
Seasonal Liza aurata tissue-specific DNA integrity in a multi-contaminated coastal lagoon (Ria de...
7
Seasonal Liza aurata tissue-specific DNA integrity in a multi-contaminated coastal lagoon (Ria de Aveiro, Portugal) M. Oliveira a, * , V.L. Maria b , I. Ahmad a , M. Pacheco a , M.A. Santos a a CESAM & Department of Biology, University of Aveiro, 3810-193 Aveiro, Portugal b CIMA & Faculty of Marine and Environmental Sciences, University of Algarve, Campus de Gambelas, 8005-139 Faro, Portugal article info Keywords: Liza aurata DNA strand breaks Gill Kidney Liver Blood abstract In this study, the DNA integrity of golden grey mullet (Liza aurata) collected in differently contaminated sites of a coastal lagoon, Ria de Aveiro (Portugal), was assessed, over the period of 1 year, using the DNA alkaline unwinding assay, in four different tissues (gill, kidney, liver and blood) and compared to a refer- ence site. The four tissues displayed different DNA integrity basal levels, clearly affected by seasonal fac- tors. Gill and kidney were, respectively, the most and least sensitive tissues. All sites demonstrated the capacity to interfere with DNA integrity. The sites displaying the highest and lowest DNA damage capa- bility were, respectively, Barra (subject to naval traffic) and Vagos (contaminated with polycyclic aro- matic hydrocarbons). In terms of seasonal variability, autumn seems to be the more critical season (more DNA damage) unlike summer when no DNA damage was found in any tissue. Data recommend the continued monitoring of this aquatic system. Ó 2010 Elsevier Ltd. All rights reserved. 1. Introduction The aquatic environment is often the ultimate recipient of a large number of contaminants, mostly resultant from human activ- ities, a large proportion of which potentially genotoxic and carcin- ogenic. The quantification of contaminants in the environment though frequently used in environmental studies may not provide reliable information on the risk to biota, considering the possible presence of unknown substances, the presence of substances in concentrations beyond detection limits, the potential interactions between contaminants as well as differences in bioavailability that may lead to discrepancies between the predictable risk of environ- mental contaminants, based on chemical analyses, and the actual effects. In this perspective, biological systems, that are the target of toxicant action, may provide important information not readily available from chemical analyses (Jha et al., 2000). Biota exposure to contaminants may disrupt normal cellular processes and lead to structural modifications to DNA which may cause subsequent problems for the cell. A high prevalence of unrepaired DNA lesions may lead to incomplete transcription, cellular dysfunction, growth inhibition, aging, weakened immunity and diseases in the organ- ism itself (Woo et al., 2006). The analysis of DNA alterations in aquatic organisms has been shown a suitable method for evaluat- ing the presence of genotoxic contamination, allowing the detection of exposure to low concentrations of contaminants in a wide range of species (Siu et al., 2003; Frenzilli et al., 2009). Thus, its use in monitoring multi-contaminated environments is recommended. Some of the most common DNA lesions are single strand breaks which have been classified as potentially pre-mutagenic lesions (Emmanouil et al., 2007). DNA strand breaks may occur due to di- rect DNA damage caused by exogenous agents, the indirect action of pro-genotoxic agents following biotransformation, oxidative stress and inhibition of DNA synthesis and repair (Eastman and Barry, 1992; Speit and Hartmann, 1995; Lee and Steinert, 2003). Many contaminants may cause damage to DNA by more than one mechanism. Thus, DNA strand breaks may provide information on the presence of a wide range of genotoxic chemicals such as polycyclic aromatic hydrocarbons (PAHs), polychlorinated biphe- nyls (PCBs) and metals (Everaarts, 1995; Mitchelmore and Chip- man, 1998) individually or in mixtures (Bihari and Fafandel, 2004). The genotoxic potential of chemicals depends on the exposure route as well as the properties of the target tissue, namely in terms of accumulation capacity, metabolic activity and basal antioxidant defense levels, cell type heterogeneity, cell cycle, turnover fre- quency and DNA repair efficiency (Lee and Steinert, 2003; Raisud- din and Jha, 2004; Jha et al., 2005). In this perspective, fish tissues such as gill, kidney, liver and blood may respond differently to environmental contaminants, displaying different susceptibility. A previous biomonitoring study focusing on the responses of a native fish species, golden grey mullet (Liza aurata) caught at differently contaminated sites of a coastal lagoon Ria de Aveiro (Portugal) revealed no significant differences between the critical 0025-326X/$ - see front matter Ó 2010 Elsevier Ltd. All rights reserved. doi:10.1016/j.marpolbul.2010.06.011 * Corresponding author. Tel.: +351 234370965; fax: +351 234426408. E-mail address: [email protected] (M. Oliveira). Marine Pollution Bulletin 60 (2010) 1755–1761 Contents lists available at ScienceDirect Marine Pollution Bulletin journal homepage: www.elsevier.com/locate/marpolbul
Transcript of Seasonal Liza aurata tissue-specific DNA integrity in a multi-contaminated coastal lagoon (Ria de...

Marine Pollution Bulletin 60 (2010) 1755–1761
Contents lists available at ScienceDirect
Marine Pollution Bulletin
journal homepage: www.elsevier .com/locate /marpolbul
Seasonal Liza aurata tissue-specific DNA integrity in a multi-contaminatedcoastal lagoon (Ria de Aveiro, Portugal)
M. Oliveira a,*, V.L. Maria b, I. Ahmad a, M. Pacheco a, M.A. Santos a
a CESAM & Department of Biology, University of Aveiro, 3810-193 Aveiro, Portugalb CIMA & Faculty of Marine and Environmental Sciences, University of Algarve, Campus de Gambelas, 8005-139 Faro, Portugal
a r t i c l e i n f o
Keywords:Liza aurataDNA strand breaksGillKidneyLiverBlood
0025-326X/$ - see front matter � 2010 Elsevier Ltd. Adoi:10.1016/j.marpolbul.2010.06.011
* Corresponding author. Tel.: +351 234370965; faxE-mail address: [email protected] (M. Oliveira)
a b s t r a c t
In this study, the DNA integrity of golden grey mullet (Liza aurata) collected in differently contaminatedsites of a coastal lagoon, Ria de Aveiro (Portugal), was assessed, over the period of 1 year, using the DNAalkaline unwinding assay, in four different tissues (gill, kidney, liver and blood) and compared to a refer-ence site. The four tissues displayed different DNA integrity basal levels, clearly affected by seasonal fac-tors. Gill and kidney were, respectively, the most and least sensitive tissues. All sites demonstrated thecapacity to interfere with DNA integrity. The sites displaying the highest and lowest DNA damage capa-bility were, respectively, Barra (subject to naval traffic) and Vagos (contaminated with polycyclic aro-matic hydrocarbons). In terms of seasonal variability, autumn seems to be the more critical season(more DNA damage) unlike summer when no DNA damage was found in any tissue. Data recommendthe continued monitoring of this aquatic system.
� 2010 Elsevier Ltd. All rights reserved.
