
DNA damage and lipid peroxidation vs. protection responses in the gill of Dicentrarchus labrax L....
10
DNA damage and lipid peroxidation vs. protection responses in the gill of Dicentrarchus labrax L. from a contaminated coastal lagoon (Ria de Aveiro, Portugal) I. Ahmad a, ⁎ , V.L. Maria b , M. Oliveira a , A. Serafim b , M.J. Bebianno b , M. Pacheco a , M.A. Santos a a CESAM & Department of Biology, University of Aveiro, 3810-193 Aveiro, Portugal b CIMA & Faculty of Marine and Environmental Sciences, University of Algarve, 8005-139 Faro, Portugal ARTICLE INFO ABSTRACT Article history: Received 19 February 2008 Received in revised form 12 June 2008 Accepted 16 June 2008 Available online 21 July 2008 The present research work aimed to investigate the damage vs. protection responses in gill of European sea bass (Dicentrarchus labrax) captured at a polluted coastal lagoon, Ria de Aveiro (Portugal), as a tool to evaluate the human impacts on environmental health. Damage was assessed as DNA strand breakage and lipid peroxidation (LPO) whereas protection was evaluated by measuring catalase (CAT), glutathione peroxidase (GPX), glutathione reductase (GR), glutathione S-transferase (GST), total glutathione (GSHt), thiols and metallothioneins (MT). Fish were caught at five locations: Gafanha (GAF), Rio Novo do Príncipe (RIO), Laranjo (LAR) and Vagos (VAG) presenting each a different recognized source of contamination, and Torreira (TOR), assumed as reference site. Among the surveyed sites, gill damage was observed only at GAF, as measured by DNA integrity loss and LPO increase. An overall induction in enzymatic antioxidant protection was perceptible in fish from GAF and VAG, expressed as higher GPX, GR and GST activities. In addition, LAR fish showed elevated GST and CAT activities. Thiols content was higher in all study sites irrespective to the pollution spectrum, whereas GSHt increase was only observed at GAF and VAG. The highest MT level was detected in fish from VAG and the lowest level from RIO suggesting a low contamination degree on this particular site. Globally, the results expressed site-specific response patterns, signalling two critical areas — GAF and VAG. Additionally, a joint analysis (damage vs. protection) allowed the following ordering of surveyed sites according to the contamination degree and risk to fish health; GAF N VAG N LAR N RIO N TOR. D. labrax gill responses demonstrated their efficacy as early warning signals of the contaminants presence. Moreover, the adopted approach, considering simultaneously protection responses and damaging effects, also revealed its usefulness on the pollution extent assessment. © 2008 Elsevier B.V. All rights reserved. Keywords: Dicentrarchus labrax L. Genotoxicity Oxidative stress Metallothionein Damage vs. Protection Ria de Aveiro 1. Introduction Coastal environments are subjected to several forms of disturbance, among which chemical pollution associated with high urbanization is of major concern. Inventory-based chemical monitoring of these impacted environments is restricted to the identification of a limited number of substances without providing information on their biological significance (Livingstone et al., 1992). To overcome this limitation, increasing emphasis has been placed on the SCIENCE OF THE TOTAL ENVIRONMENT 406 (2008) 298 – 307 ⁎ Corresponding author. Tel.: +351 234370965; fax: +351 234372587. E-mail address: [email protected] (I. Ahmad). 0048-9697/$ – see front matter © 2008 Elsevier B.V. All rights reserved. doi:10.1016/j.scitotenv.2008.06.027 available at www.sciencedirect.com www.elsevier.com/locate/scitotenv
Transcript of DNA damage and lipid peroxidation vs. protection responses in the gill of Dicentrarchus labrax L....

S C I E N C E O F T H E T O T A L E N V I R O N M E N T 4 0 6 ( 2 0 0 8 ) 2 9 8 – 3 0 7
ava i l ab l e a t www.sc i enced i r ec t . com
www.e l sev i e r. com/ loca te / sc i to tenv
DNA damage and lipid peroxidation vs. protection responses inthe gill of Dicentrarchus labrax L. from a contaminated coastallagoon (Ria de Aveiro, Portugal)
I. Ahmada,⁎, V.L. Mariab, M. Oliveiraa, A. Serafimb, M.J. Bebiannob,M. Pachecoa, M.A. Santosa
aCESAM & Department of Biology, University of Aveiro, 3810-193 Aveiro, PortugalbCIMA & Faculty of Marine and Environmental Sciences, University of Algarve, 8005-139 Faro, Portugal
A R T I C L E I N F O
⁎ Corresponding author. Tel.: +351 234370965;E-mail address: [email protected] (I. Ahmad).
0048-9697/$ – see front matter © 2008 Elsevidoi:10.1016/j.scitotenv.2008.06.027
A B S T R A C T
Article history:Received 19 February 2008Received in revised form12 June 2008Accepted 16 June 2008Available online 21 July 2008
The present research work aimed to investigate the damage vs. protection responses in gillof European sea bass (Dicentrarchus labrax) captured at a polluted coastal lagoon, Ria deAveiro (Portugal), as a tool to evaluate the human impacts on environmental health.Damage was assessed as DNA strand breakage and lipid peroxidation (LPO) whereasprotection was evaluated by measuring catalase (CAT), glutathione peroxidase (GPX),glutathione reductase (GR), glutathione S-transferase (GST), total glutathione (GSHt), thiolsand metallothioneins (MT). Fish were caught at five locations: Gafanha (GAF), Rio Novo doPríncipe (RIO), Laranjo (LAR) and Vagos (VAG) presenting each a different recognized sourceof contamination, and Torreira (TOR), assumed as reference site. Among the surveyed sites,gill damage was observed only at GAF, as measured by DNA integrity loss and LPO increase.An overall induction in enzymatic antioxidant protection was perceptible in fish from GAFand VAG, expressed as higher GPX, GR and GST activities. In addition, LAR fish showedelevated GST and CAT activities. Thiols content was higher in all study sites irrespective tothe pollution spectrum, whereas GSHt increase was only observed at GAF and VAG. Thehighest MT level was detected in fish from VAG and the lowest level from RIO suggesting alow contamination degree on this particular site. Globally, the results expressed site-specificresponse patterns, signalling two critical areas — GAF and VAG. Additionally, a jointanalysis (damage vs. protection) allowed the following ordering of surveyed sites accordingto the contamination degree and risk to fish health; GAFNVAGNLARNRIONTOR. D. labrax gillresponses demonstrated their efficacy as early warning signals of the contaminantspresence. Moreover, the adopted approach, considering simultaneously protectionresponses and damaging effects, also revealed its usefulness on the pollution extentassessment.
© 2008 Elsevier B.V. All rights reserved.
Keywords:Dicentrarchus labrax L.GenotoxicityOxidative stressMetallothioneinDamage vs. ProtectionRia de Aveiro
1. Introduction
Coastal environments are subjected to several forms ofdisturbance, among which chemical pollution associatedwith high urbanization is of major concern. Inventory-based
fax: +351 234372587.
er B.V. All rights reserved
chemical monitoring of these impacted environments isrestricted to the identification of a limited number ofsubstances without providing information on their biologicalsignificance (Livingstone et al., 1992). To overcome thislimitation, increasing emphasis has been placed on the
.

