
Effects of extrusion and inclusion level of soybean meal on diet digestibility, performance and...
15
This article was originally published in a journal published by Elsevier, and the attached copy is provided by Elsevier for the author’s benefit and for the benefit of the author’s institution, for non-commercial research and educational use including without limitation use in instruction at your institution, sending it to specific colleagues that you know, and providing a copy to your institution’s administrator. All other uses, reproduction and distribution, including without limitation commercial reprints, selling or licensing copies or access, or posting on open internet sites, your personal or institution’s website or repository, are prohibited. For exceptions, permission may be sought for such use through Elsevier’s permissions site at: http://www.elsevier.com/locate/permissionusematerial
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Effects of extrusion and inclusion level of soybean meal on diet digestibility, performance and...

This article was originally published in a journal published byElsevier, and the attached copy is provided by Elsevier for the
author’s benefit and for the benefit of the author’s institution, fornon-commercial research and educational use including without
limitation use in instruction at your institution, sending it to specificcolleagues that you know, and providing a copy to your institution’s
administrator.
All other uses, reproduction and distribution, including withoutlimitation commercial reprints, selling or licensing copies or access,
or posting on open internet sites, your personal or institution’swebsite or repository, are prohibited. For exceptions, permission
may be sought for such use through Elsevier’s permissions site at:
http://www.elsevier.com/locate/permissionusematerial

Autho
r's
pers
onal
co
py
Effects of extrusion and inclusion level of soybean meal on dietdigestibility, performance and nutrient utilization
of gilthead sea bream (Sparus aurata)
B. Venou, M.N. Alexis ⁎, E. Fountoulaki, J. Haralabous
Hellenic Center for Marine Research, Ag. Kosmas, Helliniko, Athens 16604, Greece
Received 27 March 2006; received in revised form 20 July 2006; accepted 20 July 2006
Abstract
The objective of this study was to evaluate the effects of soybean meal inclusion and treatment on the performance of two sizesof gilthead sea bream (small, 9 g and large 50 g mean initial weight). Seven diets were fed containing soybean meal, before (SBM)and after (SBMex) its extrusion, at increasing levels (0% as control and about 20, 30, and 40% for both soy forms used). For largefish effects on digestibility, as well as on trypsin activity levels and moisture at upper (UI), mid (MI) and lower (LI) intestine werealso evaluated, following the growth trial. All experiments were conducted at 22±2 °C. Specific growth rate and whole bodycomposition were not affected by soy inclusion, however an adverse effect of inclusion level on feed conversion ratio and proteinproductive value was apparent for both soy products and fish sizes. Extrusion of soy affected differently large and small fishimproving both protein productive value and feed conversion ratio of large fish, but only protein productive value of small fish.Plasma cholesterol decreased with inclusion level of both types of soy, the effect being significant only at the higher level. Liverlipid and glycogen deposition in large fish appeared to be mostly affected by digestible starch intake, increasing with increasedintakes, the same being apparent for hepatosomatic index. Soy inclusion did not appear to have any effect on nutrient digestibilities.Trypsin activity (TA) was decreased from UI to LI for all diets and was significantly higher in C diet compared to all other diets atall segments. Extrusion of SBM increased significantly TA levels. Hydration of digesta in LI increased for low soy levels anddecreased for high soy levels compared to the control diet.© 2006 Elsevier B.V. All rights reserved.
Keywords: Gilthead sea bream; Soybean meal; Extrusion; Digestibility; Trypsin activity
1. Introduction
SBM is one of the most interesting alternatives tofishmeal for fish diets due to high protein content, sat-isfactory amino acid profile, reasonable price and steadysupply. Soybean meal contains however a number of
antinutrients, which limit its utilization by fish (Franciset al., 2001). Certain of those exert an influence on theprocess of digestion reducing nutrient digestibility. Ne-gative effects on protein digestibility have been attribut-ed to trypsin inhibitors and phytic acid, the first bindingto digestive enzymes and the second making complexeswith protein molecules (Spinelli et al., 1983; Franciset al., 2001). Negative effects on lipid digestibility werealso found (Refstie et al., 1998, 2000; Storebakken et al.,1998b; Tomás et al., 2005) and attributed either to the
Aquaculture 261 (2006) 343–356www.elsevier.com/locate/aqua-online
⁎ Corresponding author. Tel.: +30 210 9856722; fax: +30 2109829239.
E-mail address: [email protected] (M.N. Alexis).
0044-8486/$ - see front matter © 2006 Elsevier B.V. All rights reserved.doi:10.1016/j.aquaculture.2006.07.030
Autho
r's
pers
onal
co
py
alcohol extractable fraction (Arnesen et al., 1989; Olliand Kroghahl, 1995), or to the non-starch polysacchar-ides (NSP) content of SBM (Refstie et al., 1999;Storebakken et al., 2000). The alcohol extractable frac-tion is also involved in alterations in the distal part ofintestine of salmonids (Refstie et al., 2000, 2001;Storebakken et al., 2000; Krogdahl et al., 2003) andphytic acid limits phosphate and mineral absorption(Robaina et al., 1998; Storebakken et al., 1998a; Sujiuraet al., 2001). Hypocholesterolemia has been reported inmany studies with soybean meal inclusion (Kaushiket al., 1995, 2004; Refstie et al., 1999; Regost et al.,1999; Tulli et al., 1999) and attributed to a number ofconstituents like, protein, saponins, NSP or isoflavonesinvolved in a multiplicity of mechanisms of cholesterolcontrol (Ikegami et al., 1990; Sugano et al., 1990; Potter,1995; Refstie et al., 1999; Rubio et al., 2004).
A number of studies indicated palatability problemsfor the feeds when SBM was included in the diets(Fowler, 1980; Hajen et al., 1993) this effect being moreimportant for the first period of fish feeding (Refstieet al., 1997, 1998). No effect (Kaushik et al., 1995;Boonyaratpalin et al., 1998; Khan et al., 2003), or evenan increase in consumption was observed (Tomás et al.,2005; Ogunkoya et al., 2006) however in other studies.While differences in dietary formulations could be oneof the reasons contributing to differences in feedingresponse of fish to soybean meal, species differences isanother possible factor.
There generally appears to be some variability be-tween species in the maximum dietary levels of SBMtolerated (Kaushik et al., 1995; Robaina et al., 1995;Nengas et al., 1996; McGoogan and Gatlin, 1997;Refstie et al., 2000) indicating different sensitivities tosoybean meal inclusion. Furthermore stronger negativeeffects have been reported for fish of smaller size, butthe relevant studies are very few (Murai et al., 1989;Choi et al., 2004).
Gilthead sea bream is one of the most importantcultured species in Mediterranean aquaculture. Fishmealreplacement by SBM was the subject of a number ofstudies. In a preliminary experiment with gilthead seabream fingerlings, a significant effect on growth wasobserved at dietary inclusions of SBM of 30% or higher(Nengas et al., 1996). In another study, inclusion of up to30% SBM did not significantly affect growth, feed orprotein efficiency (Robaina et al., 1995). The differentsizes of fish used and quality of FM replaced could havecontributed to the different responses observed in theseexperiments. Both studies involved common SBM notsubjected to further heat treatment since laboratorypelleted diets were used. Heat treatment destroys a large
number of antinutritional factors contained in plant de-rived ingredients and improves their utilization by fish(Tacon, 1997; Refstie et al., 1998; Arndt et al., 1999;Francis et al., 2001). Extrusion involves heat treatment,pressure and shear forces and it has been found to havebeneficial effects in nutritional value of many plantingredients (Pfeffer et al., 1991; Burel et al., 2000a,b;
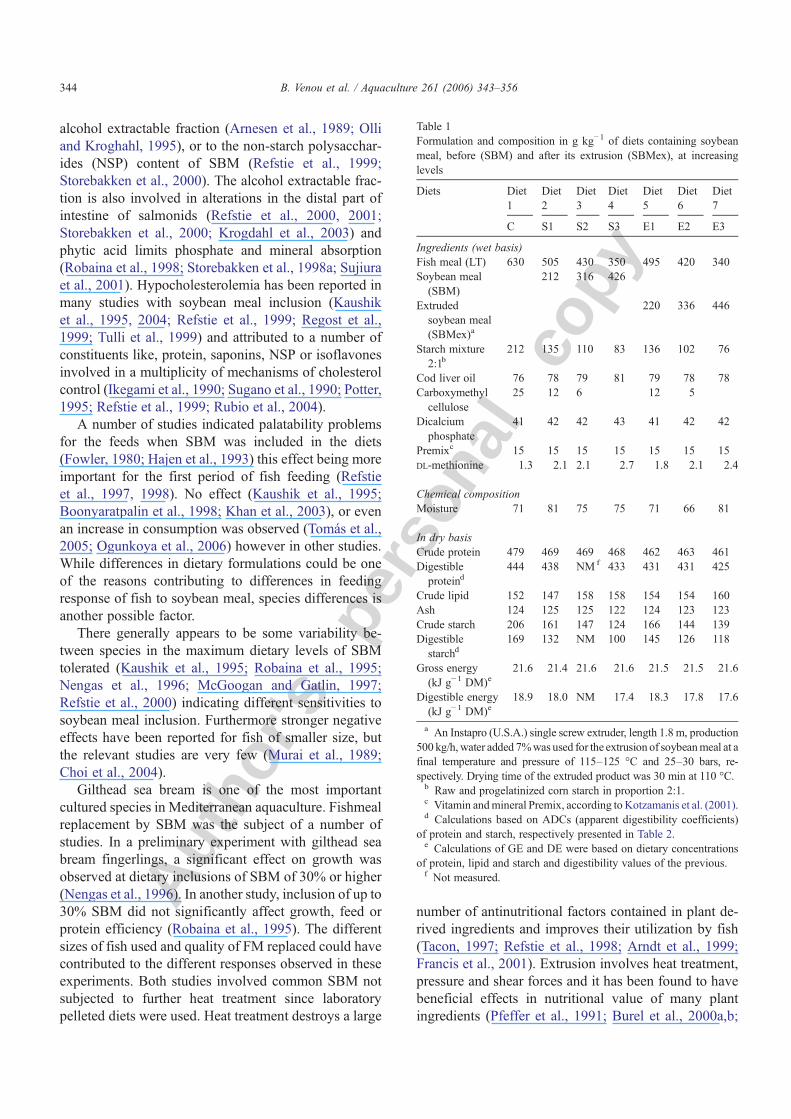
Table 1Formulation and composition in g kg−1 of diets containing soybeanmeal, before (SBM) and after its extrusion (SBMex), at increasinglevels
Diets Diet1
Diet2
Diet3
Diet4
Diet5
Diet6
Diet7
C S1 S2 S3 E1 E2 E3
Ingredients (wet basis)Fish meal (LT) 630 505 430 350 495 420 340Soybean meal(SBM)
212 316 426
Extrudedsoybean meal(SBMex)a
220 336 446
Starch mixture2:1b
212 135 110 83 136 102 76
Cod liver oil 76 78 79 81 79 78 78Carboxymethylcellulose
25 12 6 12 5
Dicalciumphosphate
41 42 42 43 41 42 42
Premixc 15 15 15 15 15 15 15DL-methionine 1.3 2.1 2.1 2.7 1.8 2.1 2.4
Chemical compositionMoisture 71 81 75 75 71 66 81
In dry basisCrude protein 479 469 469 468 462 463 461Digestibleproteind
444 438 NMf 433 431 431 425
Crude lipid 152 147 158 158 154 154 160Ash 124 125 125 122 124 123 123Crude starch 206 161 147 124 166 144 139Digestiblestarchd
169 132 NM 100 145 126 118
Gross energy(kJ g−1 DM)e
21.6 21.4 21.6 21.6 21.5 21.5 21.6
Digestible energy(kJ g−1 DM)e
18.9 18.0 NM 17.4 18.3 17.8 17.6
a An Instapro (U.S.A.) single screw extruder, length 1.8 m, production500 kg/h, water added 7%was used for the extrusion of soybeanmeal at afinal temperature and pressure of 115–125 °C and 25–30 bars, re-spectively. Drying time of the extruded product was 30 min at 110 °C.b Raw and progelatinized corn starch in proportion 2:1.c Vitamin andmineral Premix, according to Kotzamanis et al. (2001).d Calculations based on ADCs (apparent digestibility coefficients)
of protein and starch, respectively presented in Table 2.e Calculations of GE and DE were based on dietary concentrations
of protein, lipid and starch and digestibility values of the previous.f Not measured.
