
The cell coat of the developing olfactory epithelium in the chick
Upload
independentCategory
view
2download
0
Cell Differentiation, 11 (1982) 217--228 217 Elsevier/North-Holland Scientific Publishers, Ltd.
TEMPORAL AND SPATIAL DISTRIBUTION OF FIBRONECTIN DURING DEVELOPMENT OF THE EMBRYONIC CHICK LIMB BUD
ROBERT A. KOSHER, KORDASEY H. WALKER and PHILIP W. LEDGER
Department of Anatomy, University of Connecticut Health Center, Farmington, CT 06032, U.S.A.
(Accepted 17 February 1982)
Indirect immunofluorescence has been used to study the distribution of fibronectin during the course of em- bryonic chick limb morphogenesis and differentiation. At all stages of development from 19 through 25, fibro- nectin is distributed throughout the non-differentiating mesenchymal tissue directly subjacent to the apical ecto- dermal ridge (AER), i.e. the mesenchyme extending 0.15 mm or so from the AER. Fibronectin is also distributed throughout the proximal condensing central core of the limb during the early stages of cartilage differentiation. In fact, fibronectin persists as a major component of the intercellular matrix in the central core of the limb fol- lowing overt chondrogenic differentiation, since it is present as late as stage 27 throughout the well-differentiated cartilage rudiments of the radius, ulna, and humerus. The detection of fibronectin in differentiated cartilage is facilitated by pre-treatment of the sections with testicular hyaluronidase prior to immunofluorescent staining. In contrast to the chondrogenic central core of the limb, in the peripheral dorsal and ventral (myogenic) regions in which muscle differentiation is progressing, there is a progressive and striking diminution in fibronectin staining. By stage 27, little, if any, is present in the well-differentiated muscle primordia. Finally, at all stages of develop- ment, there is an accumulation of fibronectin at the ectodermal--mesenchymal interface, suggesting it is a compo- nent of embryonic limb basement membranes. On the basis of these observations, the possible role of fibronectin in limb cartilage and muscle differentiation and in other aspects of limb morphogenesis is discussed.
apical ectodermal ridge cartilage muscle differentiation fibronectin limb development
1. Introduction
The apical ectodermal ridge (AER) which extends along the distal periphery of the em- bryonic chick limb bud is required for the outgrowth and formation of distal limb struc- tures by the mesodermal cells that constitute the bulk of the limb bud (for review, see Saunders, 1977). A variety of recent studies in vivo (Stark and Searls, 1973; Summerbell et al., 1973) in organ culture (Kosher and Savage, 1980; Kosher et al., 1979a,b), and in monolayer culture (Globus and Vethamany- Globus, 1976; Solursh et al., 1981b) have demonstrated that one of the major functions of the AER is to maintain mesenchymal cells directly subjacent to it in an actively outgrow- ing, labile undifferentiated condition.
We have been investigating the mechanism
by which the AER exerts its negative effect on the chondrogenic differentiation of subja- cent mesenchymal cells and the mechanism by which differentiation is triggered when the cells leave the AER's influence. A variety of agents that elevate cyclic AMP levels elicit a dose<lependent stimulation of the already precocious chondrogenic differentiation that subridge mesoderm explants undergo in organ culture in the absence of the AER (Kosher et al., 1979b). These agents also promote the chondrogenic differentiation of limb mesen- chymal cells in high~lensity monolayer cul- ture (Ahrens et al., 1977), and stimulate the chondrogenesis of cells from the non-chon- drogenic, peripheral regions of the limb (Solursh et al., 1981a). Furthermore, in the presence of cyclic AMP derivatives, subridge mesoderm explants cultured in the presence
0045-6039/82/0000--0000/$02.75 © Elsevier/North-Holland Scientific Publishers, Ltd.

218
of the AER fail to undergo the striking proxi- mal to distal outgrowth and contour changes characteristic of control explants, and the ces- sation of AER-directed morphogenesis in the presence of these agents is accompanied by precocious cartilage differentiation (Kosher and Savage, 1980). Thus, agents that elevate cyclic AMP levels enable subridge mesen- chymal cells to overcome the negative influ- ences on cartilage differentiation and the posi- tive influences on morphogenesis being im- posed upon them by the AER (Kosher and Savage, 1980). It has, therefore, been sug- gested that when limb mesenchymal cells leave the AER's influence, their cyclic AMP con- tent increases triggering chondrogenic differ- entiation (Kosher and Savage, 1980). The increase in cyclic AMP content is thought to result from the cellular condensation and resultant intimate cell--cell interaction that precedes overt cartilage formation (Kosher and Savage, 1980; see also Solursh et al., 1978).
We have recently demonstrated that there is a gradation of hyaluronate accumulation along the proximal--distal axis of the embry- onic chick limb which correlates with the dis- tance of cells from the AER and the state of differentiation of the cells (Kosher et al., 1981). Hyaluronate is the major glycosamino- glycan accumulated by the mesenchymal cells directly subjacent to the AER, and there is a progressive decline in hyaluronate accumula- tion by more proximal cells which correlates with the initiation of the critical condensation phase of chondrogenesis (Kosher et al., 1981). We have, therefore, suggested that the AER may inhibit cartilage differentiation by caus- ing subjacent cells to secrete large amounts of hyaluronate which prevent the cell--cell inter- action necessary to trigger differentiation and/or which feed back on the cells inhibiting their production of cartilage matrix compo- nents (Kosher et al., 1981; see also Toole, 1973; Singley and Solursh, 1981). In turn, the drastic decline in hyaluronate accumulation that occurs as the cells leave the AER's influ- ence may facilitate the condensation process.
We have also suggested that the high rate of hyaluronate synthesis by subridge mesenchy- mal cells is maintained by the relatively large amount of hyaluronate that is synthesized and secreted by the AER itself (Kosher and Savage, 1981).
