
Regional trends for bud burst and flowering of woody plants in Norway as related to climate change

Antagonistic effects of FGF4 on BMP induction of apoptosis andchondrogenesis in the chick limb bud
R.A. Buckland*, J.M. Collinson1, E. Graham, D.R. Davidson, R.E. Hill
MRC Human Genetics Unit, Western General Hospital, Crewe Road, Edinburgh EH4 2XU, Scotland, UK
Received 3 November 1997; revised version received 5 January 1998; accepted 5 January 1998
Abstract
In an effort to define the roles of bone morphogenic proteins (BMPs) and fibroblast growth factors (FGFs) during chick limb developmentmore closely, we have implanted beads impregnated with these growth factors into chick limb buds between stages 20 and 26. Embryoswere sacrificed at the time the bone chondrocyte condensations first appear (stages 27–28). Implantation of beads containing BMPs at theearlier stages (20–22) caused apoptosis to occur, in the most severe cases leading to complete limb degeneration. Application of FGF4,either in the same, or in a different bead, prevented the BMP-induced apoptosis. We argue that the apoptosis observed on removal of theAER prior to stage 23 of development could be brought about by BMPs. The action of epithelial FGF in preventing BMP-mediatedapoptosis in the mesenchyme would define a novel aspect of epithelial-mesenchymal interactions. Implanting the BMP4 beads into the coreof the limb bud a day later (stages 25–26) caused intense chondrogenesis rather than apoptosis. FGF4 could again nullify this effect and byitself caused a reduction in bone size. This is the reverse of the functional relationship these growth factors have in mouse toothspecification (where it is BMP4 that inhibits the FGF8 function), and suggests that the balance between the effects of FGFs and BMPscould control the size of the chondrocyte precursor cell pool. In this way members of these two growth factor families could control the sizeof appendages when they are initially formed. 1998 Elsevier Science Ireland Ltd.
Keywords:Limb bud; BMPs; FGFs; Apoptosis; Chondrogenesis; Bone
1. Introduction
Signalling centres have been identified which consist ofan epithelium and adjacent mesenchyme whose interactiondirects the local development of embryonic structures. Onesuch centre is the distal tip of the growing vertebrate limb.This is bounded distally by the apical ectodermal ridge(AER), the removal of which results in limb truncations(Saunders, 1948). However, replacement of the AER withacrylic beads which have been pre-soaked in fibroblastgrowth factor 4 (FGF4) apically and on the posterior marginof the bud results in normal outgrowth (Niswander et al.,1993). The main function of the AER is therefore to controllimb outgrowth by producing FGF4 (and other FGFs, e.g.Crossley et al., 1996). The cells immediately beneath the
AER constitute the progress zone, a region of uncommittedprogenitor cells with a higher mitotic index than more prox-imal cells (Hornbruch and Wolpert, 1970; Summerbell etal., 1973). The zone is maintained by the AER, and as out-growth occurs the cells on the proximal margin move awayfrom the influence of the AER. It is on leaving the progresszone that cells are thought to read their positional values andbecome committed (although initially not irreversibly) to aparticular cell fate (Tickle et al., 1975).
Cells in the progress zone initially synthesise bone mor-phogenic proteins (BMP2 and BMP4). These are membersof the TGFb family of growth factors, which were originallyidentified as chondrogenic/osteoblastic agents from adultcattle bones that could induce bone at ectopic sites in rats(Urist, 1965; Wozney et al., 1988). Mutations causing trun-cations to theBmp5andGdf5genes (the latter a closeBmprelative) have bone phenotypes (Kingsley et al., 1992; Stormet al., 1994), showing that bone formation is a main functionof these particular family members during embryogenesis.
Mechanisms of Development 71 (1998) 143–150
0925-4773/98/$19.00 1998 Elsevier Science Ireland Ltd. All rights reservedPII S0925-4773(98)00008-2
* Corresponding author. Tel. +44 0131 3322471; fax: +44 0131 3432620.1 Present address: Department of Pre-clinical Veterinary Sciences, Royal
(Dick) Veterinary School, Summerfield, Edinburgh, Scotland, UK.