344 B. Venou et al. / Aquaculture 261 (2006) 343–356

Autho
r's
pers
onal
co
py
Allan and Booth, 2004) as well as for soybean meal(Pongmaneerat and Watanabe, 1992, 1993). The effectof extrusion on improving dietary quality of soybeanmeal for gilthead sea bream has not been studied.
The objectives of this study were, therefore, to eval-uate the effects of both dietary level and extrusion ofsoybean meal on diet acceptance, growth, nutrient uti-lization and body and liver composition of gilthead seabream. The effect of size, in differentiating the responsesto soybean inclusion, was also studied by using fish of9 g and 50 g in weight. Finally changes in digestibilityand digesta characteristics for the larger size group weremeasured to give further insight to the effects of SBM ondigestive physiology of gilthead sea bream and the ef-fects of extrusion. A fishmeal of highest quality was usedas basic protein supplier, so to avoid uncertainties relatedto the quality of fishmeal replaced.
2. Materials and methods
2.1. Experimental diets-fish
Defatted soybean meals with hulls, before (SBM) andafter extrusion (SBMex), as well as low temperature fishmeal (FM LT, Denmark) were used as the main ingre-dients of the diets.
Seven diets (Table 1) were formulated to be ap-proximately isonitrogenous (47% crude protein) andisocaloric (21 kJ/g DM). FM LT was the sole proteinsource in the control diet (C) and both forms of soybeanmeal were incorporated in levels ranging from about 20to 45% in the other diets as substitutes for FM (diets Sfor SBM and E for SBMex). The dietary lipid and starchsources were fish oil and a mixture of raw and pro-gelatinized cornstarch (2/1) respectively. DL-methioninewas also supplied in diets to increase the Met+Cysconcentration to 4.3 g/16 g N.
In preparing the test diets all the dietary ingredientswereblended thoroughly, moistened to produce a dense pasteand passed through amincer (Hobart foodmixer, model noA 120, Hobart manufacturing company LTD). Diets wereair-dried (b40 °C) to amoisture content of about 7–8%andstored frozen (−18 °C) until used. For the digestibility trial1% chromic oxide was incorporated in the diets.
Growth performance was evaluated in two popula-tions of gilthead sea bream of different initial size, of 9 g(small fish) and 50 g (large fish) mean initial weight inexperiments I and II, respectively. Fish at the end of thefeeding trial in experiment II were also used formeasuring both digestibility (experiment III) and trypsinactivity (experiment IV). Gilthead sea bream of bothsizes were obtained from a commercial hatchery and
acclimated to the laboratory conditions for two weeks,before distributing to the experimental tanks.
2.2. Experiments I and II — growth performance andnutrient utilization
Duplicate groups of 30 and 20 fish of “small” and“large” gilthead sea bream (Sparus aurata) were assignedto 14–100 l cylindrical and 12–120 l cylindro-conicalfiberglass tanks, respectively at the fish nutrition aquar-ium of the Hellenic Center for Marine Research. Thephotoperiod was operated on a 10:14 h L/D cycle and thetemperature of the open flow seawater was 22±2 °C. Theflow ratewas adjusted to replace tankwater once per hour.The water quality was checked periodically. PH rangedfrom 7.8 to 8.0 and total ammonia levels did not exceed0.2–0.3 mg l−1. Fish were weighed at the beginning, atthe end of the test period and every two weeks throughoutthe rearing period, after 24 h of fasting. Diets were fed byhand to apparent satiation, three times per day, for 60 and66 days for the small and the large fish, respectively. DietS2 was not tested in experiment II due to the limitednumber of tanks available. Feed consumption was reg-istered for each population of gilthead sea bream.
A sample of 10 fish was removed from each initialpopulation and sacrificed by lethal anesthesia for bodycomposition analyses after 24 h fasting, before distrib-uting the fish to the tanks. In the same way 16 fish pertank were removed at the end of the feeding period inexperiment I. Ten of them were pooled for body com-position analysis and 6 of them used for hepatosomaticindex and liver composition analysis. Six fish wereremoved from each tank in experiment II and bloodtaken with a heparinized syringe from the caudal vein.Plasma was collected after centrifugation (3000 rpm,20 min) and stored frozen (−70 °C) for cholesterolanalysis. Livers were weighed for hepatosomatic index(HSI) determination. A small portion of liver was re-moved and frozen (−70 °C) for lipid and glycogencontent measurements and the rest was homogenizedwith fish bodies for body composition analysis.
The parameters calculated were: specific growth rateas SGR=100×[(lnWF)− (lnWI)]×D
−1, where WI andWF are the initial and final weights (tank means), re-spectively and D are the rearing days; daily percent feedor nutrient intake as: 100×FD×WM
−1, where FD is theaverage dry feed (or nutrient) consumption per fish perday andWM is mean weight per fish per tank obtained asan average of the initial and final weight; feed conversionratio as: FCR=FT×WG
−1, where WG is the weight gainper fish and FT is the total dry feed consumption per fishover the whole rearing period; protein efficiency ratio as
345B. Venou et al. / Aquaculture 261 (2006) 343–356

Autho
r's
pers
onal
co
py
PER=WG×PT−1, where PT is the total protein consump-
tion per fish. Protein productive value as PPV=(PF
−PI)×PT−1, where PI and PF are the protein content per
fish (in g) at the beginning and at the end of theexperiment and PT as before. Hepatosomatic index as:HSI=100×(liver weight)× (body weight)−1.
2.3. Experiment III — digestibility
After sampling in experiment II, the rest of the fishwere kept in the same tanks, which were equipped withfecal traps (Nengas et al., 1995) and fed their respectivediets containing Cr2O3, three times a day, totaling 1.8%of their body weight. Then the system was cleaned fromfeeds and feces and feces were collected up to nextmorning for a period of 12 days and pooled for 3 daysresulting in 4 samples per tank. A 4-day acclimatizationperiod to the diets preceded the collection period. Watertemperature was 22±2 °C. Apparent digestibilitycoefficients (ADC) of nutrients were calculated as:ADC (%)=100×[1−DCr2O3×FCr2O3
−1 ×FN×DN−1],where DCr2O3 and FCr2O3 were the concentrations ofindicator in diet and feces and FN and DN were thenutrient concentrations in feces or diet, respectively.
2.4. Experiment IV — trypsin activity in three intestinalsegments
Following the digestibility trial fish fed the diets, C,S1, S3, E1, E3 were left unfed for 24 h and then a singlemeal (1.5% of the wet body weight) was offered in themorning for a period of about 20–30 min. Three fish pertank were sacrificed 4 h after feeding by lethal anes-
thesia. The abdominal cavity was then opened and thedigestive tract ligated at the pylorus and anus and care-fully freed from connective and adipose tissue, afterstomach removal. The intestine was also ligated in twospecific points, being so separated in three segments:upper intestine (UI), the section between the stomach'spyloric sphincter and the first bend of the digestivetrack, mid-intestine (MI), the section between the firstbend of the digestive track and the ring-shaped nar-rowing at the lower region of the intestine (Cataldi et al.,1987) and lower intestine (LI), the section after thisnarrowing and the anus.
The intestinal segments were then opened and di-gestive contents (DCs) of each fish collected separatelyin test tubes and weighted. DCs were centrifuged for12 min at 6500 ×g (4 °C), the supernatant removed andits volume measured using a graduated pipette of ap-propriate volume. The supernatant was then kept frozen(−70 °C) and used for trypsin determinations. Hydrationof digesta was expressed as the volume of supernatantmeasured (in ml) per 100 g of total digesta in eachintestinal segment. This expression was expected tocorrespond better to hydration values, than a weight perweight expression, since supernatant was not pure waterand contained hydrolysis products and excreta.
