
Effects of additive iron on growth, tissue distribution, haematology and immunology of gilthead sea...
12
Effects of additive iron on growth, tissue distribution, haematology and immunology of gilthead sea bream, Sparus aurata George Rigos • Alexandros Samartzis • Morgane Henry • Eleni Fountoulaki • Efthimia Cotou • John Sweetman • Simon Davies • Ioannis Nengas Received: 28 April 2009 / Accepted: 13 January 2010 / Published online: 3 February 2010 Ó Springer Science+Business Media B.V. 2010 Abstract The comparative effects of iron-supplemented levels on growth, tissue distri- bution, haematology and immunology of gilthead sea bream, Sparus aurata (2 g) were investigated, using four organic (50, 100, 200, 300 mg ORG/kg diet) and one inorganic iron source (200 INOR mg/kg diet). Fish were treated for 12 weeks with the experimental diets and maintained at a water temperature of 19–22°C. Growth (final weight and specific growth rate), tissue distribution (spleen, liver and muscle), haematological parameters (red blood cells, haematocrit, haemoglobin and mean corpuscular haemoglobin concentration) and non-specific immune indexes (respiratory burst activity and antibacterial activity of serum) were analysed. No significant differences were found in growth and iron tissue distribution among the tested groups. Red blood cell counting was statistically higher in fish given 50 ORG, 100 ORG, 200 ORG and 200 INOR feeds. However, haematocrit, haemoglobin and mean corpuscular haemoglobin concentration were not significantly affected by increasing dietary iron. Fish receiving the 100 ORG diet had the best per- formance with respect to the respiratory burst activity and significantly higher values for antibacterial activity of serum were obtained in fish fed with the 300 ORG diet. The present findings provided no clear evidence of the optimum iron concentration. However, there was adequate indication that iron supplementation enhanced the performance of gilthead sea bream, mainly from a haematological and immunological point of view. Keywords Gilthead sea bream Growth performance Sparus aurata Mineral concentration Antibacterial activity Haematological parameters Chemiluminescence G. Rigos (&) A. Samartzis M. Henry E. Fountoulaki E. Cotou I. Nengas Laboratory of Fish Nutrition and Pathology, Institute of Aquaculture, Hellenic Centre for Marine Research, Aghios Kosmas, 16777 Ellinikon, Attiki, Greece e-mail: [email protected] A. Samartzis S. Davies School of Biological Sciences, Plymouth University, Plymouth, Devon PL48AA, UK J. Sweetman Alltech Aqua, Samoli, Livadi, 28200 Lixouri, Cephalonia, Greece 123 Aquacult Int (2010) 18:1093–1104 DOI 10.1007/s10499-010-9326-7
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Effects of additive iron on growth, tissue distribution, haematology and immunology of gilthead sea...

Effects of additive iron on growth, tissue distribution,haematology and immunology of gilthead sea bream,Sparus aurata
George Rigos • Alexandros Samartzis • Morgane Henry •
Eleni Fountoulaki • Efthimia Cotou • John Sweetman •
Simon Davies • Ioannis Nengas
Received: 28 April 2009 / Accepted: 13 January 2010 / Published online: 3 February 2010� Springer Science+Business Media B.V. 2010
Abstract The comparative effects of iron-supplemented levels on growth, tissue distri-
bution, haematology and immunology of gilthead sea bream, Sparus aurata (2 g) were
investigated, using four organic (50, 100, 200, 300 mg ORG/kg diet) and one inorganic
iron source (200 INOR mg/kg diet). Fish were treated for 12 weeks with the experimental
diets and maintained at a water temperature of 19–22�C. Growth (final weight and specific
growth rate), tissue distribution (spleen, liver and muscle), haematological parameters (red
blood cells, haematocrit, haemoglobin and mean corpuscular haemoglobin concentration)
and non-specific immune indexes (respiratory burst activity and antibacterial activity of
serum) were analysed. No significant differences were found in growth and iron tissue
distribution among the tested groups. Red blood cell counting was statistically higher in
fish given 50 ORG, 100 ORG, 200 ORG and 200 INOR feeds. However, haematocrit,
haemoglobin and mean corpuscular haemoglobin concentration were not significantly
affected by increasing dietary iron. Fish receiving the 100 ORG diet had the best per-
formance with respect to the respiratory burst activity and significantly higher values for
antibacterial activity of serum were obtained in fish fed with the 300 ORG diet. The present
findings provided no clear evidence of the optimum iron concentration. However, there
was adequate indication that iron supplementation enhanced the performance of gilthead
sea bream, mainly from a haematological and immunological point of view.
Keywords Gilthead sea bream � Growth performance � Sparus aurata �Mineral concentration � Antibacterial activity � Haematological parameters �Chemiluminescence
G. Rigos (&) � A. Samartzis � M. Henry � E. Fountoulaki � E. Cotou � I. NengasLaboratory of Fish Nutrition and Pathology, Institute of Aquaculture, Hellenic Centre for MarineResearch, Aghios Kosmas, 16777 Ellinikon, Attiki, Greecee-mail: [email protected]
A. Samartzis � S. DaviesSchool of Biological Sciences, Plymouth University, Plymouth, Devon PL48AA, UK
J. SweetmanAlltech Aqua, Samoli, Livadi, 28200 Lixouri, Cephalonia, Greece
123
Aquacult Int (2010) 18:1093–1104DOI 10.1007/s10499-010-9326-7

Introduction
Iron is one of the essential minerals in living organisms involved in several biological
processes such as oxygen transport, cellular respiration and lipid oxidation reactions (Lee
et al. 1981). As in higher vertebrates, the mechanisms involved in the absorption and
regulation of iron in fish are poorly understood (Lall 2002).
