
Mast cell responses to Ergasilus (Copepoda), a gill ectoparasite of sea bream
9
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Mast cell responses to Ergasilus (Copepoda), a gill ectoparasite of sea bream

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Mast cell responses to Ergasilus (Copepoda), a gill ectoparasite of sea bream
Bahram S. Dezfuli a,*, Luisa Giari a, Alice Lui a, Massimo Lorenzoni b, Edward J. Noga c
aDepartment of Biology & Evolution, University of Ferrara, St. Borsari 46, 44123 Ferrara, ItalybDepartment of Cellular and Environmental Biology, University of Perugia, 06123 Perugia, ItalycDepartment of Clinical Sciences, College of Veterinary Medicine, North Carolina State University, 4700 St. Hillsborough, Raleigh, NC 27606, USA
a r t i c l e i n f o
Article history:Received 29 November 2010Received in revised form4 February 2011Accepted 6 February 2011Available online 18 February 2011
Keywords:Sea breamCrustacean ectoparasiteGill cell responsesMast cellsPiscidin
a b s t r a c t
Immunocytochemical, light microscopy and ultrastructural studies were conducted on gill of sea bream,Sparus aurata L., naturally parasitized with the important parasitic copepod Ergasilus sp. to assesspathology and cellular responses. Thirty-seven S. aurata were examined from a fish farm; 26 (70%) wereparasitized, with infection intensity ranging from 3 to 55 parasites per fish. Hosts were divided into twogroups, lightly infected fish (<15 parasites per fish) and heavily infected fish (>15 parasites per fish). Inhistological sections, the copepod encircled gill lamellae with its second antennae, compressed theepithelium, provoked hyperplasia and hemorrhage, occluded arteries and often caused lamellardisruption. Fusion of the secondary lamellae due to epithelial hyperplasia was common in all infectedfish; heavily infected fish showed more intense branchial inflammation. In both healthy and infected fish,mast cells (MCs) were free within the connective tissue inside and outside the blood vessels of theprimary lamellae and made close contact with vascular endothelial cells, mucous cells and rodlet cells(RCs). MCs were irregular in shape with a cytoplasm filled by numerous electron-dense, membrane-bound granules. Immunostaining of primary and secondary gill filaments with an antibody against theantimicrobial peptide (AMP) piscidin 3 (anti-piscidin 3 antibody, anti-HAGR) revealed a subpopulation ofMCs that were positive. These MCs were more abundant in gills of heavily infected fish than in eitherlightly infected or uninfected fish (ANOVA, P< 0.05). Our report documents the response of gill toectoparasite infection and provides further evidence that mast cells and their AMPs may play a role inresponding to branchial ectoparasite infections.
� 2011 Elsevier Ltd. All rights reserved.
1. Introduction
One of the most common groups of marine ectoparasites of fishare crustaceans belonging to the order Copepoda. While parasiticcopepods are common on cultured and wild marine fish [1], it hasonly been with the development of semi-intensive and intensiveaquaculture that their importance as disease-causing agents hasbecome evident [2e4]. Ergasilids (Ergasilidae) are commoncopepod parasites of fish, primarily infecting the gills [5]. Appar-ently, feeding activity and mode of attachment of these copepodsare the main reason for host damage and has been addressed inprevious studies [6,7]. According to Johnson et al. (2004) [4],outbreaks of disease due to ergasilids are a major source ofcopepod-induced mortality in brackish and freshwater fish cultureof several countries, including Israel [8], Taiwan [9], Japan [10],China [11], the USA [12], South Africa [13] and Hungary [14]. In Italy,
in our study area, in summer 2006, the copepod, Ergasilus sp.induced mortality of 40% of the juvenile sea bream, Sparus aurata,in a semi-intensively cultured fish farm. Subsequently, productionof S. aurata has been impaired by annual losses of w5e8% of thejuvenile stock each summer since 2007. These losses have beenattributed to heavy infections of Ergasilus sp.
Understanding S. aurata’s immune response to Ergasilus sp.might allow a better assessment of not only host damage but alsoprovide a better understanding of how this fish defends itselfagainst this parasite, so that potential mitigation strategies mayeventually be developed. In the present study, the occurrence ofmast cells (MCs), mucous cells and rodlet cells (RCs) was docu-mented, especially near the Ergasilus sp. attachment site. Changesin the number of mast cells and mucous cells have been reported asa response to the attachment of parasitic copepods in both fresh-water and marine fish [15e17].
With regard to RCs, their presence within the epithelial tissuesof most species of teleost fish examined, has been known for almost120 years. Their widespread distribution within and across manyspecies of marine and freshwater fish and their morphological
* Corresponding author. Tel.: þ39 0532 455701; fax: þ39 0532 455715.E-mail address: [email protected] (B.S. Dezfuli).
Contents lists available at ScienceDirect
Fish & Shellfish Immunology
journal homepage: www.elsevier .com/locate / fs i
1050-4648/$ e see front matter � 2011 Elsevier Ltd. All rights reserved.doi:10.1016/j.fsi.2011.02.005
Fish & Shellfish Immunology 30 (2011) 1087e1094

Author's personal copy
features are well established, but, the function of these cellscontinues to be hotly debated [18,19]. Published records on thepresence of RCs in relation to copepod ectoparasites are rare, withthe only available information referring to Ergasilus sieboldi-infec-ted gills of Abramis brama [17].
The initial response of a host in defending against a parasite/pathogen is via non-specific immunity [20]. Mast cells have beenrecognized as important components of non-specific immunedefence in many vertebrates [21e23]. Mast cells occur in skin, gills,and digestive tract of several species of fish, and in fish infectedwith metazoan parasites [19,24e27]. Antimicrobial peptides(AMPs), host defense effector molecules occur in virtually all lifeforms and are a key component of this defense [28].
Increasing evidence supports their significant role in protectinganimals from infections, including fish [22]. Piscidins are theprototypical AMP present in piscine MCs [16,21e23,27,29e31].Piscidins have potent, broad-spectrum antibacterial and antifungalactivity and have strong antiparasitic activity [16,21,27]. Withregard to their mechanism of action, piscidins are thought to inhibitthe synthesis of the cell wall, nucleic acids, and proteins or eveninhibit enzymatic activity [32]. Piscidins were encountered in mastcells of different organs of uninfected fish [23,30,31,33]. Indeed,piscidin 3 was detected in mast cells of infected gills of Europeansea bass, Dicentrarchus labrax [27] and Latris lineata [16].