Trypsin activity (TA) was assayed at 25.9 °C using1.09 M DL-BApNA (Sigma B 4875) as a substrate, in0.05 M Tris buffer, pH 8.2 containing 10 mM CaCl2(Erlanger et al., 1961). The production of p-nitroaniline(pNA)wasmeasured at 410 nmusing a (BeckmanDU-64)spectrophotometer. One unit of trypsin activity was de-fined as 1 μmol of pNA released per min (U ml−1), using8800 as extinction coefficient. Activity of trypsin in
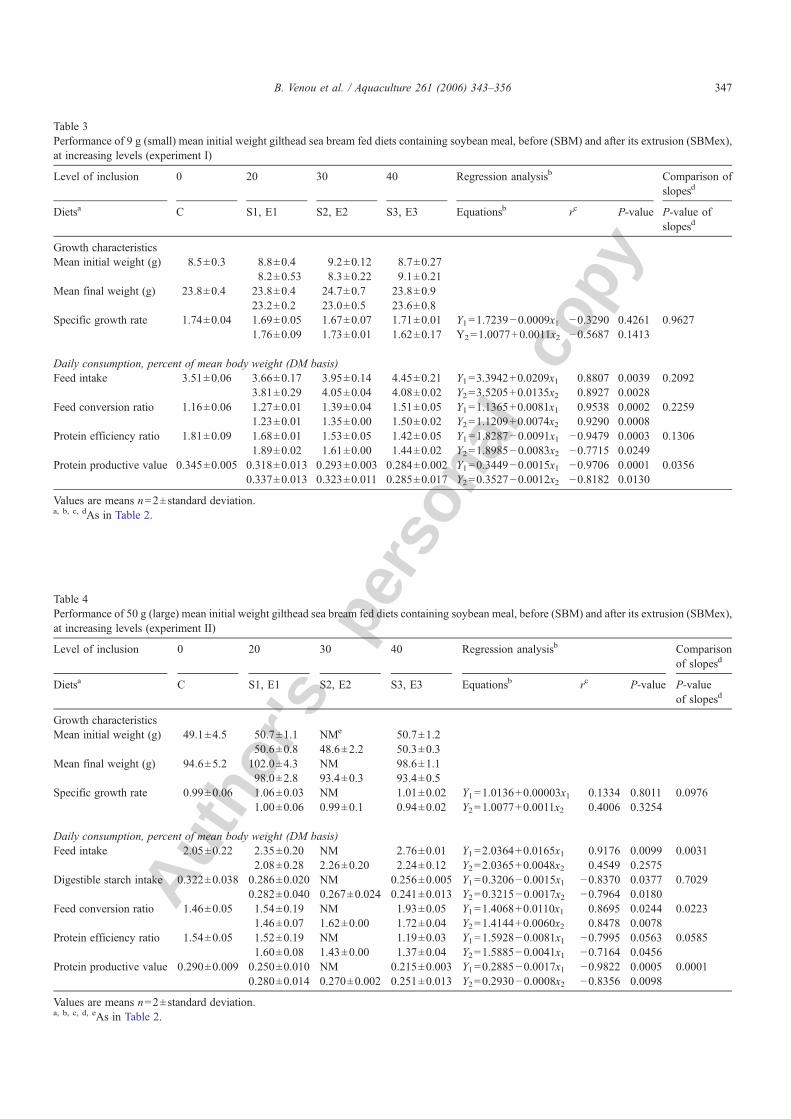
Table 2Apparent digestibility coefficients (%) of diets containing soybean meal, before (SBM) and after its extrusion (SBMex), at increasing levels, tested forgilthead sea bream of 50 g (large) mean initial weight (experiment III)
Level of inclusion 0 20 30 40 Regression analysisb Comparison of slopesd
Dietsa C S1, E1 S2, E2 S3, E3 Equationsb rc P-value P-value of slopesd
ADCs (%)Protein 92.6±0.5 93.3±0.4 NMe 92.4±0.3 Y1=92.8957−0.0064x1 −0.2313 0.6593 0.9701
93.2±0.9 93.0±0.2 92.1±0.3 Y2=92.9383−0.0080x2 −0.2305 0.5828Lipid 90.8±1.3 92.3±1.8 NM 88.0±2.1 Y1=91.7476−0.0661x1 −0.5369 0.2721 0.9873
92.1±2.8 89.1±1.9 88.2±2.2 Y2=91.6185−0.0619x2 −0.4894 0.2184Starch 82.1±0.7 82.2±1.2 NM 80.6±1.1 Y1=82.3366−0.0344x1 −0.5833 0.2243 0.0018
87.7±0.5 87.6±0.2 84.7±0.1 Y2=83.6366+0.0737x 0.5238 0.1827
Values are means n=2±standard deviation.a First line: results for SBM diets. Second line: results for SBMex diets.b Changes in parameter values in respect to soybean meal inclusion. Y1 and Y2 parameter values for SBM and SBMex, respectively.c Correlation coefficients.d Comparison of slopes of the regression lines.e Not measured.
346 B. Venou et al. / Aquaculture 261 (2006) 343–356
Autho
r's
pers
onal
co
py
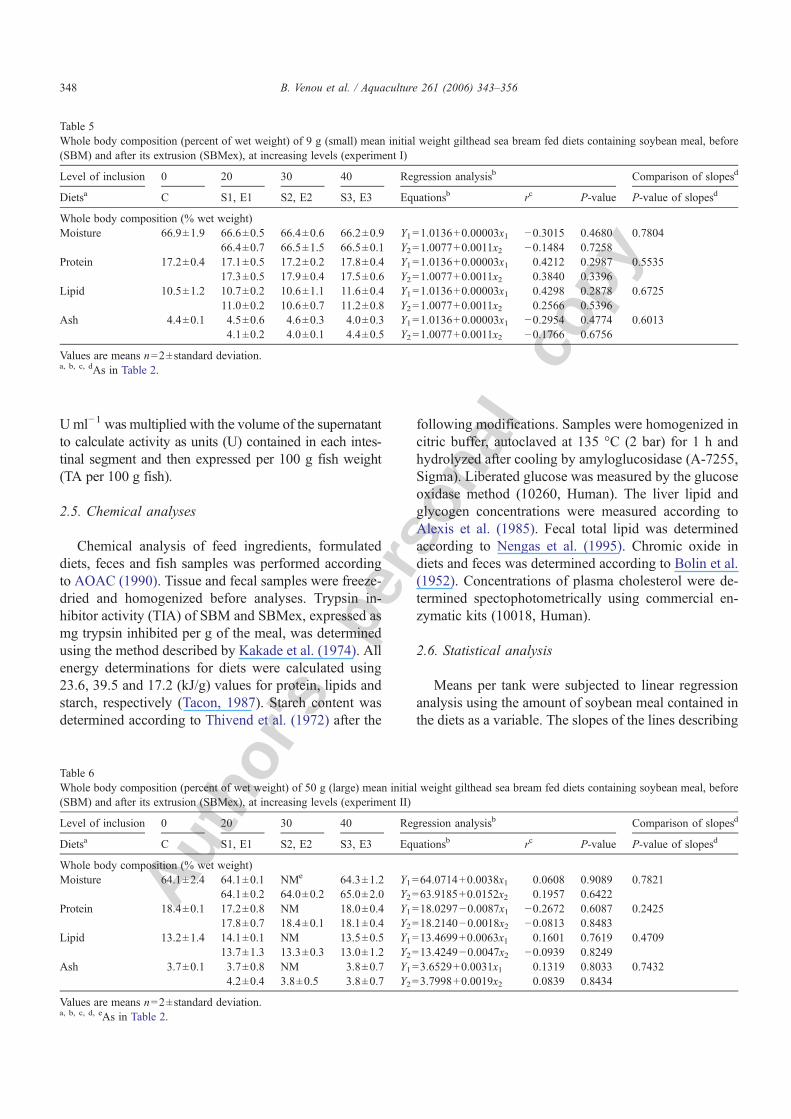
Table 3Performance of 9 g (small) mean initial weight gilthead sea bream fed diets containing soybean meal, before (SBM) and after its extrusion (SBMex),at increasing levels (experiment I)
Level of inclusion 0 20 30 40 Regression analysisb Comparison ofslopesd
Dietsa C S1, E1 S2, E2 S3, E3 Equationsb rc P-value P-value ofslopesd
Growth characteristicsMean initial weight (g) 8.5±0.3 8.8±0.4 9.2±0.12 8.7±0.27
8.2±0.53 8.3±0.22 9.1±0.21Mean final weight (g) 23.8±0.4 23.8±0.4 24.7±0.7 23.8±0.9
23.2±0.2 23.0±0.5 23.6±0.8Specific growth rate 1.74±0.04 1.69±0.05 1.67±0.07 1.71±0.01 Y1=1.7239−0.0009x1 −0.3290 0.4261 0.9627
1.76±0.09 1.73±0.01 1.62±0.17 Y2=1.0077+0.0011x2 −0.5687 0.1413
Daily consumption, percent of mean body weight (DM basis)Feed intake 3.51±0.06 3.66±0.17 3.95±0.14 4.45±0.21 Y1=3.3942+0.0209x1 0.8807 0.0039 0.2092
3.81±0.29 4.05±0.04 4.08±0.02 Y2=3.5205+0.0135x2 0.8927 0.0028Feed conversion ratio 1.16±0.06 1.27±0.01 1.39±0.04 1.51±0.05 Y1=1.1365+0.0081x1 0.9538 0.0002 0.2259
1.23±0.01 1.35±0.00 1.50±0.02 Y2=1.1209+0.0074x2 0.9290 0.0008Protein efficiency ratio 1.81±0.09 1.68±0.01 1.53±0.05 1.42±0.05 Y1=1.8287−0.0091x1 −0.9479 0.0003 0.1306
1.89±0.02 1.61±0.00 1.44±0.02 Y2=1.8985−0.0083x2 −0.7715 0.0249Protein productive value 0.345±0.005 0.318±0.013 0.293±0.003 0.284±0.002 Y1=0.3449−0.0015x1 −0.9706 0.0001 0.0356
0.337±0.013 0.323±0.011 0.285±0.017 Y2=0.3527−0.0012x2 −0.8182 0.0130
Values are means n=2±standard deviation.a, b, c, dAs in Table 2.
Table 4Performance of 50 g (large) mean initial weight gilthead sea bream fed diets containing soybean meal, before (SBM) and after its extrusion (SBMex),at increasing levels (experiment II)
Level of inclusion 0 20 30 40 Regression analysisb Comparisonof slopesd
Dietsa C S1, E1 S2, E2 S3, E3 Equationsb rc P-value P-valueof slopesd
Growth characteristicsMean initial weight (g) 49.1±4.5 50.7±1.1 NMe 50.7±1.2
50.6±0.8 48.6±2.2 50.3±0.3Mean final weight (g) 94.6±5.2 102.0±4.3 NM 98.6±1.1
98.0±2.8 93.4±0.3 93.4±0.5Specific growth rate 0.99±0.06 1.06±0.03 NM 1.01±0.02 Y1=1.0136+0.00003x1 0.1334 0.8011 0.0976
1.00±0.06 0.99±0.1 0.94±0.02 Y2=1.0077+0.0011x2 0.4006 0.3254
Daily consumption, percent of mean body weight (DM basis)Feed intake 2.05±0.22 2.35±0.20 NM 2.76±0.01 Y1=2.0364+0.0165x1 0.9176 0.0099 0.0031
2.08±0.28 2.26±0.20 2.24±0.12 Y2=2.0365+0.0048x2 0.4549 0.2575Digestible starch intake 0.322±0.038 0.286±0.020 NM 0.256±0.005 Y1=0.3206−0.0015x1 −0.8370 0.0377 0.7029
0.282±0.040 0.267±0.024 0.241±0.013 Y2=0.3215−0.0017x2 −0.7964 0.0180Feed conversion ratio 1.46±0.05 1.54±0.19 NM 1.93±0.05 Y1=1.4068+0.0110x1 0.8695 0.0244 0.0223
1.46±0.07 1.62±0.00 1.72±0.04 Y2=1.4144+0.0060x2 0.8478 0.0078Protein efficiency ratio 1.54±0.05 1.52±0.19 NM 1.19±0.03 Y1=1.5928−0.0081x1 −0.7995 0.0563 0.0585
1.60±0.08 1.43±0.00 1.37±0.04 Y2=1.5885−0.0041x1 −0.7164 0.0456Protein productive value 0.290±0.009 0.250±0.010 NM 0.215±0.003 Y1=0.2885−0.0017x1 −0.9822 0.0005 0.0001
0.280±0.014 0.270±0.002 0.251±0.013 Y2=0.2930−0.0008x2 −0.8356 0.0098
Values are means n=2±standard deviation.a, b, c, d, eAs in Table 2.
