
Food bloggers experience agriculture on Kansas Food Farm ...
Upload
becomeanailtechnicianCategory
view
1download
0
This article was downloaded by: [112.253.13.165]On: 22 March 2014, At: 07:13Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Bird StudyPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/tbis20
Influence of environmental factors and energetic valueof food on Little Tern Sterna albifrons chick growthand food delivery: Capsule Chick diet and early growthparameters differed between birds breeding on natural(sandy beaches) and alternative (salinas) habitats.Vítor H. Paiva , Jaime A. Ramos , Teresa Catry , Patrícia Pedro , Renata Medeiros & JorgePalmaPublished online: 29 Mar 2010.
To cite this article: Vítor H. Paiva , Jaime A. Ramos , Teresa Catry , Patrícia Pedro , Renata Medeiros & Jorge Palma (2006)Influence of environmental factors and energetic value of food on Little Tern Sterna albifrons chick growth and food delivery:Capsule Chick diet and early growth parameters differed between birds breeding on natural (sandy beaches) and alternative(salinas) habitats., Bird Study, 53:1, 1-11, DOI: 10.1080/00063650609461410
To link to this article: http://dx.doi.org/10.1080/00063650609461410
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of theContent. Any opinions and views expressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon andshould be independently verified with primary sources of information. Taylor and Francis shall not be liable forany losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoeveror howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use ofthe Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

Variation in food supply has a marked impact on breeding parameters and productivity of small seabirdssuch as terns (Monaghan et al. 1989), because they possess little spare capacity in their energy budgets andspend a larger proportion of their time foraging compared to other seabird species (Pearson 1968).Variation in food supply has been shown to influencechick growth and productivity (Monaghan et al. 1989),
due to variation in the availability of fish, in the vicinity of the tern colonies. Prey species can also varyin energy content (Hislop et al. 1991, Anthony et al.2000), raising the hypothesis that energy limitationmay occur if the most abundant prey available are lowin energy (Dahdul & Horn 2003). Of all the temperatetern species Little Tern Sterna albifrons is arguably themost estuarine and the species that forages closest tothe breeding colonies (Cramp 1985, Fasola & Bogliani1990). Their estuarine foraging areas are characterized
Bird Study (2006) 53, 1–11
© 2006 British Trust for Ornithology
Influence of environmental factors and energeticvalue of food on Little Tern Sterna albifrons chickgrowth and food delivery
VÍTOR H. PAIVA1*, JAIME A. RAMOS1, TERESA CATRY1, PATRÍCIA PEDRO1, RENATA MEDEIROS1
and JORGE PALMA2
1Institute of Marine Research (IMAR), Departamento de Zoologia, Universidade de Coimbra, 3004-517Coimbra, Portugal and 2CCmar, Universidade do Algarve, FCMA, Campus de Gambelas, 8005-139, Faro, Portugal
Capsule Chick diet and early growth parameters differed between birds breeding on natural (sandybeaches) and alternative (salinas) habitats.Aims To describe diet and growth of Little Tern chicks, to examine the influence of tide and wind speedon food delivered to chicks in southern Portugal, and to determine whether diet and early chick growthparameters differed between birds breeding in natural (sandy beaches) and man-made alternative (salinas) habitats. Methods Nests were marked and visited regularly. Chicks were ringed and weighed at hatching.Between 12 and 14 nests were enclosed by a fence in order to measure and weigh chicks every day.Prey items delivered to fenced chicks were observed daily from hides, in different tidal periods and windspeed conditions. Prey items dropped in the colonies were collected. Foraging efficiency of adults wasevaluated in relation to tide and wind speed. Fresh prey items were collected to determine their energeticcontent using a calorimeter. Results A chicks grew faster than B chicks, but both attained the same weight, tarsus-length and wing-length at 19 days old; C chicks attained the same values at day 21. There were significant differencesin early growth parameters. A and B chicks grew faster on sandy beaches than on salinas. The main preyitems were Atherina spp., Fundulus sp. and shrimps in salinas and Sardina pilchardus, Atherina spp. andBelone belone on sandy beaches. There was no correlation between ingestion rate and chick age, butmean length of prey ingested by chicks increased significantly with age. Conclusion Tide and wind speed influenced both size and type of prey delivered to chicks. Parents metthe increasing energy demands of the growing chicks by adjusting prey size rather than prey delivery frequency. Differences in chick growth between salinas and sandy beaches could be related to bothparental quality and diet. The presence of a high energy-content euryhaline fish species in the salinas,together with more sheltered conditions and no tidal influences, may be important in explaining the useof salinas by Little Terns.
*Correspondence author. Email: [email protected]
Dow
nloa
ded
by [
112.
253.
13.1
65]
at 0
7:13
22
Mar
ch 2
014

by shallow and clear waters (Allcorn et al. 2003).Growth and chick food provisioning of estuarine foraging terns are very sensitive to variations in foodavailability due to wind speed and tide (Frank 1992),but very little is know about the effects of these two factors on Little Tern chick food provisioning.
Norman (1992) suggested that the type and size ofavailable food is the main factor influencing growthrates in Little Tern. However, very little is known aboutgrowth of Little Terns in other areas of Europe, especially in southern Europe. Moreover, Norman(1992) did not distinguish between growth rates of thedifferent chicks within a brood.
In Algarve, southern Portugal, where this study wasconducted, Little Terns breed in natural (sandybeaches) and man-made alternative (salinas) habitats(Catry et al. 2004). Foraging Little Terns feed in natural(estuarine lagoon system and adjacent sea) and man-made (salinas and artificial channels) habitats. Severalbreeding parameters such as egg size and clutch sizewere larger in natural habitats, suggesting that these arepreferred to man-made alternative habitats (Catry et al.2004).
Here, we describe the diet and growth of Little Tern chicks in Ria Formosa, Algarve, and determinewhether diet and early chick growth parameters differed between birds breeding in natural and man-made alternative habitats. On estuaries and coastalareas, tide and wind speed are considered the main factors affecting the accessibility of prey to terns (Frank1992, Dunn 1972), and we decided to evaluate their influence on the provisioning of Little Tern chicks inRia Formosa. We examined whether the calorific con-tent of prey delivered to Little Tern chicks differedbetween birds breeding in sandy beaches and salinas.This was justified because, for instance, birds breeding on salinas were observed foraging in water-tanks used for salt production, where the compositionof prey is likely to differ from that available in thelagoon and sea that surround the colonies in naturalhabitats.
METHODS
Ria Formosa Natural Park is situated in the south coastof Portugal, Algarve, and consists of a complex tidalsystem of natural and semi-natural channels (lagoon),marshland and barrier islands covering an area of 18 400 ha along 60 km of coast. In the margins of themarshland area, there are man-made salt-gatheringtanks (salinas) and extensive fish farms. Close to the
2 V.H. Paiva et al.
© 2006 British Trust for Ornithology, Bird Study, 53, 1–11
Park’s northern borders, there are three large towns,Faro, Olhão and Tavira.
