivorous insectivorous insectivorous
insectivorous insectivorous
frugivorous frugivorous
insectivorous
migratory strong, direct flight low foraging migratory, rapid flight
low level foraging low level foraging
carry large fruit loads short tail membrane
very fast, migratory
Acetate template (bottom layer)
Skin sample
Fiti. 3. Illustration of specimen in gripping template. Excised skin samples are glued to bottom layer of acetate template. then top template layer is folded down to enclose the specimen (along path of curved arrows). Once the three layers (skin sandwiched between top and bottom layers of template) are securely positioned within the materials testing appdratUS. both layers of the acetate template are cut at the indicated site to free the skin sample.

BAT WING SKIN MECHANICAL PROPERTIES 363
insectivorous and frugivorous species, and samples almost an order of magnitude range of body mass. We use the term specimen to refer to each individual bat, and sample to designate each excised piece of skin. Samples from the plagiopatagium were cut from the portion of the wing between the radius and the fifth metacarpal. Samples from the dactylopatagium were removed from between the fourth and fifth digit, and uropatagial samples were taken from any area large enough to sample; we exercised caution to avoid regions of the uropatagium enclosing the calcar, a cartilaginous extension from the bat heel. We used standard dumb-bell-shaped die to cut our samples, to direct tissue failure to the narrow central region of the sample (Glaser et al., 1965; Ridge & Wright, 1966; Veronda & Westman, 1970; Shadwick, Russell & Lauff, 1992) (Fig. 3). Because even slight defects can lead to large stress concentrations, we took care to ensure that samples were not nicked. Each sample was 10 mm in length and about 2 mm wide at the narrowest region. We kept specimens hydrated at all times with normal saline. All samples were tested within 12 hours of excision.
Each sample was painted with a white square in the centre of the neck region. With the 4 corners as registration points, we used videotapes of each test to determine sample distortion during testing and to compare elongation within the sample itself to displacement of the grips. We found no evidence of grip slippage or other end effects. We also used the decrease in specimen width observed in these video records to calculate changing sample cross-sectional area for determination of true strain.
To investigate anisotropy in skin mechanical properties, we quantified specimen orientation relative to anatomical landmarks. We measured sample anisotropy in 2 ways. First, we measured the angle between the sample's long axis and the orientation of the primary fibres of the fibre network within the neck of the hourglass; in many taxa, the orientation of the fibres in most areas of the wing membrane is clearly visible. At each site, we cut samples at approximately O", 45", and 90" to primary fibre orientation, yielding a
I Fibre or I- ~ _.. corrugation I L-. -\-- angle I
I
I
Fibres or corrugations
Fibres or corrugations
, ,
Fibre or corrugation
axis
I
Sample axis
,
FIG. 4 . Illustration of measurement of fibre or corrugation angle relative to specimen orientation.

364 S . M . SWARTZ ET A L .
maximum of 9 samples per specimen (3 each from the plagio-, dactylo-, and uropatagium). To quantify this angle more accurately after the sample was cut, each was mounted on a light box under a video camera with a macro lens; this image was then input into a computer, and image analysis software (ImageAnalyst v. 8.1, Billerica, MA) was used to measure accurately the magnified view of the specimen (Fig. 4). We define this characteristic as fibre angle (FA).
Secondly, we measured specimen orientation with respect to the fine corrugations running through the plagio- and dactylopatagium. These corrugations directly reflect normal patterns of skin folding, and fan outward in a primarily craniocaudal direction, roughly parallel to the adjacent metacarpals and phalanges (Plate I ) . Hence. specimens whose long axes are cut parallel to the long axis of the hand bonds are also roughly parallel to the corrugation lines. We found this angle was readily quantified in an objective way using the methods described above. We define this characteristic as corrugation angle (CA). We found that the uropatagium, the portion of the membrane that does not have a consistent pattern of folding when the animals wings are retracted, had only randomly oriented corrugations, so we could not measure corrugation angle from these specimens. The uropatagium is also the smallest region of the wing membrane, and in some specimens we were unable to harvest more than a single test specimen of uropatagial skin.
To facilitate placement of the samples between testing grips, samples were attached to acetate templates using cyanoacrylate adhesive. These templates resemble window frames, and were used to orient and secure the specimen in a stable fashion so that a minimum of specimen handling was required in loading the sample into the testing apparatus (Fig. 3). This is particularly critical because of the tendency of skin samples to curl after being cut owing to tension in the elastic fibres. After we closed the hydraulically controlled grips on the materials testing machine (MTS 810, St. Paul, MN) around the sandwich of skin and acetate farming, we cut the sides of each frame so that only the skin bridged the space between grips (Fig. 3).
We expect that the aerodynamic forces exerted against the membrane during flight place it under tension, so we tested specimens in this loading mode. Furthermore, most published information concerning mechanical properties of skin have used uniaxial tensile tests, and our use of this method facilitates comparisons of bat wings to skin from other taxa. It is likely that biaxial tests would better reflect the in vivo mechanical behaviour of wing membrane tissue, particularly as unrestricted uniaxial tests may overestimate sample extension. However, apparatus for biaxial testing was not readily available; we believe the results of the uniaxial tests reported here allow valid comparisons within our sample, as well as to other studies. We preconditioned specimens in tensions by applying cyclical loads (10 cycles at 100% strain/min for 6 sec [to 10% strain]), to establish a consistent test baseline. This strain rate probably underestimates in vivo strain rates for wing membranes, but allows comparisons with published values for skin and other soft tissues. Following this preconditioning regimen, the load-displacement curve shifted to the right, showing a small increase in stiffness. No further changes in the curve occurred after 10 cycles, and 4-5 cycles were usually sufficient to precondition the specimens fully. We followed preconditioning with a 10-minute stress relaxation period to ensure that our tests measured only the effect of the applied test load and not residual internal stresses from the preconditioning load cycles. Each specimen was then loaded to rupture in tension at the strain rate used for preconditioning; strain rate was kept constant to failure. Force-displacement output from the materials testing apparatus were plotted on an X-Y plotter and sampled at 60Hz by computer and stored to disk for later analysis.
To distinguish between structural properties (dependent on sample geometry) and material properties (constant for a given material regardless of structural shape), we accurately measured specimen geometry. Length and width were uniform for all specimens since they were cut with a single custom-machined die; thickness was measured to the nearest 0.01 mm with a digital micrometer. When the wing membrane is stretched by hand, it undergoes considerable extension and then becomes extremely stiff; at this point it becomes difficult to extend the skin further without rupture. Because the thickness of wing skin varies with degree of membrane extension, we made all thickness measurements with the wing membrane taut. We made all thickness measurements with the wing membrane stretched to this point of greatly increased stiffness: these measurements were highly repeatable (less than 4% variation among trials of a given specimen). We used these thickness measurements to test allometric predictions concerning skin thickness.

