
The gait ofAustralopithecus
23
The Gait of Australopithecus C. OWEN LOVEJOY,’j2 KINGSBURY G. HEIPLE L2 AND ALBERT H. BURSTEIN 2 1 Department of Anthropology, Kent State University, Kent, Ohio 44242 and 2 Biomechanics Laboratory, Division of Orthopaedic Surgery, Case Western Reserue University, Cleveland, Ohio 441 06 KEY WORDS Australopithecus . Gait . Hominids . Locomo- tion . Stride. ABSTRACT A biomechanical analysis of the pelvic and femoral samples available for Australopithecus is presented. No feature of these samples was found to distinguish their gait pattern from that of modern man or to differ in the two presently recognized allomorphs of Australopithecus. Morphological dif- ferences between Australopithecus and modern man appear to be the result of different degrees of encephalization rather than any difference in locomotor adaptation. Almost all who have discussed australo- pithecine locomotion have concluded that these extinct hominids were at least par- tially bipedal. They have not, however, considered them capable of a striding gait - a specialization usually considered to be restricted to Homo erectus and Homo sa- piens (Le Gros Clark, ’67; Napier, ’67). Napier has also stressed the importance of differences between the locomotor patterns of the “robust” and “gracile” forms (‘64). Although such conclusions are in agree- ment with those of a majority of other authors (Mednick, ‘55; Kern and Straus, ’49; Washburn, ’60; Straus, ’62; Chopra, ’62; Campbell, ’66; Day, ’69; Zhilman, ’69; Preuschoft, ’71), they are not in ac- cord with those of Dart (’49, ’57, ’58). In previous papers (Lovejoy and Heiple, ’70, ’72; Heiple and Lovejoy, ’71) it has been demonstrated that some of these earlier conclusions are based upon errors of ob- servation and a failure to take into account the normal range of variation of Homo sapiens. In this paper we shall present a bio- mechanical analysis of australopithecine locomotion. Basing our arguments upon the morphological and biomechanical re- quirements of the striding gait of modern man, we believe that the antiquity of the striding gait can be shown to be as great as the terminal Pliocene. Since the argument to which we address ourselves in this paper is stride, we define it for this study as: the ability to (1) walk in an upright bipedal progression, succes- sively placing each foot closely under the center of gravity; (2) stabilize the pelvis against rotation by an abductor mecha- nism adequate to eliminate large trunk shifts; and, (3) utilize the principle of the compound pendulum for the lower extrem- ity so that the advance of each limb is ac- complished by gravity and short bursts of muscle contraction at accelerations and decelerations. Such energy conservation is an essen- tial feature of man’s locomotor efficiency and can in large part be considered the sine qua non of stride. While the available skeletal material of Australopithecus is not complete enough for the type of biomechanical analysis now possible for Homo sapiens, a sufficient amount of material has been recovered to 1 The definition of stride presented here is an opera- tional one for the purposes of this paper. It utilizes strictly anatomical criteria. “Stride” may also be de- fined on the basis of energy expenditure and gait effi- ciency. In this case the relative contributions of muscu- lar contraction and momentum to locomotion must be considered. This has been presented in Lovejoy (’72). AM. J. PHYS. ANTHROP., 38: 757-780. 757
Transcript of The gait ofAustralopithecus

The Gait of Australopithecus
C. OWEN LOVEJOY,’j2 KINGSBURY G. HEIPLE L 2 AND ALBERT H. BURSTEIN 2
1 Department of Anthropology, Kent State University, Kent, Ohio 44242 and 2 Biomechanics Laboratory, Division of Orthopaedic Surgery, Case Wes tern Reserue University, Cleveland, Ohio 441 06
KEY WORDS Australopithecus . Gait . Hominids . Locomo- tion . Stride.
ABSTRACT A biomechanical analysis of the pelvic and femoral samples available for Australopithecus is presented. N o feature of these samples was found to distinguish their gait pattern from that of modern man or to differ in the two presently recognized allomorphs of Australopithecus. Morphological dif- ferences between Australopithecus and modern man appear to be the result of different degrees of encephalization rather than any difference in locomotor adaptation.
Almost all who have discussed australo- pithecine locomotion have concluded that these extinct hominids were at least par- tially bipedal. They have not, however, considered them capable of a striding gait - a specialization usually considered to be restricted to Homo erectus and Homo sa- piens (Le Gros Clark, ’67; Napier, ’67). Napier has also stressed the importance of differences between the locomotor patterns of the “robust” and “gracile” forms (‘64). Although such conclusions are in agree- ment with those of a majority of other authors (Mednick, ‘55; Kern and Straus, ’49; Washburn, ’60; Straus, ’62; Chopra, ’62; Campbell, ’66; Day, ’69; Zhilman, ’69; Preuschoft, ’71), they are not in ac- cord with those of Dart (’49, ’57, ’58). In previous papers (Lovejoy and Heiple, ’70, ’72; Heiple and Lovejoy, ’71) it has been demonstrated that some of these earlier conclusions are based upon errors of ob- servation and a failure to take into account the normal range of variation of Homo sapiens.
In this paper we shall present a bio- mechanical analysis of australopithecine locomotion. Basing our arguments upon the morphological and biomechanical re- quirements of the striding gait of modern man, we believe that the antiquity of the
striding gait can be shown to be as great as the terminal Pliocene.
Since the argument to which we address ourselves in this paper is stride, we define it for this study as: the ability to (1) walk in an upright bipedal progression, succes- sively placing each foot closely under the center of gravity; (2) stabilize the pelvis against rotation by an abductor mecha- nism adequate to eliminate large trunk shifts; and, (3) utilize the principle of the compound pendulum for the lower extrem- ity so that the advance of each limb is ac- complished by gravity and short bursts of muscle contraction at accelerations and decelerations.
Such energy conservation is an essen- tial feature of man’s locomotor efficiency and can in large part be considered the sine qua non of stride.
While the available skeletal material of Australopithecus is not complete enough for the type of biomechanical analysis now possible for Homo sapiens, a sufficient amount of material has been recovered to
1 The definition of stride presented here is an opera- tional one for the purposes of this paper. It utilizes strictly anatomical criteria. “Stride” may also be de- fined on the basis of energy expenditure and gait effi- ciency. In this case the relative contributions of muscu- lar contraction and momentum to locomotion must be considered. This has been presented in Lovejoy (’72).
AM. J. PHYS. ANTHROP., 38: 757-780. 757

758 C. 0. LOVEJOY, K . G. HEIPLE AND A. H. BURSTEIN
allow reasonable reconstruction of those functions necessary for stride in bipeds.’ The present analysis will be carried out via the following headings:
1 . The Ilium 2. Femoral Neck Length 3. Femoral Head Size and Hip Joint Pres-
sure 4. Hip Flexion 5. Hip Rotation during Stride 6. The Foot in Stance Phase 7. Hip Extension during Stride 8. The Ischium
T h e il ium
As has been pointed out by numerous authors, the conformation of the australo- pithecine ilium is strikingly hominid. The iliac crest, rather than being short and relatively straight as in the quadruped, is long and shows the characteristic sig- moid curve of H. sapiens in all samples thus far recovered (SK-50; MLD-7; MLD-
Le Gros Clark, ’55, ’67). The ilium is very broad and thus provides an extensive area of origin for the abductors. Dart pointed out that “the area of the lateral or gluteal aspect [of MLD-251 . . . was actually about 10% greater than that of a male juvenile Bush ilium of the same age” (‘57: 75). While there is general agreement with respect to the hominid configuration of the australopithecine ilium, several fea- tures by which it differs from the modern ilium have been cited as evidence of in- efficient bipedalism. These include the lat- eral iliac flare, the supposed absence of the “iliac pillar” (Mednick, ’55) and tu- berosity of the ilium (“cristal tubercle”) and the prolongation of the anterior su- perior spine. We believe, to the contrary, that these features are adaptively inter- related and indicative of an advanced type of bipedalism including the ability to stride as defined above.
25; TM-1605; STS-14; STS-65) (Dart, ’49;
Lateral iliac flare
The distinct lateral flare of the australo- pithecine ilium has been a consistently noted feature of both the gracile and ro- bust forms (Dart, ’49, ’58; Napier, ’64; Le Gros Clark, ’67; Zhilman, ’69; Chopra, ’62). Such lateral flaring places the origins of the gluteus medius and gluteus mini- mus more distant from the acetabulum in
Australopithecus than in modern man since it is produced to a great extent by a greater lateral angulation of the iliac blade relative to the sagittal plane of the body. Put in other terms, the iliac blades of H. sapiens are more nearly vertical and thus the distance from the acetabulum to the crest of the ilium along the horizontal plane is shorter in modern man (cf. fig. 5).
The iliac pillar While there is general agreement with
regard to lateral iliac flare, it has also been asserted by some authors (but not all: see Dart, ’57, ’58; Day, ’59; Zhilman, ’69) that no iliac pillar is present in the austra- lopithecine ilia. However, cross sections of casts of the available innominates dem- onstrate that a significant iliac thickening is clearly present in all (fig. 1). This has been graphically represented in figure 2. The slightly anterior position of the pillar in Australopithecus may have been respon- sible for its frequent oversight. Because the ilium of Australopithecus is more lat- erally flared than it usually is in modern man, a more anterior position of the pillar is to be expected in their ilia. It indicates that the compression forces of the abduc- tors were developed in the same manner as in H. sapiens. Put in other terms, if the position of the ilia in modern man were to be altered by more pronounced lateral flar- ing, such action would move the functional position of the abductors posteriorward (cf. fig. 3). Were the position of the iliac pillar in Australopithecus the same as it is in modern man, the functional position of the line of action of the abductors would have been more posterior to the hip joint in this biped. However,. the anterior posi- tion of the pillar indicates that the abduc- tor’s line of action in this early hominid was homologous to that of modern man.
2 The materials used for the preparation of this paper are as follows: SK-50, Wenner-Gren cast F-SK17; SK-82, Wenner-Gren cast F-SK25; Penn. Mus. cast SK-82; SK- 97, Penn. Mus. cast SK-83; STS-34, Wenner-Gren cast F-ST10; TM-1513, Wenner-Gren cast F-ST19; STS-14, (right innominate), Wenner-Gren cast F-ST6; MLD-8, Penn. Mus. cast 707; MLD-7, Penn. Mus. cast 708; MLD-25, Penn. Mus. cast 750; Bushman Innominate, Wenner-Gren cast F-CP18; OH-20 Cast supplied to the authors by Dr. Michael Day; STS-65, Scaled photographs supplied to the authors by Dr. Loring Brace.
All measurements used in this paper of SK-50, SK-82, SK-97, STS-14 (innominate, femur, and reconstructed pelvis), STS-65 and MLD-17 were made on the original specimens either by Dr. Loring Brace or by Dr. Milford Wolpoff.

