
histo-physiological studies on the pituitary gland of some ...
Upload
independentCategory
view
2download
0
ARTICLE IN PRESS
0147-6513/$ - se
doi:10.1016/j.ec
$This invest
of the Univer
manipulations
European Unio�CorrespondE-mail addr
Ecotoxicology and Environmental Safety 70 (2008) 400–410
www.elsevier.com/locate/ecoenv
Histo-cytological responses of Dicentrarchus labrax (L.) followingmercury exposure$
L. Giaria, E. Simonia, M. Manerab, B.S. Dezfulia,�
aDepartment of Biology and Evolution, University of Ferrara, Via Borsari, 46, 44100 Ferrara, ItalybDepartment of Food Science, University of Teramo, I-64100 Teramo, Italy
Received 19 December 2006; received in revised form 18 July 2007; accepted 24 August 2007
Available online 22 October 2007
Abstract
This work deals with the damaging effects of mercury (Hg concentrations 251, 355, 501 mg l�1) on the structure and ultrastructure of
gills, liver, intestine and kidney of farmed European sea bass (Dicentrarchus labrax L., 1758) acutely treated for 24 and 48 h. The
histoarchitecture of the gills of exposed fish was highly modified due to severe oedema, telangiectasia and secondary lamellar fusion. In
hepatocytes and enterocytes hydropic cell swelling, alterations to the endoplasmic reticulum and mitochondria were noted, in addition to
an abundance of myelinoid bodies which were frequently encountered following treatment. In the intestine and renal tubules of exposed
European sea bass, rodlet cells (RCs) displayed ultrastructural modifications.
Statistical analyses were conducted on the number and the size of selected cell types and structures. Following exposure to mercury for
24 and 48 h, the number of chloride cells, RCs and macrophage aggregates were found to have increased significantly in the gills, the
intestine and the head kidney.
r 2007 Elsevier Inc. All rights reserved.
Keywords: Dicentrarchus labrax; Mercury; Histo-cytopathology; Rodlet cells; Biomarker
1. Introduction
Chemicals derived from agricultural operations (pesti-cides and herbicides) and industrial effluents, such asmetals, ultimately find their way into a variety of differentwater bodies and can produce a range of toxic effects inaquatic organisms, ranging from alterations to a single cell,up to changes in whole populations (Bernet et al., 1999).Mercury is a widespread and persistent pollutant occurringin several chemical forms in both freshwater and marineecosystems and is particularly dangerous to living organ-isms because of its ability to accumulate and its toxicity atvery low concentrations (Satoh, 2000; Devlin, 2006).Mercury that accumulates in aquatic systems commonlyresults from the burning of domestic and industrial refuse
e front matter r 2007 Elsevier Inc. All rights reserved.
oenv.2007.08.013
igation was supported by grants from the Italian Ministry
sity and Scientific Research and Technology. All fish
were done in accordance with the Guidelines of the
n Council (86/609/EU) for the use of laboratory animals.
ing author. Fax: +39 0532 291715.
ess: [email protected] (B.S. Dezfuli).
and fossil fuels, and from the activities of agriculture and awide range of industrial and commercial activities(Tchounwou et al., 2003). The adverse consequences thispollutant can effect on aquatic biota, and indirectly tohumans, is of international concern and in 2005, mercurywas ranked third on the CERCLA Priority List ofHazardous Substances.A range of different biomarkers have been used to
indicate the biological effects of certain xenobiotics onaquatic organisms, both in natural environments andunder experimental conditions (Ferrando et al., 2006).Besides the measurement/evaluation of chemical andphysical parameters, evidence arising from histo-cyto-pathological examinations has been increasingly recognisedas valuable tool to evaluate the impact of pollutants onfish (Bernet et al., 1999; Au, 2004). The presence andabundance of certain cell types, such as rodlet cells (RCs),chloride cells (CCs) in tissue sections, and structures suchas macrophage aggregates (MAs), are indicators ofstressful conditions which in the absence of definitivehistopathological changes, could provide information

ARTICLE IN PRESSL. Giari et al. / Ecotoxicology and Environmental Safety 70 (2008) 400–410 401
regarding the general health status of fish (for more detailssee Iger and Abraham, 1997; Agius and Roberts, 2003;Dezfuli et al., 2003a, 2006; Manera and Dezfuli, 2004;Giari et al., 2007). Quantitative studies of morphology areimportant to detect sub-acute effects and to facilitatecomparison of responses at the cell and tissue levels (Jagoeet al., 1996). Histological changes are medium-termresponses to sub-lethal stressors, and histology representsa rapid method to detect effects of toxicants in variousorgans (Bernet et al., 1999; Au, 2004). The gills (Alazemiet al., 1996), liver (Braunbeck, 1998) and kidney (Bernetet al., 1999) are commonly the primary target organs formany chemicals principally because of the role they servewithin the body. The gills are multifunctional; given theirlarge surface area, they are responsible for respiration,osmoregulation, acid–base balance and nitrogenous wasteexcretion, which also makes them extremely sensitive towater contamination (Au, 2004). Biotransformation oforganic xenobiotics, excretion of harmful trace metals,food digestion and storage, and the metabolism of sexhormones are the main hepatic functions (Hinton et al.,2001). Many contaminants, therefore, tend to accumulatein fish liver, exposing the tissues of this organ tocomparatively higher levels than those experienced byother organs (Heath, 1995). In marine fish, kidneys play animportant role in osmotic homeostasis and the excretion ofmetabolites. The kidney, being a highly perfused organ, ispotentially exposed to relatively high levels of toxicchemicals (Au, 2004).
Histopathological changes to various fish due to theaction of mercury has been the subject of numerous studies(Banerjee and Bhattacharya, 1995; Jagoe et al., 1996;Oliveira Ribeiro et al., 2000, 2002; Devlin, 2006) and yetthe damage this heavy metal inflicts on the organs ofEuropean sea bass is largely unknown. The presentinvestigation is part of a set of experiments designed todocument the effects of different stressors on the tissuesof Dicentrarchus labrax, particularly rodlet cells and theirmodifications as suitable biomarkers. The test speciesis the first marine non-salmonid species commerciallycultured in Mediterranean areas (FAO, 2003) and a keyspecies for toxicological testing/evaluation as recom-mended by the Italian Environmental Protection Agency.Earlier studies by the current authors evaluated the cellularresponses of European sea bass to osmotic shock (Giariet al., 2006), to an acute exposure of terbuthylazineand to cadmium (respectively Dezfuli et al., 2006 andGiari et al., 2007). The main objectives of the current studywere, therefore, to (1) characterise the structural andultrastructural effects of mercury on the gills, liver,intestine and kidney of D. labrax, (2) to expand ourknowledge on the toxicity of this metal in fish, (3) toprovide a set of baseline data for the potential use of RCsresponses (either qualitatively or quantitatively) as indica-tors of a changing health status in D. labrax (i.e. asbiomarkers). This study also sets out to look at a numberof other criteria including the presence of histopathological
lesions, the number and the size of MAs, includingcomments on density of CCs.
2. Materials and methods
2.1. Fish maintenance
Twenty-one D. labrax (118.971.5mm (mean LT7S.E.); 14.970.5 g
(mean mass7S.E.)) were obtained from a local fish farm ‘‘Valle Ca’
Zuliani’’ (Pila di Porto Tolle, Rovigo, Italy) and acclimated for 2 weeks in
a 200 l aerated aquarium containing 22 ppt saltwater maintained at
19.670.1 1C under a constant 12:12 L:D photoperiod. Fish were fed daily
with a commercial pelleted feed (crude protein 62%; crude fat 11%; fibre
0.8%; ash 10%; phosphorus 1.1%) but were starved for 48 h prior to the
start of the experiment and throughout its duration.