The main aim of this investigation was to examine gill cellularresponses of S. aurata to Ergasilus sp. infection in relationship to theintensity of infection. We also provide further evidence for thepossible importance of piscidin-expressing MCs in host defenseagainst ectoparasitic copepods. We also relate the observationsmade on the histopathology and ultrastructure of the immune cellsof infected gill with their involvement in the gill inflammatoryresponse.
2. Materials and methods
2.1. Animals
Thirty-seven sea bream, S. aurata, measuring 18.16� 5.22 cm intotal length (mean� S.D.) andweighing73.36� 25.23 g (mean� S.D.)were obtained froma local semi-intensivefish farm, “Valle Ca’Zuliani”(Pila di Porto Tolle, Rovigo, Italy) from September to December 2009.On this site, sea bream are artificially reproduced in the hatcheryunit and the fry grown on in internal concrete tanks. Once the fryreach between 2 and 5 g in weight, they are then transferred to, andreared in, earthernpondswhich are exposed to seawater. It is believedthat infection is acquired at this point, directly from the water source.Infected fish can be found all year round but the prevalenceand intensity of infection is higher in the summer months. In 2009,the water temperature in the ponds ranged from 3.3 �C to 30.3 �C(16.6� 7.7 C, mean� S.D., n¼ 12) whilst the salinity over thesameperiodwas 12.6 ppt to 38.0 ppt (25.5� 4.3,mean� S.D., n¼ 12).
2.2. Histology and electron microscopy
The fish were transported live to the laboratory where theywere anesthetized using MS222 (Sandoz, Basel, Switzerland) andtheir spinal cords severed. Immediately after euthanasia, the entiregill was placed in a dish of saline solution and, using a steromicro-scope, the total number of Ergasilus sp. were counted on the entiregill, including that of the left and right sides of the fish. The infectedfilaments were then fixed in chilled (4 �C) 10% neutral bufferedformalin for 8 h. The samples were then paraffin-embeddedfollowing standard procedures and 5 mm sections were stainedwith Giemsa. For light and electron microscopy, infected gill fila-ments up to 7 mm in diameter were fixed in 2.5% glutaraldehyde in
0.1 M cacodylate buffer for 3 h at 4 �C before post-fixing them in 1%osmium tetroxide in the same buffer for 3 h. The specimenswere dehydrated through a graded acetone series before beingembedded in Epoxy resin (Durcupan ACM, Fluka, Buchs,Switzerland). Semi-thin sections (1.5 mm) were cut on a ReichertOm U 2 ultramicrotome using glass knives and then stained withtoluidine blue. Ultra-thin sections (90 nm) were stained with a 4%uranyl acetate solution in 50% ethanol and Reynold’s lead citrateand examined using a Hitachi H-800 electron microscope.
For comparative purposes, gills of 11 uninfected sea breamweresimilarly processed. Light photomicrographs were taken usinga Nikon Eclipse 80i microscope (Nikon, Tokyo, Japan). Images of gillsections showing the overall distribution of mast cells anddimensions (150 MCs) were obtained using computerized imageanalysis software (Nis Elements AR 3.0, Nikon, Tokyo, Japan).
2.3. Immunohistochemistry
Anti-piscidin 3 (anti-HAGR) antibody was produced bya commercial laboratory (Bethyl Laboratories, Montgomery, TX)using the company’s standard procedures. Briefly, 2 mg of a 12-merpeptide constituting the C-terminus of piscidin 3 (HAGRSIGRFLTG)was conjugated to keyhole limpet hemocyanin (KLH) using mal-eimide chemistry, which linked the peptide to KLH via a cysteineadded to the N-terminal histidine. The conjugation via the terminalamino acid allows tertiary conformation of the peptide that may beexpected to mimic that in the native peptide, thus eliciting anti-conformational antibodies important for recognizing the nativepeptide. The immunogen was mixed with Complete Freund’sAdjuvant (1:1) and the KLH-conjugated peptide was injected intotwo New Zealand white rabbits biweekly at 5 subcutaneous sites(0.2 mL per site) using the following immunization schedule(100 mg/injection): Days 0, 14, 28 and 42. Thirty ml of antiserumwas collected from each rabbit on days 35 and 45. The antiserawerepooled and were then affinity-purified using the piscidin fragmentconjugated to cyanogen bromide-activated agarose as an immu-nosorbent (10.5 mg of piscidin fragment was reacted with 15 g ofagarose). One hundred and twenty ml of antiserum (two 30 mLbleeds from two rabbits) was loaded onto each affinity column(Uniflow 4, Sterogene, Carlsbad, CA). After washing, the affinity-purified antibody was eluted and concentrated.
This method produces greater than 0.1 mg of peptide-specificantibody per mL of antiserum, as determined by recovered affinity-purified antibody. Antibodywas greater than 95% IgG, as determinedby immunoelectrophoresis using antibodies specific for rabbit IgG,IgM and serum proteins. The titer of the antibodies was determinedvia ELISA, using the piscidin fragment as the antigen coated ontoa microtiter plate. The 12-mer peptide (10 mgmL�1 in PBS, pH7.2e7.5) was coated onto microtiter plates at room temperature for1 h. The plate was thenwashed and post-coated with 1% BSA in PBSfor 30 min. The plate was washed and then dilutions of antibody in1%BSA/PBS/0.01%Tween20were added, beginningat 1 mg antibody/mL. After incubation for 1 h, the plate was washed, followed byaddition of peroxidase-conjugated goat anti-rabbit IgG (h&l) in 1%BSA/PBS/0.01% Tween 20. After incubation for 1 h, the plate waswashed and peroxidase substrate (Tetramethylbenzidene PeroxideSingle Reagent [KPL]) was added, incubated for 15 min, and thenstoppedwith 1 NHCl (1:1). The absorbancewas then read at 450 nm.The titer was read as the reciprocal of the antibody dilution (dilutionof a 1 mg/mL solution) that produced a net optical density of 1.0,compared to a blank (peptide-coatedwellwithno antibody [diluentsonly] added),which hadanOD< 0.1. The ELISA titer of the antibodiesused in all assays was approximately 1:18,000.
The peptide-specific antibody is less than 1% cross-reactive withunrelated antigens by ELISA, where 1% cross-reactivity means that,
B.S. Dezfuli et al. / Fish & Shellfish Immunology 30 (2011) 1087e10941088
Author's personal copy
using the same concentrations of antigen as for the piscidin frag-ment, 100 times more antibody would be required to produce thesame optical density with either free KLH, or a free peptide thatshared less than 3 amino acids in the sequence.