1. Introduction
The aquatic environment is often the ultimate recipient of alarge number of contaminants, mostly resultant from human activ-ities, a large proportion of which potentially genotoxic and carcin-ogenic. The quantification of contaminants in the environmentthough frequently used in environmental studies may not providereliable information on the risk to biota, considering the possiblepresence of unknown substances, the presence of substances inconcentrations beyond detection limits, the potential interactionsbetween contaminants as well as differences in bioavailability thatmay lead to discrepancies between the predictable risk of environ-mental contaminants, based on chemical analyses, and the actualeffects. In this perspective, biological systems, that are the targetof toxicant action, may provide important information not readilyavailable from chemical analyses (Jha et al., 2000). Biota exposureto contaminants may disrupt normal cellular processes and lead tostructural modifications to DNA which may cause subsequentproblems for the cell. A high prevalence of unrepaired DNA lesionsmay lead to incomplete transcription, cellular dysfunction, growthinhibition, aging, weakened immunity and diseases in the organ-ism itself (Woo et al., 2006). The analysis of DNA alterations inaquatic organisms has been shown a suitable method for evaluat-ing the presence of genotoxic contamination, allowing thedetection of exposure to low concentrations of contaminants in a
ll rights reserved.
: +351 234426408..
wide range of species (Siu et al., 2003; Frenzilli et al., 2009). Thus,its use in monitoring multi-contaminated environments isrecommended.
Some of the most common DNA lesions are single strand breakswhich have been classified as potentially pre-mutagenic lesions(Emmanouil et al., 2007). DNA strand breaks may occur due to di-rect DNA damage caused by exogenous agents, the indirect actionof pro-genotoxic agents following biotransformation, oxidativestress and inhibition of DNA synthesis and repair (Eastman andBarry, 1992; Speit and Hartmann, 1995; Lee and Steinert, 2003).Many contaminants may cause damage to DNA by more thanone mechanism. Thus, DNA strand breaks may provide informationon the presence of a wide range of genotoxic chemicals such aspolycyclic aromatic hydrocarbons (PAHs), polychlorinated biphe-nyls (PCBs) and metals (Everaarts, 1995; Mitchelmore and Chip-man, 1998) individually or in mixtures (Bihari and Fafandel, 2004).
The genotoxic potential of chemicals depends on the exposureroute as well as the properties of the target tissue, namely in termsof accumulation capacity, metabolic activity and basal antioxidantdefense levels, cell type heterogeneity, cell cycle, turnover fre-quency and DNA repair efficiency (Lee and Steinert, 2003; Raisud-din and Jha, 2004; Jha et al., 2005). In this perspective, fish tissuessuch as gill, kidney, liver and blood may respond differently toenvironmental contaminants, displaying different susceptibility.
A previous biomonitoring study focusing on the responses of anative fish species, golden grey mullet (Liza aurata) caught atdifferently contaminated sites of a coastal lagoon Ria de Aveiro(Portugal) revealed no significant differences between the critical

1756 M. Oliveira et al. / Marine Pollution Bulletin 60 (2010) 1755–1761
sites and reference site in gill DNA integrity (Oliveira et al., 2009),despite the significant DNA integrity loss found in kidney at twosites (Oliveira et al., 2010). Despite the known presence of genotox-icants in Ria de Aveiro, Oliveira et al. (submitted for publication-a)found seasonal variations of oxidative DNA damage and erythro-cyte nuclear abnormalities. With the exception of the previouslymentioned studies, the DNA damage dynamics in L. aurata hasnot been studied in complex environments with different contam-ination profiles despite the knowledge that environmental condi-tions may affect xenobiotics bioavailability and input as well asfood availability to fish (Noyes et al., 2009) which may have a dra-matic influence on enzyme systems that activate or detoxify geno-toxicants. Thus, the objective of this study was to assess the tissuespecific (gill, kidney, liver and blood) DNA integrity variations of L.aurata seasonally caught at differently contaminated sites of Ria deAveiro (Portugal) and select the most reliable tissue for genotoxi-city assessment in a multi-contaminated environment. Addition-ally we aimed to rank the assessed sites in terms of the presenceof genotoxicants.
2. Material and methods
2.1. Study area
Ria de Aveiro (Fig. 1) is a coastal lagoon 45 km long (NNE–SSW)and 8.5 km wide, covering a wetland area of approximately 66 (lowtide) to 83 km2 (high tide) which is permanently connected to theocean through a narrow channel (Dias et al., 2001). Sampling siteswere selected on a geographic distribution basis taking into ac-count the various types and sources of contamination as well asthe selection of an unpolluted reference point. Sampling sites were
Fig. 1. Map of Ria de Aveiro (Portugal) with locations of fish-capture sites (j). Therespective coordinates are: reference site (TOR) – 40�44002N, 008�41044W; BAR –40�37042.00N, 8�44035.00W GAF – 40�38038N, 008�41042W; RIO – 40�41008N,008�39041W; LAR – 40�43030N, 008�37043W and VAG – 40�33059N, 008�40055W.
Torreira (TOR), an intermediate region of the longest channel(S. Jacinto-Ovar channel), far from the main polluting sourcesand thus assumed as reference site; Barra (BAR), the initial partof the Mira channel close to the lagoon entrance and subject toconsiderable naval traffic; Gafanha (GAF) situated in the vicinityof a deep-sea fishing port and dry-docks, also connected with themain channel coming from Aveiro city carrying domestic dis-charges; Rio Novo do Principe (RIO), located at the terminal areaof the Vouga River, 6.5 km distant from a pulp/paper mill effluentoutlet, that discharged to this water course during nearly five dec-ades (until the year 2000); Laranjo (LAR), close to a chlor-alkaliplant (6 km), an important source of metal contamination (mainlymercury); Vagos (VAG), located at the terminal part of the Ílhavochannel, receiving municipal and domestic effluents with high lev-els of PAHs.
2.2. Sampling
Liza aurata seasonal samplings were carried out between May2006 and March 2007 in spring (15–18 May 2006), summer(6–11 September 2006), autumn (4–7 December 2006) and winter(6–9 March 2007), using a traditional beach-seine net named‘‘chincha”. L. aurata specimens (n = 6 per site), selected on the basisof their size, had an average length of 14.0 ± 3.0 cm and weighed21.4 ± 3.6 g. Immediately after catching blood was collected fromthe posterior cardinal vein using a heparinised Pasteur pipette, fishwas sacrificed and gill, kidney, liver were removed. All tissues wereimmediately frozen in liquid nitrogen.
At each sampling site, abiotic parameters such as depth, turbid-ity, dissolved oxygen, temperature, pH and salinity were assessed(Table 1) as per the guidelines of APHA (1998).