2.2. Experimental design
Following acclimation and prior to the start of the trial (pre-exposure,
t0), three control fish were euthanased and prepared for histology. The
remaining fish were randomly divided into three equal groups (n ¼ 6 per
group) and allocated to a 20 l static polycarbonate tank filled with the
appropriate concentration of an aqueous solution of Hg (standard
solution for atomic absorbance spectophotometry, Code 497551, Carlo
Erba, Italy). The European sea bass were exposed to nominal mercury
concentrations of 251mg l�1 (tank 1), 355mg l�1 (tank 2), 501mg l�1 (tank3), respectively, and maintained for 48 h with aeration. These sub-lethal
doses were chosen on the basis of preliminary toxicity tests (Dezfuli,
unpublished data) suggestive of inducing toxic effects but not lethally so.
Conditions within each experimental tank were monitored daily with the
concentration of mercury being determined alongside the concentration of
dissolved oxygen (DO) 7.270.6mg l�1, the temperature 19.570.1 1C, pH
7.7070.20 and salinity 2270.0 ppt.
At 24 h (t24) and 48 h (t48) post-exposure, three fish from each tank were
sampled. Fish were killed by a blow to the head and then pithed before
dissection where representative tissue samples from the main organs were
quickly taken.
Summarising, one control group (Ctr) and six exposure groups
(251mg l�1 at t24, 355mg l�1 at t24, 501mg l
�1 at t24 and 251mg l�1 at t48,
355mg l�1 at t48, 501mg l�1 at t48) with three specimens each were enclosed
in the experimental trial.
2.3. Tissue processing
For light microscopy, representative samples of the gills, liver, intestine
and kidney were fixed by immersion in Bouin’s fluid for 8 h, dehydrated in
a graded series of ethanol, paraffin embedded and then cut taking 5 mm-
thick sections which were subsequently stained using haematoxylin/eosin
or Periodic Acid-Schiff (PAS). For electron microscopy, small pieces of
the appropriate tissue were fixed at 4 1C for 2 h in 2% glutaraldehyde
solution buffered with 0.1M sodium cacodylate pH 7.2, then post-fixed in
1% osmium tetroxide in the same buffer for 2 h, dehydrated in a graded
series of ethanol, transferred to propylene oxide and then embedded in an
Epon–Araldite mixture. Semi-thin sections (1.5 mm) were cut on a Reichert
Om U2 ultramicrotome with glass knives and stained with toluidine blue.
Ultra-thin sections were contrasted in a 50% alcohol–uranyl acetate
solution and lead citrate before examination on a Hitachi H-800 electron
microscope operated at 80 kV.
2.4. Biometric and statistical data elaboration
The number of RCs, CCs and the global area occupied by MAs were
evaluated using light microscopy and computerised image analyser
software (Lucia G 4.8, Laboratory Imaging, Praha, Czech Republic).
Twenty, randomly selected, tissue areas ranging from 25 000 to 34 000mm2

ARTICLE IN PRESSL. Giari et al. / Ecotoxicology and Environmental Safety 70 (2008) 400–410402
in dimensions in each fish were screened to count the number of RCs in
paraffin sections taken from gill, kidney and intestine tissues. For counting
CCs, 20 branchial tissue areas measuring between 25 000 and 34 000mm2 in
size were randomly selected from the semi-thin sections of the gills made
on two sea bass in each experimental group and compared to the results
obtained for the control fish. Finally, MA numbers and area, in 10
randomly selected head kidney tissue areas between 25 000 and 34 000mm2
in all the fish, were screened. RC, CC and MA numbers were then
projected to 25 000mm2 to permit inter-organ and inter-cellular compar-
isons. Gaussian distributions of data (i.e. normality) were assessed using
the Komogorov–Smirnov test and normalised using a mean of square root
transformation. Homogeneity of variance was assessed using Levene’s test
and subsequently analysed using parametric tests (one-way ANOVA,
GLM). A Bonferroni post-hoc test and a po0.05 level of significance
selected was used. Given the low number of replicates, the non-parametric
tests, Kruskal–Wallis and Jonckheere–Terpstra tests, were also used for
several independent samples with Monte Carlo exact test extension (SPSS
14.0.2, SPSS Inc., Chicago, IL, USA). Because both assumptions of
normality and homogeneity of variance were met in square root
transformed data, a parametric test was used. In particular, General
Linear Model (GLM) Univariate Analysis procedure was performed using
cells number or MAs area as dependant variable and exposure groups as
factor (one-way ANOVA). Exposure groups were codified as following:
pre-exposure fish ¼ Ctr, 251mg l�1 at t24 ¼ t24 tank 1, 355mg l�1 at
t24 ¼ t24 tank 2, 501mg l�1 at t24 ¼ t24 tank 3, 251mg l�1 at t48 ¼ t48 tank
1, 355mg l�1 at t48 ¼ t48 tank 2, 501mg l�1 at t48 ¼ t48 tank 3. Bonferroni
was adopted as post-hoc test. The level of significance selected was
po0.05. Moreover, the low number of replicates requested also the use of
non-parametric tests: in particular Kruskal–Wallis and Jonckheere–Terp-
stra tests for several independent samples with Monte Carlo exact test
extension were performed. SPSS 14.0.2 (SPSS Inc., Chicago, IL, USA) was
adopted as statistical package.
3. Results
There were no mortalities during the experiment,however, while the control fish displayed a normalswimming behaviour, those of the test groups was erraticand showed respiratory distress.
Fig. 1. (a–f) Tissue structure within the control group of Dicentrarchus
labrax a. Light micrograph showing the gills of unexposed (control) D.
labrax which possess distinct, regular secondary lamellae. Scale
bar ¼ 60mm. b. Transmission electron micrograph of the primary lamella
with few intact CCs (arrows). Scale bar ¼ 5.8mm. c. Liver hepatocytes
3.1. Histo-cytopathology
A histopathological and ultrastructural analysis of thecontrol and mercury-exposed D. labrax was conducted toassess whether the presence of sub-lethal concentrations ofmercury effected changes in the structure of the differenttarget organs, with particular attention being paid to thedensity of CCs, hepatocytes, enterocytes, tubular epithelialcells and RCs.
(arrows) with central spherical nuclei surrounded by an electron dense
cytoplasm (asterisks) rich in organelles. In the peripheral area of the cell
abundant glycogen and a few lipid droplets (arrowheads) can be seen.
Scale bar ¼ 6.4mm. d. Micrograph of intestinal tissue showing mucous
cells (black arrowheads) and intact enterocytes with clearly visible
intercellular junctions (white arrowheads); asterisks indicate microvilli.
Scale bar ¼ 4.6 mm e. A rodlet cell within the intestine, normal in
appearance. Note the capsule (arrow), the basal nucleus (N) and the
typical inclusions (rodlets, asterisks). Scale bar ¼ 1.45mm. f. Micrograph
of kidney tissue showing tubular epithelial cells containing numerous,
well-preserved mitochondria (arrows). Scale bar ¼ 2.9 mm.
3.2. Control fish
Gills had a normal morphological structure: a row ofdistinct and regular secondary lamellae running perpendi-cular to the upper and lower surfaces of each primarylamellae (Fig. 1a). With reference to the different cell types,the epithelial cells covered both types of lamellae, CCs werevisible along the primary lamellae (Fig. 1b) and were rarelyseen in the secondary lamellae. Finally, the mucous cellswere sparsely distributed in the primary lamellar epithe-lium and less frequently along the secondary lamellae.
