
The effect of modifications in photoperiod on spawning time, ovarian development and egg quality in...
15
Aquaculture, 81 (1989) 351-365 351 Elsevier Science Publishers B.V., Amsterdam - Printed in The Netherlands The Effect of Modifications in Photoperiod on Spawning Time, Ovarian Development and Egg Quality in the Sea Bass (Dicentrarchus lab-ax L.) M. CARRILLO’, N. BROMAGE’, S. ZANUY’, R. SERRANO’ and F. PRAT’ ‘Institute de Acuicultura de Torre de la Sal, C.S.I.C., Ribera de Cabanes, 12545 Castellon (Spain) 21nstitute of Aquaculture, University of Stirling, Stirling FK9 4LA (Great Britain) (Accepted 15 February 1989) ABSTRACT Carillo, M., Bromage, N., Zanuy, S., Serrano, R., and Prat, F., 1989. The effect of modifications in photoperiod on spawning time, ovarian development and egg quality in the sea bass (Dicen- trarchus labrux L.). Aquaculture, 81: 351-365. Under simulated natural conditions female sea bass (Dicentrurchus labrax) in eastern Spain (latitude 4O”N and longitude 0” ) spawned over a 6-8-week period in February and March, with each female spawning on more than one occasion. Generally, male fish began spermiation up to 2 months before, and continued for at least 1 month after the spawning period of the female fish. Exposure of fish to 1 month of long days (LD 15/9) from either 2nd May (Group C), 3rd June (Group D) or 3rd July (Group E) in an otherwise constant short day (LD 9/15) photoperiod regime, speeded up the rates of maturation, thus increasing the proportions of oocytes entering exogenous vitellogenesis during October and November, and also brought forward the timings of ovulation and spawning. In contrast, constant long days from 2nd May (Group B) delayed ma- turation and spawning time by 2-3 months. Fish maintained under constant short days through- out the experiment (Group A) spawned up to 6 weeks in advance of the control fish, suggesting that endogenous timing mechanisms may operate in this species. Spawning occurred naturally in all the experimental and control fish without the necessity for induction with pituitary or hypothalamic hormones. For the control fish, egg quality (defined as the proportion of eggs spawned which were floating and viable), hatching rate and survival to first feeding (both expressed as percentages of the numbers of floating or good quality eggs) averaged 78.3 + 7.7% (mean + se. ), 84.8 + 3.3% and 54.6 + 20% respectively. With the exception of the eggs and fry from Group B, these measures of quality and survival appeared unaffected by the photoperiod treatments. The eggs and fry pro- duced by the fish maintained under long days (Group B) towards the end of their spawning period (i.e. in May), showed much more variable egg quality and survivals through hatch and first- feeding, possibly because of the higher sea water temperatures which prevailed at the time of their delayed spawning. Fecundities expressed as numbers of eggs per kg of post-spawning fish weight ranged from 247- 305 OOO/kg for Groups A, C, D and E and the controls, but were significantly reduced in Group B fish. Egg sizes, which ranged from 1.147-1.176 mm in diameter, were unaffected by the photope- riodic manipulations of spawning time. 0044-8486/89,‘$03.50 0 1989 Elsevier Science Publishers B.V.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of The effect of modifications in photoperiod on spawning time, ovarian development and egg quality in...

Aquaculture, 81 (1989) 351-365 351
Elsevier Science Publishers B.V., Amsterdam - Printed in The Netherlands
The Effect of Modifications in Photoperiod on Spawning Time, Ovarian Development and Egg Quality in the Sea Bass (Dicentrarchus lab-ax L.)
M. CARRILLO’, N. BROMAGE’, S. ZANUY’, R. SERRANO’ and F. PRAT’
‘Institute de Acuicultura de Torre de la Sal, C.S.I.C., Ribera de Cabanes, 12545 Castellon
(Spain)
21nstitute of Aquaculture, University of Stirling, Stirling FK9 4LA (Great Britain)
(Accepted 15 February 1989)
ABSTRACT
Carillo, M., Bromage, N., Zanuy, S., Serrano, R., and Prat, F., 1989. The effect of modifications in photoperiod on spawning time, ovarian development and egg quality in the sea bass (Dicen-
trarchus labrux L.). Aquaculture, 81: 351-365.
Under simulated natural conditions female sea bass (Dicentrurchus labrax) in eastern Spain (latitude 4O”N and longitude 0” ) spawned over a 6-8-week period in February and March, with each female spawning on more than one occasion. Generally, male fish began spermiation up to 2 months before, and continued for at least 1 month after the spawning period of the female fish.
