
Evangelist Charles S. Price - Flower Pentecostal Heritage ...
Upload
independentCategory
view
2download
0
FHYSIOLOGIA PLANTARUM 82: 33^338. Copenbagen 1991
Flower formation in vitro in a quantitative short-day tobacco:interrelation between photoperiod and infructescence development
Maria Maddalena Altamura, Gabriella Pasqua, Barbara Motiacelli, Marisa Tomassi, Tiziana Archilletti andGiuseppina Falasca
Altamura, M.M., Pasqua, G., Monacelli, B.. Tomassi, M., Archilletti, T. andFalasca, G. 1991. Flower formation in vitro in a quantitative short-day tobacco:interrelation between photoperiod and infructescence development. - Physiol. Plant.82: 333-338.
The flowering response of thin layers excised from branch internodes of Nicotianatabacum cv. Maryland Catterton (quantitative short-day plant for induction) wasstudied under three photoperiodic treattnents. The explants were excised from in-florescences bearing flowers only, flowers and green fruits, or from infructescenceswith green fruits only. The aim of the study was to investigate the post-inductivephotoperiodic effects on in vitro flower bud formation in a quantitative short-daytobacco and the relation with infructescence development. Short days quantitativelyenhanced the flower bud regeneration capacities of explants in all stages of develop-ment, both as number of explants induced to produce flowers and as mean number offlowers per explant. There was no significant difference in flower bud formation onexplants of the first two stages, which produced much more flowers than those of thethird stage. Observations in planta showed that, during the 20 days separating thesecond stage from the first stage, there was no significant difference in the number offloral buds and flowers present on the inflorescence; however, the branch internodeslengthened, as did the floral buds and flowers. During the iO days leading to the thirdstage, the number of capsules did not change significantly, but a high rate of floralabscission occurred. The present results show that in Nicotiana tabacum cv. MarylandCatlerton short day quantitatively controls not only the inductive step of the flower-ing process, but also affects the capacity to regenerate flower buds during the latepost-inductive phases. The responsiveness to the photoperiodic signal decreases onlywhen the plant exhibits oniy fruits.
Key words - Flowering in vitro, infructescence development, Nicotiana tabacum,photoperiod, tobacco.
M.M. Altamura (corresponding author) et al., Dipartimento di Biologia Vegetate,Univ. "La Sapienza", P. le A. Moro, 5, I-O01S5 Rome, Italy,
the flowering process, studies on flower formation inintroduction vitro were carried otjt with several cultivars of tobacco.The habit of a fiowering platit, the development of using thiti layer expiants {Monacelli et al, 1984, Rajee-flowers, and fruit set and retentioti are markedly in- van and Lang 1987, Altamura et al, 1989). Thin layers,fluenced not only by short- and long-distance interac- consisting ofepidennal,subepidertnat and cortical cells,tions among the plant's organs (Trehame et al, 1985), are capable of initiating and developing flowers whenbut also by environmental factors (e,g, photoperiod). In excised from the floral regions of the piant and culturedspecies with post-inductive photoperiodic requirements, on a suitable medium (Kamate et al, 1981).unfavourable photoperiods induce varied reproductive In a day-neutral cultivar of tobacco {Nicotiarm taba-abnormalities (ICinet et al, 1985) and reversion of cum cv, Samsun), floral neoformation from thin layersflowers to vegetative growth (Krishnamoorthy and is quantitatively affected by the physiological stage ofNatida 1968). In an attempt to determine the photope- the donor plant (Pasqua et al, 1990). Furthermore, theriodic requirements during the post-inductive phases of order of branching on the tobacco inflorescence is an-
Received 13 December, 1990; revised 25 March, 1991
22' Physiol. Plant. 82, lWl
other important factor in the aptitude of the thin layersfor flower regeneration. In a short day tobacco {Nico-tiana tahacum cv. White Burley), the flowering and thevegetative response of branch internode explants areintermediate between the response of explants takenfrom the pedicel (the best excision site for flower budregeneration) and those taken from the rachis (the bestsite for caulogenesis) (Altamura et al. 1989).