347B. Venou et al. / Aquaculture 261 (2006) 343–356
Autho
r's
pers
onal
co
py
Uml−1 was multiplied with the volume of the supernatantto calculate activity as units (U) contained in each intes-tinal segment and then expressed per 100 g fish weight(TA per 100 g fish).
2.5. Chemical analyses
Chemical analysis of feed ingredients, formulateddiets, feces and fish samples was performed accordingto AOAC (1990). Tissue and fecal samples were freeze-dried and homogenized before analyses. Trypsin in-hibitor activity (TIA) of SBM and SBMex, expressed asmg trypsin inhibited per g of the meal, was determinedusing the method described by Kakade et al. (1974). Allenergy determinations for diets were calculated using23.6, 39.5 and 17.2 (kJ/g) values for protein, lipids andstarch, respectively (Tacon, 1987). Starch content wasdetermined according to Thivend et al. (1972) after the
following modifications. Samples were homogenized incitric buffer, autoclaved at 135 °C (2 bar) for 1 h andhydrolyzed after cooling by amyloglucosidase (A-7255,Sigma). Liberated glucose was measured by the glucoseoxidase method (10260, Human). The liver lipid andglycogen concentrations were measured according toAlexis et al. (1985). Fecal total lipid was determinedaccording to Nengas et al. (1995). Chromic oxide indiets and feces was determined according to Bolin et al.(1952). Concentrations of plasma cholesterol were de-termined spectophotometrically using commercial en-zymatic kits (10018, Human).
2.6. Statistical analysis
Means per tank were subjected to linear regressionanalysis using the amount of soybean meal contained inthe diets as a variable. The slopes of the lines describing
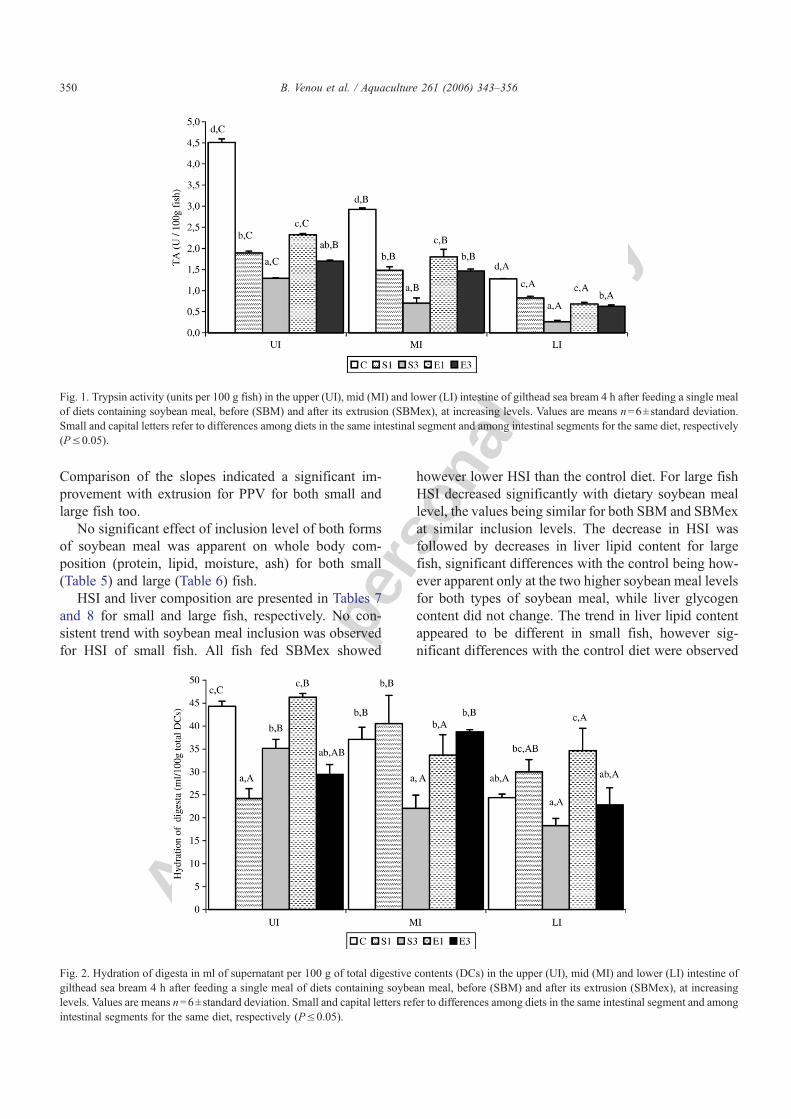
Table 5Whole body composition (percent of wet weight) of 9 g (small) mean initial weight gilthead sea bream fed diets containing soybean meal, before(SBM) and after its extrusion (SBMex), at increasing levels (experiment I)
Level of inclusion 0 20 30 40 Regression analysisb Comparison of slopesd
Dietsa C S1, E1 S2, E2 S3, E3 Equationsb rc P-value P-value of slopesd
Whole body composition (% wet weight)Moisture 66.9±1.9 66.6±0.5 66.4±0.6 66.2±0.9 Y1=1.0136+0.00003x1 −0.3015 0.4680 0.7804
66.4±0.7 66.5±1.5 66.5±0.1 Y2=1.0077+0.0011x2 −0.1484 0.7258Protein 17.2±0.4 17.1±0.5 17.2±0.2 17.8±0.4 Y1=1.0136+0.00003x1 0.4212 0.2987 0.5535
17.3±0.5 17.9±0.4 17.5±0.6 Y2=1.0077+0.0011x2 0.3840 0.3396Lipid 10.5±1.2 10.7±0.2 10.6±1.1 11.6±0.4 Y1=1.0136+0.00003x1 0.4298 0.2878 0.6725
11.0±0.2 10.6±0.7 11.2±0.8 Y2=1.0077+0.0011x2 0.2566 0.5396Ash 4.4±0.1 4.5±0.6 4.6±0.3 4.0±0.3 Y1=1.0136+0.00003x1 −0.2954 0.4774 0.6013
4.1±0.2 4.0±0.1 4.4±0.5 Y2=1.0077+0.0011x2 −0.1766 0.6756
Values are means n=2±standard deviation.a, b, c, dAs in Table 2.
Table 6Whole body composition (percent of wet weight) of 50 g (large) mean initial weight gilthead sea bream fed diets containing soybean meal, before(SBM) and after its extrusion (SBMex), at increasing levels (experiment II)
Level of inclusion 0 20 30 40 Regression analysisb Comparison of slopesd
Dietsa C S1, E1 S2, E2 S3, E3 Equationsb rc P-value P-value of slopesd
Whole body composition (% wet weight)Moisture 64.1±2.4 64.1±0.1 NMe 64.3±1.2 Y1=64.0714+0.0038x1 0.0608 0.9089 0.7821
64.1±0.2 64.0±0.2 65.0±2.0 Y2=63.9185+0.0152x2 0.1957 0.6422Protein 18.4±0.1 17.2±0.8 NM 18.0±0.4 Y1=18.0297−0.0087x1 −0.2672 0.6087 0.2425
17.8±0.7 18.4±0.1 18.1±0.4 Y2=18.2140−0.0018x2 −0.0813 0.8483Lipid 13.2±1.4 14.1±0.1 NM 13.5±0.5 Y1=13.4699+0.0063x1 0.1601 0.7619 0.4709
13.7±1.3 13.3±0.3 13.0±1.2 Y2=13.4249−0.0047x2 −0.0939 0.8249Ash 3.7±0.1 3.7±0.8 NM 3.8±0.7 Y1=3.6529+0.0031x1 0.1319 0.8033 0.7432
4.2±0.4 3.8±0.5 3.8±0.7 Y2=3.7998+0.0019x2 0.0839 0.8434
Values are means n=2±standard deviation.a, b, c, d, eAs in Table 2.
348 B. Venou et al. / Aquaculture 261 (2006) 343–356

Autho
r's
pers
onal
co
py
the relation between the dependent variable and eitherSBM or SBMex content were also compared. Regres-sions were performed using the statistical programStatgraphics Plus for Windows 2.1. The linear fits andthe comparison of slopes were tested at a significancelevel P≤0.05.
In the case where measurements were made on indi-vidual fish, results were subjected to nested analysis ofvariance using the general linear model. If significant dif-ferences were detected, means were compared usingDuncan's multiple range test (P≤0.05). The statisticalprogram SPSS for Windows (release 11.0) was used. Theresults are presented as mean±S.D. (standard deviation ofthe mean).
3. Results
3.1. Digestibility
No effect of either SBM or SBMex inclusion level wasapparent on ADCs of diets (Table 2). This apparentlyresulted from the fact that digestibilities measured for thefirst level of soybean meal were similar or better thanthose of the control diet. When a two way ANOVAwasused for comparing the effect of inclusion and treatment,using only soybean meal containing diets, then asignificant effect was observed only for starch digestibil-ity, which was much stronger with extrusion than with
soybean meal level (76% of the variation explained byprocessing and 17.4% by inclusion).
3.2. Growth experiments
No mortality was observed and gilthead sea bream ofboth sizes accepted well all experimental diets in bothexperiments.