GAIT OF A U S T R A L O P I T H E C U S 759
H.S. 1
HA 2
H.s.3
I
H.s.4
S K 5 0 - TM 1605
STS 14 I
MLD25
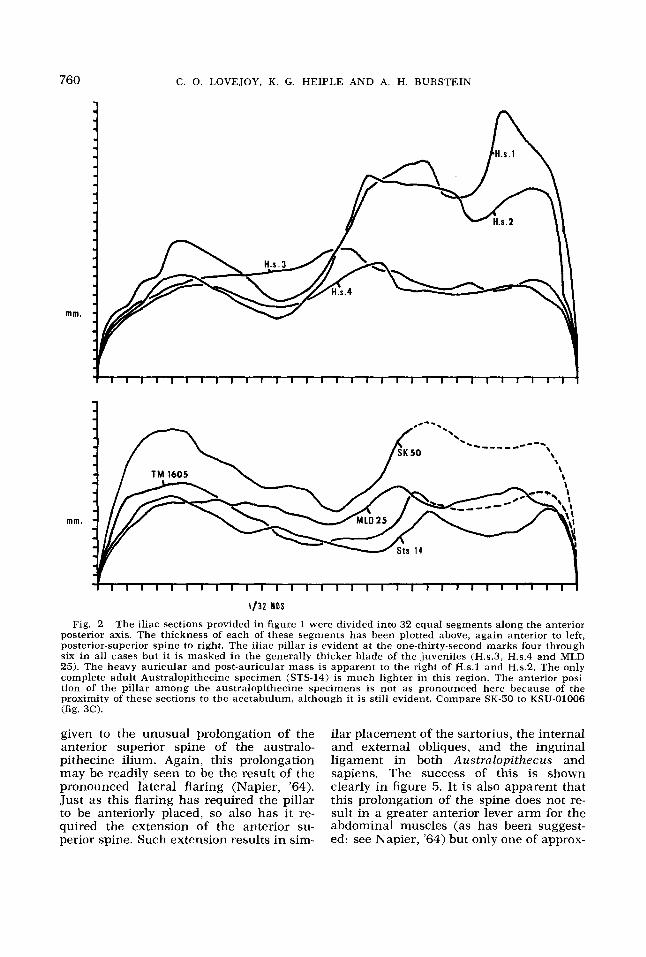
Fig. 1 The Iliac Pillar. These iliac cross sec- tions were taken from negative casts through a line from the posterior superior spine to the deep- est point in the notch below the anterior superior spine (cf. line X-Y in fig. 10). Anterior is to the left and lateral is to the top. The double marks indi- cate the position of the anterior-most and poste- rior-most extent of the auricular surface. H o m o sapiens I is a large Amerindian female. H o m o sapiens 2 is a small Amerindian female. H o m o sap- iens 3 is an Amerindian child of about five years. H o m o sapiens 4 is a Bushman adolescent (Wen- ner-Gren cast). The posterior portions of SK-50 and TM-1605 (Wenner-Gren Casts) have required extensive reconstruction (cross hatched) and have been included only to indicate the relative posi- tion of the pillar in each. The thickest point at the far left is the iliac pillar in each case. H.s. 3 and MLD 25 (Pennsylvania Museum Cast), the juve- niles, have no discernible pillar, but show a dis- tinctive thickness in the region of the pillar.
It should also be pointed out that Dart, during study of the MLD-7 ilium, was able to observe the internal structure of the bone in the region of the pillar. After com- parison with a similarly sectioned Bush- man ilium he concluded that “the princi- pal difference is seen to lie in the greater thickness of the compact bone both inter- nally and externally in the australopithe-
cine ilium” (’57: 77). He also reported that the natural split-lines formed in MLD- 7 and MLD-25 are almost identical to those found by Mednick (‘55) in the ilia of modern man, and Tappen (’69) has shown such weathering cracks to be fully homologous to split lines induced in mod- ern specimens.
T h e cristal tubercle
In addition to the general negation of an iliac pillar by some authors, the “cristal tubercle” (the thickening at the pillar’s iliac margin) has also been described as absent in the australopithecine pelvis. It is difficult to determine the evidence for this negative ascription since the critical portions of the iliac crest are missing in the two adult samples for which casts are available (STS-14 - right side; SK-50). The two adolescent ilia, MLD-7 and MLD- 25, both have thickening in this area but are from individuals below the.age of fu- sion of the iliac apophysis which was not recovered for either sample. As the tuber- cle is formed from this apophysis and is much less prominent in modern individ- uals of equivalent osteological age, these are therefore unreliable samples with re- gard to this morphological feature. The only remaining sample which could at present provide evidence with regard to the cristal tubercle is the left iliac fragment of STS-14. This specimen is also incom- plete but does provide some evidence. There appear to have been two thickened regions on the crest of this specimen, separated by a thinner space. The ilium is thickened in the region of the pillar (ar- row, fig. 4) but unfortunately the crest is “broken away just where the pillar meets it” (Wolpoff, personal communication). Posterior to this the ilium first thins but then a second thickening of a shelf-like type occurs which is perhaps related to the insertion of the external and/or internal oblique. In any case, the clearly demon- strable pillar (or a thickening which adum- brates it in immature specimens) is ample evidence of considerable abductor power in Australopithecus, and is consistently present in all available samples.
The anterior superior spine
Considerable attention has also been
760 C. 0. LOVEJOY, K . G. HEIPLE AND A. H. BURSTEIN
mm.
mm.
\
\
1 /32 NOS
Fig. 2 The iliac sections provided in figure 1 were divided into 32 equal segments along the anterior posterior axis. The thickness of each of these segments has been plotted above, again anterior to left, posterior-superior spine to right. The iliac pillar is evident at the one-thirty-second marks four through six in all cases but it is masked in the generally thicker blade of the juveniles (H.s.3, H.s.4 and MLD 25). The heavy auricular and post-auricular mass is apparent to the right of H.s.1 and H.s.2. The only complete adult Australopithecine specimen (STS-14) is much lighter in this region. The anterior posi- tion of the pillar among the australopithecine specimens is not as pronounced here because of the proximity of these sections to the acetabulum, although it is still evident. Compare SK-50 to KSU-01006 (fig. 3 0 .
given to the unusual prolongation of the anterior superior spine of the australo- pithecine ilium. Again, this prolongation may be readily seen to be the result of the pronounced lateral flaring (Napier, '64). Just as this flaring has required the pillar to be anteriorly placed, so also has it re- quired the extension of the anterior su- perior spine. Such extension results in sim-
ilar placement of the sartorius, the internal and external obliques, and the inguinal ligament in both Australopithecus and sapiens. The success of this is shown clearly in figure 5. It is also apparent that this prolongation of the spine does not re- sult in a greater anterior lever arm for the abdominal muscles (as has been suggest- ed: see Napier, '64) but only one of approx-
GAIT OF AUSTRALOPITHECUS 76 1
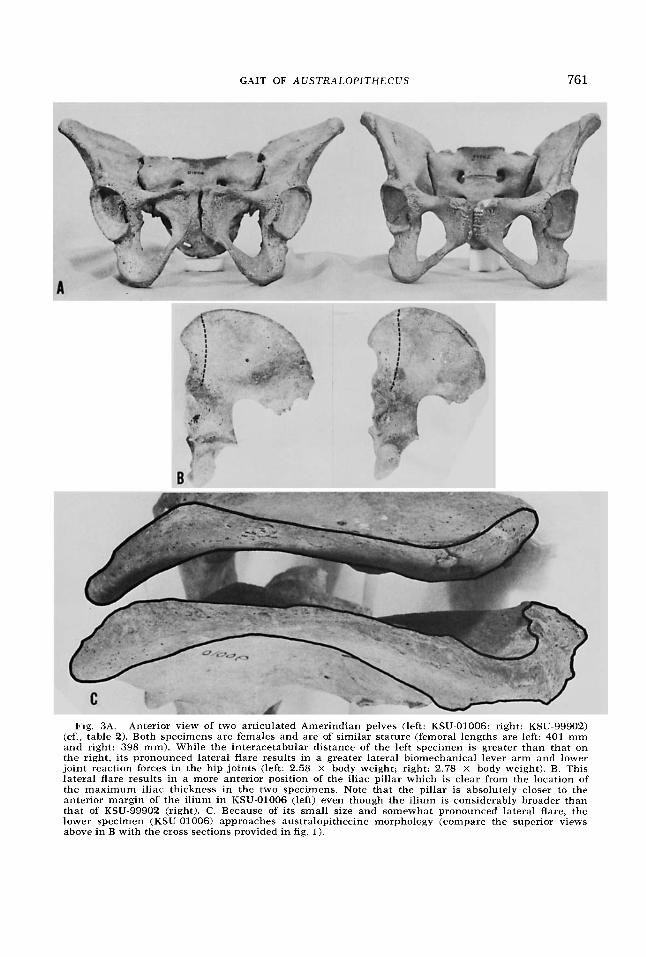
Fig. 3A. Anterior view of two articulated Amerindian pelves (left: KSU-01006; right: KSU-99902) (cf., table 2) . Both specimens are females and are of similar stature (femoral lengths are left: 401 mm and right: 398 mm). While the interacetabular distance of the left specimen is greater than that on the right, its pronounced lateral flare results in a greater lateral biomechanical lever arm and lower joint reaction forces in the hip joints (left: 2.58 X body weight; right: 2.78 X body weight). B. This lateral flare results in a more anterior position of the iliac pillar which is clear from the location of the maximum iliac thickness in the two specimens. Note that the pillar is absolutely closer to the anterior margin of the ilium in KSU-01006 (left) even though the ilium is considerably broader than that of KSU-99902 (right). C. Because of its small size and somewhat pronounced lateral flare, the lower specimen (KSU-01006) approaches australopithecine morpholcgy (compare the superior views above in B with the cross sections provided in fig. 1).
762 C. 0. LOVEJOY, K. G. HEIPLE AND A. H. BURSTEIN
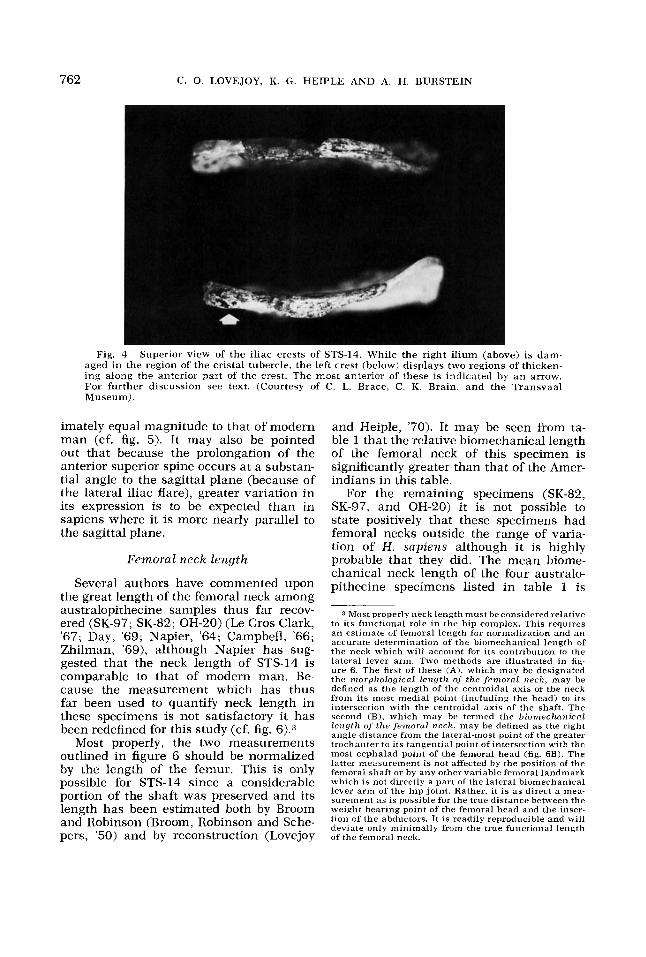
Fig. 4 Superior view of the iliac crests of STS-14. While the right ilium (above) is dam- aged i n the region of the cristal tubercle, the left crest (below) displays two regions of thicken- ing along the anterior part of the crest. The most anterior of these is indicated by an arrow. For further discussion see text. (Courtesy of C. L. Brace, C. K. Brain, and the Transvaal Museum).
imately equal magnitude to that of modern man (cf. fig. 5). It may also be pointed out that because the prolongation of the anterior superior spine occurs at a substan- tial angle to the sagittal plane (because of the lateral iliac flare), greater variation in its expression is to be expected than in sapiens where it is more nearly parallel to the sagittal plane.
Femoral neck length
Several authors have commented upon the great length of the femoral neck among australopithecine samples thus far recov- ered (SK-97; SK-82; OH-20) (Le Gros Clark, '67; Day, '69; Napier, '64; Campbell, '66; Zhilman, '69), although Napier has sug- gested that the neck length of STS-14 is comparable to that of modern man. Be- cause the measurement which has thus far been used to quantify neck length in these specimens is not satisfactory it has been redefined for this study (cf. fig. 6).3
Most properly, the two measurements outlined in figure 6 should be normalized by the length of the femur. This is only possible for STS-14 since a considerable portion of the shaft was preserved and its length has been estimated both by Broom and Robinson (Broom, Robinson and Sche- pers, '50) and by reconstruction (Lovejoy
and Heiple, '70). It may be seen from ta- ble 1 that the relative biomechanical length of the femoral neck of this specimen is significantly greater than that of the Amer- indians in this table.
For the remaining specimens (SK-82, SK-97, and OH-20) it is not possible to state positively that these specimens had femoral necks outside the range of varia- tion of H . sapiens although it is highly probable that they did. The mean biome- chanical neck length of the four australo- pithecine specimens listed in table 1 is
3 Most properly neck length must be considered relative to its functional role i n the hip complex. This requires a n estimate of femoral length for normalization and a n accurate determination of' the biomechanical length of the neck which will account for its contribution to the lateral lever arm. Two methods are illustrated in fig- ure 6. The first of these (A), which may be designated the morphological length of the femoral neck, may be defined as the length of the centroidal axis of the neck from its most medial point (including the head) to its intersection with the centroidal axis of the shaft. The second (B), which may be termed the biomechanicril length of the femorul neck, may be defined as the right angle distance from the lateral-most point of the greater trochanter to its tangential point of intersection with the most cephalad point of the femoral head (fig. 6B). The latter measurement is not affected by the position of the femoral shaft or by any other variable femoral landmark which is not directly a part of the lateral biomechanical lever arm of the hip joint. Rather, it is as direct a mea- surement as is possible for the true distance between the weight bearing point of the femoral head and the inser- tion of the abductors. It is readily reproducible and will deviate only minimally from the true functional length of the femoral neck.