Caulogenesis is enhanced under phototreatments un-favourable for flowering (Cousson and Tran Thanh Van1983, Altamura et al. 1986, 1989, Rajeevan and Lang1987), In the present study, thin layers from branchinternodes were used, because the organogenic re-sponse of these explanls may be directed toward eitherflower bud formation or caulogenesis, depending on thephotoperiod. The organogenic response of the thinlayers was examined under three photoperiodic treat-ments. The explants were excised from plants of Nico-tiana tabacum cv. Maryland Catterton (quantitativeshort day for induction) at different reproductive stages.The aim was to determine whether there is a post-inductive photoperiodic requirement in this genotypewith respect to flower bud regeneration in vitro in rela-tion to infructescence development.
Abbreviations - A stage, plants with inflorescences with flora!buds and flowers; B stage, plants with inflorescences withflowers and green fruits; C stage, plants with infructescenceswith alt green fruits; IB, branch internode; Veg, vegetative.
Materials and methods
Nicotiana tabacum cv. Maryland Catterton plants weregrown in pots, in the Botanical Garden of the Uni-versity "La Sapienza" of Rome. They were cultured in agreenhouse to the rosette stage (Goodspeed 1954), andthen transferred to open air. The seeds were kindlyprovided by the Istituto Sperimentale dei Tabacchi, Sca-fati, Italy.
The morphological characters of the inflorescenceand the infructescence were examined in two successiveyears, without relevant differences in the results (dataof the second year shown).
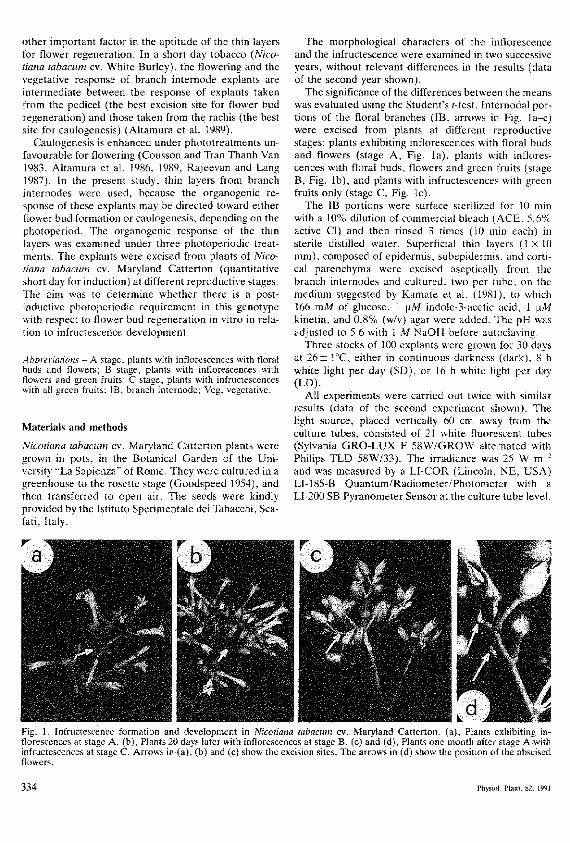
The significance of the differences between the meanswas evaluated using the Student's f-test. Intertiodal por-tions of the floral branches (IB, arrows in Fig. la-c)were excised from plants at different reproductivestages; plants exhibiting inflorescences with floral budsand flowers (stage A, Fig. la), plants with inflores-cences with floral buds, flowers and green fruits (stageB, Fig. lb), and plants with infructescences with greenfruits only (stage C, Fig. lc).
The IB portions were surface sterilized for 10 minwith a 10% dilution of commercial bleach (ACE, 5.6%active Cl) and then rinsed 3 times (10 min each) insterile distilled water. Superficial thin layers (1 x 10mm), composed of epidermis, subepidermis, and corti-cal parenchyma were excised aseptically from thebranch internodes and cultured, two per tube, on themedium suggested by Kamate et al. (1981), to which166 mM of glucose, 1 \iM indole-3-acetic acid, 1 |iiWkinetin, and 0,8% (w/v) agar were added. The pH wasadjusted to 5.6 with 1 M NaOH before autoclaving.
Three stocks of 100 explants were grown for 30 daysat 26 ± 1°C, cither in continuous darkness (dark), 8 hwhite light per day (SD), or 16 h white light per day(LD).
All experiments were carried out twice with similarresults (data of the second experiment showti). Thelight source, placed vertically 60 cm away from theculture tubes, consisted of 21 white fluorescent tubes(Sylvania GRO-LUX F 58W/GROW alternated withPhilips TLD 58W/33). The irradiance was 25 W m'-and was measured by a LI-COR (Lincoln, NE, USA)LI-185-B Quantum/Radiometer/Photometer with aLI-200 SB Pyranometer Sensor at the culture tube level.