Performance parameters as well as nutrient utili-zation values are indicated in Table 3 for small andTable 4 for large fish respectively. SGR ranged between1.62–1.76 for small and 0.94–1.06 for large size gilt-head sea bream. Increasing the level of either type ofSBM had no significant effect on SGR for fish of bothsizes. Mean daily feed intake ranged between 3.51–4.45for small and 2.05–2.76 for large fish. For small fishfeed consumption increased significantly with inclusionlevel for both forms of soybean meal and differencesamong slopes were not significant. Large fish respondedalso linearly to SBM inclusion, however no responsewas apparent to SBMex inclusion, this leading to asignificant difference among the slopes of the two lines.Inclusion level of both forms of soybean meal increasedsignificantly FCR for both small and large fish. Com-parison of the slopes for FCR indicated a significantimprovement with extrusion only for large fish. Soy-bean meal inclusion decreased PER and PPV signifi-cantly for both fish sizes and types of soybean meal.
Table 7Liver composition (percent of wet weight) and hepatosomatic index of 9 g (small) mean initial weight gilthead sea bream fed diets containing soybeanmeal, before (SBM) and after its extrusion (SBMex), at increasing levels (experiment I)
Diets C S1 S2 S3 E1 E2 E3
Liver composition (% wet weight)Glycogen 7.0±0.5b 5.8±0.3a,b 5.6±0.0a 5.5±0.4a 5.7±0.1a 6.2±0.1a,b 6.8±0.0a,b
Total lipid 15.2±0.6b,c,d 14.3±0.5a,b 15.0±0.4b,c 16.5±0.08d 13.5±0.08a 14.4±0.2a,b 16.1±0.1c,d
Hepatosomatic index 2.32±0.06b 2.20±0.02b 1.86±0.06a 2.20±0.01b 1.93±0.03a 1.96±0.08a 1.93±0.04a
Values are means n=6 per tank±standard deviation.Means with common superscript are not significantly different (P≤0.05).
Table 8Liver composition (percent of wet weight), hepatosomatic index and plasma cholesterol (mg/dl) of 50 g (large) mean initial weight gilthead sea breamfed diets containing soybean meal, before (SBM) and after its extrusion (SBMex), at increasing levels (experiment II)
Diets C S1 S3 E1 E2 E3
Liver composition (% wet weight)Glycogen 7.0±0.4a 6.4±0.06a 6.3±0.04a 7.0±0.2a 6.8±0.1a 6.3±0.04a
Total lipid 19.6±0.3b 19.5±0.4b 17.7±0.2a 19.1±0.2b 17.9±0.1a 17.4±0.1a
Hepatosomatic index 2.60±0.03c 2.32±0.04b 2.01±0.04a 2.34±0.04b 2.15±0.03a,b 2.05±0.06a
Plasma cholesterol (mg/dl) 49.8±3.2c 44.6±0.3b,c 32.6±2.0a 44.7±3.3b,c 43.8±2.6b 42.0±4.9b
Values are means n=6 per tank±standard deviation.Means with common superscript are not significantly different (P≤0.05).
349B. Venou et al. / Aquaculture 261 (2006) 343–356
Autho
r's
pers
onal
co
py
Comparison of the slopes indicated a significant im-provement with extrusion for PPV for both small andlarge fish too.
No significant effect of inclusion level of both formsof soybean meal was apparent on whole body com-position (protein, lipid, moisture, ash) for both small(Table 5) and large (Table 6) fish.
HSI and liver composition are presented in Tables 7and 8 for small and large fish, respectively. No con-sistent trend with soybean meal inclusion was observedfor HSI of small fish. All fish fed SBMex showed
however lower HSI than the control diet. For large fishHSI decreased significantly with dietary soybean meallevel, the values being similar for both SBM and SBMexat similar inclusion levels. The decrease in HSI wasfollowed by decreases in liver lipid content for largefish, significant differences with the control being how-ever apparent only at the two higher soybean meal levelsfor both types of soybean meal, while liver glycogencontent did not change. The trend in liver lipid contentappeared to be different in small fish, however sig-nificant differences with the control diet were observed
Fig. 1. Trypsin activity (units per 100 g fish) in the upper (UI), mid (MI) and lower (LI) intestine of gilthead sea bream 4 h after feeding a single mealof diets containing soybean meal, before (SBM) and after its extrusion (SBMex), at increasing levels. Values are means n=6±standard deviation.Small and capital letters refer to differences among diets in the same intestinal segment and among intestinal segments for the same diet, respectively(P≤0.05).
Fig. 2. Hydration of digesta in ml of supernatant per 100 g of total digestive contents (DCs) in the upper (UI), mid (MI) and lower (LI) intestine ofgilthead sea bream 4 h after feeding a single meal of diets containing soybean meal, before (SBM) and after its extrusion (SBMex), at increasinglevels. Values are means n=6±standard deviation. Small and capital letters refer to differences among diets in the same intestinal segment and amongintestinal segments for the same diet, respectively (P≤0.05).
350 B. Venou et al. / Aquaculture 261 (2006) 343–356

Autho
r's
pers
onal
co
py
only in the case of E1 diet, all other diets giving nostatistically different values compared to the control. Nodifferences in liver glycogen content were observedamong SBM or SBMex containing diets. Significantlylower values than the respective control were observedfor S2, S3 and E1 diets.
Plasma cholesterol measured for large fish decreasedwith soybean level for both types of soybean meal, theeffect being significant, compared to the control diet, atthe two higher inclusion levels. S3 diet indicated thelowest value among all diets tested (Table 8).
3.3. Digesta characteristics
Trypsin activity was decreased from UI to LI for alldiets (Fig. 1). Control diet contained significantly higherlevels of TA than all soybean meal containing diets at allintestinal segments. Among soybean meal containingdiets TAwas highest for E1 at both UI and MI, while S3indicated the lowest TA levels. S1 and E3 diets resultedin similar TA levels in the UI and MI.
Hydration of digesta was affected by soybean mealinclusion as well as from intestinal segment (Fig. 2). Thediets containing both soy products were less hydratedthan the control at the UI with the exception of E1 diet.This changed at the MI, where similar hydrations wereobserved with the exception of S3 diet. At the LI hy-dration of E1 remained significantly higher than that ofthe control.
4. Discussion
Protein digestibility was not affected by either levelor treatment of SBM in accordance with a number ofstudies (Pongmaneerat and Watanabe, 1992, 1993;Allan and Booth, 2004; Tomás et al., 2005) for differentfish species. This could be attributed to the generallylow values of 2–6 mg/g (Snyder and Kwon, 1987) ofTIA resulting from the heat treatment applied duringthermal processing of commercial SBM (Liener, 1980;Francis et al., 2001). Effects of inhibitor levels lowerthan 5 mg/g of feed were found to be negligible forAtlantic salmon (Olli et al., 1994). TIA levels measuredin the present work were 5.4 and 3.6 for SBM andSBMex, respectively, and dietary levels would be lessthan half of these values. However other authors haveobserved a decrease in protein digestibility (Robainaet al., 1995; Refstie et al., 1999) and attributed mainly tothe presence of phytate.
The results of the present study would suggest thatsoybean meal does not affect lipid digestibility by giltheadsea bream, this being in accordance with certain studies for
other species (Pongmaneerat and Watanabe, 1993;Sintayehu et al., 1996; Refstie et al., 2000). However asmall but clear effect could be seen for the highest level ofinclusion for both extruded and non-extruded materialsindicating the possible existence of an effect at high levelsof soybeanmeal. This however needs further confirmation.
Inclusion of SBM in the diets reduced the dietarystarch levels and due to that an increase of starch di-gestibility was expected, as observed in many species(Wilson, 1994; Krogdahl et al., 2005) as well as ingilthead sea bream (Fountoulaki et al., 2005b). An in-crease in the digestibility of starch contained in SBMwas also expected with extrusion due to its gelatiniza-tion (Bergot and Breque, 1983; Pfeffer, 1995; Peres andOliva Teles, 2002; Venou et al., 2003; Krogdahl et al.,2005). Values of starch digestibility tended to be higherfor the extruded product, however the expected increasewith SBM inclusion was not observed. Values at thehighest levels of inclusion of soy products seemed to beeven lower than those at the lowest level indicating apossible negative effect of SBM on starch digestibility.This agrees with the results of Pongmaneerat andWatanabe (1993), which have found, using extrudeddiets, reduction of starch digestibility when 30% SBMwas included in a FM control diet (69.7 and 86.9%respectively).
No palatability problems were apparent for any of thediets in the present study. Robaina et al. (1995) reportedsimilar feed consumption for all SBM levels used in theirstudy, while in the present study feed consumption wasincreased with increasing SBM inclusion. Since it isgenerally accepted that fish eat to satisfy their energeticneeds (Jobling andWandsvik, 1983; Kaushik and Oliva-Teles, 1985; Boujard and Medale, 1994; Paspatis andBoujard, 1996) the increase in feed consumption couldresult from reduced available energy content as SBMinclusion increased. SBM contains about 20% of NSPand 10% oligosaccharides (Snyder and Kwon, 1987;Bach Knudsen, 1997), which are considered indigest-ible. Calculation of the digestible energy content of thefeeds indicated a decrease with SBM inclusion (Table 1).This agrees in general with the changes in feed con-sumption observed although the last were of a largermagnitude.
Extrusion of SBM reduced feed consumption of largefish and values measured were almost constant withSBMex level. This could indicate an improvement ofnutritional value of SBM with extrusion leading to lowerconsumption. However respective values of digestibleenergy content of SBM and SBMex containing feedsindicated only a small improvement with extrusion. Cal-culation of digestible energy values was performed taking
351B. Venou et al. / Aquaculture 261 (2006) 343–356

Autho
r's
pers
onal
co
py
into account only digestible protein fat and starch levels,since digestibility of energy was not measured separatelydue to the small amount of fecal material available. NSPand oligosaccharides are a substrate for microbial flora atthe hindgut and can supply an amount of energy to the fishthrough transformation to low molecular weight fattyacids (Kihara and Sakata, 2002; Mountfort et al., 2002).Oligosaccharides, mainly raffinose, were found to bedigestible by Atlantic salmon and NSP showed also asmall but measurable digestion (Refstie et al., 2005).Utilization of NSP depends on their solubility, solubleNSP being better utilized than insoluble ones (Montagneet al., 2003). Extrusion could increase the soluble fibercontent of SBM as observed in other raw materials(Björck et al., 1984) and that might differentiate their useby microflora and availability of energy to the fish. Inaddition to that soluble NSP are known to retard feedpassage through the gut (Storebakken, 1985) and thiscould be an additional factor affecting the results. Thedifferentiation of utilization of NSP with treatment is ofgreat importance in production of diets of optimalcharacteristics and merits further study. No significantchange in feed consumption was however observed forsmall gilthead sea bream, indicating that small fish couldnot benefit from any change in properties of SBM re-sulting from extrusion.