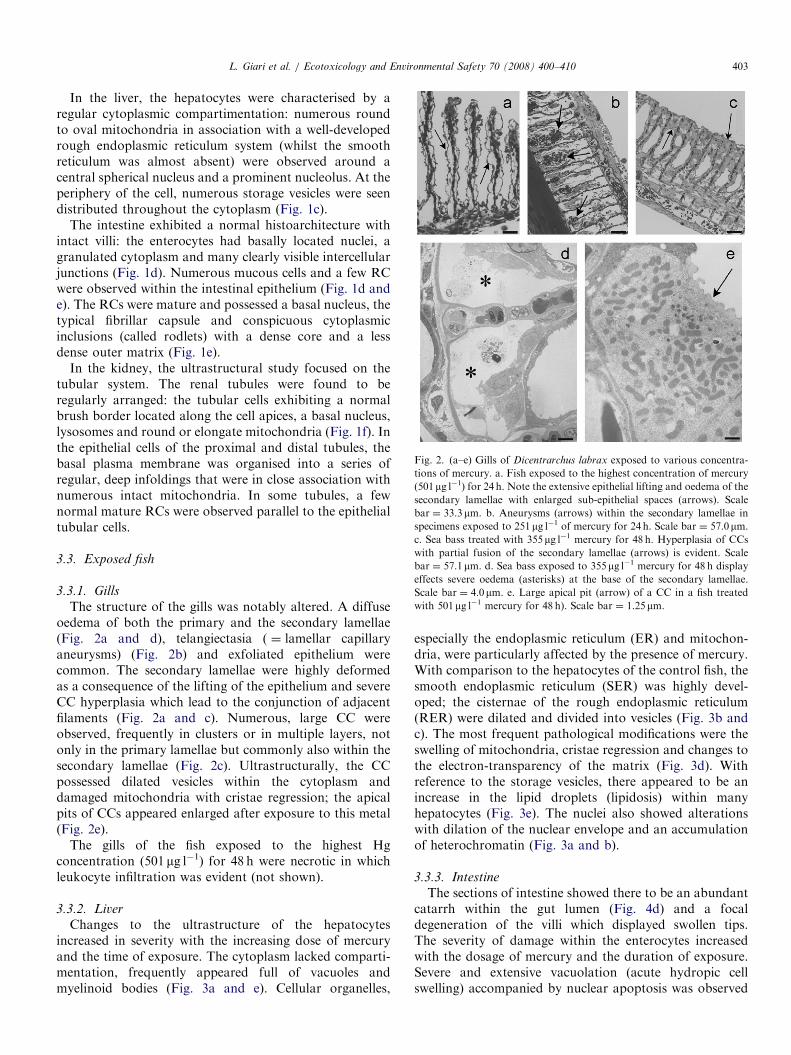
ARTICLE IN PRESS
Fig. 2. (a–e) Gills of Dicentrarchus labrax exposed to various concentra-
tions of mercury. a. Fish exposed to the highest concentration of mercury
(501mg l�1) for 24 h. Note the extensive epithelial lifting and oedema of the
secondary lamellae with enlarged sub-epithelial spaces (arrows). Scale
bar ¼ 33.3mm. b. Aneurysms (arrows) within the secondary lamellae in
specimens exposed to 251mg l�1 of mercury for 24 h. Scale bar ¼ 57.0mm.
c. Sea bass treated with 355mg l�1 mercury for 48 h. Hyperplasia of CCs
with partial fusion of the secondary lamellae (arrows) is evident. Scale
bar ¼ 57.1mm. d. Sea bass exposed to 355mg l�1 mercury for 48 h display
effects severe oedema (asterisks) at the base of the secondary lamellae.
Scale bar ¼ 4.0mm. e. Large apical pit (arrow) of a CC in a fish treated
with 501mg l�1 mercury for 48 h). Scale bar ¼ 1.25mm.
L. Giari et al. / Ecotoxicology and Environmental Safety 70 (2008) 400–410 403
In the liver, the hepatocytes were characterised by aregular cytoplasmic compartimentation: numerous roundto oval mitochondria in association with a well-developedrough endoplasmic reticulum system (whilst the smoothreticulum was almost absent) were observed around acentral spherical nucleus and a prominent nucleolus. At theperiphery of the cell, numerous storage vesicles were seendistributed throughout the cytoplasm (Fig. 1c).
The intestine exhibited a normal histoarchitecture withintact villi: the enterocytes had basally located nuclei, agranulated cytoplasm and many clearly visible intercellularjunctions (Fig. 1d). Numerous mucous cells and a few RCwere observed within the intestinal epithelium (Fig. 1d ande). The RCs were mature and possessed a basal nucleus, thetypical fibrillar capsule and conspicuous cytoplasmicinclusions (called rodlets) with a dense core and a lessdense outer matrix (Fig. 1e).
In the kidney, the ultrastructural study focused on thetubular system. The renal tubules were found to beregularly arranged: the tubular cells exhibiting a normalbrush border located along the cell apices, a basal nucleus,lysosomes and round or elongate mitochondria (Fig. 1f). Inthe epithelial cells of the proximal and distal tubules, thebasal plasma membrane was organised into a series ofregular, deep infoldings that were in close association withnumerous intact mitochondria. In some tubules, a fewnormal mature RCs were observed parallel to the epithelialtubular cells.
3.3. Exposed fish
3.3.1. Gills
The structure of the gills was notably altered. A diffuseoedema of both the primary and the secondary lamellae(Fig. 2a and d), telangiectasia ( ¼ lamellar capillaryaneurysms) (Fig. 2b) and exfoliated epithelium werecommon. The secondary lamellae were highly deformedas a consequence of the lifting of the epithelium and severeCC hyperplasia which lead to the conjunction of adjacentfilaments (Fig. 2a and c). Numerous, large CC wereobserved, frequently in clusters or in multiple layers, notonly in the primary lamellae but commonly also within thesecondary lamellae (Fig. 2c). Ultrastructurally, the CCpossessed dilated vesicles within the cytoplasm anddamaged mitochondria with cristae regression; the apicalpits of CCs appeared enlarged after exposure to this metal(Fig. 2e).
The gills of the fish exposed to the highest Hgconcentration (501 mg l�1) for 48 h were necrotic in whichleukocyte infiltration was evident (not shown).
3.3.2. Liver
Changes to the ultrastructure of the hepatocytesincreased in severity with the increasing dose of mercuryand the time of exposure. The cytoplasm lacked comparti-mentation, frequently appeared full of vacuoles andmyelinoid bodies (Fig. 3a and e). Cellular organelles,
especially the endoplasmic reticulum (ER) and mitochon-dria, were particularly affected by the presence of mercury.With comparison to the hepatocytes of the control fish, thesmooth endoplasmic reticulum (SER) was highly devel-oped; the cisternae of the rough endoplasmic reticulum(RER) were dilated and divided into vesicles (Fig. 3b andc). The most frequent pathological modifications were theswelling of mitochondria, cristae regression and changes tothe electron-transparency of the matrix (Fig. 3d). Withreference to the storage vesicles, there appeared to be anincrease in the lipid droplets (lipidosis) within manyhepatocytes (Fig. 3e). The nuclei also showed alterationswith dilation of the nuclear envelope and an accumulationof heterochromatin (Fig. 3a and b).