Another extracellular and cell surface mole- cule which may play an important role in limb chondrogenesis is the high molecular weight, adhesive glycoprotein, fibronectin (Yamada and Olden, 1978). During the initial aggregation or condensation phase the mesen- chymal cells of mouse or chick limb buds un- dergo in high density monolayer culture, they synthesize fibronectin and accumulate it on their surfaces (Hassell et al., 1978; Lewis et al., 1978). However, later in the culture peri- od, after the cells have deposited a cartilage matrix, the synthesis of fibronectin is no longer detectable and it is no longer detected on the surface of the cells (Lewis et al., 1978; Hassell et al., 1978). Similarly, when isolated chondrocytes are subjected to monolayer cul- ture, they initially accumulate fibronectin on their surfaces as they form epithelioid colo- nies, but as cartilage matrix deposition pro- ceeds, fibronectin accumulation ceases (Dessau et al., 1978). Thus, it has been sug- gested that fibronectin may be involved in promoting cell--cell adhesions during the con- densation phase of chondrogenesis, but that the loss of fibronectin may be required for chondrogenesis to occur and for maintenance of the chondrocytic phenotype (Hassell et al., 1978; Lewis et al., 1978; see also Penny- packer et al., 1979; West et al., 1979).
On the basis of these in vitro studies, we felt it would be important to examine the dis- tribution of fibronectin during all phases of limb chondrogenic differentiation in vivo. We were also very much interested in determining if fibronectin was present in the pre-chondro- genic mesenchymal tissue of the limb prior to the initiation of condensation and cartilage differentiation. We have suggested that hyal- uronate produced by the pre-chondrogenic mesenchymal cells directly subjacent to the AER is involved in maintaining the cells un-

differentiated (Kosher et al., 1981), and it has recently been demonstrated that fibronectin has a specific hyaluronate binding site (Yamada et al., 1980). We, therefore, have utilized indirect immunofluorescence to study the distribution of fibronectin during the vari- ous stages of limb morphogenesis and differ- entiation. We have found that fibronectin is distributed throughout the pre-chondrogenic mesenchymal tissue in the subridge region of the limb at all stages of development we have examined, as well as being present in the proximal central core regions of the limb in which condensation is occurring. Further- more, fibronectin remains a major component of the extracellular matrix in the central core of the limb following overt cartilage differen- tiation. In contrast, a striking diminution and eventual disappearance of fibronectin occurs in the dorsal--ventral myogenic regions of the limb. These results are compared to other recent, conflicting immunofluorescent studies on fibronectin distribution in the limb that were published while our investigation was in progress (Dessau et al., 1980; Melnick et al., 1981; see also Silver et al., 1981).
2. Materials and methods
2.1. Preparation o f tissue
Forelimb buds were removed from stage 19, 21, 23, 24, 25, and 27 (Hamburger and Hamilton, 1951) embryos of White Leghorn chicks. Whole stage 19 and 21 limb buds were fixed and processed as described in detail be- low. Stage 23 and older limb buds were usu- ally dissected into various well-defined regions, and the different limb regions fixed and processed separately. This was done to facilitate determining the precise location in the limb at which a particular pattern of fibronectin staining was observed. This is im- portant since the pattern of fibronectin dis- tribution depends upon the degree of differ- entiation, and there is a gradation of differen- tiation along the proximal--distal axis of the
219
limb, such that at all early stages of develop- ment differentiation is more advanced proxi- mally than distally.
Stage 23 and 24 limb buds were usually dissected into two regions, the subridge region (which consists of approximately the distal 0.45 mm of limb mesoderm that is capped by the AER and surrounded dorsally and ven- trally by ectoderm; see Fig. 1 in Kosher et al., 1979a), and the remaining proximal portion of the limb. At stages 25 and 27, following removal of the subridge region, the remaining proximal portion of the limb was in turn sep- arated into equally sized proximal and distal segments. In one experiment, the subridge region of stage 25 limb buds was dissected into distal and proximal regions and the remaining proximal portion of the limb dis- sected into four additional 0.3 mm segments along the proximal--distal axis as described in detail in Kosher et al. {1981; see particularly Figs. 1 and 2 in Kosher et al. (1981) for dia- grams and photographs of the various regions into which the limb bud has been dissected).
Following fixation, processing, and embed- ding as described below, whole stage 19 and 21 limb buds and the subridge regions of stage 23 and older limb buds were serially sectioned frontally and occasionally sagittally. The proximal regions were usually serially sec- tioned transversely, since this enabled us to concomitantly compare the pattern of fibro- nectin accumulation in the chondrogenic cen- tral core of the limb with that in the dorsal and ventral myogenic regions and in the lateral post-axial and pre-axial regions.
2.2. Fixation and processing o f tissues
In preparation for immunofluorescent staining, the various regions of the limb de- scribed above were processed as described by Sainte-Marie (1962). Briefly, the tissues were fixed in cold 95% ethanol, dehydrated with four changes of cold absolute ethanol, cleared with three changes of cold xylene, warmed to room temperature in xylene, passed through four baths of 56--57°C paraplast, embedded

220
with the orientation described above, and seri- ally sectioned.
2.3. Immunofluorescen t staining
Anti-fibronectin antiserum was prepared from rabbits immunized with human plasma fibronectin purified by gelatin-Sepharose affinity chromatography followed by prepara- tive sodium dodecyl sulfate (SDS) gel electro- phoresis as described in detail previously (Ledger et al., 1980). The antiserum formed a single precipitin band with purified human fibronectin or human plasma as determined by Ouchterlony double immunodiffusion. In addition, immunoprecipitat ion analysis dem- onstrated that the antiserum specifically pre- cipitated fibronectin from cell homogenates of metabolically labeled human skin fibro- blasts in culture (see Ledger et al., 1980). Furthermore, preincubation of the antiserum with purified fibronectin eliminated the spe- cific fluorescent immunostaining described below.