Piscidin 3-positive cells were detected in gill histologicalsections using the indirect immunohistochemical method (perox-idase-anti-peroxidase immunocomplex). Briefly, sections (5 mm)were deparaffinised in xylene, rehydrated, then endogenousperoxidase activity and non-specific staining were blocked in 3%H2O2 (10 min) and in goat normal serum (1:20) (30 min). Afterincubation with the primary antibody (anti-HAGR diluted 1:400)for 3 h at room temperature, sections were incubated for 30 minwith goat anti-rabbit immunoglobulins 1:100 (DAKO, Milan, Italy),and then for 30 min with rabbit peroxidase-antiperoxidase 1:200(DAKO). The sections were then developed using DAB (3,30-dia-minobenzidine 0.04% w/v in TBS 0.05 M, pH 7.4) and H2O2 (0.005%),rinsed and counterstained with Alcian Blue and Harris’s haema-toxylin. Non immune rabbit serum and diluent only sections wereused as negative controls. Positive control tissue was striped bass(Morone saxatilis Walbaum) intestine.
2.4. Statistical analysis
The number of mast cells (MCs) positive to anti-HAGR wascounted in the gills via light microscopy, using computerized imageanalyser software (Nis Elements AR 3.0). For comparative purposes,the cells were screened at 400x magnification in 10 areas of21,000 mm2 (5 randomly selected areas of primary lamella and 5randomly selected areas of secondary lamella) for each fish of thethree groups: uninfected fish (n¼ 11), lightly infected fish (3 to 15parasites per fish, 8.00� 4.97, mean� SD, n¼ 7), heavily infectedfish (39 to 55 parasites per fish, 46.78� 4.89, mean� SD, n¼ 9). TheGaussian distributions (i.e. normality) of the data were assessed
using the KolmogoroveSmirnov Test. Using the software packageSTATISTICA 7, ANOVA repeated measures was performed to detectsignificant differences in the number of the above mentioned cellsamong groups (uninfected vs. lightly infected vs. heavily infected).A Bonferroni post-hoc test and a P< 0.05 level of significanceselected was used.
3. Results
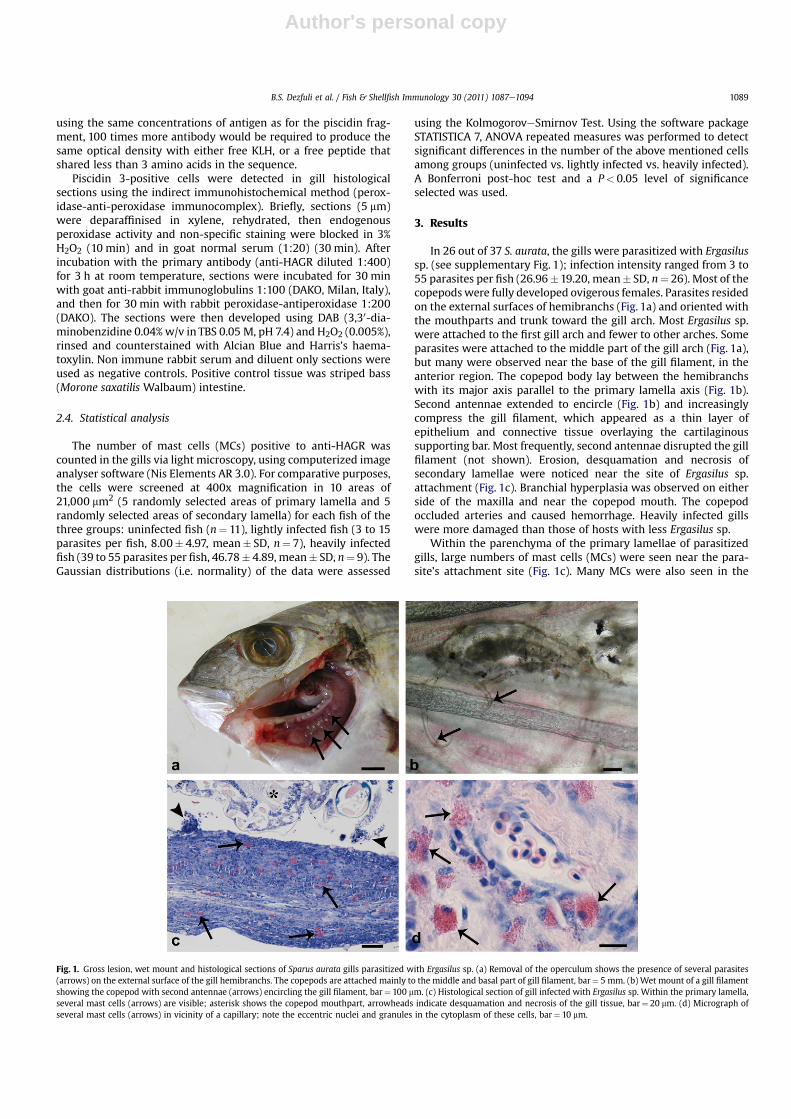
In 26 out of 37 S. aurata, the gills were parasitized with Ergasilussp. (see supplementary Fig. 1); infection intensity ranged from 3 to55 parasites per fish (26.96�19.20, mean� SD, n¼ 26). Most of thecopepodswere fully developed ovigerous females. Parasites residedon the external surfaces of hemibranchs (Fig. 1a) and oriented withthe mouthparts and trunk toward the gill arch. Most Ergasilus sp.were attached to the first gill arch and fewer to other arches. Someparasites were attached to the middle part of the gill arch (Fig. 1a),but many were observed near the base of the gill filament, in theanterior region. The copepod body lay between the hemibranchswith its major axis parallel to the primary lamella axis (Fig. 1b).Second antennae extended to encircle (Fig. 1b) and increasinglycompress the gill filament, which appeared as a thin layer ofepithelium and connective tissue overlaying the cartilaginoussupporting bar. Most frequently, second antennae disrupted the gillfilament (not shown). Erosion, desquamation and necrosis ofsecondary lamellae were noticed near the site of Ergasilus sp.attachment (Fig. 1c). Branchial hyperplasia was observed on eitherside of the maxilla and near the copepod mouth. The copepodoccluded arteries and caused hemorrhage. Heavily infected gillswere more damaged than those of hosts with less Ergasilus sp.