299S C I E N C E O F T H E T O T A L E N V I R O N M E N T 4 0 6 ( 2 0 0 8 ) 2 9 8 – 3 0 7
evaluation of the causal relationships between contaminantexposure and observable biological effects in aquatic organ-isms. Consequently, the use of biomarkers in sentinel specieshas become amajor issue in environmental quality evaluationand risk assessment. This approach is also important toinvestigate the mechanisms by which the environmentalinsult causes complex molecular–cellular alterations as wellas their interdependence.
Among the numerous ecotoxicological biomarkers pro-posed in the last decades, those based on the imbalancebetween pro-oxidant and antioxidant status as well as on itsconsequences namely DNA damage, lipid peroxidation (LPO)and enzyme inhibition, represent earlier signals of environ-mental disturbance (Livingstone, 2001; Ahmad et al., 2000;2004; Regoli et al., 2004; Santos et al., 2004). Previously, usingAnguilla anguilla L. exposed to metals and complex environ-mental mixtures of contaminants, it was shown that anti-oxidants modulation (induction or inhibition) can be usefulbiomarkers of pollution (Ahmad et al., 2005; 2006a, b).
Metallothioneins (MTs) are heat stable non-enzymaticproteins, with low molecular weight, high cysteine contentand a lack of aromatic amino acids, that have been thoroughlyevaluated as a marker for metal-induced stress (Ahmad et al.,2000; Haq et al., 2003; Bebianno et al., 2007; Fernandes et al.,2007). As with many other defence mechanisms, MTs synth-esis is inducible and involved not only in metal detoxification/homeostasis, but also in scavenging free radicals duringoxidative damage (Viarengo et al., 2000; Atif et al., 2006). LPOwas also found to have a high predictive value as revealedfrom a number of research papers describing its suitability as abiomarker of effect (Lackner, 1998; Livingstone, 2001; Ahmadet al., 2004; 2005). Moreover, the level of DNA strand breakage,in particular, has been proposed as a sensitive indicator ofgenotoxicity in both field and laboratory conditions (Mariaet al., 2004a, b; Regoli et al., 2004; Ahmad et al., 2006a).
In the past decade, a remarkable increment of scientificarticles dealing with pollutant-induced antioxidants modula-tion, MTs protection, genotoxicity and peroxidative damage infish was perceptible. Despite some studies dealing with thecausal and mechanistic links between some of the previousevents (Ahmad et al., 2000; 2006a; Amado et al., 2006;Fernandes et al., (2007), no studies provided evidence linkingall the previous events under realistic field conditions in fish.
Gill plays a central role on fish physiological balance basedonavariety of functions suchasgaseousexchange, osmotic andionic regulation, acid–base balance, and nitrogenouswaste. Thefilaments and lamellae of gill are covered by epithelial cells viz.,pavement cells, chloride cells and mucous cells supported by acomplex system of blood vessels. Hence, the gill epitheliumprovides an extensive surface of contact with the environment.Ironically, it also facilitates faster absorption of toxicantspresent on aqueous phase (Perry, 1997). The gill suitability forenvironmental biomonitoring came into lighton thebasis of ourprevious findings on A. anguilla responses to a variety ofstressors. Thus, following fish in situ exposure either to harbour(Ahmad et al., 2004) or pulp mill effluent affected areas (Santoset al., 2004), gill responded earlier than kidney and liver in termsof antioxidants and lipid peroxidation endpoints. Gill alsoshowed earlier stimulationmeasured as oxidative burst activityby adhered phagocytes when compared to peritoneal exudates
andhead kidney phagocytes following exposure to the previouscontaminated areas (Ahmad et al., 2004; Santos et al., 2006), aswell as in laboratory exposures to naphthalene (A. anguilla)(Ahmad et al., 2003) and paper mill effluent (Heteropneustesfossilis) (Fatima et al., 2000).
Historically, Ria de Aveiro (Portugal) has been the mainreceptor of anthropogenic discharges resulting mainly fromchlor–alkali and pulp/paper plants, harbour and dry-dockactivities as well as municipal and domestic effluents.Previous chemical analyses displayed the presence of PAHs,organometallic compounds and heavy metals, as an indica-tion of water quality deterioration in some critical areas of thisestuarine system (Pacheco et al., 2005; Sousa et al., 2007).Moreover, this lagoon, like other estuaries, is a nursery area forjuvenile stages of several fish species including European seabass (Dicentrarchus labrax L.). Besides the ecological key role ofthis species on coastal systems, it has a high commercial valuebeing the basis of professional and recreational fishingactivities. Thus, the detection of early effects disturbing theindividual fitness of sea bass in a multi-pollution context canprovide crucial information to address human impacts on theaquatic environment as well as the impact of contaminants inthe human environment keeping in view a sustainablemanaging of biotic resources. Pursuing these purposes, thedichotomy between damage and protection responses in gillof D. labrax captured in different sites of Ria de Aveiro wasinvestigated. The following parameters were assessed: DNAstrand breakage and LPO increase as markers of damage;catalase (CAT), glutathione peroxidase (GPX), glutathionereductase (GR), glutathione S-transferase (GST) as enzymaticprotection; and non-protein thiols (NP-SH), total glutathione(GSHt), MT contents as non-enzymatic protection. In addition,it was intended to evaluate the efficacy of gill responses asearly warning signals of the contaminants presence.
2. Materials and methods
2.1. Chemicals
Genomic DNA purification kit was purchased from Fermentas.Butylated hydroxytoluene (BHT), diethylenetriaminepenta-acetic acid (DTPA), 1-chloro, 2,4, dinitrobenzene (CDNB), 2,5,dithiobis-tetranitrobenzoic acid (DTNB), 2-thiobarbituric acid(TBA), trichloroacetic acid (TCA), β-nicotinamide adeninedinucleotide phosphate reduced tetrasodium salt (NADPH),ethylene diamine tetraacetic acid tetrasodium salt hydrate(EDTA), sodium azide (NaN3), glutathione reductase (GR),glutathione reduced (GSH), glutathione oxidized (GSSG) andpurified rabbit liver MT were purchased from Sigma. Otherroutine chemicals and reagents (analytical grade) werepurchased from local sources.
2.2. Sampling — animals and stations
The sampling was carried out in October 2005 and juvenileEuropean sea bass (D. labrax L.) were collected (n=6) from theRia de Aveiro, Portugal. The sampling sites were selected on ageographic distribution basis along the main channels, takinginto account the various types and sources of contamination

300 S C I E N C E O F T H E T O T A L E N V I R O N M E N T 4 0 6 ( 2 0 0 8 ) 2 9 8 – 3 0 7
aswell as the selection of a (theoretically) unpolluted referencepoint (Fig. 1). The sampling sites were: Torreira (TOR) as areferencepoint at the stretch of the longest channel (S. Jacinto–Ovar channel) having non-point pollution inputs and far fromthe main polluting sources; Gafanha (GAF) situated in thevicinity of a deep-sea fishing port and dry-docks, alsoconnected with the main channel coming from Aveiro cityreceiving domestic discharges; Rio Novo do Principe (RIO)located at the terminal area of the Vouga river and the mainlagoons' fresh water source, 6.5 km distant from a pulp/papermill effluent outlet, that was exposed during nearly fivedecades to its discharges; Laranjo (LAR) close to a chlor–alkaliplant, an important source of mercury contamination; Vagos(VAG) at the terminal part of the Ílhavo channel, receivingmunicipal and domestic effluents.
