
Wild juvenile Dicentrarchus labrax L. liver antioxidant and damage responses at Aveiro Lagoon,...
10
Wild juvenile Dicentrarchus labrax L. liver antioxidant and damage responses at Aveiro Lagoon, Portugal V.L. Maria a,b , I. Ahmad a , M. Oliveira a , A. Serafim b , M.J. Bebianno b , M. Pacheco a , M.A. Santos a, a CESAM & Biology Department of Aveiro University, 3810-193 Aveiro, Portugal b CIMA & Faculty of Marine and Environmental Sciences, University of Algarve, 8005-139 Faro, Portugal article info Article history: Received 16 October 2008 Received in revised form 13 May 2009 Accepted 9 June 2009 Available online 5 July 2009 Keywords: Fish D. labrax Genotoxicity Oxidative stress Metalothioneins Liver Aveiro Lagoon abstract The Aveiro Lagoon, at the north-western coast of Portugal, has been under considerable anthropogenic pressure for the last 5 decades. In order to perform an adequate survey of the effects induced by the contaminants in presence, wild juveniles Dicentrarchus labrax (sea bass) were selected. Thus, sea bass was captured at five sites: Torreira (TOR, as reference site), Gafanha (GAF), Rio Novo Pr´ ıncipe (RIO), Laranjo (LAR) and Vagos (VAG) in autumn 2005. Liver defence responses such as catalase (CAT), glutathione peroxidase (GPx), glutathione reductase (GR), glutathione S-transferase (GST), total glutathione (GSHt), total non-protein thiols (NP-SH) and metallothioneins (MT) were measured. Liver damage was determined as lipid peroxidation (LPO) and DNA integrity loss. RIO, LAR and VAG presented lower CAT, GR, GST activities and NP-SH and GSHt depletion in comparison to TOR. VAG and LAR showed higher GPx activity when compared to TOR. The highest MT level was found at GAF and VAG. The NP-SH and DNA integrity decreased at GAF compared to TOR. This field study demonstrated that not only antioxidant induction but also inhibitory responses must be considered as a signal of contamination. & 2009 Elsevier Inc. All rights reserved. 1. Introduction Aveiro Lagoon is a coastal system (47km 2 of maximum surface area) permanently connected to the sea, located in the northern west region of Portugal, surrounded by several urban aggregates and industrial centres. Historically, this lagoon has been the main receptor of anthropogenic discharges resulting mainly from chlor- alkali and pulp/paper plants, harbour, dry-dock and agriculture activities as well as municipal and domestic effluents. Hence, there are some critical areas revealing the imperative need of an effective biomonitoring programme. According to Bucheli and Fent (1995), biomarkers, representing toxicant-induced changes in biological systems, can serve as links between an environ- mental contamination (cause) and its effects, providing therefore unique information on the ecosystem health. The integrated use of a wide battery of biomarkers allow different levels of investigation and resolution depending on the sensitivity and biological significance of analysed responses (Regoli et al., 2002b). Different responses will be expected concerning eventual additive, synergistic or antagonistic interactions whenever aquatic species face complex mixtures of contaminants. Many studies on the toxicological effects of aquatic contaminants have been performed on a variety of bioindicators, especially in fish (Pacheco et al., 2005; Ahmad et al., 2006b; Maria et al., 2006, 2008). In the current work, Dicentrarchus labrax L. (sea bass) were chosen as a top predator species entering the Aveiro Lagoon and occurring in the entire system. Sea bass also represents a keystone species with ecological and economic importance in the Eastern Atlantic Ocean, Mediterranean and Black Sea. Moreover, different D. labrax biomarkers, namely phase I and phase II biotransformation (Gravato and Santos, 2003), intermediary metabolism and general stress (Teles et al., 2006), vitelogenin levels (Teles et al., 2007), DNA and lipid damage (Maria et al., 2008) have been successfully applied for the evaluation of environmental water contamination. Under normal physiological conditions, liver metabolism of the chemicals in the body can generate reactive oxygen species (ROS), which may be removed by cellular defence systems, which include enzymatic and non-enzymatic antioxidants. The most important antioxidant enzymes are superoxide dismutase (SOD), which catalyses the dismutation of superoxide anion (O 2 d ) to hydrogen peroxide (H 2 O 2 ), catalase (CAT) and glutathione perox- idase (GPx) that are in charge of reducing H 2 O 2 to H 2 O(Harris, 1992; Fridovich, 1997). Whenever, antioxidant defences are inefficient combating the action of ROS, the result is oxidative stress, leading to lipid peroxidation (LPO) and DNA damage (Livingstone, 2001). Antioxidant enzymes and substrates, LPO and DNA damage responses in fish have been commonly and successfully employed ARTICLE IN PRESS Contents lists available at ScienceDirect journal homepage: www.elsevier.com/locate/ecoenv Ecotoxicology and Environmental Safety 0147-6513/$ - see front matter & 2009 Elsevier Inc. All rights reserved. doi:10.1016/j.ecoenv.2009.06.001 Corresponding author. Fax: +351234 426 408. E-mail addresses: [email protected] (V.L. Maria), [email protected] (I. Ahmad), [email protected] (M. Oliveira), aserafi[email protected] (A. Serafim), [email protected] (M.J. Bebianno), [email protected] (M. Pacheco), [email protected] (M.A. Santos). Ecotoxicology and Environmental Safety 72 (2009) 1861–1870
Transcript of Wild juvenile Dicentrarchus labrax L. liver antioxidant and damage responses at Aveiro Lagoon,...

ARTICLE IN PRESS
Ecotoxicology and Environmental Safety 72 (2009) 1861–1870
Contents lists available at ScienceDirect
Ecotoxicology and Environmental Safety
0147-65
doi:10.1
� Corr
E-m
miguelo
(M.J. Be
journal homepage: www.elsevier.com/locate/ecoenv
Wild juvenile Dicentrarchus labrax L. liver antioxidant and damage responsesat Aveiro Lagoon, Portugal
V.L. Maria a,b, I. Ahmad a, M. Oliveira a, A. Serafim b, M.J. Bebianno b, M. Pacheco a, M.A. Santos a,�
a CESAM & Biology Department of Aveiro University, 3810-193 Aveiro, Portugalb CIMA & Faculty of Marine and Environmental Sciences, University of Algarve, 8005-139 Faro, Portugal
a r t i c l e i n f o
Article history:
Received 16 October 2008
Received in revised form
13 May 2009
Accepted 9 June 2009Available online 5 July 2009
Keywords:
Fish
D. labrax
Genotoxicity
Oxidative stress
Metalothioneins
Liver
Aveiro Lagoon
13/$ - see front matter & 2009 Elsevier Inc. A
016/j.ecoenv.2009.06.001
esponding author. Fax: +351 234 426 408.
ail addresses: [email protected] (V.L. Maria),
[email protected] (M. Oliveira), [email protected]
bianno), [email protected] (M. Pacheco), mon
a b s t r a c t
The Aveiro Lagoon, at the north-western coast of Portugal, has been under considerable anthropogenic
pressure for the last 5 decades. In order to perform an adequate survey of the effects induced by the
contaminants in presence, wild juveniles Dicentrarchus labrax (sea bass) were selected. Thus, sea bass
was captured at five sites: Torreira (TOR, as reference site), Gafanha (GAF), Rio Novo Prıncipe (RIO),
Laranjo (LAR) and Vagos (VAG) in autumn 2005. Liver defence responses such as catalase (CAT),
glutathione peroxidase (GPx), glutathione reductase (GR), glutathione S-transferase (GST), total
glutathione (GSHt), total non-protein thiols (NP-SH) and metallothioneins (MT) were measured. Liver
damage was determined as lipid peroxidation (LPO) and DNA integrity loss. RIO, LAR and VAG presented
lower CAT, GR, GST activities and NP-SH and GSHt depletion in comparison to TOR. VAG and LAR showed
higher GPx activity when compared to TOR. The highest MT level was found at GAF and VAG. The NP-SH
and DNA integrity decreased at GAF compared to TOR. This field study demonstrated that not only
antioxidant induction but also inhibitory responses must be considered as a signal of contamination.
& 2009 Elsevier Inc. All rights reserved.
1. Introduction
Aveiro Lagoon is a coastal system (47 km2 of maximum surfacearea) permanently connected to the sea, located in the northernwest region of Portugal, surrounded by several urban aggregatesand industrial centres. Historically, this lagoon has been the mainreceptor of anthropogenic discharges resulting mainly from chlor-alkali and pulp/paper plants, harbour, dry-dock and agricultureactivities as well as municipal and domestic effluents. Hence,there are some critical areas revealing the imperative need of aneffective biomonitoring programme. According to Bucheli andFent (1995), biomarkers, representing toxicant-induced changesin biological systems, can serve as links between an environ-mental contamination (cause) and its effects, providing thereforeunique information on the ecosystem health. The integrated useof a wide battery of biomarkers allow different levels ofinvestigation and resolution depending on the sensitivity andbiological significance of analysed responses (Regoli et al., 2002b).Different responses will be expected concerning eventual additive,synergistic or antagonistic interactions whenever aquatic speciesface complex mixtures of contaminants. Many studies on the
ll rights reserved.