Exposure of fish to 1 month of long days (LD 15/9) from either 2nd May (Group C), 3rd June (Group D) or 3rd July (Group E) in an otherwise constant short day (LD 9/15) photoperiod regime, speeded up the rates of maturation, thus increasing the proportions of oocytes entering exogenous vitellogenesis during October and November, and also brought forward the timings of ovulation and spawning. In contrast, constant long days from 2nd May (Group B) delayed ma- turation and spawning time by 2-3 months. Fish maintained under constant short days through- out the experiment (Group A) spawned up to 6 weeks in advance of the control fish, suggesting that endogenous timing mechanisms may operate in this species.
Spawning occurred naturally in all the experimental and control fish without the necessity for induction with pituitary or hypothalamic hormones.
For the control fish, egg quality (defined as the proportion of eggs spawned which were floating and viable), hatching rate and survival to first feeding (both expressed as percentages of the numbers of floating or good quality eggs) averaged 78.3 + 7.7% (mean + se. ), 84.8 + 3.3% and 54.6 + 20% respectively. With the exception of the eggs and fry from Group B, these measures of quality and survival appeared unaffected by the photoperiod treatments. The eggs and fry pro- duced by the fish maintained under long days (Group B) towards the end of their spawning period (i.e. in May), showed much more variable egg quality and survivals through hatch and first- feeding, possibly because of the higher sea water temperatures which prevailed at the time of their delayed spawning.
Fecundities expressed as numbers of eggs per kg of post-spawning fish weight ranged from 247- 305 OOO/kg for Groups A, C, D and E and the controls, but were significantly reduced in Group B fish. Egg sizes, which ranged from 1.147-1.176 mm in diameter, were unaffected by the photope- riodic manipulations of spawning time.
0044-8486/89,‘$03.50 0 1989 Elsevier Science Publishers B.V.

352 M. CARRILU) ET AL.
It is concluded that artificial control of daylength can improve the supply of eggs and fry for commercial on-growing without any adverse effects on the fecundity of the broodstock or the quality and survival of the eggs and fry.
INTRODUCTION
In Europe the sea bass is a much prized fish, both as a source of food and for angling (Bromage et al., 1988). Supplies of this fish from conventional hunting or trawling, however, remain limited and as a consequence there is a continuing interest in its potential for culture. One aspect which specifically constrains the farming of this species is an ability to control fully its reproduction, in particular for the provision of eggs or fry for on-growing. Manipulation of day- length has been used to alter spawning time in this species (Girin and Devau- chelle, 1978; Barnabe and Paris, 1984; Devauchelle, 1984; Zanuy et al., 1986), although the mechanism of this control and the possible requirements for as- sociated injections of pituitary hormones to induce final maturation remain unclear (Barnabe and Paris, 1984; Barnabe and Barnabe-Quet, 1985; Zanuy et al., 1986). There is also disagreement in the literature relating to the effects of photoperiod treatments on fecundity, fertility and egg quality (Girin and De- vauchelle, 1978; Zanuy et al., 1986; Devauchelle, 1987). These questions are addressed in the present investigation.
MATERIALS AND METHODS
Beginning on 2 January, groups of lo-12 2-year-old previously immature female sea bass (Dicentrurchus Zubralc L. ) , 335 ? 15 g (mean 2 s.e. ) in weight and 31.8 t- 0.45 cm in length, were maintained in aerated running sea water (37.8%0 ) in light-proof 2-m-diameter circular (2000 1 capacity) tanks and ex- posed to one of the following photoperiod regimes: Group A: LD 9/15 (9 h light followed by 15 h dark in each 24 h cycle), i.e. constant short days. Group B: LD 9/15 until 2nd May followed by LD 15/9 (constant long days). Group C: LD 9/15 until 2nd May, LD 15/9 until 3rd June and then LD 9/15. Group D: LD 9/15 until 3rd June, LD 15/9 until 3rd July and then LD 9/15. Group E: LD 9/15 until 3rd July, LD 15/9 until 3rd August and then LD 9/15.
Daylength was controlled by electronic timeclocks with tungsten bulbs pro- viding an artificial day light of 480 lux at the water’s surface. The photoperiod regimes employed, together with the water temperature which was uncon- trolled and hence fluctuated with ambient condition, are shown in Fig. 1. An- other group of fish, maintained in an adjacent tank under ambient temperature and light (40’ N; 0 ’ ), served as controls.

THE EFFECT OF MODIFICATIONS IN PHOTOPERIOD ON THE SEA BASS 353
Hr tight/day
a ‘It-
1
JFMAMJJASOND
Fig. 1. Photoperiodic regimes (h light/day) received in Groups A, B, C, D and E (a-e) together with the ambient light cycle (at 40”N (f) and the seasonal temperatures to which all the fish were exposed (g). Horizontal axis, months of the year January (J) to December (D).