2.3. DNA integrity assessment
DNA integrity was tested using DNA alkaline unwinding assay.Tissues were placed in TNES (Tris–HCl 10 mM, NaCl 125 mM, EDTA10 mM, SDS 1%, pH 7.5) – urea (5 M) buffer with proteinase K solu-tion (final concentration 0.8 mg/ml). DNA isolation was performedusing a genomic DNA purification kit (Fermentas). DNA integritymeasurements were performed according to Rao et al. (1996) asadopted by Maria et al. (2002). Data from DNA unwinding tech-nique were expressed as F-value [DNA integrity (%)], determinedby applying the following equation:
F ¼ dsdsþ ss
� 100
where ss is the relative fluorescence (measured with a Jasco FP 750spectrofluorometer) of the single-stranded eluent of a sampleminus the single-stranded control blank fluorescence value and dsis the relative fluorescence of the corrected double stranded eluentof the same sample.
2.4. Statistical analysis
Results are expressed as means ± SE (standard error). The DNAintegrity data was transformed prior to statistic analyses accordingto the formula arcsen
pp. Statistical data analysis was done using
Statistica software (StatSoft, Inc., Tulsa, OK, USA). The assumptionsof normality and homogeneity of data were verified. FactorialANOVA was performed in order to assess significant effects at eachsite and seasonal differences. This analysis was followed by posthoc Tukey test to signal significant differences between groups(Zar, 1999). Significance of results was ascertained at a = 0.05.The relationship between DNA integrity loss and abiotic factorswas studied using multiple regression analyses.
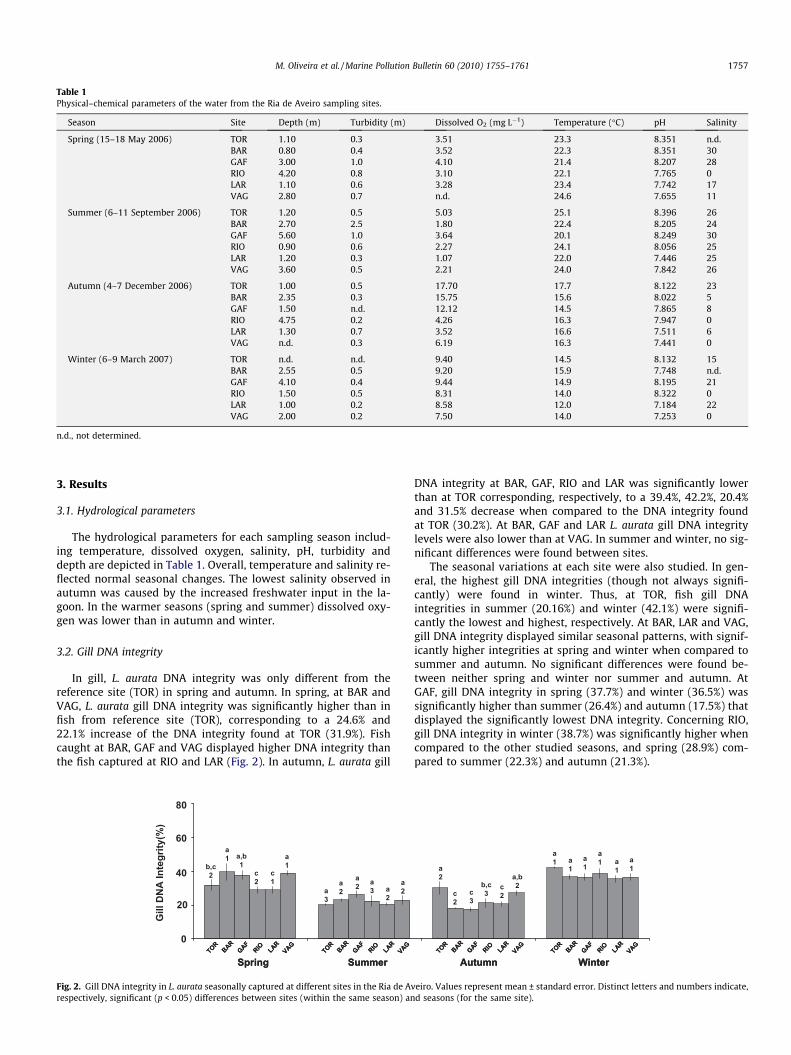
Table 1Physical–chemical parameters of the water from the Ria de Aveiro sampling sites.
Season Site Depth (m) Turbidity (m) Dissolved O2 (mg L�1) Temperature (�C) pH Salinity
Spring (15–18 May 2006) TOR 1.10 0.3 3.51 23.3 8.351 n.d.BAR 0.80 0.4 3.52 22.3 8.351 30GAF 3.00 1.0 4.10 21.4 8.207 28RIO 4.20 0.8 3.10 22.1 7.765 0LAR 1.10 0.6 3.28 23.4 7.742 17VAG 2.80 0.7 n.d. 24.6 7.655 11
Summer (6–11 September 2006) TOR 1.20 0.5 5.03 25.1 8.396 26BAR 2.70 2.5 1.80 22.4 8.205 24GAF 5.60 1.0 3.64 20.1 8.249 30RIO 0.90 0.6 2.27 24.1 8.056 25LAR 1.20 0.3 1.07 22.0 7.446 25VAG 3.60 0.5 2.21 24.0 7.842 26
Autumn (4–7 December 2006) TOR 1.00 0.5 17.70 17.7 8.122 23BAR 2.35 0.3 15.75 15.6 8.022 5GAF 1.50 n.d. 12.12 14.5 7.865 8RIO 4.75 0.2 4.26 16.3 7.947 0LAR 1.30 0.7 3.52 16.6 7.511 6VAG n.d. 0.3 6.19 16.3 7.441 0
Winter (6–9 March 2007) TOR n.d. n.d. 9.40 14.5 8.132 15BAR 2.55 0.5 9.20 15.9 7.748 n.d.GAF 4.10 0.4 9.44 14.9 8.195 21RIO 1.50 0.5 8.31 14.0 8.322 0LAR 1.00 0.2 8.58 12.0 7.184 22VAG 2.00 0.2 7.50 14.0 7.253 0
n.d., not determined.
M. Oliveira et al. / Marine Pollution Bulletin 60 (2010) 1755–1761 1757
3. Results
3.1. Hydrological parameters
The hydrological parameters for each sampling season includ-ing temperature, dissolved oxygen, salinity, pH, turbidity anddepth are depicted in Table 1. Overall, temperature and salinity re-flected normal seasonal changes. The lowest salinity observed inautumn was caused by the increased freshwater input in the la-goon. In the warmer seasons (spring and summer) dissolved oxy-gen was lower than in autumn and winter.