Limited iron passage is possible across the gills from the fresh water environment and
thus diet is the major source for iron especially in marine farmed fish (Bury and Grosell
2003). The difference in water-derived capacity for iron absorption between freshwater and
seawater is due to the adaptive physiology and thus variations in the lumen chemistry of
fish inhabiting the two environments (Bury and Grosell 2003). In order to fight dehydra-
tion, marine fish compensate the high accumulation of calcium and magnesium from the
continuous consumption of water by secreting large quantities of bicarbonate resulting in
the formation of alkaline precipitates, which are hostile for the transport of ferrous iron (the
absorvable form) in the intestine (Cooper et al. 2006). Conversely, freshwater fish in their
hypo-osmotic environment aim to reduce water intake and posses low bicarbonate levels
and mildly acidic environment in their gut (Wilson 1999).
The key factors influencing dietary iron absorption in fish include nutrient intake,
chemical form, proportion of organic and inorganic dietary components, digestibility of the
diet, interactions with other dietary components, animal age and health status of the digestive
tract (Andersen et al. 1996; Lall 2002). It is generally accepted that the organic forms of iron
can be absorbed better compared to the inorganic sources (Watanabe et al. 1997).
The requirements of iron have been determined for several species. The minimum
dietary iron requirement has been reported to be between 60 and 100 mg/kg in Atlantic
salmon, Salmo salar (Andersen et al. 1996), 30 mg/kg in channel catfish, Ictaluruspunctatus (Gatlin and Wilson 1986), 170 mg/kg in Japanese eel, Anguilla japonica (Nose
and Arai 1979) and 150 mg/kg in red sea bream, Pagrus major (Sakamoto and Yone
1976). Deficiency of iron induces haematological suppression, reduced growth and poor
feed conversion in farmed fish (Tacon 1992). In particular, experimentally induced lack of
iron may cause microcytic anaemia or low haemoglobin levels in salmonids (Kawatsu
1972; Andersen et al. 1996), channel catfish (Gatlin and Wilson 1986), Japanese eel (Nose
and Arai 1979), yellowtail, Seriola quinqueradiata (Ikeda et al. 1973) and red sea bream.
On the other hand, excessive amounts of dietary iron may not always be beneficial to
farmed fish (Salte et al. 1994; Andersen et al. 1997) and can be connected with increased
bacterial load (Fouz et al. 1994).
Determining the optimum iron feed concentration is therefore a necessary task for the
performance of any farmed fish of interest. Such data is however unavailable for Medi-
terranean mariculture species. Moreover, gilthead sea bream, Sparus aurata, the most
commercialised farmed fish species in this industry, suffers regularly from anaemia and
substantial seasonal mortalities due to infestations of haematophagus monogenean gill
parasites (Microcotyle spp.) (Athanassopoulou et al. 2005). Preliminary in situ results
showed a significant reduction of mortalities with the incorporation of iron supplementa-
tion in the diet (200 mg/kg diet) due to a possible improvement of fish blood parameters
(Nengas, personal communication). Consequently, there is an increasing interest in using
iron dietary supplementation in order to improve disease resistance in gilthead sea bream,
Sparus aurata.
The aim of this study was therefore to investigate the effect of additive iron on gilthead
sea bream, Sparus aurata performances and determine the type (organic vs. inorganic) and
optimum level of iron supplementation in commercial diets in relation to growth,
1094 Aquacult Int (2010) 18:1093–1104
123
haematology, tissue distribution and immunology of this species. This information is to be
used in future experiments involving possibly challenge by natural infections of haema-
tophagus monogenean gill parasites.
Materials and methods
All experiments were carried out in the facilities of the Laboratory of Fish Nutrition &
Pathology of the Institute of Aquaculture in the Hellenic Centre for Marine Research in
Athens (HCMR).
Fish
Two thousand gilthead sea bream, Sparus aurata of an initial average weight of about
1.8 ± 0.1 g and length 2.8 ± 0.2 cm (n = 50) were transported from a local farm (Evia,
Central Greece) at a biomass of 2 kg/m3 with continuous supply of oxygen. Fish were
acclimatised for 1 month before the start of the experiment and were fed with the control
diet used in the experiment (Tables 1, 2). Triplicate groups of 50 fish for each of the six
treatments were weighed and randomly distributed in 18 fibre glass tanks (100 l) receiving
seawater of 38% salinity and maintained at a water temperature of 19–22�C.