However, gene-targetedBmp4 homozygous null mutantmice have a variable phenotype, the most severely affectedembryos degenerating without forming mesoderm (Winnieret al., 1995) demonstrating that BMP4 has functions otherthan specifying bone (for review see Hogan, 1996).
The Bmp4genes are initially transcribed in the cells ofstage 16–17 chick wing fields, in both the ectoderm andmesenchyme, and expression continues in the whole ofthe limb bud until stages 18–19, whereupon it is graduallylost from the centre of the mesenchyme of the developingbud. By stage 24 very much lower levels of transcripts arepresent directly beneath the AER, and by stage 25 levels
along the posterior margin of the limb bud are also reduced(Francis et al., 1994).Bmp2, however, is only expressed inthe posterior mesenchyme, and is more prominentlyexpressed in the ectoderm, and for longer in the AER thanBmp4(Francis et al., 1994).
In the early chick limb bud, BMP2 (but not BMP4) hasbeen shown to have an inhibitory effect on the mitogenicactivity of FGF4 (Niswander and Martin, 1993). At a similarstage of development (stages 20–22) BMP2-producing cellsimplanted under the anterior AER were partially able toreproduce the properties of Shh and the zone of polarisingactivity (ZPA) (Duprez et al., 1996a), suggesting that BMPs
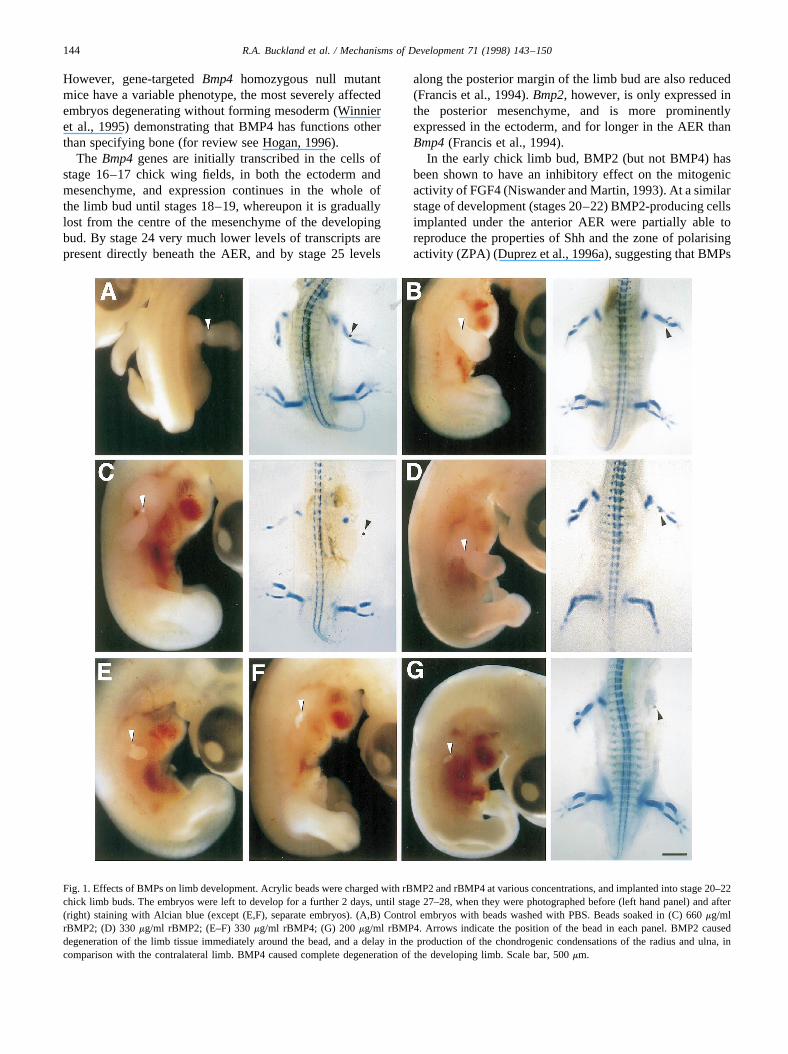
Fig. 1. Effects of BMPs on limb development. Acrylic beads were charged with rBMP2 and rBMP4 at various concentrations, and implanted into stage 20–22chick limb buds. The embryos were left to develop for a further 2 days, until stage 27–28, when they were photographed before (left hand panel) and after(right) staining with Alcian blue (except (E,F), separate embryos). (A,B) Control embryos with beads washed with PBS. Beads soaked in (C) 660mg/mlrBMP2; (D) 330mg/ml rBMP2; (E–F) 330mg/ml rBMP4; (G) 200mg/ml rBMP4. Arrows indicate the position of the bead in each panel. BMP2 causeddegeneration of the limb tissue immediately around the bead, and a delay in the production of the chondrogenic condensations of the radius and ulna, incomparison with the contralateral limb. BMP4 caused complete degeneration of the developing limb. Scale bar, 500mm.