Sections were deparaffinized in two conse- cutive baths of cold xylene, and washed in three consecutive baths of cold 95% ethanol followed by three consecutive baths of cold phosphate-buffered saline (PBS) (Sainte- Marie, 1962}. The sections were then trans- ferred to room temperature PBS for 10 rain, after which they were treated for 30 min at room temperature with the anti-fibronectin antiserum which was usually diluted 1 : 2 0 with PBS. The sections were then washed for a total of 30 min with three changes of PBS, after which they were stained for 30 min at room temperature with fluorescein-conju- gated goat anti-rabbit IgG (Cappel Laborato- ries, Cochranville, PA) diluted 1 : 5 0 with PBS. After staining, the sections were washed three times for a total of 30 min with PBS, and then mounted under coverslips in glycerol/PBS ( 1 : 1 , v/v). The sections were examined with a Zeiss Photomicroscope III equipped with a III-RS epi-illuminator. In some experiments, prior to antiserum treat- ment the sections were treated for 30 min at
37°C with 1.25% bovine testicular hyaluron- idase (Sigma, Type 1-S) in PBS. After hyalu- ronidase treatment, the sections were washed three times for a total of 30 min with PBS followed by antiserum treatment and subse- quent t reatments as described above.
In control experiments, the anti-fibronectin serum was substi tuted with preimmune serum, anti-fibronectin serum that had been preincubated with purified fibronectin (see above), or plain PBS. Each of the above treat- ments resulted in the elimination of the spe- cific fluorescent staining seen with anti-fibro- nectin serum.
3. Results
3.1. Fibronectin distribution in the subridge region o f the limb
The pattern of fibronectin distribution in the subridge region and in progressively more proximal regions of the limb at stages 24, 25, and 23 is shown in Figs. 1, 2, and 3B, C, respectively. Fibronectin distribution in whole stage 21 limb buds is shown in Fig. 3A.
Note first of all that the pattern of fibro- nectin staining in whole stage 21 limb buds (Fig. 3A) and in the subridge region of stage 23 (Fig. 3B), stage 24 (Fig. 1A, B), and stage 25 (Fig. 2A) limb buds is virtually identical. At all of these stages fibronectin is present throughout the subridge mesoderm, but its distribution is different in the distal and prox- imal portions of the subridge region.
Fibronectin is uniformly and diffusely dis- t r ibuted throughout the distal subridge region which consists of the non<tifferentiating mes- enchymal tissue directly subjacent to the AER, i.e. the mesenchyme extending 0.15 mm or so from the AER (see Kosher et al., 1981). A uniform distribution of f ibronectin throughout the non<iifferentiating mesen- chyme directly subjacent to the AER is de- tectable as early as stage 19. Note that , in ad- dition to being present throughout the distal mesenchymal tissue, at all of these stages there
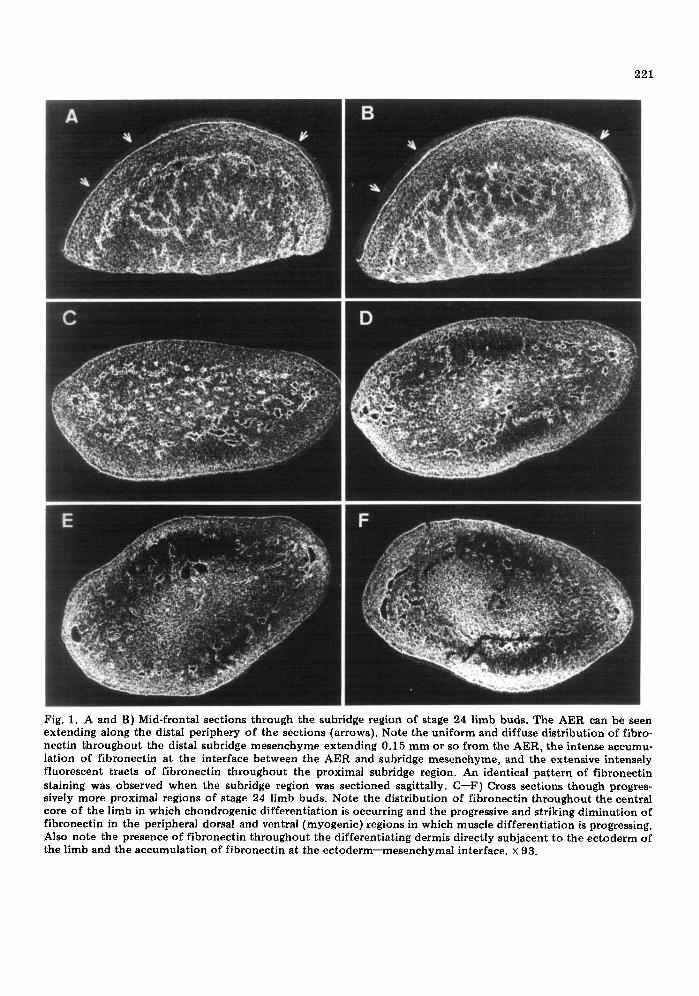
221
Fig. 1. A and B) Mid-frontal sections through the subridge region of stage 24 limb buds. The AER can be seen extending along the distal periphery of the sections (arrows). Note the uniform and diffuse distribution of fibro- nectin throughout the distal subridge mesenchyme extending 0.15 mm or so from the AER, the intense accumu- lation of fibronectin at the interface between the AER and subridge mesenchyme, and the extensive intensely fluorescent tracts of fibronectin throughout the proximal subridge region. An identical pattern of fibronectin staining was observed when the subridge region was sectioned sagittally. C--F) Cross sections though progres- sively more proximal regions of stage 24 limb buds. Note the distribution of fibronectin throughout the central core of the limb in which chondrogenic differentiation is occurring and the progressive and striking diminution of fibronectin in the peripheral dorsal and ventral (myogenic) regions in which muscle differentiation is progressing. Also note the presence of fibronectin throughout the differentiating dermis directly subjacent to the ectoderm of the limb and the accumulation of fibronectin at the ectoderm--mesenchymal interface. × 93.