Within the parenchyma of the primary lamellae of parasitizedgills, large numbers of mast cells (MCs) were seen near the para-site’s attachment site (Fig. 1c). Many MCs were also seen in the
Fig. 1. Gross lesion, wet mount and histological sections of Sparus aurata gills parasitized with Ergasilus sp. (a) Removal of the operculum shows the presence of several parasites(arrows) on the external surface of the gill hemibranchs. The copepods are attached mainly to the middle and basal part of gill filament, bar¼ 5 mm. (b) Wet mount of a gill filamentshowing the copepod with second antennae (arrows) encircling the gill filament, bar¼ 100 mm. (c) Histological section of gill infected with Ergasilus sp. Within the primary lamella,several mast cells (arrows) are visible; asterisk shows the copepod mouthpart, arrowheads indicate desquamation and necrosis of the gill tissue, bar¼ 20 mm. (d) Micrograph ofseveral mast cells (arrows) in vicinity of a capillary; note the eccentric nuclei and granules in the cytoplasm of these cells, bar¼ 10 mm.
B.S. Dezfuli et al. / Fish & Shellfish Immunology 30 (2011) 1087e1094 1089
Author's personal copy
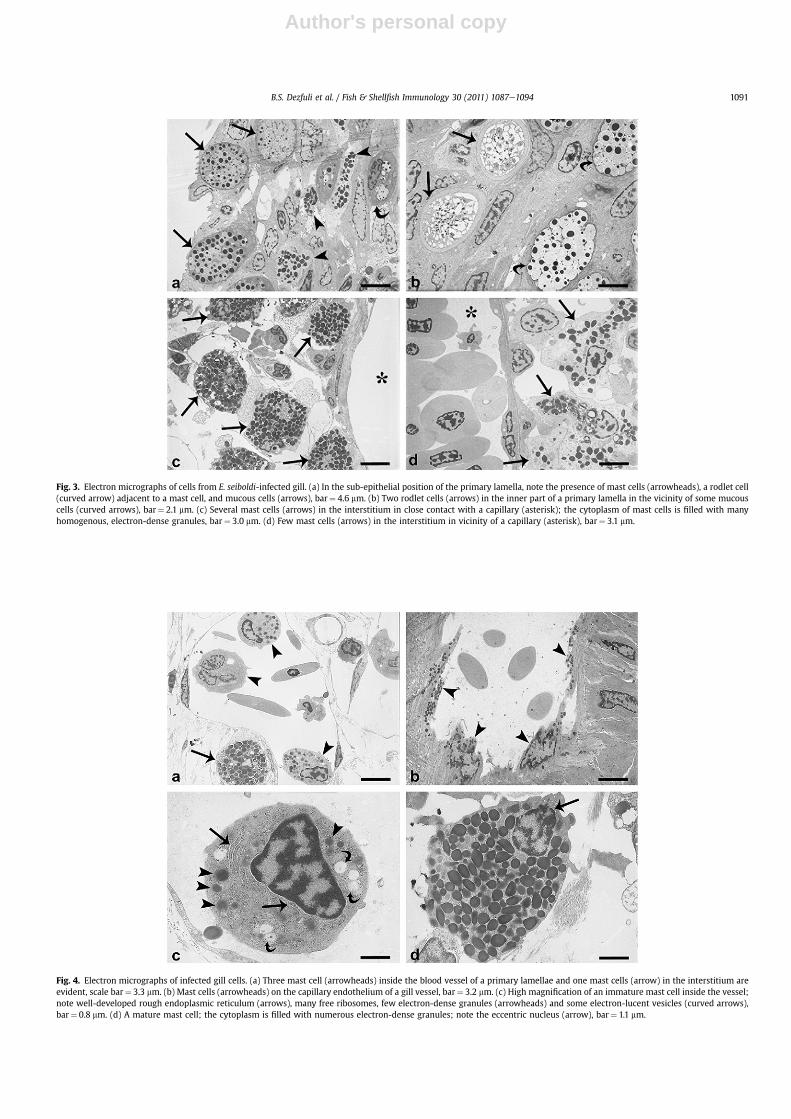
primary lamellae in close contact with a capillary, or in thesurrounding interstitium (Figs. 1c and d, 3c and d), with some MCsin the lumen of the vessel (Figs. 2f, 4a and c). In both parasitized anduninfected fish, the mast cells in the primary lamellae were basal tothe mucous cells (Fig. 3a) and rodlet cells. Mast cells (major axis11.10�1.96 mm, mean� SD, n¼ 150) were irregular in shape, withan eccentric, polar nucleus (Figs. 1d, 4a, c and d); their cytoplasmwas filled with numerous large, electron-dense, membrane-boundgranules (Figs. 1d, 4d and e).
Transmission electron microscopy revealed that the cytoplasmof MCs of the interstitium contained very few mitochondria, aninconspicuous Golgi apparatus and many homogeneous and elec-tron-dense granules (Figs. 3c and 4d). In contrast, all mast cellsinside blood vessels seemed to be immature and had well-devel-oped rough endoplasmic reticulum (Fig. 4c), many free ribosomes,several spherical-round mitochondria, low numbers of electron-dense granules (Fig. 4a and c) and some electron-lucid vesicles
(Fig. 4c). In the vast majority of infected gills, several mast cellswere inside the gill blood vessels on the capillary endothelium(Fig. 4b). In some vessels, the mast cells seemed to be detachingfrom the vessel wall (Fig. 4a and c) andmoving toward the center ofthe lumen. In several instances mast cells were present inside theinterstitium, in the vicinity of a capillary (Fig. 3d). However, MCdegranulation was not observed in either uninfected or infectedtissues.