Our observations took place in both natural (sandybeaches of the barrier islands Armona, Faro andBarreta) and alternative (salinas around Santa Luzia,Tavira) breeding habitats (Catry et al. 2004). The closest foraging areas were the lagoon and the sea forbirds breeding in natural habitats, and the lagoon andsalinas for those breeding in alternative habitats.
Chick growth
Nests in the salinas were marked in May and June 2002and 2003, and visited every two days until hatching,and daily thereafter. Chicks were weighed and ringedon the day of hatching (day 0). Their wing-length (tothe nearest 1 mm) and tarsus-length (to the nearest 0.1mm) were measured on the day of hatching and there-after whenever chicks were found. Fourteen nests in2002 and twelve in 2003 from Santa Luzia wereselected within an area of approximately 1500 m2 andfenced with hexagonal wire, 0.3 m high and 1 cm2
mesh, to keep chicks near the nest-site. Fences wereabout 1.5 m in diameter. Inside the fences, we placedsome rocks for use as perches by the parents, as well asa wooden shelter and some vegetation to providechicks with protection against the sun and possibleavian predators. Chicks were designated as A, B and Caccording to hatching order (when two chicks hatchedon the same day, we considered the A chick as the onewith higher mass, greater egg volume and greater tarsus-length). Each chick (A, B and C) of a nest wasmarked, using non-toxic paint, with a different colouron the head and breast. These chicks (n = 32 in 2003and n = 25 in 2002) were weighed daily between 08:00and 11:00 hours, until they were 20 to 22 days old.Daily or on alternate days in 2003, the chicks ofunfenced nests (see Fig. 2) were also weighed in order to verify the impact of the fences on chickgrowth.
We verified that mass data of Little Tern chicks fordays 0–4 were fitted to a quadratic regression model.a
Two parameters were calculated to characterizegrowth of the 2003 chicks after the age of four days: linear growth rate (LGR) and asymptotic mass (AM).LGR was defined as the slope of a regression line ofchick mass against date (Nisbet et al. 1995) during thelinear growth period (5–14 days). AM was defined asthe mean of all masses measured during the period ofnear-constant mass (15–22 days) (Nisbet et al. 1998).Linear growth rate and asymptotic mass were calcu-
Dow
nloa
ded
by [
112.
253.
13.1
65]
at 0
7:13
22
Mar
ch 2
014

lated for all fenced chicks (weighed daily) with at leastfour and two data points, respectively.
Food delivery
During regular visits to the colonies on sandy beaches(Armona, Barreta, Praia de Faro) and salinas (SantaLuzia I, Santa Luzia II, Arraial Ferreira Neto, ValeCarangueijo) in 2002 and 2003, 596 dropped preyitems (this includes all prey that were not ingested)were collected and identified. Observation of fooddelivery to fenced chicks occurred between 11 June and5 July 2003. From the fenced nests, ten were selectedaccording to their location. Two portable hides, placed2–7 m from these nests, were used to observe and identify prey delivered to chicks, using 10 × 40 binocu-lars. A total of 72 hours of observation were made,divided into periods of one to ten hours (median = twohours), in different tidal phases (low, high, incomingand receding). We considered that each tidal phaselasted three hours (1.5 hours before and after each tidepeak). At the beginning of each observation period wemeasured wind speed with a field anemometer and later classified it into three categories: 0–2, 2–4, >4m/s. Water surface conditions were evaluated as: calm,small undulation, high undulation.
For each prey item offered to chicks we registered thenumber of the nest, chick (A, B or C), time, prey species,prey length and fate. The fate of each prey was classifiedas ingested or not ingested (the latter category includedprey robbed by adults, eaten by parents or dropped). Wewere familiar with the main prey species in the area(Muzavor et al. 1993, Sobral & Gomes 1998), havingcollected prey dropped in colonies and made observa-tions in 2002. Tests to identify prey and establish theirlength by the several observers were made during thecourtship phase. When it was not possible to identifyprey species, only the genus or family name was given, orthey were classified as non-identified. Gobiidae andBlenniidae were difficult to separate and were includedin the same category (Gobiidae/Blenniidae) with theexception of the Gobiidae Pomatoschistus sp., which waseasily identified. Prey length was determined in relationto the average Little Tern bill length (mean culmen =29.85 mm, se = ±0.26, n = 26 Little Tern adults capturedin 2003, at the barrier island of Armona). Throughoutthis paper, size is used as a synonym of length. ‘Bill-units’were used with a division of 0.25 ‘bills’. Prey itemsdropped were picked up after each observation periodand used to clarify identification of some prey deliveredto chicks.
© 2006 British Trust for Ornithology, Bird Study, 53, 1–11
Little Tern chick growth and diet 3
Foraging behaviour in relation to tide
Fifteen transects were made between 12 June and 12July 2002, and 33 between 15 May and 19 June 2003,to collect data on foraging adults in relation to tide.Transects were made along the salinas of Santa Luziaand adults actively foraging were assigned to habitattype (salinas or lagoon) during four tidal phases (low,high, incoming, receding) in 2003 and two tide categories (low, high) in 2002.
In 2003 we observed 45 adults diving (for at least oneminute each) in the lagoon habitat and registered thenumber of dives per minute and the number of preycaptured per minute in relation to tidal phase.
Calorific content
Fresh prey items found in the colonies were measured(standard length, maximum width), weighed, washedwith fresh water and frozen. To determine the calorificcontent, items were thawed and dried in the oven at60°C. Once in the oven, prey items were weighed dailyuntil constant mass values were obtained, and theneach item was crushed to dust in a mortar. A smallquantity of the samples (0.058 to 0.200 g) were converted into discs in a press and used to determinethe calorific content of each prey species in a PARR1425 calorimeter. Three discs of each sample weremade and the mean value calculated. The calorificcontent was given by the calorimeter in calories pergram of dry weight (cal/g dry wt) (Henken et al. 1986,Fraschetti et al. 1994); this value was converted tojoules per gram of dry weight (J/g dry wt) by multi-plying by 4.187.
Statistical analysis
Data from sandy beaches (Barreta, Praia de Faro andArmona) were grouped. We evaluated differences bothin the linear growth rate and the asymptotic massbetween A and B chicks in the salinas during 2003using Student’s t-tests. One-way ANOVA, followed by a post-hoc test (Tukey test), was used to test for differences in the early chick growth parameters (A andB chicks were grouped) among salinas in 2002, salinasin 2003, and beaches in 2003. ANOVA was also used totest the null hypothesis that the mean calorific contentis similar between prey species. We used Pearson correlation coefficients to verify: (1) whether the proportion of each prey item dropped in the salinas wassimilar to that ingested by the chicks; and (2) whether
Dow
nloa
ded
by [
112.
253.