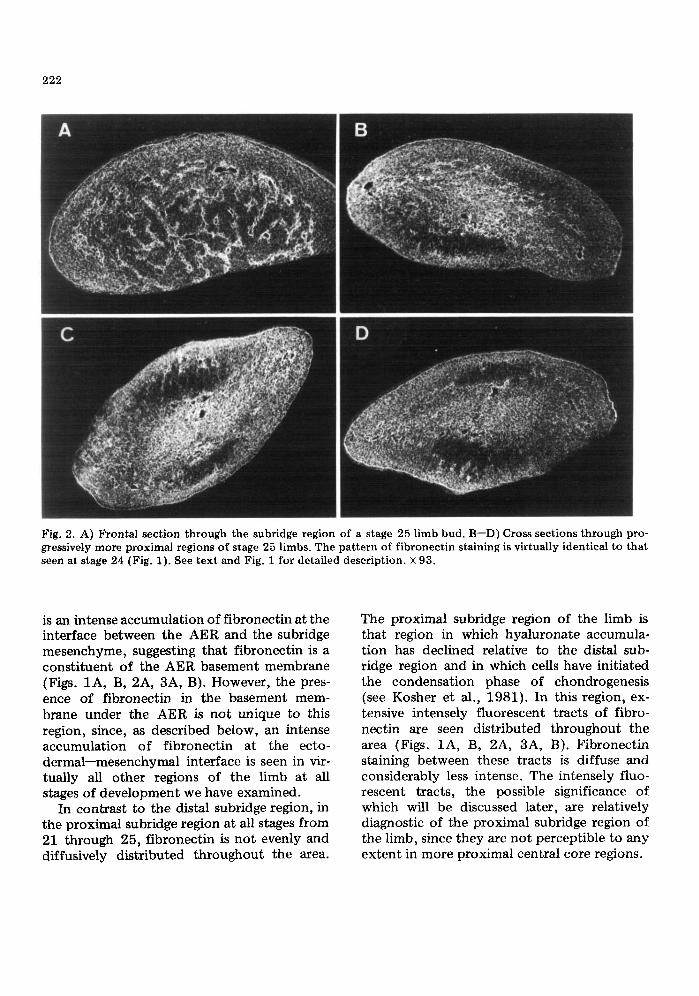
222
Fig. 2. A) Frontal section through the subridge region of a stage 25 limb bud. B--D) Cross sections through pro- gressively more proximal regions of stage 25 limbs. The pattern of fibronectin staining is virtually identical to that seen at stage 24 (Fig. 1). See text and Fig. i for detailed description. × 93.
is an intense accumulation of fibronectin at the interface between the AER and the subridge mesenchyme, suggesting that fibronectin is a consti tuent of the AER basement membrane (Figs. 1A, B, 2A, 3A, B). However, the pres- ence of fibronectin in the basement mem- brane under the AER is not unique to this region, since, as described below, an intense accumulation of fibronectin at the ecto- dermal--mesenchymal interface is seen in vir- tually all other regions of the limb at all stages of development we have examined.
In contrast to the distal subridge region, in the proximal subridge region at all stages from 21 through 25, fibronectin is not evenly and diffusively distributed throughout the area.
The proximal subridge region of the limb is that region in which hyaluronate accumula- t ion has declined relative to the distal sub- ridge region and in which cells have initiated the condensation phase of chondrogenesis {see Kosher et al., 1981). In this region, ex- tensive intensely fluorescent tracts of fibro- nectin are seen distributed throughout the area (Figs. 1A, B, 2A, 3A, B). Fibronectin staining between these tracts is diffuse and considerably less intense. The intensely fluo- rescent tracts, the possible significance of which will be discussed later, are relatively diagnostic of the proximal subridge region of the limb, since they are not perceptible to any extent in more proximal central core regions.
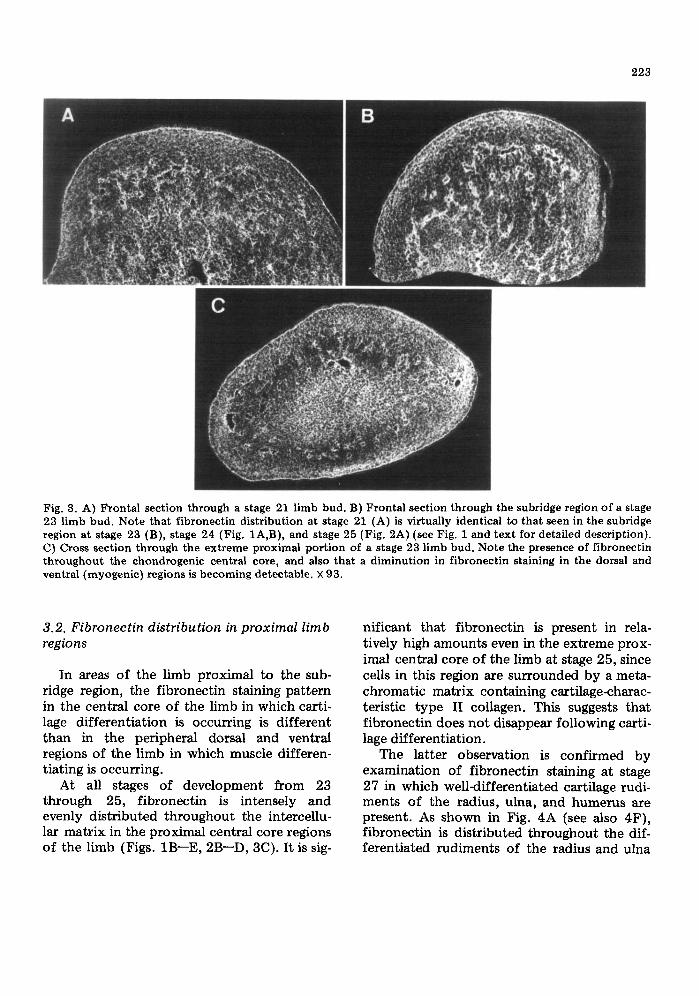
223
Fig. 3. A) Frontal section through a stage 21 limb bud. B) Frontal section through the subridge region of a stage 23 limb bud. Note that fibronectin distribution at stage 21 (A) is virtually identical to that seen in the subridge region at stage 23 (B), stage 24 (Fig. 1A,B), and stage 25 (Fig. 2A) (see Fig. 1 and text for detailed description). C) Cross section through the extreme proximal portion of a stage 23 limb bud. Note the presence of fibronectin throughout the chondrogenic central core, and also that a diminution in fibronectin staining in the dorsal and ventral (myogenic) regions is becoming detectable, × 93.