BAT WING SKIN MECHANICAL PROPERTIES 365
We constructed stress-strain curves from the test data sampled for each specimen (Fig. 5) and computed tensile strength (maximum true stress recorded before failure), maximum load, failure strain (strain at failure stress), elastic modulus (stiffness), and energy absorption before failure (total area under the stress- strain curve). Modulus was calculated for the linear region of the stress-strain curve. Our complete data set comprises values for each of these material characteristics at several orientations and for each of three major wing regions. All data were natural log transformed to improve normality of distributions.
Results
Comparisons to other mammals
The range of variation in mechanical properties is large within bats compared to the total range of variation for other vertebrates, especially mammals, although only limited comparative data are available at present (Table 11). Strain at failure for mammals studied to date typically ranges from 0.80 to 1.00; with somewhat lower values for the white rhinoceros (Ceratotherium simum). In bats, we found variation in failure strain from 0.13 to 2.32. Similarly, bat wings show twice the range of variation in strength observed in other mammalian skin, and twice the variation in
0.0 0.2 0.4 0.6 0.8 1 .o Strain
FIG. 5. Representative stress-strain curve for a sample of bat skin. This sample, from the plagiopatagium of Tuduridu brasiliensis, shows the J-shaped loading curve typical of collagenous tissues, with low stiffness at low stress, followed by a region of significantly higher elastic modulus before failure occurs.

366 S. M. SWARTZ E T A L .
TABLE I1 Summary of tensile material clzaracteristics of bat wing skin deterniined in comparison to those of non-flying vertebrates. For rhinoceroses and onagers, standard errors in parentheses folloic means; for bats, frogs. and snakes, range values in
parentheses follows means
Tissue source ~ ~ ~ - ~ ~ -
Failure strain Strength (MPa) Young's modulus (MPa)
Human abdomen: Human abdomen- Rat back' Rat back4 White rhinoceros (C. simurn)
dorsolateral body? White rhinoceros belly5 Onager (Equus hernionrts)' Hippopotamus (Hippopotamus
amphibius)' back and flank Hippopotamus belly' Snake body' Frog (Xenopus laevi .~)~ Eel9 Norfolk spot" Skipjack tuna" Bat wing
1 .oo 0.90 0.80 1 .oo
0.24 (0.01)
0.33 (0.02) 0.86 (0.08)
__ 0.43 (0.17-0.69) 0.93 (0.59-1.26)
~
-
0.68 (0.13-2.32)
14.00 5.00 4.00
12.50 30.5 (1.08)
14.5 (1.59)
35 1.01 (1.63)
17
13.28 (1 1.41-16.45) 22.39 (5.00-82.95)
~
~
7.41 (1.14-26.38)
22.4 6.4
~
44 237.3 (9.75)
107.8 (13.9) 31.1 (3.4)
~
129.11 (15.40-584.50) 21.74 (10.39-38.39) 3.54a, 14.7', 156' 2.41", 16.4b, 75.0' 6.92a, 60.2b, 36.2'
23.92 (1.02-250.26)
a = longitudinal directio?. = hoop direction. = on-fibre direction ' Dunn & Silver. 1983, - Dal , 1982. 'Trelstad & Silver, 1981, 4Vogel, 1979, Shadwick et al., 1992, 'Harkness & J, Harkness, 1964. ' Jayne, 1988, Greven. Zanger & Schwinger. 1995. Hebrank. 1980, lo Hebrank & Hebrank, 1986
elastic modulus when the highly specialized protective skin of rhinos is excluded from com- parison. Both maximum strength and maximum stiffness of wing membranes are comparable to those of the rhinoceros dermal amour . Mean energy absorbed was 1.92 MJ/m' (range: 0.19-6.42), in comparison to mean values of 2.9-3.7 for large mammals (Shadwick et al., 1992). The variation in mechanical properties of wing skin reflects both variation in material properties within a single individual and variation among bat taxa (see below, Comparisons among taxa). The only vertebrates studied to date with comparable variation in mechanical properties are snakes (Jayne. 1988; Table 11).
Material characteristics show a distinct pattern of intercorrelation among variables (Table 111). All material property variables show significant correlations with corrugation angle; bat skin
T A B L E 111 Pearson correlation 1natri.v for ull bat data. r values that are not significantly different from zero are indicatd as -.
Correlations with corrugation angle are displayed in bold print
corrugation angle fibre angle In CSA In strength In failure c In modulus In energy
fibre angle -0.39
In strength -0.56 - -0.24 In failure e 0.74 0.21 -0.37 In modulus -0.12 - -0.23 0.77 -0.78
- 0.49 0.57 In energy 0.29 - ~
__ - In CSA
~
In max. load --0.50 0.44 0.76 -0.20 0.55 0.39