13.1
65]
at 0
7:13
22
Mar
ch 2
014

the proportion of each dropped prey item was similarbetween 2002 and 2003 (data were arcsine trans-formed). To test the null hypothesis that the meanpercentage (data arcsine transformed) of the mainitems in the diet of A and B chicks was similar, we usedStudent’s t-tests. A Mann–Whitney test was used totest for differences between delivery and ingestionrates. The relationships between both ingestion rateand length of prey delivered in relation to chick agewere examined with Spearman correlations.Differences in the delivery rate due to tide and windspeed were examined using a two-way ANOVA test. Theinfluence of environmental factors (i.e. tide state andwind speed) in the rate and length of prey deliveredwas tested using Kruskal–Wallis non-parametricANOVA. Differences in the number of adults foraging inthe lagoon in relation to tide were examined withMann–Whitney (2002) and Kruskal–Wallis (2003)tests. One-way ANOVA, followed by a post-hoc test(Tukey test), was used in capture success data to test for differences between the four tidal phases. A significance level of P ≤ 0.05 was set a priori for all statistical tests. We present values as mean ± se. Allanalyses were conduced using STATISTICA version 6.0(Statsoft 1996).
RESULTS
Chick growth
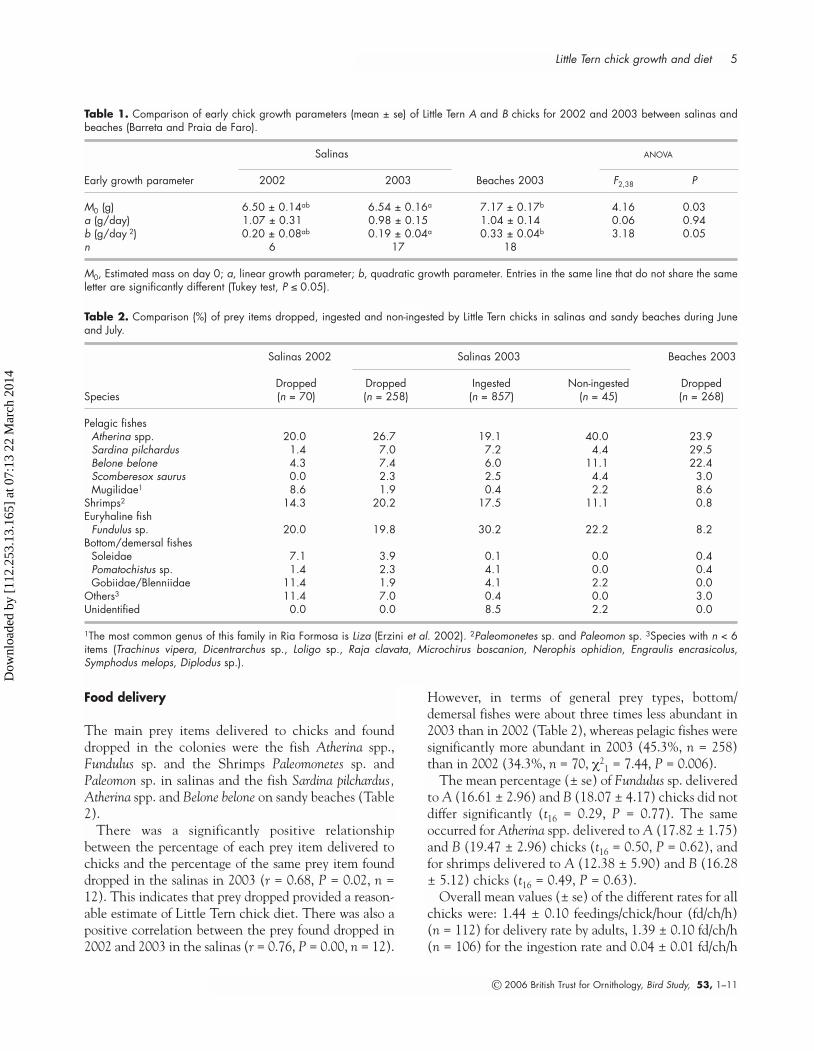
The growth curves of A, B and C chicks, in terms ofchick mass, wing-length and tarsus-length, are shownin Fig. 1. A chicks grew faster than B chicks but bothattained the same weight, tarsus-length and wing-length at 19 days, whereas C chicks reached the sameweight and wing-length two days later. There were significant differences in the estimated hatching mass(M0) and the quadratic growth parameter (b) of A andB chicks among the salinas in 2002, salinas in 2003 andbeaches in 2003 (Table 1). On the other hand, the linear growth parameter (a) did not differ significantlyamong these three categories (Table 1).
In 2003, both linear growth rate (LGR, mean ± se)and asymptotic mass (AM, mean ± se) did not differsignificantly between A (LGR: 2.53 ± 0.09 g/day, n =12; AM: 41.00 ± 1.48 g, n = 10) and B (LGR: 2.31 ±0.17 g/day, n = 10; AM: 39.48 ± 1.23 g, n = 6) chicks(LGR: t20 = 1.16, P = 0.26; AM: t14 = 0.71, P = 0.49).Only two out of eight C chicks survived in the fencednests from five days of age onwards (LGR: 2.28 ± 0.04g/day).
4 V.H. Paiva et al.
© 2006 British Trust for Ornithology, Bird Study, 53, 1–11
There was no apparent effect of fences on chickgrowth because the mean mass values of fenced chickswere similar to those of non-fenced chicks (Fig. 2).Although chick growth curves in terms of weight weresimilar between 2002 and 2003, variability was muchhigher in 2002 (Fig. 2).
Figure 1. Growth curves (mean ± se; the curves were adjusted bypolynomial regression) for Little Tern chicks (n = 32) in terms of: (a)mass; (b) tarsus-length; (c) wing-length. ��, A chicks, n = 14; �, Bchicks, n = 11; ��, C chicks, n = 2–7 (median = 2; after day 5 therewere only two C chicks).
Dow
nloa
ded
by [
112.
253.
13.1
65]
at 0
7:13
22
Mar
ch 2
014
Food delivery
The main prey items delivered to chicks and founddropped in the colonies were the fish Atherina spp.,Fundulus sp. and the Shrimps Paleomonetes sp. andPaleomon sp. in salinas and the fish Sardina pilchardus,Atherina spp. and Belone belone on sandy beaches (Table2).
There was a significantly positive relationshipbetween the percentage of each prey item delivered tochicks and the percentage of the same prey item founddropped in the salinas in 2003 (r = 0.68, P = 0.02, n =12). This indicates that prey dropped provided a reason-able estimate of Little Tern chick diet. There was also apositive correlation between the prey found dropped in2002 and 2003 in the salinas (r = 0.76, P = 0.00, n = 12).