GAIT OF AUSTRALOPITHECUS 763
Fig. 5 Superior view of the left ilium of an im- mature australopithecine (MLD-7, shaded) articu- lated with an adolescent Bushman pelvis. The symmetrical coordinate lines are inserted to clarify the relative positions of the anterior superior spines (solid arrows) on both ilia. The greater lateral flare of the iliac crest and the prominent ilio-psoas groove of MLD-7 (open arrow) are clear. (Drawn from a photograph in Dart, '49).
65.5 mm. The average circumference of their femoral shafts (taken immediately below the lesser trochanter but not in- cluding any portion of it) is 84.4. Thus in these specimens the biomechanical neck length is 78% of the shaft circumference. For the Amerindians also listed in table 1, biomechanical neck length is only 73% of shaft circumference. Femoral circumfer- ence is a highly variable parameter, how- ever, and will vary not only with the length of the femur but with its general robustic- ity as well. It is important to note there- fore, that in the one specimen in which the relative biomechanical neck length can be calculated (because a sufficient portion of the shaft and a complete pelvis were recovered with it allowing reasonable esti- mation of its overall length), that of STS- 14, the neck can be shown to be signifi- cantly longer than that of H . sapiens (cf. table 1). Statistical demonstration of long- er neck lengths in other specimens must await more complete characterization of their length either by recovery of more complete femora, or recovery of sufficient portions to allow reasonable reconstruc- tion of their lengths.
An additional difference that has been noted between the proximal femora of Australopithecus and those of modern man is the lack of lateral flare of the greater trochanter in the former (Napier, '64;
Day, '69) and this appears to be a consis- tent feature. While the femoral neck usual- ly is loaded only in compression during gait, it is the site of frequent torsional strain and occasional traumatic fracture. Selection may therefore have acted to re-
t-
Fig. 6 Two reproducible measurements of the femoral neck. Distance A (morphological length of the femoral neck) is measured from the most me- dial extension of the centroidal axis of the neck (in- cluding the head) to the centroidal axis of the femoral shaft and is thus a measure of the length of the centroidal axis of the neck. It is most accu- rately obtained from sharp-shadow tracings (Love- joy and Heiple, '70). Distance B (biomechanical length of the femoral neck) is taken perpendicu- lar to the centroidal axis of the shaft. It is the length of a line from the lateral-most point on the greater trochanter to its tangential point of inter- section with the most cephalad point on the fem- oral head. This measurement is most accurately determined directly from the femur. Previous meth- ods used for determining femoral neck length re- quire a subjective judgement of the origin of the base of the neck from the trochanteric mass.