144 R.A. Buckland et al. / Mechanisms of Development 71 (1998) 143–150

produced in the ZPA might be involved in pattern forma-tion. At slightly later stages (stages 25–26) BMP2 andBMP4 promote chondrogenesis (Duprez et al., 1996b).We wished to assess more closely the role of BMPs at theearly stages of limb development, and define the possibleinteraction with FGF4. To this end acrylic beads werecharged with the different growth factors and implantedinto stage 20–26 chick limb buds. We show that there is ashort window, between stages 20 and 22 when ectopic appli-cation of BMPs to the core of the limb bud causes apoptosis.A day later (stages 25–26) BMP4 promotes chondrogenesis.Both the apoptotic and chondrogenic activities of BMP4 canbe inhibited by FGF4, demonstrating the requirement for thecorrect balance between these growth factors to be main-tained, both spatially and temporally, for normal limb devel-opment.
2. Results
2.1. Application of rBMPs causes limb abnormalities
After bead implantation at stages 20–22, embryos were
left to develop for 2 days (stages 27–28) before photogra-phy and Alcian blue staining of chondrogenic condensa-tions. The operation to implant control beads (containingphosphate-buffered saline (PBS) only) had no harmfuleffect on the developing limb (Fig. 1A,B). Beads containing660 mg/ml recombinant human BMP2 (rBMP2), however,caused severe degeneration of the tissue around the bead,leading to a constriction at this point in an almost normal-ised limb (Fig. 1C). There was no chondrogenesis beyondthe shoulder girdle condensation in the treated limb bud atthis stage, resulting in a gap between shoulder girdle andbead (Fig. 1C). At half this concentration, rBMP2 caused areduction in the size of the humerus (Fig. 1D) in five out offive cases showing that, at these doses, the effect was con-centration dependent.
Application of rBMP4 (330mg/ml) resulted in completedegeneration of the developing limb (Fig. 1E,F). It thereforehad a greater effect than BMP2 (at least with the particularpreparations we were using). This degree of degenerationcould also be achieved at a dosage level of 200mg/ml (Fig.1G) and so this concentration was used in all subsequentexperiments. Usually no chondrogenesis was observed, butoccasionally (two out of twelve) shoulder girdle condensa-
Fig. 2. Cell death in BMP4-treated limb buds. Beads charged with (A) PBS, (B) 200mg/ml rBMP4, (C) 400mg/ml rFGF4 and 200mg/ml rBMP4, wereimplanted into stage 20–22 chick limb buds and fixed 24 h later (stages 25–26). Limbs treated with BMP4 beads were approximately half the size of thecontrol limb at this time. Transverse sections cut through the level where the beads were located were subjected to the TUNEL assay for DNA degradation.Left hand panels, haematoxylin/eosin stained sections and FITC-stained dead cells; scale bar, 500mm. Right hand panels,×2 enlargements of FITC-stainedlimb buds. The apoptosis triggered by rBMP4 application was inhibited by rFGF4 at a dose twice that of the rBMP4.