3.2. Gill DNA integrity
In gill, L. aurata DNA integrity was only different from thereference site (TOR) in spring and autumn. In spring, at BAR andVAG, L. aurata gill DNA integrity was significantly higher than infish from reference site (TOR), corresponding to a 24.6% and22.1% increase of the DNA integrity found at TOR (31.9%). Fishcaught at BAR, GAF and VAG displayed higher DNA integrity thanthe fish captured at RIO and LAR (Fig. 2). In autumn, L. aurata gill
60
40
20
80
0
Gill
DN
A In
tegr
ity(%
)
a1 a,b
1a1b,c
2 c2
c1
a3
a2
a2 a
3a2a
2
Spring SummerSpring Summer
TOR
BARGAFRIO LARVAG
TOR
BARGAFRIO LARVAG
TOR
BARGAFRIO LARVAG
TOR
BARGAFRIO LARVAG
Fig. 2. Gill DNA integrity in L. aurata seasonally captured at different sites in the Ria de Avrespectively, significant (p < 0.05) differences between sites (within the same season) an
DNA integrity at BAR, GAF, RIO and LAR was significantly lowerthan at TOR corresponding, respectively, to a 39.4%, 42.2%, 20.4%and 31.5% decrease when compared to the DNA integrity foundat TOR (30.2%). At BAR, GAF and LAR L. aurata gill DNA integritylevels were also lower than at VAG. In summer and winter, no sig-nificant differences were found between sites.
The seasonal variations at each site were also studied. In gen-eral, the highest gill DNA integrities (though not always signifi-cantly) were found in winter. Thus, at TOR, fish gill DNAintegrities in summer (20.16%) and winter (42.1%) were signifi-cantly the lowest and highest, respectively. At BAR, LAR and VAG,gill DNA integrity displayed similar seasonal patterns, with signif-icantly higher integrities at spring and winter when compared tosummer and autumn. No significant differences were found be-tween neither spring and winter nor summer and autumn. AtGAF, gill DNA integrity in spring (37.7%) and winter (36.5%) wassignificantly higher than summer (26.4%) and autumn (17.5%) thatdisplayed the significantly lowest DNA integrity. Concerning RIO,gill DNA integrity in winter (38.7%) was significantly higher whencompared to the other studied seasons, and spring (28.9%) com-pared to summer (22.3%) and autumn (21.3%).
a1a
2 a,b2b,c
3c2
c3
c2
a1 a
1
a1 a
1a1
Autumn WinterAutumn Winter
TOR
BARGAFRIO LARVAG
TOR
BARGAFRIO LARVAG
TOR
BARGAFRIO LARVAG
TOR
BARGAFRIO LARVAG
eiro. Values represent mean ± standard error. Distinct letters and numbers indicate,d seasons (for the same site).

1758 M. Oliveira et al. / Marine Pollution Bulletin 60 (2010) 1755–1761
3.3. Kidney DNA integrity
In spring, at GAF and VAG, L. aurata kidney DNA integrity washigher than at TOR (26.1%), 81.4% and 84.4%, respectively. BAR dis-played the (significantly) lowest DNA integrity (20.0%) whereasGAF and VAG displayed the highest DNA integrity (Fig. 3).
In summer, no kidney DNA integrity loss was found. In fact, kid-ney DNA integrity was significantly higher than TOR, that dis-played a DNA integrity of 28.8%, at all studied sites except BAR(31.2%), respectively, 50.5%, 29%, 41.8% and 40.4% at GAF, RIO,LAR and VAG, respectively.
In autumn, significant differences to TOR (that displayed a DNAintegrity value of 53.9%) were only found at BAR which displayed16.4% higher kidney DNA integrity than TOR. Fish captured atBAR also displayed higher kidney DNA integrity that the fish cap-tured at RIO (54.6%) and VAG (53.3%). Kidney DNA integrity atGAF (61.2%) was also higher than at VAG (53.3%).
In winter, L. aurata kidney DNA integrity at GAF (41.8%) was sig-nificantly lower than at TOR (48.9%).
Concerning seasonal differences, TOR displayed the highestDNA integrities in autumn and winter which were significantlyhigher than spring and summer. At BAR, kidney DNA integritywas, in spring and autumn, respectively, significantly lower andhigher than in the other seasons. At GAF, in autumn, kidney DNAintegrity (61.2%) was significantly higher than in the other studiedseasons, spring (47.3%), summer (43.4%) and winter (41.9%). Thelowest kidney DNA integrities of the fish captured at RIO andLAR (27.1% and 30.0%), were found in spring. At RIO, kidney DNAintegrity in autumn (54.6%) and winter (48.2%) was significantlyhigher than in summer (37.2%) whereas at LAR, kidney DNA in au-tumn (58.6%) was significantly higher than all studied seasons. At
60
40
20
80
0
Kid
ney
DN
A In
tegr
ity(%
)
a2
a1
b2
b3
b3
c4
a2
a,b2
a2
a2
b,c3
c2
Spring SummerSpring Summer
TOR
BARGAFRIO LARVAG
TOR
BARGAFRIO LARVAG
TOR
BARGAFRIO LARVAG
TOR
BARGAFRIO LARVAG
Fig. 3. Kidney DNA integrity in L. aurata seasonally captured at different sites in the Rindicate, respectively, significant (p < 0.05) differences between sites (within the same s
60
40
Spring SummerSpring Summer
20
80
0
Live
r DN
A In
tegr
ity(%
)
a3a,b
3a,b3
a,b3
a,b3b
3
a2
a2
a2 a
2
a2 a
2
TOR
BARGAFRIO LA
RVAG
TOR
BARGAFRIO LA
RVAG
TOR
BARGAFRIO LARVAG
TOR
BARGAFRIO LARVAG
Fig. 4. Liver DNA integrity in L. aurata seasonally captured at different sites in the Riaindicate, respectively, significant (p < 0.05) differences between sites (within the same s
VAG, kidney DNA integrity in summer (40.5%) was significantlylower when compared to the other studied seasons.
3.4. Liver DNA integrity
In spring, L. aurata caught at VAG displayed higher liver DNAintegrity than the fish caught at TOR, corresponding to a 32.1% in-crease in the DNA integrity found at TOR (20.7%) (Fig. 4). No signif-icant differences between sites were found in summer and winter.In autumn, liver DNA integrity in the fish caught at GAF (42.3%),LAR (41.3%) and VAG (43.8%) was significantly lower than at TOR(50.7%). Moreover, at GAF, LAR and VAG liver DNA integrity wasalso lower than at BAR (52.9%).
A general seasonal pattern was found in liver DNA integrity atall sites, pointing out that DNA integrity in spring was significantlylower than in the other seasons. At TOR, BAR, RIO and VAG, theliver DNA integrity of L. aurata caught in autumn and winter wassignificantly higher than in summer. At GAF and LAR, the liverDNA integrity of the fish caught in winter was significantly higherthan in summer.