BAT WING SKIN MECHANICAL PROPERTIES 361
L
a - m / - * m l e
-1 .. -. I. :>-:I
h
$ - %
* 0
8 I II 1.
O N 7
i"
T-
o

368 S . M . SWARTZ E T A L .
properties are highly orientation dependent (Fig. 6). However, fibre angle is significantly correlated only with corrugation angle, and not with any other mechanical parameters. Cross- sectional area, a direct reflection of skin thickness given that all specimens were of uniform shape and size, is not significantly correlated with corrugation angle; a specific skin region is characterized by a particular thickness, regardless of the orientation of the soft tissue skeleton of the sample.
This extreme orientation dependence of wing membrane skin in mechanical properties differs from the much less anisotropic skin of other mammals and reptiles (Conabere & Hall, 1950; Kanagy el al., 1952; Harkness & Harkness, 1964; Ridge & Wright, 1966; Bauer, Russell & Shadwick, 1989). In most mammalian skin, collagen fibres are arranged in a complex feltwork, with little preferential orientation, and strength differences in various directions differ less than two-fold (Harkness, 1968). Wing membrane anisotropy relates directly to the wing's skeletal architecture (Table 111, Plate I, Fig. 7). Corrugation lines in the plagio- and dactylopatagium are roughly parallel to adjacent wing bones, so CA of 0" corresponds to a sample whose long axis runs craniocaudally along the finger bones' axes, and CA of 90" indicates that the sample axis runs mediolaterally, perpendicular to the bones. Variation in mechanical properties with respect to CA thus also reflects variation relative to the bone axes. The significant negative correlations among modulus, strength, maximum load, and CA indicate that these values decrease as sample orientation deviates from the craniocaudal axis, reaching their minima as orientation approaches perpendicular to the wing bones. In contrast, maximum strain at failure is positively correlated with CA, and hence is greatest perpendicular to the bones.
-failure // strain /
FIG 7 , Schematic of anisotropic variation in mechanical properties related to the wing's anatomical framework. Modulus, strength. maximum. and cross-sectional area are greatest parallel to the fifth digit; failure strain, in contrast, is greatest perpendicular to the fifth digit.