The increase in feed consumption resulted to similargrowths of fish, irrespective of SBM inclusion levels.Adjustment of feed intake of gilthead sea bream result-ing to similar growths has been also observed in pre-vious studies, in which different protein and energylevels were used (Marti-Palanka et al., 1996; Nengaset al., 1996; Santinha et al., 1999; Fountoulaki et al.,2005a).
The increase in FCR with increasing levels of bothtypes of SBM, for both large and small fish, is inaccordance with other studies (Pongmaneerat andWatanabe, 1992; Arndt et al., 1999; Fagbenro andDavies, 2001; Choi et al., 2004), in which a range of fishsizes was studied. In the present study extrusion de-creased significantly the slope of the curve describingthis relationship for large fish but not for small, con-firming the difference in response of small compared tolarge fish to extrusion.
The only parameter significantly affected by treat-ment for both small and large fish, improving with theSBM extrusion, was retention of protein, in accordancewith other works (Pongmaneerat and Watanabe, 1993;Deguara, 1997; Booth et al., 2002). As discussed beforeprotein digestibility was not significantly affected byextrusion and therefore could not be the factor con-tributing to better protein utilization. Although it has
been shown that low levels of TIA in the diets promoteincreased production of protein digesting enzymes bypancreas, to counteract the binding of enzymes bytrypsin inhibitors (Krogdhal et al., 1994; Olli et al.,1994; Haard et al., 1996), a decreased loss of proteasesdue to decreased TIA levels (Dabrowski et al., 1989;Krogdahl et al., 2003) of extruded meal could possiblycreate this difference. Digestive enzymes are rich insulfur amino acids (Liener, 1980) and their loss couldlead to deficiencies. Methionine was supplied in thediets; however methionine supplementation might beless efficient in satisfying needs for sulphur amino acidsthan cysteine (Nordrum et al., 2000). Changes in thephysicochemical characteristics of the diets due to soyextrusion, could also affect the rate of absorption of freemethionine added, increasing its utilization (Watanabeet al., 2001; Marcouli et al., 2004).
Whole body composition of gilthead sea bream wasnot affected either by inclusion level or treatment ofsoybeanmeal in agreement with several studies with otherfish species (Pongmaneerat and Watanabe, 1992, 1993;Kaushik et al., 1995; Davies andMorris, 1997; Carter andHauler, 2000; Fagbenro and Davies, 2001; Refstie et al.,2001) as well as with gilthead sea bream (Robaina et al.,1995; Nengas et al., 1996). The decrease in plasma cho-lesterol levels was also in agreement with studies withother fish species (Kaushik et al., 1995, 2004; Refstieet al., 1999; Regost et al., 1999; Tulli et al., 1999).
Dietary carbohydrate intake was found to affect lipiddeposition in the liver (Hemre et al., 1989; Brauge et al.,1994; Wilson, 1994; Lanari et al., 1999; Venou et al.,2003; Krogdahl et al., 2004). In the present study asignificant correlation (P=0.023, CC=0.647) was foundbetween digestible starch intake and liver lipid content,for large fish, indicating that starch and not soybeanintake was the main factor affecting this parameter. Thelipid content affected more strongly HSI values than theglycogen content (P=0.0001, CC=0.895 and P=0.026and CC=0.638, respectively). Several studies in manyfish species (Hemre et al., 1989; Brauge et al., 1994;Suarez et al., 1995; Keembiyehetty and Wilson, 1998;Robaina et al., 1998; Venou et al., 2003; Krogdahl et al.,2004) have associated high HSI with increased liver lipidand/or glycogen content. Such effects were not howeverobserved for small fish in the present study.
The changes in TA measured in different intestinalsegments indicated a strong reduction with the level ofsoybean meal either extruded or non-extruded. The in-tensity of the effect was however depended on treatmentand level of inclusion. TA measured in the supernatant ismainly free trypsin. Bound trypsin is the portion of theenzyme bound by feed protein molecules as well as by
352 B. Venou et al. / Aquaculture 261 (2006) 343–356

Autho
r's
pers
onal
co
py
protein inhibitors contained in feed. As previously dis-cussed, low levels of TIA in the diets, as those suppliedby SBM, promote increased production of protein di-gesting enzymes by pancreas, to counteract the bindingof enzymes by TIA (Krogdhal et al., 1994; Olli et al.,1994; Haard et al., 1996). From the results of the presentstudy it appears that this increase was not adequate toincrease TA levels to those obtained for the control diet,even at the low inclusion level of the extruded product.Krogdahl et al. (2003) suggested that there is an ex-haustion of the synthesizing capacity of the pancreas atSBM levels above 15%. The levels of TA attained, in thepresent study, for soy containing diets, in comparison tothose of the control, indicate that production of trypsinby pancreas had also approached a limiting value. Thelowest average values of TA were observed at the LI.This was expected since LI has an important role in theabsorption of intact protein as well as of endogenousdigestive components (Hofer, 1982; McLean et al.,1999; Nordrum et al., 2000).
Increased hydration of the feces of rainbow trout(Refstie et al., 1997) has been found, when fed dietscontaining SBM compared to a control FM diet. Anincrease in hydration at the low levels of soy inclusion,being significant for the extruded diet, was observed in theLI of gilthead sea bream (the composition of which iscloser to the composition of feces). For the higher level atendency to decrease was apparent. This discrepancymight be due to the different ways used to calculatehydration in the present study or to the differences indietary formulations and needs further verification.
5. Conclusions
The data obtained in the present study indicate anadverse effect of inclusion of the SBM products used onfeed conversion ratio and protein productive value forboth fish sizes of gilthead sea bream. Extrusion of soyaffected differently large and small fish improving bothprotein productive value and feed conversion ratio oflarge fish, but only protein productive value of smallfish. Extrusion is a common procedure in fish feedproduction. The results obtained with SBMex couldtherefore give an indication of the contribution of SBMto any changes in FCR and protein retention with itslevel, when it is included in practical extruded diets.From the regression equations and within the range ofsubstitutions used, increasing SBMex dietary level by10% decreased FCR by about 4.2% for large fish and6.6% for small fish, while respective values for proteinproductive value were 2.7% and 3.4%. Since reductionof production cost depends not only on feed cost but
also on feed conversion rate, the increase in FCR shouldbe taken into account when planning fish meal sub-stitution by SBM. More information is however re-quired for the effects of SBM inclusion over prolongedfeeding of gilthead sea bream.
References
Allan, G.F., Booth, M.A., 2004. Effects of extrusion processing ondigestibility of peas, lupins, canola meal and soybean meal in silverperchBidyanus bidyanus (Mitchell) diets. Aquac. Res. 35, 981–991.
Alexis, M.N., Papaparaskeva-Papoutsoglou, E., Theochari, V., 1985.Formulation of practical diets for rainbow trout (Salmo gairdneri)made by partial or complete substitution of fish meal by poultryby-products and certain plant by-products. Aquaculture 50, 61–73.
AOAC, 1990. In: Helrich, K. (Ed.), Official Methods of Analysis, 15thedn. Association of Official Analytical Chemists, Arlington, VA.
Arndt, R.E., Hardy, R.W., Suguira, S.H., Dong, F.M., 1999. Effects ofheat treatment and substitution level on palatability and nutritionalvalue of soy defatted flour in diets for coho salmon, Oncorhynchuskisutch. Aquaculture 180, 129–145.
Arnesen, P., Brattaas, L.E., Olli, J.J., Krogdhal, Å., 1989. Soybeancarbohydrates appear to restrict the utilization of nutrients byAtlanticsalmon (Salmo salar L.). In: Takeda, M., Watanabe, T. (Eds.), TheCurrent Status of Fish Nutrition in Aquaculture. Proc. 3rd Int. Symp.on Feeding and Nutrition in Fish. Toba, Japan, 28 August–1September 1990, pp. 273–280.
Bach Knudsen, K.E., 1997. Carbohydrate and lignin contents of plantmaterials used in animal feeding. Anim. Feed Sci. Technol. 67,319–338.
Bergot, F., Breque, J., 1983. Digestibility of starch by rainbow trout:effects of the physical state of starch and of the intake level.Aquaculture 22, 81–96.
Björck, I., Nyman, M., Asp, N.G., 1984. Extrusion cooking anddietary fiber: effects on dietary fiber content and on degradation inthe rat intestinal tract. Cereal Chem. 61, 174–179.
Bolin, D.W., King, R.P., Klosterman, W.W., 1952. A simplifiedmethod for the determination of chromic oxide (Cr2O3) when usedas an inert substance. Science 116, 634–635.
Boonyaratpalin, M., Suraneiranat, P., Tunpibal, T., 1998. Replacementof fish meal with various types of soybean products in diet for theAsian seabass, Lates calcarifer. Aquaculture 161, 67–78.
Booth, M.A., Allan, G.L., Evans, A.J., Gleeson, V.P., 2002. Effects ofsteam pelleting or extrusion on digestibility and performance ofsilver perch Bidyanus bidyanus. Aquac. Res. 33, 1163–1173.
Boujard, T., Medale, F., 1994. Regulation of voluntary feed intake injuvenile rainbow trout fed by hand or by self feeders with dietscontaining two different protein/energy ratios. Aquat. Living Resour.4, 211–215.
Brauge, C., Medale, F., Corraze, G., 1994. Effect of dietary carbohydratelevels on growth, body composition and glycaemia in rainbow trout,Oncorhynchus mykiss, reared in seawater. Aquaculture 123,109–120.
Burel, C., Boujard, T., Tulli, F., Kaushik, S.J., 2000a. Digestibility ofextruded peas, extruded lupin and rapeseed meal in rainbow trout(Oncorhynchus mykiss) and turbot (Psetta maxima). Aquaculture188, 285–298.
Burel, C., Boujard, T., Kaushik, S.J., Bœuf, G., van der Geyten, S.,Mol, K.A., Kuhn, E.R., Quinsac, A., Krouti, M., Ribaillier, D.,2000b. Potential of plant-protein sources as fish meal substitutes in
353B. Venou et al. / Aquaculture 261 (2006) 343–356

Autho
r's
pers
onal
co
py
diets for turbot (Psetta maxima): growth, nutrient utilization andthyroid status. Aquaculture 188, 363–382.
Carter, C.G., Hauler, R.C., 2000. Fish meal replacement by plant mealsin extruded feeds for Atlantic salmon, Salmo salar L. Aquaculture185, 299–311.