764 C. 0. LOVEJOY, K. G. HEIPLE AND A. H. BURSTEIN
TABLE 1
Metric prirtrmeters of Aiistrnlopithrciizr trnd Amerindinn femora (All mecisirrements in millimeter5 except where otherzurse noted)
Relative hio- mechanical neck length (hio. L/max. fem. length)
Relative mor- phological
neck length (morph. L/ max. fem.
length)
Circu mfer- ence of fem-
oral shaft be- low the lesser
trochanter
Neck-shaft (collo-di-
aphyseal) an- gle
Biomechani- cal length of femoral neck
(hio. L.)
64 67 78 3
53
Morphological length of fem-
oral neck (mor. L.)
61 66 75 3
51
Specimen Maximum femoral length
SK-82 1
OH-20 2 SK-97 1
STS-14 1
Am-eerindian
Amerindian X ( N = 40)
S.D.
- - -
0.189 0.171
- - -
0.182 0.165
1190 115' 117'
118'
89 93 87
68
-
280 4
310 5
454
22
67.7
6.9
68.5
4.7
0.151
0.007
0.149
0.012
128"
3.6'
93.3
5.4
1 Measurements of SK-82, SK-97, and STS-14 made on the original specimens by M. H. Wolpoff (personal communication). 2 Measurements of OH-20 made on a cast supplied to the authors by M. H. Day. 8 The femoral head of this specimen was not preserved. This is an estimated minim~m value, assuming that the head be-
4 Estimate from Lovejoy and Heiple ('70). 5 Estimate by Broom and Robinson (Broom et al., '50).
gan immediately at the point of breakage, and that it was the same size as that of SK-97.
TABLE 2
Morphologirctl dcrtci mid cctlcztlntiorzs o n the pelvis tzndf'emzrr of STS-I 4 a n d 20 A m t ~ i . i z ~ t l i < i , i s
(J.R.F.) (fem. Inter-fern. Max. di- length13 head dis.
Catalog oral head Abductor reaction femoral femoral (fem. head length of fem- max. fem. number distance P angle force 1 length head Sex diam)2 oral neck
Case Skeleton Inter-fem- Joint Maximum ameter Biomechanical
length
STS 14 mm mm 132 36 15" 2.46 152 51 16" 2.26 140 47 15" 2.26 156 52 13" 2.28 160 52 13' 2.32 160 52 12O 2.32 156 50 14" 2.33 172 53 14' 2.39 156 47 18' 2.42 164 50 9' 2.42 158 48 11" 2.42 158 46 13' 2.49 176 51 1l0 2.50 166 47 15" 2.53 164 46 12O 2.55 160 44 15' 2.58 170 45 14" 2.66 180 47 8' 2.69 172 43 15" 2.76 154 38 15' 2.78 180 41 9' 2.96 163 48 13" 2.50 9.8 3.9 2.5' .19
mm 280 452 445 436 467 496 44 5 468 403 453 42 7 408 490 493 467 401 457 462
mm 31.2 48 44 46 45 48 47 48 40 46 41 40 49 49 45 42 41 48 45
55.5 91
103 89
116 123 93
107 99
106 112 106 123 126 128 94
151 115 121 128 159 112.5 18.7
mm 53 73 67
2.49 2.08 2.09 2.36 2.13
~ _ _ ~~
1 KSU03257 2 KSU03237 3 KSU10017
M M M 66
75 70 73 73 61 70 63 63 70 75 68 56 64 72 66
4 KSU02219 5 KSU02012 6 KSU02247
M M 2.29
2.15 2.37
M F F M
7 KSU0416F 8 KSU08206 9 KSU06071
2.56 2.34 2.45 2.51 2.50 2.21 2.43 2.86 2.64 2.50
10 KSU04043 11 KSU05209 12 KSU05206
F F M F F F
13 KSU03307 14 KSU01231 15 KSU01006 16 KSU0423F 17 KSU0425E 18 KSU00235 19 KSU99902 20 KSU01230 x S.D.
F F F 446
398 477 450
~~
37 46 45
F F
.~
58 69 68 2.42
.21 29 3.4 5.3
1 In units of body weight.
duce its length during the Pleistocene, adaptive change would have been a medi- especially if body size were increasing. alward shift in the position of the femo- However, reduction of femoral neck length ral shaft. In this way the neck could be would have substantially increased femo- reduced in apparent length without con- ral head pressures (see below). A more current reduction of the abductor lever
GAIT OF A U S T R A L O P I T H E C U S 765
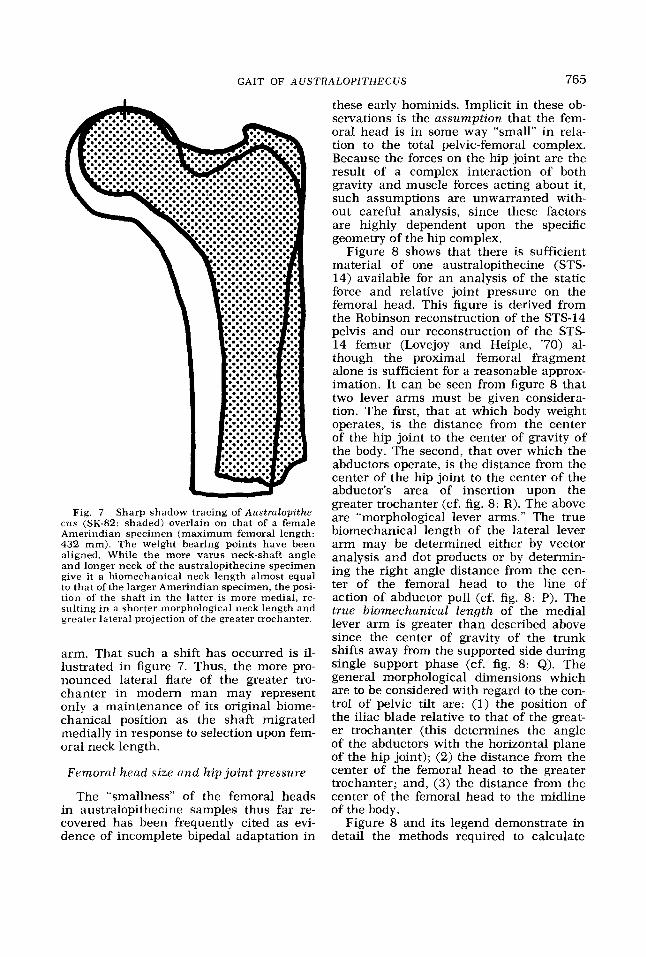
Fig. 7 Sharp shadow tracing of Aus t ra lop i the - cus (SK-82: shaded) overlain on that of a female Amerindian specimen (maximum femoral length: 432 mm). The weight bearing points have been aligned. While the more varus neck-shaft angle and longer neck of the australopithecine specimen give it a biomechanical neck length almost equal to that of the larger Amerindian specimen, the posi- tion of the shaft i n the latter is more medial, re- sulting in a shorter morphological neck length and greater lateral projection of the greater trochanter.
arm. That such a shift has occurred is il- lustrated in figure 7. Thus, the more pro- nounced lateral flare of the greater tro- chanter in modern man may represent only a maintenance of its original biome- chanical position as the shaft migrated medially in response to selection upon fem- oral neck length.
Femoral head size and h ip joint pressure
The “smallness” of the femoral heads in australopithecine samples thus far re- covered has been frequently cited as evi- dence of incomplete bipedal adaptation in
these early hominids. Implicit in these ob- servations is the assumpt ion that the fem- oral head is in some way “small” in rela- tion to the total pelvic-femoral complex. Because the forces on the hip joint are the result of a complex interaction of both gravity and muscle forces acting about it, such assumptions are unwarranted with- out careful analysis, since these factors are highly dependent upon the specific geometry of the hip complex.
Figure 8 shows that there is sufficient material of one australopithecine (STS- 14) available for an analysis of the static force and relative joint pressure on the femoral head. This figure is derived from the Robinson reconstruction of the STS-14 pelvis and our reconstruction of the STS- 14 femur (Lovejoy and Heiple, ’70) al- though the proximal femoral fragment alone is sufficient for a reasonable approx- imation. It can be seen from figure 8 that two lever arms must be given considera- tion. The first, that at which body weight operates, is the distance from the center of the hip joint to the center of gravity of the body. The second, that over which the abductors operate, is the distance from the center of the hip joint to the center of the abductor’s area of insertion upon the greater trochanter (cf. fig. 8: R). The above are “morphological lever arms.” The true biomechanical length of the lateral lever arm may be determined either by vector analysis and dot products or by determin- ing the right angle distance from the cen- ter of the femoral head to the line of action of abductor pull (cf. fig. 8: P). The true biomechanical l eng th of the medial lever arm is greater than described above since the center of gravity of the trunk shifts away from the supported side during single support phase (cf. fig. 8: Q). The general morphological dimensions which are to be considered with regard to the con- trol of pelvic tilt are: (1) the position of the iliac blade relative to that of the great- er trochanter (this determines the angle of the abductors with the horizontal plane of the hip joint); (2) the distance from the center of the femoral head to the greater trochanter; and, (3) the distance from the center of the femoral head to the midline of the body.
Figure 8 and its legend demonstrate in detail the methods required to calculate

766
5
Body a
C. 0. LOVEJOY, K. G. HEIPLE AND A. H. BURSTEIN
Figure 8

76 7
both the relative joint reaction force and the relative femoral head pressure €or STS-14. Calculations made by identical methods upon 20 Amerindian skeletons
GAIT OF A U S T R A L O P I T H E C U S
-
yield a mean of 2.50 times body weight for joint reaction force while our calcula- tion for STS-14 is 2.46 times body weight. However, when the pressure upon the fem-
Fig. 8 A frontal projection of the STS-14 pelvis (restored by Robinson) and our reconstruction of the STS-14 femur in mid-stance phase.‘: At this instant the pelvis is level and the foot would lie beneath the midline. The approximate extent of the abductors is indicated and their centroid is projected cephalad (Line A). The direction of the abductor muscle force is indicated by the arrow on Line A. Line D is the midline of the body. Line C, a vertical through the center of gravity, is displaced slightly to the opposite side because the weight of the entire right lower extremity (one-sixth body weight) must be eliminated when the remain- der of the body is considered as a free body (Frankel and Burstein, ’70). The direction of this weight force is also indicated by an arrow. The line of action of the joint reaction force (Line B: the negative of the sum of the abductor and body weight vectors) can be drawn from the center of the femoral head to the intersection of Line A and Line C with direction as indicated. That Line B must pass through the point of intersection of Lines A and C is a requirement of moment equilib- rium.
Since the direction of all three forces acting about the hip joint on this free body in equilibrium and the magnitude of one of them (five-sixths body weight) are known a vector diagram can be con- structed. From this we can determine the static load (in multiples of body weight) by which the femoral head is compressing the acetabulum at this instant of stance phase. Such a vector diagram has been constructed to the left of the primary figure. A weight vector (“5/6 X body weight”) has been drawn an arbitrary length of 14.1 cm. The joint reaction force vector produced by adding the muscle force vector and constructing the force equilibrium triangle is 41.6 cm. By calculation:
Joint Reaction Force = 41‘6 = (6/5) (14.1)
2.46 X Body Weight (1)
This would be the relative joint load on the fem- oral head of STS-14 during stationary weight bearing on one foot. Pauwels (’35) has made sim- ilar calculations for H o m o sap iens and concluded that the value of this force for modern man was 2.92 X body weight, while Frankel and Burstein (‘70) from similar calculations obtained a value of 2.75 X body weight for modern man. It was neces- sary, however, to recalculate this joint loading for modern man in a fashion similar to that used for STS-14. The force triangle is subject to trigonomet- ric solution. We have used this method to obtain the figures provided for 20 Amerindian specimens i n Table 2. The joint loading of modern man was found to be equal to that of STS-14 (z of 20 H o m o supiens = 2.50). Thus it would appear that the shifts in the geometry of the STS-14 hip and pel-
vis (longer femoral neck, lower neck-shaft angle, laterally flared ilium, relatively greater interace- tabular distance) all combine to produce a leverage system that results in total forces on the femoral head within the range of H o m o snp iens .
However, before we can compare pressure on the femoral heads of STS-14 and modern man, the total force must be related to body size and fem- oral head diameter. The mean femoral head diam- eter of the 20 Amerindian specimens is 45 mm and their mean femoral length is 450 mm. Our reconstruction of the STS-14 femur (Lovejoy and Heiple, ’70) yielded a length of about 28 cm. The reconstructed head (based upon the size of the acetabulum of STS-14 is 31.2 mm. (table 2). Since stature and weight are more closely correlated with femoral length than with any other long bone dimension, we have used these lengths to obtain the relative femoral head pressures of STS-14 and the 20 Amerindians by the following calculations:
Total body weight = (femoral length13 X K
For H . scipie?is (%) = (45)3 K (constant) (2)
= 91.125 K ~~
For STS-14 = (28P K = 21,952 K
Total load on femoral head = relative joint load X body weight (3)
For H . sczpicvzs (x) = 2.50 X 91,125 K = 227,812 K
For STS-14 = 2.46 X 21,952 K = 54,002 K
Relative femoral head area = (femoral head
For H . snpiens (x) = (45Yk (constant) diameter)* (4)
= 2,025 k For STS-14 = (31.2)Zk
= 973k Relative pressure on femoral head =
total load (3) relative area (4)
(5)
For H . snpiens (x) = 227’812 2,025 k
= 112.5 K/k
54,002 K = 55.5 K/k For STS-14 = 973 k
Ratio of femoral head pressure in H . snp iens and STS-14: (6)
112.5 ___- K’k - 2.03 (constants cancel) 55.5 K/k
Thus when the joint loads of STS-14 and the mod- ern specimens are normalized for the total body weights of each it can be seen that the relative pressure on the articular cartilage of the hip joint of H . snp iens is twice that of STS-14. Conversely, the hip joint of STS-14 carried only 50% of the static pressure carried by the Amerindian hip joints used in this sample.
* The reconstruction which we presented (Lovejoy and Heiple, ’70) has been criticized because our measure- ments of the pelvis were determined from scaled photographs in published sources. Measurements on the original specimen (STS-14) recently supplied to us by Dr. Mllford Wolpoffdid not significantly alter our results.