Diets and sampling
Six practical diets were prepared in the facilities of HCMR. Diets were supplemented with
four levels of organic iron source (Bioplex, Alltech Inco) at 50, 100, 200 and 300 mg/kg
diet inclusion level and one level of inorganic iron source (Ferrous sulphate, DCM) at
200 mg/kg diet inclusion level (50 ORG, 100 ORG, 200 ORG, 300 ORG and 200 INOR,
respectively). A control diet was also used where no iron was added in the premix. The
composition and iron analysis of the experimental diets are given in Tables 1 and 2,
respectively. Triplicate groups of fish were fed with each one of the diets for 12 weeks at a
constant ratio close to satiation. At the end of the experiment, the total population of fish in
each tank was weighted and growth parameters were calculated. Six fish from each
replicate tank (n = 18) were anesthetised and were used for haematological evaluation
(red blood cell count, haematocrit and haemoglobin concentration), tissue concentration
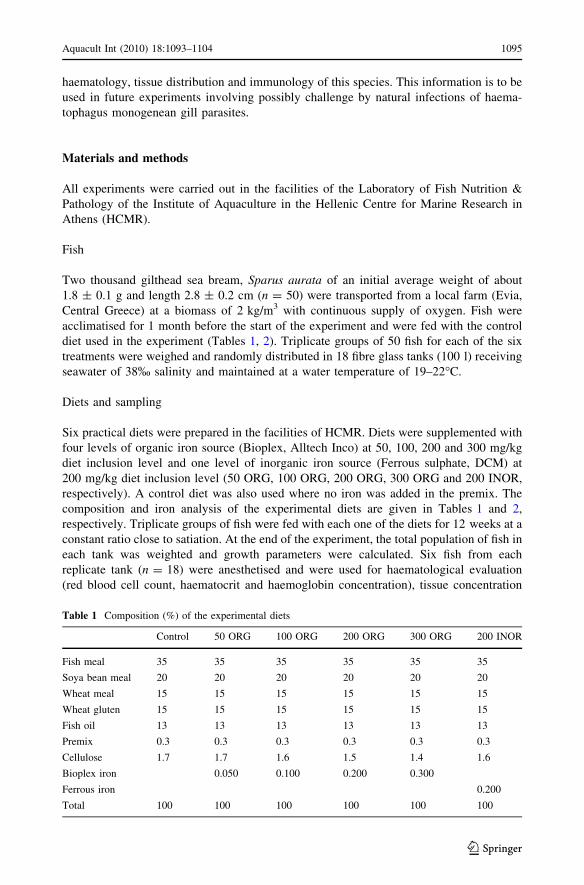
Table 1 Composition (%) of the experimental diets
Control 50 ORG 100 ORG 200 ORG 300 ORG 200 INOR
Fish meal 35 35 35 35 35 35
Soya bean meal 20 20 20 20 20 20
Wheat meal 15 15 15 15 15 15
Wheat gluten 15 15 15 15 15 15
Fish oil 13 13 13 13 13 13
Premix 0.3 0.3 0.3 0.3 0.3 0.3
Cellulose 1.7 1.7 1.6 1.5 1.4 1.6
Bioplex iron 0.050 0.100 0.200 0.300
Ferrous iron 0.200
Total 100 100 100 100 100 100
Aquacult Int (2010) 18:1093–1104 1095
123

analysis (muscle, liver and spleen) and measurement of the immune status of the fish
(whole blood respiratory burst activity and serum antibacterial activity).
Blood sampling
Approximately 1 ml of blood was drawn by a caudal puncture using 23G needles (Mi-
crolance 23G 11/4 0.6 9 30, Becton Dickinson, Saragosa, Spain). Heparin-treated blood
was used for the measure of the respiratory burst activity and for the determination of the
haematological parameters, whereas antibacterial activity was assessed in serum (unhep-
arinised blood was left to coagulate overnight at 4�C, centrifuged twice at 9,000g for
10 min and serum was kept at -80�C until assayed).
Feed and tissue analysis
Feed and tissues (muscle, liver and pooled spleen) were freeze-dried with a CHRIST
GAMMA 1-20 lyophilisator. Spleen samples were pooled due to small quantities collected.
One gram of dried tissue was digested with 10 ml of HNO3 in a microwave-device (CEM
MDS 2100). The drying factor varied from 22 to 25% of the fresh weight. Following
digestion, the sample was diluted with distilled water to 20 ml. A Varian AA 20 Plus flame
Atomic Absorption Spectrophotometer was used for the determination of the metal con-
centrations. The accuracy and precision of the analytical methodology was tested with the
reference material NRCC-Dorm-2 of homogenised dogfish muscle.
Growth
Fish growth was evaluated by calculating the final weight of fish and specific growth rate
(SGR).
SGR ð%=dayÞ ¼ ½lnðW1Þ � lnðW0Þ�total days
� 100:
Haematology
Red blood cell count
Red blood cell count (RBC) was based on Blaxhall (1972). Blood was diluted (1:500) in
Dacie’s solution, and red blood cells were counted in 5 squares of a neubauer chamber.
Cell concentration per ml of blood was deduced.
Haemoglobin concentration
The haemoglobin concentration (Hb) was determined by a miniaturised method using the
Drabkin’s reagent (Drabkin and Austin 1935). In brief, 2 ll of blood was added to 500 ll
Table 2 Measurement of iron concentration (mg/kg) in the experimental diets following preparation
Control 50 ORG 100 ORG 200 ORG 300 ORG 200 INOR
Iron concentration 22.12 71.34 110.54 217.69 308.98 220.24
1096 Aquacult Int (2010) 18:1093–1104
123

of Drabkin’s reagent (Sigma) and 200 ll was transferred to duplicate wells of a transparent
96-flat bottomed well microplate (Greiner). The haemoglobin concentration of the blood
sample was calculated from a standard curve prepared from bovine haemoglobin (Fluka).
The absorbance was determined at 540 nm by a GeniosPro luminometer (TECAN, Aus-
tria). Results were expressed as g/dl blood.
Haematocrit
The haematocrit (Hct) was measured in 18 fish per treatment using duplicate heparinised
75-ll capillary tubes per fish. Capillary tubes were centrifuged at 13,000g for 10 min and
read against a percentage scale. Results were expressed as percentage haematocrit.
Mean corpuscular haemoglobin concentration
Mean corpuscular haemoglobin concentration (MCHC) expressed in g/dl blood was
deduced from the haemoglobin and haematocrit results (MHCH = Hb 9 100/Hct).