3.2. Fibronectin distribution in proximal limb regions
In areas of the limb proximal to the sub- ridge region, the fibronectin staining pattern in the central core of the limb in which carti- lage differentiation is occurring is different than in the peripheral dorsal and ventral regions of the limb in which muscle differen- tiating is occurring.
At all stages of development from 23 through 25, fibronectin is intensely and evenly distributed throughout the intercellu- lar matrix in the proximal central core regions of the limb (Figs. 1B--E, 2B--D, 3C). It is sig-
nificant that fibronectin is present in rela- tively high amounts even in the extreme prox- imal central core of the limb at stage 25, since cells in this region are surrounded by a meta- chromatic matrix containing cartilage-charac- teristic type II collagen. This suggests that fibronectin does not disappear following carti- lage differentiation.
The latter observation is confirmed by examination of fibronectin staining at stage 27 in which weU-differentiated cartilage rudi- ments of the radius, ulna, and humerus are present. As shown in Fig. 4A (see also 4F), fibronectin is distributed throughout the dif- ferentiated rudiments of the radius and ulna
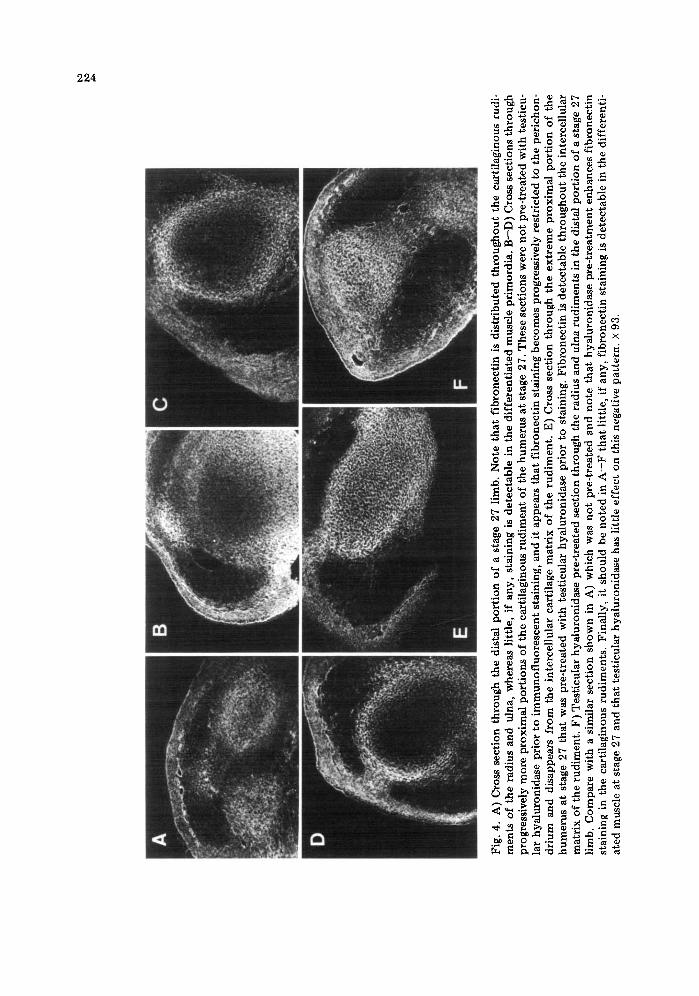
Fig
. 4.
A
) C
ross
sec
tio
n t
hro
ug
h t
he
dist
al p
ort
ion
of
a st
age
27 l
imb.
No
te t
hat
fib
ron
ecti
n i
s d
istr
ibu
ted
th
rou
gh
ou
t th
e ca
rtil
agin
ou
s ru
di-
men
ts o
f th
e ra
dius
and
ul
na,
whe
reas
lit
tle,
if
any
, st
aini
ng i
s d
etec
tab
le i
n th
e d
iffe
ren
tiat
ed m
usc
le p
rim
ord
ia.
B--
D)
Cro
ss s
ecti
on
s th
rou
gh
pr
ogre
ssiv
ely
mo
re p
rox
imal
po
rtio
ns
of
the
cart
ilag
ino
us
rud
imen
t o
f th
e h
um
eru
s at
sta
ge 2
7. T
hes
e se
ctio
ns
wer
e n
ot
pre
-tre
ated
wit
h t
esti
cu-
lar
hy
alu
ron
idas
e pr
ior
to i
mm
un
ofl
uo
resc
ent
stai
ning
, an
d it
app
ears
th
at f
ibro
nec
tin
sta
inin
g b
eco
mes
pro
gre
ssiv
ely
res
tric
ted
to
th
e p
eric
ho
n-
dri
um
an
d di
sapp
ears
fr
om
th
e in
terc
ellu
lar
cart
ilag
e m
atri
x o
f th
e ru
dim
ent.