768 C. 0. LOVEJOY, K. G . HEIPLE AND A. H . BURSTEIN
oral head is calculated the magnitude for STS-14 is only half that of the Amerin- dians; i.e., the articular cartilage of the femoral head of STS-14 would have suf- fered pressures only 50% those of mod- ern man.
The joint reaction forces of the 20 Amerindians (table 2) show a significant distribution with regard to some of those factors which we discussed above in rela- tion to Australopithecus. Perhaps the most interesting is the substantial difference between hip joint reaction forces in males and females. These were found to differ significantly a t the P = 0.001 level by use of the t-test. Inspection of the data in ta- ble 2 shows that the causes of this differ- ence are complex, although some general trends can be recognized. The significant- ly greater hip joint reaction force in fe- males can be related to the relatively greater inter-acetabular distance and gen- erally shorter stature in this group. As the breadth of the lesser pelvis must of neces- sity be great in females, the result is a greater abductor force requirement for the prevention of pelvic tilt. These factors re- sult in greater joint reaction forces. Inter- acetabular distance alone, however, is insufficient to predict accurately the re- sultant joint reaction force. It is necessary also to consider the extent of lateral flaring of the ilia and the length of the femoral neck, both of which tend to reduce pres- sure as each is increased. The principal effects of these features can be generally illustrated and are shown in figure 3 and described in its legend.
In Australopithecus, we find that the joint reaction force in STS-14 is lower (2.46 X body weight) than the mean of the Amerindian females (2.60 X body weight). This is the case despite the rel- atively pronounced interacetabular dis- tance of STS-14 (Lovejoy and Heiple, '70). It may be noted from table 2 that the dis- tance between the centers of the femoral heads in STS-14 is 132 mm, while the fem- oral length of this specimen is only about 28 cm. (Lovejoy and Heiple, '70). A simple ratio would predict for a sapiens female (for example the smallest individual in ta- ble 2: Case 19, KSU99902: femoral length of 398 mm) a distance between the centers of the femoral heads of 188 mm, rather
than its actual value of 154 mm. In fact, the projected value (188 mm) is greater than any inter-femoral head distance among the sapiens pelves, even the largest (table 2). Yet even with its relatively pro- nounced medial lever arm, STS-14 has maintained a joint reaction force substan- tially smaller than the average Amerindian female. This has been possible only through the relatively greater femoral neck length and laterally flared ilium which together offset the elongated me- dial lever arm. It is interesting in this re- spect that McLeish and Charnley ('70), in studying living individuals by methods similar to those which have been applied to STS-14 and the Amerindians, found ab- ductor force and femoral head pressure to increase substantially with shortening of the femoral neck.
The problem of femoral head size in the australopithecine femurs without ac- companying pelves must be approached in a somewhat different fashion. Assuming their femoral head pressures to be similar to those of the Amerindians in table 2, we may bracket possible femoral lengths com- mensurate with the head diameter in each australopithecine femur. This can be done in the following manner. The relative in- tensity of hip joint pressure (cf. fig. 8) was calculated for each of the 20 Amerindians in table 2. These calculations yield a mean of 112.5 with a standard deviation of 18.7.4 If we set an upper limit of joint loading at two standard deviations above the mean and a lower limit at two stan- dard deviations below the mean we may then use the known australopithecine head diameters and these relative load factors to calculate a range of possible lengths for the femora from which each comes. Using the Amerindian hip joint reaction force 2.50 and rewriting equations 3, 4, and 5 from the legend of figure 8 we ob- tain:
J(112.5 3 [18,7] 1 (head diameter)* - - 2.50
femoral length
which yields the following projected lengths :
4 There are no units to the relative femoral head pres- sure figures since these are only relative, i.e., a ratio de- pendent upon both the femoral head size and multiples of body weight.

GAIT OF AUSTRALOPITHECUS 769
Range of Sample Head diameter projected lengths
mm cm SK-82 34 31 to 39 SK-97 37 33 to 42 MLD-17 37.5 33 to 42
Thus, if the femur for each specified head diameter fell within the calculated project- ed lengths, pressures on the femoral heads of these specimens would have been within the range of modern man. These projected lengths are reasonable in light of the gen- eral morphology of the intact proximal femoral fragments, but we have assumed the joint reaction force in these australo- pithecines to be the same as that of the Amerindian mean. There is reason to be- lieve that this is not the case and that the joint reaction force in larger australo- pithecines was less than that of modern man. The two critical dimensions deter- mining joint reaction force are the relative lengths of the lateral and medial lever arms. These may be expressed as a simple ratio:
Inter-femoral head distance Biomechanical fem. neck length
= R
It may be noted (cf. fig. 8) that the lower the value of R, the greater the advantage of the abductors in controlling pelvic tilt and the lower the joint reaction force in stance phase. This ratio for the 20 Amer- indians in table 2 yields a mean R of 2.42. If we use this ratio to estimate inter- femoral head distance for each australo- pithecine specimen for which a neck length is known, the following data are obtained:
Projected inter- Specimen Biomechanical femoral head
neck length distance
mm mm SK-82 64 155 SK-97 67 162 OH-20 78 189
The mean projected inter-femoral head distance calculated by the above ratio (R) for Austrdopithecus is thus 169 mm while the mean of the sapiens in table 2 is only 163 mm. We believe that these projected distances are therefore too great since the adult cranial capacity of this early horninid (about one third that of modern man) indicates that the cranial di-
mensions of the full term fetus would have also been significantly less than in modern man and the requirements for the dimensions of the parturition canal would have therefore also been substantially less than in H . sapiens. It is therefore likely that the joint reaction forces in these australopithecines were substantially low- er than the Amerindian mean (cf. table 2).
H i p ftexion
In modern man the swing phase of gait is initiated by the contraction of two pow- erful flexors of the hip, the rectus femoris and the psoas major. Both leave promi- nent evidence of their action upon the pel- vis. The straight head of the rectus takes origin from the anterior inferior iliac spine. This same landmark, absent in the quad- ruped, is also the site of origin of the ilio- femoral ligament, a structure which pre- vents overextension of the trunk (by passive tension) when i t is held erect as in modern man. In the australopithecine innominates this spine is prominent and is as well developed as it is in H . sapiens. Although the femoral region of insertion of the ilio-femoral ligament has been claimed to be less well developed among australopithecine samples than in modern man, this has been recently shown not to be the case (Lovejoy and Heiple, '72).
A well defined ilio-psoas groove is pres- ent in both the robust and gracile pelvic samples (fig. 5) (LeGros Clark, '67). This not only serves as an indication that loco- motor posture was consistently erect in these bipeds but also indicates the promi- nent role of the ilio-psoas in their gait cycle.
Considerable attention has been paid to the position of the lesser trochanter on the femoral samples of both the robust and gracile australopithecines. Many authors have claimed that its position in these early hominid femora is more lateral than its position in modern femora (Broom and Robinson, in Broom et al., '50; Day, '69; Napier, '64; LeGros Clark, '67). We have recently shown, however, that the position of the lesser trochanter in the australo- pithecine samples falls well within one standard deviation of the mean of its posi-