All fish were individually marked with a brand cooled in liquid nitrogen on their undersurface, and fed ad libitum each day with fresh chopped trash fish (Boops boops) . Male fish were present in all tanks throughout in order to main- tain normal sexual behaviour and spawning and to ensure fertilization of any eggs produced.
At the beginning of each month, between 11.00 and 13.00 h, all fish were anaesthetized in a solution of 100 ppm MS-222, weighed and fork length re- corded, and examined for the development of secondary sexual characteristics and the presence of spermiation, i.e. sperm release on gentle hand pressure or “stripping”. All fish were starved for 24 h before anaesthetization. In addition, from September onwards biopsies were removed through a cannula inserted into the ovary via the vent. Biopsies were not carried out on fish until Septem- ber because of the relatively small size of the developing gonad up to this date.
Ovarian biopsies were examined under a microscope and the oocytes staged

354 M. CARRILLO ET AL.
according to Zanuy et al. (1986) to establish whether the fish were ready to spawn; subsequently, biopsied tissue was fixed in Bouin-Hollande fixative for histological processing and detailed examination. Fixed ovarian biopsies were embedded in the methacrylate plastic Historesin (LKB ), sectioned at 4-5 /urn and stained in either Ehrlich’s haematoxylin and eosin or Cleveland Wolf’s. After histological processing, counts were made of the proportions of oocytes in previtellogenesis, the vesicle (“endogenous yolk” or cortical alveolus) stage, exogenous vitellogenesis (incorporation of yolk), final maturation or ovula- tion and atresia. Males were not cannulated but spermiation was a useful pre- dictor of impending female maturation because sperm could generally be ex- pressed from males 2 months before final maturation of female fish.
In order to time ovulation and spawning, which occurred naturally in all the experimental and control fish, a fine gauge net was placed across the outflow from each tank, allowing all the spawned eggs to be collected. By checking the outflow nets every 24 h, the number of days on which spawning had occurred could be established for all groups. Subsequently, at the monthly samplings ovarian biopsies allowed the exact number of fish which had spawned over the previous month to be determined. It was not possible to ascertain the exact number of spawnings which had been carried out by individual females because the eggs from each tank were pooled in the outflow collection nets; nor was it possible to establish how many fish had spawned, unless, at the subsequent monthly examination, only one fish had spawned during that month.
From the eggs collected each day, the percentage which were of good quality was determined by measuring the proportion of floating (good quality) to sink- ing (poor quality) eggs in a measuring cylinder of sea water. Sinking eggs in- cluded both infertile and damaged ova whereas the floating group from both control and experimental fish consistently exhibited fertilization rates in ex- cess of 90%. Egg size was determined by measuring the diameter of 100 good quality eggs.
Subsequently, the number of good quality floating eggs which survived to hatch and then to first-feeding (yolk-sac reabsorption) was also recorded, as were the percentage of fish in each group which spawned, the total number of eggs produced and the relative fecundity (i.e. the number of egg/kg of post- spawned female body weight). Differences in egg quality and viability were statistically tested using Scheffe’s multiple range analysis and the Kruskal- Wallis test, respectively. Arcsine transformations were used as appropriate to normalise percentage distributions.
RESULTS
Oocytes in exogenous vitellogenesis were clearly distinguishable from pre- vitellogenic oocytes and those of the vesicle stage of development in the sec- tions of ovaries of experimental and control fish (Fig. 2a, b and c) . In control

THE EFFECT OF MODIFICATIONS IN PHOTOPERIOD ON THE SEA BASS 355
Fig. 2. Photomicrographs of 4-5 pm thick sections of ovaries of sea bass under natural conditions (controls) stained with Ehrlich’s haematoxylin and eosin, showing previtellogenic (a), vesicle stage (b), exogenously vitellogenic (c), atretic (d) and ovulated (e) oocytes. Magnification (a,b) x 190; (c,d,e) x 120.


THE EFFECT OF MODIFICATIONS IN PHOTOPERIOD ON THE SEA BASS 357
100
1 GROUP D
80
Y&J@
60
40
20
”
cm m EC JAN FEE MAR
GROUP E
DEC JAN
CONTROLS
FEE MAR
CCT NW EC JAN FE8 MAR APR
Fig. 3. Proportions (% ages ) of previtellogenic, vesicle stage, exogenously vitellogenic, fully mature or ovulated and atretic oocytes (mean 5 s.e. of mean) in the ovary of female sea bass exposed to photoperiods A, B, C, D and E and in control fish.