145R.A. Buckland et al. / Mechanisms of Development 71 (1998) 143–150

tions developed. The limb bud continued to grow after beadimplantation, but after one day post-implantation, degenera-tion had started. The degree of apoptosis was assessed usingthe terminal transferase assay (TUNEL) (Gavrieli et al.,1992). The beads without growth factors (Fig. 2A) did notcause any additional cell death, whereas beads soaked in200mg/ml rBMP4 caused considerable cell death through-out the limb bud and some of the adjacent tissue (Fig. 2B).
2.2. rFGF4 can rescue normal development in limb budstreated with rBMP4
Bmp4mRNA is produced by most of the mesenchyme ofthe stage 18–19 chick limb bud (Francis et al., 1994).Assuming that this is successfully translated and that thenecessary receptors are present to transduce the signal,there must be some inhibition of the apoptotic activity ofthe BMP4 protein in this mesenchyme. As FGF4 can replacethe AER in stages 20–22 limb buds in producing normaloutgrowth, and it (along with FGF8) overlies theBmp2and
Bmp4expression domains in the progress zone, FGF4 wastested for such an activity. Beads soaked in a 2:1 ratio ofrFGF4:rBMP4 (400:200mg/ml) were able to prevent apop-tosis from occurring (Fig. 2C) and to rescue normal devel-opment (Fig. 3A and Table 1). The duplicate exampleshown in Fig. 3B is classed as ‘normal’ despite havingonly one chondrogenic condensation in the forearm, asdigit condensations are clearly being laid down. In threecases, rescue was incomplete, with evidence of humeruscondensations but no radius, ulna or autopod condensations(Table 1).
If these growth factors were delivered in separate beads,the results were much more variable, with only five out ofthe twelve rFGF4 beads capable of rescuing the condition(Fig. 3C,D), five of partial rescue, and two similar to thepositive controls, that is, no limb structures having devel-oped beyond a shoulder girdle condensation (Table 1).Therefore rescue was still possible when the factors weredelivered separately, and had overlapping, but not identicaldistributions.
Fig. 3. Rescue of BMP4-mediated degeneration by FGF4. Each row contains duplicate embryos (A,B.) each with a bead containing 400mg/ml rFGF4 and 200mg/ml rBMP4, (C,D) with the same concentration growth factors administered in separate beads, (E,F) with a single bead containing 200mg/ml of bothgrowth factors, implanted at stages 20–22. Two days later (stages 27–28) the embryos were fixed and stained with Alcian blue. A similar degree of outgrowthwas achieved with a high dose of FGF4 as in the control limbs (left hand side). Lower concentrations were not sufficient to achieve this. Scale bar, 1000mm.
146 R.A. Buckland et al. / Mechanisms of Development 71 (1998) 143–150

If, as these results suggest, FGF4 is alleviating thedeleterious effect of BMP4, then it should be possible totitrate out this activity of FGF4 by simply reducing theamount of rFGF4 loaded into the beads. When beadscontaining a 1:1 ratio (200:200mg/ml) of the two growthfactors were implanted in limb buds, the abnormalities pro-duced were similar to the severest effect seen with rBMP2(Fig. 1B), that is, a normal-sized limb, but a constriction ofthe tissue around the bead, with very little (or no) chondro-genesis beyond the shoulder girdle condensation (Fig.3E,F).
2.3. rBMP4 causes chondrogenesis in 5-day chick limbbuds
Because BMPs promoted apoptosis in the early limb bud,we were interested to determine the duration of the windowwhen BMPs have deleterious effects on limb development.Beads loaded with 200mg/ml rBMP4 were implanted in thelimb buds of 5-day chick embryos (stages 25–26), and leftfor a further day (stages 27–28) before staining with Alcianblue. Under these conditions, that is, implanting the beadjust one day after BMPs disrupt limb development, rBMP4caused intense chondrogenesis to occur in six out of sixcases (Fig. 4A,B). The chondrogenesis was usually, butnot always, around the bead, which merely reflects the cen-tral location within the limb bud of most of the beads.