Fig. 1. Infructescetice formation and developrrtent in Nicotiana tabacum cv. Maryland Catterton. (a). Plant* , \hi l ' ' i i ig in-florescences at stage A. (b). Plants 20 days later with inflorescences at stage B. (c) and (d). Plants one month after stage A withinfructescences at stage C. Arrows it̂ (a), (b) and (c) show the excision sites. The arrows in (d) show the position of the abscisedflowers.
334 Physiol. Planl. 82. 1991

Tab, 1. ReSationship among the tnain characters of inflorescences at stages A and B and infructescences at stage C. The values areexpressed as mean lengths and diameters (in mm ± SE) or mean number (± SE) of capsules, flowers and floral buds.
Character
Length of capsule pedicelLength of flower pedicelLength of floral bud pedicelDiameter of capsule pedicelDiameter of flower pedicelDiameter of floral bud pedicelLength of branch internodeDiameter of branch internodeCapsule lengthCapsule diameterFlower lengthLength of flower calyxLength of flower corollaDiameter of flower calyxDiameter of flower corollaFloral bud lengthLength of floral bud calyxLength of floral bud corollaDiatneter of flora) bud calyxCapsule numberFlower numberFloral bud number
A
_
8.0±0.23.6±0.2
-1.7±0.0l.l±0.06.2±0.42.1±0.1
-—
45.8±0.516.9±0.332.6±0.57.5±0.1
n,4±0.921.0±1.314.8+0.38.3±L46.4±0.20.0±0.08,0±0.9
15.6+2.9
Developmental stage
B
12.6±0.38.1 ±0.34,3±0.31.9±0,01.6+0,01.2±0.09.7±0.82.3±0.0
12.2±0.56.9±0,2
47.4±0,519.1 ±0.234.8±0.58.6±a.l
17.8±1.027.5±2.316.4±0.315.3±2.47.3±0.2
33.3±2.311.0±l.l8.7±1.2
C
13.1±O,3——
2,0±0.0—-
9.7±0.92.3±0.0
14.4±0.57.9±0.2
--—------
37.0±1.40.0±0.00,0±0.0
Results
Observations in jrianta
We examined the morphological characters of both theterminal inflorescence (stages A and B, Fig, la-b) andthe terminal infructescence (stage C, Fig, lc) of Nico-tiana tabacum cv, Maryland Catterton plants. Onehundred measurements per parameter were randomlycarried out on three stocks of 10 plants each. The pa-rameters considered are shown in Tab, 1, During theapproximately 20 days separating stage A from B, thetotal length of the flowers changed with a low level ofsignificance (P<0,05) . However, at stage B, both thelength and the diameter of the flowers' calyces in-creased significantly {P < 0.01), as did the length of thecorollas (P<0.01) . The floral buds of the stage B in-florescences were longer (P < 0.05) than those at stageA (Tab. 1), At stage B, the length and the diameter ofthe branch internodes increased significantly( f < 0 , 0 1 ) , with respect to the previous stage. In con-trast, both the size of the pedicels attached to flowersand the size of those attached to the floral buds did notchange significantly (Tab, 1), Furthermore, from stageA to B, the number of floral buds did not decreasesignificantly, and the number of flowers did not increasesignificantly (Tab, 1), At stage B, the fruits were visible(Fig. lb), and had noticeably longer and thickerpedicels (P < 0,01) in comparison to the flowers, whosepedicels were longer (P<0,01) than those near thefloral buds (Tab, 1), An additional 10 days were neces-sary for the inflorescence to become an infructescencewith all green fruits (Fig. lc). The size of the branch
Physiol. Plant. 82, 1991
internodes in this new stage did not change (C, Tab, 1);however, both the length and the diameter of the cap-sules increased significantly (P<0.01) . The attachedpedicels, in comparison to those near the capsules ob-served at stage B, thickened significantly (P<0,01) ,without significant increase in length (Tab, 1). At thisstage, fruit pedicels were slightly incurved, and the re-gion of insertion on the branch node thickened greatly(Fig. Id). There was no significant difference in thenumber of capsules between B and C stages (Tab, 1),This was due to the high rate of abscission of floral budsand flowers (arrows in Fig. Id), which occurred only inthe interval between stages B and C. These results showthat several of the morphological characters of the in-florescence change during the month between its fullflowering and its transformation into an infructescence.