[email protected] (I. Ahmad),
(A. Serafim), [email protected]
[email protected] (M.A. Santos).
toxicological effects of aquatic contaminants have been performedon a variety of bioindicators, especially in fish (Pacheco et al.,2005; Ahmad et al., 2006b; Maria et al., 2006, 2008). In thecurrent work, Dicentrarchus labrax L. (sea bass) were chosen as atop predator species entering the Aveiro Lagoon and occurring inthe entire system. Sea bass also represents a keystone specieswith ecological and economic importance in the Eastern AtlanticOcean, Mediterranean and Black Sea. Moreover, different D. labrax
biomarkers, namely phase I and phase II biotransformation(Gravato and Santos, 2003), intermediary metabolism and generalstress (Teles et al., 2006), vitelogenin levels (Teles et al., 2007),DNA and lipid damage (Maria et al., 2008) have been successfullyapplied for the evaluation of environmental water contamination.Under normal physiological conditions, liver metabolism of thechemicals in the body can generate reactive oxygen species (ROS),which may be removed by cellular defence systems, whichinclude enzymatic and non-enzymatic antioxidants. The mostimportant antioxidant enzymes are superoxide dismutase (SOD),which catalyses the dismutation of superoxide anion (O2
�d) tohydrogen peroxide (H2O2), catalase (CAT) and glutathione perox-idase (GPx) that are in charge of reducing H2O2 to H2O (Harris,1992; Fridovich, 1997). Whenever, antioxidant defences areinefficient combating the action of ROS, the result is oxidativestress, leading to lipid peroxidation (LPO) and DNA damage(Livingstone, 2001).
Antioxidant enzymes and substrates, LPO and DNA damageresponses in fish have been commonly and successfully employed

ARTICLE IN PRESS
Fig. 1. Map of Aveiro Lagoon; the sampling sites were selected on a geographic
distribution basis along the main channels and at the lagoon entrance, taking into
account the various types and sources of contamination as well as the selection of
(theoretically) unpolluted reference points. The respective coordinates are:
reference site (TOR)—40144002 N, 008141044 W; GAF—40138038 N, 008141042 W;
RIO—40141008 N, 008139041 W; LAR—40143030 N, 008137043 W; and VAG
—40133059 N, 008140055 W.
V.L. Maria et al. / Ecotoxicology and Environmental Safety 72 (2009) 1861–18701862
in field studies aimed to characterise impacted areas, wherecomplex mixtures of pollutants are usually present (Livingstone,2001; Santos et al., 2004; Amado et al., 2006). Antioxidantenzymes and non-enzymatic antioxidants may be increased(Ahmad et al., 2006a; Guilherme et al., 2008) or decreased(Bagnyukova et al., 2005; Ahmad et al., 2006b) under chemicalstress. Thus, an increased activity can be considered as anadaptation, where the biological systems partially or totallyovercame stress resulting from exposure to an unsafe environ-ment. On the other hand, an inactivation in the defence system bythe chemical reactive species, which act as antioxidant enzymesinhibitors reduce the cell protection and the organism fitness, willbecome precarious (Kono and Fridovich, 1982; Escobar et al.,1996).
Toxicant-specific biomarkers, such as metallothioneins (MT),have also been widely employed to monitor fish environmentalexposure to metals such as cadmium (Cd), copper (Cu), nickel (Ni),zinc (Zn), lead (Pb) and mercury (Hg) (Li et al., 2004; Gigu�ereet al., 2006; Bebianno et al., 2007; Fernandes et al., 2007).However, procymidone, a non-metal fungicide, also demonstratedto be a causative agent of MT induction in fish (Paris-Palacios etal., 2003).
The assessment of aquatic species exposure to environmentalgenotoxicants is a complex issue because of the diversity ofpotential pollutants that are often found in aquatic systems ascomplex mixtures (Mitchelmore and Chipman, 1998; Shailaja etal., 2006). Nevertheless, the degree of DNA integrity has beenfrequently and successfully used as a sensitive indicator ofgenotoxicity and an effective biomarker for environmentalmonitoring (Regoli et al., 2004; Pacheco and Santos, 2002; Mariaet al., 2004a, b, 2006; Ahmad et al., 2006b).
Taking into account the potential severity of aquatic pollutionin the Aveiro Lagoon, the present research will focus onautochthonous D. labrax liver responses including CAT, GPx, GRand GST (antioxidant enzymes), non-protein thiols (NP-SH), totalglutathione (GSHt), and MT contents (non-enzymatic defencesystems) but also LPO and DNA strand breakage as damageindicators.
2. Materials and methods
2.1. Chemicals
Butylated hydroxytoluene (BHT), diethylenetriaminepenta-acetic acid (DTPA),
1-chloro, 2,4, dinitrobenzene (CDNB), 2,5, dithiobis-tetranitrobenzoic acid (DTNB),
2-thiobarbituric acid (TBA), trichloroacetic acid (TCA), b-nicotinamide adenine
dinucleotide phosphate reduced tetrasodium salt (NADPH), ethylene diamine
tetraacetic acid tetrasodium salt hydrate (EDTA), sodium azide, glutathione
reductase, reduced glutathione (GSH), oxidised glutathione (GSSG) and purified
rabbit liver MT were purchased from Sigma (Spain). Other routine chemicals and
reagents (analytical grade) were purchased from local sources.
2.2. Animals and study area
Juvenile specimens of European sea bass (D. labrax L.) were captured (n ¼ 6) in
October 2005 at the Aveiro Lagoon (Portugal) using a fishing-rod. The sampling
sites selection was based on a geographic distribution, along the main channels
and at the lagoon entrance, taking into account the various types and sources of
contamination as well as the selection of a (theoretically) unpolluted reference
point (Fig. 1). Thus, the capture sites were: Torreira (TOR) as a reference point at
the stretch of the longest channel (S. Jacinto-Ovar) having non-point pollution
inputs and far from the main polluting sources; Barra (BAR) at the initial part of the
Mira channel, close to the sea-side lagoon entrance and subjected to a considerable
naval traffic; Gafanha (GAF) situated in the vicinity of a deep-sea fishing port
and dry-docks, also connected with the main channel coming from Aveiro city and
receiving domestic discharges; Rio Novo do Prıncipe (RIO) located at the terminal
area of the Vouga River, 6.5 km distant from a pulp/paper mill effluent outlet,
was exposed to its discharges since nearly five decades up to 2000; Laranjo (LAR)
presenting a well-described mercury contamination gradient as a result of
chlor-alkali plant discharges (Pereira et al., 1998); Vagos (VAG) at the terminal
part of the �Ilhavo Channel, receiving municipal and domestic effluents.
Fish biometrical parameters, such as weight and total length ranged from
9.73–41.96 g and 10.7–16.5 cm, respectively. Hydrological abiotic parameters such
as temperature, dissolved oxygen, salinity, pH, turbidity and depth were also
measured (Table 1).
The current study was conducted in accordance with national and interna-
tional guidelines (Directive 2007/526/Ecv of the European Commission) for the
protection of animal welfare.
2.3. Tissue sampling and fractionation
Immediately after capture, fish were sacrificed and liver was then removed,
divided into three sets, frozen in liquid nitrogen and later stored at �80 1C. The first
set was used for further process in oxidative stress studies, the second set for MT
measurement and the third and last set for DNA strand breaks measurement.
The first set of tissue was homogenised, using a Potter-Elvehjem homogeniser,
in chilled phosphate buffer (0.1 M, pH 7.4) (1 g of tissue/15 ml buffer ¼ 15%). This
homogenate was then divided in three aliquots for LPO and glutathione
measurements as well as for post-mitochondrial supernatant (PMS) isolation for
further antioxidant parameters. The first aliquot was immediately used for
thiobarbituric acid reactive substances (TBARS) estimation. The second aliquot of
the homogenised was separated for NP-SH and GSHt measurements. Finally, the
PMS preparation was accomplished by centrifugation in a refrigerated ultracen-
trifuge (Optima TL, Beckman) at 13,400g for 20 min at 4 1C. Aliquots of PMS were
stored in microtubes, frozen in liquid nitrogen and stored at �80 1C until analyses.
2.4. Estimation of lipid peroxidation (LPO)
LPO was determined in the previously prepared tissue homogenate by the
procedure of Ohkawa et al. (1979) and Bird and Draper (1984) with some
ARTICLE IN PRESS
Table 1
Hydrological characteristics determined on different study sites at Aveiro Lagoon (Torreira—TOR, Gafanha—GAF, Rio Novo do Prıncipe—RIO, Laranjo—LAR and
Vagos—VAG): depth, turbidity, dissolved oxygen, water temperature (Temp.), pH and salinity.
Sites Depth (m) Turbidity (m) Dissolved O2 (mg/L) Temperature (1C) pH Salinity
TOR 1.2 0.8 6.88 18.9 8.172 32.5
GAF 6.9 0.7 5.78 17.9 8.28 34
RIO 4.8 1 6.37 19 8.199 21
LAR 1.2 0.3 4.96 18.6 7.735 29
VAG 3.5 0.3 5.18 18.5 7.88 33
V.L. Maria et al. / Ecotoxicology and Environmental Safety 72 (2009) 1861–1870 1863
modifications. Briefly, to a 50ml of 15% homogenate, 3ml of 4% BHT in methanol,
was added and mixed well. To this aliquot, 0.5 ml of 12% TCA in aqueous solution,
0.45 ml Tris–HCl (60 mM, pH 7.4; and 0.1 mM DTPA) and 0.5 ml 0.73% TBA were
added and mixed well. The mixture was heated for 1 h in a water bath set at boiling
temperature. The test tubes were then removed and cooled to room temperature.
The content from each tube was decanted and centrifuged at 13,400g for 3 min.