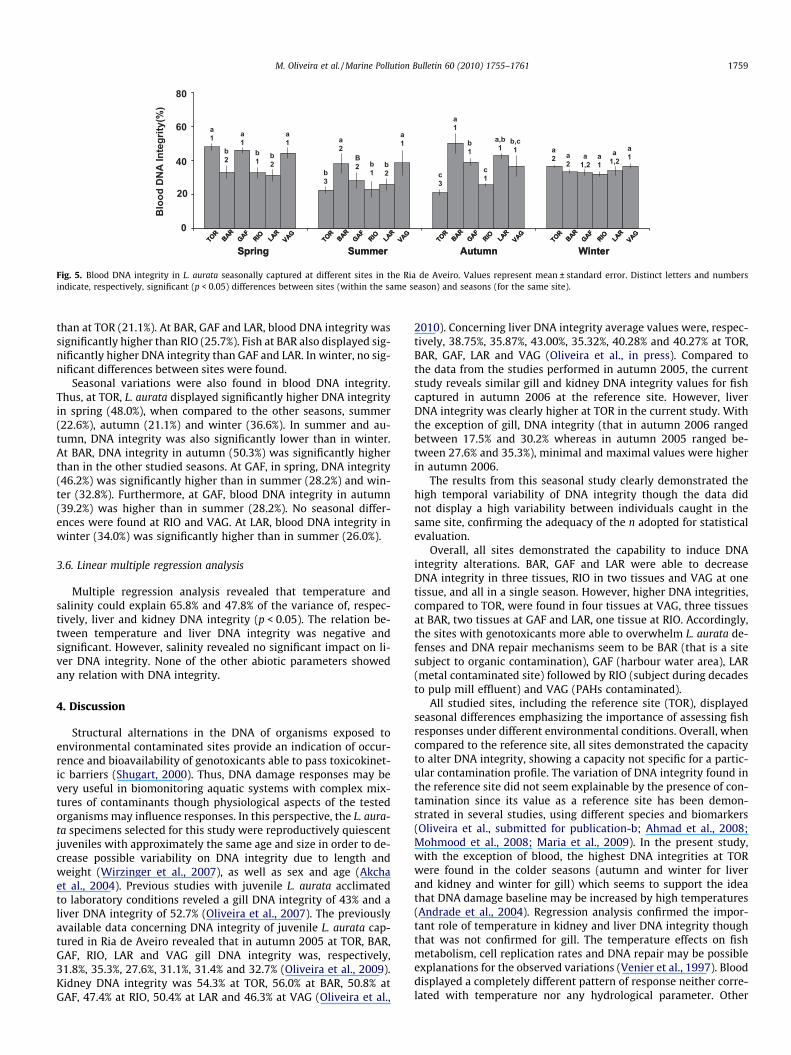
3.5. Blood DNA integrity
In spring, L. aurata blood DNA integrity at BAR (33.0%), RIO(32.8%) and LAR (31.4%) was significantly lower than at TOR(48.0%) as well VAG (44.6%) (Fig. 5). However, in summer, L. auratablood DNA integrity at BAR (38.3%) and VAG (38.7%) was signifi-cantly higher than at TOR (22.6%). The DNA integrity of fish capturedat BAR and GAF was also higher than at GAF (28.2%), RIO (23.3%) andLAR (26.0%). In autumn, L. aurata blood DNA integrity at BAR (50.3%),GAF (39.1%), LAR (25.7%) and VAG (42.3%) was significantly higher
a,b,c1
b,c1
a1
a,b1b,c
1c1 a
1 a,b2
a,b1
a,b2
a,b1b
2
Autumn WinterAutumn Winter
TOR
BARGAFRIO LARVAG
TOR
BARGAFRIO LARVAG
TOR
BARGAFRIO LARVAG
TOR
BARGAFRIO LARVAG
ia de Aveiro. Values represent mean ± standard error. Distinct letters and numberseason) and seasons (for the same site).
Autumn WinterAutumn Winter
a1
a1 a,b,c
1c
1,2 c1,2
c1
a1
a1 a
1a1 a
1
a1
TOR
BARGAFRIO LA
RVAG
TOR
BARGAFRIO LA
RVAG
TOR
BARGAFRIO LARVAG
TOR
BARGAFRIO LARVAG
de Aveiro. Values represent mean ± standard error. Distinct letters and numberseason) and seasons (for the same site).
60
40
20
80
0
Blo
od D
NA
Inte
grity
(%)
a1
a1 a
1b2
b1
b2
a2
a1
b3
B2 b
1b2
a1
a,b1b
1b,c1
c3
c1
a2 a
2a
1,2a1
a1,2
a1
Spring Summer Autumn WinterSpring Summer Autumn Winter
TOR
BARGAFRIO LARVAG
TOR
BARGAFRIO LARVAG
TOR
BARGAFRIO LARVAG
TOR
BARGAFRIO LARVAG
TOR
BARGAFRIO LARVAG
TOR
BARGAFRIO LARVAG
TOR
BARGAFRIO LARVAG
TOR
BARGAFRIO LARVAG
Fig. 5. Blood DNA integrity in L. aurata seasonally captured at different sites in the Ria de Aveiro. Values represent mean ± standard error. Distinct letters and numbersindicate, respectively, significant (p < 0.05) differences between sites (within the same season) and seasons (for the same site).
M. Oliveira et al. / Marine Pollution Bulletin 60 (2010) 1755–1761 1759
than at TOR (21.1%). At BAR, GAF and LAR, blood DNA integrity wassignificantly higher than RIO (25.7%). Fish at BAR also displayed sig-nificantly higher DNA integrity than GAF and LAR. In winter, no sig-nificant differences between sites were found.
Seasonal variations were also found in blood DNA integrity.Thus, at TOR, L. aurata displayed significantly higher DNA integrityin spring (48.0%), when compared to the other seasons, summer(22.6%), autumn (21.1%) and winter (36.6%). In summer and au-tumn, DNA integrity was also significantly lower than in winter.At BAR, DNA integrity in autumn (50.3%) was significantly higherthan in the other studied seasons. At GAF, in spring, DNA integrity(46.2%) was significantly higher than in summer (28.2%) and win-ter (32.8%). Furthermore, at GAF, blood DNA integrity in autumn(39.2%) was higher than in summer (28.2%). No seasonal differ-ences were found at RIO and VAG. At LAR, blood DNA integrity inwinter (34.0%) was significantly higher than in summer (26.0%).
3.6. Linear multiple regression analysis
Multiple regression analysis revealed that temperature andsalinity could explain 65.8% and 47.8% of the variance of, respec-tively, liver and kidney DNA integrity (p < 0.05). The relation be-tween temperature and liver DNA integrity was negative andsignificant. However, salinity revealed no significant impact on li-ver DNA integrity. None of the other abiotic parameters showedany relation with DNA integrity.