Immunology
Antibacterial activity of serum
The antibacterial activity of serum was assessed against a strain of Escherichia colitransformed with a luciferase gene (Nikoskelainen et al. 2002). Fish sera were diluted with
Phosphate Buffer Saline (PBS) and 50 mM Mg2? Ca2? (MgCa, pH 7.4) and increasing
amount (0–100 ll) of the diluted serum were added to 12 wells of a white 96-wells flat-
bottomed microplate (Nunc) giving final serum concentration ranging from 0 to 40 ll/ml
for each fish. Volume was adjusted in each well to 100 ll with PBS and MgCa so that
equal amount of MgCa was present in all wells. Bacteria (E. coli K12pEGFLPLucTet;
kindly provided by S. Verho University of Turku, Finland) were grown overnight to log-
phase at 37�C in LB-broth containing 10 lg tetracycline/ml. Bacterial concentration was
then adjusted to OD450nm of 0.1 and 50 ll were added to the sera. A 3-h incubation at 23�C
allowed bacterial killing, then 100 ll of D-luciferin (Synchem, Germany) at 0.5 mM (in
0.1 M citrate buffer, pH 5.0) were added to each well. The emitted luminescence (RLU)
measured by GeniosPro luminometer (Tecan, Austria) was proportional to the number of
live bacteria. Curves representing RLU as a function of serum concentration (ll/ml)
starting by a plateau and rapidly decreasing to a final plateau were plotted using the Origin
software. From these curves, IC50 (concentration of serum necessary for 50% of bacterial
killing) was calculated, and bactericidal activity of serum in Units/ml was deduced (1000/
IC50).
Chemiluminescence
Induction of the respiratory burst (RB) activity in blood leucocytes was measured directly
from heparinised blood following the method described by Nikoskelainen et al. (2005)
with some modifications. Briefly, 275 ll of diluted blood (1:200) in gHBSS-hep-luminol
(Hanks’ Balanced Salt Solution containing 0.1% gelatin, 5 U/ml of heparin, 100 I.U./ml of
penicillin/streptomycin, 5 lg/ml of oxytetracycline, 0.19 mM luminol, pH 7.4) was dis-
pensed in triplicate wells of a white flat-bottomed 96-well microplate. The background
Aquacult Int (2010) 18:1093–1104 1097
123
luminescence was allowed to stabilise for 10 min. The reaction started with the addition of
25 ll of zymosan at 5 mg/ml in each well, and luminescence was recorded for 2 h every
3 min at 23�C. The peak chemiluminescence was determined and expressed in relative
luminescent units (RLU).
Statistical analysis
Data from each measured parameter was checked for homogeneity of variances (Levene)
and normality (Kolmogorov–Smirnov) prior to statistical analysis. The triplicate tanks of
each diet showed no significant difference (One-Way ANOVA) and could be pooled for
each parameter analysed. Statistical analysis between dietary groups was performed by
One-Way ANOVA followed by a Tukey’s multiple range comparison test using STAT-
GRAPHICS Plus. A student t-test was used to compare the effect of organic and inorganic
source of iron for 200 ORG and 200 INOR. The level of significance for all tests was set at
P \ 0.05.
Results
Iron feed and tissue concentration
The analysis of the iron concentrations in the tested diets revealed values of expected
magnitude, since the measured levels were slightly above the supplemented concentrations
with concentration in the control non-supplemented diet of 22.12 mg/kg (Table 2). Iron
concentrations in the analyzed tissues are shown in Table 3. The iron levels in the three
tested tissue of fish fed with the 200 ORG diet displayed the highest values with however
no statistical difference in muscle (P = 0.507) and liver (P = 0.369) among the experi-