Fish biometrical parameters, such as weight and lengthranged from 18.71–41.96 g and 12.9–16.5 cm. Hydrologicalabiotic parameters such as temperature, dissolved oxygen,salinity, pH were measured at sub-surface level. Turbidity(Secchi disc method) and depth were also determined.
2.3. Organs handling and preparation of post-mitochondrialsupernatant (PMS) fraction
Immediately after catching, fish were sacrificed and gills wereremoved and frozen in liquid nitrogen. In laboratory, each fish
Fig. 1 –Map of Ria de Aveiro (Portugal) with locations offish-capture sites (■). The respective coordinates are:reference site (TOR)—40°44′02 N, 008°41′44 W; GAF—40°38′38 N, 008°41′42 W; RIO—40′41′08 N, 008°39′41 W; LAR—40°43′30 N, 008°37′43 W; and VAG—40°33′59 N, 008°40′55 W.
gill was divided in three sets of tissue for all the assays per-formedand stored at −80 °C: first set corresponding to one archof gill for later DNA strand breaks measurement; second setcorresponding to the sameportion of gill forMTmeasurement;and the third set corresponding to the rest of gill for furtherprocess in oxidative stress studies. Thus, the third set washomogenized, using a Potter–Elvehjem homogenizer, inchilled phosphate buffer (0.1 M, pH 7.4) (1 g of tissue/15 mlbuffer=15%). This homogenate was then divided in threealiquots for LPO, glutathionemeasurements and PMS prepara-tion. The PMSpreparationwas accomplished by centrifugationin a refrigerated ultracentrifuge (Optima TL, Beckman) at13,400 g for 20 min at 4 °C. Aliquots of PMS were divided inmicrotubes and stored at −80 °C until analyses.
2.4. DNA strand breaks evaluation
Genotoxicity was tested using the DNA alkaline unwindingassay. The previous stored gill portions were placed in TNES(Tris–HCl 10mM, NaCl 125mM, EDTA 10mM, SDS 1%, pH 7.5) –urea (5 M) buffer with proteinase K solution (final concentra-tion 0.8 mg/ml). Gill DNA isolation was performed using agenomic DNA purification kit (Fermentas). DNA integritymeasurements were performed according to Rao et al. (1996)with minor modifications, as adopted by Maria et al. (2002).DNA single and double strands were separated by hydro-xyapatite (20%) “batch” elution and quantified with a DNA-specific fluorescent dye (2.5 nM bisbenzimide) after alkalineunwinding. Data from DNA unwinding technique wereexpressed as DNA integrity (%), determined by applying thefollowing equation:
DNA integrity kð Þ ¼ ½ Double stranded DNAð Þ=ðDouble stranded DNA
þ Single stranded DNAÞ� � 100:
2.5. Estimation of lipid peroxidation (LPO)
LPO was determined in the previously prepared tissue homo-genate by the procedure of Ohkawa et al. (1979) and Bird andDraper (1984)with somemodifications. Briefly, to a 50 μl of 15%homogenate, 3 μl of 4%BHT inmethanol,was addedandmixedwell. To this aliquot, 0.5 ml of 12% TCA in aqueous solution,0.45 ml Tris–HCl (60 mM, pH 7.4; and 0.1 mM DTPA) and 0.5 ml0.73% TBA were added and mixed well. The mixture washeated for 1 h in a water bath set at boiling temperature. Thetest tubeswere then removedand cooled to roomtemperature.The contents from each tube were decanted into 2 ml micro-tubes and centrifuged at 13,400 g for 3 min. The absorbance ofeach aliquot was measured at 535 nm. The rate of LPO wasexpressed as nmol of thiobarbituric acid reactive substances(TBARS) formed per milligram of protein using a molar ex-tinction coefficient of 1.56×105 M−1cm−1.
2.6. Catalase (CAT) activity measurement
CAT activity was assayed in PMS fraction by the method ofClaiborne (1985) with some modifications. Briefly, the assaymixture consisted of 1.99 ml phosphate buffer (0.05 M, pH 7.0),1 ml H2O2 (30 mM) and 0.010 ml PMS in a final volume of 3 ml.Change in absorbance was recorded at 240 nm at room

301S C I E N C E O F T H E T O T A L E N V I R O N M E N T 4 0 6 ( 2 0 0 8 ) 2 9 8 – 3 0 7
temperature. CAT activity was calculated in terms of µmolH2O2 consumed/min/mg protein using a molar extinctioncoefficient of 43.5 M−1 cm−1.
2.7. Glutathione peroxidase (GPX) activity measurement
GPX activity was assayed in PMS fraction according to themethod described by Mohandas et al. (1984) with somemodification. The assay mixture consisted of 0.720 ml phos-phate buffer (0.05 M, pH 7.0), 0.050 ml EDTA (1 mM), 0.050 mlsodium azide (1 mM), 0.025 ml GR (1 IU/ml), 0.050 ml GSH(4 mM), 0.050 ml NADPH (0.8 mM), 0.005 ml H2O2 (1.0 mM) and0.050 ml of PMS (15%) in a total volume of 2 ml. Oxidation ofNADPH was recorded spectrophotometrically (Jasco UV/VIS,V-530) at 340 nm at room temperature. The enzyme activitywas calculated as nmol NADPH oxidized/min/mg of protein,using a molar extinction coefficient of 6.22×103 M−1 cm−1.
2.8. Glutathione reductase (GR) activity measurement
GR activity was assayed by the method of Cribb et al. (1989)with somemodifications. Briefly, the assaymixture contained0.025ml of PMS fraction and 0.925ml of NADPH (0.2mM), GSSG(1 mM) and DTPA (0.5 mM). The enzyme activity wasquantified at room temperature by measuring the disappear-ance of NADPH at 340 nm during 3 min. The enzyme activitywas calculated as nmol NADPH oxidized/min/mg proteinusing a molar extinction coefficient of 6.22×103 M−1cm−1.
2.9. Glutathione S-transferase (GST) activity measurement
GST activity was determined in PMS fraction, following theconjugation of GSHwith 1-chloro-2,4-dinitrobenzene (CDNB) bythe method of Habig et al. (1974) with some modifications. Thereaction mixture consisted of 1.85 ml sodium phosphate buffer(0.1 M, pH 7.4), 0.050 ml reduced glutathione (1 mM), 0.050 mlCDNB (1 mM) and 0.050 ml PMS in a total volume of 2 ml. Thechange in absorbance was recorded at 340 nm and enzymeactivity calculated as mmol CDNB conjugate formed/min/mgprotein using a molar extinction coefficient of 9.6 mM−1 cm−1.