Analysis of tlie response in vitro
Qualitative analysisAt the end of the culture period of 30 days the morpho-genic response of IB expiants was examined with adissecting microscope.
Floral neoformation (Fig, 2a-h), caulogenesis (Fig,2i), and callogenesis (associated or not with organ for-mation) were observed in all stages and for all treat-ments. The time course of morphogenesis showed that,in both stages A and B , floral neoformation was preco-cious. That is, the first floral domes were observed at7-8 days in culture, regardless of the treatment; butonly SD treatment resulted in a high percentage ofexplants with flowers (50% of expiants in stage A and
335

\-i'^. 2 r I oral neoformation, caulogenesis and callogenesis on IB explants at the end of the culture period. Bars — 0.5 mm. (a),Siit!;li.- flowers with sepal and petal primordja (SD, stage A), (b), Explants with single flowers with fully developed anthers plusl:;:lli!̂ I'tiark stage C). (c). Regular flowers at foil developroent (ca, calyx; co, corolla; and a, androeciutn are shown). SD, stage B.(J). .Sessile flowers on callus (dark, stage A), (e). Single flowers with pedicel elongation (SD, stage A), (f). Inflorescences newlyformed uttder SD (stage A), (g). Anomalous flower with only one anther. Perianth and anther tnetamorphosed by callus (dark,stage C). (h). Anomalous flower with an increased number of anthers and carpels. Carpels onl> partially fused (dark, stage B). (i).Vegetative buds on C explants (dark).
60% of explants in stage B), In contrast, the explantsfrom stage C showed the first floral domes between day10 and day 13, and the number of explants with flowersincreased slowly during the rest of the culture period inall treatments.
At the end of the culture, flowers were observed incombination or not (Fig, 2a-c) with vegetative buds onthe same explant. Frequently flowers and/or vegetativebuds were observed on explants showing also callus(Fig. 2b,d,e,i). The most abundant callus formation.
336 Physiol. Plant. 82. 1991

Tab. 2. Effect of photoperiod on the percentage of explantswith flowers, vegetative buds only, callus only or with noresponse in vitro. The numbers within brackets show the per-centages of explants with flowers only, of the total number offlowering explants.
Developmental
ABC
ABC
ABC
ABC
Phototreatment
Dark SD
605529
148
12
161712
102047
Flowers(42) 80 (71)(31) 86 (62)(34) 48 (43)
Vegetative buds only000
Callus only1245
No response8
1047
LD
65 (51)61 (34)25 (36)
75
11
15105
132459
per explant, was observed in darkness. Under all treat-ments and for all stages we observed, on the sameexplant, single flowers at different degrees of develop-ment (i.e from the stage of floral dome with sepal andpetal primordia [Fig, 2a] to the stage of flowers with fivedeveloped anthers and two fusing carpels [Fig. 2b,c]).Pedicel elongation occurred only on flowers of the Aand B explants, with the most favourable treatmentbeing SD (Fig. 2e). Under all treatments, A explantsdemonstrated the capacity to form de novo inflores-cences (Fig. 2f); this occurred frequently only under SD(not shown). Furthermore, SD was the only treatmentunder which inflorescences developed on B explants; noinflorescence was ever found in C explants.
Under darkness, and occasionally under LD, with norelevant difference among the stages, some floral anom-alies were found (i.e, partial or total substitution ofsome whorls by caJltis; decreased or increased numberof anthers, abnormal [Fig. 2g] or normal; and increasednumber and only partial fusion of carpels [Fig. 2h]).
Quantitative analysisStage A and B explants were much more productivethan stage C expiants (Tab. 2); the flowering responseof A and B explants was better than that of C explants,regardless of the photoperiodic treatment (Tabs 2-3),SD strongly enhanced the capacity to flower in vitro inexplants of all stages (Tab. 2). An enhanced floweringresponse in tobacco thin layers is associated with thereduction or the repression of vegetative budding (Al-tamura et ai. 1989). Consequently, the promotive effectof SD on the flower bud formation concurs with theabsence of vegetative bud formation on A and B as wellas C explants (Tab. 2). Furthermore, there was a greaternumber of explants with only flowers under SD, thanunder darkness or LD; however, under SD, the numberof A and B explants with only flowers was higher thanthat of C explants (Tab. 2, numbers in brackets). Re-garding the mean number of flowers per expiant, therewas no significant difference between the responses ofA and B explants, under the same treatment (Tab, 3),In explants of both stages, the number of flowers pro-duced was significantly (P < 0.01) greater than the num-ber of vegetative buds. Both the greatest number{P < 0,01) of flowers and the least number (P < 0,01) ofvegetative buds were produced under SD (Tab. 3), Cexplants produced less flowers than both A explants(/'<0,05) and B explants (P<0.01 for SD, /><0,05for dark), and there was no significant difference be-tween the flowering and the vegetative response of Cexplants, in all treatments (Tab. 3). Nevertheless, thenumber of flowers produced was greater under SD thanunder the other treatments, which showed no significantdifference between them (Tab, 3, "Flowers", coiumnC).