mental groups.
Growth
The results concerning the growth performance of tested fish are given in Table 4. Growth
was not affected by iron supplementation, since no statistical differences were observed in
final weight (P = 0.423) and SGR (P = 0.511) between the experimental groups.
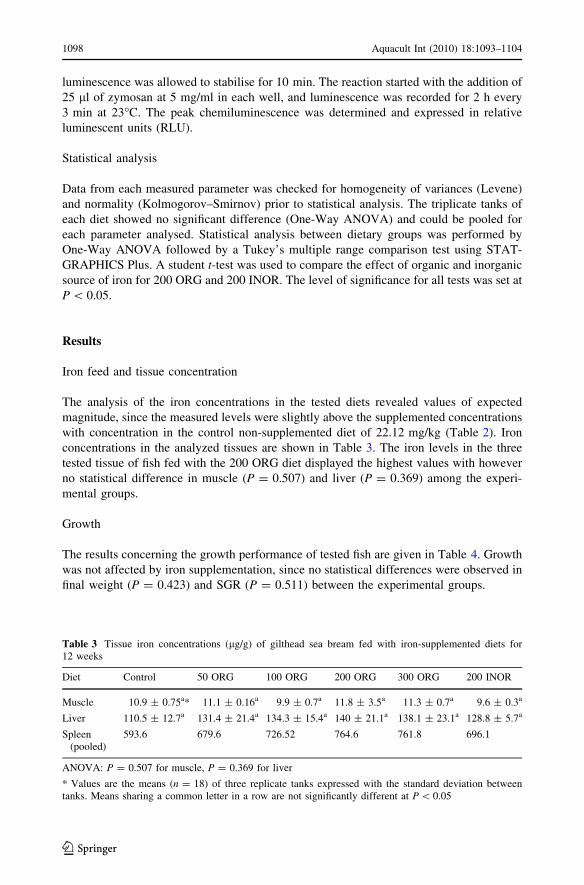
Table 3 Tissue iron concentrations (lg/g) of gilthead sea bream fed with iron-supplemented diets for12 weeks
Diet Control 50 ORG 100 ORG 200 ORG 300 ORG 200 INOR
Muscle 10.9 ± 0.75a* 11.1 ± 0.16a 9.9 ± 0.7a 11.8 ± 3.5a 11.3 ± 0.7a 9.6 ± 0.3a
Liver 110.5 ± 12.7a 131.4 ± 21.4a 134.3 ± 15.4a 140 ± 21.1a 138.1 ± 23.1a 128.8 ± 5.7a
Spleen(pooled)
593.6 679.6 726.52 764.6 761.8 696.1
ANOVA: P = 0.507 for muscle, P = 0.369 for liver
* Values are the means (n = 18) of three replicate tanks expressed with the standard deviation betweentanks. Means sharing a common letter in a row are not significantly different at P \ 0.05
1098 Aquacult Int (2010) 18:1093–1104
123
Haematological analysis
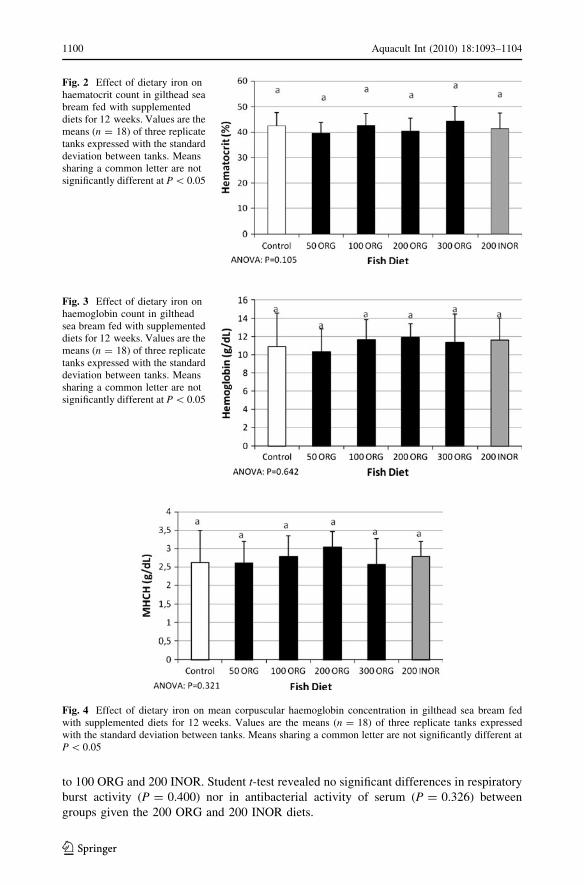
The haematological analysis showed significantly higher levels (P = 0.003) of RBC in fish
fed with the 50, 100, 200 ORG and 200 INOR diets compared to the fish receiving the
control feed (Fig. 1), whereas Hct (P = 0.105; Fig. 2), Hb (P = 0.642; Fig. 3) and MHCH
(P = 0.321; Fig. 4) measurements showed no significant differences between the experi-
mental groups. A tendency for increased Hb and MCHC was observed in fish fed with 200
ORG. Student t-test revealed no significant differences in Htc (P = 0.435), in Hb
(P = 0.687) nor in MHCH (P = 0.101) between groups given the 200 ORG and 200
INOR diets.
Immunological parameters
Respiratory burst activity showed no significant increase in response to the inclusion of
dietary iron (P = 0.404; Fig. 5). A tendency for increased levels was however present in
fish fed with 100 ORG and 200 ORG, but large standard deviations between fish masked
any significant differences. Antibacterial activity of serum was significantly higher
(P = 0.004; Fig. 6) in fish given the highest dosage of organic iron (300 ORG) compared
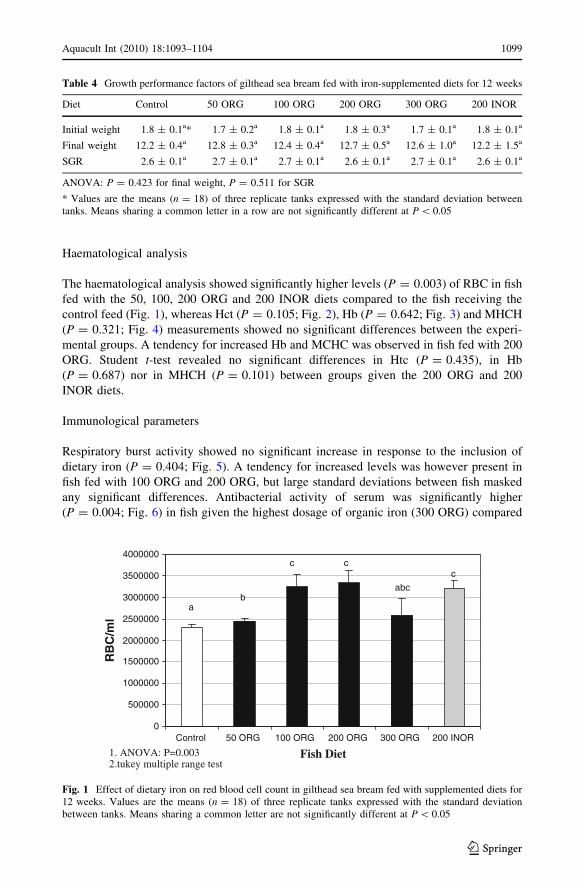
Table 4 Growth performance factors of gilthead sea bream fed with iron-supplemented diets for 12 weeks
Diet Control 50 ORG 100 ORG 200 ORG 300 ORG 200 INOR
Initial weight 1.8 ± 0.1a* 1.7 ± 0.2a 1.8 ± 0.1a 1.8 ± 0.3a 1.7 ± 0.1a 1.8 ± 0.1a
Final weight 12.2 ± 0.4a 12.8 ± 0.3a 12.4 ± 0.4a 12.7 ± 0.5a 12.6 ± 1.0a 12.2 ± 1.5a
SGR 2.6 ± 0.1a 2.7 ± 0.1a 2.7 ± 0.1a 2.6 ± 0.1a 2.7 ± 0.1a 2.6 ± 0.1a
ANOVA: P = 0.423 for final weight, P = 0.511 for SGR
* Values are the means (n = 18) of three replicate tanks expressed with the standard deviation betweentanks. Means sharing a common letter in a row are not significantly different at P \ 0.05
0
500000
1000000
1500000
2000000
2500000
3000000
3500000
4000000
Control 50 ORG 100 ORG 200 ORG 300 ORG 200 INOR
Fish Diet
RB
C/m
l
a
c
b
c
abc
c
1. ANOVA: P=0.003 2.tukey multiple range test
Fig. 1 Effect of dietary iron on red blood cell count in gilthead sea bream fed with supplemented diets for12 weeks. Values are the means (n = 18) of three replicate tanks expressed with the standard deviationbetween tanks. Means sharing a common letter are not significantly different at P \ 0.05
Aquacult Int (2010) 18:1093–1104 1099
123
to 100 ORG and 200 INOR. Student t-test revealed no significant differences in respiratory
burst activity (P = 0.400) nor in antibacterial activity of serum (P = 0.326) between
groups given the 200 ORG and 200 INOR diets.