3.3.3. Intestine
The sections of intestine showed there to be an abundantcatarrh within the gut lumen (Fig. 4d) and a focaldegeneration of the villi which displayed swollen tips.The severity of damage within the enterocytes increasedwith the dosage of mercury and the duration of exposure.Severe and extensive vacuolation (acute hydropic cellswelling) accompanied by nuclear apoptosis was observed

ARTICLE IN PRESS
Fig. 3. (a–e) The livers of Dicentrarchus labrax exposed to various
concentrations of mercury. a. A highly degenerated hepatocyte in a fish
exposed to 355mg l�1 mercury for 24 h. Acute cell swelling (asterisks) and
accumulation of heterochromatin (arrows) is evident. Scale bar ¼ 1.1 mm.
b. Hepatocyte of a sea bass exposed to 251mg l�1 mercury for 48 h. Note
the development of SER (arrows) and the dilation of the nuclear envelope
(arrowhead). N ¼ nucleus. Scale bar ¼ 1.25mm. c. Hepatocyte within a
fish treated with 501mg l�1 mercury for 48 h which, at high magnification,
displays dilation and vesiculation of the reticulum cisternae. Scale
bar ¼ 0.3 mm. d. Degenerated mitochondria within the hepatocyte of a
sea bass exposed to 355mg l�1 mercury for 48 h. Scale bar ¼ 1.25mm. e.
Presence of numerous and large lipid droplets (asterisks) and myelinoid
bodies (arrows) within a hepatocyte of a fish exposed to 251mg l�1 mercury
for 48 h. Scale bar ¼ 1.7mm.
Fig. 4. (a–e) Alterations to the intestinal tissue of Dicentrarchus labrax
exposed to various concentrations of mercury. a. A rodlet cell in the
intestine of D. labrax exposed to 251mg l�1 mercury for 48 h. The
dissolution of rodlets (arrowheads) and the deformation of the rodlet sacs
(arrow) are evident. Scale bar ¼ 0.85mm. b. Area of damaged intestinal
epithelium in a fish treated with 355mg l�1 mercury for 24 h. Two RCs
(arrows) with highly vacuolated cytoplasms (asterisks) are present. Note
the accumulation of heterochromatin (arrowhead) in the nucleus of one
RC and the anomalous orientation of the rodlet core (star) in the other
cell. Scale bar ¼ 2.7mm. c. Micrograph shows the presence of three rodlet
cells (arrows) and the expulsion of some rodlets (arrowheads) into the gut
lumen of a fish exposed to 251mg l�1 mercury for 24 h. Scale bar ¼ 5.0mm.
d. The gut lumen of sea bass exposed to 501mg l�1 mercury for 48 h which
is filled with catarrh (C) and in which an expulsed rodlet cell is visible.
IM ¼ intestinal mucosa. Scale bar ¼ 3.6 mm. e. Acute cell swelling
(asterisks) of enterocytes in sea bass exposed to 501mg l�1 mercury. An
apoptotic nucleus can be seen (arrow). Scale bar ¼ 3.5 mm.
L. Giari et al. / Ecotoxicology and Environmental Safety 70 (2008) 400–410404
in the intestinal epithelial cells, notably within the fishexposed to the higher concentrations of mercury (Fig. 4e).Cristolysis and myelinoid bodies derived from involutingmitochondria were also commonly observed (not shown).
RC were also affected by exposure to mercury andexhibited cellular modifications irrespective of concentra-tion or the duration of exposure. The occurrence of RCs inthe intestinal epithelium was notably within the moredamaged areas (Fig. 4b) and discharge of their rodlets intothe gut lumen was common (Fig. 4c). RC within the fishexposed to the lowest dose of mercury (251 mg l�1) for 24 hdisplayed alterations especially to their mitochondria,inclusions and nuclei. The cytoplasm was characterisedby intracellular oedema and an abundance of myelinoidbodies whilst the nuclear envelope appeared dilated withan accumulation of heterochromatin within the nuclei(Fig. 4b). The RC displayed different degrees of degenera-
tion ranging from coarctation to deformation of the rodletsac and dissolution of the matrix (Fig. 4a). In someinstances, the rodlet core showed an anomalous orientationand was extruded towards the basal part of the cell, next tothe nucleus (Fig. 4b).
3.3.4. Kidney
In the same section, severe degeneration was noticednear to areas with moderate and reversible changes, in sum,the renal tubules were particularly affected by mercury,especially after 48 h exposure. The intensity of damage seenin the tubular epithelial cells appeared to be linked to thedose of mercury whilst the cellular alterations to the RCsappeared to be the same under the three concentrations ofmercury tested here. The tubular lumen was often occupiedby detached apical expansion of tubular epithelial cells

ARTICLE IN PRESSL. Giari et al. / Ecotoxicology and Environmental Safety 70 (2008) 400–410 405
(‘‘blebs’’) and cellular debris (Fig. 5a). Within the tubularepithelial cells, two different ultrastructural changes to themitochondria were noted; swelling and lysis with electron-transparent matrix and not well outlined cristae (Fig. 5b)or condensation with an electron-dark appearance.Changes to the ER were also marked, with the cisternaeenlarged or vesiculated (Fig. 5c). There was also thepresence of a high number of myelinoid bodies (Fig. 5c)and lysosomes, some of which were degenerated andcoarcted (Fig. 5d). At the highest concentration of mercuryand period of exposure (501 mg l�1 t48), the folded basalpart of the plasma membrane was irregularly arranged andpartially disintegrated (not shown).
The RC exhibited the same ultrastructural alterationsas described previously within intestinal tissue, namelyrarefaction of the cytoplasm around the rodlets (Fig. 5e).
Fig. 5. (a–e) Kidney tissues of Dicentrarchus labrax exposed to various
concentrations of mercury. a. Micrograph showing the tubular lumen
filled with cellular debris (asterisk). Scale bar ¼ 3.3mm. b. Epithelial
tubular cell of a sea bass treated with 355mg l�1 mercury for 48 h. Note the
cytoplasm rich in swollen mitochondria (arrows). Scale bar ¼ 1.25mm. c.
Fish exposed to 355mg l�1 mercury for 48 h showing the presence of
myelinoid bodies (arrowheads) and RER dilated cisternae (arrows) within
an epithelial tubular cell. Scale bar ¼ 0.6 mm. d. Epithelial tubular cells in
the kidney of a sea bass treated with 355mg l�1 mercury for 48 h exhibiting
coarctation of lysosomes (arrowheads). Scale bar ¼ 2.0mm.e. Kidney
tissue of a fish treated with 355mg l�1 mercury for 48 h which shows a
portion of an RC with numerous myelinoid bodies in formation (arrows)
and a dilated nuclear envelope (thick arrow). The rarefaction of the
cytoplasm around the rodlets can be observed (arrowheads). Scale
bar ¼ 0.7 mm.
3.4. Biometry
Figs. 6–10 summarises graphically the enumeration ofthe different cell types in the gills, liver and intestine ofunexposed (control) and mercury exposed (251–501 mg l�1)sea bass.In gills, the number of CCs increased 24 h after exposure
to mercury (treated vs controls, po0.05), with no apparentfurther increases thereafter (Fig. 6). The 501 mg l�1 (t24)treated fish differed significantly from the other treatedgroups (po0.05) (Fig. 6).
t48
tank3
t48
tank2
t48
tank1
t24
tank3
t24
tank2
t24
tank1
Ctr
exposure group
2.40
2.10
1.80
1.20
1.50
95%
CI R
Cs n
Fig. 7. The 95% confidence interval of the intestinal RCs number
(projected to 25 000mm2 and square root transformed) in relation to
exposure group.
t48
tank3
t48
tank2
t48
tank1
t24
tank3
t24
tank2
t24
tank1
Ctr
exposure group
6.00
5.00
4.00
3.00
95
% C
I C
Cs
n
Fig. 6. CCs 95% confidence interval of the branchial CCs number
(projected to 25 000mm2 and square root transformed) in relation to the
exposure group.