7 70 C. 0. LOVEJOY, K. G. HEIPLE AND A. H. BURSTEIN
tion in Amerindians when it is metrically assessed (Lovejoy and Heiple, '72).
When all evidence has been considered there are thus no reasonable grounds for the suggestion that the psoas major, rectus femoris, or ilio-femoral ligament were not as well developed in Australopithecus as they are in modern man, nor is there any reason to believe that their locomotor func- tions were not homologous or identical to their respective roles in the gait cycle of H . sapiens.
Hip rotation during stride
Two important elements of the human stride complex are the position of the foot in stance phase and the rotation of the pelvis in the horizontal plane (about the hip joint which is in stance phase). The latter may also be referred to as internal rotation of the supporting limb. Zhilman has recently concluded:
. . . The rotation component was lost when the quadrupedal pelvis became upright and had not yet been reacquired in Azrstrtrlo- pithecus.
Regardless of how these muscles are re- constructed [gluteus inedius and gluteus minimus], i t is apparent from the lateral position of the anterior superior iliac spine that they could have been only weak inter- nal rotators at best. . . . Because of this inef- fective rotation mechanism, it i s probable that the arms were used more in locomo- tion to aid in balance and in transferring weight on to the advancing limb. . . , ('69: 81).
While it is true that the lateral iliac flare in Australopithecus would have also placed the abductors in a more lateral functional position (see above), their ability to in- ternally rotate would have been unaffected because commensurate anatomical shifts were already present in other parts of the hip, This is clear from the previous dis- cussions of hip abduction and flexion. The longer femoral neck carries the in- sertion of the abductors to a more lateral position, restoring the internal rotative power of the minimus and medius.
In addition, the femora of Australopith- ecus almost certainly displayed some an- teversion. Torsion of the femoral shaft al- ters the position of the greater trochanter - the greater the anteversion, the more posterior the placement of the greater trochanter in functional anatomical posi-
tion (assuming the axis of the knee joint is maintained in the coronal plane). Ante- version thus increases the internal rota- tive power of the gluteus minimus and medius.
A second point can be made with respect to the above discussion. Since anteversion is so highly variable in H. sapiens, varying from - 5" to 40 + O , and since it does have a significant effect upon the functional position of the greater trochanter, such variability indicates that the internal rota- tive function of these muscles is not criti- cal to the striding gait,
The negation of internal rotative power in australopithecine hips also fails to con- sider the basic dynamics of stride and evi- dence supplied by electromyography. First, swing phase is initiated by contraction of the hip flexors and plantarflexors. During the period between foot flat and toe off, the ground reaction force has a forwardly directed component. This component de- velops a moment which tends to rotate the limb around the opposite (support- ing) hip joint. I t i s this momen t which initiates pelvic rotation, i.e., internal rota- tion is produced by the dynamics of stride, not by any specific internal rotator. As the limb nears the completion of swing phase this original pelvic rotative energy will have been reduced and additional input may be required. By this time, however, the abductors of the supporting limb have ceased contraction, while the ilio-psoas, which is a more powerful internal rotator than the gluteals, does contract. Evidence that this muscle was well developed in Australopithecus is seen in the well defined ilio-psoas groove discussed above.
Further evidence of full capacity to in- ternally rotate among australopithecines also comes from electromyographic stud- ies of spastic children with internal rota- tion gait. These have shown that a large proportion of the excessive internal rota- tion in such children is produced by the contraction of the internal hamstrings and the gracilis (Southerland, et al., '69), and that a portion of such internal rotation could be corrected by surgical transfer of these muscles to the posterior surface of the lateral condyle. Given the high bi- condylar angle of Australopithecus (see below), it is very probable that these same muscles would have also been efficient in-
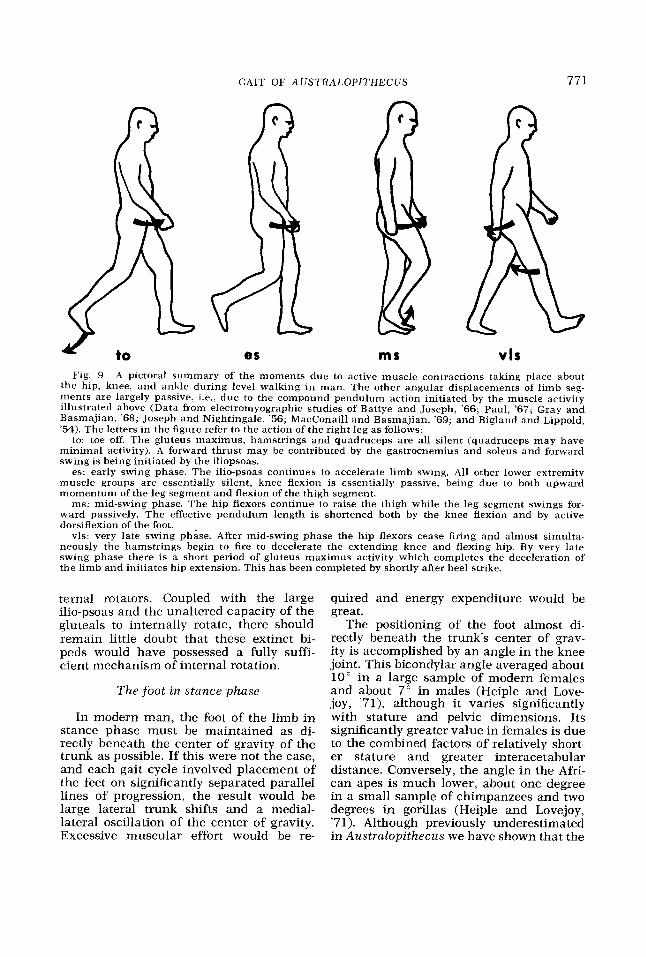
GAIT OF AUSTRALOPITHECUS 771
es ms vls Fig. 9 A pictoral summary of the moments due to active muscle contractions taking place about
the hip, knee, and ankle during level walking in man. The other angular displacements of limb seg- ments are largely passive, i.e., due to the compound pendulum action initiated by the muscle activity illustrated above (Data from elwtromyographic studies of Battye and Joseph, '66; Paul, '67; Gray and Basmajian, '68; Joseph and Nightingale, '56; MacConaill and Basmajian, '69; and Bigland and Lippold, '54). The letters in the figure refer to the action of the right leg as follows:
to: toe off. The gluteus maximus, hamstrings and quadruceps are all silent (quadruceps may have minimal activity). A forward thrust may be contributed by the gastrocnemius and soleus and forward swing is being initiated by the iliopsoas.
es: early swing phase. The ilio-psoas continues to accelerate limb swing. All other lower extremity muscle groups are essentially silent, knee flexion is essentially passive, being due to both upward momentum of the leg segment and flexion of the thigh segment.
ms: mid-swing phase. The hip flexors continue to raise the thigh while the leg segment swings for- ward passively. The effective pendulum length is shortened both by the knee flexion and by active dorsiflexion of the foot.
vls: very late swing phase. After mid-swing phase the hip flexors cease firing and almost simulta- neously the hamstrings begin to fire to decelerate the extending knee and flexing hip. By very late swing phase there is a short period of gluteus maximus activity which completes the deceleration of the limb and initiates hip extension. This has been completed by shortly after heel strike.
ternal rotators. Coupled with the large ilio-psoas and the unaltered capacity of the gluteals to internally rotate, there should remain little doubt that these extinct bi- peds would have possessed a fully suffi- cient mechanism of internai rotation.
The foot in stance phase
In modern man, the foot of the limb in stance phase must be maintained as di- rectly beneath the center of gravity of the trunk as possible. If this were not the case, and each gait cycle involved placement of the feet on significantly separated parallel lines of progression, the result would be large lateral trunk shifts and a medial- lateral oscillation of the center of gravity. Excessive muscular effort would be re-
quired and energy expenditure would be great.
The positioning of the foot almost di- rectly beneath the trunk's center of grav- ity is accomplished by a n angle in the knee joint. This bicondylar angle averaged about 10" in a large sample of modern females and about 7" in males (Heiple and Love- joy, '71), although it varies significantly with stature and pelvic dimensions. Its significantly greater value in females is due to the combined factors of relatively short- er stature and greater interacetabular distance. Conversely, the angle in the Afri- can apes is much lower, about one degree in a small sample of chimpanzees and two degrees in gorillas (Heiple and Lovejoy, '71). Although previously underestimated in Australopithecus we have shown that the

772 C. 0. LOVEJOY, K. G. HEIPLE AND A. H. BURSTEIN
most accurate method of estimation yields very high bicondylar angles in the distal femoral samples of STS-34 and TM-1513 - about 14" and 15" respectively (Heiple and Lovejoy, '71).
The existence of such great bicondylar angles in these femoral fragments posi- tively confirms the above discussions con- cerning abduction and internal rotation. Either the placement of the shank and foot during stance phase was almost directly beneath the center of gravity of the trunk in australopithecine locomotion (i.e., ef- fective abduction and internal rotation were present) or the knee joint in these early hominids was simply not adapted to locomotion at all, i.e., placement of the knee joint in a more lateral position with such a high bicondylar angle would have resulted in very high shear in the knee joint. That this angle should be greater in STS-34 and TM-1513 than in most modern females is not surprising, if we once more consider the longer lateral lever arm and somewhat shorter stature which probably characterized these individuals.
Hip extension
The great breadth of the ilium, deep sciatic notch, and extensive retroauricu- lar part of the sacro-pelvic surface (Dart, '49, '57, '58) adequately demonstrate that the gluteus maximus was an efficient hip extensor in both the gracile and robust forms of Australopithecus. It has none the less been argued that the conformation of the ischium in these early hominids is in- consistent with a striding gait. The posi- tion and length of the ischium have been accorded far more significance than is warranted with respect to the locomotion of Australopithecus. In man the hamstrings are not powerful hip extensors. The sciatic fibers of the adductor magnus add only a small amount of additional extensor mass. The diminuitive condition of these muscles is related to both the massive development of the gluteus maximus and to the bio- mechanics of bipedal gait.
Contrary to the requirements of the striding gait, jogging and running necessi- tate almost constant activity in the mus- cles of the hip, and an additional propul- sive force is to be found in active hip extension. The hamstrings are poor ex-
tensors for this purpose. They are not only relatively gracile, but have little leverage (the length of the lever arm of these mus- cles rapidly approaches zero as the limb is extended), and they cross both the hip and knee joints, which greatly reduces propul- sive efficiency in a biped. It is clear from the anatomy and biomechanics of hominid bipeds that a more powerful extensor (glu- teus maximus) is a necessity for rapid and active locomotion with orthograde posture. The cross sectional area of this muscle (which is directly proportional to its max- imum force output) is consistently greater than that of any other hip muscle. Ye t the gluteus max imus has been shown to be silent during the propulsive phase of hu- m a n stride (cf. fig. 9 and references there- in).
Gravity, inertia, and the mechanics of compound pendula are the primary ele- ments of striding gait. Indeed, the great efficiency of this type of locomotion is predicated upon the greatly reduced re- quirements for muscular contraction. In progressive forward stride, locomotion is accomplished by movement of the trunk forward over the extended hip and not by active movement of the leg and hip pos- teriorward. In fact, electromyographic studies show that extensor activity occurs only during the early part of stance phase, and these muscles are thus not a source of propulsion in stride (cf. fig. 9 and refer- ences therein). The leg, after heel strike, remains in almost complete extension as the trunk's momentum carries it over the somewhat stiffened limb. Its momentum is supplied by the alternate contraction of the plantarflexors of each foot while the flexors of the hip contribute to the alter- nate progressive swing of the legs. The role of the hip extensors in this process is minimal and the key anatomical correlates of the striding gait are thus to be found on the pelvis and femur in the form of those features which we have already dis- cussed. The negation of stride on the basis of differences in the ischia of Australo- pithecus and modern man thus cannot be substantiated.
T h e ischium
There appears to have been throughout the literature a rather uniform judgment