Fig. 2 Effect of dietary iron onhaematocrit count in gilthead seabream fed with supplementeddiets for 12 weeks. Values are themeans (n = 18) of three replicatetanks expressed with the standarddeviation between tanks. Meanssharing a common letter are notsignificantly different at P \ 0.05
Fig. 3 Effect of dietary iron onhaemoglobin count in giltheadsea bream fed with supplementeddiets for 12 weeks. Values are themeans (n = 18) of three replicatetanks expressed with the standarddeviation between tanks. Meanssharing a common letter are notsignificantly different at P \ 0.05
Fig. 4 Effect of dietary iron on mean corpuscular haemoglobin concentration in gilthead sea bream fedwith supplemented diets for 12 weeks. Values are the means (n = 18) of three replicate tanks expressedwith the standard deviation between tanks. Means sharing a common letter are not significantly different atP \ 0.05
1100 Aquacult Int (2010) 18:1093–1104
123

Discussion
The present study represents the first investigation on the effects of iron supplementation in
Mediterranean farmed fish species. No mortalities were recorded in any of the treated
groups during experimentation. Iron addition either as organic (up to 300 mg/kg diet) or
inorganic (200 mg/kg) form was not found to improve final weight (P = 0.423) and SGR
(P = 0.511) of gilthead sea bream, Sparus aurata indicating that growth cannot be used as
a criterion for evaluating iron supplementation in this species. In accordance with this
finding, Atlantic salmon (Andersen et al. 1996, 1997, 1998) and red sea bream (Sakamoto
and Yone 1976) showed no significant weight gain when receiving additional iron in their
feeds, whereas channel catfish showed a growth retardation when given a purified diet
without iron supplementation (Gatlin and Wilson 1986). These data suggested that iron
present in the dietary fish meal provides the fish with iron levels sufficient for adequate
growth of Sparus aurata.
Fig. 5 Effect of dietary iron on respiratory burst activity in blood of gilthead sea bream fed withsupplemented diets for 12 weeks. Values are the means (n = 18) of three replicate tanks expressed with thestandard deviation between tanks. Means sharing a common letter are not significantly different at P \ 0.05
Fig. 6 Effect of dietary iron on antibacterial activity in serum of gilthead sea bream fed with supplementeddiets for 12 weeks. Values are the means (n = 18) of three replicate tanks expressed with the standarddeviation between tanks. Means sharing a common letter are not significantly different at P \ 0.05
Aquacult Int (2010) 18:1093–1104 1101
123

Although no significant differences were revealed in iron tissue concentrations among
the experimental groups (P = 0.507 and P = 0.369 for muscle and liver, respectively), an
increasing trend was apparent with the increase of dietary organic iron (up to 200 mg/kg
diet) with, however, lower levels measured in fish receiving the highest organic iron
concentration (300 mg/kg diet). It was previously demonstrated that iron absorption
capability and thus tissue levels in Atlantic salmon are not proportional to the dietary levels
(Andersen et al. 1996). In particular, the latter study revealed that a stabilisation in iron
storage occurred when high dietary iron was supplemented (100–400 mg/kg). This was in
accordance with the previous findings in the same species studied by Rasmussen (1994),
where similar amounts of stored iron were measured despite wide differences in dietary
iron intake (210–307 mg/kg). These results support a previous hypothesis that there is a
regulation mechanism for iron metabolism in fish that is not thoroughly known yet
(Andersen et al. 1996). Even if not significantly, organic iron sources in the present work
showed higher availability compared to the inorganic iron form as seen in all three tissues
tested confirming the fact that organic iron is more available than inorganic sources in fish.
Haematological analyses revealed clear differences among the treated groups only in
RBC counts (P = 0.003) where iron levels from both organic and inorganic sources up to
200 mg/kg feed displayed a positive effect on this parameter. This probably indicates that
the iron content of the control feed (22 mg/kg) may cover some of the haematological
requirements of healthy gilthead sea bream, Sparus aurata, which however may not be
adequate during infestations of haematophagus monogeneans. Andersen et al. (1996) found
that RBC count in Atlantic salmon was not significantly affected by the supplemental iron
levels of 10–400 mg/kg (given as iron sulphate) for 12 weeks, but a tendency for increased
values was observed. Other blood parameters such as Hb and Hct were actually affected by
iron addition in the latter study, whereas Andersen et al. (1998) reported that the hae-
matological parameters of the same fish species were not influenced by iron treatment for
20 weeks (iron sulfate; 400 mg/kg). Iron supplementation in gilthead sea bream, Sparusaurata did not significantly affect Hb concentrations (P = 0.642) but its concentration in
fish fed with the 200 ORG feed increased almost 10% in relation to the control diet.