358 M. CARRILLO ET AL.
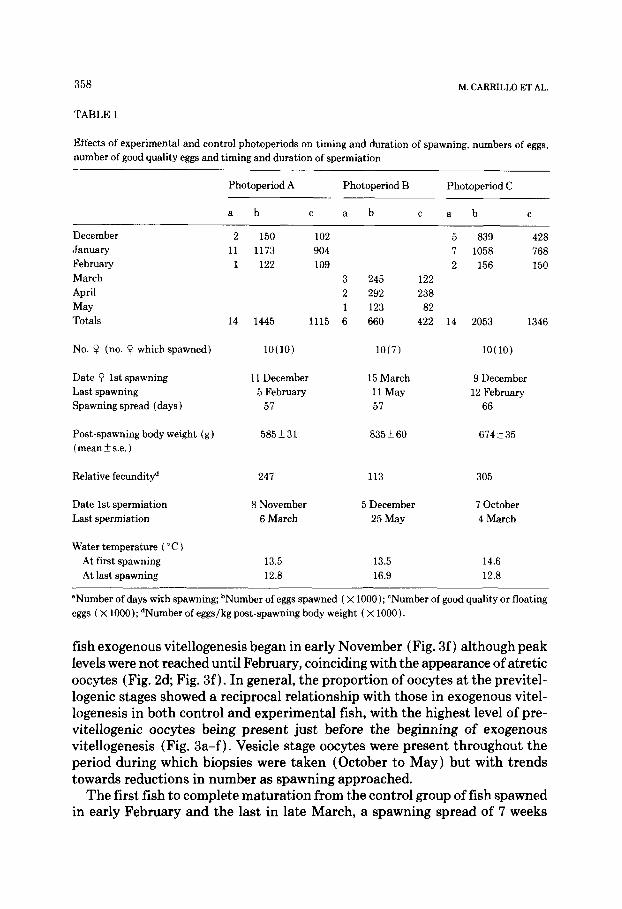
TABLE 1
Effects of experimental and control photoperiods on timing and duration of spawning, numbers of eggs,
number of good quality eggs and timing and duration of spermiation
Photoperiod A Photoperiod B Photoperiod C
a b C a b c a b C
December 2 150 102 5 839 428
January 11 1173 904 I 1058 768
February 1 122 109 2 156 150
March 3 245 122
April 2 292 238
May 1 123 82
Totals 14 1445 1115 6 660 422 14 2053 1346
No. 9 (no. 0 whichspawned) lO(10)
Date 9 1st spawning
Last spawning
Spawning spread (days )
11 December
5 February
57
Post-spawning body weight (g)
(mean _+ s.e. ) 585231
Relative fecundityd 247
Date 1st spermiation
Last spermiation
8 November
6 March
Water temperature ( “C ) At first spawning
At last spawning
13.5 13.5 14.6
12.8 16.9 12.8
lO(7) lO( 10)
15 March
11 May
57
9 December
12 February
66
835 ?I 60 674 + 35
113 305
5 December
25 May
7 October
4 March
“Number of days with spawning; bNumber of eggs spawned ( X 1000); ‘Number of good quality or floating
eggs ( x 1000); dNumber of eggs/kg post-spawning body weight ( X 1000).
fish exogenous vitellogenesis began in early November (Fig. 3f) although peak levels were not reached until February, coinciding with the appearance of atretic oocytes (Fig. 2d; Fig. 3f). In general, the proportion of oocytes at the previtel- logenic stages showed a reciprocal relationship with those in exogenous vitel- logenesis in both control and experimental fish, with the highest level of pre- vitellogenic oocytes being present just before the beginning of exogenous vitellogenesis (Fig. 3a-f) . Vesicle stage oocytes were present throughout the period during which biopsies were taken (October to May) but with trends towards reductions in number as spawning approached.
The first fish to complete maturation from the control group of fish spawned in early February and the last in late March, a spawning spread of 7 weeks

THE EFFECT OF MODIFICATIONS IN PHOTOPERIOD ON THE SEA BASS 359
Photoperiod D Photoperiod E Control
a b C a b C a b c
I 1000 537 2 303
8 938 680 9 925
1 14 10 6 704
16 1952
lO(10)
4 December
13 February
72
647 + 43
302
3 October
2 March
14.8 14.6 11.9
11.5 11.0 13.8
1227 17 1927
lO(10)
9 December
26 February
80
649 k 29
297
5 November
2 April
163
493
130
786
5
6
11
977 821
1060 689
2037 1510
9(9)
2 February
20 March
46
811+54
279
2 December
20 April
(Table 1). Control male fish were ‘running’ (emitting sperm on gentle hand pressure) from early December to late April.
The photoperiodic treatments induced significant changes in the timing of both the different stages of oocyte development and of spawning. Vitellogen- esis was most advanced in the fish exposed to one month of long days in May (Group C; Fig. 3c ) and most delayed in those which were on constant long days from May onwards (Group B; Fig. 3b). Although there was only a 1 month separation between the initial appearance of exogenous vitellogenic oocytes in the ovaries of Group B (November) and those of Group C (October) fish, peak levels of exogenously vitellogenic oocytes were found in December in Group C but not until February in Group B.