GAIT OF AUSTRALOPITHECUS 773
Fig. 10 Three-quarter lateral views of the STS-14 innominate (left) and young adult Bushman of equal size (right) (Wenner-Gren Casts). The lower portions, below the mid- acetabular line, are in equal amounts of rotation. The apparent relative narrowness of the upper blade of the ilium of Azistrctlopithecus is an artifact of foreshortening due to the more laterally flared anterior portion (compare with fig. 1). The true (functional) length of the ischium is indicated (B and B’) from the center of rotation of the acetabulum to the mid- point of the tuberosity. The vertical height of the ilium (with the innominate in approximate anatomical position) is also indicated (A and A’). Although the ischium of STS-14 is visibly more gracile than that of the Bushman t it is neither absolutely nor relatively (A/B:A’/B’) longer (cf. table 3) . Comparison of these casts and modern human material with published figures for ischial length make it apparent that the ischial length has frequently been mea- sured as the distance from the rim of the acetabulum to the margin of the ischial tuberosity. Such a measurement is unrelated to the biomechanical, anatomical, or embryological length of the ischium. See also figure 11 and its legend.
t Care must he taken with observations on the Wenner-Gren cast of the right innominate of STS-14. Dr. Wolpoff reports that the right ischial ramus and pubis are distorted, being crushed laterally and uuwards. A measurement from the top of the svmphysis to the acetabular rim gives 63.2 mm for the left . - side hut only 44.6 mm for the right side.
that the ischium of Australopithecus is significantly longer than that of modern man. These conclusions have been based upon three specimens: SK-50; MLD-8; and, STS-14. Inspection of figure 10, however, shows that there is a serious problem with regard to what is to be considered the “length’ of the ischium. Upon compari- son of the ischia of either SK-50 or STS-14 and that of a Bushman it is apparent that the absolute length of the interval from the margin of the acetabulum to the margin of the hamstring impression is
greater in the australopithecine samples than it usually is in modern man. Schultz has pointed out the important distinction between anatomical variations which are distinct in two separate groups as opposed to those which have overlapping ranges of variation (‘50). Specimens from recent human populations which display the “australopithecine type” of long ischium (acetabular margin to hamstring impres- sion) are not inf’requent (fig. l l ) , and al- though the ischial apophysis of MLD-8 has not yet fused, this specimen appears to

774 C. 0. LOVEJOY, K. G. HEIPLE AND A. H. BURSTEIN
Fig. 11 Two ischia taken from a small sample (N = 39) of innominates from one Amerindian skeletal population (left: KSU-02061; right: KSU-08201). The acetabula are of the same size and the functional ischial lengths are 52 mm (left) and 76 mm (right) (cf. fig. 10). The distance from the lip of the acetabulum to the border of the hamstring impression in the right specimen is 28 mm which is greater than in SK-50 despite a greater acetabular diameter in this specimen (50 mm) than in SK-50. Moreover, the distance is nine times greater in the right specimen than in the left one, showing this measurement can have little functional significance.
have had an ischial “length” more typi- cal of modern man.
The greater distance between the ace- tabular margin and hamstring impression in some australopithecine samples is ac- countable by consideration of other ana- tomical variants of the innominate. First, as was noted above, the relative size of the femoral head of Australopithecus ap- pears to be somewhat smaller than that of
modern man. The shorter relative diam- eter of the acetabula of AustTaZopithecus probably results in a longer segment of bone from its margin to that of the ham- string impressions. Secondly, the morphol- ogy of the ischial tuberosity of STS-14 and SK-50 is somewhat different than that usually characteristic of modern innomi- nates. In the modern innominate, the ischial tuberosity is most often separated

GAIT OF AUSTRALOPITHECUS 775
into superior and inferior portions by a distinct angulation (this appears also to have been the case in MLD-8). The more even-surfaced superior portion bears the impression of the hamstrings, while the more inferior portion bears the impression of the adductor magnus. In the australo- pithecine innominates (SK-50 and STS-14) the tuberosity lacks this regional differen- tiation via angulation, i.e., in these two specimens the origin of the hamstrings is not “wrapped around” as is most often the case in modern man. This again results in a greater distance between the margin of the acetabulum and that of the ham- strings.
If we are to attempt some measure of the “length’ of the ischium in Australo- pithecus such a measure must be of some functional significance. The total length of the ischium when treated as a morpholog- ical lever arm (distance from the center of the acetabulum to the center of the hamstring impression) in STS-14 is 42 mm. In a Bushman specimen of similar size (Wenner-Gren Cast) this distance is 43 mm. If the relative length of this morpho- logical lever arm is to be determined, another comparable measurement of some other morphological lever arm is necessary. A reasonable comparison is the length of the morphological lever arm of the ilium. This may be defined as the distance from the center of the acetabulum to the max- imum height of the ilium, taken along the maximum thickness of the iliac pillar (fig. 10). The ratio of the lengths of these two morphological lever arms (ischial length/ iliac height) is almost identical to that of a sample of modern innominates (table 3).
Thus far we have only discussed the morphological length of the ischium. How- ever the true biomechanical length of this lever arm depends upon the degree of hip flexion at which it is measured, and varies significantly with ordinary changes in erect posture. Slight alterations in either of these two variables have pronounced ef- fects upon its length. For example, Thieme (‘50) found that a plane determined by the pubis and anterior superior iliac spines varied t 10” from the vertical among a small series of erect medical students. The effect of even this small range of varia- tion upon the moment arm of the ham- strings is shown in figure 12. It can be
TABLE 3
Functional and relative ischial length of Australopithecus and H . sapiens
Ischial Iliac Specimen length (A) height (B) AIB
mm mm STS-14 1 43 92 0.47 SK-50 1 60 MLD-8 2 0.48 Bushman 3 42 89 0.47 Homo sapiens 60.2 125.6 0.48 Homo sapiens S.D. 5.31 8.75 0.034
N = 25
1 Measurements on original specimens. 2 Measurement from Drawing in Dart (’49). 3 Measurement on Wenner-Gren cast.
seen from this figure that despite an un- changing ischial length, the moment arm of the hamstrings can vary by more than a 3:2 ratio within this slight range of pos- tural variation. One would suspect, given this data, that the hamstrings would only be important to gait when their biomechan- ical lever arm was consistently favorable to their contraction. This is indeed the case, as is shown by the electromyographic evidence (see references in fig. 9). The brief period of contraction recorded for the hamstrings in normal stride corre- sponds to that phase in which the hip is most flexed (i.e., the end of swing phase and beginning of stance phase). There is no evidence that the hamstrings of Austra- lopithecus would have a significantly dif- ferent moment arm than those of modern man during this phase of stride.
Summary of the evidence
The following list summarizes those points of the anatomical evidence from the hip and knee joints of Australopithecus which are important with regard to stride. A notation follows each indicating whether it is known to characterize the gracile form, robust form, or both.
1. The ilium is broad and of generally hominid configuration. A well-formed sig- moid curve is present as well as a deep sciatic notch. The sacroiliac joint is ori- ented similarly to that of modern man (Le Gros Clark, ’55, ’67). (Gracile and Robust)
2. The morphological length of the fem- oral neck appears to be significantly great- er than that of modern man (table 1) and the neck-shaft angle lies significantly be-

776 C. 0. LOVEJOY, K. G. HEIPLE AND A. H. BURSTEIN
Fig. 12 Lateral views of the pelvis and proximal femur of Homo sapiens, drawn both in 10’ of anterior and 10” of posterior tilt from the “normal” vertical (the plane of the anterior-most part of the symphysis and the anterior tip of the superior spine). hf indicates the direction of hamstring force in both instances, while il is the true ischial length, identical in both figures. The ischial moment arm (im) (the right angle distance from the line of hamstring force to the center of rotation of the aceta- bulum) varies markedly with these small changes in pelvic position. It is 1.6 times greater in the right hand diagram than in the left.
low the mean of Homo sapiens. These fea- tures in combination produce a biomechan- ical length of the femoral neck relatively greater than that found in modern man. (Gracile and Robust)
3. The ilium is laterally flared relative to that of sapiens. The lateral biomechan- ical lever arm is thus relatively greater than that of sapiens. (Gracile and Robust)
4. Hip joint pressure in STS-14 was probably only half that of modern man. (Gracile)
5. The biomechanical length of the fem- oral necks of SK-82, SK-97, and OH-20 indicates that their joint reaction forces were probably less than the average of modern man. (Robust)
6. A prominent iliac pillar is present in all ilia thus far recovered. (Gracile and Robust)
7. The position of the iliac pillar is slightly anterior to that of modern man. In conjunction with lateral iliac flare, this
indicates that the function of the abduc- tors was homologous in Australopithecus and modern man. (Gracile and Robust)
8. A cristal tubercle was probably pres- ent in the only iliac sample sufficiently preserved to exhibit evidence of it. (Grac- ile)
9. A cross section of the ilia in the re- gion of the pillar shows considerable thick- ening of the iliac cortical bone. (Gracile)
10. The natural split lines formed in some ilia (MLD-7 and MLD-25) are almost identical to those of modern man. (Gracile)
11. The prominent anterior superior iliac spine, in conjunction with the later- ally flared ilium, places the origins of the sartorius, internal and external obliques, and inguinal ligament in the same rela- tive position as in modern man. (Gracile and Robust)
12. The anterior inferior iliac spine is well-formed and prominent. (Gracile and Robust)

GAIT OF AUSTRALOPITHECUS 777
13. The development of the intertro- chanteric line of the femur is within the range of variation of modern man. These last two features indicate a prominent role in locomotion of the straight head of the rectus femoris and a well developed ilio-femoral ligament. (Gracile and Robust)
14. The prominent ilio-psoas groove in- dicates that locomotion was habitually erect and that the ilio-psoas muscle was a prominent flexor and internal rotator. (Gracile and Robust)
15. The position of the lesser trochanter is similar to that of modern man and is consistent with anteversion of the femur. (Gracile and Robust)
16. There is a high bicondylar angle indicating that the foot was placed be- neath the body's center of gravity during the gait cycle. (Gracile)
17. The deep inter-condylar groove is related to the prevention of patellar dis- location in the completely extended knee with a high bicondylar angle. (Heiple and Lovejoy, '71). (Gracile)
18. The medial condylar projections of the distal femoral fragments indicate a se- lective increase of true cartilage contact in the habitually extended knee (Heiple and Lovejoy, '71). (Gracile)
19. The biomechanical length of the ischial moment arm is within the range of variation of modern man. (Gracile and Ro- bust)
20. An obturator externus depression is present on the posterior aspect of the neck of OH-20, SK-82, and SK-97 (Day, '69), indicating consistent complete extension of the femur during stance and gait. (Ro- bust)
21. A well developed ischial spine is present on the only sufficiently preserved innominate for which casts are available, indicating a well developed sacrospinal ligament. (Gracile)
Encephalization and pelvic-femoral evolution
It has thus far been found that the hip complex of Australopithecus shows virtual- ly complete adaptation to the bipedal gait, including that form known as stride. In comparison with Homo sapiens it was found that femoral head pressures were prob- ably equal to or less than those of modern
man because of the more advantaged posi- tion of the abductors in these early bi- peds. The question must therefore neces- sarily arise as to why the hip complexes of Australopithecus and Homo sapiens dif- fer from one another morphologically. Why should the highly advantageous position of the abductors be sacrificed in Pleisto- cene hominid evolution? The reduction of lateral flare of the ilium (and of its com- mensurate morphological features: ante- rior iliac pillar position, longer biomechan- ical neck length, anterior superior spine position, etc.) leads to an increase in the pressure suffered by the articular carti- lage of the hip joint.
Our answer to this question is the rapid encephalization which took place during the Pleistocene. There would have been during this period a significant selection upon the capacity of the female to main- tain adequate parturition dimensions in the face of a constantly increasing fetal cranial size. Just how intense was this se- lection is difficult to assess, since many factors are obviously in operation. Socio- cultural intervention into the birth prac- tice itself, retardation of the fetal matura- tion rate, and the maintenance of the bipedal gait are perhaps three of the most important. All such factors must have interplayed during the Pleistocene in re- sponse to hominid encephalization. If we assume, however, that despite those adap- tations which tended to reduce the selec- tion pressure of fetal head size at birth, there was still a need to enlarge the pelvic ring of the female, some other changes in the morphology of the hip would be re- quired in the evolution of Pleistocene hom- inids.
First, there is certainly a maximum total pelvic breadth which can be effi- ciently maintained in a biped of any given stature. For example, beyond certain pel- vic breadthlstature ratios rapid locomotion becomes cumbersome and awkward and the efficiency of stride is lost. The pelvic breadth of Australopithecus was already great because of the laterally flared ilium and relatively broad interacetabular dis- tance (compared to that of quadrupeds). In the face of increasing fetal cranial di- mensions, adjustments which increased the pelvic ring but which did not increase total pelvic breadth would have been fa-