The absorbance of each aliquot was measured at 535 nm. The rate of LPO was
expressed as mM of TBARS formed per gram of fresh tissue using a molar extinction
coefficient of 1.56�105 M�1 cm�1.
2.5. Catalase (CAT) activity measurement
CAT activity was assayed in PMS fraction by the method of Claiborne (1985)
with some modifications. Briefly, the assay mixture consisted of 1.99 ml phosphate
buffer (0.05 M, pH 7.0), 1 ml H2O2 (30 mM) and 0.010 ml PMS in a final volume of
3 ml. Change in absorbance was recorded at 240 nm at 25 1C. CAT activity was
calculated in terms of mmol H2O2 consumed/min/mg protein using a molar
extinction coefficient of 43.5 M�1 cm�1.
2.6. Glutathione reductase (GR) activity measurement
GR activity was assayed by the method of Cribb et al. (1989) with some
modifications. Briefly, the assay mixture contained 0.025 ml of PMS fraction and
0.925 ml of NADPH (0.2 mM), GSSG (1 mM) and DTPA (0.5 mM). The enzyme
activity was quantified at 25 1C by measuring the disappearance of NADPH at
340 nm during 3 min. The enzyme activity was calculated as nmole NADPH
oxidised/min/mg protein using a molar extinction coefficient of 6.22�103 M�1
cm�1.
2.7. Glutathione S-transferase (GST) activity measurement
GST activity was determined in PMS fraction, following the conjugation of GSH
with CDNB by the method of Habig et al. (1974) with some modifications. The
reaction mixture consisted of 1.85 ml sodium phosphate buffer (0.1 M, pH 7.4),
0.050 ml GSH (1 mM), 0.050 ml CDNB (1 mM) and 0.050 ml PMS in a total volume
of 2 ml. The change in absorbance was recorded at 340 nm and 25 1C. Enzyme
activity was calculated as mmol CDNB conjugate formed/min/mg protein using a
molar extinction coefficient of 9.6 mM–1 cm�1.
2.8. Glutathione peroxidase (GPx) activity measurement
GPx activity was assayed in PMS fraction according to the method described by
Mohandas et al. (1984) with some modification. The assay mixture consisted of
0.720 ml phosphate buffer (0.05 M, pH 7.0), 0.050 ml EDTA (1 mM), 0.050 ml
sodium azide (1 mM), 0.025 ml GR (1 IU/ml), 0.050 ml GSH (4 mM), 0.050 ml
NADPH (0.8 mM), 0.005 ml H2O2 (1.0 mM) and 0.050 ml of PMS in a total volume of
2 ml. Oxidation of NADPH was recorded spectrophotometrically at 340 nm at 25 1C.
The enzyme activity was calculated as nmol NADPH oxidised/min/mg of protein,
using a molar extinction coefficient of 6.22�103 M�1 cm�1.
2.9. Non-protein thiols (NP-SH) content
Protein content in the tissue homogenised was precipitated with sulfosalicylic
acid (5%) for 1 h and then centrifuged at 13,400g for 20 min at 4 1C (deproteinated
PMS). NP-SH was determined by the method of Sedlak and Lindsay (1968) as
adopted by Parvez et al. (2003). The assay mixture contained 0.5 ml of filtered
aliquot, 2.3 ml of sodium phosphate buffer (0.1 M, pH 7.4) and 0.2 ml DTNB
(stock ¼ 100 mM in 0.1 M sodium phosphate buffer, pH 7.4) in a total volume of
3 ml. The optical density of reaction product was read immediately at 412 nm on a
spectrophotometer.
2.10. Total glutathione (GSHt) content
GSHt content was determined in the deproteinated PMS adopting the
enzymatic recycling method using GR excess, whereby the sulfhydryl group of
GSH reacts with DTNB (Ellman’s reagent) producing a yellow coloured 5-thio-2-
nitrobenzoic acid (TNB). The rate of TNB production is directly proportional to this
recycling reaction, which is in turn directly proportional to the concentration of
GSH in the sample (Tietze, 1969; Baker et al., 1990). Formation of TNB was
measured by spectrophotometry at 412 nm. It should be noted that GSSG is
converted to GSH by GR in this system, which consequently measures total
glutathione (GSHt). The results were expressed as mmol TNB formed/min/mg
protein using a molar extinction coefficient of 14.1�103 M�1 cm�1.
2.11. Metallothioneins (MT) determination
MT levels were determined after homogenisation in 20 mM Tris buffer (pH 8.6)
containing 150 mM of NaCl. The homogenates were centrifuged for 45 min at
30,000g at 4 1C. Aliquots of the supernatant were heat-treated at 80 1C for 10 min
and re-centrifuged at 30,000g for 45 min at 4 1C. MT quantification was made by
differential pulse polarography (DPP) according to the method described by
Bebianno and Langston (1989) using a 646VA Processor autolab type II and an ECO
Chemie IME663 mercury drop electrode. Comparisons of peak heights with those
of standard additions of purified rabbit MT enabled the quantification. Accuracy of
the DPP analytical method was obtained by internal calibration using commercial
rabbit liver MT-I. Recovery studies of cytosols spiked with the MT-I rabbit liver
standards revealed good recovery results ranging from 97% to 103%. Results were
expressed as mg MT/mg protein.
2.12. DNA strand breaks determination
Genotoxicity was tested using the DNA alkaline unwinding assay. The previous
stored liver portions were placed in TNES (Tris–HCl 10 mM, NaCl 125 mM, EDTA
10 mM, SDS 1%, pH 7.5)—urea (5 M) buffer with proteinase K solution (final
concentration 0.8 mg/ml). DNA isolation was performed using a genomic DNA
purification kit (Fermentas). DNA integrity measurements were performed
according to Rao et al. (1996) with minor modifications, as adopted by Maria et
al. (2002). DNA single and double strands were separated by hydroxyapatite (20%)
‘‘batch’’ elution and quantified with a DNA-specific fluorescent dye (2.5 nM
bisbenzimide) after alkaline unwinding. Data were expressed as DNA integrity (%),
determined by applying the following equation:
DNA integrity ð%Þ ¼ ½ðDouble� stranded DNAÞ=ðDouble� stranded DNAþ Single� stranded DNAÞ� � 100:
2.13. Protein measurement
Total protein contents were determined according to the Biuret method
(Gornall et al., 1949), using bovine serum albumin (E. Merck-Darmstadt, Germany)
as a standard.
2.14. Statistical analysis
Statistica software (StatSoft Inc., Tulsa, OK, USA) was used for statistical
analyses. Data analysis followed standard statistical procedures (Zar, 1999). Data
were tested for goodness of fit to normal distribution and requirements of
homogeneity of variances were also determined. Analysis of variance (ANOVA) on
ranks were performed followed by all pairwise multiple comparison procedures
(Tukey test). Differences between means were considered significant at po0.05.
The relationship between biomarkers applied to D. labrax liver was explored using
linear regression analyses. The correlation coefficient (r) was calculated and its
statistical significance (p) was determined from table of r critical values in Zar (1999).

ARTICLE IN PRESS
V.L. Maria et al. / Ecotoxicology and Environmental Safety 72 (2009) 1861–18701864
3. Results
No important variations on physico-chemical parameters werefound between the studied sites (Table 1). Temperature rangedfrom 17.9 to 19.0 1C, pH ranged from 7.7 to 8.2, dissolved oxygenranged from 5 to 6.9 and salinity ranged from 29 to 34, withexception for RIO where it was 21. Depth ranged from 1.2 (TOR
Fig. 2. Represents enzymatic protective responses CAT (A), GR (B), GST (C), GPx (D) i
(Torreira—TOR, Gafanha—GAF, Rio Novo do Prıncipe—RIO, Laranjo—LAR and Vagos—VA
common letter are statistically different between groups (po0.05).
and LAR) to 6.9 (GAF). The highest turbidity levels (0.3 m) weremeasured at LAR and GAF, whereas the other sites ranged from 0.7to 1.0 m (Table 1).
The pollutants effects on enzymatic defence system arerepresented in the Fig. 2. Thus, fish liver CAT (Fig. 2A), GR(Fig. 2B) and GST (Fig. 2C) activities at RIO, LAR and VAG weresignificantly lower when compared to the reference site (TOR) and
n liver of Dicentrarchus labrax L. collected at different sites in the Aveiro Lagoon
G). Values represent the means7S.E. (n ¼ 6). For a given enzyme, values without a
ARTICLE IN PRESS
V.L. Maria et al. / Ecotoxicology and Environmental Safety 72 (2009) 1861–1870 1865
GAF. Moreover, when compared to TOR values, 75%, 69% and 66%decreases in CAT activity were noticed in fish taken from RIO, LARand VAG, respectively. Hence, GR activity decreased 74% (RIO) and80% (LAR and VAG). The reduction observed for GST was 68% (RIOand LAR) and 77% (VAG). The liver GPx activity (Fig. 2D) wassignificantly increased in sea bass caught at LAR (106%) and VAG(232%) when compared to fish collected from TOR. Moreover, fishfrom VAG also showed significant GPx increases compared to thefish collected from RIO and GAF.