360 M. CARRILLO ET AL.
TABLE 2
Effects of experimental and control photoperiods on egg size, egg quality, hatching rate and survival of fry to first-feeding
A B C D E Controls
Egg diameter (pm) 1160.8&34.8 1144.9k34.3 1164.7k24.4 1147.1?24.1 1150.7k43.7 1176.3k47.0
Egg quality (% )” (Number of blanks ) b
Hatching rate (% )’ (Number of blanks)d
Survival to first feeding
(%)”
80.12 4.3 58.7f 12.6 68.5 f 8.3 64.1f7.7 47.6f 11.8 78.3 t 7.7
(1) (1) (1) (2) (6) (0) 83.2 & 6.7 21.4k2.8 81.2 + 6.3 64.5 ? 9.7 78.9 f 7.5 84.8 i 3.3
(1) (1) (0) (2) (1) (0) 56.9 f 19.7 8.6 f 2.4 58.8 i 6.5 36.6 f 28.6 58.3 f 8.7 54.6 i 20.8
“Number of eggs spawned which floated i.e. were of good quality. bBlanks are the number of spawnings in which the %age of good eggs was < 5.0%.
“Hatching rate and survival to first-feeding both expressed as percentages of the number of good quality or floating eggs.
dThe number of blanks at hatching does not include those which were blanks at fertilization. All values are means + s.e. of the mean. No significant differences in egg quality; the number of egg quality blanks significantly increased in Group E (Pi 0.05); hatching rate and survival to first feeding significantly reduced in Group B (bothP<O.OOl).
Groups D and E, with 1 month of long days in June and July respectively, were in turn marginally delayed with regards to the timing of exogenous vitel- logenesis (Figs. 3d and 3e) when compared with Group C, with peak levels being reached in December/January and January respectively. Exogenously vitellogenic oocytes did not appear in the ovaries of Group E fish until Novem- ber (Fig. 3e), one month behind Groups C and D. These differences were re- flected in the profile of spawning of the three groups which showed concentra- tions in December and January for Groups C and D respectively, but not until January/February for group E (Table 1) . Fish maintained under constant short days throughout the experiment (Group A) showed similar rates of develop- ment to Group E with oocytes in exogenous vitellogenesis appearing for the first time in November but peak levels not being reached until December/ January (Fig. 3a). Spawning in Group A began in December but like Group E fish there was a concentration of spawnings in January (Table 1).
Male fish showed parallel changes to the females in the timing of maturation in response to the different photoperiod regimes with Groups C and D males beginning ‘running’ (commencing spermiation) before Groups A and E and the fish under both these treatments ‘running’ before group B and the controls.
The proportion of female fish which matured was reduced in Group B whereas in all the other treatments and the controls, all fish spawned. This difference was also reflected in the total numbers of eggs produced and in the apparent relative fecundities (Table 1); the values for Group B fish being less than half those of the other experimental and control fish.
There was little difference in the spawning spread of the different groups with the first female in each treatment spawning approximately 6325 (mean 2 s.e.) days before the last in each group, and the males running with

THE EFFECT OF MODIFICATIONS IN PHOTOPERIOD ON THE SEA BASS 361
milt for 143 + 5 days, generally commencing spermiation l-2 months in ad- vance and finishing up to 1 month after the spawning period of the correspond- ing group of females.
Although the experimental protocol of this investigation did not allow the number of spawnings of each individual fish to be determined, because the eggs spawned over the previous 24 h from each of the experimental and control tanks were pooled, it averaged l-2 spawnings per fish.
Eggs from Groups A, C and D had a similar quality (i.e. % floating) to those of controls (Table 2 ) . Groups B and E showed a significantly greater variabil- ity in egg quality (PC 0.05) having approximately 20% fewer good eggs than the others. This included, for the fish in Group E, six spawnings in which the percentages of good eggs were less than 5% (Table 2); these are often referred to as blanks. The eggs and fry from Group B showed significantly poorer levels of survival to hatch (PC 0.001) and to the time of first-feeding or yolk sac resorption (P < 0.001) than the remaining experimental and control fish.
DISCUSSION
The present results show clearly that the rates of maturation and timing of spawning in the sea bass can be modified by photoperiodic change. This con- firms and, by using light regimes of constant lengths, extends the findings of Girin and Devauchelle (1978)) Devauchelle (1984)) Barnabe and Paris (1984) and Barnabe and Barnabe-Quet (1985) who only studied the effects of modi- fied seasonally changing light cycles.