Cataldi, E., Cataudella, S., Monaco, G., Rossi, A., Tancioni, L., 1987.A study of the histology and morphology of the digestive track ofthe sea-bream, Sparus aurata L. J. Fish Biol. 30, 135–145.
Choi, S.M., Wang, X., Park, G.-J., Lim, S.-R., Kim, K.-W., Bai, S.C.,Shin, I.-S., 2004. Dietary dehulled soybean meal as a replacementfor fish meal in fingerling and growing olive flounder Para-lichthys olivaceus (Temminck et Schlegel). Aquac. Res. 35,410–418.
Dabrowski, K., Poczyczynski, P., Köck, G., Berger, B., 1989. Effectof partially and totally replacing fish meal protein by soybeanmeal protein on growth, food utilization and proteolytic enzymeactivities in rainbow trout (Salmo gairdneri). New in vivo test forexocrine pancreatic secretion. Aquaculture 77, 29–49.
Davies, S.J., Morris, P.C., 1997. Influence of multiple amino acidsupplementation on the performance of rainbow trout,Oncorhynchusmykiss (Walbaum), fed soya based diets. Aquac. Res. 28, 65–74.
Deguara, S., 1997. Evaluation of different pressed and extruded fishmeal based diets on the growth of gilthead seabream, Sparus aurataL. In: Tacon, A., Barusco, B. (Eds.), Feeding Tomorrow's Fish,vol. 22. Cashiers Options Méditerranéennes, Institut AgronomiqueMéditerranéen de Zaragoza, Spain, pp. 123–139.
Erlanger, B., Kokowsky, N., Cohen, W., 1961. The preparation andproperties of two new chromogenic substrates of trypsin. Arch.Biochem. Biophys. 95, 271–278.
Fagbenro, O.A., Davies, S.J., 2001. Use of soybean flour (dehulled,solvent-extracted soybean) as a fish meal substitute in practical dietsfor African catfish,Clarias gariepinus (Burchell 1822): growth, feedutilization and digestibility. J. Appl. Ichthyol. 17, 64–69.
Fountoulaki, E.E., Alexis, M.N., Nengas, I., 2005a. Protein and energyrequirements of gilthead bream (Sparus aurata L.) fingerlings.Preliminary results. Cah. Options Mediterr. 63, 19–26.
Fountoulaki, E., Alexis, M.N., Nengas, I., Venou, B., 2005b. Effect ofdiet composition on nutrient digestibility and digestive enzymelevels of gilthead sea bream (Sparus aurata L.). Aquac. Res. 36,1243–1251.
Fowler, L.G., 1980. Substitution of soybean and cottonseed products forfish meal in diets fed to chinook and coho salmon. Prog. Fish-Cult.42, 86–91.
Francis, G., Makkar, H.P.S., Becker, K., 2001. Antinutritional factorspresent in plant derived alternative fish feed ingredients and theireffects in fish. Aquaculture 199, 197–227.
Haard, N.F., Dimes, L.E., Arndt, R.E., Dong, F.M., 1996. Estimationof protein digestibility-IV. Digestive proteinases from the pyloriccaeca of coho salmon (Oncorhynchus kisutch) fed diets containingsoybean meal. Comp. Biochem. Physiol. 115B, 533–540.
Hajen, W.E., Higgs, D.A., Beames, R.M., Dosanjh, B.S., 1993.Digestibility of various feedstuffs by post juvenile chinook salmon(Oncorhynchus tshawytscha) in sea water: measurement of digest-ibility. Aquaculture 112, 333–348.
Hemre, G.-I., Lie, Ø., Lied, E., Lambertsen, G., 1989. Starch as anenergy source in feed for cod (Gadus morhua): digestibility andretention. Aquaculture 80, 261–270.
Hofer, R., 1982. Protein digestion and proteolytic activity in the digestivetract of an omnivorous cyprinid. Comp. Biochem. Physiol. 72A,55–63.
Ikegami, S., Tsuchihashi, F., Harada, H., Tsuchihashi, N., Nishide, E.,Innami, S., 1990. Effect of viscous indigestible polysaccharides on
pancreatic-biliary secretion and digestive organs in rat. J. Nutr.120, 353–360.
Jobling, M., Wandsvik, A., 1983. An investigation of factors con-trolling food intake in Arctic charr, Salvelinus alpinus L. J. FishBiol. 23, 397–404.
Kakade, M.L., Rackis, J.J., McGhee, J.E., Puski, G., 1974. Trypsininhibitors in soy: collaborative analysis of an improved procedure.Cereal Chem. 51, 376–382.
Kaushik, S.J.,Oliva-Teles,A., 1985. Effect of digestible energy onnitrogenand energy balance in rainbow trout. Aquaculture 50, 89–101.
Kaushik, S.J., Cravedi, J.P., Lalles, J.P., Sumpter, J., Fauconneau, B.,Laroche, M., 1995. Partial or total replacement of fish meal bysoybean protein on growth, protein utilization, potential estro-genic or antigenic effects, cholesterolemia and flesh qualityin rainbow trout, (Oncorhynchus mykiss). Aquaculture 133,257–274.
Kaushik, S.J., Covès, D., Dutto, G., Blanc, D., 2004. Almost totalreplacement of fish meal by plant protein sources in the diet of amarine teleost, the European seabass, Dicentrarchus labrax.Aquaculture 230, 391–404.
Keembiyehetty, C.N., Wilson, R.P., 1998. Effect of water temperatureon growth and nutrient utilization of sunshine bass (Moronechrysops ♀×Morone saxatiles ♂) fed diets containing differentenergy/protein ratios. Aquaculture 166, 151–162.
Khan, M.A., Jafri, A.K., Chadha, N.K., Usmani, N., 2003. Growth andbody composition of rohu (Labeo rohita) fed diets containingoilseed meals: partial or total replacement of fish meal with soy-bean meal. Aquac. Nutr. 9, 391–396.
Kihara, M., Sakata, T., 2002. Production of short-chain fatty acids andgas from various oligosaccharides by gut microbes of carp(Cyprinus carpio L.) in microscale batch culture. Comp. Biochem.Physiol. 132A, 333–340.
Kotzamanis, Y.P., Alexis, M.N., Andriopoulou, A., Castritsi-Cathar-iou, Fotis, G., 2001. Utilization of waste material resulting fromtrout processing in gilthead bream (Sparus aurata L.) diets. Aquac.Res. 32, 1–9.
Krogdhal, Å., Lee, T.B., Olli, J.J., 1994. Soybean protease inhibitorsaffect intestinal trypsin activities and amino acids digestibilities inrainbow trout (Oncorhyncus mykiss). Comp. Biochem. Physiol.107A, 215–219.
Krogdahl, Å., Bakke-Mckellep, A., Baeverfjord, G., 2003. Effects ofgraded levels of standard soybean meal on intestinal structure,mucosal enzyme activities, and pancreatic response in Atlanticsalmon (Salmo salar L.). Aquac. Nutr. 9, 361–371.
Krogdahl, Å., Sundby, A., Olli, J.J., 2004. Atlantic salmon (Salmosalar L.) and rainbow trout (Oncorhynchus mykiss) digest andmetabolise nutrients differently. Effects of water salinity and die-tary starch level. Aquaculture 229, 335–360.
Krogdahl, Å., Hemre, G.-I., Mommsen, T.P., 2005. Carbohydrates infish nutrition: digestion and absorption in postlarval stages. Aquac.Nutr. 11, 103–122.
Lanari, D., Poli, B.M., Ballestrazzi, R., Lupi, P., D'Agaro, E., Mecatti,M., 1999. The effects of dietary fat and NFE levels on growingEuropean sea bass (Dicentrarchus labrax). Growth rate, body andliver composition, carcass traits and nutrient retention efficiency.Aquaculture 179, 351–364.
Liener, I.E., 1980. Toxic Constituents of Plant Foodstuffs. AcademicPress, New York 10003, NY, pp. 1–502.
Marcouli, P.A., Alexis,M.N., Andriopoulou, A., Iliopoulou-Georgudaki,J., 2004. Development of a reference diet for use in indispensableamino acid requirement studies of gilthead sea bream Sparus aurataL. Aquac. Nutr. 10, 335–344.
354 B. Venou et al. / Aquaculture 261 (2006) 343–356

Autho
r's
pers
onal
co
py
Marti-Palanka, H., Martinez-Barbera, J.P., Pendon, C., Valdivia, M.M.,Perez-Sanchez, J., Kaushik, S.J., 1996.Growth hormone as a functionof age and dietary protein: energy ratio in amarine teleost, the giltheadsea bream (Sparus aurata L.). Growth Regul. 6, 253–259.
McGoogan,B.B.,Gatlin III,D.M., 1997.Effects of replacing fishmealwithsoybean meal in diets for red drum, Sciaenops ocellatus and potentialfor palatability enhancement. J. World Aquac. Soc. 28, 374–385.
McLean, E., Rønsholdt, B., Sten, C., Najamuddin, 1999. Gastrointes-tinal delivery of peptide and protein drugs to aquacultured teleosts.Aquaculture 177, 231–247.
Montagne, L., Pluske, J.R., Hampson, D.J., 2003. A review of inter-actions between dietary fibre and the intestinal mucosa, and theirconsequences on digestive health in young non-ruminant animals.Anim. Feed Sci. Technol. 108, 95–117.
Mountfort, D.O., Campbell, J., Clements, K.D., 2002. Hindgut fer-mentation in three species of marine herbivorous fish. Appl.Environ. Microbiol. 68, 1374–1380.
Murai, T., Ogata, H., Villaneda, A., Watanabe, T., 1989. Utilization ofsoy flour by fingerling rainbow trout having different body size.Bull. Jpn. Soc. Sci. Fish. 55, 1067–1073.
Nengas, I., Alexis, M., Davies, S.J., Petichakis, G., 1995. An in-vestigation to determine the digestibility coefficients of variousraw materials in diets for the gilthead sea bream, Sparus aurata L.Aquac. Res. 26 (3), 185–194.
Nengas, I., Alexis, M., Davies, S.J., 1996. Partial substitution offish meal with soybean meal products and derivatives in dietsfor the gilthead sea bream Sparus aurata L. Aquac. Res. 27,147–156.
Nordrum, S., Krogdahl, Å., Rosjo, C., Olli, J.J., Holm, H., 2000.Effect of methionine cysteine and medium chain triglycerides onnutrient digestibility absorption of amino acids along the intestinaltract and nutrient retention in Atlantic salmon (Salmo salar L.)under pair-feeding regime. Aquaculture 186, 341–360.