2.10. Total non-protein thiols (NP-SH) content
NP-SH was determined in a different aliquot in relation toGSHt by the method of Sedlak and Lindsay (1968) as adoptedby Parvez et al. (2003). PMS (0.3 ml) was precipitated with0.3 ml of sulfosalicylic acid (5.0%). The samples were kept at4 °C for 1 h and then subjected to centrifugation at 13,400 g for15 min at 4 °C. The assay mixture contained 0.5 ml of filteredaliquot, 2.3 ml of sodium phosphate buffer (0.1 M, pH 7.4) and0.2 ml DTNB (stock=100 mM in 0.1 M sodium phosphatebuffer, pH 7.4) in a total volume of 3 ml. The optical density ofreaction product was read immediately at 412 nm on aspectrophotometer.
2.11. Total glutathione (GSHt) content
Protein content in the tissue lysate was precipitated withsulfosalicylic acid (5%) for 1 h and then centrifuged at 13,400 gfor 20min at 4 °C. GSHt contentwas determined in the resulting
supernatant (deproteinated PMS) adopting the enzymatic re-cycling method using GR excess, whereby the sulfhydryl groupof GSH reacts with DTNB (Ellman's reagent) producing a yellowcoloured 5-thio-2-nitrobenzoic acid (TNB). The rate of TNBproduction is directly proportional to this recycling reaction,which is in turn directly proportional to the concentration ofGSH in the sample (Tietze, 1969; Baker et al., 1990). Formation ofTNB was measured by spectrophotometry at 412 nm. It shouldbe noted that GSSG is converted to GSH by GR in this system,which consequently measures total GSH. The results wereexpressed as µmol TNB formed/min/mg protein using a molarextinction coefficient of 14.1×103 M−1 cm−1.
2.12. Metallothioneins (MT) determination
MT levels were determined after gill homogenization in 20 mMTris buffer (pH 8.6) containing 150 mM of NaCl. The homo-genates were centrifuged for 45min at 30,000 g at 4 °C. Aliquotsof the supernatant were heat-treated at 80 °C for 10min and re-centrifuged at 30,000 g for 45min at 4 °C. MT quantification wasmade by differential pulse polarography (DPP) according to themethod described by Bebianno and Langston (1989) using a646VA Processor autolab type II and an ECO Chemie IME663mercury drop electrode. Comparisons of peak heights withthose of standard additions of purified rabbit MT enabled thequantification. Accuracy of the DPP analytical method wasobtained by internal calibration using commercial rabbit liverMT-I. Recovery studies of our cytosols spiked with the MT-Irabbit liver standards revealed good recovery results rangingfrom 97 to 103%. Results were expressed as µg MT/g wet weight(ww)/mg protein.
2.13. Protein measurement
Total protein contents were determined according to theBiuret method (Gornall et al., 1949), using bovine serum al-bumin (E. Merck-Darmstadt, Germany) as a standard.
2.14. Statistical analysis
Statistica software (StatSoft, Inc., Tulsa, OK, USA) was used forstatistical analyses. The final results were based on themeansof the six individual fish by each group (n=6). Data were firsttested for normality and homogeneity of variance to meetstatistical demands. ANOVA analysiswas used to compare theresults of the various fish groups, followed by the Tukey's testin order to compare results between fish groups (Zar, 1999)and the significance of the results was ascertained at pb0.05.
3. Results
The hydrological abiotic parameters showed, in general, noimportant differences between the surveyed sites. Hence,temperature ranged from17.9 to 19.0 °C, pH from7.7 to 8.2, anddissolved oxygen from 5 to 6.9. Considering salinity, all thesites were in the range 29–34, excluding RIOwhere it was 21. Interms of turbidity, LAR and GAF displayed the highest levels(0.3 m) while the other sites ranged from 0.7 to 1.0 m. Depthranged from 1.2 (TOR and LAR) to 6.9 (GAF).

302 S C I E N C E O F T H E T O T A L E N V I R O N M E N T 4 0 6 ( 2 0 0 8 ) 2 9 8 – 3 0 7
3.1. Gill damage (Fig. 2)
3.1.1. Genotoxicity (DNA integrity loss)Among the sampled sites, gill genotoxicity was observed in GAFfish, as measured by significant DNA integrity decrease whencompared to fish collected from TOR, as well as to RIO and LAR.The decrease was 50% in comparison to the TOR.
3.1.2. LPO inductionFish collected from GAF showed a significant LPO increasecomparing with those from TOR, RIO and LAR. The observedLPO increase was 1.8 folds when compared to TOR.
3.2. Gill protection responses (Fig. 3)
3.2.1. Enzymatic antioxidants protectionAn overall induction in enzymatic protection is perceptible inthe current study. CAT activity at LAR was significantly higherwhen compared to fish from TOR, RIO and VAG sites. Theincrease was 2.3 folds comparing to TOR values. The GPXactivity was significantly increased in fish collected from GAF(1.7 folds) and VAG (1.9 folds) sites when compared to fishcollected from TOR.
When the gill GST activities were compared in all thestudied sites, fish collected from GAF, LAR and VAG showed asignificant increase in comparison to the fish collected fromTOR site. The increased values were 2.0 (GAF), 1.8 (LAR) and 1.9(VAG) folds comparing to TOR values, respectively.
The GR enzyme showed a significantly increased responsein gill of fish from GAF and VAG, when compared to TOR aswell as RIO and LAR sites. The increased activity levels were2.9 folds at VAG and 3.0 folds at GAF in comparison to TOR.
3.2.2. Non-enzymatic protectionTheNP-SHcontentswere significantly increased ingill ofD. labraxcollected from all the study sites in comparison to TOR. Theincrease in NP-SH content was 3 folds at GAF, 1.4 folds at RIO, 2.3folds at LAR and 3.0 folds at VAG. However, a decrease in NP-SHcontent was recorded at RIO in comparison to GAF and VAG.
GSHt levels were significantly increased in fish collectedfrom GAF and VAG sites. The increase was 1.8 fold at GAF and2.2 fold at VAG when compared to the GSHt values of fish
Fig. 2 –Represents damage [DNA integrity (%) and lipid peroxidatsites in the Ria de Aveiro (Torreira — TOR, Gafanha — GAF, Rio NValues represent the means±S.E. (n=6). Distinct letters indicate s(pb0.05). a vs. TOR; b vs. RIO; c vs. LAR.
collected from TOR. Moreover, fish collected from VAG alsoshowed a significant increase in comparison to RIO and LAR.
As regards gill MT content, the significant elevation wasobserved in fish from VAG (vs. TOR, GAF, RIO and LAR). Theincrease was 14.8 (VAG) folds in comparison to TOR. However,fish from RIO showed a significant decrease in MT contentwhen compared to TOR, GAF, LAR and VAG.
4. Discussion
A theoretically ideal fish species to assess pollutant-inducedeffects should be (i) sensitive enough to detect effects of a widerange of pollutants at low doses, (ii) widespread in theconsidered ecosystems, and (iii) abundant in terms of naturalpopulations, to allow detraction of individuals without endan-gering their conservation. Taking into account the previouscriteria and the current goals, D. labrax suffice every require-ment justifying its selection.Moreover, juvenile specimensweresurveyed in order to aside the gender interference and diminishthe background of previous exposures to pollutants.