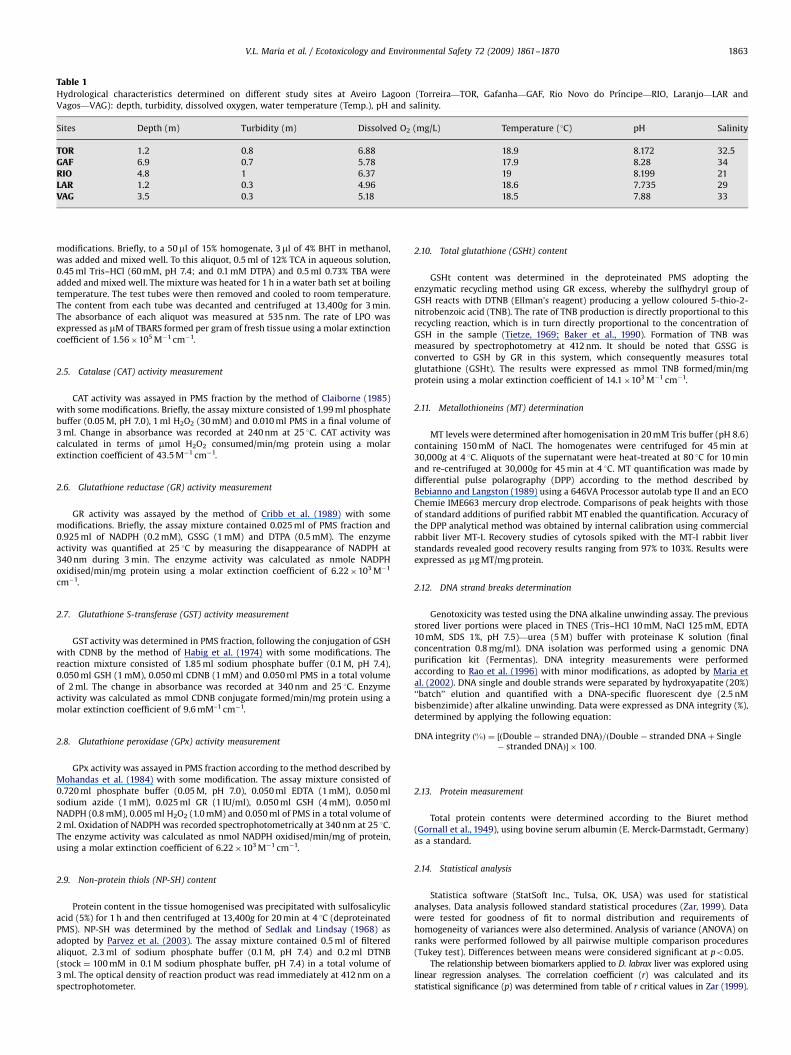
The pollutants effects on non-enzymatic antioxidants contentare described in the Fig. 3. The non-protein thiols (NP-SH) contentwas significantly lower in sea bass caught in all sampling siteswhen compared to TOR with reductions of 55% (GAF), 87% (RIO),77% (LAR) and 69% (VAG) (Fig. 3A). Significantly lower totalglutathione (GSHt) levels were also observed in fish collectedfrom RIO, LAR and VAG sites compared to TOR with reductions of70% (RIO), 63% (LAR) and 67% (VAG) (Fig. 3B). In relation to MT(Fig. 3C), a significantly higher content was observed in fishcollected from GAF and VAG when compared to TOR, representingan increment of 546% and 3078%, respectively. In addition, MTcontent observed in fish collected from VAG was significantly
Fig. 3. Represents non-enzymatic protective responses NP-SH (A), GSHt (B) and MT (C
(Torreira—TOR, Gafanha—GAF, Rio Novo do Prıncipe—RIO, Laranjo—LAR and Vagos—VA
a common letter are statistically different between groups (po0.05).
higher when compared to all the other sites. GAF fish showedhigher MT levels in fish compared to RIO and LAR.
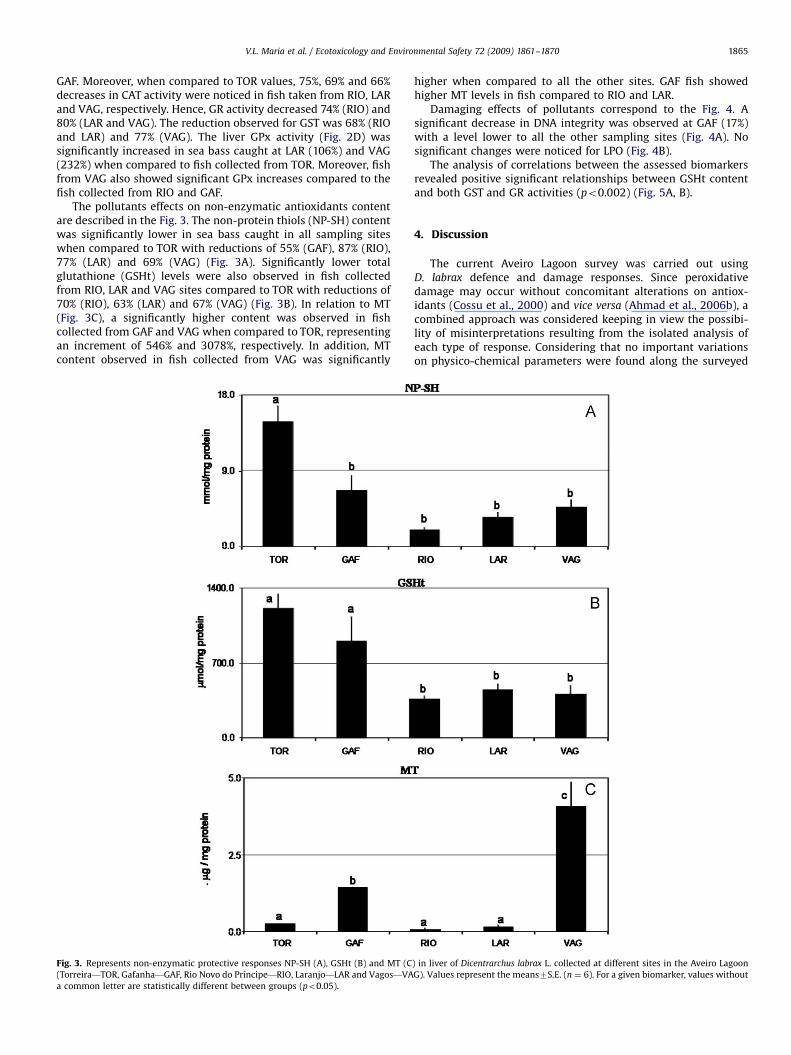
Damaging effects of pollutants correspond to the Fig. 4. Asignificant decrease in DNA integrity was observed at GAF (17%)with a level lower to all the other sampling sites (Fig. 4A). Nosignificant changes were noticed for LPO (Fig. 4B).
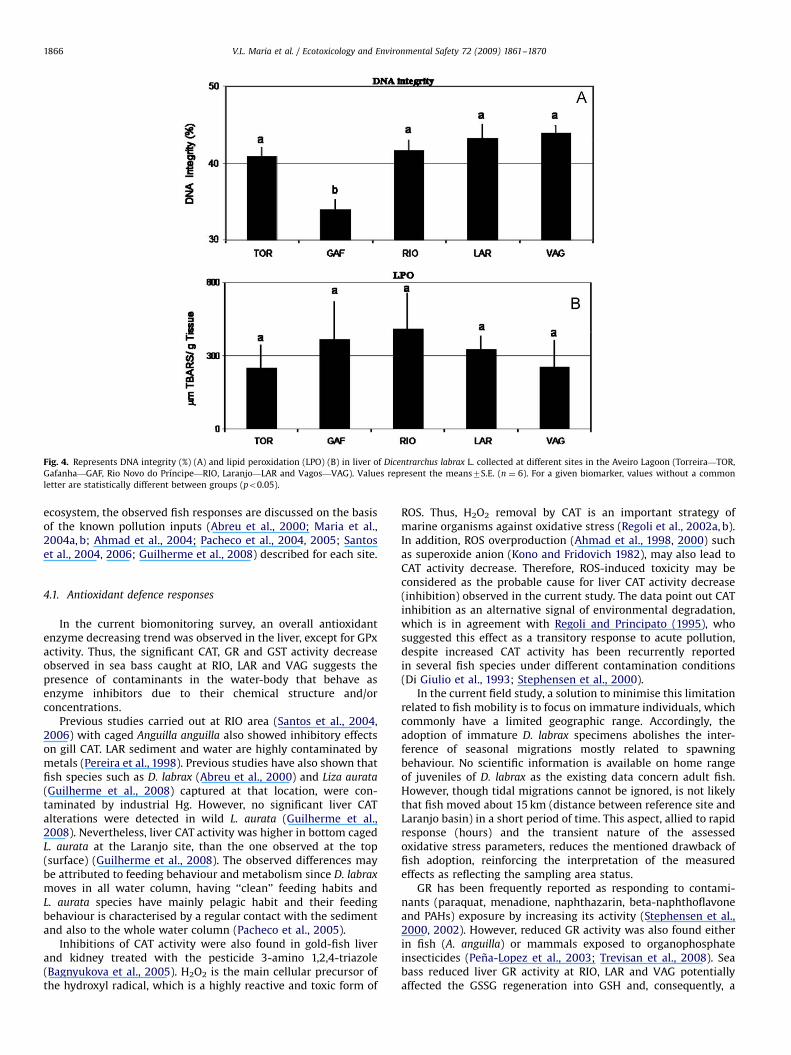
The analysis of correlations between the assessed biomarkersrevealed positive significant relationships between GSHt contentand both GST and GR activities (po0.002) (Fig. 5A, B).