2.4. rFGF4 inhibits chondrogenesis
Beads loaded with a 2:1 ratio of rFGF4 and rBMP4(400:200mg/ml, respectively) and implanted into the coreof stage 25–26 wing buds, generally caused little change inlimb morphology (three out of five cases; Fig. 4E) althoughin some embryos there was evidence of either extra orreduced chondrogenesis (e.g. Fig. 4F). Beads containingjust rFGF4 caused a reduction in chondrogenesis in thearea where the bead had been implanted in six out ofeight cases (Fig. 4C,D), indicating FGF4 can block cellsfrom undergoing chondrogenesis.
3. Discussion
The fact that FGF4 and BMP2 have antagonistic activitieshas been shown previously, where BMP2 was shown toinhibit the effect of FGF4 on proliferation of mesenchymecells in a mouse limb bud assay (Niswander and Martin,1993). These findings have recently been extended toshow that BMP2 and BMP4 can inhibit the ability ofFGFs to inducePax9 expression in the mandibular andmaxillary arches, which determines the position of forma-tion of individual teeth (Neu¨buser et al., 1997). Here weshow the reverse, namely, that FGF4 can inhibit the abilityof BMP4 to induce apoptosis and chondrogenesis in thechick limb bud.
3.1. BMP4 and apoptosis during limb bud development
Bmpsare expressed in the anterior and posterior necroticzones of the stage 23–25 chick wing bud (Francis et al.,1994), and as they have been shown to promote apoptosisof neural crest cells in rhombomeres 3 and 5 of the stage 9chick hindbrain (Graham et al., 1994), they have also beenimplicated in the cell death that occurs in the necrotic zonesof the limb buds. Here, we show that ectopic application ofBMP2 and BMP4 to the core of stage 20–22 chick limbbuds can cause severe apoptosis, leading to complete degen-eration of the limb bud. The doses of growth factors usedhere are similar to those used in other in vivo bead implantexperiments, such as in the application of the FGF4 in theAER replacement experiments of Niswander et al. (1993)and in the dose of BMPs required to achieve expressionlevels similar to those of the endogenousMsx genes onectopic application to cranial mesenchyme (Barlow andFrancis-West, 1997).
Apoptosis also occurs in the interdigital regions of thehand- and footplates, and both the direct application ofBMP4 (Ganan et al., 1996), and dominant negative muta-tions introduced intoBmpr-1Aand Bmpr-1B indicate thatBMPs are instrumental in causing apoptosis in the interdi-gital regions (Zou and Niswander, 1996; Yokouchi et al.,1996).
The sequence of limb development in the chick has beenestablished by sequential removal of the AER (Saunders,1948; Summerbell et al., 1973). Until stage 23 this initiallyresults in a certain degree of apoptosis in the progress zone(Rowe et al., 1982). We have shown here that the transitionperiod between the early action of BMP4 (apoptosis) andthe late function (chondrogenesis) is between stages 23 and24. At this time also, expression ofBmp4is ceasing at thedistal tip of the progress zone (Francis et al., 1994). As AERfunction can be performed by exogenously-applied FGF4,withdrawal from the effects of FGF4 has, until now, seemedthe most likely explanation for the apoptosis resulting fromAER removal. Our experiments, however, suggest thatBMPs in the mesenchyme are responsible for this, andthat their action is normally suppressed in the progress
Table 1
Frequency of limb abnormalities produced by different combinations ofgrowth factors applied to stage 20–22 chick embryo limb buds
Limb category
Growth factorsloaded (mg/ml)
Normal limb Abnormal limba Amelia
BMP4 (200) 0 2 10FGF4/BMP4
(400:200)9 3 0
FGF4/BMP4(400:200) intwo beads
5 5 2
aGenerally, the presence of shoulder girdle and humerus only, unless statedotherwise in the text.