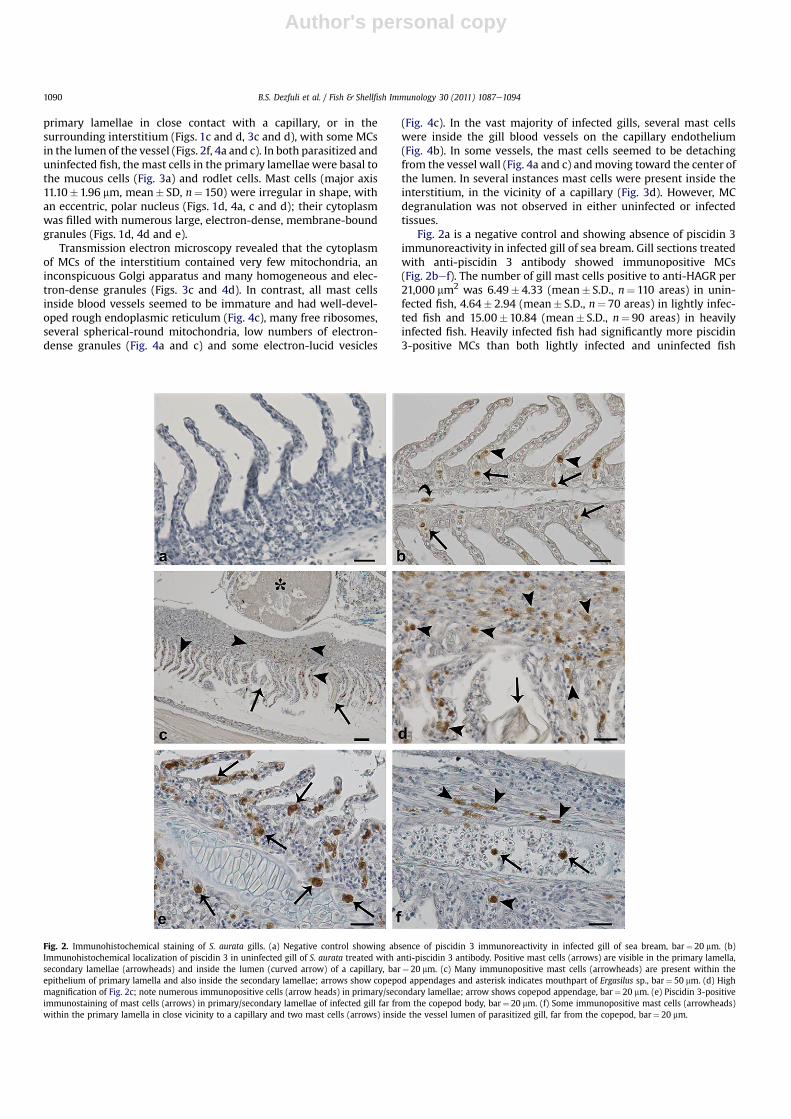
Fig. 2a is a negative control and showing absence of piscidin 3immunoreactivity in infected gill of sea bream. Gill sections treatedwith anti-piscidin 3 antibody showed immunopositive MCs(Fig. 2bef). The number of gill mast cells positive to anti-HAGR per21,000 mm2 was 6.49� 4.33 (mean� S.D., n¼ 110 areas) in unin-fected fish, 4.64� 2.94 (mean� S.D., n¼ 70 areas) in lightly infec-ted fish and 15.00�10.84 (mean� S.D., n¼ 90 areas) in heavilyinfected fish. Heavily infected fish had significantly more piscidin3-positive MCs than both lightly infected and uninfected fish
Fig. 2. Immunohistochemical staining of S. aurata gills. (a) Negative control showing absence of piscidin 3 immunoreactivity in infected gill of sea bream, bar¼ 20 mm. (b)Immunohistochemical localization of piscidin 3 in uninfected gill of S. aurata treated with anti-piscidin 3 antibody. Positive mast cells (arrows) are visible in the primary lamella,secondary lamellae (arrowheads) and inside the lumen (curved arrow) of a capillary, bar¼ 20 mm. (c) Many immunopositive mast cells (arrowheads) are present within theepithelium of primary lamella and also inside the secondary lamellae; arrows show copepod appendages and asterisk indicates mouthpart of Ergasilus sp., bar¼ 50 mm. (d) Highmagnification of Fig. 2c; note numerous immunopositive cells (arrow heads) in primary/secondary lamellae; arrow shows copepod appendage, bar¼ 20 mm. (e) Piscidin 3-positiveimmunostaining of mast cells (arrows) in primary/secondary lamellae of infected gill far from the copepod body, bar¼ 20 mm. (f) Some immunopositive mast cells (arrowheads)within the primary lamella in close vicinity to a capillary and two mast cells (arrows) inside the vessel lumen of parasitized gill, far from the copepod, bar¼ 20 mm.
B.S. Dezfuli et al. / Fish & Shellfish Immunology 30 (2011) 1087e10941090
Author's personal copy
Fig. 3. Electron micrographs of cells from E. seiboldi-infected gill. (a) In the sub-epithelial position of the primary lamella, note the presence of mast cells (arrowheads), a rodlet cell(curved arrow) adjacent to a mast cell, and mucous cells (arrows), bar¼ 4.6 mm. (b) Two rodlet cells (arrows) in the inner part of a primary lamella in the vicinity of some mucouscells (curved arrows), bar¼ 2.1 mm. (c) Several mast cells (arrows) in the interstitium in close contact with a capillary (asterisk); the cytoplasm of mast cells is filled with manyhomogenous, electron-dense granules, bar¼ 3.0 mm. (d) Few mast cells (arrows) in the interstitium in vicinity of a capillary (asterisk), bar¼ 3.1 mm.
Fig. 4. Electron micrographs of infected gill cells. (a) Three mast cell (arrowheads) inside the blood vessel of a primary lamellae and one mast cells (arrow) in the interstitium areevident, scale bar¼ 3.3 mm. (b) Mast cells (arrowheads) on the capillary endothelium of a gill vessel, bar¼ 3.2 mm. (c) High magnification of an immature mast cell inside the vessel;note well-developed rough endoplasmic reticulum (arrows), many free ribosomes, few electron-dense granules (arrowheads) and some electron-lucent vesicles (curved arrows),bar¼ 0.8 mm. (d) A mature mast cell; the cytoplasm is filled with numerous electron-dense granules; note the eccentric nucleus (arrow), bar¼ 1.1 mm.
B.S. Dezfuli et al. / Fish & Shellfish Immunology 30 (2011) 1087e1094 1091

Author's personal copy
(ANOVA, P< 0.05). But, there was no significant difference innumber of piscidin 3-positive MCs lightly infected fish versusuninfected fish (P> 0.05). Mast cells immunopositive for piscidin 3were observed both close to the copepod (Fig. 2c and d) and faraway from the parasite (Fig. 2e, f). In both uninfected and parasit-ized gills, piscidin 3-positive mast cells were observed in theprimary lamellae (Fig. 2bed), as well as in the apex, middle andbasal part of the secondary lamellae (Fig. 2d and e). Piscidin-posi-tive mast cells were also adjacent to capillaries and in someinstances, within them (Fig. 2b and f).
Rodlet cells were seen adjacent to mast cells and mucous cells.RCs were ovoid, had a basal nucleus, and a very distinct sub-plas-malemmar capsule (Fig. 3b). The cytoplasm contained severalrodlets. Rodlet cells were observed in sub-epithelial positions of theprimary lamellae (Fig. 3b), and rarely in secondary lamellae. In bothuninfected and parasitized gills, no piscidin 3 immunoreaction wasseen in RCs and mucous cells.