E)
Cro
ss s
ecti
on
th
rou
gh
th
e ex
trem
e p
rox
imal
po
rtio
n o
f th
e h
um
eru
s at
sta
ge 2
7 th
at w
as p
re-t
reat
ed w
ith
tes
ticu
lar
hy
alu
ron
idas
e p
rio
r to
sta
inin
g. F
ibro
nec
tin
is
det
ecta
ble
th
rou
gh
ou
t th
e in
terc
ellu
lar
mat
rix
of
the
rud
imen
t. F
) T
esti
cula
r h
yal
uro
nid
ase
pre
-tre
ated
sec
tio
n t
hro
ug
h t
he
radi
us a
nd u
lna
rud
imen
ts i
n th
e di
stal
po
rtio
n o
f a
stag
e 27
li
mb.
Co
mp
are
wit
h a
sim
ilar
sec
tio
n s
ho
wn
in
A)
whi
ch w
as n
ot
pre
-tre
ated
and
no
te t
hat
hy
alu
ron
idas
e p
re-t
reat
men
t en
han
ces
fib
ron
ecti
n
stai
ning
in
the
cart
ilag
inou
s ru
dim
ents
. F
inal
ly,
it s
ho
uld
be
no
ted
in
A--
F t
hat
lit
tle,
if
any
, fi
bro
nec
tin
sta
inin
g i
s d
etec
tab
le i
n th
e d
iffe
ren
ti-
ated
mus
cle
at s
tage
27
and
that
tes
ticu
lar
hy
alu
ron
idas
e ha
s li
ttle
eff
ect
on
thi
s ne
gati
ve p
atte
rn.
X 9
3.

225
in the distal region of the limb. These rudi- ments also stain intensely with monoclonal antibodies to type II collagen (Kosher and Walker, 1982). In the more proximal regions of stage 27 limbs in which the cartilaginous rudiment of the humerus is present, it appears that fibronectin staining becomes progres- sively restricted to the perichondrium and dis- appears from the intercellular cartilage matrix of the rudiment (Fig. 4B--D). However, if sec- tions from these limb regions are treated with testicular hyaluronidase prior to immunofluo- rescent staining, fibronectin becomes detect- able throughout the intercellular matrix of the rudiments (Fig. 4A, F). This confirms the observations of others that in mature carti- lage, fibronectin staining is masked by proteo- glycans in the matrix (Weiss and Reddi, 1980, 1981; Melnick et al., 1981).
In contrast to the chondrogenic central core of the limb in which fibronectin persists following cartilage differentiation, in the dor- sal and ventral {myogenic) regions in which muscle differentiation is progressing, there is a progressive and striking diminution in f ibro- nectin accumulation. This sequence is particu- larly evident at stages 24 and 25. During these stages, in the region of the limb immediately subjacent to the proximal subridge region, fibronectin staining persists in the peripheral dorsal and ventral areas, although it is dimin- ished relative to the central core and lateral areas (Fig. 1B). However, in progressively more proximal dorsal and ventral myogenic areas, there is a striking diminution in fibro- nectin, and little, if any, is seen in the ex- treme proximal myogenic regions (Figs. 1C--E, 2B--D). Temporally, a diminution in fibronectin staining in the dorsal and ventral myogenic regions is detectable as early as stage 23 in the extreme proximal portion of the limb {Fig. 3C). By stage 27, little, if any, staining in the well~lifferentiated muscle pri- mordia is observed (Fig. 4).
Finally, it should be noted that in the prox- imal portion of the limb at all stages of devel- opment, fibronectin is present throughout the differentiating dermis directly subjacent to
the ectoderm of the limb. In addition, as pre- viously described, there is an intense accumu- lation of fibronectin at the ectodermal-- mesenchymal interface throughout the prox- imal portion of the limb, suggesting that fibronectin is a constituent of embryonic limb basement membranes.
4. Discussion
Our results demonstrate that at all stages of development from 19 through 25, fibronectin is present throughout the distal subridge region of the limb which consists of the undif- ferentiating mesenchymal tissue directly sub- jacent to the AER. Our detection of fibronec- tin in the limb mesenchyme as early as stage 19 confirms the study of Dessau et al. (1980), who also reported the presence of fibronectin in the interstitial space of the mesenchyme at this early stage of development, but is incon- sistent with the recent study of Melnick et al. (1981), in which fibronectin was not detect- able in any region of the limb until at least stage 22. It should also be noted that, in a recent abstract, Tomasek et al. (1980) have also reported that the mesenchyme at the distal tip of the embryonic chick and duck wing bud is very rich in fibronectin.