4. Discussion
The current Aveiro Lagoon survey was carried out usingD. labrax defence and damage responses. Since peroxidativedamage may occur without concomitant alterations on antiox-idants (Cossu et al., 2000) and vice versa (Ahmad et al., 2006b), acombined approach was considered keeping in view the possibi-lity of misinterpretations resulting from the isolated analysis ofeach type of response. Considering that no important variationson physico-chemical parameters were found along the surveyed
) in liver of Dicentrarchus labrax L. collected at different sites in the Aveiro Lagoon
G). Values represent the means7S.E. (n ¼ 6). For a given biomarker, values without
ARTICLE IN PRESS
Fig. 4. Represents DNA integrity (%) (A) and lipid peroxidation (LPO) (B) in liver of Dicentrarchus labrax L. collected at different sites in the Aveiro Lagoon (Torreira—TOR,
Gafanha—GAF, Rio Novo do Prıncipe—RIO, Laranjo—LAR and Vagos—VAG). Values represent the means7S.E. (n ¼ 6). For a given biomarker, values without a common
letter are statistically different between groups (po0.05).
V.L. Maria et al. / Ecotoxicology and Environmental Safety 72 (2009) 1861–18701866
ecosystem, the observed fish responses are discussed on the basisof the known pollution inputs (Abreu et al., 2000; Maria et al.,2004a, b; Ahmad et al., 2004; Pacheco et al., 2004, 2005; Santoset al., 2004, 2006; Guilherme et al., 2008) described for each site.
4.1. Antioxidant defence responses
In the current biomonitoring survey, an overall antioxidantenzyme decreasing trend was observed in the liver, except for GPxactivity. Thus, the significant CAT, GR and GST activity decreaseobserved in sea bass caught at RIO, LAR and VAG suggests thepresence of contaminants in the water-body that behave asenzyme inhibitors due to their chemical structure and/orconcentrations.
Previous studies carried out at RIO area (Santos et al., 2004,2006) with caged Anguilla anguilla also showed inhibitory effectson gill CAT. LAR sediment and water are highly contaminated bymetals (Pereira et al., 1998). Previous studies have also shown thatfish species such as D. labrax (Abreu et al., 2000) and Liza aurata
(Guilherme et al., 2008) captured at that location, were con-taminated by industrial Hg. However, no significant liver CATalterations were detected in wild L. aurata (Guilherme et al.,2008). Nevertheless, liver CAT activity was higher in bottom cagedL. aurata at the Laranjo site, than the one observed at the top(surface) (Guilherme et al., 2008). The observed differences maybe attributed to feeding behaviour and metabolism since D. labrax
moves in all water column, having ‘‘clean’’ feeding habits andL. aurata species have mainly pelagic habit and their feedingbehaviour is characterised by a regular contact with the sedimentand also to the whole water column (Pacheco et al., 2005).
Inhibitions of CAT activity were also found in gold-fish liverand kidney treated with the pesticide 3-amino 1,2,4-triazole(Bagnyukova et al., 2005). H2O2 is the main cellular precursor ofthe hydroxyl radical, which is a highly reactive and toxic form of
ROS. Thus, H2O2 removal by CAT is an important strategy ofmarine organisms against oxidative stress (Regoli et al., 2002a, b).In addition, ROS overproduction (Ahmad et al., 1998, 2000) suchas superoxide anion (Kono and Fridovich 1982), may also lead toCAT activity decrease. Therefore, ROS-induced toxicity may beconsidered as the probable cause for liver CAT activity decrease(inhibition) observed in the current study. The data point out CATinhibition as an alternative signal of environmental degradation,which is in agreement with Regoli and Principato (1995), whosuggested this effect as a transitory response to acute pollution,despite increased CAT activity has been recurrently reportedin several fish species under different contamination conditions(Di Giulio et al., 1993; Stephensen et al., 2000).
In the current field study, a solution to minimise this limitationrelated to fish mobility is to focus on immature individuals, whichcommonly have a limited geographic range. Accordingly, theadoption of immature D. labrax specimens abolishes the inter-ference of seasonal migrations mostly related to spawningbehaviour. No scientific information is available on home rangeof juveniles of D. labrax as the existing data concern adult fish.However, though tidal migrations cannot be ignored, is not likelythat fish moved about 15 km (distance between reference site andLaranjo basin) in a short period of time. This aspect, allied to rapidresponse (hours) and the transient nature of the assessedoxidative stress parameters, reduces the mentioned drawback offish adoption, reinforcing the interpretation of the measuredeffects as reflecting the sampling area status.
GR has been frequently reported as responding to contami-nants (paraquat, menadione, naphthazarin, beta-naphthoflavoneand PAHs) exposure by increasing its activity (Stephensen et al.,2000, 2002). However, reduced GR activity was also found eitherin fish (A. anguilla) or mammals exposed to organophosphateinsecticides (Pena-Lopez et al., 2003; Trevisan et al., 2008). Seabass reduced liver GR activity at RIO, LAR and VAG potentiallyaffected the GSSG regeneration into GSH and, consequently, a
ARTICLE IN PRESS
Fig. 5. Correlation between GST and GSHt (A) as well as GSHt and GR (B) in liver of Dicentrarchus labrax. Statistical significance and correlation coefficient are represented
by p and r, respectively.
V.L. Maria et al. / Ecotoxicology and Environmental Safety 72 (2009) 1861–1870 1867
diminished antioxidant defence capacity is expected. The positiveand significant correlation found between GR and GSHt indicatesthat the increase in the last parameter also represented anincrease in the content of oxidised form and thus, requiring higherGR activity.
Detoxification enzymes, such as GST, help in eliminatingreactive compounds by forming conjugates with glutathione andsubsequently eliminating them as mercapturic acid, therebyprotecting cells against ROS-induced damage (Rodriguez-Arizaet al., 1991). Although GST induction has been widely demon-strated following exposure to some organic contaminants (P�erez-Lopez et al., 2000, 2002), its inhibition has also been reported infish either as a specific response to endosulfan (Ballesterosa et al.,2009) or as a non-specific response to chemical challenge (Regoliet al., 2003). The current reduction observed in GST activity couldnot be dissociated from the concomitant reduction observed inthe GSHt (and NP-SH) levels, which is corroborated by thesignificant correlation found. Santos et al. (2004, 2006) alsodemonstrated gill GST inhibition and GSH content reduction inA. anguilla caged at RIO site in Aveiro Lagoon. However, nosignificant alterations in the GST activity were observed byGuilherme et al. (2008) in LAR site using wild and caged L. aurata
specimens. Another non-exclusive option that also explains thelowering of GST may be related to gene expression. The migrationof the nuclear transcription factor erythroid 2p45 (NF-E2)-relatedfactor 2 (NRf2) is the marker for the expression of important genesto the antioxidant defence, including those coding for the catalytic
subunit of g-glutamate cysteine ligase (gGCL) and several GSTisoenzymes (Lee and Surh, 2005). Hence, is important to identifywhich are the chemopreventive agents present in the studiedAveiro Lagoon sites, since some recognised chemicals can induceNRf2, namely oltipraz, sulforaphane, curcumin and b-naphtho-flavone (Lee and Surh, 2005).
In the present research, liver GPx increased activity at LAR andVAG indicates its involvement as an antioxidant defence andsensitivity towards reactive contaminants. This induction can belinked to the main classes of contaminants previously detected inthose areas, i.e. PAHs for VAG (Pacheco et al., 2005) and metals,namely mercury, for LAR (Pereira et al., 1998), which isresponsible for the increased production of H2O2. In spite of theobserved GPx induction, we may not exclude the possibility of alimited increased activity by low GSH availability. Moreover, thislimitation could have played a determinant role in GPx responseat RIO. Thus, the hypothesis concerning the enzymes GPx and alsoGR that have different susceptibilities towards inhibitory effectmay arise. The presence of mixtures of chemicals in Aveiro Lagoonwater may affect the crucial role of GPx protection and therefore,current GPx responses reflect changes on the GPx inhibitors/inducers balance along the lagoon favouring the later. Consideringthe GPx and CAT similarity in their final action towards H2O2
degradation, the occurrence of CAT decrease gives to GPx anaugmented protective importance. Moreover, a significant GPxincrease along-with the decrease of other antioxidants iscorroborated by Ahmad et al. (2006b) and may reflect the

ARTICLE IN PRESS
V.L. Maria et al. / Ecotoxicology and Environmental Safety 72 (2009) 1861–18701868
enzyme-inducing effect of H2O2, as suggested by Aksnes and Njaa(1981).
In the current study, the combined analysis of CAT, GR, GSTreduction and GPx increased activity indicates a complexcontamination scenario. The overall liver enzyme antioxidantsresponses may be due to a balance between inhibitors andinducers present at those Aveiro Lagoon sites. However, a differentGPx sensitivity should be considered since CAT, GR and GST wereinhibited.
Under laboratory conditions, fish exposed to a variety ofchemical compounds (buthionine sulfoximine, diethyl maleate,herbicide molinate, fungicide prochloraz, chromium (Cr) sufferedGSH depletion, which is considered as a biomarker of environ-mental stress (Gallagher et al., 1992; Pena-Lopez et al., 2001;Ahmad et al., 2006a; Sanchez et al., 2007). Almar et al. (1998) alsoobserved GSH depletion in the freshwater fish Gobio gobio andRutilus arcasii caught downstream and upstream of severalchemical industries located at river Bernesga (Spain). In thepresent work, the significant decrease in NP-SH and GSHtcontents observed at RIO, LAR and VAG sea bass, may be explainedby the inhibition of the enzymes involved in glutathionesynthesises. Considering also the inhibition observed for CAT,GR and GST, the inhibition of the glutathione synthesises enzymes(g-glutamylcysteine synthetase and GSH synthetase) must behypothesised as a cause of the impediment in the GSH reposition.Another possible explanation can be related to NRf2 alterationand consequently gene expression, as was previously mentioned.