4. Discussion
Structural alternations in the DNA of organisms exposed toenvironmental contaminated sites provide an indication of occur-rence and bioavailability of genotoxicants able to pass toxicokinet-ic barriers (Shugart, 2000). Thus, DNA damage responses may bevery useful in biomonitoring aquatic systems with complex mix-tures of contaminants though physiological aspects of the testedorganisms may influence responses. In this perspective, the L. aura-ta specimens selected for this study were reproductively quiescentjuveniles with approximately the same age and size in order to de-crease possible variability on DNA integrity due to length andweight (Wirzinger et al., 2007), as well as sex and age (Akchaet al., 2004). Previous studies with juvenile L. aurata acclimatedto laboratory conditions reveled a gill DNA integrity of 43% and aliver DNA integrity of 52.7% (Oliveira et al., 2007). The previouslyavailable data concerning DNA integrity of juvenile L. aurata cap-tured in Ria de Aveiro revealed that in autumn 2005 at TOR, BAR,GAF, RIO, LAR and VAG gill DNA integrity was, respectively,31.8%, 35.3%, 27.6%, 31.1%, 31.4% and 32.7% (Oliveira et al., 2009).Kidney DNA integrity was 54.3% at TOR, 56.0% at BAR, 50.8% atGAF, 47.4% at RIO, 50.4% at LAR and 46.3% at VAG (Oliveira et al.,
2010). Concerning liver DNA integrity average values were, respec-tively, 38.75%, 35.87%, 43.00%, 35.32%, 40.28% and 40.27% at TOR,BAR, GAF, LAR and VAG (Oliveira et al., in press). Compared tothe data from the studies performed in autumn 2005, the currentstudy reveals similar gill and kidney DNA integrity values for fishcaptured in autumn 2006 at the reference site. However, liverDNA integrity was clearly higher at TOR in the current study. Withthe exception of gill, DNA integrity (that in autumn 2006 rangedbetween 17.5% and 30.2% whereas in autumn 2005 ranged be-tween 27.6% and 35.3%), minimal and maximal values were higherin autumn 2006.
The results from this seasonal study clearly demonstrated thehigh temporal variability of DNA integrity though the data didnot display a high variability between individuals caught in thesame site, confirming the adequacy of the n adopted for statisticalevaluation.
Overall, all sites demonstrated the capability to induce DNAintegrity alterations. BAR, GAF and LAR were able to decreaseDNA integrity in three tissues, RIO in two tissues and VAG at onetissue, and all in a single season. However, higher DNA integrities,compared to TOR, were found in four tissues at VAG, three tissuesat BAR, two tissues at GAF and LAR, one tissue at RIO. Accordingly,the sites with genotoxicants more able to overwhelm L. aurata de-fenses and DNA repair mechanisms seem to be BAR (that is a sitesubject to organic contamination), GAF (harbour water area), LAR(metal contaminated site) followed by RIO (subject during decadesto pulp mill effluent) and VAG (PAHs contaminated).
All studied sites, including the reference site (TOR), displayedseasonal differences emphasizing the importance of assessing fishresponses under different environmental conditions. Overall, whencompared to the reference site, all sites demonstrated the capacityto alter DNA integrity, showing a capacity not specific for a partic-ular contamination profile. The variation of DNA integrity found inthe reference site did not seem explainable by the presence of con-tamination since its value as a reference site has been demon-strated in several studies, using different species and biomarkers(Oliveira et al., submitted for publication-b; Ahmad et al., 2008;Mohmood et al., 2008; Maria et al., 2009). In the present study,with the exception of blood, the highest DNA integrities at TORwere found in the colder seasons (autumn and winter for liverand kidney and winter for gill) which seems to support the ideathat DNA damage baseline may be increased by high temperatures(Andrade et al., 2004). Regression analysis confirmed the impor-tant role of temperature in kidney and liver DNA integrity thoughthat was not confirmed for gill. The temperature effects on fishmetabolism, cell replication rates and DNA repair may be possibleexplanations for the observed variations (Venier et al., 1997). Blooddisplayed a completely different pattern of response neither corre-lated with temperature nor any hydrological parameter. Other

1760 M. Oliveira et al. / Marine Pollution Bulletin 60 (2010) 1755–1761
studies have also found a positive correlation between water tem-perature and DNA integrity in other aquatic species such as zebramussels (Buschini et al., 2003). Autumn seems to be the more crit-ical season, considering that it was the season in which L. auratadisplayed damage at more sites, i.e. gill at four sites and liver atthree sites, probably due to a conjugation of factors such as in-creased bioavailability of sediment associated contaminants linkedwith increased turbidity. In spring, DNA integrity decrease wasonly found in kidney (at one site – GAF) and in blood, that only dis-played DNA integrity decrease in this season (at BAR, RIO and LAR).However, none of the studied tissues displayed significant DNAintegrity decrease in summer. The lower re-suspension of sedi-ment associated contaminants may result in lower bioavailabilityof contaminants. Wirzinger et al. (2007) also found the lowestDNA integrity decrease in summer in three-spined sticklebacks(Gasterosteus aculeatus). Winter was the season when DNA integ-rity was less responsive (both in terms of increased and decreasedintegrity) only displaying a DNA integrity decrease at 1 site – GAF.The lower variability in terms of DNA integrity in winter may berelated with a decreased activity of fish with low water filtrationthrough gill presenting a lower metabolism and consequent de-creased uptake of contaminants. In the four studied tissues, DNAdamage was never found in two consecutive seasons at the samesite. Seasons associated with more extreme temperatures (summerand winter) displayed less damage.
Higher susceptibility to short-term exposure to a PAH (phenan-threne) in terms of oxidative stress was found in L. aurata gill fol-lowed by liver and kidney (Oliveira et al., 2008). However, DNAintegrity loss was found in liver and not in gill (Oliveira et al.,2007) probably related with higher cell regeneration rate of gill.Kilemade et al. (2004) found that juvenile turbot (Scophthalmusmaximus) gill DNA was the more sensitive to damage caused by in-ter-tidal contaminated sediments exposure, followed by liver andblood. On the other hand, Belpaeme et al. (1998) found in S. max-imus exposed to ethyl methanesulphonate that gill cells seemedmore sensitive than blood and the responsiveness of liver cellswas even lower. Overall, the present data suggests that L. auratakidney is the tissue least vulnerable to DNA damage since DNAintegrity loss, though displayed in two seasons (spring and winter),was only found at two sites (BAR in spring and GAF in summer)with similar contamination profile (associated with naval trafficand harbour activities, respectively). This supports the hypothesisof Kilemade et al. (2004) that haematopoietic tissues could bethe least sensitive or responsive cell type. Gill was the most sensi-tive tissue towards DNA damage, displaying damage at more sites,which may be considered expected since they are the first organ tobe exposed to waterborne contaminants displaying low levels ofbasal defenses when compared to kidney (Oliveira et al., 2008).The DNA integrity loss found in blood cells at RIO and LAR withoutconcomitant damage in other tissues does not support the ideathat damage in blood may be reflected in other tissues or vice ver-sa. Kilemade et al. (2004) found correlations between blood and li-ver DNA damages as well as between blood and gill and suggestedthat blood could act as a suitable predictor of DNA damage in thewhole organism.
The data from this study confirms the tissue-specific sensitivityto contaminants that was highly variable with seasons. The use ofgenotoxic responses of resident species in biomonitoring studies,though logistically easy may be also influenced by physiologicaladaptations which potentially render chronically exposed speci-mens less responsive to genotoxic impacts (Frenzilli et al., 2009;Katsumiti et al., 2009) as observed for example in VAG site, thatis contaminated with genotoxicants (Pacheco et al., 2005). Thehigher DNA integrity found in critical sites compared to the refer-ence site may be the result of a more efficient DNA repair system,stimulated by the chronic exposure.