BAT WING SKIN MECHANICAL PROPERTIES 369
In our sample, skin thickness showed no significant correlation with body mass. However, analysis of data from a diversity of mammalian skins has demonstrated that total skin mass (M,) is related to body mass (M) as M, = 0.106 M0.94 (M in kg; r2 = 0.992, N = 42) and skin surface area (A) as A = 0.11 1 M0.65 m2 (r2 = 0.903; Calder, 1984). Following Shadwick et al. (1992), we assumed skin density of 1100 kg/m3, and calculated skin thickness as skin volume/area = 0.86 M0.29 mm. This relationship predicts a single thickness of skin at 0.22 mm for a 10 g bat, and 0.36mm for a 50g bat. Because wing membranes comprise two skin layers sandwiched together, membrane thickness represents two skin layers, and predicted skin thickness by these equations becomes 0.44 mm and 0.72 mm, respectively. Actual skin thickness measured in this study ranged from 0.039 to 0.267 mm, with the mean for Myotis (< 10 g body mass) at 0.1 1 mm, and for Artibeus jamaicensis (approximately 50 g) equal to 0.14 mm. Thus, wing membrane skin is 4-10 times thinner than would be predicted for bat-sized mammals on the basis of body size alone.
Comparisons among wing regions
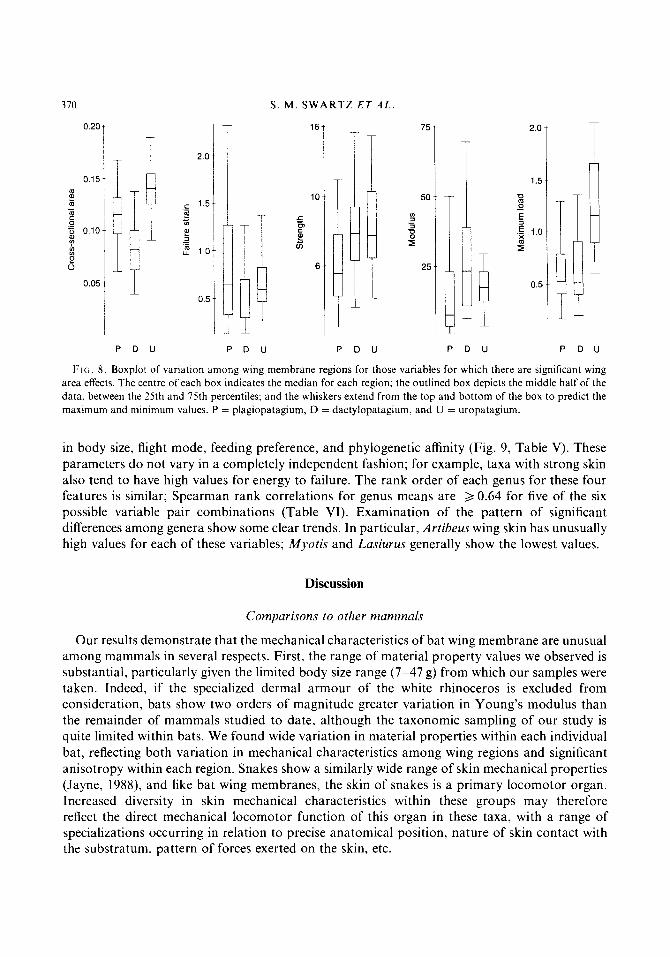
We carried out ANOVAs for each variable to determine which mechanical characteristics showed significant variation among wing regions; for those variables with significant differences, further tests of individual contrasts (plagiopatagium vs. dactylopatagium, plagiopatagium vs. uropatagium, and dactylopatagium vs. uropatagium) were carried out to specify which regions were responsible for group differences. We found that all mechanical characteristics of bat wing skin except energy stored to failure vary significantly among wing regions (Table IV, Figs 7 & 8). In particular: 1) skin thickness is greatest in the uropatagium and least in the dactylopatagium; 2) maximum supportable load is greatest in the uropatagium and roughly the same in the plagio- and dactylopatagium; 3) the plagiopatagium is the weakest part of the wing; and 4) the dactylopatagium is the least extensible part of the wing.
Comparisons among taxa
To test for differences among genera, we carried out ANOVAs with both genus and wing region as main effects. In no case were interactions among genus and wing region significant, so the final analyses include only the main effects. Scheffk post hoc tests for significant differences among subgroups show that most material properties (maximum load, strength, failure strain, modulus, and energy absorbed before failure) showed significant variation among taxa that differ
TABLE IV Matrix of P-values for significance tests of mean differences by wing region; significant results are highlighted in bold print. Only variables for which F-tests of multiple means showed signijicant differences are included here
plagiopatagium vs. plagiopatagium vs. dactylopatagium vs. dactylopatagium uropatagium uropatagium
In CSA 4 0.0001 < o.oO01 < 0.0001 In strength 0.0002 0.0011 0.7807 In failure strain 0.0212 0.9974 0.0083 In modulus 0.0012 0.0529 0.2260 In maximum load 0.7868 4 0.0001 < o.Ooo1
S . M . SWARTZ E T A L 370
0.20
0.15 m F - m 0 .- 5 0.10
I 5
0.05
T
I i
0.5
P D U P D U P D U
75
50
Y)
3 0 -
2. 25
2.0
1.5
P D U
11 P D U
FIG. 8 . Boxplot of variation among wing membrane regions for those variables for which there are significant wing area effects. The centre of each box indicates the median for each region; the outlined box depicts the middle half of the data, between the 25th and 75th percentiles; and the whiskers extend from the top and bottom of the box to predict the maximum and minimum values. P = plagiopatagium, D = dactylopatagium. and U = uropatagium.
in body size, flight mode, feeding preference, and phylogenetic affinity (Fig. 9, Table V). These parameters do not vary in a completely independent fashion; for example, taxa with strong skin also tend to have high values for energy to failure. The rank order of each genus for these four features is similar; Spearman rank correlations for genus means are 2 0.64 for five of the six possible variable pair combinations (Table VI). Examination of the pattern of significant differences among genera show some clear trends. In particular, Artibeus wing skin has unusually high values for each of these variables; Myotis and Lasiurus generally show the lowest values.
Discussion
Comparisons to other mammals
Our results demonstrate that the mechanical characteristics of bat wing membrane are unusual among mammals in several respects. First, the range of material property values we observed is substantial, particularly given the limited body size range (7-47 g) from which our samples were taken. Indeed, if the specialized dermal armour of the white rhinoceros is excluded from consideration, bats show two orders of magnitude greater variation in Young’s modulus than the remainder of mammals studied to date, although the taxonomic sampling of our study is quite limited within bats. We found wide variation in material properties within each individual bat, reflecting both variation in mechanical characteristics among wing regions and significant anisotropy within each region. Snakes show a similarly wide range of skin mechanical properties (Jayne, 1988), and like bat wing membranes, the skin of snakes is a primary locomotor organ. Increased diversity in skin mechanical characteristics within these groups may therefore reflect the direct mechanical locomotor function of this organ in these taxa, with a range of specializations occurring in relation to precise anatomical position, nature of skin contact with the substratum, pattern of forces exerted on the skin, etc.

BAT WING SKIN MECHANICAL PROPERTIES
(a) Plagiopatagiurn
(b) Dactylopatagiurn 1.4
1 .o
0.6
- 5.00
2.50
1.25
I
A E L M P T U (c) Uropatagiurn
3.0
2.0
1 .o
- 6.0
4.5
3.0 $ a,, - 1.5
A E M P T U
1 1.61
16
12
a
4
14
A E L M P T U
1 1.25
A ~ M P T U
A E L M P T U
i
A E M P T U
15
12
9
6
371
- I
Maximum load Energy absorbed Failure strain Strength (MPa) before failure (MJrn”) (MN)
FIG. 9 . Boxplot of variation among genera, by region. (a) Plagiopatagium. (b) Dactylopatagium. (c) Uropatagium. A = Artibeus, E = Eptesicus, L = Lasiurus, M = Myotis, P = Pteronotus, T = Tadarida, U = Uroderma.