4. Discussion
Parasitic copepods naturally occur on wild fish. With regard tocultured fish, the action of these crustaceans may result indecreased flesh quality, mortality and a need for increased drugtreatment, adding as much as 6% to production costs [34e36].Previous studies have reported that the mode of attachment andfeeding activity of parasitic copepods were the main reasons ofdamage to the gills [6,7]. Indeed, it was mentioned that ergasilidcopepod infections can reduce gill function, resulting in decreasedgrowth rate due to impaired respiration, morbidity and oftensubstantial mortality in brackish water fish [11,13,14].
Mortality due to E. sieboldi has been reported in [37,38]. In ourstudy area, low mortality (w2%) of young S. aurata due to Ergasilussp. has been noticed in winter during the past few years (Dezfuliunpublished data). Nonetheless, in the semi-intensive farm wherewe sampled, since 2007, there has been an annual loss ofw5e8% ofthe juvenile stock of sea bream each summer, attributed to heavyinfections of Ergasilus sp.. At present, economic losses due to thiscopepod are primarily from the costs of treatments, implementedmanagement strategies, reduced growth rates and carcass down-grading at harvest.
With reference to the gills, this is a major route of pathogenentry after epithelial damage [39]. Attachment to the gill can resultin severe local damage such as inflammation, infiltration, andhyperplasia of rodlet cells, mucous cells and mast cells[16,30,40e43]. Herein, in gill of sea bream parasitized with Erga-silus sp., some RCs were encountered. The same finding has beenreported in infected gill of A. brama [17] and in parasitized gills ofD. labrax [24]. Rodlet cells are exclusive to fish and represent aninflammatory cell type closely linked to other piscine inflammatorycells [44e47]. There are many studies that consistently report anassociation between RC proliferation and the presence of a varietyof parasites [45e49].
In our current study in S. aurata, fish heavily infected withErgasilus sp. had significantly more piscidin 3-positive mast cellsthan either lightly infected or uninfected fish. These findings aresomewhat similar to that of Andrews et al. [16], who reported thatmast cells (MCs) were absent in healthy gills of L. lineata, whilenumerous MCs, many of which were piscidin-3 positive, were seenin gills infected with the copepod Chondracanthus goldsmidi.However, in our previous study on D. labrax, we noticed that whilethere was no significant difference in the number of mast cellspositive for anti-piscidin 3 antibody in gills parasitized witha monogenean compared to uninfected gills, the intensity of theantibody reaction strongly suggested that there was greaterexpression of piscidin 3 in the MCs of infected fish [27]. Thus, the
cumulative evidence suggests that piscidin-expressing MCs mayplay an important role in defending against this important group ofparasites, but the method in which this expression occurs varieswith either the specific host, parasite, or both.
Innate immunity of vertebrates relies on different type of cells,and one of the most common is mast cells [50]. Mast cells arepresent in most species of teleost and in a variety of tissues (e.g.,alimentary canal, gills and skin) [51,52]. Mast cells are motile[19,53] and have been recognized in regions of active inflammatoryresponse especially due to bacterial and parasitic infections[25,54,55]. Gill is one of the tissues first exposed to environmentalchallenges and pathogens, so an ability to mount an immunedefense is crucial [16,51]. In S. aurata, MCswere documented inside,and/or in close proximity to capillaries and they were observedthroughout the loose connective tissue of the gill arch. Withreference to the presence of mast cells in loose connective tissuesome authors have suggested a resident population for these cells[51,56].
In all vertebrates, mast cells likely are strategically positioned atperivascular sites to regulate inflammatory responses [57]. Theclose association of mast cells with the capillary endothelial cellssuggests that they may migrate across the endothelium [51,58].Furthermore, mast cells migrate within the gill microcirculation,which may facilitate rapid movement throughout the gill tissue[58]. These observations support the suggestion that the gill isprimed with the innate immune system, enabling an immediateresponse to pathogenic or parasitic organisms [59,60]. Severalrecords have documented degranulation of mast cells in responseto exposure to a variety of known degranulating agents and path-ogens with release of their contents [25,51e53,58]. However, ingills of S. aurata, we never observed evidence of degranulation inmast cells.
The functional similarity between mammalian and fish mastcells has stimulated studies into identifying the contents of gran-ules in fish MCs. According to Murray et al. [51], mast cells could bedirectly involved in the destruction of pathogens and they providefurther evidence for their multifunctional role in teleosts. Investi-gations have shown that mast cells of fish intestine and gills areinvolved in the production of specific AMPs, namely pleurocidin[51] and piscidins [21,23,30,33,61]. Piscidins have potent, broad-spectrum antifungal and antibacterial activity and recently beenfound to have strong antiparasitic activity andwere reported in gillsof fish infected with three protistan ectoparasites [21], gills para-sitized with a copepod [16], and gills infected with a monogenean[27].
The occurrence of antimicrobial peptide piscidin 3 in site ofErgasilus sp. infection might be a combined immune responseresulting from both the copepod’s attachment and feeding activi-ties and the threat of pathogens entering through the disruption ofthe gill lamella. The same assumption was adopted by Andrewset al. [16] on piscidin 3 expression in mast cells of gill of the stripedtrumpeter infected with a copepod. Our data are in agreement withthe suggestion that piscidins play an important role in the non-specific immune defense of many teleosts [33].
Acknowledgments
We are indebt to Samantha Squerzanti, Silvia Fabbri, and AndreaMargutti from the University of Ferrara for technical assistance.This study was supported by grants from the Italian Ministry of theUniversity and Scientific Research and Technology.
Conflict of interestThe authors declare that there is no conflict of interest.
B.S. Dezfuli et al. / Fish & Shellfish Immunology 30 (2011) 1087e10941092

Author's personal copy
Appendix. Supplementary data
Supplementary data associated with this article can be found inthe online version, at doi:10.1016/j.fsi.2011.02.005.
References
[1] Costello MJ. Ecology of sea lice parasitic on farmed and wild fish. TrendsParasitol 2006;22:475e83.
[2] Bravo S. Sea lice in Chilean salmon farms. Bull Eur Ass Fish Path 2003;23:197e200.
[3] Guo FC, Woo PTK. Selected parasitosis in cultured and wild fish. Vet Parasitol2009;163:207e16.
[4] Johnson SC, Treasurer JW, Bravo S, NagasawaK, Kabata Z. A review of the impactof parasitic copepods on marine aquaculture. Zool Stud 2004;43:229e43.