© 2006 British Trust for Ornithology, Bird Study, 53, 1–11
Little Tern chick growth and diet 5
However, in terms of general prey types, bottom/demersal fishes were about three times less abundant in2003 than in 2002 (Table 2), whereas pelagic fishes weresignificantly more abundant in 2003 (45.3%, n = 258)than in 2002 (34.3%, n = 70, χ2
1 = 7.44, P = 0.006).The mean percentage (± se) of Fundulus sp. delivered
to A (16.61 ± 2.96) and B (18.07 ± 4.17) chicks did notdiffer significantly (t16 = 0.29, P = 0.77). The sameoccurred for Atherina spp. delivered to A (17.82 ± 1.75)and B (19.47 ± 2.96) chicks (t16 = 0.50, P = 0.62), andfor shrimps delivered to A (12.38 ± 5.90) and B (16.28± 5.12) chicks (t16 = 0.49, P = 0.63).
Overall mean values (± se) of the different rates for allchicks were: 1.44 ± 0.10 feedings/chick/hour (fd/ch/h)(n = 112) for delivery rate by adults, 1.39 ± 0.10 fd/ch/h(n = 106) for the ingestion rate and 0.04 ± 0.01 fd/ch/h
Table 1. Comparison of early chick growth parameters (mean ± se) of Little Tern A and B chicks for 2002 and 2003 between salinas andbeaches (Barreta and Praia de Faro).
Salinas ANOVA
Early growth parameter 2002 2003 Beaches 2003 F2,38 P
M0 (g) 6.50 ± 0.14ab 6.54 ± 0.16a 7.17 ± 0.17b 4.16 0.03a (g/day) 1.07 ± 0.31 0.98 ± 0.15 1.04 ± 0.14 0.06 0.94b (g/day 2) 0.20 ± 0.08ab 0.19 ± 0.04a 0.33 ± 0.04b 3.18 0.05n 6 17 18
M0, Estimated mass on day 0; a, linear growth parameter; b, quadratic growth parameter. Entries in the same line that do not share the sameletter are significantly different (Tukey test, P ≤ 0.05).
Table 2. Comparison (%) of prey items dropped, ingested and non-ingested by Little Tern chicks in salinas and sandy beaches during Juneand July.
Salinas 2002 Salinas 2003 Beaches 2003
Dropped Dropped Ingested Non-ingested DroppedSpecies (n = 70) (n = 258) (n = 857) (n = 45) (n = 268)
Pelagic fishesAtherina spp. 20.0 26.7 19.1 40.0 23.9Sardina pilchardus 1.4 7.0 7.2 4.4 29.5Belone belone 4.3 7.4 6.0 11.1 22.4Scomberesox saurus 0.0 2.3 2.5 4.4 3.0Mugilidae1 8.6 1.9 0.4 2.2 8.6
Shrimps2 14.3 20.2 17.5 11.1 0.8Euryhaline fish
Fundulus sp. 20.0 19.8 30.2 22.2 8.2Bottom/demersal fishes
Soleidae 7.1 3.9 0.1 0.0 0.4Pomatochistus sp. 1.4 2.3 4.1 0.0 0.4Gobiidae/Blenniidae 11.4 1.9 4.1 2.2 0.0
Others3 11.4 7.0 0.4 0.0 3.0Unidentified 0.0 0.0 8.5 2.2 0.0
1The most common genus of this family in Ria Formosa is Liza (Erzini et al. 2002). 2Paleomonetes sp. and Paleomon sp. 3Species with n < 6items (Trachinus vipera, Dicentrarchus sp., Loligo sp., Raja clavata, Microchirus boscanion, Nerophis ophidion, Engraulis encrasicolus,Symphodus melops, Diplodus sp.).
Dow
nloa
ded
by [
112.
253.
13.1
65]
at 0
7:13
22
Mar
ch 2
014
(n = 6) for the non-ingested prey rate items. The delivery and ingestion rates were very similar (Mann–Whitney: Z = 0.39, P = 0.70, n = 44). The mean rate offood delivered to A, B and C chicks of all ages was 1.38± 0.10 fd/h (n = 101), 1.26 ± 0.10 fd/h (n = 86) and 1.16± 0.24 fd/h (n = 27), respectively. There was no correla-tion between ingestion rate and chick age for A and Bchicks (rs = –0.26, P = 0.25, n = 22, Fig. 3) but meanlength of prey ingested by chicks increased significantlywith age (rs = 0.81, P = 0.0001, n = 22, Fig. 3).
Two environmental variables (wind speed and watersurface condition) were positively correlated in thisstudy (rs = 0.78, P = 0.0001, n = 20). Considering allprey items delivered to chicks, there was no significantdifference in the delivery rate among the four tidestates (Kruskal–Wallis: H3,170 = 3.75, P = 0.29) norwith different wind categories (H2,112 = 0.72, P = 0.70).
6 V.H. Paiva et al.
© 2006 British Trust for Ornithology, Bird Study, 53, 1–11
A two-way ANOVA on the effect of these two variableson the delivery rate showed no significant influence oftide (F3,170 = 0.15, P = 0.93), wind speed (F2,170 = 2.72,P = 0.07) and no interaction (F6,170 = 0.57, P = 0.75).
The mean length of prey delivered to chicksincreased significantly with chick age for Fundulus sp.(rs = 0.75, P = 0.0001, n = 21) and shrimps (rs = 0.67,P = 0.003, n = 18). Other prey species presented a positive but non-significant relationship: Atherina spp.(rs = 0.42, P = 0.06, n = 20), Gobiidae/Blenniidae (rs =0.16, P = 0.56, n = 15), Scomberesox saurus (rs = 0.46, P= 0.14, n = 12), Belone belone (rs = 0.34, P = 0.17, n =18), Sardina pilchardus (rs = 0.05, P = 0.86, n = 14).Pomatoschistus sp. (rs = –0.32, P = 0.24, n = 15) presented a negative but not significant correlation.