ARTICLE IN PRESS
1.25
1.00
0.75
0.50
0.25
95%
CI M
As n
L. Giari et al. / Ecotoxicology and Environmental Safety 70 (2008) 400–410406
In the intestine, the number of RCs increased signifi-cantly after exposure to mercury (treated vs controls,po0.05) with the exception of 251 mg l�1 (t24) (Fig. 7), ascontrary, the augmentation of RCs in the renal tubules wassignificant only in fish of the latter group (t24 tank1 vscontrols, po0.05) (Fig. 8).
Mercury induced a significant augmentation to theglobal area occupied by MAs in the head kidney in allthe mercury exposed fish (treated vs controls, po0.05) withthe exception of 251 mg l�1 (t24) (Fig. 9). This organ showedalso an increment of MA numbers in treated European seabass in comparison to controls (treated vs controls,po0.05), without significant differences between exposuregroups (Fig. 10).
t48
tank3
t48
tank2
t48
tank1
t24
tank3
t24
tank2
t24
tank1
Ctr
exposure group
300.00
250.00
150.00
200.00
100.00
50.00
0.00
95%
CI M
As a
rea
Fig. 9. The 95% confidence interval of the global area of kidney MAs
(projected to 25 000mm2) in relation to the exposure group.
t48
tank3
t48
tank2
t48
tank1
t24
tank3
t24
tank2
t24
tank1
Ctr
exposure group
0.80
1.00
0.60
0.40
0.20
0.00
95%
CI R
Cs n
Fig. 8. The 95% confidence interval of the renal RCs number (projected
to 25 000mm2 and square root transformed) in relation to exposure group.
t48
tank3
t48
tank2
t48
tank1
t24
tank3
t24
tank2
t24
tank1
Ctr
exposure group
Fig. 10. The 95% confidence interval of the kidney MAs number
(projected to 25 000mm2 and square root transformed) in relation to the
exposure group.
4. Discussion
Because of the deleterious impact of pollutants inaquatic ecosystems, the histo-cytological responses of fishesto various classes of xenobiotics need to be determined andcharacterised (Hinton et al., 1978). Following the definitionby Hinton and Lauren (1990), biomarkers will beconsidered as any contaminant-induced physiologicaland/or biochemical modification in a not-too-sensitiveorganism, which leads to the formation of altered structure(lesion) in the cells, tissues or organs.Many changes in water quality are rapidly reflected in
fish gill structure, since this organ is continuously anddirectly exposed to the external environment (Tkatchevaet al., 2004). Numerous types of gill damage have beendocumented in fish experimentally exposed to toxicants orin populations sampled from polluted environments(Alazemi et al., 1996; Pawert et al., 1998; Thophon et al.,2003). Most of the gill histopathological changes arelargely non-specific as confirmed by the occurrence ofsimilar alterations under a wide range of toxicant-exposureconditions (see review of Mallatt, 1985). Hyperplasia withlamellar fusion, telangiectasia, oedema with epitheliallifting and desquamation, as documented in the presentsurvey, are typical lesions of gills in response to organo-chlorines, petroleum compounds, organophosphates, car-bamates, herbicides and heavy metals (Alazemi et al., 1996;GlobalTox, 1997; Jiraungkoorskul et al., 2003; Thophonet al., 2003; Dezfuli et al., 2006; Giari et al., 2007) andsuggest an impairment to the respiratory and osmoregula-tory functioning of the gills (Au, 2004). With reference tothe necrotic gill epithelium noted in this study, Mallatt(1985) reported that this lesion was more often associatedwith heavy metals in comparison to other toxicants andmercury seems especially cytotoxic. The results of this work

ARTICLE IN PRESSL. Giari et al. / Ecotoxicology and Environmental Safety 70 (2008) 400–410 407
confirm the high sensitivity of D. labrax gills to mercury, anobservation that reflects the damage inflicted to the gillswhen exposed to cadmium (Giari et al., 2007). Exposure toheavy metals produces morphological and functionalmodifications in the branchial epithelium (Alazemi et al.,1996; Mauceri et al., 2005); mercury inhibits gill respirationat sub-acute and acute levels (Jagoe et al., 1996).
Almost all previous studies relating to the mercury-induced alterations in gill tissues have been dealt with in apurely qualitative morphological approach, however, thestudy of Jagoe et al. (1996) and the present work considerquantitative changes to the gill tissues by enumeratingchanges in certain cell types such as the number of CC. It iswell documented that high concentrations of toxic sub-stances, particularly ionic stressors, may modify thenumber of this peculiar cell type which is involved inosmoregulation (Oronsaye and Brafield, 1984; Pereira,1988; Giari et al., 2007). In a study using mosquitofish,Gambusia holbrooki, the number of CCs also increased andshowed larger apical surfaces after 14 days exposure to300 nM Hg (Jagoe et al., 1996).
Changes to the ultrastructure of fish livers have provento be suitable and sensitive signs of toxicant-induced injuryand have been used as biomarkers of chemicals inenvironmental risk assessments (Braunbeck and Volkl,1991; Alazemi et al., 1996). There have been numerousreports on histo-cytopathological modifications in livers offish exposed to a wide range of organic compounds andheavy metals (Hinton and Lauren, 1990; GlobalTox, 1997;Au, 2004). The loss of the regular cytoplasmic comparti-mentation is a typical unspecific ultrastructural reaction offish hepatocytes which indicates disturbance of hepatocel-lular homeostasis (Braunbeck, 1998). Some of the altera-tions observed in the hepatic cells in the present study, suchas vacuolar degeneration, dilation of ER and lipid dropletaccumulation, are consistent with those documented inspecimens of D. labrax, Lates calcarifer and Carassius
carassius acutely treated with other heavy metals (e.g. leadand cadmium see Franchini et al., 1991; Thophon et al.,2003, 2004; Giari et al., 2007). A wide spectrum of RERmodifications, appearance of myelinoid bodies, deforma-tion of mitochondria and the nuclear envelope are alsoherbicide-induced alterations in hepatocytes of rainbowtrout, Oncorhynchus mykiss, and carp, Cyprinus carpio, asreported respectively by Braunbeck et al. (1992) and Szareket al. (2000). According to Franchini et al. (1991), theincrease in heterochromatin in the nuclei, in addition to thepathological changes within RER, suggest an impairmentto the synthetic and secretory activities of the cell. Withreference to the extensive development of SER in the seabass exposed to mercury in this study, the same phenom-enon has been seen in the hepatocytes of this and other fishspecies when treated with lead, benzopyrene and certainpesticides (Franchini et al., 1991; Lemaire et al., 1992;Braunbeck, 1998).