Glutathione represents more than 90% of the cells NP-SHcontent. Nevertheless, it was demonstrated that other thiolsbesides GSH, namely N-acetyl-L-cysteine (NAC), can also play animportant role as antioxidant (Pena-Lopez et al., 2003). Therefore,it is recommendable the joint analysis of both NP-SH and GSHt inorder to improve the knowledge on non-enzymatic antioxidantresponse. Considering all the Aveiro Lagoon sites, the currentresults showed similar patterns for these two parameters,suggesting that glutathione represents the main NP-SH in fishfrom the RIO, LAR and VAG. Despite no alterations in GSHt levelshave been found at GAF, the involvement of other thiols issupported by the significant NP-SH decrease observed. This maybe explained by the complex regulation of glutathione homo-eostasis. An alteration in the consuming and regenerating GSHpathways may be hindered by de novo synthesis of the tripeptide.As changes in glutathione status may be transient, in order todetect environmental stressors more accurately, GSH should bemeasured in parallel with glutathione peroxidases and reductase,which are involved in its turnover (Cossu et al., 2000).
An overall interpretation of the GSHt and NP-SH declineconcomitantly with the GST, GR and CAT activities inhibitionobserved in the RIO, LAR and VAG, revealed a potential risk to fishinhabiting those locations, due to antioxidant defence decreasedefficiency. Therefore, the measurement of a single antioxidanteffect is not enough to define a complete antioxidant defenceresponse under experimental/laboratory and field conditions(Zanette et al., 2006).
The majority of research concerning MT in bivalves and fishindicates that its synthesis can be induced by exposure to metals,such as Cd, Cr, Cu, Zn, Hg and consequently they have been used asspecific biomarkers of metal contamination (Bebianno andLangston, 1989; Fernandes et al., 2007). In the present field study,MT elevation was only observed at GAF and VAG sites. Aspreviously referred, these sites did not seem to be contaminatedby heavy metals. GAF is a harbour area affected by moderate levelsof PAHs (Pacheco et al., 2005) and high levels of organometallicssuch as tributyltin (TBT) (Sousa et al., 2007). On the other hand,the MT elevation observed at VAG, an area also marked by non-metallic contamination, should not be considered as surprising
since non-metals, such as fungicides and herbicides have alreadybeen shown as causative agents for MT induction in aquaticorganisms (Davis and Cousins 2000). As an example, Paris-Palacios et al. (2003) observed increased hepatic MT levels inRutilus rutilus exposed to the fungicide procymidone. Similarly,Mosleh et al. (2004, 2005a, b) also found that the exposure ofTubifex tubifex to the herbicide isoproturon or the fungicidefenhexamid increased MT levels. On opposite, the absence of MTinduction in LAR sea bass was not expected considering Hgcontamination in that location. A low residence time caused byhydrological changes in that area may explain the unchanged MTlevels. MT response represents a long-lasting adaptation and theirinduction requires longer exposures (days) to inducers and,thereby, longer permanence in each surveyed area. In this case,tidal migrations could assume greater importance and difficultthe data interpretation. However, though tidal migrations cannotbe ignored, is not likely that juvenile sea bass moved approxi-mately 20 or 25 km from Laranjo (area highly contaminated withmetals) to Gafanha or Vagos, respectively.
D. labrax feeding habits and its mobility through entire watercolumn must also be considered for MT results. Therefore, wehypothesised that the MT induction observed in fish captured atGafanha and Vagos is attributable to non-metallic contaminants.
4.2. Damaging effects of pollutants
In the present field survey, the lowest liver DNA integrity wasfound in sea bass caught at GAF, suggesting the presence of DNAdamaging compounds in the water and sediments. Previously,European eel (A. anguilla) in situ caging exposure, carried out in2001, demonstrated liver DNA integrity loss at the same site (GAF)(Maria et al., 2004a). Thus, genotoxic and/or pro-genotoxiccompounds present in GAF were able to induce early (8 hexposure) DNA strand breaks in eel’s blood cells and a delayed(48 h exposure) response in liver and kidney. However, in othersurveys concerning erythrocytic nuclear abnormalities (ENAs), noDNA alterations were observed in Atherina presbyter (Pacheco etal., 2004) and L. aurata (Pacheco et al., 2005) captured at GAF.Considering the results of Maria et al. (2004a) as a first signal ofthe genotoxicants presence at that particular site, current resultsdefinitely demonstrated the risk for wild fish population. Theseparticular results at GAF agree with a continued degradation ofenvironmental water quality in the last half decade (from 2000 to2005). As previously referred, studies at GAF site detectedmoderate levels of PAHs and high levels of TBT (Pacheco et al.,2004, 2005; Sousa et al., 2007). Overall, the observed genotoxicresponse should be attributed to the mixture of different classes ofcontaminants rather than to single chemical. Accordingly, Micaelet al. (2007) reported that chronic exposure to binary mixtures oflow levels of TBT plus ethinylestradiol or TPT (triphenyltin) plusethinylestradiol were genotoxic to zebrafish. The genotoxicityinduction at GAF was hypothesised to be related to a failure in thedetoxification process where some unconjugated reactive meta-bolites may damage DNA (Maria et al., 2004a).
Unexpectedly, no statistical differences in liver LPO measure-ments were observed, suggesting that fish were able to deal withthe pollution stress, despite the depletion of several antioxidantdefences at RIO, LAR and VAG. This is in agreement with Cossuet al. (2000), who stated that LPO increase cannot be predictedonly on the basis of antioxidant depletion. Furthermore, Ferreira-Cravo et al. (2007) demonstrated that the low antioxidantcompetence against HOd radicals was not concomitant with highLPO values in estuarine polychaeta Laeonereis acuta sampled froma contaminated site, suggesting that other macromolecules couldbe affected by the higher ROS levels.

ARTICLE IN PRESS
V.L. Maria et al. / Ecotoxicology and Environmental Safety 72 (2009) 1861–1870 1869
Liver damaging effects resulting from enzymatic and non-enzymatic antioxidants depletion at RIO, LAR and VAG were notconfirmed considering the LPO and DNA integrity responses.However, the low DNA integrity level and the absence ofantioxidant inhibition effect (excluding the NP-SH) observed inGAF seems to indicate that the genotoxic response may be relatedwith direct genotoxic action rather than to ROS overproduction.
5. Conclusion
On the basis of wild juvenile D. labrax responses concerningenzymatic (CAT, GR and GST activities) and non-enzymatic (GSHt,NP-SH) antioxidant defences decrease at RIO, LAR and VAG, theAveiro Lagoon water contamination by xenobiotics was demon-strated at those particular sites. However, DNA integrity lossmeasured as a damage indicator signalled GAF as the mostimpacted site. Thus, the efficacy of a combined strategy encom-passing sets of defence and damage indicators was demonstratedsince it was confirmed that the evaluation of only one of theseaspects may be misleading. Despite the consensually assumedantioxidant induction as a sign of exposure to contaminants, itwas demonstrated that the decrease in antioxidant responsesmust also be considered in order to achieve a more efficientenvironmental health assessment.
Conflict of interest statement
The authors declare that there are no conflicts of interest.
Acknowledgments
The financial supports from FCT (Foundation for the Scienceand Technology), POCI/MAR/61195/2004, SFRH/BPD/26970/2006,SFRH/BPD/34326/2006, SFRH/BD/27584/2006, by the Aveiro Uni-versity Research Institute/CESAM and CIMA & Faculty of Marineand Environmental Sciences, University of Algarve are gratefullyacknowledged.
References
Abreu, S.N., Pereira, E., Vale, C., Duarte, A.C., 2000. Accumulation of Mercury in SeaBass from a Contaminated Lagoon (Ria de Aveiro, Portugal). Mar. Pollut. Bull.40, 293–297.
Ahmad, I., Fatima, M., Athar, M., Khan, N.Z., Raisuddin, S., 1998. Responses ofcirculating fish phagocytes to paper mill effluent exposure. Bull. Environ.Contam. Toxicol. 61, 746–753.
Ahmad, I., Hamid, T., Fatima, M., Chand, H.S., Jain, S.K., Athar, M., Raisuddin, S.,2000. Induction of hepatic antioxidants in freshwater fish (Channa punctatusB.) is a biomarker of paper mill effluent exposure. Biochim. Biophys. Acta. 1523,37–48.
Ahmad, I., Maria, V.L., Oliveira, M., Santos, M.A., 2006a. Anguilla anguilla L. gill andkidney oxidative stress and genotoxic response to chromium exposure withand without b-naphthoflavone—pre-exposure. Mutat. Res. 608, 16–28.
Ahmad, I., Pacheco, M., Santos, M.A., 2006b. Anguilla anguilla L. oxidative stressbiomarkers: An in situ study of freshwater wetland ecosystem (Pateira deFermentelos, Portugal). Chemosphere 65, 952–962.
Ahmad, I., Pacheco, M., Santos, M.A., 2004. Enzymatic and non-enzymaticantioxidants as an adaptation to phagocyte-induced damage in Anguillaanguilla L. following in situ harbor water exposure. Ecotoxicol. Environ. Saf.57, 290–302.