7 78 C. 0. LOVEJOY, K. G. HEIPLE AND A. H. BURSTEIN
Total pelvic breadth I
Fig. 13 Schematic representation of pelvic evolution during the Pleistocene (anterior view). While maximum pelvic breadth remains un- changed, the interacetabular distance increases with encephalization. This increases the torque developed about the hip by body weight in stance phase and at the same time decreases the lever arm of the abductors. As a result, the diameter of the femoral head must be markedly larger in Homo sapiens than in Australopithecus in order to maintain femoral head pressure within physiolog- ical limits. These changes also necessitate greater iliac robusticity in Homo sapiens as the total ab- ductor force will be much greater.
vored. The result would have been those changes in pelvic morphology which we have already discussed above. This is il- lustrated in figure 13. It can be seen from this figure that if stature and total pelvic breadth remain unchanged relative to one another, and the dimensions of the partur- ition canal are to be increased, the only avenue of change is an increase in the medial lever arm length and commensur- ate reduction in the length of the lateral lever arm. Such changes would lead to substantially increased femoral head pres- sures and perhaps the only adaptation available in response to such an increase would be an enlargement of the femoral head. In addition, the resulting increase in the muscle force of the abductor mech- anism would require a commensurate in- crease in the iliac pillar and general robus- ticity of the ilium. It is precisely these changes in addition to those which have been discussed above which can be seen between the hip complexes of Austrulo- pithecus and Homo sapiens.
CONCLUSIONS
The evidence which has been present- ed in this paper leads us to the following conclusions :
1. There is no morphological or bio-
mechanical feature by which the two pres- ently recognized allomorphs of Australo- pithecus differ significantly with respect to their locomotor adaptations. Rather, both clearly conform to the same total morphological pattern and more signifi- cantly, total biomechanical pattern. Fur- thermore, this pattern is as fully commen- surate with erect striding as is that of modern man.
2. Those morphological differences be- tween Austrulopithecus and modern man which are of biomechanical significance appear to be related to the combination of a fully bipedal striding gait with differ- ent degrees of encephalization, rather than to differences in the gait pattern itself.
ACKNOWLEDGMENTS
We wish to thank Dr. Milford Wolpoff, Dr. C. Loring Brace, and Dr. Michael Day for their aid in supplying critical data necessary for the analyses presented. We are indebted to the Cleveland Museum of Natural History for their permission to study the Hamann-Todd Collection and to Dr. C. K. Brain and the Transvaal Muse- um, for permission to use data supplied by Dr. Brace and Dr. Wolpoff and to reproduce figure 4. We thank Mr. Richard S . Meindl, Dept. of Anthropology, Kent State Univer- sity for aid in calculations of hip joint pressure. The photographs (other than fig. 4) are by Mr. Lawrence Rubin.
LITERATURE CITED
Aitken, J. T., G. Causey, J. Joseph and J. Z. Young 1966 A Manual of Human Anatomy. Living- stone, Edinburgh.
Battye, C. K., and J. Joseph 1966 An investiga- tion by telemetering of the activity of some mus- cles in walking. Med. and Biol. Enging., 4: 125- 134.
Bigland, B., and 0. C. J. Lippold 1954 The re- lation between force, velocity, and integrated electrical activity in human muscles. J. Physiol., 192: 773-800.
Broom, R., J. T. Robinson and G. W. H. Schepers 1950 Sterk-fontein Ape-Man, Plesianthropus. Transvaal Mus. Mem., No. 4.
Campbell, B. G. 1966 Human Evolution. Aldine, Chicago.
Chopra, S. R. K. 1962 The innominate bone of the Australopithecinae and the problem of erect posture. Bibl. Primat., 1 : 93-102.
Dart, R. 1949 Innominate fragments of Austra- lopithecus prometheus. Am. J. Phys. Anthrop., 7: 301-334.
1957 The second adolescent (female) ilium ofAustralopithecus prometheus. J. Palaeon. SOC. India, 2: 73-82.

GAIT OF AUSTRALOPITHECUS 779
~ 1958 A further adolescent ilium from Makapansgat. Am. J . Phys. Anthrop., 16: 473- 479.
Day, M. H. 1959 Guide to Fossil Man. Camelot Press, London.
1969 Femoral fragment of a robust australopithecine from Olduvai Gorge, Tanzania. Nature, 221 ; 23G233.
Frankel, V. F., and A. H. Burstein 1970 Ortho- paedic Biomechanics. Lea and Febiger, Phila- delphia.
Gray, E. G., and J. V. Basmajian 1968 Electro- myography and cinematography of leg and foot (“Normal” and flat) during walking. Anat. Rec., 161 : 1-16.
Heiple, K. G., and C. 0. Lovejoy 1971 The distal femoral anatomy of Australopithecus. Am. J . Phys. Anthrop., 35: 75-84.
Joseph J., and A. Nightingale 1956 Electromyog- raphy of muscles of posture: leg and thigh mus- cles in women, including the effects of high heels. J. Physiol., 132: 46-68.
Kern, H. H., and W. L. Straus 1949 The femur of Plesianthropus transvualensis. Am. J . Phys. Anthrop., 7: 53-77.
Le Gros Clark, W. E. 1955 The 0s innominatum of the recent pongidae with special reference to that of the Australopithecinae. Am. J. Phys. Anthrop., 13: 1!+27.
Man-apes or Ape-Men. Holt, Rine- hart and Winston, New York.
Lovejoy, C. 0. 1972 The biomechanics of stride and their bearing on the gait ofAustralopithecus. Paper presented at the forty-first annual meet- ing of the American Association of Physical A n - thropologists.
Lovejoy, C . O., and K. G. Heiple 1970 A recon- struction of the femur of Australopithecus afri- cnnus . Am. J. Phys. Anthrop., 32: 3%40.
1972 Proximal femoral anatomy of Australopithecus. Nature, 235: 175-1 76.
Mac Conaill, M. A,, and J. V. Basmajian 1969 Muscles and Movements. Williams and Wilkins, BaItimore.
McLeish, R. D., and J. Charnley 1970 Abduction forces in one-legged stance. J. Biomechanics, 3: 191-210.
Mednick, L. W. 1955 The evolution of the human ilium. Am. J. Phys. Anthrop., 13: 203-216.
1967
Napier, J. R. 1964 The evolution of bipedal walk- ing in the hominids. Arch. Biol. (Liege), 75 (Suppl): 673-708.
The antiquity of human walking. Sci. Amer., 216: 56-66.
Paul, J. P. 1967 Forces at the human hip joint. Ph.D. Thesis, University of Glasgow.
Pauwels, F. 1935 Der Schenkelhalsbruck ein Mechanische Problem. Ferd. Enke., Stuttgart.
Preuschaft, H. 1971 Body Posture and Mode of Locomotion in Early Pleistocene Hominids. Folia primat., 14: 209-240.
Reynolds, E. 1931 The evolution of the human pelvis in relation to the mechanics of the erect posture. Pap. Pea. Mus. Amer. Arch. Ethn. Har- vard Univ., 1 I : 25.5382.
Schultz, A. H. 1950 The physical distinctions of man. Proc. Am. Phil. SOC., 94: 428449.
Simons, E. L. 1967 The significance of primate palaeontology for anthropoIogical studies. Am. J. Phys. Anthrop., 27: 307-332.
Simpson, G. G. 1963 The meaning of taxonomic statements. In: Classification and Human Evolu- tion. S. L. Washburn, ed. Aldine, Chicago,
Southerland, D. H., E. R. Schottstaedt, L. J. Lar- sen, R. K. Ashley, J. N. Callander and P. M. James 1969 Clinical and electromyographic study of seven spastic children with internal rotation gait. J. Bone Jt. Surg., 51-A: 1070-1082.
Straus, W. L. 1962 Fossil evidence for the evolu- tion of the erect bipedal posture. Clin. Ortho- paed., 25: 9-19,
Tappen, N. C. 1969 The relationship of weather- ing cracks to split line orientation in bone. Am. J. Phys. Anthrop., 31 : 191-198.
Thieme, F. P. 1950 Lumbar breakdown caused by erect posture in man. Anthrop. Pap. Mus. Anthrop. Univ. Michigan, 4: 1-44.
Washburn, S. L. 1950 The analysis of primate evolution with particular reference to the origin of man. Cold Spring Harbor Symp., 15: 67-78.
1960 Tools and human evolution. Sci. Amer., 203: 6%75.
Zhilman, A. L. 1969 Human Locomotion: A re- appraisal of the functional and anatomical evi- dence. Ph.D. Thesis, University of California, Berkeley.
1967
pp. 1-31.
Note added in proof: This article was submitted prior to the publication of de- scriptions of the new East African post-cranial samples. These specimens will be discussed in a later presentation.




