Hypertrophy of the ER is a classical response of teleoststo pollutants and is linked to hepatic detoxication
mechanisms (Hawkes, 1980; Lemaire et al., 1992). Themitochondrial structural alterations noticed within thehepatocytes and enterocytes of mercury-exposed sea basswere commonly found in different organs following toxicinjury (Biagianti-Risbourg, 1992; Lemaire et al., 1992), buttheir significance is difficult to evaluate (Biagianti-Risbourgand Bastide, 1995).Based on the cytopathological changes to the liver
documented here, the susceptibility of D. labrax to mercuryis suggested to be high. According to Braunbeck (1998),however, ‘‘for these changes the term ‘‘general intoxicant
adaptation syndrome’’ is proposed, since this set of ultra-
structural modifications of fish hepatocytes may be expected
following exposure to any toxicant and many other disturb-
ing environmental factors other than chemical substances’’.The kidney is one of the main target organs for mercury,
which for a long time has been recognised as nephrotoxicchemical in different fish species (Bano and Hasan, 1990).The swelling of mitochondria and ER cisternae in theepithelial tubular cells of treated D. labrax, could beexplained by the fact that various properties of cellmembranes are impaired by metals with a consequentinhibition of the transporters and the overall osmoregula-tory processes (Ballatori and Boyer, 1996). Proliferation oflysosomes which was observed in this study may occurafter treatment with different xenobiotics and is a typicalresponse to heavy metal exposure (Braunbeck, 1998;Thophon et al., 2004). Their increase in number constitutesone of the primary phases of intoxication processes andmanifestation of cell injury, and reflects the attempt madeby the lysosomes to digest the toxicant and to remove anydamaged organelles (Lemaire et al., 1992; Cheville, 1994).In addition to the proliferation of lysosomes, the formationof numerous myelinoid bodies and an abundance ofcellular debris (all reported in the results) are signs of anincreased turnover of the cellular components underconditions of chemical-induced stress (Hinton et al.,1978; Ghadially, 1997; Braunbeck, 1998).The kidney is one organ in which MAs are known to
occur. MAs are groups of pigment-containing cells, whichare characteristic of heterothermic vertebrates, and in fishare generally localised in the stroma of the haemopoietictissue of the spleen and the head kidney. One of thefunctions of MAs is the sequestration of endogenous andexogenous materials (Agius and Roberts, 2003). As seen inthe current study, an increase in the abundance of renalMAs was registered after exposure to mercury. In otherstudies, hyperplasia of MAs has been generally reported infish species that inhabit low quality environments or inthose that have been exposed to elevated levels of chemicals(Couillard and Hodson, 1996; Meinelt et al., 1997; Dezfuliet al., 2006; Giari et al., 2007). An augmentation of MAs instressed fish might be linked to the increased volume ofcellular debris that results from necrosis or the degenera-tion of tissues as a consequence of the pathological andinflammatory conditions (Au, 2004). Although a numberof confounding biological and physical factors might be

ARTICLE IN PRESSL. Giari et al. / Ecotoxicology and Environmental Safety 70 (2008) 400–410408
responsible for the increase in MAs in fish (Manera et al.,2000), their presence has been proven to be useful butrepresent non-specific cytological indicators of exposure todegraded aquatic environments (Rice, 2001; Dezfuli et al.,2006).
While many studies have documented the histomorpho-logical alterations to the gills, kidneys and livers of fish,especially following exposure to heavy metals (Fracacioet al., 2003; Au, 2004), only a few have investigated theeffect they exert on the intestine (Banerjee and Bhattachar-ya, 1995; Kamunde et al., 2001; Ferrando et al., 2006). Thegastrointestinal tract represents a major route of entry for awide variety of toxicants present in the diet or in the waterthat the fish inhabit (Bano and Hasan, 1990; Lemaire et al.,1992; Banerjee and Bhattacharya, 1995); nonetheless thereis little information relating to the protective mechanismsadopted by the intestine epithelial surfaces of the fishagainst mercury uptake (Oliveira Ribeiro et al., 2002).Light microscopy-based investigations have demonstratedthat there are alterations to the gut of Channa punctatus
and Heteropneustes fossilis following mercury intoxication(Bano and Hasan, 1990; Banerjee and Bhattacharya, 1995)but surprisingly no histopathological changes were de-scribed in Salvelinus alpinus following exposure to inor-ganic mercury and methyl mercury in their feed (OliveiraRibeiro et al., 2002). Only Hawkes (1980), Lemaire et al.(1992), Dezfuli et al. (2006), Giari et al. (2007) and thepresent paper document, with the aid of a TEM, the effectsof different xenobiotics (petroleum, chlorinated biphenyl,benzopyrene, terbuthylazine, cadmium) on the intestinalepithelium of fish. The main perturbations include thenotable presence of vacuoles within the cytoplasm alongwith various inclusions, myelinoid bodies and modifica-tions to the endoplasmic reticulum and mitochondria. Cellswelling is one of the earliest and most conspicuousfeatures of cellular injury after exposure to inorganicmercury (Hg++) or various organomercurial compounds(Kinter and Pritchard, 1977) and was observed frequentlyin the enterocytes of exposed sea bass in this study.According to Ferrando et al. (2006), ‘‘the knowledge of the
gut morphological alterations, both at macroscopic and
microscopic level, can be used as a rapid, easy to detect, and
cost-effective biomarker for a preliminary assessment of
contamination’’.With regard to the RCs, it is shown that these cells
display a number of ultrastructural alterations and increasesignificantly in number within the intestine followingmercury exposure. RCs are exclusive to fish, occur invarious tissues (Manera and Dezfuli, 2004) and theirpresence appears to be linked to stressful situations such asparasitic infestations (Leino, 1996; Dezfuli et al., 2000,2003b; Reite and Evensen, 2006), exposure to chemicals(Iger and Abraham, 1997; Pawert et al., 1998; Balabanova,2000), and physical and mechanical disturbances (Iger andAbraham, 1997; Giari et al., 2006). Numerical augmenta-tion of RCs, supported by statistical analyses, has beenreported in various body organs of Leuciscus cephalus and
D. labrax experimentally exposed to herbicides (Stams
M-4 and terbuthylazine) and cadmium (Dezfuli et al.,2003a, 2006; Giari et al., 2007). Recently, a field study byTkatcheva et al. (2004) documented that branchial RCswere significantly more numerous in perch Perca fluviatilis
from a polluted lake in Russia than in those of thereference sites. All the above-mentioned data tally with thesuggestion of Smith et al. (1995) who considered RCs as acell type produced and recruited in response to sub-lethalenvironmental conditions.Several studies have recorded an increase in the number
of RCs in stressed fishes, whereas very limited informationis available on the effects of xenobiotics on the ultra-structure of these cells. Nonetheless, useful data can befound in Hawkins (1984), Dezfuli et al. (2003a, 2006) andGiari et al. (2007). A high secretory and discharge activityof RCs was common in fishes affected by chemicals inboth natural and laboratory conditions (Hawkins, 1984;Tkatcheva et al., 2004; Giari et al., 2007; Dezfuli, personalobservations) and was also observed in the present survey.Leino (1996) assumed that the phenomenon of rodletsexpulsion could be a consequence of tissue injury. Severalmodifications within RCs of mercury-treated sea bass (i.e.the formation of numerous myelinoid bodies, vacuolisa-tion, granule dissolution and deformation) are suggested torepresent unspecific symptoms of stress, as these cells havealso been recorded in a range of fish species exposed toorganic and inorganic compounds or under pathologicalconditions (Ghadially, 1997; Dezfuli et al., 2003a, 2006;Giari et al., 2007).The present results on RCs confirm the findings from
early experimental investigations with D. labrax (seeDezfuli et al., 2006; Giari et al., 2006, 2007), in that withinalmost all the organs that were examined, the number ofRCs was observed to increase significantly followingexposure to the stressor. The intestine appears to be thebest organ for biometric analyses because RCs are easilyrecognisable here and their high numeric increase linked toconcentration of mercury and the period of exposure. Theultrastructural changes of RCs in exposed fish were similarin the different organs that were examined and were foundin fish exposed to the lowest concentrations of mercury foran arguably short period of exposure (i.e. within 24 h).These findings support the suggestion of Manera andDezfuli (2004) that RCs might be considered sensitivebiomarkers of exposure and provide further evidence forthe involvement of RCs in cellular non-specific responses ofthe fish immune system (Reite and Evensen, 2006).
5. Conclusions
Our findings reveal that, structurally and ultrastructu-rally, the gills and the liver of D. labrax were the mostaffected by exposure to mercury, in terms of both in theseverity of the observed alterations to the tissue and theextent of damage.

ARTICLE IN PRESSL. Giari et al. / Ecotoxicology and Environmental Safety 70 (2008) 400–410 409
Many of the lesions documented in this study agree withthose reported in other fish species under a wide range ofexposure conditions, therefore, it seems plausible that theseeffects reflect physiological adaptation to stress rather thanas specific toxic responses to the concentrations of mercuryconsidered here.