372 S. M . SWARTZ E T A L
TABLE V Matrices of genus contrast Srheffe post-hoc significance test.s,for A N 0 V A with genus and wing area as main factors. P-values for significant test.s are given; bold tjpe indicates statistical significance at < 0.05. Top row indicates3rst letter abbreviation
.for genus names
A E
In maximum load Arfibeus Epresicus Lasiurus
Pterono t us Tadarida Uroderma
In strength Artibeus Epresicus Lasiurus Myofis Pferonofus Tadar ida Uroderma
In failure strain Art ibeus Epfesicus Lasiurus Mjmis Pteronotus Tadarida Uroderma
A rtibeus Eptesicus Lusiurtrs Myoti.c Ptcronotus Tadaricia Uroderma
,wj,ot;s
In energy absorbed
0.002 0.001
< 0.001 < 0.001
0.032 0.009
0.121 0.185 0.002 0.004 0.136 0.012
0.243 0.116 0.324 0.162 0.057 0.256
0.122 0.056 0.002 0.006 0.056 0.397
0.046 0.002 0.287 0.999 0.999
0.728 0.004 0.014 0.831 0.067
0.725 0.985 0.454 0.016 0.084
0.273 0.003 0.021 0.372 0.663
L
0.999 0.999 0.037 0.998
0.246 0.345 0.844 0.336
0.736 0.589 0.008 0.049
0.594 0.882 0.618 0.238
M
0.953 0.013 0.783
0.716 0.045 0.963
0.465 0.023 0.106
0.521 0.120 0.027
P
0.406 0.999
0.090 0.833
0.017 0.060
0.319 0.074
T
0.999
0.142
0.547
0.337
Beyond the within-individual and intraspecific variation, we found significant variation among taxa. We propose that this variation reflects the unique mechanical role of skin in bats in comparison to other mammals. In bats, skin is central to locomotor function. A wide range of critical aspects of locomotor performance may be influenced by skin’s mechanical characteristics. We would expect, then, to see variation in these characteristics in relation to flight function,
TABLE V I Spc~arnrati rank curre/ation.s ofgpnus nreans.for variables sl7ort.itig significanr variation among genera
In In energy maximum In failure to failure load strain In strength
-~ __ In energy to failure 1000 In mdximum lodd 0 786 1000 In failure Strain 0 750 0 750 1 000 In strength 0 643 0 750 0 286 1 000

BAT WING SKIN MECHANICAL PROPERTIES 373
rather than relatively uniform mechanical characteristics from skin over a large portion of the body.
The proximate cause of this large variation in mechanical properties cannot yet be conclusively demonstrated. The skin’s collagen network, particularly the size of fibres and the degree of cross-linking present, are believed to be the primary determinants of skin strength (Harkness, 1970; Viidik, 1980). Indeed, Fry, Harkness & Harkness (1963) have shown that rat skin increases in strength and decreases in extensibility as collagen content and degree of cross-linking increase during ageing. The variation we see within bats, both within and among individual wings, may be due to variation in collagen content, structural organization, cross-linking, or a combination of these effects. Preliminary data from our lab suggest that bat taxa differ from one another in the gross spatial organization of the fibre network; we observe interfamilial variation in fibre density, geometry of fibre tract organization, and, to a lesser degree, size of individual fibre bundles (Swartz & KO, unpubl. data). Given that elastic fibres are also particularly prominent in wing skin, there may also be variation among taxa and/or wing regions in the amount and organization of elastic tissue within collagen/elastin bundles.
We find that wing membrane skin is far more anisotropic than other vertebrate skins. For example, rhinoceros skin varies in mechanical properties between the dorsolateral and belly regions, but shows no anisotropy (Shadwick et al., 1992). The anisotropy is almost certainly a function of the architectural organization of the unique bat wing skin collagen-elastin network: in contrast, most skin collagen is organized as a randomly arrayed feltwork of convoluted collagen fibres (Shelac, 1957; Gibson, 1977). This is not, however, the only known case in which specialization of skin function for a particular ecological role has led to a great increase in anisotropy in the specialized lineage. Bauer et al. (1989) found that fragile-skinned gekkonid lizards, in which the superficial skin layers readily release from underlying tissue as a predator escape mechanism, have highly anisotropic skin in the regions from which skin breaks off easily (Bauer et al., 1989). In bat wings, however, the anisotropy is even more pronounced, with stiffness varying close to 250-fold.
We suggest that the marked anisotropy of bat skin is related to skeletal function. Bones are strongest along their long axes, with much less ability to resist forces or deformations perpendi- cular to the diaphysis. We find here that wing membrane skin is strongest and least extensible in the same orientation as the greatest strength and stiffness of the bones. Because of the particularly elongated, slender form of bat wing bones, any forces applied to the bones that are not directed along the longitudinal axis can result in substantial deflections. We propose that if wing skin were not modified for increased stiffness parallel to the bones’ long axes, intense loading of the wing skin by aerodynamic forces during downstroke would cause large skin extensions parallel to the digits. Since the bones are very stiff relative to skin, they will deform little. At the interface of the bone and skin, differences in extensibility would tend to lead to shearing of wing skin off the bones. To the extent that skin stiffness can be increased parallel to the bone axes, the disadvantageous consequences of this shearing can be reduced.
Skin stiffness in bats is not only relatively higher than the typical mammalian condition along the bone long axes, it is significantly lower perpendicular to the bones. This reduced stiffness serves to facilitate the large shape changes the wing experiences during the wingbeat cycle and during unfolding and folding of wing membranes at the beginning and conclusion of flights. The large extensions possible in the mediolateral direction allow a large amount of energy to be absorbed before failure, reaching maximum values of over 6 MJ/m3, contributing to the reported puncture