It would appear that the sequence of changes of daylength provided by the photoperiod regimes C, D and E offered similar zeitgebers or environmental cues to those present in the natural seasonal light cycle. Clearly photoperiodic regime B did not provide the appropriate series of light cues, for spawning was delayed in this group. Similar results with light regimes involving a sequence of 3-4 different constant photoperiods during the year have also been reported for the brook trout (Henderson, 1963), rainbow trout (Whitehead and Brom- age, 1980; Bromage et al., 1984; Bromage and Duston, 1986) and masu salmon (Takashima and Yamada, 1984). Specifically, Bromage et al. (1984) showed that rainbow trout exposed to constant long days after the summer solstice were delayed in their spawning; in parallel with the present results the last authors also showed that only 6 weeks of long days in May/June in an other- wise short day photoperiod regime is all that is required to advance spawning time.
One important question which has been addressed in the present investiga- tion is that eggs whose maturation has been advanced by photoperiodic ma- nipulation can undergo natural spawning in the same way as controls. Other authors have had to use hCG or LHRH-A injections in conjunction with pho- toperiod treatments (Barnabe and Paris, 1984; Barnabe and Barnabe-Quet,

362 M. CARRILLO ET AL.
1985 ), thus bringing into question the relative efficacies of the hormonal and environmental methods of spawning advancement.
An important finding of the present study is that different light cues are not required for the completion of reproduction, for spawning still occurred in Group A fish which were maintained throughout under constant short days. This strongly suggests that endogenous mechanisms may be involved in the timing of reproduction. Recently, Bromage and Duston (1986) and Duston and Bromage (1986) have reported that such mechanisms operate to coordinate maturation and spawning in the rainbow trout. Extension of the present work over more than one reproductive cycle will, however, be necessary before the presence of an endogenous rhythm or clock can be firmly established for the sea bass. The advancements of spawning in Groups C and D are, however, compatible with a hypothesis which involves the entrainment of an endoge- nous rhythm or clock by photoperiodic change, the direction of change of pho- toperiod in relation to the phase of the internal rhythm producing phase changes in the endogenous clock. Under photoperiodic regimes C and D the mainte- nance of fish under short days until May and June respectively would have resulted in a phase delay of the clock; this would have been followed in both C and D by successive phase advances, firstly when daylength was increased in either May and June and then when it was reduced in June and July respec- tively. Similar results after exposure to long and then short days have also been reported for other fish (Henderson, 1963; Bromage et al., 1984; Takashima and Yamada, 1984).
Further support for this hypothesis also comes from a re-examination of the work of Devauchelle (1984) on the sea bass. In a series of experiments extend- ing over 6 years this author progressively advanced the spawning of sea bass by exposing fish firstly to three lo-month condensed seasonal light cycles and then to three 12-month seasonal light cycles which were 6 months out-of-phase with ambient daylength. Initially, the timing of spawning of the experimental group showed a phase delay in relation to the accelerated cycle in which they were being maintained, with spawning occurring at a slightly longer photope- riod than would have been expected if there had been a direct inductive effect of daylength on maturation. Gradually, with the progression of the fish onto the succession of 12-month cycles, spawning occurred at exactly the same day- length as the controls. Such phase delays in response to ‘driving zeitgebers’ have been widely reported in descriptions of circadian rhythms but only re- cently have such suggestions been applied to the behaviour of annual or cir- cannual rhythms in fish (Bromage and Duston, 1986; Duston and Bromage, 1986,1987,1988).
An important consideration if photoperiod techniques are to be used effec- tively to control the spawning times of farmed stocks of sea bass is that there must be little or no associated loss in the quality i.e. survival of the eggs and fry and/or the fecundity of the broodstock. Devauchelle and co-workers (Girin

THE EFFECT OF MODIFICATIONS IN PHOTOPERIOD ON THE SEA BASS 363
and Devauchelle, 1978; Devauchelle, 1984, 1987 ) all reported slightly poorer survivals amongst the eggs from fish which had been photoperiodically ad- vanced. With the possible exception of group E no such loss in egg quality was experienced with the photoperiod advancements of spawning in the present work. This is in agreement with studies of both Barnabe and Paris (1984) and Barnabe and Barnabe-Quet (1985). Like the eggs from Group E fish, those from the photoperiodically delayed Group B also showed extremely variable quality and patterns of survival. It is difficult to explain why Groups E and B should exhibit such variability in egg quality. However, it is possible that this was due to the inappropriately high sea water temperature extant during the spring and early summer when most of the spawning of the Group B fish and the last of those of Group E occurred. Zanuy et al. (1986) also suggested that high sea temperatures were the reason for the poor egg survivals of fish whose spawning had been delayed by photoperiodic treatment. Although this might account for the poor quality of the eggs from Group B fish it can not explain the results of the Group E fish which spawned at approximately the same time as controls but with a markedly poorer egg quality. Clearly, this question will only be answered when further investigations are made of the effects on egg quality of photoperiodic alterations in spawning time whilst maintaining tem- perature at seasonally constant levels. It is possible that the sperm quality of the male fish from Group E deteriorated during the spawning season; such a change has been reported by Billard et al. (1977) and Billard (1984). Unfor- tunately, at present there is no entirely suitable means of assessing sperm qual- ity or viability; also it was not possible to include any estimations of the fertil- ization rate of the poor quality or sinking eggs in the present study.