[5] Muzzal PM, Hudson PL. Occurrence of Ergasilus megaceros Wilson, 1916, inthe sea lamprey and other fishes from North America. J Parasitol 2004;90:184e5.
[6] Jónsdóttir H, Bron JE, Wootten R, Turnbull JF. The histopathology associatedwith the pre-adult and adult stages of Lepeophtherius salmonis on the Atlanticsalmon, Salmo salar L. J Fish Dis 1992;15:521e7.
[7] Nolan DT, Ruane NM, van der Heijden Y, Quabius ES, Costelloe J. WendelaarBonga SE. Juvenile Lepeophtherius salmonis (Kroyer) affect the skin and gills ofrainbow trout Oncorhynchus mykiss (Walbaum) and the host response toa handling procedure. Aquac Res 2000;31:823e33.
[8] Paperna I. Parasites and disease of the grey mullet (Mugilidae) with specialreference to the seas of the near east. Aquaculture 1975;16:173e5.
[9] Lin CL, Ho JS. Two species of Caligidae (Copepoda) parasitic on cultured inTaiwan. J Nat His 1998;31:1483e500.
[10] Yamashita K. Parasitic copepoda (Ergasilus sp.) found on the epidermis oflarval fish of red sea bream (Pagrus major). Fish Pathol 1980;15:91e4.
[11] Wang GT, Li WW, Yao WJ, Nie P. Mortalities induced by the copepod Siner-gasilus polycolpus in farmed silver and bighead carp in a reservoir. Dis AquatOrg 2002;48:237e9.
[12] Benetti DD, Leingang AJ, Russo R, Powell TM, Cleary D, Grabe SW, et al.Development of aquaculture methods for southern flounder, Paralichthyslethostigma: II. Nursery abd Grow-out. J Appl Aquac 2001;11:135e46.
[13] Tsotetsi AM, Avenant-Oldewage A, Mashego SN. Aspects of the pathology ofLamproglena clariae (Copepoda: Learnaeidae) of gills of Clarias gariepinus fromthe Vaal River system, South Africa. Afr Zool 2005;40:169e78.
[14] Molnar K, Székely C. Occurrence and pathology of Sinergasilus lieni (Cope-poda: Ergasilidae), a parasite of the silver carp bighead, in Hungarian ponds.Acta Vet Hung 2004;52:51e60.
[15] Alston S, Lewis JW. The Ergasilid parasites (Copepoda: Poecilostomatoida) ofBritish freshwater fish. In: Pike AW, Lewis JW, editors. Parasitic of disease offish. Dyfed: Samara Publishing Limited; 1994. p. 171e88.
[16] Andrews M, Battaglene S, Cobcroft J, Adams M, Noga E, Nowak B. Hostresponse to the chondracanthid copepod Chondracanthus goldsmidi, a gillparasite of the striped trumpeter, Latris lineata (Forster), in Tasmania. J FishDis 2010;33:211e20.
[17] Dezfuli BS, Giari L, Konecny R, Jaeger P, Manera M. Immunohistochemistry,ultrastructure and pathology of gills of Abramis brama (L.) from lake Mondsee(Austria) due to Ergasilus sieboldi (Copepoda). Dis Aquat Org 2003;53:257e62.
[18] Manera M, Dezfuli BS. Rodlet cells in teleosts: a new insight into their natureand functions. J Fish Biol 2004;65:597e619.
[19] Reite OB, Evensen Ø. Inflammatory cells of teleostean fish: a review focusingon mast cells/eosinophilic granule cells and rodlet cells. Fish ShellfishImmunol 2006;20:192e208.
[20] Beutler B. Innate immunity: an overview. Mol Immunol 2004;40:845e59.[21] Colorni A, Ullal A, Heinisch G, Noga EJ. Activity of the antimicrobial poly-
peptide piscidin 2 against fish ectoparasites. J Fish Dis 2008;31:423e32.[22] Jia X, Patrzykat A, Devlin RH, Ackerman PA, Iwama GK, Hancock RE. Antimi-
crobial peptides protect Coho salmon from Vibrio anguillarum infections. ApplEnviron Microbiol 2000;66:1928e32.
[23] Silphaduang U, Noga E. Peptide antibiotics in mast cells of fish. Nature2001;414:268e9.
[24] Dezfuli BS, Giari L, Simoni E, Menegatti R, Shinn AP, Manera M. Gill histopa-thology of cultured European sea bass, Dicentrarchus labrax (L.), infected withDiplectanum aequans (Wagener 1857) Diesing 1958 (Diplectanidae: Mono-genea). Parasitol Res 2007;100:707e13.
[25] Dezfuli BS, Giovinazzo G, Lui A, Giari L. Inflammatory response to Dentitruncustruttae (Acanthocephala) in the intestine of brown trout. Fish ShellfishImmunol 2008;24:724e33.
[26] Dezfuli BS, Lui A, Giovinazzo G, Boldrini P, Giari L. Intestinal inflammatoryresponse of powan Coregonus lavaretus (Pisces) to the presence of acantho-cephalan infections. Parasitology 2009;136:929e37.
[27] Dezfuli BS, Pironi F, Giari L, Noga EJ. Immunocytochemical localizationof piscidinin mast cells of infected seabass gill. Fish Shellfish Immunol 2010;28:476e82.
[28] Zasloff M. Antimicrobial peptides of multicellular organisms. Nature 2002;415:389e95.
[29] Corrales J, Gordon WL, Noga EJ. Development of an ELISA for quantification ofthe antimicrobial peptide piscidin 4 and its application to assess stress in fish.Fish Shellfish Immunol 2009;27:154e63.
[30] Corrales J, Mulero I, Mulero V, Noga EJ. Detection of antimicrobial peptidesrelated to piscidin 4 in important aquacultured fish. Dev Comp Immunol2010;34:331e43.
[31] Mulero I, Noga EJ, Meseguer J, Garcìa-Ayala A, Mulero V. The antimicrobialpeptides piscidins are stored in the granules of professional phagocyticgranulocytes of fish and are delivered to the bacteria-containing phagosomeupon phagocytosis. Dev Comp Immunol 2008;32:1531e8.
[32] Campagna S, Saint N, Molle G, Aumelas A. Structure and mechanism of actionof the antimicrobial peptide piscidin. Biochemistry 2007;46:1771e8.
[33] Silphaduang U, Colorni A, Noga EJ. Evidence for widespread distribution ofpiscidin antimicrobial peptides in teleost fish. Dis Aquat Org 2006;72:241e52.
[34] Costello MJ. The global economic cost of sea lice to the salmonid farmingindustry. J Fish Dis 2009;32:115e8.