314 S. M . S W A R T Z ET A L .
resistance of wing membrane (Studier, 1972). This is particularly important given the constraint imposed on extensibility by the need to limit skin deformations in parallel with the wing bones.
It is also possible that material anisotropy in wing membranes is relevant for elastic energy storage and release. As aerodynamic forces are exerted against the wing membrane during downstroke, the membrane will stretch in all directions, but, owing to the large differences in skin stiffness parallel and perpendicular to the bone axes, deformation will be far greater in the perpendicular direction (mediolaterally in the region between the fourth and fifth digits and in the plagiopatagium). The elastic tissues oriented in this direction, or with some component in this direction, are thus stretched by the forces of downstroke. At the end of downstroke, the orientation of the wing surface approaches vertical, and the force exerted perpendicular to the wing surface drops. The wing membrane may recoil elastically as the aerodynamic force drops in magnitude. This recoil may have a role in drawing the digits together in preparation for the upstroke, during which the wing is typically flexed and folded relative to the downstroke.
We also observed that fibre angle and corrugation angle showed a significant negative correlation. It has been suggested that the corrugations in bat wing membranes arise due to tension in the primary fibre bundles, and hence should always be oriented at right angles to them (Holbrook & Odland, 1978). However, our data show that the correlation between fibre and corrugation orientation is weak (r2 = 0.152). None the less, it is the orientation of our samples with respect to corrugations, not fibre tracts, that has the strongest relation to material characteristics. We propose that the tensioning of the crossed-fibre network is not uniform throughout the mesh, resulting in a complex biaxial stress field within the membrane plane. As a consequence, corrugation angle may vary somewhat independently of fibre architecture. Our observations of independence of mechanical properties and fibre bundle orientation support the view that the stress distribution within the network is complex. In a simple collagenous tissue such as tendon, stiffness and strength are greatest in parallel with the predominant fibre orientation; the absence of this customary pattern in wing skin may indicate interactions among the crossed-fibre arrays, or unusual interactions between the fibre bundles and the extracellular matrix.
Our results show that bat wing skin is considerably thinner than would be predicted from the regular pattern of skin thickness scaling described for other mammals (Calder, 1984). It is plausible that this serves to decrease significantly overall mass to be transported by metabolically expensive powered flight, given that each wing contributes 8-15% of the bat’s total body mass and that skin and other soft tissues can make up 60% of total wing mass (Thollesson & Norberg, 1991; Swartz, unpubl. data). Of that 6O%, a significant fraction is muscle, but the skin remains a sizeable component of wing mass, particularly in the distal wing, where there is little musculature and where reduction of wing mass can most dramatically influence the wing mass moment of inertia. It is also possible that the thickness of bat wing skin is driven by aerodynamic considerations. Wing skin may be thin to ensure that it will assume a three-dimensional configuration with particular aerodynamic performance characteristics when exposed to pressure gradients of the magnitude generated during bat flight. Finally, wing membranes also serve an important thermoregulatory function in bats, and the reduced thickness may help promote heat dissipation.
Comparisons among wing regions
We predicted that the regional differentiation in function among the arm, hand, and tail wing should be reflected in adaptations of wing skin to their primary tasks. We found that mechanical

BAT WING SKIN MECHANICAL PROPERTIES 375
characteristics differ distinctly among regions of the bat wing. The plagiopatagium is the weakest, most extensible region, while the uropatagium is the thickest and supports the greatest load before failure. We propose that these differences probably relate directly to the functional differentiation of the proximal, distal, and tail portions of the wing membrane.
The uropatagium generates some lift during flight (Vaughan, 1970), although its contribution is far less than that of the rest of the wing. It also serves an important role in prey capture among insectivorous bats. As these bats approach flying prey items, they pull the uropatagium forward with the legs, and may scoop or catapult a moth or other flying insect up toward the open mouth. Because the uropatagium is responsible for forcibly propelling prey, the skin must be strong enough to perform this task without failure. Some insects may also have pointed projections on their body or limb surfaces, maximizing the importance of puncture resistance in this region.
The plagiopatagium, immediately adjacent to the bat’s body and containing the aerodynamic centre of pressure, plays the major role in generating lift (Vaughan, 1970; Altenbach, 1979; Norberg, 1990). Its greater extensibility may allow greater billowing during downstroke, thereby increasing wing camber and improving the airfoil function of the wing. In contrast, the surface contours of the carpal and metacarpal joint surfaces permit supination and pronation of the dactylopatagium, as well as flexion and extension at the carpometacarpal, metacarpophalangeal, and interphalangeal joints during flight. These motions position the wing in such a way that a component of the lift force is directed forward; this portion of the wing therefore plays a great role in providing thrust and controlling manoeuvrability (Vaughan, 1970). A relatively stiff membrane in this region may allow it to minimize drag more effectively during these movements, reducing the energy required to carry out the requisite wing motions and maximizing the thrust produced.
Bat taxa differ in the degree to which different portions of the wing participate in these contrasting functions; in some species there is little or no pronation/supination of the handwing, and in others these movements are particularly important and involve a large fraction of the wing surface (Norberg, 1990). It would be possible to explore our ideas further relating mechanical characteristics to aerodynamic function in future studies by comparing skin from various subregions of the wing in taxa that differ specifically in the details of their flight kinematics.
Comparisons among taxa
Despite our limited sample, we found significant differences among bats in cross-sectional area (skin thickness), load and energy absorbed to failure and strength. However, our taxonomic sampling is limited in diversity, and sample sizes for each species are small, so our data are therefore insufficient to resolve fully sources of variation in mechanically relevant parameters. Comparisons at the genus and family levels, for example, are dependent upon the number of species in these levels among the various groups. Similarly, separating dietary from phylogenetic effects requires, at minimum, that each dietary group be reasonably well represented among multiple phylogenetic lineages. Although the data reported here did not resolve a number of important issues, we observed several clear trends, particularly in relation to wing loading and to body mass.
Artibeus, the largest bat in our sample and the species with the highest wing loading, withstood greater loads and absorbed more energy before failure. These characteristics may well be driven by the variation in skin thickness; skin that is thicker will have a greater volume of tissue for a given surface area and a greater tissue volume for any cross-section of the wing. Thus, all else being equal, thick skin will bear more load and absorb more energy before failure. Thickness