The results suggest that changes in the ultrastructure andnumber of RCs may provide useful information regardingthe environmental conditions and monitoring changes tothem, as specific biomarkers, may provide insights intoevaluating the general health and stress status of fish (Igerand Abraham, 1997; Dezfuli et al., 2003a, 2006; Maneraand Dezfuli, 2004; Giari et al., 2007). MA, similarly, havebeen confirmed as good indicators of exposure tocontaminants (Rice, 2001).
Acknowledgments
The authors would like to thank Mrs Paola Boldrinifrom the Electron Microscopy Centre at the University ofFerrara for technical assistance. We also thank Dr AndyShinn from the Institute of Aquaculture, University ofStirling, UK for comments on this manuscript and helpwith our English.
References
Agius, C., Roberts, R.J., 2003. Melano-macrophage centres and their role
in fish pathology. J. Fish Dis. 26, 499–509.
Alazemi, B.M., Lewis, J.W., Andrews, E.B., 1996. Gill damage in the
freshwater fish Gnathonemus petersii (Family: Mormyridae) exposed to
selected pollutants: an ultrastructural study. Environ. Technol. 17,
225–238.
Au, D.W.T., 2004. The application of histocytopathological biomarkers in
marine pollution monitoring: a review. Mar. Pollut. Bull. 48, 817–834.
Balabanova, L.V., 2000. Rodlet cells of fish in calcium deficiency in the
external environment. Tsitologiia 42 (11), 1048–1052.
Ballatori, N., Boyer, J.L., 1996. Disruption of cell volume regulation by
mercuric chloride is mediated by an increase in sodium permeability
and inhibition of an osmolyte channel in skate hepatocytes. Toxicol.
Appl. Pharmacol. 140, 404–410.
Banerjee, S., Bhattacharya, S., 1995. Histopathological changes induced
by chronic nonlethal levels of elsan, mercury, and ammonia in the
small intestine of Channa punctatus (Bloch). Ecotoxicol. Environ. Saf.
31, 62–68.
Bano, Y., Hasan, M., 1990. Histopathological lesions in the body organs
of cat-fish (Heteropneustes fossilis) following mercury intoxication. J.
Environ. Sci. Health B 25 (1), 67–85.
Bernet, D., Schmidt, H., Meier, W., Burkhardt-Hol, P., Wahli, T., 1999.
Histopathology in fish: proposal for a protocol to assess aquatic
pollution. J. Fish Dis. 22, 25–34.
Biagianti-Risbourg, S., 1992. Interet (eco)toxicologique d’une approche
histocytologique pour la comprehension des responses hepatiques de
Mugilides (Teleosteens) a des contaminations sub-aigues par l’atrazine.
Ichtyophysiol. Acta 15, 205–223.
Biagianti-Risbourg, S., Bastide, J., 1995. Hepatic perturbations induced
by a herbicide (atrazine) in juvenile grey mullet Liza ramada
(Mugilidae, Teleostei): an ultrastructural study. Aquat. Toxicol. 31,
217–229.
Braunbeck, T., 1998. Cytological alterations in fish hepatocytes following
in vivo and in vitro sublethal exposure to xenobiotics—structural
biomarkers of environmental contamination. In: Braunbeck, T.,
Hinton, D.E., Streit, B. (Eds.), Fish Ecotoxicology. Birkhauser Verlag,
Basel, Switzerland, pp. 61–140.
Braunbeck, T., Volkl, A., 1991. Induction of biotransformation in the liver
of eel (Anguilla anguilla L.) by sublethal exposure to dinitro-o-cresol:
an ultrastructural and biochemical study. Ecotoxicol. Environ. Saf. 21,
109–127.
Braunbeck, T., Burkhardt-Holm, P., Gorge, G., Nagel, R., Negele, R.D.,
Storch, V., 1992. Rainbow trout and zebrafish, two models for
continuous toxicity tests: relative sensitivity, species and organ
specificity in cytopathologic reaction of liver and intestines to atrazine.
Schr. Ver Was. Boden Luft. 89, 109–145.
Cheville, N.F., 1994. Ultrastructural Pathology. Iowa State University
Press, Ames.
Couillard, C.M., Hodson, P.V., 1996. Pigmented macrophage aggregates:
a toxic response in fish exposed to bleached-Kraft mill effluent?
Environ. Toxicol. Chem. 15, 1844–1854.
Devlin, E.W., 2006. Acute toxicity, uptake and histopathology of aqueous
methyl mercury to fathead minnow embryos. Ecotoxicology 15,
97–110.
Dezfuli, B.S., Simoni, E., Rossi, R., Manera, M., 2000. Rodlet cells and
other inflammatory cells of Phoxinus phoxinus infected with Raphi-
dascaris acus (Nematoda). Dis. Aquat. Org. 43, 61–69.
Dezfuli, B.S., Giari, L., Simoni, E., Palazzi, D., Manera, M., 2003a.
Alteration of rodlet cells in chub caused by the herbicide Stams M-4
(Propanil). J. Fish Biol. 63, 232–239.
Dezfuli, B.S., Giari, L., Konecny, R., Jaeger, P., Manera, M., 2003b.
Immunohistochemistry, ultrastructure and pathology of gills of
Abramis brama (L.) from lake Mondsee (Austria) due to Ergasilus
sieboldi (Copepoda). Dis. Aquat. Org. 53, 257–262.
Dezfuli, B.S., Simoni, E., Giari, L., Manera, M., 2006. Effects of
experimental terbuthylazine exposure on the cells of Dicentrarchus
labrax (L.). Chemosphere 64, 1684–1694.
FAO, Food and Agriculture Organisation, 2003. Yearbook of Fishery
Statistics: Summary Tables. FAO of the United Nations, Rome.
Ferrando, S., Malsano, M., Parrino, V., Ferrando, T., Girosi, L.,
Tagliaferro, G., 2006. Gut morphology and metallothionein immu-
noreactivity in Liza aurata from different heavy metal polluted
environments. Ital. J. Zool. 73, 7–14.
Fracacio, R., Fenerich Verani, N., Gaeta Espiandola, E.L., Rocha, O.,
Rigolin-Sa, O., Andrade, A., 2003. Alteration on growth and gill
morphology of Danio rerio (Pisces, Ciprinidae) exposed to the toxic
sediments. Braz. Arch. Biol. Technol. 46, 685–695.
Franchini, A., Barbanti, E., Fantin, A.M., 1991. Effects of lead on
hepatocyte ultrastructure in Carassius carassius (L.) var. auratus.
Tissue Cell 23 (6), 893–901.
Ghadially, F.N. (Ed.), 1997. Ultrastructural Pathology of the Cell and
Matrix. Butterworths, London.
Giari, L., Manera, M., Simoni, E., Dezfuli, B.S., 2006. Changes
to chloride and rodlet cells in gills, kidney and intestine of
Dicentrarchus labrax (L.) exposed to reduced salinities. J. Fish Biol.
69, 590–600.
Giari, L., Manera, M., Simoni, E., Dezfuli, B.S., 2007. Cellular alterations
in different organs of European sea bass Dicentrarchus labrax (L.)
exposed to cadmium. Chemosphere 67, 1171–1181.
GlobalTox, 1997. Technical evaluation of histopathology as an environ-
mental monitoring tool for the mining industry in Canada. Report
Prepared for Aquatic Effects Technology Evaluation (AETE) Pro-
gram, Ottawa. Natural Resources Canada by GlobalTox International
Consultants Inc., Ottawa, 153pp.
Hawkes, J.W., 1980. The effects of xenobiotics on fish tissues:
morphological studies. Fed. Proc. 39 (14), 3230–3236.