Similarly, Hct (P = 0.105) and MHCH (P = 0.321) were not significantly affected by iron
supplementation in the treated groups and this finding was possibly related to the fact that
the actual iron concentration was up to 22 mg/kg higher than the supplementation level in
the experimental diets due to the supply of iron through the fishmeal present in all diets
including the control. The decrease in RBC count, Hb concentration and MHCH in fish fed
with 300 ORG compared to fish fed with 200 ORG, although not significant, suggested that
supplementation doses above 200 mg/kg should be avoided as they may reduce the ben-
eficial effect of iron supplementation on the fish haematological parameters.
Berger (1996) reported that either a deficiency or an excess of iron could compromise
the immune system. In the present study, no immunosuppression was observed compared
to control diet even at the highest dietary iron concentration tested (300 ORG). Immu-
nostimulation was even observed in the current study after 12 weeks feeding of gilthead
sea bream, Sparus aurata in the antibacterial activity of serum of fish fed with 300 ORG
compared to fish fed with 100 ORG or 200 INOR (P = 0.004). Also, a tendency for
increased respiratory burst activity was observed in fish fed with 100 and 200 ORG
compared to other diets (P = 0.405). Given the fact that the actual iron content of control
diet was 22 mg/kg, this treatment appeared to induce no iron-deficiency. The reduction of
respiratory burst activity at dietary iron levels of 300 ORG compared to 100 ORG,
although not significant, suggested that higher iron levels may reduce the extent of
immunostimulation. However, the antibacterial activity of serum was still high at the
1102 Aquacult Int (2010) 18:1093–1104
123

highest concentrations tested (200 and 300 ORG). In Atlantic salmon, the serum total
antibody, specific haemolytic complement activity and spontaneous haemolytic activity
and serum, head–kidney and spleen lysozyme activities were not significantly influenced
by dietary iron administration for 8 and 20 weeks (Andersen et al. 1998). In channel
catfish, iron-deficiency had no effect on the specific antibody response (Sealey et al. 1997)
but decreased macrophage chemotaxis (Lim and Klesius 1997; Sealey et al. 1997) and
accelerated the onset of mortality in channel catfish exposed to Edwardsiella ictaluri (Lim
and Klesius 1997; Lim et al. 2000). Cumulative mortality results were either not influenced
by dietary levels of iron (Lim and Klesius 1997; Lim et al. 2000) or increased in iron-
deficient fish (Sealey et al. 1997). In Atlantic salmon, a significant association between
high concentrations of serum iron and mortality of Atlantic salmon infected with Vibrioanguillarum was reported (Sherman 1992). Ravndal et al. (1994) indicated that a delicate
balance exists between the need of iron for host defence mechanisms and the need for iron
to sustain microbial growth. The results of the respiratory burst activity in the present study
suggested that a dose of 100–200 mg/kg gives satisfactory stimulation and higher doses
may reduce the extent of the immunostimulation, although the antibacterial activity of
serum was still positively influenced by 300 mg/kg.
In conclusion, the present findings provided no clear evidence respecting the optimum
iron concentration. However, there was indication that iron supplementation enhanced the
performance of gilthead sea bream, Sparus aurata, mainly from a haematological and
immunological point of view. Possibly a longer than 12 weeks experimentation with
additive dietary iron could have induced more apparent effects. The superiority of organic
vs inorganic iron sources can be only suggested as higher values of 200 ORG (in muscle,
liver and spleen iron concentrations, final fish weight and respiratory burst activity) were not
significantly different from values obtained with 200 INOR. It can be preliminary proposed
that an iron concentration between 100–200 mg/kg diet should be the recommended
additive level in feeding schedules of gilthead sea bream, Sparus aurata. Considering the
cost of the ingredient, the 100 mg/kg additive iron concentration should be considered on a
yearly basis for the dietary schedules of this species, while the 200 mg/kg concentration
should be used as a preventive measure when attacks of haematophagus monogeneans are
expected and when the water oxygen levels are low and the haematological status of the fish
might become highly compromised. Further experiments will investigate iron dietary
supplementation in association with hypoxia and/or parasitical natural challenges.
Acknowledgment This work was funded by Alltech, Department of Aquaculture Nutrition and Health.