The presence of fibronectin throughout the pre~hondrogenic mesenchyme directly subja- cent to the AER is of considerable interest in view of the high rate of accumulation of hyal- uronate by cells in this region (Kosher et al., 1981). We have suggested that this extracellu- lar hyaluronate is involved in inhibiting differ- entiation by preventing a cell--cell interaction necessary to trigger differentiation and/or by feeding back upon the cells and inhibiting their production of cartilage matrix compo- nents (Kosher et al., 1981). It has recently been demonstrated that fibronectin has a specific hyaluronate binding site (Yamada et al., 1980). That a developmentally important interaction between extracellular hyaluronate and cell surface fibronectin may be occurring in the distal subridge region is an intriguing,

226
though highly speculative, possibility. Our results also demonstrate that fibronec-
tin is present in the proximal subridge region of the limb, and a striking feature of fibronec- tin distribution in this region is the extensive intensely fluorescent tracts that are present throughout the area. Since in the proximal subridge region the critical condensation phase of chondrogenesis is being initiated, it is tempt- ing to speculate that the tracts o f fibronectin may represent a re-organization of cell surface fibronectin in response to the initiation of the condensation process and/or may reflect the cellular re-organization occurring during this process. An alternate, perhaps more likely possibility, however, is that the tracts repre- sent fibronectin associated with the endo- thelial lining of b lood vessels. Observation of Fig. 9 in Caplan (1981) (see also Caplan and Koutroupas, 1973) indicates that the prox- imal subridge region is an area of the limb containing a relatively rich capillary network, and it has been demonstrated that f ibronectin is synthesized and secreted by endothelial cells in culture (Birdwell et al., 1978; Jaffe and Mosher, 1978a,b; Macarek et al., 1978). Furthermore, immunofluorescence studies have shown that the subendothelial basement membrane region of blood vessels in the chick embryo is rich in fibronectin (Linder et al., 1975). In addition, fibronectin is localized in association with endothelial cells during the vascular invasion of cartilage that precedes osteogenesis (Weiss and Reddi, 1980). Deter- mining, however, the precise nature of the tracts of fibronectin in the proximal subridge region will require a technique with more resolution than indirect immunofluorescence.
At stages 23 through 25, fibronectin is evenly distributed throughout the avascular condensed central core of the limb directly subjacent to proximal subridge region. An ac- cumulation of fibronectin along the surfaces of closely apposed cells during the aggregation or condensation phase limb mesenchymal cells undergo in high density monolayer cul- ture has also been reported, and it has been suggested that f ibronectin may be involved in
promoting cell adhesions during condensation Hassell et al., 1978; Lewis et al., 1978; New- man and Frisch, 1979). In this regard, it is of interest that Dessau et al. (1980) and Silver et al. (1981) have detected the presence of type I collagen as well as fibronectin in the con- densing core of the limb, and suggested that an interaction between type I collagen and fibronectin which has a high-affinity binding site for type I collagen may be involved in the critical condensation phase of chondrogenesis.
At stages 24 and 25, fibronectin continues to be present throughout the intercellular matrix in the central core of the limb in which the cells are progressively accumulating cartilage matrix. In fact, we continue to detect the presence of fibronectin at stage 27 throughout the well-differentiated cartilage rudiments of the radius, ulna, and humerus which consist of chondrocytes surrounded by a metachromatic matrix containing type II col- lagen. The detect ion of fibronectin in differ- entiated cartilage is facilitated by pre-treat- ment of the sections with testicular hyaluroni- dase prior to immunofluorescent staining as has been reported by others (Weiss and Reddi, 1980; 1981; Melnick et al., 1981). The presence of fibronectin during the critical early stages of cartilage differentiation as well as in mature cartilage indicates that in vivo the disappearance of fibronectin is not required for cartilage differentiation to occur or for maintenance of the chondrocyt ic phenotype as others have suggested (see Hassell et al., 1978; Lewis et al., 1978; Penny- packer et al., 1979; West et al., 1979). It is noteworthy, however, that exogenous fibro- nectin appears to modulate the phenotypic expression of chondrocytes in monolayer cul- ture (Pennypacker et al., 1979; West et al., 1979).
In contrast to the chondrogenic central core of the limb in which fibronectin persists following differentiation, in the dorsal and ventral (myogenic) regions in which muscle differentiation is progressing, there is a pro- gressive and striking diminution in fibronectin. In their Abstract, Tomasek et al. (1980)have

also reported a marked reduction of fibro- nectin in the proximal myogenic regions of chick and duck limbs. We find a diminution in fibronectin as early as stage 23 in the extreme proximal dorsal and ventral regions of the limb. This diminution thus occurs precisely at the stage Hilfer et al. (1973) have detected cells in proximal dorsal and ventral regions of the limb becoming arranged and aligned in the clusters and cords which are the initial struc- tural changes characteristic o f muscle differ- entiation. The initiation of the synthesis of muscle troponin also occurs during this period (Llewellyn-Smith, 1978). It is noteworthy that the diminishment in fibronectin pre- cedes myoblast fusion which is first detect- able at stage 26 (Hilfer et al., 1973) .
The striking reduction in fibronectin which we observe during the progression of muscle differentiation in vivo also seems to occur during muscle differentiation in vitro. Chen (1977) and Furcht et al. (1978} have observed a marked reduction in fibronectin fol lowing the differentiation and fusion of myoblasts in monolayer culture. Furthermore, it has been demonstrated that exogenous fibronectin de- lays fusion, suggesting that the diminution of fibronectin may be necessary for terminal muscle differentiation (Podleski et al., 1979) .
Finally, it is noteworthy that, in all regions of the limb at all stages of development we have examined, fibronectin is present at the ectodermal--mesenchymal interface, suggest- ing that it is a constituent of embryonic limb basement membranes. Fibronectin has been localized in basement membranes at the inter- face between epithelial and mesenchymal components in several other developing sys- tems (Linder et al., 1975; Wartiovaara et al., 1978; Kurkinen et al., 1979; Thesleff et al., 1979) .
Acknowledgements
This work was supported by Grant PCM- 7925907 from the National Science Founda- tion to RAK. We thank Dr. D.K. Morest for use of his fluorescence microscope.
227
References
Ahrens, P.B., M. Solursh and R.S. Reiter: Dev. Biol. 60, 69--82 (1977).
Birdwell, C.R., D. Gospodarowicz and G.L. Nicolson: Proc. Natl. Acad. Sci. U.S.A. 75, 3273--3277 (1978).
Caplan, A.I.: In: Levels of Genetic Control in Devel- opment, eds. S. Sublelny and U.K. Abbott (Alan R. Liss, New York) pp. 37--68 (1981).