[35] Heuch PA, Bjørn PA, Finstad B, Holst JC, Asplin L, Nilsen F. A review of theNorwegian ‘National Action Plan Against Salmon Lice on Salmonids’: theeffect on wild salmonids. Aquaculture 2005;246:79e92.
[36] Rae GH. Sea louse control in Scotland, past and present. Pest Manag Sci2002;58:515e20.
[37] Fryer G. The parasitic Copepoda and Branchiura of British freshwater fishes,a hand book and key. Freshwater Biological Association; 1982. p. 87.
[38] Schäperclaus W. Fish diseases, vols. 1 & 2. Berlin, Germany: Akademie-Verlag;1986. p. 1398.
[39] Palzenberger M, Pohla H. Gill surface area of water-breathing freshwater fish.Rev Fish Biol Fish 1992;2:187e216.
[40] Bennet SM, Bennet MB. Pathology of attachment and vascular damage asso-ciated with larval stages of Dissonus manteri Kabata, 1966 (Copepoda: Dis-sonidae) on the gills of coral trout, Plectropomus leopardus (Lacépède)(Serranidae). J Fish Dis 1994;17:447e60.
[41] Bennet SM, Bennet MB. Gill pathology caused by infestation of adult andpreadult Dissonus manteri Kabata (Copepoda: Dissonidae) on coral trout,Plectropomus leopardus (Lacépède), (Serranidae). J Fish Dis 2001;24:523e33.
[42] Covello JM, Bird S, Morrison RN, Battaglene SC, Secombes CJ, Nowak BF. Cloninoand expression analysis of three striped trumpeter (Latris lineata) pro-imflam-matory cytokines, TNF-a, IL-1b and IL 8, in response to infection by the ecto-parasitic, Chondracathus goldsmidi. Fish Shellfish Immunol 2009;26:773e86.
[43] Roubal FR. Extent of gill pathology in the toadfish Tetractenos hamiltoni causedby Naobranchia variavilis (Copepoda: Naobranchiidae). Dis Aquat Org1999;35:203e11.
[44] Dezfuli BS, Simoni E, Rossi R, Manera M. Rodlet cells and other inflammatorycells of Phoxinus phoxinus infected with Raphidascaris acus (Nematoda). DisAquat Org 2000;43:61e9.
[45] Matisz CE, Goater CP, Bray D. Density and maturation of rodlet cells in braintissue of fathead minnows (Pimephales promelas) exposed to trematodecercariae. Int J Parasitol 2010;40:307e12.
[46] Reite OB. The rodlet cells of teleostean fish: their potential role in host defencein relation to the role of mast cells/eosinophilic granule cells. Fish ShellfishImmunol 2005;19:253e67.
[47] Vigliano FA, Bermúdez R, Nieto JM, Quiroga MI. Development of rodlet cells inthe gut of turbot (Psetta maxima L.): relationship between their morphologyand S100 protein immunoreactivity. Fish Shellfish Immunol 2009;26:146e53.
[48] Dezfuli BS, Giari L, Shinn AP. The role of rodlet cells in the inflammatoryresponse in Phoxinus phoxinus brains infected with Diplostomum. Fish Shell-fish Immunol 2007;23:300e4.
[49] Leino RL. The effects of periodic acid-silver methenamine staining andprotease digestion on the secretory granules of rodlet cells. In: Bailey GW,editor. Thirty-seventh annual meeting of the electron microscopy society ofAmerica, Baton Rouge, LA; 1979. p. 310e1.
[50] Murray HM, Gallant JW, Douglas SE. Cellular localization of pleurocidin geneexpression and synthesis in winter flounder gill using immunohistochemistryand in situ hybridization. Cell Tissue Res 2003;312:197e202.
[51] Murray HM, Leggiadro CT, Douglas SE. Immunocytochemical localization ofpleurocidin to the cytoplasmic granules of eosinophilic granular cells from thewinter flounder gill. J Fish Biol 2007;70:336e45.
[52] Ellis AE. Eosinophilic granular cells (EGC) and histamine responses to Aero-monas salmonicida toxins in rainbow trout. Dev Comp Immunol 1985;9:251e60.
[53] Vallejo AN, Ellis AE. Ultrastructural study of the response of eosinophilic granulecells to Aeromonas salmonicida extracellular products and histamine liberators inrainbowtrout, SalmogairdneriRichardson.DevComp Immunol1989;13:133e48.
[54] Reimschuessel R, Bennett RO, May EB, Lipsky MM. Eosinophilic granular cellresponse to a microsporidian infection in a sergeant major fish, Abudefdufsaxatilis L. J Fish Dis 1987;10:319e22.
[55] Sharp GJE, Pike AW, Secombes CJ. The immune response of wild rainbow troutSalmo gairdneri Richardson to naturally acquired plerocercoid of Diphyllo-bothrium dendriticun (Nitzsch 1824) and D. ditremun (Creplin 1825). J Fish Biol1989;35:781e94.
[56] Noya M, Lamas J. Response of eosinophilic granule cells of gilthead seabream(Sparus aurata, Teleostei) to bacteria and bacteria product. Cell Tissue Res1997;287:223e30.
[57] Mekori YA. The mastocyte: the “other” inflammatory cell in immunopatho-genesis. J Allergy Clin Immunol 2004;114:52e7.
B.S. Dezfuli et al. / Fish & Shellfish Immunology 30 (2011) 1087e1094 1093

Author's personal copy
[58] PowellMD,WrightGM, Burka JF. Eosinophilic granule cells in thegills of rainbowtrout, Oncorhynchus mykiss: evidence of migration? J Fish Biol 1990;37:495e7.
[59] Campos-Perez JJ, Ward M, Grabowski PS, Ellis AE, Secombes CJ. The gills are animportant site of iNOS expression in rainbow trout Oncorhynchus mykiss afterchallenge with the Gram-positive pathogen Renibacterium salmoninarum.Immunology 2000;99:153e61.
[60] Flaño E, Lopez-Fierro P, Razquin BE, Villena A. In vitro differentiationof eosinophilic granular cells in Renibacterium salmoninarum-infected gillcultures from rainbow trout. Fish Shellfish Immunol 1996;6:173e84.
[61] Noga EJ, Silphaduang U. Piscidins: a novel family of peptide antibiotics fromfish. Drug News Perspect 2003;16:87e92.
B.S. Dezfuli et al. / Fish & Shellfish Immunology 30 (2011) 1087e10941094




