Ogunkoya, A.E., Page, G.I., Adewolu, M.A., Bureau, D.P., 2006.Dietary incorporation of soybean meal and exogenous enzymecocktail can affect physical characteristics of faecal materialegested by rainbow trout (Oncorhynchus mykiss). Aquaculture254, 466–475.
Olli, J.J., Kroghahl, Å., 1995. Alcohol soluble components of soy-beans seem to reduce fat digestibility in fish-meal-based diets forAtlantic salmon, Salmo salar L. Aquac. Res. 26, 831–835.
Olli, J.J., Hjelmeland, K., Kroghahl, Å., 1994. Soybean trypsininhibitors in diets for Atlantic salmon (Salmo salar, L.): effects onnutrient digestibilities and trypsin in pyloric caeca homogenate andintestinal content. Comp. Biochem. Physiol. 109A, 923–928.
Paspatis, M., Boujard, T., 1996. A comparative study of automaticfeeding and self-feeding in juvenile Atlantic salmon (Salmo salar)fed diets with different energy levels. Aquaculture 145, 245–247.
Peres, H., Oliva Teles, A., 2002. Utilization of raw and gelatinizedstarch by European sea bass (Dicentrarchus labrax) juveniles.Aquaculture 205, 287–299.
Pfeffer, E., 1995. Carbohydrate utilization and its determination. J. Appl.Ichthyol. 11, 175–182.
Pfeffer, E., Beckmann-Toussaint, J., Henrichfreise, B., Jansen, H.D.,1991. Effect of extrusion on efficiency of utilization of maize starchby rainbow trout (Oncorhynchus mykiss). Aquaculture 96, 293–303.
Pongmaneerat, J., Watanabe, T., 1992. Utilization of soybean meal asprotein source in diets for rainbow trout. Nippon Suisan Gakkaishi58, 1983–1990.
Pongmaneerat, J., Watanabe, T., 1993. Effect of extrusion processingon the utilization of soybean meal diets for rainbow trout. NipponSuisan Gakkaishi 59, 1407–1414.
Potter, S.M., 1995. Overview of proposed mechanisms for thehypocholesterolemic effect of soy. J. Nutr. 125, 606–611.
Refstie, S., Helland, J., Storebakken, T., 1997. Adaptation to soybeanmeal in diets for rainbow trout, Oncorhynhus mykiss. Aquaculture153, 263–272.
Refstie, S., Storebakken, T., Roem, A., 1998. Feed consumption andconversion in Atlantic salmon (Salmo salar) fed diets with fishmeal, extracted soybeanmeal or soybeanmeal with reduced contentof oligosaccharides, trypsin inhibitors, lectins and soya antigens.Aquaculture 162, 301–312.
Refstie, S., Svihus, B., Shearer, K.D., Storebakken, T., 1999. Nutrientdigestibility in Atlantic salmon and broiler chickens related to viscosityand non-starch polysaccharide content in different soyabean products.Anim. Feed Sci. Technol. 79, 331–345.
Refstie, S., Korsøen, Ø.J., Storebakken, T., Baeverfjord, G., Lein, I.,Roem, A.J., 2000. Differing nutritional responses to dietary soy-bean meal in rainbow trout (Oncorhynchus mykiss) and Atlanticsalmon (Salmo salar). Aquaculture 190, 49–63.
Refstie, S., Storebakken, T., Baeverfjord,G., Roem,A.J., 2001. Long-termprotein and lipid growth of Atlantic salmon (Salmo salar) fed dietswith partial replacement of fish meal by soy protein products atmedium or high lipid level. Aquaculture 193, 91–106.
Refstie, S., Sahlstrom, S., Brathen, E., Baeverfjord, G., Krogedal, P.,2005. Lactic acid fermentation eliminates indigestible carbohy-drates and antinutritional factors in soybean meal for Atlanticsalmon (Salmo salar). Aquaculture 246, 331–345.
Regost, C., Arzel, J., Kaushik, S.J., 1999. Partial or total replacementof fish meal by corn gluten meal in diet for turbot (Psetta maxima).Aquaculture 180, 99–117.
Robaina, L., Izquierdo, M.S., Moyano, F.J., Socorro, J., Vergara, J.M.,Montero, D., Fernández-Palacios, H., 1995. Soybean and lupinmeals as protein sources in diets for gilthead sea bream (Sparusaurata): nutritional and histological implications. Aquaculture130, 219–233.
Robaina, L., Izquierdo, M.S., Moyano, F.J., Socorro, J., Vergara, J.M.,Montero, D., 1998. Increase of the dietary n-3/n-6 fatty acid ratioand addition of phosphorus improves liver histological alterationsinduced by feeding diets containing soybean meal to gilthead seabream, Sparus aurata. Aquaculture 161, 281–293.
Rubio, L.A., Rodriguez, J., Fernández, C., Crespo, J.F., 2004. Storageproteins: physiological and antigenic effects. In: Muzquiz, M., Hill,G.D., Cuadrado, C., Pedrosa, M.M., Burbano, C. (Eds.), RecentAdvances of Research in Antinutritional Factors in Legume Seedsand Oilseeds. Proceedings of the 4th International Workshop onAntinutritional Factors in Legume Seeds and Oilseeds. Toledo,Spain, March 2004. EAAP Publication, vol. 110. WageningenAcademic Publishers, pp. 159–175.
Santinha, P.J.M., Medale, F., Corraze, G., Gomes, E.F.S., 1999.Effects of dietary protein/lipid ratio on growth and nutrientutilization in gilthead sea bream (Sparus aurata L.). Aquac. Nutr.5, 147–156.
Sintayehu, A., Mathies, E., Meyer-Burgdorff, K.-H., Rosenow, H.,Günther, K.-D., 1996. Apparent digestibilities and growth experi-ments with tilapia (Oreochromis niloticus) fed soybean meal,cottonseed meal and sunflower seed meal. J. Appl. Ichthyol. 12,125–130.
Snyder, H.E., Kwon, T.W., 1987. Soybean Utilization. Van NostrandReinhold, New York, NY, USA. 346 pp.
Spinelli, J., Houle, C.R., Wekell, J.C., 1983. The effect of phytates onthe growth of rainbow trout (Salmo gairdneri) fed purified dietscontaining various quantities of calcium and magnesium. Aqua-culture 30, 71–83.
355B. Venou et al. / Aquaculture 261 (2006) 343–356

Autho
r's
pers
onal
co
py
Storebakken, T., 1985. Binders in fish feeds. I: effect of alginate andguar gum on growth, digestibility, feed intake and passage throughthe gastrointestinal track of rainbow trout. Aquaculture 47, 11–26.
Storebakken, T., Shearer, K.D., Roem, A.J., 1998a. Availability ofprotein, phosphorus, zinc and other elements in fishmeal, soy-proteinconcentrate and phytase-treated soy-protein-concentrate-based dietsto Atlantic salmon, Salmo salar. Aquaculture 161, 365–379.
Storebakken, T., Kvien, I.S., Shearer, K., Grisdale-Helland, B.,Helland, S.J., Berge, G.M, 1998b. The apparent digestibility ofdiets containing fish meal, soybean meal or bacterial meal fed toAtlantic salmon (Salmo salar): evaluation of different faecal col-lection methods. Aquaculture 169, 195–210.
Storebakken, T., Refstie, S., Ruyter, B., 2000. Soy products as fat andprotein sources in fish feeds for intensive aquaculture. In:Drackley, J.K. (Ed.), Soy in Animal Nutrition. Fed. Anim. Sci.Soc., Savoy, IL, USA, pp. 127–170.
Suarez, M.D., Hidalgo, M.C., Gallego, M.G., Sanz, A., de la Higuera,M., 1995. Influence of the relative proportions of energy yieldingnutrients on liver intermediary metabolism of the European eel.Comp. Biochem. Physiol. 111A, 421–428.
Sugano, M., Goto, S., Yamada, Y., Yoshida, K., Hashimoto, Y., Matsuo,T., Kimoto, M., 1990. Cholesterol lowering activity of variousundigested fractions of soybean protein in rats. J. Nutr. 120, 977–985.
Sujiura, S.H., Gabaudan, J., Dong, F.M., Hardy, R.W., 2001. Dietarymicrobial phytase supplementation and the utilisation of phospho-rus, trace minerals and protein by rainbow trout (Oncorhynchusmykiss Walbaum) fed soybean-meal based diets. Aquac. Res. 32,583–592.
Tacon, A.G.J., 1987. The nutrition and feeding of farmed fish andshrimp—a training manual 1. The essential nutrients. Food andAgriculture Organization of the United Nations, GCP/RLA/075/ITA, Brazil, p. 117.
Tacon, A.G.J., 1997. Fishmeal replacers: review of antinutrients withinoilseeds and pulses — a limiting factor for the aquafeed greenrevolution? In: Tacon, A., Barusco, B. (Eds.), Feeding Tomorrow'sFish, vol. 22. Cashiers Options Méditerranéennes, Institut Agrono-mique Méditerranéen de Zaragoza, Spain, pp. 153–182.
Thivend, P., Mercier, C., Guilbot, A., 1972. Determination of starchwith glucoamylase. In: Whistler, R.L., Bemiller, J.N. (Eds.),Methods in Carbohydrate Chemistry, vol. VI. Academic Press,New York, pp. 100–105.
Tomás, A., De la Gándara, F., García-Gomez, A., Pérez, A., Jover, L.,2005. Utilization of soybean meal as an alternative protein sourcein the Mediterranean yellowtail, Seriola dumerili. Aquac. Nutr. 11,333–340.
Tulli, F., Tibaldi, E., Comin, A., 1999. Dietary protein sources differentlyaffect plasma lipid levels and body fat deposition in juvenile sea bass.In: Piva, G., Bertoni, G., Masoero, F., Bani, P., Calamari, L. (Eds.),Recent Progress in Animal Production Science. Franco Angeli,Milano, Italia, pp. 782–784.
Venou, B., Alexis, M.N., Fountoulaki, E., Nengas, I., Apostolopoulou,M., Castritsi-Cathariou, I., 2003. Effect of extrusion ofwheat and cornon gilthead sea bream (Sparus aurata) growth, nutrient utilizationefficiency, rates of gastric evacuation and digestive enzyme activities.Aquaculture 225, 207–223.
Watanabe, T., Akimoto, A., Aoki, H., Shimeno, S., 2001. Effects ofphysical properties of diets on evacuation time of digesta and plasmafree amino acid patterns in yellowtail. Fish. Sci. 67, 456–460.
Wilson, R.P., 1994. Utilization of dietary carbohydrate by fish.Aquaculture 124, 67–80.
356 B. Venou et al. / Aquaculture 261 (2006) 343–356




