Although it was not possible to determine the relative fecundities of individ- ual fish in the present work, because of the way in which eggs were collected, the average levels of fecundity/kg post-spawning female body weight (i.e. rel- ative fecundities) for the fish from Groups A, C, D and E were similar to the controls, all of them approximately 250-300 000 eggs/kg; only Group B fish had significantly lower relative fecundities. Levels of 300 OOO/kg were also re- corded by Devauchelle (1984) with no reported effects of photoperiod treat- ment. In contrast Barnabe and Paris (1984) estimated that the relative fe- cundities of photoperiodically delayed fish were higher than for fish under ambient conditions, although their comparisons were made with groups spawning in different years and twice the size of the experimental fish. Possi- bly, the different fish sizes may have contributed to this difference in fecundity, as increased relative fecundities have been reported with smaller fish size in other species (Springate and Bromage, 1984; Springate et al., 1985; Bromage and Cumaranatunga, 1988). Zohar et al. (1984) gave values of 200 000 eggs/ kg which are similar to those provided by Barnabe (1973,198O) and Barnabe and Paris (1984); Barnabe (1980) stated that 250 000 eggs/kg was the maxi- mum number which could be expected from sea bass with 200 000 being the

364 M. CARRILLO ET AL.
norm. Somewhat higher relative fecundities were recorded by Kennedy and Fitzmaurice (1972) although their determinations of fecundity were carried out on the fixed ovaries of pre-spawning fish. Thus, it would appear that the relative fecundities described in the present work for fish which had their spawning advanced by photoperiod manipulation, although a little higher than that found in the controls, were generally similar to those found in other stud- ies of sea bass. Although in other species of fish increased numbers of eggs are sometimes achieved by a reduction in the size of egg produced (Springate and Bromage, 1984; Springate et al., 1985) there was no evidence that such a “trade- off” was operating in the present experiments for the eggs were of similar size to that described by Boulineau (1974) and Barnabe (1980).
In summary, it will appear that simple methods of photoperiodic control would enable supplies of eggs or fry to be made available at exactly those times required by on-growing sea bass farms. The high fecundities of sea bass mean that only small numbers of broodstock would need to be maintained under photoperiod control in order to achieve the necessary hatchery production ob- jectives. Although, in general, there would appear to be no adverse effects of these treatments as far as the fecundity of the broodstock and the survival of the eggs and fry are concerned, at present we are investigating the possibility that inappropriately high temperatures at spawning may reduce egg quality.
ACKNOWLEDGEMENTS
Our thanks to J. Llorens, R. Simo and A. Ibanez for their technical assis- tance. This work was supported by an Anglo-Spanish Joint Research Pro- gramme (SPN/99/3/44) and CAICYT Research Grant (3/83).
REFERENCES
Barnabe, G., 1973. Mass rearing of the bass Dicentrarchus labrar. FAO Fish Rep., 141: 29-30. Barnabe, G., 1980. Expose synoptique des don&es biologiques sur le loup ou bar Dicentrarchus
labrax (Linne 1748). Synopsis FAO sur les pbches, No. 126 (FIR/S126). FAO, Rome, pp. l- 70.
Barnabe, G. and Paris, J., 1984. Ponte avancee et ponte normale du loup Dicentrarchus labrax (L) a la Station de Biologie Marine et Lagunaire de Sete. In: G. Barnabe and R. Billard (Ed- itors), L’Aquaculture du Bar et des Sparides. INRA Pub]., Paris, pp. 63-72.
Barnabe, G. and Barnabe-Quet, R., 1985. Avancement et amelioration de la ponte induite chez le loup Dicentrarchus labran (L) a l’aide dun analogue de LH-RH inject& Aquaculture, 49(2): 125-132.
Billard, R., 1984. La conservation des gametes et l’insemination artificielle chez le bar et la daur- ade. In: G. Barnabe and R. Billard (Editors), L’Aquaculture du Bar et des Sparides. INRA Publ., Paris, pp. 95-116.
Billard, R., DuPont, J. and Barnabe, G., 1977. Diminution de la motilite et de la duke de conser- vation du sperme de Dicentrarchus labrax L. (Poisson Teleosteen) pendant la periode de sper- miation. Aquaculture, 11: 363-367.