Discussion
SD enhanced flower neoformation in thin layers of fSo-ral branch internodes of Nicotiana tabacum cv, Mary-land Catterton plants (Tabs 2-3), The photomorpho-genic response was quantitative (Tabs 2-3), and theresponsiveness of the explant to the photoperiodic sig-nal varied according to the stage of infructescence de-velopment on the donor plant, Croes et al, (1985) foundthat tissue aging negatively affects tobacco thin layer'sability to regenerate flower buds, Croes et al, (1986)found that the aging process is induced by fruit forma-
Tab. 3. Effect of photoperiod on the floral and vegetative productivity of branch internode explants excised from inflorescences atstages A and B and from infructescences at stage C. The productivity is evaluated as the average number of flowers or vegetativebuds per explant (± SE). Inflorescences are evaluated as single flowers.
Treatment
DarkSDLD
A
10.4±1.719.7±2.O10.0±0.6
Flowers
B
8.0±0.819.3±L210.9±l.l
C
5.0±0.5U.0±1.97.0±].l
A
5.7±1.12.0±0.24.1 ±0.5
Vegetative buds
B
7,6±0.85.0±0,48.0±0.8
C
7.0±l,26.0±L06.3+2.S
Physiol. PUrn. 82, 1991 337

tion. During the 20 day interval separating stage A fromstage B, fruit setting occurred; however, contrary toCroes et al,, a fruit-induced aging seemed not to affectthe flower bud regenerating capacity of B explants. Infact, there was no significant difference, under anytreatment, in the aptitude of A and B explants to regen-erate floral buds (Tabs 2-3), In contrast, in comparisonto A and B explants, there was a decrease in flower budformation on explants from infructescences with greenfruits only (see C explants. Tabs 2-3), This decreasecannot be attributed to fniit-induced aging because, inplanta, there was no increase in the number of fruits atstage C, with respect to stage B (Tab, 1), We hypothe-size that the decline in the capacity of C explants toregenerate floral buds may be due to the abscisic acidresponsable for the high rate of floral abscission occur-ring in this cultivar during the interval between stages Band C (Fig. Id, arrows). The negative effects of absdsicacid on flower bud formation in vitro have, in fact, beendemonstrated (Barendse et al, 1985), Our hypothesis isalso supported by the fact that in a tobacco cultivarwithout significant floral abscission (i.e, Nicotiana taba-cum cv, Samsun), explants from infructescences withonly green fruits develop a large number of flowers invitro (Pasqua et al, 1990),
The presence of flowers on the inflorescence (Tab. 1)may explain the strong flowering capacity of A and Bexplants (Tabs 2-3). In planta, the open flowers mayrelease promotive factors which stimulate the develop-ment of other flowers on the inflorescence, as hypothe-sized by Kinet et al. (1985) for tomato. Moreover, in N.tabacum cv. Samsun, pedicel expiants retain most oftheir flowering capacities in vitro if the presence offlowers in planta is prolonged by emasculation (Croes etal. 1986), A and B branch internodes could benefit fromthe "promotive factors" released by the flowers presentin the plant, and produce, in comparison to stage C, agreater number of flowers in vitro. The lack of a signif-icant difference between the flowering responses of Aand B explants (Tabs 2-3) may be due to the lack of asignificant differetice, between the two stages, in thenumber of floral buds and flowers on the inflorescence(Tab, 1),
In N, tabacum cv. White Burley, the promotive effectof SD on the flower formation of the thin layers resultedin a reduction in the number of newly formed vegetativebuds (Altamura et a!, 1989), SD also enhanced flower-ing and decreased vegetative budding in our genotype(Tabs 2-3); however, its effect was greater on A and Bexplants than on C explants (Tabs 2-3), In the C ex-plants, abscisic acid may have counteracted the promo-tive effects of SD, In fact, in Phaseolus vulgaris P47,application of abscisic acid to differentiating floral budsresulted in the inhibition and abscission of many buds,even in plants cultured under the photoperiodic treat-ment inductive for floral development (Bentley et al,1975),
Edited by C.H. Bornman
338
Based on the flowering response of explants consist-ing of tissues generated after the plant's transition toflowering, we conclude that in N, tabacum cv. MarylandCatterton, SD quantitatively affects the capacity to re-generate flower buds during the late post-inductivephases of the flowering process. The responsiveness tothe photoperiodic signal is high when flowers are pre-sent on the inflorescence and decreases when the platitexhibits fruits only.