Peroxidative damage to unsaturated fatty acids is a typicalreaction during oxidative stress and has been convenientlyused as effect biomarker (Pandey et al., 2001; Ahmad et al.,2004; Santos et al., 2006). Malonaldehyde, one of the productsof LPO has been shown to be highly reactive and an importantmediator of DNA damage (Wang and Liehr, 1995). Thus, DNAintegrity decrease may be associated with degenerativeprocesses initiated by LPO, supporting the importance of ROSas a causative agent of DNA damage (Wang and Liehr, 1995).On the other hand, fish tends to adapt to oxidative conditionsby mobilizing antioxidant defences (Lackner, 1998) pointingthe presence of redox active chemicals. Thus, CAT, GPX andGR induction provides an indication of the presence of redoxactive chemicals (Stephensen et al., 2000). GST helps ineliminating reactive compounds by forming conjugates withGSH, thereby protecting cells against ROS induced damage(Rodriguez-Ariza et al., 1991). Moreover, its role as antioxidanthas also been reported by Wang and Ballatori (1998) statingthat the reduction of organic hydroperoxides by GSH may becatalyzed by GST of α class. Glutathione represents the bulk ofNP-SH content of the cells, but it was demonstrated that other
ion (LPO)] in gill of Dicentrarchus labrax L. collected at differentovo do Príncipe — RIO, Laranjo — LAR and Vagos — VAG).ignificant differences between sites according to Tukey's test
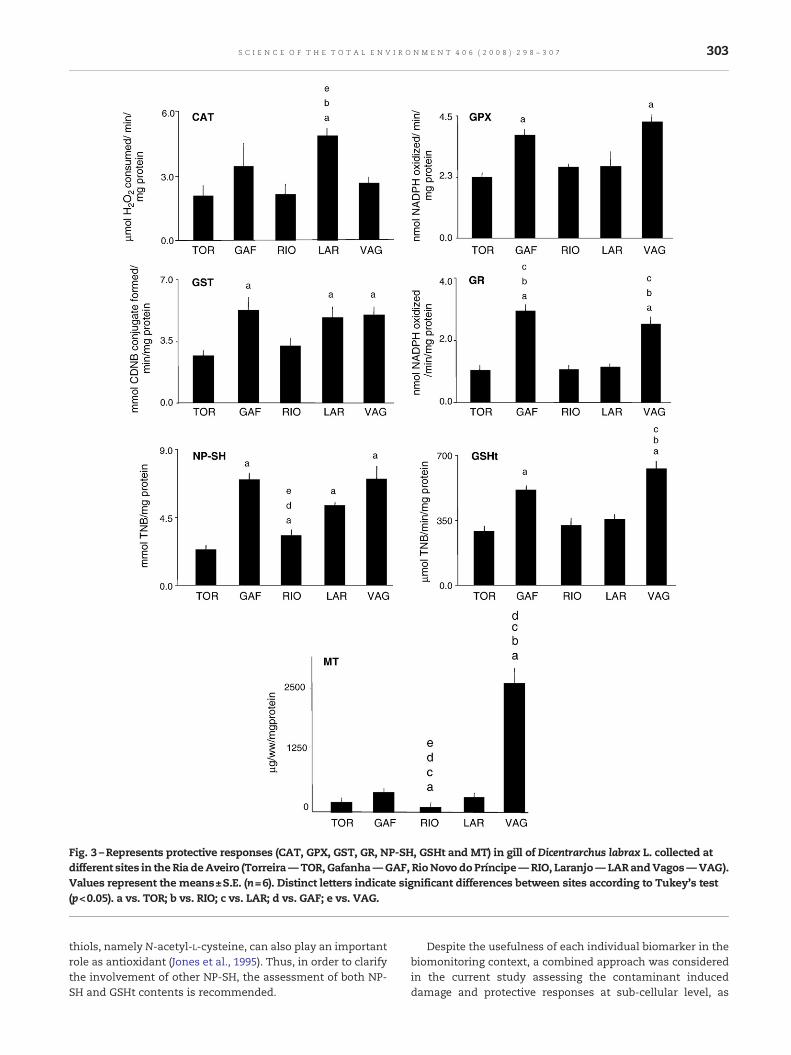
Fig. 3 –Represents protective responses (CAT, GPX, GST, GR, NP-SH, GSHt and MT) in gill of Dicentrarchus labrax L. collected atdifferent sites in theRia deAveiro (Torreira—TOR, Gafanha—GAF, RioNovo doPríncipe—RIO, Laranjo— LARandVagos—VAG).Values represent the means±S.E. (n=6). Distinct letters indicate significant differences between sites according to Tukey's test(p<0.05). a vs. TOR; b vs. RIO; c vs. LAR; d vs. GAF; e vs. VAG.
303S C I E N C E O F T H E T O T A L E N V I R O N M E N T 4 0 6 ( 2 0 0 8 ) 2 9 8 – 3 0 7
thiols, namely N-acetyl-L-cysteine, can also play an importantrole as antioxidant (Jones et al., 1995). Thus, in order to clarifythe involvement of other NP-SH, the assessment of both NP-SH and GSHt contents is recommended.
Despite the usefulness of each individual biomarker in thebiomonitoring context, a combined approach was consideredin the current study assessing the contaminant induceddamage and protective responses at sub-cellular level, as

304 S C I E N C E O F T H E T O T A L E N V I R O N M E N T 4 0 6 ( 2 0 0 8 ) 2 9 8 – 3 0 7
well as its interdependence as a tool to evaluate the ecosystemhealth. It was previously observed that peroxidative damagemay occur without concomitant alterations of antioxidants(Cossu et al., 2000) and vice versa (Ahmad et al., 2006b). Hence,the adopted strategy reduces the risk to misinterpretationsresulting from the isolated analysis of each aspect.
Taking into account that hydrological abiotic conditionsdid not substantially differ between the surveyed sites, currentdiscussion will be made on the basis of the predictable conta-mination scenario at each particular site. An exception wasconsidered for salinity at RIO site.
At GAF area, the overall results pointed out a cell defenceoverwhelm resulting on the initiation of damage on cellconstituents. The activation of all the protection responses(except CAT and MT) was not enough to prevent both LPO andDNA damage in gill cells. Previous studies on GAF areadetected moderate levels of PAHs (Pacheco et al., 2005) andhigh levels of organometallics such as tributyltin (TBT) (Sousaet al., 2007). Moreover, due to its location, the occurrence ofxenoestrogens must be also considered. Taking into accountthe previous pollution scenario, the genotoxicity and LPOincrease observed in fish collected from GAF should beattributed to the combination of different classes of com-pounds rather than to individual chemicals. The currentresults in terms of damage evidence are in agreement withprevious findings concerning A. anguilla caged in the samelocation in 2001. Ahmad et al. (2004) found LPO increase in gill,while Maria et al. (2004b) detected DNA integrity loss in theassessed eel tissues (blood, liver and kidney). Differently,A. anguilla caged at GAF displayed gill antioxidant depression(GSH, GPX, CAT and GST) (Ahmad et al., 2004). Though theprevious and current results are not directly comparable dueto differences on the mode of exposure, fish species andselected organ, the potential risk for wild fish population atGAF was strengthened, also suggesting that environmentalhealth in this particular site was not improved in 2005 inrelation to 2001 concerning the presence of genotoxicants andpro-oxidants. A failure in the detoxification process wheresome unconjugated-reactive metabolites may damage DNAwas presented by Maria et al. (2004b) as a possible mechanismfor genotoxicity induction at GAF. Alternatively, ROS involve-ment should also be considered since Ahmad et al. (2004)found the presence of ROS over-production in gill of A. anguillacaged at GAF associated to oxidative burst activity (OBA) in gilladhered phagocytes.