THE EFFECT OF MODIFICATIONS IN PHOTOPERIOD ON THE SEA BASS 365
Boulineau, F., 1974. Ponte naturelle et ponte induite hormonalement chez Dicentrarchus labrax
(L.) en captivitk. Publ. CNEXO. Ser: Actes de Colloq., NO. 1: 151-160. Bromage, N.R. and Duston, J., 1986. The control of spawning in the rainbow trout (Salmo gaird-
neri) using photoperiod techniques. Rep. Inst. Freshwater Res. Drottningholm, 63: 26-35. Bromage, N.R. and Cumaranatunga, P.R.T., 1988. Egg production in the rainbow trout. In: J.
Muir and R. Roberts (Editors), Advances in Aquaculture, Vol. 3. Chapman and Hall, London, pp. 63-138.
Bromage, N.R., Elliott, J.A., Springate, J.R.C. and Whitehead, C., 1984. The effects of constant photoperiods on the timing of spawning in the rainbow trout. Aquaculture, 43: 213-223.
Bromage, N.R., Carrillo, M. and Zanuy, S., 1988. Light controls spawning in the sea bass. Fish Farm Int., 15: 22.
Devauchelle, N., 1984. Reproduction decalee du bar (Dicentrarchus labrax) et de la daurade (Spa- rus aurata). In: G. Barnabe and R. Billard (Editors) L’Aquaculture du Bar et des Sparides. INRA Publ., Paris, pp. 53-61.
Devauchelle, N., 1987. Four marine spawners in European hatcheries. In: Production Control&e en Ecloserie. Synthese des papiers present& dans le cadre du MEDRAP a Roviny-Zadar (Yu-
goslavia), 18-28 January 1986. Rapport FAO, Rome, 50 pp. Duston, J. and Bromage, N.R., 1986. Photoperiodic mechanisms and rhythms of reproduction in
the female rainbow trout. Fish. Physiol. Biochem., 2: 35-51. Duston, J. and Bromage, N.R., 1987. Constant photoperiod regimes and the entrainment of the
annual cycle of reproduction in the female rainbow trout (Salmo gairdneri). Gen. Comp. En- docrinol., 65: 733-784.
Duston, J. and Bromage, N., 1988. The entrainment and gating of the endogenous circannual rhythm of reproduction in the female rainbow trout (Salmo gairdneri). J. Comp. Physiol. A, 164: 259-268.
Girin, M. and Devauchelle, N., 1978. Decalage de la periode de reproduction par raccourcissement des cycles photoperiodiques et thermiques chez des poissons marins. Ann. Biol. Anim. Biochim. Biophys., 18: 1059-1065.
Henderson, N.R., 1963. Influence of light and temperature on the reproductive cycle of the eastern brook trout (Salvelinus fontinalis Mitchell). J. Fish Res. Board Can., 20: 859-897.
Kennedy, M. and Fitzmaurice, P., 1972. The biology of the bass Dicentrarchus labrax, in Irish waters. J. Mar. Biol. Assoc. U.K., 52: 597.
Springate, J.R.C. and Bromage, N.R., 1984. Egg size and number - it’s a trade-off. Fish Farmer, 7: 12-14.
Springate, J.R.C., Bromage, N.R. and Cumaranatunga, P.R.T., 1985. The effects of different ra- tion on fecundity and egg quality in the rainbow trout (Salmo gairdneri). In: C.B. Cowey, A.M. Mackie and J.G. Bell (Editors), Feeding and Nutrition in Fish. Proc. Fish. Sot. Br. Isles. Academic Press, London, pp. 371-391.
Takashima, F. and Yamada, Y., 1984. Control of maturation in masu salmon by manipulation of photoperiod. Aquaculture, 43: 243-257.
Whitehead, C. and Bromage, N.R., 1980. Effects of constant long and short day photoperiods on the reproductive physiology and spawning of the rainbow trout. J. Endocrinol., 87: 6-7.
Zanuy, S., Carrillo, M. and Ruiz, F., 1986. Delayed gametogenesis and spawning of sea bass (Di- centrarchus labran L.) kept under different photoperiod and temperature regimes. Fish Phys- iol. Biochem., 2: 53-63.
Zohar, Y., Billard, R. and Weil, C., 1984. La reproduction chez la daurade (Sparus aurata) et Ie bar (Dicentrarchus labrax): Connaissance du cycle sexuel et controle de la gametogenese et de la ponte. In: G. Barnabe and R. Billard (Editors), L’Aquaculture du Bar et des Sparides. INRA Publ., Paris, pp. 3-24.




