Acknowledgements - Research supported by National Re-search Council of Italy, Special Project RAISA, Sub-projectN. 2, Paper N. 34.
References
Altamura, M.M., Pasqua, G. & Monacelli, B. 1986. In vitrofloral morphogenesis in a doubled haploid tobacco. - PlantSci. 46; 69-75.
- , Monacelh, B. & Pasqua, G. 1989. The effect of photo-period on flower formation in vitro in a quantitative short-day cultivar of Nicotiana tabacum. — Physiol. Plant. 76:233-239.
Barendse, G. W. M., Croes, A, F., van den Ende, G., Bosveld,M. & Creemers, T. 1985. Role of hormones on flower budformation in thin-layer explants of tobacco. - Biol. Plant.27; 40^-412.
Bentley, B., Morgan, C.B., Morgan, D.G. & Saad, F. A.1975. Plant growth substances and effects of photoperiodon flower bud development in Phaseolus vulgaris. - Nature256: 121-122.
Cousson, A. & Tran Thanh Van, K. 1983. Light- and sugar-mediated control of direct de novo flower differentiationfrom tobacco thin cell layers. - Plant Physio!. 72: 33-36.
Croes, A. F., Creemers-Molenaar, T., van den Ende, G.,Kemp, A. & Barendse, G. W. M. 1985, Tissue age as anendogenous factor controlling in vitro bud formation onexplants from the inflorescence of Nicotiana tabacum L. -J. Exp. Bot. 36: 1771-1779.
- , Derksen, R., Kemp, A., van Wezel, H. & Barendse,G. W. M. 1986. Influence of the developing fruit on agingof the floral stalk tissue with respect to flower bud regener-ation in vitro. - J, Plant Physiol. 125; 61-68.
Goodspeed, T. H. 1954. The Genus Nicotiana. - The ChronicaBotanica Company, Waltham, MA, USA. p. 61, p. 177.
Kamate, K,, Cousson, A., Trinh, T. H. & Tran Thanh Van, K.1981. Influence des facteurs genetique et physiologiqueChez le Nicotiana sur la neoformation in vitro de fleurs apartir d assises cellulaires ^pidermiques et sous-epider-miques. - Can. J. Bot. 59: 775-781.
Kinet, J,-M., Sachs, R. M, & Bemier, G, 1985, The Physiologyof Flowering, Vol, IJI. - CRC Press, Boca Raton, FL, pp.68-71, 108-111, 113-114. ISBN 0-8493-5711-X.
Krishnamoorthy, H. N. & Nanda, K.K. 1968. Floral bud re-version in Impatiens balsamina under non-inductive photo-periods. - Planta 80: 43-51.
Monacelh, B., Pasqua, G., Altamura, M, M. & Mazzolani, G.1984. Photocontrol of the morphogenesis in vitro in a short-day tobacco. - Ann. Bot, (Roma) 42: 1-5.
Pasqua, G., Altamura, M, M. & Monacelli, B. 1990. Differentphases of the reproductive process in ptanta quantitativelyaffect floral and vegetative neoformation in vitro. - Ann.Bot. (Roma) 48; 93-100.
Rajeevan, M, S, & Lang, A, 1987. Comparison of de novoflower-bud formation in a photoperiodic and a day-neutraltobacco. - Planta 171: 560-564.
Trehame, K.J., Ouinlan, J.D., Knight, J.N. & Ward, D A .1985. Hormonal regulation of fruit development in apple:'A mini review'. - Plant Growth Regul. 3: 125-132.
Physiol. Ptant. 82. 1991

Copyright © 2022 FDOKUMEN




