Aksnes, A., Njaa, L.R., 1981. Catalase, glutathione peroxidase and superoxidedismutase in different fish species. Comp. Biochem. Physiol. 69B, 893–896.
Almar, M., Otero, L., Santos, C., Gallego, J., 1998. Liver glutathione content andglutathione-dependent enzymes of two species of freshwater fish asbioindicators of chemical pollution. J. Environ. Sci. Health B 33 (6), 769–783.
Amado, L.L., Robaldo, R.B., Geracitano, L., Monserrat, J.M., Bianchini, A., 2006.Biomarkers of exposure and effect in the Brazilian flounder Paralichthysorbignyanus (Teleostei: Paralichthyidae) from the Patos Lagoon estuary(Southern Brazil). Mar. Pollut. Bull. 52 (2), 207–213.
Bagnyukova, T., Storey, K.B., Lushchak, V.I., 2005. Adaptive response of antioxidantenzymes to catalase inhibition by aminotriazole in goldfish liver and kidney.Comp. Biochem. Phys. B 142, 335–341.
Baker, M.A., Cerniglia, G.J., Zaman, A., 1990. Microtiter plate assay for themeasurement of glutathione and glutathione disulfide in large numbers ofbiological samples. Anal. Biochem. 190, 360–365.
Ballesterosa, M.L., Wunderlinb, D.A., Bistonia, M.A., 2009. Oxidative stressresponses in different organs of Jenynsia multidentata exposed to endosulfan.Ecotoxicol. Environ. Saf. 72 (1), 199–205.
Bebianno, M.J., Langston, W.J., 1989. Quantification of metallothioneins in marineinvertebrates using differential pulse polarography. Portugaliae Electrochim.Acta 7, 511–524.
Bebianno, M.J., Santos, C., Can�ario, J., Gouveia, N., Sena-Carvalho, D., Vale, C., 2007.Hg and metallothionein-like proteins in the black scabbardfish Aphanopuscarbo. Food Chem. Toxicol. 45, 1443–1452.
Bird, R.P., Draper, A.H., 1984. Comparative studies on different methods ofmalondyhaldehyde determination. Methods Enzymol. 90, 105–110.
Bucheli, T.D., Fent, K., 1995. Induction of cytochrome-450 as a biomarker forenvironmental contamination in aquatic ecosystems. Crit. Rev. Environ. Sci.Technol. 25, 201–268.
Claiborne, A., 1985. Catalase activity. In: Greenwald, RA (Ed.), CRC Handbook ofMethods in Oxygen Radical Research. CRC Press, Boca Raton, FL, pp. 283–284.
Cossu, C., Doyotte, A., Babut, M., Exinger, A., Vasseur, P., 2000. Antioxidantbiomarkers in freshwater bivalves, Unio tumidus, in response to differentcontamination profiles of aquatic sediments. Ecotoxicol. Environ. Saf. 45,106–121.
Cribb, A.E., Leeder, J.S., Spielberg, S.P., 1989. Use of a microplate reader in an assayof glutathione reductase using 5,50-dithiobis(2-nitrobenzoic acid). Anal.Biochem. 183, 195–196.
Davis, S.R., Cousins, R., 2000. Metallothionein expression in animals: a physiolo-gical perspective on function. RANS 130 (5), 1085–1088.
Giulio, R.T., Habig, C., Gallagher, E.P., 1993. Effects of black rock harbour sedimentson indices of biotransformation, oxidative stress and DNA integrity in channelfish. Aquat. Toxicol. 26, 1–22.
Escobar, J.A., Rubio, M.A., Lissi, E.A., 1996. SOD and catalase inactivation by singletoxygen and peroxyl radicals. Free Radical. Bio. Med. 20 (3), 285–290.
Fernandes, D., Porte, C., Bebianno, M.J., 2007. Chemical residues and biochemicalresponses in wild and cultured European sea bass (Dicentrarchus labrax L.).Environ. Res. 103, 247–256.
Ferreira-Cravo, M., Piedras, F.R., Moraes, T.B., Ferreira, J.L.R., Freitas, D., Machado,M., Geracitano, L., Monserrat, J.M., 2007. Antioxidant responses and reactiveoxygen species generation in different body regions of the estuarinepolychaeta Laeonereis acuta (Nereididae). Chemosphere 66, 1367–1374.
Fridovich, I., 1997. Superoxide anion radical, superoxide dismutases and relatedmatters. J. Biol. Chem. 272, 18515–18517.
Gallagher, E.P., Hasspieler, B.M., Giulio, R.T., 1992. Effects of buthionine sulfoximineand diethyl maleate on glutathione turnover in the channel catfish. Biochem.Pharmacol. 43, 2209–2215.
Gigu�ere, A., Campbell, P.G.C., Hare, L., Couture, P., 2006. Sub-cellular partitioning ofcadmium, copper, nickel and zinc in indigenous yellow perch (Perca flavescens)sampled along a polymetallic gradient. Aquat. Toxicol. 77, 178–189.
Gornall, A.C., Bardawill, C.J., David, M.M., 1949. Determination of serum proteins bymeans of the biuret reaction. J. Biol. Chem. 177, 751–766.
Gravato, C., Santos, M.A., 2003. Dicentrarchus labrax biotransformation andgenotoxicity responses after exposure to a secondary treated industrial/urbaneffluent. Ecotoxicol. Environ. Saf. 55 (3), 300–306.
Guilherme, S., V�alega, M., Pereira, M.E., Santos, M.A., Pacheco, M., 2008.Antioxidant and biotransformation responses in Liza aurata under environ-mental mercury exposure—relationship with mercury accumulation andimplications for public health. Mar. Pollut. Bull. 56, 845–859.
Habig, W.H., Pabst, M.J., Jakoby, W.B., 1974. Glutathione-S-transferases, the firstenzymatic step in mercapturic acid formation. J. Biol. Chem. 249, 7130–7139.
Harris, E.D., 1992. Regulation of antioxidant enzymes. FASEB J. 6 (9), 2675–2683.Kono, Y., Fridovich, I., 1982. Superoxide radical inhibit catalase. J. Biol. Chem. 257,
5751–5754.Lee, J.S., Surh, Y.J., 2005. Nrf2 as a novel molecular target for chemoprevention.
Cancer Lett. 224, 171–184.Li, Wing-Han, Chan, P.C.Y., Chan, K.M., 2004. Metal uptake in zebrafish embryo-
larvae exposed to metal-contaminated sediments. Mar. Environ. Res. 58,829–832.
Livingstone, D.R., 2001. Contaminant stimulated reactive oxygen species produc-tion and oxidative damage in aquatic organisms. Mar. Pollut. Bull. 8, 656–666.
Maria, V.L., Ahmad, I., Santos, M.A., 2008. Juvenile sea bass (Dicentrarchus labrax L.)DNA strand breaks and lipid peroxidation response following 17beta-estradioltwo mode of exposures. Environ. Int. 34 (1), 23–29.
Maria, V.L., Pacheco, M., Santos, M.A., 2006. Anguilla anguilla L. genotoxic responsesafter in situ exposure to freshwater wetland (Pateira de Fermentelos, Portugal).Environ. Int. 32, 510–515.
Maria, V.L., Correia, A.C., Santos, M.A., 2004a. Genotoxic and biochemical responsesin caged eel (Anguilla anguilla L.) after short-term exposure to harbour waters.Environ. Int. 29, 923–929.
Maria, V.L., Teles, M., Pacheco, M., Correia, A., Santos, M.A., 2004b. Biomarkerresponses in polluted river: possible effects of paper mill contaminants oncaged Anguilla anguilla L. Fresen Environ. Bull. 13 (4), 317–325.
Maria, V.L., Correia, A.C., Santos, M.A., 2002. Anguilla anguilla L. biochemical andgenotoxic responses to benzo(a)pyrene. Ecotoxicol. Environ. Saf. 53, 86–92.

ARTICLE IN PRESS
V.L. Maria et al. / Ecotoxicology and Environmental Safety 72 (2009) 1861–18701870
Micael, J., Reis-Henriques, M.A., Carvalho, A.P., Santos, M.M., 2007. Genotoxiceffects of binary mixtures of xenoandrogens (tributyltin, triphenyltin) and axenoestrogen (ethinylestradiol) in a partial life-cycle test with Zebrafish (Daniorerio). Environ. Int. 33, 1035–1039.
Mitchelmore, C.L., Chipman, J.K., 1998. Detection of DNA strand breaks in browntrout (Salmo trutta) hepatocytes and blood cells using the single cell gelelectrophoresis (comet) assay. Aquat. Toxicol. 41, 161–182.
Mohandas, J., Marshall, J.J., Duggins, G.G., Horvath, J.S., Tiller, D., 1984. Differentialdistribution of glutathione and glutathione related enzymes in rabbit kidney.Possible implications in analgesic neuropathy. Cancer Res. 44, 5086–5091.
Mosleh, Y.Y., Paris-Palacios, S., Couderchet, M., Biagianti-Risbourg, S., Vernet, G.,2005a. Metallothionein induction, antioxidative responses, glycogen andgrowth changes in Tubifex tubifex (oligochaete) exposed to the fungicide,fenhexamid. Environ. Pollut. 135, 73–82.
Mosleh, Y.Y., Paris-Palacios, S., Couderchet, M., Biagianti-Risbourg, S., Vernet, G.,2005b. Effects of the herbicide isoproturon on metallothioneins, growth andantioxidativedefenses in the aquatic worm Tubifex tubifex (Oligochaeta,Tubificidae). Ecotoxicology 14, 1–13.