Effects of tidal phase and wind speed on the delivery of specific prey species
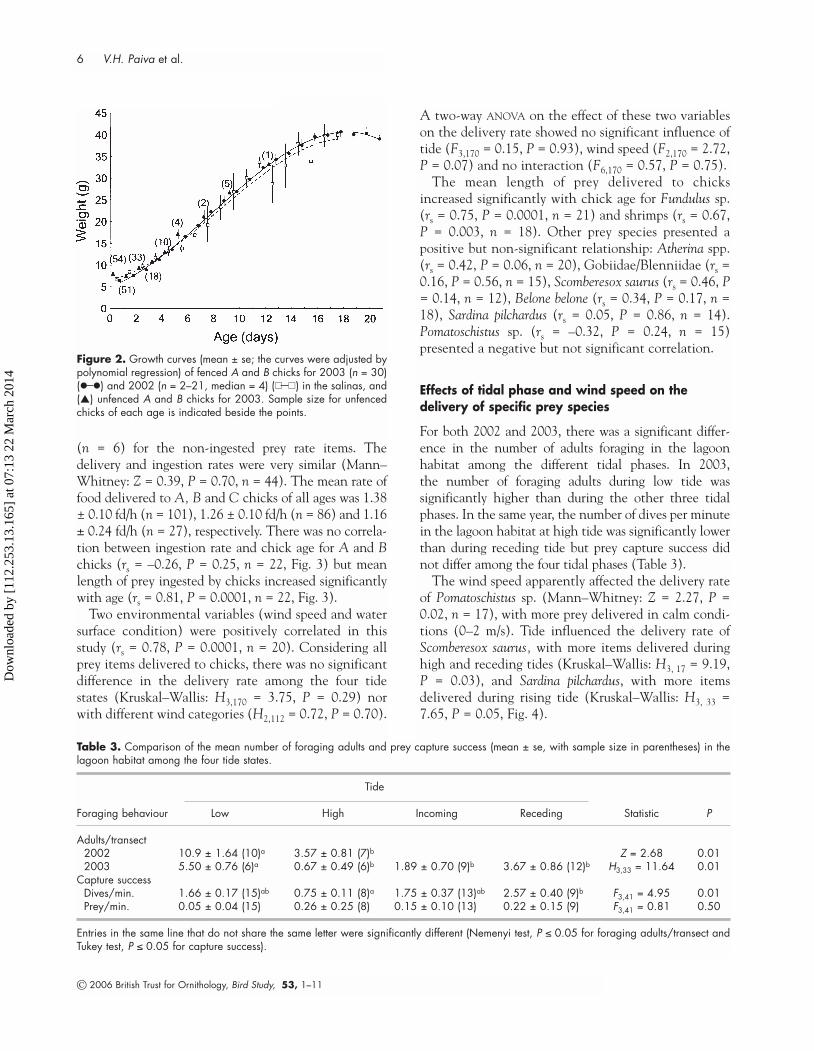
For both 2002 and 2003, there was a significant differ-ence in the number of adults foraging in the lagoonhabitat among the different tidal phases. In 2003, the number of foraging adults during low tide was significantly higher than during the other three tidalphases. In the same year, the number of dives per minutein the lagoon habitat at high tide was significantly lowerthan during receding tide but prey capture success didnot differ among the four tidal phases (Table 3).
The wind speed apparently affected the delivery rateof Pomatoschistus sp. (Mann–Whitney: Z = 2.27, P =0.02, n = 17), with more prey delivered in calm condi-tions (0–2 m/s). Tide influenced the delivery rate ofScomberesox saurus, with more items delivered duringhigh and receding tides (Kruskal–Wallis: H3, 17 = 9.19,P = 0.03), and Sardina pilchardus, with more itemsdelivered during rising tide (Kruskal–Wallis: H3, 33 =7.65, P = 0.05, Fig. 4).
Figure 2. Growth curves (mean ± se; the curves were adjusted bypolynomial regression) of fenced A and B chicks for 2003 (n = 30)(�—�) and 2002 (n = 2–21, median = 4) (��—��) in the salinas, and(�) unfenced A and B chicks for 2003. Sample size for unfencedchicks of each age is indicated beside the points.
Table 3. Comparison of the mean number of foraging adults and prey capture success (mean ± se, with sample size in parentheses) in thelagoon habitat among the four tide states.
Tide
Foraging behaviour Low High Incoming Receding Statistic P
Adults/transect2002 10.9 ± 1.64 (10)a 3.57 ± 0.81 (7)b Z = 2.68 0.012003 5.50 ± 0.76 (6)a 0.67 ± 0.49 (6)b 1.89 ± 0.70 (9)b 3.67 ± 0.86 (12)b H3,33 = 11.64 0.01
Capture successDives/min. 1.66 ± 0.17 (15)ab 0.75 ± 0.11 (8)a 1.75 ± 0.37 (13)ab 2.57 ± 0.40 (9)b F3,41 = 4.95 0.01Prey/min. 0.05 ± 0.04 (15) 0.26 ± 0.25 (8) 0.15 ± 0.10 (13) 0.22 ± 0.15 (9) F3,41 = 0.81 0.50
Entries in the same line that do not share the same letter were significantly different (Nemenyi test, P ≤ 0.05 for foraging adults/transect andTukey test, P ≤ 0.05 for capture success).
Dow
nloa
ded
by [
112.
253.
13.1
65]
at 0
7:13
22
Mar
ch 2
014

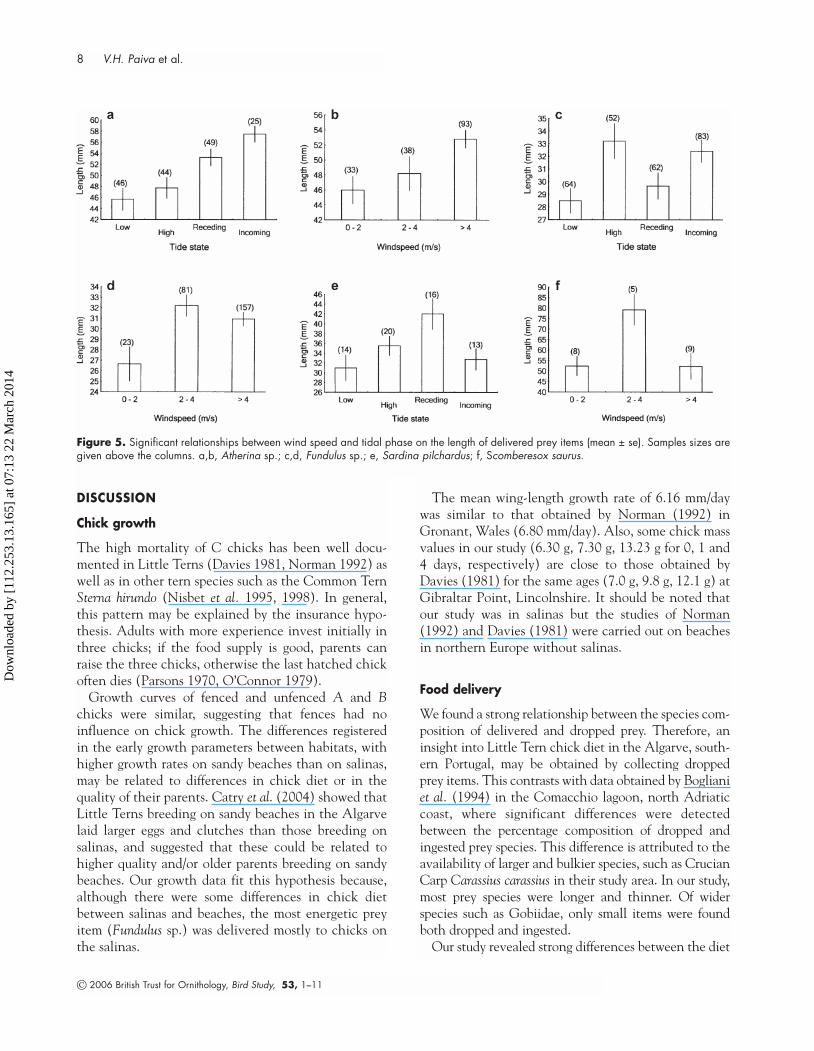
The size of Atherina spp., Fundulus sp. and Sardinapilchardus delivered to chicks showed significant differ-ences among tidal phases. Larger items were deliveredduring receding and incoming tides for Atherina spp.(Kruskal–Wallis: H3, 164 = 22.52, P = 0.001), high and
© 2006 British Trust for Ornithology, Bird Study, 53, 1–11
Little Tern chick growth and diet 7
incoming tides for Fundulus sp. (H3, 261 = 14.64, P =0.002), and receding tide for Sardina pilchardus (H3, 63 =9.68, P = 0.02). Larger Fundulus sp. were delivered during intermediate and high wind speed (2–4 and >4m/s; Kruskal–Wallis: H2, 261 = 10.08, P = 0.01). ForScomberesox saurus larger prey items were delivered during intermediate wind speed (2–4 m/s; Kruskal–Wallis: H2, 22 = 8.48, P = 0.01). In the case of Atherinaspp., larger items were delivered during higher windspeed (>4 m/s; Kruskal–Wallis: H2, 164 = 10.02, P = 0.01,Fig. 5).