Concerning the responses of fish captured at RIO, LAR andVAG sites, it was evident an effective protection able toprevent genetic and peroxidative damage. Thus, NP-SH actionwas enough to prevent damage at RIO without any interven-tion of enzymatic protection. In LAR, besides NP-SH, CAT andGST activities also participated on the protection strategy.Finally, at VAG the contamination scenario seems to be morechallenging for gill cell protection mechanisms, since all thestudied parameters (except CAT) were induced.
The ability of pulpmill effluent components accumulated inRIO sediments to induce DNA damage in blood and liver cells(Maria et al., 2004c), LPO and OBA increase in gill (Santos et al.,2006), aswell as antioxidants (GSH,GPX,CATandGST)depletionin gill (Santos et al., 2004) was demonstrated on the basis ofcaged A. anguilla experiments carried out in 2002. Despite the
obvious differences between previous and present studies, theabsence of antioxidants alterations and damaging effects maysuggest an improvement on RIO water quality, probably as aresult of the pulp mill effluent outlet deviation towards the seacarried out in 2000. RIO site displayed a lower salinity in relationto all the other sites. However, this fluctuation is within rangestolerable to sea bass. It was demonstrated in estuarine speciesthat MT induction is higher in low salinity (Leung et al., 2002).Therefore, the lowMTlevels observed ingill of seabass fromRIOcannot be explained by the salinity decrease and probablycorrespond to the basal levels of MT in gill of this species.Decreases in salinity are also associated with increasedbioavailability and uptake rates of both metals (Monserratet al., 2007) and organic contaminants (Ramachandran et al.,2006). In the light of the previous statements, the idea of a lowcontamination degree at RIO area is reinforced.
LAR is recognizedashighlypollutedbymercury (Pereiraet al.,1997), and previous studies have shown that different species,including D. labrax captured (Abreu et al., 2000) and Liza auratacaged at this location had been contaminated by industrialmercury (Guilherme et al., 2008a, b). According to Stohs andBagchi (1995), mercury is a pro-oxidant that exerts oxidativestress via H2O2 production, and thus confirming the current CATincreased activity. GST induction observed at LAR suggests itsantioxidant action against metals rather than to its phase IIdetoxification action since metals have not been reported assubstrate to GST. Most research on MT indicates that thesestress-related proteins can be induced by heavymetals and thatthey may therefore be used as specific biomarkers of metalcontamination. Surprisingly, in the present study no MTinductionwas observed in LAR area, raising a question concern-ing the usefulness of MT assessment as an indication of metals'presence on the basis of fish gill. The non-appearance of DNAdamage in sea bass from LAR seems to be in disagreement withthe findings of Guilherme et al. (2008a) who showed erythrocyticnuclear abnormalities (ENA) induction in L. aurata captured inthe same area and in the same year. Among other differences,the higher cell renewal rate of gill in relation to blood should beconsidered. Taking into account that mercury is mostly accu-mulated in the sediment (Guilherme et al., 2008a), differentfeeding behaviours cannot be overlooked since L. aurata as amud feeder is potentially more vulnerable to mercury effectscomparing to D. labrax.
In relation to VAG, despite the efficacy of protective mecha-nisms onpreventing damage on lipidmembranes andDNA, thepresence of threatening levels of oxidant compounds isapparent. The elevated GST activity observed at VAG suggestsan activation of gill detoxification processes probably due to thepresence of organic contaminants, namely PAHs, previouslymeasured in high concentrations (Pacheco et al., 2005). The MTstimulation observed at VAG, an area contaminated with non-metal compounds, should not be considered as unexpectedsincenon-metals have already been shownas a causative agentfor MT induction in aquatic organisms, namely biocides (Paris-Palacios et al., 2003; Mosleh et al. 2004, 2005a, b). Moreover, astudy on rats treated with paraquat demonstrated that the ROSscavenging potential of MT can be comparable to that of GSH(Sato and Bremner, 1993). Taking into consideration theprevious statements and the present findings, MT role as ROSscavengers and their inducibility by non-metals must not be

305S C I E N C E O F T H E T O T A L E N V I R O N M E N T 4 0 6 ( 2 0 0 8 ) 2 9 8 – 3 0 7
overlooked for the interpretation of MT over-expression in fieldsurveys. In contrast with the current findings showing no DNAdamage at VAG site, a L. aurata field survey carried out in 2000demonstrated ENA induction which was associated with highlevels of PAHs (Pacheco et al., 2005).
As per the pattern of gill DNA integrity and LPO, a high simi-larity is evident on both parameters for all sites. A consistentincrease in thiol contentswas foundat all thepresumedpollutedsites irrespectively to the pollution scenario, while the otherstudiedparameters showedpunctual responses suggesting theirhigher specificity. Hence, GSHt increase was detected only atGAF and VAG sites, suggesting the relevance of other thiolsbesides GSH at RIO and LAR sites. Though GR activity was in-creased at GAF and VAG, the GSHt elevation detected at thosesites cannot be explained by the increased regeneration ofoxidizedglutathione carriedout by thisenzyme, but represents aGSH new synthesis as an adaptive response towards itsincreased use. It must be also highlighted that CAT and GPXincreaseswere not detected concomitantly, suggesting a coordi-nated regulation of these two antioxidant activities by the cell asa function of the type and degree of contamination. The overallanalysis of current gill protection responses showed antiox-idants induction coupled with or without peroxidative damage.
5. Conclusions
Present results expressed site-specific response patterns, iden-tifying two critical areas at the Ria de Aveiro — Gafanha andVagos — and reflecting a contamination by a mixture ofpollutants from point and non-point sources. A global analysisof D. labrax responses indicates that, considering the degree ofcontamination and risk to fish health, the surveyed sites can beordered in the following manner: GAFNVAGNLARNRIONTOR.
The gill susceptibility towards the imbalance between pro-oxidants challenges and antioxidants protection was demon-strated, pointing a potential risk to native ichthyofauna inha-biting the impacted areas as prolonged oxidative stress mayimpair the individual fitness. Despite the complexity underlyingthe dichotomy between protection and damage responses, ag-gravatedby thepresenceof contaminantmixtures, thisapproachrevealed its efficacy on providing early warning signals of thecontaminants presence and the assessment of the pollutionextent. Moreover, both protection and damage responses shouldbe considered simultaneously to avoidmisinterpretations result-ing from the non-occurrence of damage on cell constituents inparallel with early protection activation and vice versa.
D. labrax proved its ability for biomonitoring aquatic an-thropogenic contamination on the basis of its gill responsesand can be successfully employed as a sentinel species in bothAtlantic and Mediterranean European coastal waters.