376 S. M . SWART2 E T A L
alone, however, will not determine strength differences, since strength is normalized to sample cross-sectional area.
Artibeus’ distinctiveness could also be due to differences between frugivorous and insecti- vorous habits, since Artibeus is a fruit eater. However, the other frugivorous taxon included here, Uruderma, while similar to Artibeus in having unusually high values for skin thickness (CSA) and energy absorbed before failure, contrasts markedly with Artibeus in its low values for maximum load and strength. Artibeus is unique in our sample in its combination of high body mass, high wing loading, and fruit-based diet. At present, it appears that the mechanical aspects of increased load due to body weight are more important than frugivory and its associated increase in load carrying in dictating mechanical characteristics of skin.
Mpfk and Lusiurus stand out in contrast to Artibeus as the taxa with the lowest values for strength, load and energy to failure, and thckness. Again, some part of the load and energy dflerences are due to very thin skin in these species, and some part is due to intrinsic characteristics. Both genera are migratory, flying long distances on a regular basis. Specialization for strength related characteristics may be less important than features that improve aerodynamic performance for long duration, energetically expensive flight behaviours. It is possible that whatever specializations of skin maximize aerodynamic performance, perhaps by changing characteristics of airflow over the wing surface, compromise strength, load bearing ability, extensibility, and energy absorption.
Conclusion
A variety of mechanically important properties of bat wing membrane (cross-sectional area, stiffness, strength, extensibility) vary according to orientation within the wing, anatomical region, and across taxa. Although mammalian skin typically varies little in mechanical proper- ties, we believe these data suggest that the structural design of soft tissues are influenced by selective forces in much the same manner as architecture of skeletal elements and overall wing shape. These soft tissue characteristics may directly affect the forces acting on the wing skeleton, and help dictate the flight performance capabilities of bats. The pattern we observe indicates that selection acting to increase wing loading in bats may operate not only by alterations in wing shape (Norberg & Rayner, 1987), but also via changes in material composition and organization of membrane tissues. More thorough allometric studies are needed to discern these relationships, as well as detailed information on skin function during flight, and the variation in this function among wing regions and among taxa. With this information, it will be possible to establish more clearly the relationships among skin architecture, mechanical characteristics, and flight performance; we hope to address these issues in our future work.
Specimens used in this study were generously contributed by: Drs T. Kunz, W. Lancaster, J. Ryan, J. Simmons, and N. Simmons. Amy Ritter provided invaluable technical assistance. Ed Mullen ingeniously created the tissue sample cutting apparatus. Dave Carrier, Jennifer Gray Chickering, Kathleen Earls, Antigone Parker, and Philip Watts provided insightful comments on an earlier version of this manuscript. This work is supported by NSF IBN-9119143 to SMS.
REFERENCES
Altenbach, S. J. (1979). Locomotor morphology of the vampire bat, Desmodus rotundus. Spec. Puhl. Am. SOC. Mammal. 6
Bauer, A. M.. Russell. A. P. & Shadwick, R. E. (1989). Mechanical properties and morphological correlates of fragile skin
Calder. W. A, , 111 (1984). Size./uncrion, and life histor!,. Cambridge: Harvard University Press.
1-133.
in gekkonid lizards. J. ESP. Biol. 145 79-102.