Calorific content
Table 4 presents the energetic content of various prey items ingested by Little Tern chicks. Atherina spp. was significantly less calorific (F3, 33 = 9.71, P =0.001) than Belone belone, Fundulus sp. and Sardinapilchardus.
A and B chicks that received more than 30% ofFundulus sp. had a greater LGR (2.50 ± 0.15 g, n = 8)than those that were fed with more than 30% ofAtherina spp. (2.31 ± 0.11 g, n = 11), although this difference was not significant (t17 = 1.06, P = 0.19).There was also little difference in LGR between thechicks whose diet had more than 14% of shrimps (2.21± 0.16 g, n = 7) and those that had less than 5% ofshrimps (2.42 ± 0.14 g, n = 11; t16 = 0.96, P = 0.23).However, chicks that received more than 50% shrimps(less calorific prey) were fed at a higher rate (1.48 ±0.27 fd/ch/h, n = 44; this includes two chicks of onenest that received 53.1% of shrimps) than those thatreceived more than 50% Fundulus (more calorificitems) (0.77 ± 0.11 fd/ch/h, n = 45; see also Table 4).This suggests that parents were able to compensate forthe lower energetic value of shrimps.
Figure 3. Relationship between: (a) prey delivery rate to A and Bchicks (mean ± se) and chick age (n = 3–15, median = 9); and (b)prey length (mean ± se, n = 3–76, median = 35) and chick age.
Figure 4. Significant relationships between wind speed and tidal phase on the prey delivery rate (mean ± se). Samples sizes are given abovethe columns. a, Pomatoschistus sp.; b, Scomberesox saurus; c, Sardina pilchardus.
Dow
nloa
ded
by [
112.
253.
13.1
65]
at 0
7:13
22
Mar
ch 2
014
DISCUSSION
Chick growth
The high mortality of C chicks has been well docu-mented in Little Terns (Davies 1981, Norman 1992) aswell as in other tern species such as the Common TernSterna hirundo (Nisbet et al. 1995, 1998). In general,this pattern may be explained by the insurance hypo-thesis. Adults with more experience invest initially inthree chicks; if the food supply is good, parents canraise the three chicks, otherwise the last hatched chickoften dies (Parsons 1970, O’Connor 1979).
Growth curves of fenced and unfenced A and Bchicks were similar, suggesting that fences had no influence on chick growth. The differences registeredin the early growth parameters between habitats, withhigher growth rates on sandy beaches than on salinas,may be related to differences in chick diet or in thequality of their parents. Catry et al. (2004) showed thatLittle Terns breeding on sandy beaches in the Algarvelaid larger eggs and clutches than those breeding onsalinas, and suggested that these could be related tohigher quality and/or older parents breeding on sandybeaches. Our growth data fit this hypothesis because,although there were some differences in chick dietbetween salinas and beaches, the most energetic preyitem (Fundulus sp.) was delivered mostly to chicks onthe salinas.
8 V.H. Paiva et al.
© 2006 British Trust for Ornithology, Bird Study, 53, 1–11
The mean wing-length growth rate of 6.16 mm/daywas similar to that obtained by Norman (1992) inGronant, Wales (6.80 mm/day). Also, some chick massvalues in our study (6.30 g, 7.30 g, 13.23 g for 0, 1 and4 days, respectively) are close to those obtained byDavies (1981) for the same ages (7.0 g, 9.8 g, 12.1 g) atGibraltar Point, Lincolnshire. It should be noted thatour study was in salinas but the studies of Norman(1992) and Davies (1981) were carried out on beachesin northern Europe without salinas.
Food delivery
We found a strong relationship between the species com-position of delivered and dropped prey. Therefore, aninsight into Little Tern chick diet in the Algarve, south-ern Portugal, may be obtained by collecting droppedprey items. This contrasts with data obtained by Boglianiet al. (1994) in the Comacchio lagoon, north Adriaticcoast, where significant differences were detectedbetween the percentage composition of dropped andingested prey species. This difference is attributed to theavailability of larger and bulkier species, such as CrucianCarp Carassius carassius in their study area. In our study,most prey species were longer and thinner. Of widerspecies such as Gobiidae, only small items were foundboth dropped and ingested.
Our study revealed strong differences between the diet
Figure 5. Significant relationships between wind speed and tidal phase on the length of delivered prey items (mean ± se). Samples sizes aregiven above the columns. a,b, Atherina sp.; c,d, Fundulus sp.; e, Sardina pilchardus; f, Scomberesox saurus.
Dow
nloa
ded
by [
112.
253.
13.1
65]
at 0
7:13
22
Mar
ch 2
014

of chicks in salinas (mainly Fundulus sp. and shrimp bothin 2002 and 2003) and beaches (Sardina pilchardus andBelone belone in 2003), although Atherina spp. wereimportant in both habitats. Fundulus sp., a main item ofchicks in the salinas, is presumably captured in channelsand salinas near the Santa Luzia breeding coloniesbecause this is a euryhaline species not found in the mainlagoon or the sea (Erzini et al. 2002). Another interest-ing aspect is that Fundulus sp. was not reported in thediet of chicks in 1989 by Calado (1989) at two differentsalinas of Ria Formosa. Presumably this fish species is arecent colonist in the area.