Acknowledgements
The financial supports from FCT (Government of Portugal)provided through POCI/MAR/61195/2004 SFRH/BPD/34326/2006, SFRH/BPD/26970/2006, SFRH/BD/27584/2006 and by theAveiro University Research Institute/CESAM are gratefullyacknowledged.
R E F E R E N C E S
Abreu SN, Pereira E, Vale C, Duarte AC. Accumulation of mercuryin Sea Bass from a Contaminated Lagoon (Ria de Aveiro,Portugal). Mar Pollut Bull 2000;40:293–7.
Ahmad I, Maria VL, Oliveira M, Santos MA. Anguilla anguilla L. gilland kidney oxidative stress and genotoxic response tochromium exposure with and without β-naphthoflavonepre-exposure. Mutat Res 2006a;608:16–28.
Ahmad I, Pacheco M, Santos MA. Anguilla anguilla L. oxidativestress biomarkers: an in situ study of freshwater wetlandecosystem (Pateira de Fermentelos, Portugal). Chemosphere2006b;65:952–62.
Ahmad I, Oliveira M, Pacheco M, Santos MA. Anguilla anguilla L.oxidative stress biomarkers responses to copper exposure withor without β-naphthoflavone pre-exposure. Chemosphere2005;61:267–675.
Ahmad I, Pacheco M, Santos MA. Enzymatic and non-enzymaticantioxidants as an adaptation to phagocyte-induced damagein Anguilla anguilla L. following in situ harbor water exposure.Ecotoxicol Environ Saf 2004;57:290–302.
Ahmad I, PachecoM, SantosMA.Naphthalene-induceddifferentialtissue damage association with circulating fish phagocytesinduction. Ecotoxicol Environ Saf 2003;54:7–15.
Ahmad I, Hamid T, Fatima M, Chand HS, Jain SK, Athar M,Raisuddin S. Induction of hepatic antioxidants in freshwaterfish (Channa punctatus B.) is a biomarker of paper mill effluentexposure. Biochim Biophys Acta 2000;1523:37–48.
Amado LL, Robaldo RB, Geracitano L, Monserrat JM, Bianchinia A.Biomarkers of exposure and effect in the Brazilian flounderParalichthys orbignyanus (Teleostei: Paralichthyidae) from thePatos Lagoon estuary (Southern Brazil). Marine Pollut Bul2006;52:207–13.
Atif F, Kaur M, Yousuf S, Raisuddin S. In vitro free radicalscavenging activity of hepatic metallothionein induced in anIndian freshwater fish, Channa punctata Bloch. Chem BiolInteract 2006;162:172–80.
Baker MA, Cerniglia GJ, Zaman A. Microtiter plate assay for themeasurement of glutathione and glutathione disulfide in largenumbers of biological samples. Anal Biochem 1990;190:360–5.
Bebianno MJ, Santos C, Canário J, Gouveia N, Sena-Carvalho D,Vale C. Hg and metallothionein-like proteins in the blackscabbardfish Aphanopus carbo. Food Chem Toxicol2007;45:1443–52.
Bebianno MJ, Langston WJ. Quantification of metallothioneins inmarine invertebrates using differential pulse polarography.Portugaliae Electrochimica Acta 1989;7:511–24.
Bird RP, Draper AH. Comparative Studies on Different Methods ofMalondyhaldehyde Determination Methods Enzymol.Academic Press; 1984. p. 299–305.
Claiborne A. Catalase activity. In: Greenwald RA, editor. CRChandbook of methods in oxygen radical research. Boca Raton,FL: CRC Press; 1985. p. 283–4.
Cossu C, Doyotte A, Babut M, Exinger A, Vasseur P. Antioxidantbiomarkers in freshwater bivalves, unio tumidus, in response todifferent contamination profiles of aquatic sediments.Ecotoxicol Environ Saf 2000;45:106–21.
Cribb AE, Leeder JS, Spielberg SP. Use of a microplate reader in anassay of glutathione reductase using 5,5′-dithiobis(2-nitrobenzoic acid). Anal Biochem 1989;183:195–6.
Fatima M, Ahmad I, Sayeed I, Athar M, Raisuddin S.Pollutant-induced overactivation of phagocytes isconcomitantly associated with peroxidative damage in fishtissues. Aquat Toxicol 2000;49:243–50.
Fernandes D, Porte C, Bebianno MJ. Chemical residues andbiochemical responses in wild and cultured European sea bass(Dicentrarchus labrax L.). Environ Res 2007;103:247–56.

306 S C I E N C E O F T H E T O T A L E N V I R O N M E N T 4 0 6 ( 2 0 0 8 ) 2 9 8 – 3 0 7
Guilherme S, Válega M, Pereira ME, Santos MA, Pacheco M.Erythrocytic nuclear abnormalities in wild and caged fish(Liza aurata) along an environmental mercury contaminationgradient. Ecotoxicol Environ Saf 2008a;70:411–21.
Guilherme S, Válega M, Pereira ME, Santos MA, Pacheco M.Antioxidant and biotransformation responses in Liza aurataunder environmental mercury exposure — relationship withmercury accumulation and implications for public health. MarPollut Bul 2008b;56:845–59.
Gornall AC, Bardawill CJ, David MM. Determination of serumproteins by means of the biuret reaction. J Biol Chem1949;177:751–66.
Habig WH, Pabst MJ, Jakoby WB. Glutathione-S-transferases, thefirst enzymatic step in mercapturic acid formation. J Biol Chem1974;249:7130–9.
Haq F, Mahoney M, Koropatnick J. Signaling events formetallothionein induction. Mut Res 2003;533:211–26.
Jones DP, Maellaro E, Jiang S, Slater AG, Orrenius S. Effects ofN-acetyl-L-cysteine on T-cell apoptosis are not mediated byincreased cellular glutathione. Immunol Let 1995;45:205–9.
Lackner R. Oxidative stress in fish by environmental pollutants. In:Braunbeck TD, Hinton E, Streit B, editors. Fish Ecotoxicology.Basel: Birkhauser; 1998. p. 203–24.
Leung KMY, Svavarsson J, Crane M, Morritt D. Influence of staticand fluctuating salinity on cadmium uptake andmetallothionein expression by the dog whelk Nucella lapillus(L.). J Exp Mar Biol Ecol 2002;274:175–89.
Livingstone DR, Donkin P, Walker CH. Pollutants in marineecosystems: an overview. In: Walker CH, Livingstone DR,editors. Persistent Pollutants in Marine Ecosystems. Oxford:Pergamon Press; 1992. p. 236–63.
Livingstone DR. Contaminant stimulated reactive oxygen speciesproduction and oxidative damage in aquatic organisms.Marine Pollut Bullet 2001;8:656–66.
Maria VL, Correia AC, Santos MA. Anguilla anguilla L. genotoxic andliver biotransformation responses to abietic acid exposure.Ecotoxicol Environ Saf 2004a;58:202–10.
Maria VL, Correia AC, Santos MA. Genotoxic and biochemicalresponses in caged eel (Anguilla anguilla L.) after short-termexposure to harbour waters. Environ Int. 2004b;29:923–9.