Thus, the genotoxic effects measurements utilizing DNA strandbreaks in resident species may provide misleading results as ob-served for example in summer, where no DNA integrity loss wasfound despite the known presence of contaminants. The lack of dif-ferences between clean and polluted sites was also found in otherstudies. Akcha et al. (2004) and Large et al. (2002) found no differ-ences in mussels (Mytilus sp.) DNA strand breaks despite the differ-ent extents in contamination between sites. Everaarts et al. (1993)found a high incidence in strand breaks in an uncontaminated areain hardhead catfish (Arius felis) and no differences in DNA integrityin relation to contaminated sites. Bombail et al. (2001) also did notfind differences between sites in butterfish (Pholis gunnellus)caught along a pollution gradient. This may be due to higher de-fenses basal levels and activation of cellular defenses, which arethought to prevent accumulation of electrophilic metabolites andfree radicals and hence partially protect DNA and other cellularmacromolecules against oxidation and adduct formation (Rocheret al., 2006). Furthermore, repair enzymes, cell turnover, regulatedabsorption, detoxification, excretion and storage may also play adeterminant in the different responses displayed by L. auratatissues.
5. Conclusions
All sites demonstrated the capacity to alter DNA integrity inrelation to the reference site when compared to the reference site.L. aurata DNA integrity was most affected at BAR whereas VAG dis-played the least DNA damage.
In terms of organ specific responses, as expected, the four or-gans displayed different basal levels, which were clearly affectedby season. Gill and kidney seem to be, respectively, the most andleast sensitive tissues.
The use of resident L. aurata DNA integrity assessment in bio-monitoring studies may provide misleading results since physio-logical adaptations (more efficient defenses and repairmechanisms) may lead to DNA integrities even higher than inthe reference site. The highly dynamic nature of DNA integrity,dependent on an array of processes that may lead to its damageor repair, may limit its usefulness in pollution monitoring if not in-cluded in a set of biomarkers namely including more permanentindications of damage.
Acknowledgments
The financial support from Fundação para a Ciência e Tecnolo-gia – FCT (Government of Portugal) provided through POCI/MAR/61195/2004, SFRH/BD/27584/2006, SFRH/BPD/26970/2006, SFRH/BPD/34326/2006 and by the CESAM – Aveiro University ResearchInstitute are gratefully acknowledged.
References
Ahmad, I., Maria, V.L., Oliveira, M., Serafim, A., Bebianno, M.J., Pacheco, M., Santos,M.A., 2008. DNA damage and lipid peroxidation vs. protection responses in thegill of Dicentrarchus labrax L. from a contaminated coastal lagoon (Ria de Aveiro,Portugal). Sci. Total Environ. 406, 298–307.
Akcha, F., Leday, G., Pfohl-Leszkowicz, A., 2004. Measurement of DNA adducts andstrand breaks in dab (Limanda limanda) collected in the field: effects of biotic(age, sex) and abiotic (sampling site and period) factors on the extent of DNAdamage. Mutat. Res. 552, 197–207.
Andrade, V.M., Freitas, T.R.O., Silva, J., 2004. Comet assay using mullet (Mugil sp.)and sea catfish (Netuma sp.) erythrocytes for the detection of genotoxicpollutants in aquatic environment. Mutat. Res. 560, 57–67.
APHA, 1998. In: Clesceri, L.S., Greenberg, A.E., Eaton, A.D. (Eds.), Standard Methodsfor the Examination of Water and Waste-Water. American Public HealthAssociation, Washington.
Belpaeme, K., Cooreman, K., Kirsch-Volders, M., 1998. Development and validationof the in vivo alkaline comet assay for detecting genomic damage in marineflatfish. Mutat. Res. 415, 167–184.

M. Oliveira et al. / Marine Pollution Bulletin 60 (2010) 1755–1761 1761
Bihari, N., Fafandel, M., 2004. Interspecies differences in DNA single strand breakscaused by benzo(a)pyrene and marine environment. Mutat. Res. 552, 209–217.
Bombail, V., Aw, D., Gordon, E., Batty, J., 2001. Application of the comet andmicronucleus assays to butterfish (Pholis gunnellus) erythrocytes from the Firthof Forth, Scotland. Chemosphere 44, 383–392.
Buschini, A., Carboni, P., Martino, A., Poli, P., Rossi, C., 2003. Effects of temperatureon baseline and genotoxicant-induced DNA damage in haemocytes of Dreissenapolymorpha. Mutat. Res. 537, 81–92.
Dias, J.M., Lopes, J.F., Dekeyser, I., 2001. Lagrangian transport of particles in Ria deAveiro lagoon, Portugal. Phys. Chem. Earth B 26, 721–727.
Eastman, A., Barry, M.A., 1992. The origins of DNA breaks: a consequence of DNAdamage, DNA repair, or apoptosis? Cancer Invest. 10, 229–240.
Emmanouil, C., Sheehan, T.M.T., Chipman, J.K., 2007. Macromolecule oxidation andDNA repair in mussel (Mytilus edulis L.) gill following exposure to Cd and Cr(VI).Aquat. Toxicol. 82, 27–35.
Everaarts, J.M., 1995. DNA integrity as a biomarker of marine pollution: strandbreaks in seastar (Asterias rubens) and dab (Limanda limanda). Mar. Pollut. Bull.31, 431–438.
Everaarts, J.M., Shugart, L.R., Gustin, M.K., Hawkins, W.E., Walker, W.W., 1993.Biological markers in fish – DNA integrity, hematological parameters and liversomatic index. Mar. Environ. Res. 35, 101–107.
Frenzilli, G., Nigro, M., Lyons, B.P., 2009. The comet assay for the evaluation ofgenotoxic impact in aquatic environments. Mutat. Res. 681, 80–92.
Jha, A.N., Cheung, V.V., Foulkes, M.E., Hill, S.J., Depledge, M.H., 2000. Detection ofgenotoxins in the marine environment: adoption and evaluation of anintegrated approach using the embryo-larval stages of the marine mussel,Mytilus edulis. Mutat. Res. 464, 213–228.
Jha, A.N., Dogra, Y., Turner, A., Millward, G.E., 2005. Impact of low doses of tritiumon the marine mussel, Mytilus edulis: genotoxic effects and tissue-specificbioconcentration. Mutat. Res. 586, 47–57.
Katsumiti, A., Domingos, F.X.V., Azevedo, M., da Silva, M.D., Damian, R.C., Almeida,M.I.M., de Assis, H.C.S., Cestari, M.M., Randi, M.A.F., Ribeiro, C.A.O., Freire, C.A.,2009. An assessment of acute biomarker responses in the demersal catfishCathorops spixii after the Vicuña Oil Spill in a harbour estuarine area in SouthernBrazil. Environ. Monit. Assess. 152, 209–222.
Kilemade, M.F., Hartl, M.G.J., Sheehan, D., Mothersill, C., van Pelt, F., O’Halloran, J.,O’Brien, N.M., 2004. Genotoxicity of field-collected inter-tidal sediments fromCork Harbor, Ireland, to juvenile turbot (Scophthalmus maximus L.) as measuredby the comet assay. Environ. Mol. Mutagen. 44, 56–64.