Hawkins, W.E., 1984. Ultrastructure of rodlet cells: response to cadmium
damage in the kidneys of the spot, Leiostomus xanthurus Lacepede.
Gulf Res. Rep. 7, 365–372.
Heath, A.G., 1995. Water Pollution and Fish Physiology, second ed. CRC
Lewis Publishers, Boca Raton.
Hinton, D.E., Lauren, D.J., 1990. Liver structural alterations accompany-
ing chronic toxicity in fishes: potential biomarkers of exposure. In:

ARTICLE IN PRESSL. Giari et al. / Ecotoxicology and Environmental Safety 70 (2008) 400–410410
McCarty, J.F., Shigart, L.R. (Eds.), Biomarkers of Environmental
Contamination. Lewis Publishers, Boca Raton, pp. 15–57.
Hinton, D.E., Klaunig, J.E., Lipsky, M.M., 1978. PCB-induced altera-
tions in teleost liver: a model for environmental disease in fish. Mar.
Fish. Rev. 40, 47–50.
Hinton, D.E., Segner, H., Braunbeck, T., 2001. Toxic responses of
the liver. In: Schlenk, D., Bensen, W.H. (Eds.), Toxicity in Marine
and Freshwater Teleosts, vol. 1. Taylor & Francis, London,
pp. 224–268.
Iger, Y., Abraham, M., 1997. Rodlet cells in the epidermis of fish exposed
to stressors. Tissue Cell 29, 431–438.
Jagoe, C.H., Faivre, A., Newman, M.C., 1996. Morphological and
morphometric changes in the gills of mosquitofish (Gambusia
holbrooki) after exposure to mercury (II). Aquat. Toxicol. 34, 163–183.
Jiraungkoorskul, W., Upatham, E.S., Kruatrachue, M., Sahaphong, S.,
Vichasri-Grams, S., Pokethitiyook, P., 2003. Biochemical and histo-
pathological effects of glyphosate herbicide on Nile tilapia (Oreochro-
mis niloticus). Environ. Toxicol. 18 (4), 260–267.
Kamunde, C.N., Grosell, M., Lott, J.N.A., Wood, C.M., 2001. Copper
metabolism and gut morphology in rainbow trout (Oncorhynchus
mykiss) during chronic sublethal dietary copper exposure. Can. J. Fish.
Aquat. Sci. 58, 293–305.
Kinter, W.B., Pritchard, J.B., 1977. Altered permeability of cell
membranes. In: Lee, D.H.K. (Ed.), Handbook of Physiology—
Reactions to Environmental Agents. American Physiological Society,
Baltimore, MD, pp. 563–576.
Leino, R.L., 1996. Reaction of rodlet cells to a myxosporean infection in
kidney of the bluegill, Lepomis macrochirus. Can. J. Zool. 74, 217–225.
Lemaire, P., Berhaut, J., Lemaire-Gony, S., Lafaurie, M., 1992.
Ultrastructural changes induced by benzo[a]pyrene in sea bass
(Dicentrarchus labrax) liver and intestine: importance of the intoxica-
tion route. Environ. Res. 57 (1), 59–72.
Mallatt, J., 1985. Fish gill structural changes induced by toxicants and
other irritants: a statistical review. Can. J. Fish. Aquat. Sci. 42,
630–648.
Manera, M., Dezfuli, B.S., 2004. Rodlet cells in teleosts: a new insight into
their nature and functions. J. Fish Biol. 65, 597–619.
Manera, M., Serra, R., Isani, G., Carpene, E., 2000. Macrophage
aggregates in gilthead sea bream fed copper, iron and zinc enriched
diets. J. Fish Biol. 57, 457–465.
Mauceri, A., Fossi, M.C., Leonzio, F., Ancora, S., Minniti, F., Malsano,
M., Lo Cascio, P., Ferrando, S., Fasulo, S., 2005. Stress factors in the
gills of Liza aurata (Perciformes, Mugilidae) living in polluted
environments. Ital. J. Zool. 72, 285–292.
Meinelt, T., Kruger, R., Pietrock, M., Osten, R., Steinberg, C., 1997.
Mercury pollution and macrophage centres in pike (Esox lucius)
tissues. Environ. Sci. Pollut. Res. 4 (1), 32–36.
Oliveira Ribeiro, C.A., Pelletier, E., Pfeiffer, W.C., Rouleau, C., 2000.
Comparative uptake, bioaccumulation, and gill damages of inorganic
mercury in tropical and Nordic freshwater fish. Environ. Res. 83,
286–292.
Oliveira Ribeiro, C.A., Belger, L., Pelletier, E., Rouleau, C., 2002.
Histopathological evidence of inorganic mercury and methyl mercury
toxicity in the arctic charr (Salvelinus alpinus). Environ. Res. 90,
217–225.
Oronsaye, J.A.O., Brafield, A.E., 1984. The effect of dissolved cadmium
on the chloride cells of the gills of stickleback, Gasterosteus aculeatus
L. J. Fish Biol. 25, 253–258.
Pawert, M., Muller, E., Triebskorn, R., 1998. Ultrastructural changes in
fish gills as biomarker to assess small stream pollution. Tissue Cell 30
(6), 617–626.
Pereira, J.J., 1988. Morphological effects of mercury exposure on
windowpane flounder gills as observed by scanning electron micro-
scopy. J. Fish Biol. 33, 571–580.
Reite, O.B., Evensen, Ø., 2006. Inflammatory cells of teleostean fish.
A review focusing on mast cells/eosinophilic granule cells and rodlet
cells. Fish Shellfish Immunol. 20, 192–208.
Rice, C.D., 2001. Fish immunotoxicology: understanding mechanisms of
action. In: Schlenk, D., Bensen, W.H. (Eds.), Target Organ Toxicity in
Marine and Freshwater Teleosts, vol. 2. Taylor & Francis,
London, pp. 96–138.
Satoh, H., 2000. Occupational and environmental toxicology of mercury
and its compounds. Ind. Health 38, 153–164.
Smith, S.A., Caceci, T., Robertson, J.L., 1995. Occurrence of the rodlet
cells and associated lesions in the vascular system of freshwater
angelfish. J. Aquat. Anim. Health 7, 63–69.
Szarek, J., Siwicki, A., Andrzejewska, A., Terech-Majewska, E.,
Banaszkiewicz, T., 2000. Effects of the herbicide RoundupTM on the
ultrastructural pattern of hepatocytes in carp (Cyprinus carpio). Mar.
Environ. Res. 50, 263–266.
Tchounwou, P.B., Ayensu, W.K., Ninashvili, N., Sutton, D., 2003.
Environmental exposure to mercury and its toxicopathologic implica-
tions for public health. Environ. Toxicol. 18, 149–175.
Thophon, S., Kruatrachue, M., Upatham, E.S., Pokethitiyook, P.,
Sahaphong, S., Jaritkhuan, S., 2003. Histopathological alterations of
white sea bass, Lates calcarifer, in acute and subchronic cadmium
exposure. Environ. Pollut. 121, 307–320.
Thophon, S., Pokethitiyook, P., Chalermwat, K., Upatham, E.S.,
Sahaphong, S., 2004. Ultrastructural alterations in the liver and
kidney of white sea bass, Lates calcarifer, in acute and subchronic
cadmium exposure. Environ. Toxicol. 19, 11–19.
Tkatcheva, V., Hyvarinen, H., Kukkonen, J., Ryzhkov, L.P., Holopainen,
I.J., 2004. Toxic effects of mining effluents on fish gills in a subarctic
lake system in NW Russia. Ecotoxicol. Environ. Saf. 57, 278–289.
Copyright © 2022 FDOKUMEN




