Maria VL, Teles M, Pacheco M, Correia A, Santos MA. Biomarkerresponses in polluted river possible effects of paper millcontaminants on caged Anguilla anguilla L. Fresenius EnvironBull 2004c;13:317–25.
Maria VL, Correia AC, Santos MA. Anguilla anguilla L. biochemicaland genotoxic responses to benzo(a)pyrene. Ecotoxicol EnvironSaf 2002;53:86–92.
Mohandas J, Marshall JJ, Duggins GG, Horvath JS, Tiller D. Lowactivities of glutathione-related enzymes as factors in thegenesis of urinary bladder cancer. Cancer Res 1984;44:5086–91.
Monserrat JM, Martínez PE, Geracitano LA, Amado LL, Martins CM,Pinho GL, Chaves IS, Ferreira-Cravo M, Ventura-Lima J,Bianchini A. Pollution biomarkers in estuarine animals: criticalreview and new perspectives. Comp Biochem Physiol C ToxicolPharmacol 2007;146:221–34.
Mosleh YY, Paris-Palacios S, Arnoult F, Couderchet M, Vernet G.Metallothionein induction in the aquatic Oligochaere tubifextubifex exposed to the herbicide isoproturon. Environ Toxicol2004;19:88–93.
Mosleh YY, Paris-Palacios S, Couderchet M, Biagianti-Risbourg S,Vernet G. Metallothionein induction, antioxidative responses,glycogen and growth changes in Tubifex tubifex (oligochaete)exposed to the fungicide, fenhexamid. Environ Poll2005a;135:73–82.
Mosleh YY, Paris-Palacios S, Couderchet M, Biagianti-Risbourg S,Vernet G. Effects of the herbicide isoproturon onmetallothioneins, growth and antioxidative defenses in theaquatic worm Tubifex tubifex (Oligochaeta, Tubificidae). Eco-toxicology 2005b;14:1–13.
Ohkawa H, Ohishi N, Yagi K. Assay for lipid peroxides in animaltissues by thiobarbituric acid reaction. Anal Biochem1979;95:351–8.
Pacheco M, Santos MA, Teles M, Oliveira M, Rebelo JE, Pombo L.Biotransformation and genotoxic biomarkers in mullet species(Liza sp.) from a contaminated coastal lagoon (Ria de Aveiro,Portugal). Environ Monit Assess 2005;107:133–53.
Pandey S, Ahmad I, Parvez S, Bin-Hafeez B, Haque R, Raisuddin S.Effect of endosulfan on antioxidants of freshwater fishChanna punctatus Bloch.: 1. Protection against lipid peroxidationin liver by copper preexposure. Arch Environ Contam Toxicol2001;41:345–52.
Paris-Palacios S, Biagianti-Risbourg S, Vernet G. Metallothioneininduction related to structural perturbations and antioxidativedefences in liver of roach (Rutilus rutilus) exposed to thefungicide procymidone. Biomarkers 2003;8:128–41.
Parvez S, Sayeed I, Pandey S, Ahmad A, Hafeez B, Haque R,Ahmad I, Raisuddin S. Regulatory role of copper onnon-enzymatic antioxidants in freshwater fish Channapunctatus (Bloch.). Biological Trace Element Research2003;93:237–48.
Pereira ME, Duarte AC, Millward GE, Abreu SN, Reis MC.Distribution of mercury and other heavy metals in the Ria deAveiro. Quim Anal 1997;16(Suppl.1):S31–5.
Perry SF. The chloride cell: structure and functions in the gill offreshwater fishes. Annu Rev Physiol 1997;59:325–47.
Ramachandran SD, Sweezey MJ, Hodson PV, Boudreau M,Courtenay SC, Lee K, King T, Dixon JA. Influence of salinity andfish species on PAH uptake from dispersed crude oil. Mar PollutBull 2006;52:1182–9.
Rao SS, Neheli TA, Carey JH, Herbert A, Hansen PD. DNA alkalineunwinding assay for monitoring the impact of environmentalgenotoxins. Environ Toxicol Water Quality Int J 1996;11:351–4.
Regoli F, Frenzillib G, Bocchettia R, Annarummab F, Scarcellib V,Fattorinia D, Nigro M. Time-course variations of oxyradicalmetabolism, DNA integrity and lysosomal stability in mussels,Mytilus galloprovincialis, during a field translocationexperiment. Aquatic Toxicol 2004;68:167–78.
Rodriguez-Ariza A, Dorado G, Peinado J, Pueyo C, Lopez-Barea J.Biochemical effects of environmental pollution in fishes fromSpanish south-Atlantic littoral. Biochem Soc Trans1991;19:301S.
Santos MA, Pacheco M, Ahmad I. Anguilla anguilla L. antioxidantsresponses to in situ bleached kraft pulp mill effluent outletexposure. Environ Int 2004;30:301–18.
Santos MA, Pacheco M, Ahmad I. Responses of European eel(Anguilla anguilla L.) circulating phagocytes to an in situ closedpulp mill effluent exposure and its association withorgan-specific peroxidative damage. Chemosphere2006;63:794–801.
Sato M, Bremner I. Oxygen free radicals and metallothionein. FreeRad Biol Med 1993;14:325–37.
Sedlak J, Lindsay HR. Estimation of total, protein-bound andnonprotein sulfhydryl groups in tissues with Ellman's Reagent.Anal Biochem 1968;25:192–205.
Sousa A, Matsudaira C, Takahashi S, Tanabe S, Barroso C.Integrative assessment of organotin contamination in asouthern European estuarine system (Ria de Aveiro,NW Portugal): tracking temporal trends in order toevaluate the effectiveness of the EU ban. Mar Poll Bullet2007;54:1645–53.
Stohs SJ, Bagchi. Oxidative mechanisms in the toxicity of metalsions. Free Radical Bio Med 1995;2:321–36.
Stephensen E, Svavarsson J, Sturve J, Ericson G, Adolfsson-Erici M,Forlin L. Biochemical indicators of pollution exposure inshorthorn sculpin (Myxocephalus scorpius), caught in fourharbours on the southwest coast of Iceland. Aquat Toxicol2000;48:431–42.

307S C I E N C E O F T H E T O T A L E N V I R O N M E N T 4 0 6 ( 2 0 0 8 ) 2 9 8 – 3 0 7
Tietze F. Enzymatic method for quantitative determination ofnanogram amounts of total and oxidized glutathione. AnalBiochem 1969;27:502–22.
Viarengo A, Burlando B, Ceratto N, Panfoli I. Antioxidant role ofmetallothioneins: a comparative overview. Cell Mol Biol2000;46:407–17.
Wang MY, Liehr JG. Lipid hydroperoxide-induced endogenousDNA adducts in hamsters: Possible mechanism of lipid
hydroperoxide-mediated carcinogenesis. Arch BiochemBiophys 1995;316:38–46.
Wang W, Ballatori N. Endogenous glutathione conjugates:occurrence and biological functions. Pharmacol Rev1998;50:335–55.
Zar JH. Biostatistical Analysis. 4th. USA: Prentice HallInternational, Inc.; 1999.




