References
Andersen F, Maage A, Julshamn K (1996) An estimation of dietary requirements of Atlantic salmon, Salmosalar L., parr. Aquacult Nut 2:41–47
Andersen F, Lorentzen M, Waagbo R, Maage A (1997) Bioavailability and interactions with othermicronutrients of three dietary iron sources in Atlantic salmon, Salmo salar, smolts. Aquacult Nut3:239–246
Andersen F, Lygren B, Maage A, Waagbo R (1998) Interaction between two dietary levels of iron and twoforms of ascorbic acid and the effect on growth, antioxidant status and some non-specific immuneparameters in Atlantic salmon (Salmo salar) smolts. Aquaculture 161:437–451
Athanassopoulou F, Ragias V, Vagianou S, Di Cave D, Rigos G, Papathanasiou G, Georgoulakis J (2005)Report of Sparicotyle (Microcotyle) chrysophrii Van Beneden and Hesse 1863, Atrispinum seminalisEuzet and Maillard 1973 and Polylabris tubicirrus Paperna and Kohn 1964 (Monogenea) on captive seabream and sharp snout sea bream in coastal Greece and Italy. Bull Eur Ass Fish Pathol 25:256–262
Aquacult Int (2010) 18:1093–1104 1103
123

Baker RTM, Martin P, Davies SJ (1997) Ingression of sub-lethal levels of iron sulphate by African catfishaffects growth and tissue lipid peroxidation. Aquat Toxicol 40:51–61
Berger LL (1996) Trace minerals: key to immunity. Salt Trace Miner 28:1–4Blaxhall PC (1972) The haematological assessment of the health of freshwater fish: a review of selected
literature. J Fish Biol 4:593–604Bury N, Grosell M (2003) Iron acquisition by teleost fish. Comp Biochem Physiol Part C 135:97–105Cooper CA, Bury NR, Grosell M (2006) The effects of pH and the iron redox state on iron uptake in the
intestine of a marine teleost fish, gulf toadfish (Opsanus beta). Comp Biochem Physiol Part A143:292–298
Drabkin DL, Austin JH (1935) Spectrophotometric studies. A technique for the analysis of undiluted bloodand concentrated hemoglobin solution. J Biol Chem 112:105–115
Fouz B, Toranzo AE, Biosca EG, Mazoy R, Amaro C (1994) Role of iron in the pathogenecity of Vibriodamsela for fish and mammals. FEMS Microbiol Lett 121:181–188
Gatlin DM, Wilson RP (1986) Characterisation of iron deficiency and the dietary iron requirement offingerling channel catfish. Aquaculture 52:191–198
Ikeda Y, Ozaki H, Uematsu K (1973) Effect of enriched diet with iron in culture of yellowtail. J TokyoUniver Fish 59:91–99
Kawatsu H (1972) Studies on the anaemia of fish. 5. Dietary iron deficient anaemia in brook trout, Salvelinusfontinalis. Bull Freshwat Fish Res Lab Tokyo 22:59–67
Lall SP (2002) The Minerals. In: Halver JH, Hardy RW (eds) Fish Nutrition, 3rd edn. Academic Press, CALee YH, Layman DK, Bell RB, Norton HW (1981) Response of glutathione peroxidase and catalase to
excess dietary iron in rats. J Nutr 111:2195–2202Lim C, Klesius PH (1997) Responses of channel catfish Ictalurus punctatus fed iron-deficient and replete
diets to Edwardsiella ictaluri challenge. Aquaculture 157:83–93Lim C, Klesius PH, Li MH, Robinson EH (2000) Interaction between dietary levels of iron and vitamin C on
growth, hematology, immune response and resistance of channel catfish Ictalurus punctatus toEdwardsiella ictaluri challenge. Aquaculture 185:313–327
Nikoskelainen S, Lehtinen J, Lilius EM (2002) Bacteriolytic activity of rainbow trout (Oncorhynchusmykiss) complement. Dev Comp Immunol 26:797–804
Nikoskelainen S, Verho S, Airas K, Lilius EM (2005) Adhesion and ingestion activities of fish phagocytesinduced by bacterium (Aeromonas salmonicida) can be distinguished and directly measured fromhighly diluted whole blood of fish. Dev Comp Immunol 29:525–537
Nose T, Arai S (1979) Recent advances in studies on mineral nutrition of fish in Japan. In: Pillay TVR, DillWA (eds) Advances in aquaculture. Fishing News (Books) Ltd, Farnham
Rasmussen KJ (1994) Spray-dried blood in diets to Atlantic salmon (Salmo salar). Fisk Dir Skr SerErncering 6:151–161
Ravndal J, Lovold T, Bentsen HB, Roed KH, Gjedrem T, Rorvik KA (1994) Serum iron levels in farmedAtlantic salmon: family variation and associations with disease resistance. Aquaculture 125:37–45
Sakamoto S, Yone Y (1976) Requirement of red sea bream for dietary iron. Rep Fish Res Lab Kyushu Univ3:53–58
Salte R, Rørvik KA, Reed E, Nordberg K (1994) Winter ulcers of the skin in Atlantic salmon, Salmo salarL.: pathogenesis and possible aetiology. J Fish Dis 17:661–665
Sealey WM, Lim C, Klesius PH (1997) Influence of dietary level of iron from iron methionine and ironsulfate on immune response and resistance of channel catfish to Edwardsiella ictaluri. J WorldAquacult Soc 28:142–149
Sherman AR (1992) Zinc, copper and iron nutriture and immunity. J Nutr 122:604–609Sutton J, Balfry S, Higgs D, Huang C-H, Skura B (2006) Impact of iron-catalyzed dietary lipid peroxidation
on growth performance, general health and flesh proximate and fatty acid composition of Atlanticsalmon (Salmo salar L.) reared in seawater. Aquaculture 257:534–557
Tacon AJ (1992) Nutritional fish pathology. Morphological signs of nutrient deficiency and toxicity infarmed fish. FAO Fish Technical Paper. No. 330. Rome, FAO, p 75
Vangen B, Hemre GI (2003) Dietary carbohydrate, iron and zinc interactions in Atlantic salmon (Salmosalar). Aquaculture 219:597–611
Watanabe T, Kiron V, Satoh S (1997) Trace minerals in fish nutrition. Aquaculture 151:185–207Wilson RW (1999) A novel role for the gut of seawater teleosts in acid–base balance. In: Taylor EW, Raven
JA, Egginton S (eds) Regulation of tissue pH in plants and animals. Cambridge University Press,Cambridge
1104 Aquacult Int (2010) 18:1093–1104
123




