Mosleh, Y.Y., Paris-Palacios, S., Arnoult, F., Couderchet, M., Vernet, G., 2004.Metallothionein induction in the aquatic Oligochaere Tubifex tubifex exposed tothe herbicide isoproturon. Environ. Toxicol. 19, 88–93.
Ohkawa, H., Ohishi, N., Yagi, K., 1979. Assay for lipid peroxides in animal tissues bythiobarbituric acid reaction. Anal. Biochem. 95, 351–358.
Pacheco, M., Santos, M.A., Teles, M., Oliveira, M., Rebelo, J.E., Pombo, L., 2005.Biotranformation and genotoxic biomarkers in mullet species (Liza sp.) from acontaminated coastal lagoon (Ria de Aveiro, Portugal). Environ. Monit. Assess.107, 133–153.
Pacheco, M., Santos, M.A., Teles, M., Oliveira, M., Lopes, C., Rebelo, J.E., 2004.Assessment of biotransformation and genotoxicity biomarkers in feral sand-smelt (Atherina presbyter) collected in a contaminated coastal lagoon (Ria deAveiro, Portugal). Fres. Environ. Bull. 13 (12a), 1433–1438.
Pacheco, M., Santos, M.A., 2002. Biotransformation, genotoxic, and histopatholo-gical effects of environmental contaminants in European eel (Anguilla anguillaL.). Ecotoxicol. Environ. Saf. 53, 331–347.
Paris-Palacios, S., Biagianti-Risbourg, S., Vernet, G., 2003. Metallothionein induc-tion related to structural perturbations and antioxidative defences in liver ofroach (Rutilus rutilus) exposed to the fungicide procymidone. Biomarkers 8,128–141.
Parvez, S., Sayeed, I., Pandey, S., Ahmad, A., Hafeez, B., Haque, R., Ahmad, I.,Raisuddin, S., 2003. Regulatory role of copper on non-enzymatic antioxidantsin freshwater fish Channa punctatus (Bloch.). Biol. Trace Elem. Res. 93,237–248.
Pena-Lopez, S., Ferrando, M.D., Pena, J.B., 2003. Increased recovery of brainacetylcholinesterase activity in dichlorvos-intoxicated. Dis. Aquat. Organ. 55(3), 237–245.
Pena-Lopez, S., Pena, J.B., Sancho, E., Fernandez-Veja, C., Ferrando, M.D., 2001.Glutathione-dependent resistance of the European eel Anguilla anguilla to theherbicide molinate. Chemosphere 45, 671–681.
Pereira, M.E., Duarte, A.C., Millward, G.E., Valec, C., Abreu, S.N., 1998. Tidal export ofparticulate mercury from the most contaminated area of Aveiro’s Lagoon,Portugal. Sci. Total Environ. 213, 157–163.
P�erez-Lopez, M., Novoa Valinas, M.C., Melgar Riol, M.J., 2002. Induction of cytosolicGlutathione S-transferases from Atlantic eel (Anguilla anguilla) after intraper-itoneal treatment with polychlorinated biphenyls. Sci. Total Environ. 297 (1–3),141–151.
P�erez-Lopez, M., Anglade, P., Bec-Fert�e, M., Debrauwer, L., Perdu, E., Cravedi, J.P.,Rouimi, P., 2000. Characterization of hepatic and extrahepatic glutathione S-transferases in rainbow trout (Oncorhynchus mykiss) and their induction by3,39,4,49-tetrachlorobiphenyl. Fish Physiol. Biochem. 22, 21–32.
Rao, S.S., Neheli, T.A., Carey, J.H., Herbert, A., Hansen, P.D., 1996. DNA alkalineunwinding assay for monitoring the impact of environmental genotoxins.Environ. Toxicol. Water Qual. 11, 351–354.
Regoli, F., Frenzillib, G., Bocchettia, R., Annarummab, F., Scarcellib, V., Fattorinia, D.,Nigro, M., 2004. Time-course variations of oxyradical metabolism, DNA
integrity and lysosomal stability in mussels, Mytilus galloprovincialis, duringa field translocation experiment. Aquat. Toxicol. 68, 167–178.
Regoli, F., Winston, G.W., Gorbi, S., Frenzilli, G., Nigro, M., Corsi, I., Focardi, S., 2003.Integrating enzymatic responses to organic chemical exposure with totaloxyradical absorbing capacity and DNA damage in the European eel Anguillaanguilla. Environ. Toxicol. Chem. 22, 2120–2129.
Regoli, F., Gorbi, S., Frenzilli, G., Nigro, M., Corsi, I., Focardi, S., Winston, G.W.,2002a. Oxidative stress in ecotoxicology: from the analysis of individualantioxidants to a more integrated approach. Mar. Environ. Res. 54, 419–423.
Regoli, F., Pellegrini, D., Winston, G.W., Gorbi, S., Giuliani, S., Virno- Lamberti, C.,Bompadre, S., 2002b. Application of biomarkers for assessing the biologicalimpact of dredged materials in the Mediterranean: the relationship betweenantioxidant responses and susceptibility to oxidative stress in the red mullet(Mullus barbatus). Mar. Pollut. Bull. 44, 912–922.
Regoli, F., Principato, G., 1995. Gluatathione, glutathione-dependent and antiox-idant enzymes in mussel, Mytilus galloprovincialis, exposed to metal under fieldand laboratory conditions: implications for the use of biochemical biomarkers.Aquat. Toxicol. 31, 143–164.
Rodriguez-Ariza, A., Dorado, G., Peinado, J., Pueyo, C., Lopez-Barea, J., 1991.Biochemical effects of environmental pollution in fishes from Spanish south-Atlantic littoral. Biochem. Soc. Trans. 19 (3), 301S.
Sanchez, W., Aıt-Aıssa, S., Palluel, O., Ditche, J.-M., Porcher, J.-M., 2007. Preliminaryinvestigation of multi-biomarker responses in three-spined stickleback(Gasterosteus aculeatus L.) sampled in contaminated streams. Ecotoxicology16 (2), 279–287.
Santos, M.A., Pacheco, M., Ahmad, I., 2004. Anguilla anguilla L. antioxidantsresponses to in situ bleached kraft pulp mill effluent outlet exposure. Environ.Int. 30 (3), 301–308.
Santos, M.A., Pacheco, M., Ahmad, I., 2006. Responses of European eel (Anguillaanguilla L.) circulating phagocytes to an in situ closed pulp mill effluentexposure and its association with organ-specific peroxidative damage. Chemo-sphere 63, 794–801.
Sedlak, J., Lindsay, H.R., 1968. Estimation of total, protein-bound and nonproteinsulfhydryl groups in tissues with Ellman’s Reagent. Anal. Biochem. 25, 192–205.
Shailaja, M.S., Rajamanickam, R., Wahidulla, S., 2006. Increased formation ofcarcinogenic PAH metabolites in fish promoted by nitrite. Environ. Pollut. 143,174–177.
Sousa, A., Matsudaira, C., Takahashi, S., Tanabe, S., Barroso, C., 2007. Integrativeassessment of organotin contamination in a southern European estuarinesystem (Ria de Aveiro, NW Portugal): tracking temporal trends in order toevaluate the effectiveness of the EU ban. Mar. Poll. Bull. 54, 1645–1653.
Stephensen, E., Sturve, J., Forlin, L., 2002. Effects of redox cycling compounds onglutathione content and activity of glutathione-related enzymes in rainbowtrout liver. Comp. Biochem. Physiol. C 133 (3), 435–442.
Stephensen, E., Svavarsson, J., Sturve, J., Ericson, G., Adolfsson-Erici, M., Forlin, L.,2000. Biochemical indicators of pollution exposure in shorthorn sculpin(Myoxocephalus scorpius), caught in four harbours on the southwest coast ofIceland. Aquat. Toxicol. 48, 431–442.
Teles, M., Carrillo, M., Pacheco, M., Santos, M.A., 2007. Vitellogenin Induction inJuvenile Dicentrarchus labrax L. treated with 17 beta-Estradiol (E2)—relationwith exposure route and E2 plasmatic levels. Fres. Environ. Bull. 16 (5),495–499.
Teles, M., Pacheco, M., Santos, M.A., 2006. Biotransformation, stress and genotoxiceffects of 17 beta-estradiol in juvenile sea bass (Dicentrarchus labrax L.).Environ. Int. 32 (4), 470–477.
Tietze, F., 1969. Enzymatic method for quantitative determination of nanogramamounts of total and oxidized glutathione. Anal. Biochem. 27, 502–522.
Trevisan, R., Uliano-Silva, M., Pandolfo, P., Franco, J.L., Brocardo, P.S., Santos, A.R.S.,Farina, M., Rodrigues, A.L.S., Takahashi, R.N., Dafre, A.L., 2008. Antioxidant andacetylcholinesterase response to repeated malathion exposure in rat cerebralcortex and hippocampus. Basic Clin. Pharmacol. 102 (4), 365–369.
Zanette, J., Monserrat, J.M., Bianchini, A., 2006. Biochemical biomarkers in gills ofmangrove oyster Crassostrea rhizophorae from three Brazilian estuaries. Comp.Biochem. Physio1. C 143, 187–195.
Zar, J.H., 1999. In: Biostatistical Analysis fourth ed. Prentice-Hall International Inc., USA.




