Large, A.T., Shaw, J.P., Peters, L.D., McIntosh, A.D., Webster, L., Mally, A., Chipman,J.K., 2002. Different levels of mussel (Mytilus edulis) DNA strand breaksfollowing chronic field and acute laboratory exposure to polycyclic aromatichydrocarbons. Mar. Environ. Res. 54, 493–497.
Lee, R.F., Steinert, S., 2003. Use of the single cell gel electrophoresis/comet assay fordetecting DNA damage in aquatic (marine and freshwater) animals. Mutat. Res.544, 43–64.
Maria, V.L., Ahmad, I., Oliveira, M., Serafim, A., Bebianno, M.J., Pacheco, M., Santos,M.A., 2009. Wild juvenile Dicentrarchus labrax L. liver antioxidant and damageresponses at Aveiro Lagoon, Portugal. Ecotoxicol. Environ. Saf. 72, 1861–1870.
Maria, V.L., Correia, A.C., Santos, M.A., 2002. Anguilla anguilla L. biochemical andgenotoxic responses to benzo[a]pyrene. Ecotoxicol. Environ. Saf. 53, 86–92.
Mitchelmore, C.L., Chipman, J.K., 1998. DNA strand breakage in aquatic organismsand the potential value of the comet assay in environmental monitoring. Mutat.Res. 399, 135–147.
Mohmood, I., Maria, V.L., Oliveira, M., Ahmad, I., Pacheco, M., Santos, M.A., 2008.Seasonal assessment of a contaminated coastal lagoon (Ria de Aveiro, Portugal)using Dicentrarchus labrax L. erythrocytic nuclear abnormalities. Fresen.Environ. Bull. 17, 1924–1931.
Noyes, P.D., McElwee, M.K., Miller, H.D., Clark, B.W., Van Tiem, L.A., Walcott, K.C.,Erwin, K.N., Levin, E.D., 2009. The toxicology of climate change: environmentalcontaminants in a warming world. Environ. Int. 35, 971–986.
Oliveira, M., Ahmad, I., Maria, V.L., Ferreira, C.S.S., Serafim, A., Bebianno, M.J.,Pacheco, M., Santos, M.A., submitted for publication-a. Evaluation of oxidativeDNA lesions and erythrocytic nuclear abnormalities in wild fish (Liza aurata) asan integrated approach for genotoxicity assessment.
Oliveira, M., Ahmad, I., Maria, V.L., Pacheco, M., Santos, M., in press. Antioxidantresponses versus DNA damage and lipid peroxidation in golden grey mulletliver: a field study at Ria de Aveiro (Portugal). Arch. Environ. Contam. Toxicol.
Oliveira, M., Ahmad, I., Maria, V.L., Pacheco, M., Santos, M.A., 2010. Monitoringpollution of coastal lagoon using Liza aurata kidney oxidative stress and geneticendpoints: an integrated biomarker approach. Ecotoxicology 19, 643–653.
Oliveira, M., Maria, V.L., Ahmad, I., Serafim, A., Bebianno, M.J., Pacheco, M., Santos,M.A., 2009. Contamination assessment of a coastal lagoon (Ria de Aveiro,Portugal) using defence and damage biochemical indicators in gill of Liza aurata– an integrated biomarker approach. Environ. Pollut. 157, 959–967.
Oliveira, M., Maria, V.L., Ahmad, I., Teles, M., Serafim, A., Bebianno, M.J., Pacheco, M.,Santos, M.A., submitted for publication-b. Golden grey mullet and sea bassoxidative DNA damage and clastogenic/aneugenic responses in a contaminatedcoastal Lagoon.
Oliveira, M., Pacheco, M., Santos, M.A., 2007. Cytochrome P4501A, genotoxic andstress responses in golden grey mullet (Liza aurata) following short-termexposure to phenanthrene. Chemosphere 66, 1284–1291.
Oliveira, M., Pacheco, M., Santos, M.A., 2008. Organ specific antioxidant responses ingolden grey mullet (Liza aurata) following a short-term exposure tophenanthrene. Sci. Total Environ. 396, 70–78.
Pacheco, M., Santos, M.A., Teles, M., Oliveira, M., Rebelo, J.E., Pombo, L., 2005.Biotransformation and genotoxic biomarkers in mullet species (Liza sp.) from acontaminated coastal lagoon (Ria de Aveiro, Portugal). Environ. Monit. Assess.107, 133–153.
Raisuddin, S., Jha, A.N., 2004. Relative sensitivity of fish and mammalian cells tosodium arsenate and arsenite as determined by alkaline single-cell gelelectrophoresis and cytokinesis-block micronucleus assay. Environ. Mol.Mutagen. 44, 83–89.
Rao, S.S., Neheli, T.A., Carey, J.H., Herbert, A., Hansen, P.D., 1996. DNA alkalineunwinding assay for monitoring the impact of environmental genotoxins.Environ. Toxicol. Water 11, 351–354.
Rocher, B., Le Goff, J., Peluhet, L., Briand, M., Manduzio, H., Gallois, J., Devier, M.H.,Geffard, O., Gricourt, L., Augagneur, S., Budzinski, H., Pottier, D., André, V.,Lebailly, P., Cachot, J., 2006. Genotoxicant accumulation and cellular defenceactivation in bivalves chronically exposed to waterborne contaminants fromthe Seine River. Aquat. Toxicol. 79, 65–77.
Shugart, L.R., 2000. DNA damage as a biomarker of exposure. Ecotoxicology 9, 329–340.
Siu, W.H.L., Hung, C.L.H., Wong, H.L., Richardson, B.J., Lam, P.K.S., 2003. Exposureand time dependent DNA strand breakage in hepatopancreas of green-lippedmussels (Perna viridis) exposed to Aroclor 1254, and mixtures of B[a]P andAroclor 1254. Mar. Pollut. Bull. 46, 1285–1293.
Speit, G., Hartmann, A., 1995. The contribution of excision repair to the DNA effectsseen in the alkaline single cell gel test (comet assay). Mutagenesis 10, 555–560.
Venier, P., Maron, S., Canova, S., 1997. Detection of micronuclei in gill cells andhaemocytes of mussels exposed to benzo[a]pyrene. Mutat. Res. 390, 33–44.
Wirzinger, G., Weltje, L., Gercken, J., Sordyl, H., 2007. Genotoxic damage in field-collected three-spined sticklebacks (Gasterosteus aculeatus L.): a suitablebiomonitoring tool? Mutat. Res. 628, 19–30.
Woo, S., Kim, S., Yum, S., Yim, U.H., Lee, T.K., 2006. Comet assay for the detection ofgenotoxicity in blood cells of flounder (Paralichthys olivaceus) exposed tosediments and polycyclic aromatic hydrocarbons. Mar. Pollut. Bull. 52, 1768–1775.
Zar, J.H., 1999. Biostatistical Analysis. Prentice Hall, New Jersey.




