BAT WING SKIN MECHANICAL PROPERTIES 377
Conabere, G. 0. & Hall, R. H. (1950). Directional variations in strength and stretch in calf, goat, and sheep leather. J. Soc.
Daly, C . H. (1982). Biomechanical properties of dermis. J. Invest. Dermatol. 7 9 17s-20s. Dunn, M. G. & Silver, F. H. (1983). Viscoelastic behavior of human connective tissues: relative contribution of viscous
and elastic components. Connect. Tissue Res. 12 59-70. Fry, P., Harkness, M. & Harkness, R. D. (1963). Mechanical properties of collagenous framework of skin in rats of
different ages. Am. J. Physiol. 206: 1425-1429. Gibson, T. (1977). The physical properties of skin. In Reconstructive plastic surgery: principles and procedures in
correction. reconstruction and transplantation: 69-77. Converse, J. M. (Ed.) Philadelphia: W. B. Saunders. Glaser, A. A,, Maragoni, R. D., Must, J., Beckwith, T., Brody, G., Walker, G. &White, W. (1965). Refinements in the
methods for the measurement of the properties of unwounded and wounded skin. Med. Electon. Biol. Engng 3
Greven, H., Zanger, K. & Schwinger, G. (1995). Mechanical properties of the skin of Xenopus laevis (Anura, Amphibia).
Gupta, B. B. (1967). The histology and musculature of the plagiopatagium in bats. Mammalia 31: 313-321. Harkness, R. D. (1968). Mechanical properties ofcollagenous tissues. In Treatiseon collagen: 247-310. Gould, B. S. (Ed.).
NY: Academic Press. Harkness, R. D. (1970). Functional aspects of the connective tissues of the skin. In Chemistry and molecular biology ofthe
intercellular matrix: 1309-1340. Balazs, E. A. (Ed.). NY: Academic Press. Harkness, M. R. & Harkness, R. D. (1964). Some mechanical properties of collagenous frameworks and their functional
significance. In Proceedings of the Fourth International Congress on Biorheology, Pt 4: 477-489. Copley, A. L. (Ed.). NY: Interscience.
Hebrank, M. R. (1980). Mechanical properties and locomotor functions of eel skin. Bid. Bull. (Woods Hole) 158 58-68. Hebrank, M. R. & Hebrank, J. H. (1986). The mechanics of fish skin: lack of an “external tendon” role in two teleosts.
Holbrook, K. & Odland, G. F. (1978). A collagen and elastin network in the wing of the bat. J. Anat. 126 21-36. Jayne, B. C. (1988). Mechanical behaviour of snake skin. J. Zool., Lond. 214 125-140. Kanagy, J. R., Randall, E. B., Carter, T. J. , Kinmonth, R. A. & Mann, C. V. (1952). Variations of physical and chemical
properties within and between vegetable retanned cow and steer sides. J. Am. Leather Chem. Assoc. 47: 726-748. Norberg, U. M. (1990). Vertebrate j ight: mechanics. physiology, morphology, ecology and evolution. Berlin:
Springer-Verlag. Norberg, U. M. & Rayner, J. M. V. (1987). Ecological morphology and flight in bats (Mammalia; Chiroptera): wing
adaptations, flight performance, foraging strategy and echolocation. Philos. Trans. R. Soc. Lond. B 316 335-427. Nowak, R. M. (1991). Walker’s mammals of the world. 5th edn. Baltimore: Johns Hopkins University Press. Padian, K. & Rayner, J. M. V.{1993). The wings of pterosaurs. Am. J. Sci. 293A 91-166. Quay, W. B. (1968). Structure and evolutionary implications of the musculi arrectores pilorum in Chiroptera. Anat. Rec.
Quay, W. B. (1970). Integument and derivatives. In Biology of batsII: 2-56. Wimsatt, W. A. (Ed.). New York: Academic
Ridge, M. D. &Wright, V. (1966). The directional effects of skin: a bioengineering study of skin with particular reference
Shadwick, R. E., Russell, A. P. & Lauff, R. F. (1992). The structure and mechanical design of rhinoceros dermal amour.
Shelac, B.-J. (1957). Hides, skins, and leather under the microscope. British Leather Manufacturers’ Research Association,
Silver, F. H. (1987). Biological materials: Structure, mechanicalproperties and modelling of soft tissues. (NYU Biomedical
Sokolov, V. E. (1982). Mammal skin. Berkeley & Los Angeles: University of California Press. Strickler, T. L. (1978). Functional osteology and myology of the shoulder in Chiroptera. Basel: S. Karger. Studier, E. H. (1972). Some physical properties of the wing membrane of bats. J. Mammal. 53 623-625. Thollesson, M. & Norberg, U. M. (1991). Moments of inertia of bat wings and body. J. Exp. Bid . 158 19-35. Trelstad, R. L. &Silver, F. H. (1981). Matrix assembly. In Cellbiology of extracellular matrix: 179-215. Hay, E. D. (Ed.).
Vaughan, T. A. (1970). Flight patterns and aerodynamics. In The biology of bats 1: 195-216. Wimsatt, W. A. (Ed.).
Leather Trades. Chem. 34: 57-66.
411-419.
J . Morph. 224 15-22.
Biol. Bull. (Woods Hole) 171: 236-247.
163 587-594.
Press.
to Langer’s lines. J . Invest. Dermatol. 46: 341-346.
Philos. Trans. R. Soc. Lond. B 337: 419-428.
Egham, Surrey.
Engineering Series). New York: NYU Press.
New York: Plenum Press.
London & New York: Academic Press.

378 S . M. SWARTZ E T A L
Veronda. D. R. & Westman, R. A. (1970). Mechanical characteristics of skin finite deformations. J . Biomech. 3 11 1-124. Viidik. A. ( 1980). Interdependence between structure and function in collagenous tissues. In Biology of collagen: 257-280.
Vogel. H. G. (1979). Mechanical properties of rat skin at high and low loads: Influence of age and desmotropic drugs. In Viidik, A. & Vuust. J. (Eds). London: Academic Press.
Bioengineeririg and the skin: 97-101. Marks, R. & Payne, P. A. (Eds). Boston: MTP Press.



