The diet of A and B chicks was similar, therefore thedifference in growth rates between them was related todifferences in their ingestion rates. The delivery ratewas also related to the calorific content of prey, becausechicks that received a higher percentage of shrimpswere fed at a faster rate than those that received ahigher percentage of fish. Similar observations werereported by Davies (1981) and Norman (1992) forLittle Terns in Great Britain. The mean delivery rate ofprey to Little Tern chicks in our study was similar tothat obtained by Davies (1981) for a nest of threechicks at Gibraltar Point, Lincolnshire (0.90 fd/ch/h).In our study, mean delivery rate was not correlated with
© 2006 British Trust for Ornithology, Bird Study, 53, 1–11
Little Tern chick growth and diet 9
mean age of chicks. On the other hand, there was anincrease in mean length of prey delivered to Little Ternchicks as they grew. This suggests that either prey wereincreasing in size or parents selected larger prey itemsfor older chicks. Our study gives more support to thesecond hypothesis because there was a steady increasein the length of fish delivered as chicks grew.Therefore, parents met the increasing energy demandof their growing chicks by adjusting prey size, ratherthan prey frequency. Similar results were found byBogliani et al. (1994) for Little Terns and Stienen et al.(2000) for Sandwich Terns Sterna sandvicensis. Sinceyoung chicks cannot swallow larger prey, the avail-ability of smaller prey could determine the timing andsuccess of breeding (Safina et al. 1988).
Massias & Becker (1990) argued that shrimps are analternative energetic resource for Common Tern chicksin periods of absence of more energy-rich species. Ourstudy agrees with this because chicks fed mostly onshrimps, which had the lowest calorific content, werefed at a higher rate. We also observed shrimps beingcaptured in salinas 10–50 m away from the Santa Luziabreeding colonies, which should contribute to loweringthe energy expenditure in foraging by adults breedingin salinas.
Table 4. Energy content, length and delivery rate (mean ± se, with n in parentheses) of prey items ingested by Little Tern chicks in the sali-nas of Santa Luzia, Algarve.
Delivery rate
Species Energy (kJ/g dry wt) Length (mm) (feeds/nest/hour) (feeds/chick/hour)
Pelagic fishesAtherina spp. 18.48 ± 0.61 (22) 50.37 ± 0.95 (164) 0.91 ± 0.09 (75) 0.46 ± 0.04 (75)Sardina pilchardus 20.48 ± 0.56 (8) 35.60 ± 1.32 (63) 0.58 ± 0.08 (29) 0.32 ± 0.05 (29)Belone belone 20.32 ± 0.21 (11) 83.66 ± 1.97 (52) 0.46 ± 0.07 (34) 0.30 ± 0.04 (34)Scomberosox saurus 19.22 (1) 58.44 ± 4.13 (22) 0.53 ± 0.10 (16) 0.30 ± 0.07 (16)Mugilidae 20.35 (1)
Euryhaline fishFundulus sp. 21.95 ± 0.56 (8) 30.96 ± 0.53 (261) 1.29 ± 0.13 (72) 0.66 ± 0.08 (72)
Bottom/demersal fishes1 32.60 ± 1.58 (36) 0.38 ± 0.05 (21) 0.20 ± 0.03 (21)Parablennius galerita 21.53 (1)Dicentrarchus punctatus 19.49 ± 0.09 (2)Coryphoblennius gallerita 22.28 (1)Diplodus vulgaris 20.26 (1)Symphodus melops 20.08 (1)Pomatochistus rispidus 19.26 ± 0.12 (3) 26.76 ± 0.86 (33) 0.64 ± 0.19 (17) 0.45 ± 0.17 (17)
SquidLoligo sp. 23.61 (1)
Shrimps 25.61 ± 0.33 (150) 1.03 ± 0.14 (34) 0.71 ± 0.13 (34)Paleomonetes varians 17.38 (1)Paleomon serratus 18.71 (1)
1Length and delivery rate refer only to items of the families Blenniidae and Gobiidae.
Dow
nloa
ded
by [
112.
253.
13.1
65]
at 0
7:13
22
Mar
ch 2
014

Effects of tidal phase and wind speed on the delivery of specific prey species
Although a greater number of adults were observed foraging in the lagoon habitat at low tide (both in 2002and 2003), tide had no influence on prey delivery rateto chicks. This could be explained if adults use low tide to fulfil their daily energy requirements. However,adults dived more during incoming and receding tides,presumably when most prey are moving in the watercolumn. We suspect that low tide constrained theavailability of estuarine prey species, as suggested by thelow capture success that we registered during this tidalphase. Stienen et al. (2000) observed also that the success of Sandwich Terns foraging in the Wadden Seawas low during low tide. On the contrary, salinas andchannels around the colonies should be more con-sistent in prey availability, because they are notsubjected to tidal influences. A salina is composed ofseveral tanks separated by soil banks that protectagainst strong winds, thus providing a calmer water surface and good visibility for diving. This may explainthe delivery of larger Fundulus sp. individuals duringhigh and incoming tides (when fewer adults are foraging in the lagoon habitat) and during adverse foraging conditions in the lagoon (Brenninkmeijer etal. 2002) and in the sea, such as during windier periods(Stienen et al. 2000).
Atherina is a resident lagoon species (Muzavor et al.1993, Erzini et al. 2002), and normally the older (andlarger) individuals live at greater depths. When tidevaries between high and low, they move in the watercolumn and become more accessible to Little Ternadults. This could explain the delivery of larger itemsduring the incoming and receding tidal phases.Atherina spp. may also be more active during periods ofgreater wind speed, because the mixing of sea-surfacewaters may influence the cycle of nutrients in the watercolumn. Pomatoschistus sp. is a benthic resident lagoonspecies (Muzavor et al. 1993, Erzini et al. 2002), whichexplains why their delivery rate was higher in calmconditions. For Scomberesox saurus, a marine species(Whitehead et al. 1984), larger prey items were delivered during intermediate wind speed periods,which are considered to be optimal conditions for ternforaging in the sea (Cramp 1985, Stienen et al. 2000).On the other hand, there was a greater delivery rate ofthis prey species during high and receding tidal phases,when we observed fewer adults foraging in the lagoonhabitat.
In summary, Little Terns in the Algarve appear to
10 V.H. Paiva et al.
© 2006 British Trust for Ornithology, Bird Study, 53, 1–11
rely on several species for rearing chicks. Although onespecies, Atherina sp., was the main prey species in both2002 and 2003, there were important variations in preyspecies between habitats as well as in the importance ofpelagic and bottom/demersal fishes between 2002 and2003. The availability of prey species for the ternsdepends also on environmental conditions such aswind speed and tide that affect the availability of cer-tain prey and/or the fishing success of the parents. Thehigher energy content of Fundulus sp. and the abundance of shrimps on salinas may be important criteria for Little Terns to breed there. Nevertheless,Little Tern parents experienced no difficulties in providing sufficient food to rear two chicks in both salinas and sandy beaches in the Algarve. Disturbance,predation and flooding (Catry et al. 2004) are usuallymore important in explaining breeding failure thanfood availability.