Caplan, A.I. and S. Koutroupas: J. Embryol. Exp. Morphol. 29, 571--583 (1973).
Chen, L.B.: Cell 10,393--400 (1977). Dessau, W., J. Sasse, R. Timpl, F. Jilek and K. vonder
Mark: J. Cell Biol. 79,342--355 (1978). Dessau, W., H. yon der Mark, K. vonder Mark and S.
Fischer: J. Embryol. Exp. Morphol. 57, 51--60 (1980).
Furcht, L.T., D.F. Mosher and G. Wendelschafer- Crabb: Cell 13,263--271 (1978).
Globus, M. and S. Vethamany-Globus: Differentia- tion 6, 91--96 (1976).
Hamburger, V. and H.L. Hamilton: J. Morphol. 88, 49--92 (1951).
Hassell, J.R., J.P. Pennypacker, K.M. Yamada and R.M. Pratt: Ann. N.Y. Acad. Sci. 312, 406--409 (1978).
Hiller, S.R., R.L. Searls and V. Fonte: Dev. Biol. 30, 374--391 (1973).
Jaffe, E.A. and D.F. Mosher: Ann. N.Y. Acad. Sci. 312,122--131 (1978a).
Jaffe, E.A. and D.F. Mosher: J. Exp. Med. 147, 1779--1791 (1978b).
Kosher, R.A. and M.P. Savage: J. Embryol. Exp. Morphol. 56, 91--105 (1980).
Kosher, R.A. and M.P. Savage: Nature 291,231--232 (1981).
Kosher, R.A. and K.H. Walker: in preparation (1982). Kosher, R.A., M.P. Savage and S.-C. Chan: J. Em-
bryol. Exp. Morphol. 50, 75--97 (1979a). Kosher, R.A., M.P. Savage and S.-C. Chan: J. Exp.
Zool. 209, 221--228 (1979b). Kosher, R.A., M.P. Savage and K.H. Walker: J. Em-
bryol. Exp. Morphol. 63, 85--98 (1981}. Kurkinen, M., K. Alitalo, A. Vaheri, S. Stenman and
L. Saxen: Dev. Biol. 69, 589--600 (1979). Ledger, P.W., N. Uchida and M.L. Tanzer: J. Cell
Biol. 87,663--671 (1980). Lewis, C.A., R.M. Pratt, J.P. Pennypacker and J.R.
Hassell: Dev. Biol. 64, 31--47 (1978). Linder, E., A. Vaheri, E. Ruoslahti and J. Wartio-
vaara: J. Exp. Med. 142, 41--49 (1975). Llewellyn-Smith, I.J.: Dev. Biol. 67, 40--53 (1978). Macarak, E.J., E. Kirby, T. Kirk and N.A. Kefalides:
Proc. Natl. Acad. Sci. U.S.A. 75, 2621--2625 (1978).

228
Melnick, M., T. Jaskoll, A.G. Brownell, M. Mac- DougaU, C. Bessem and H.C. Slavkin: J. Embryol. Exp. Morphol. 63 ,193- -206 (1981).
Newman, S.A. and H.L. Frisch: Science 205, 662-- 668 (1979).
Pennypacker, J.P., J.R. Hassell, K.M. Yamada and R.M. Pratt: Exp. Cell Res. 121 ,411--415 (1979).
Podleski, T.R., I. Greenberg, J. Schlessinger and K.M. Yamada: Exp. Cell Res. 122, 317--326 (1979).
Sainte-Marie, G.: J. Histochem. Cytochem. 10, 250-- 256 (1962).
Saunders, J.W.: In: Vertebrate Limb and Somite Mor- phogenesis, eds. D.A. Ede, J.R. Hinchliffe and M. Balls (Cambridge University Press, Cambridge) pp. 1--24 (1977).
Silver, M.H., J.M. Foidar t and R.M. Pratt: Differentia- tion 18, 141--149 (1981).
Singley, C.T. and M. Solursh: Dev. Biol. 84, 102--120 (1981).
Solursh, M., P.B. Ahrens and R.S. Reiter: In vitro 14, 51--61 (1978).
Solursh, M., R.S. Reiter, P.B. Ahrens and B.M. Vertel: Dev. Biol. 83, 9--19 (1981a).
Solursh, M., C.T. Singley and R.S. Reiter: Dev. Biol. 86, 471--482 (1981b).
Stark, R.J. and R.L. Searls: Dev. Biol. 33, 138--153 (1973).
Summerbell, D., J.H. Lewis and L. Wolpert: Nature New Biol. 244,492--496 (1973).
Thesleff, I., S. Stenman, A. Vaheri and R. Timp| : Dev. Biol. 70 ,116- -126 (1979).
Tomasek, J.J., J.E. Mazurkiewicz and S.A. Newman: J. Cell Biol. 87, l 16a (1980).
Toole, B.P.: Am. Zool. 13, 1061--1065 (1973). Wartiovaara, J., I. Leivo, I. Virtanen and A. Vaheri:
Ann. N.Y. Acad. Sci. 312, 132--141 (1978). Weiss, R.A. and A.H. Reddi: Proc. Natl. Acad. Sci.
U.S.A. 77, 2074--2078 (1980). Weiss, R.A. and A.H. Redd i : J. Cell Biol. 88, 630---
636 (1981). West, C.M., R. Lanza, J. Rosenbloom, M. Lowe, H.
Holtzer and N. Avdalovic: Cell 17, 491--501 (1979).
Yamada, K.M. and K. Olden: Nature 275, 179--184 (1978).
Yamada, K.M., D.W. Kennedy, K. Kimata and R.M. Pratt: J. Biol. Chem. 255, 6055--6063 (1980).
Copyright © 2022 FDOKUMEN




