ACKNOWLEDGEMENTS
This study is a contribution from the project POCTI/BSE/37385/2001 ‘Breeding and foraging ecology of LittleTerns to provide ecological indicators in estuarine environ-ments’ financed by the Fundação Portuguesa para a Ciênciae Tecnologia and POCTI, co-funded by the EU FEDER pro-gramme. We thank Nuno Grade of the Ria Formosa NaturalPark for logistic support, Ricardo Lopes, John Bowler and two anonymous referees for valuable comments on the manuscript.
ENDNOTE
a. Md = M0 + ad + bd2, where Md = chick mass on day d, M0= estimated chick mass on day 0, d = age in days, a = lineargrowth parameter, the instantaneous slope of the growthcurve on day 0, and b = quadratic growth parameter (a measure of the curvature of the growth curve; Nisbet et al.1998). M0, a and b were estimated for each fenced andunfenced chick for which mass was measured on day 0 and atleast two other days
REFERENCES
Allcorn, R., Eaton, M.A., Cranswick, P.A., Perrow, M., Hall,C., Smith, L., Reid, J., Webb, A., Smith, K.W.S., Langston,R.H.W. & Ratcliffe, N. 2003. A Pilot Study of Breeding Tern For-aging Ranges in NW England and East Anglia in Relation toPotential Development Areas for Offshore Windfarms. RSPB/WWT/JNCC, Sandy.
Anthony, J., Roby, D. & Turco, K. 2000. Lipid content and energydensity of forage fishes from the northern Gulf of Alaska. J. Exp.Marine Biol. Ecol. 249: 53–78.
Dow
nloa
ded
by [
112.
253.
13.1
65]
at 0
7:13
22
Mar
ch 2
014

© 2006 British Trust for Ornithology, Bird Study, 53, 1–11
Little Tern chick growth and diet 11
Bogliani, G., Fasola, M., Canova, L. & Saino, N. 1994. Preyselection by parents and chicks of the Little Tern Sterna albifrons.Avocetta 18: 9–11.
Brenninkmeijer, A. Stienen, E., Klaassen, M. & Kersten, M.2002. Feeding ecology of wintering terns in Guinea-Bissau. Ibis144: 602–613.
Calado, M. 1998. Sterna albifrons no Parque Natural da Ria Formosa: alguns aspectos da ecologia alimentar. Airo 9: 53.
Catry, T., Ramos, J.A., Catry, I., Allen-revez, M. & Grade, N.2004. Are salinas a suitable alternative breeding habitat for LittleTerns? Ibis 146: 247–257.
Cramp, S. (eds) 1985. The Birds of the Western Paleartic, Vol. 4.Oxford University Press, Oxford.
Dahdul & Horn 2003. Energy allocation and postnatal growth in captive Elegant Tern (Sterna elegans) chicks: responses to high – versus low – energy diets. Auk 120: 1069–1081.
Davies, S. 1981. Development and behaviour of Little Tern chicks. Br.Birds 74: 291–298.
Dunn, E. 1972. Changes in fishing ability of terns associated withwind speed and sea surface conditions. Nature 244: 520–521
Erzini, K., Bentes, L., Coelho, R., Correia, C., Lino, P., Mon-teiro, P., Ribeiro, J., Gonçalves, J. 2002. Recruitment of SeaBreams (Sparidae) and Other Commercially Important Species in theAlgarve (Southern Portugal). Final Report to the Commission of theEuropean Communities, Faro.
Fasola, M. & Bogliani, G. 1990. Foraging ranges of an assem-blage of Mediterranean seabirds. Colon. Waterbirds 13: 72–74.
Frank, D. 1992. The influence of feeding conditions and food provi-sioning of chicks in Common Terns Sterna hirundo nesting in theGerman Wadden Sea. Ardea 80: 45–55.
Fraschetti, S., Capelli, R., Robertson, M., Albertelli, G. &Eleftheriou, A. 1994. Calorimetry: use of the Phillipsonmicrobomb. Oebalia 20: 117–127.
Henken, A., Lucas, H., Tijssen, P. & Machiels, M. 1986. A comparison between methods used to determine the energy contentof feed, fish and faeces samples. Aquaculture 58: 195–201.
Hislop, J., Harris, M. & Smith, J. 1991. Variation in the calorific value
and total energy content of the lesser sandeel (Ammodytes marinus) andother fish preyed on by seabirds. J. Zool. (Lond.) 224: 501–517.
Massias, A. & Becker, P.H. 1990. Nutritive value of food andgrowth in Common Tern Sterna hirundo chicks. Ornis Scand. 21:187–194.
Monaghan, P., Uttley, J. D., Burns, M. D., Thaine, C. & Black-wood, J. 1989. The relationship between food supply,reproductive effort and breeding success in Artic terns Sterna Par-adisaea. J. Anim. Ecol. 58: 261–274.
Muzavor, S., Arruda, L. & Andrade, J. 1993. Roteiro Ecológicoda Ria Formosa II – Peixes. Algarve em foco, Faro.
Nisbet I., Spendelow, J. & Hatfield, J. 1995. Variations in growthof Roseate Tern chicks. Condor 97: 335–344.
Nisbet, I., Hatfield, J., Zingo, J. & Gough, G. 1998. Variationsin growth of Roseate Tern chicks: II. Early growth as an index ofparental quality. Condor 100: 305–315.
Norman, D. 1992. The growth rate of Little Tern Sterna albifronschicks. Ring. Migr. 13: 98–102.
O’Connor, R.J. 1979. Egg weights and brood reduction in the Euro-pean swift (Apus apus). Condor 81: 133–145.
Parsons, J. 1970. Relationship between egg size and post-hatchingchick mortality in the herring gull (Larus argentatus). Nature 228:1221–1222.
Pearson, T. 1968. The feeding biology of sea-bird species breedingon the Farne Islands, Northumberland. J. Anim. Ecol. 37: 521–552.
Safina, C., Burger, J., Gochfeld, M. & Wagner, R. 1988. Evi-dence of prey limitation of Common tern and Roseate ternreproduction. Condor 90: 852–859.
Sobral, D. & Gomes, J. 1998. Peixes Litorais – Estuário do Sado.Instituto de Conservação da Natureza, Lisboa.
Stienen, E., Beers, P., Brenninkmeijer, A., Habraken, J.,Raaijmakers, M. & Van Tienen, P. 2000. Reflections of a specialist: Patterns in food provisioning and foraging conditions inSandwich Terns Sterna sandvicensis. Ardea 88: 33–49.
Whitehead, P., Bauchot, M., Mureau, J., Nielsen, J. & Tor-tonese, E. 1984. Fishes of the North-eastern Atlantic and theMediterranean, Vols. 1–3. UNESCO, Paris.
(MS received 2 July 2004; revised MS accepted 19 October 2004)
Dow
nloa
ded
by [
112.
253.
13.1
65]
at 0
7:13
22
Mar
ch 2
014
Copyright © 2022 FDOKUMEN




