147R.A. Buckland et al. / Mechanisms of Development 71 (1998) 143–150

zone by FGF4, or by other fibroblast growth factors that areexpressed during limb bud development (Munaim et al.,1988; Crossley et al., 1996; Ohuchi et al., 1997). Thisnovel aspect of epithelial-mesenchymal interactions mayapply to other signalling centres during development.
3.2. Induction of chondrogenesis
Implanting the BMP4 beads a day later, between stages25 and 26, caused intense chondrogenesis. Once again thiseffect was inhibited by FGF4. Stasis is achieved at a 2:1ratio of FGF4 to BMP4, which is a 1:1 ratio in terms ofmolecules. This time the FGF4 appears to be directly inhi-biting chondrogenesis. This data is consistent with the phe-notype of theFgfr-3 targeted null mutation in mice (Colvinet al., 1996; Deng et al., 1996) where mutant embryos have
enlarged long bones of the limbs and large vertebrae. Theprimary defect of these mice is an increase in chondrocyteproduction. This indicates that FGFs have a role in nega-tively regulating bone size during normal development.Confirmation of this comes from the finding that achondro-plasia, the most common form of dwarfism, is the result ofdominant negative mutations ofFgfr-3 (Shiang et al., 1994).The lack of an embryo phenotype may be a result of func-tional overlap between FGF receptors, because although thethree FGF receptors that affect bone growth have reason-ably distinct expression patterns within developing bone,with a major part of the expression ofFgfr-1 being in theperiostium, that ofFgfr-2 in the hypertrophic cartilage, andthat ofFgfr-3 in the resting cartilage, there is considerableoverlap of expression in the chondrocyte condensations ofthe early limb bud (Orr-Urtreger et al., 1991; Peters et al.,
Fig. 4. The effects on chondrogenesis of BMP4 and FGF4 when applied to limb buds of 5-day-old (stages 25–26) embryos. Beads containing (A,B) 200mg/ml rBMP4, (C,D) 400mg/ml rFGF4, (E,F) 400mg/ml rFGF4 and 200mg/ml rBMP4 were implanted into the limb buds of stages 25–26 chick embryos.The following day (stages 27–28) embryos were fixed and stained with Alcian blue. While BMP4 prompted cartilage formation, FGF4 prevented it. Amixture of these growth factors in a ratio of 2:1 allowed normal development. Scale bar, 1000mm.
148 R.A. Buckland et al. / Mechanisms of Development 71 (1998) 143–150

1992, 1993). The ectopic application of rFGF4 describedhere, being the equivalent of a localised over-expressionregime, clearly shows that FGF4 can act negatively on car-tilage formation at the earliest stages of bone formation.
The BMP type I receptors have distinct but also overlap-ping functions in bone formation, both being involved incartilage production, butBmpr-1B appearing to be moreinvolved in initiating the chondrocyte lineage, and theBmpr-1Ain its differentiation (Zou et al., 1997). The recep-tors of both families of growth factor are therefore presentand functional in the developing cartilage rudiments, andcould act antagonistically during this process to control thesize of the chondrocyte progenitor cell pool and hence thesize of limb bones in proportion to other structures in thebody of the early embryo.
4. Experimental procedures
4.1. Chick embryos
Fertile eggs were supplied by the Roslin Institute,(Roslin, Midlothian, UK) and were incubated at 37°C in ahumidified incubator, with daily turning, for the timerequired to produce the desired stage (Hamburger andHamilton, 1951) of development.
4.2. Loading beads
Heparin acrylic beads (Sigma, H5263) were washed inseveral changes of Dulbecco A solution (PBS) then trans-ferred to a 2ml drop of growth factor solution (Niswander etal., 1993). All human recombinant growth factors were gen-erously supplied by the Genetics Institute, (Boston, MA)and when necessary were diluted with PBS/0.1% bovineserum albumin (BSA) immediately prior to use. Beadswere embedded into the mesenchyme of limb buds throughslits that had been scored with tungsten dissection needles.Egg windows were re-sealed with sellotape and the eggsreturned to the incubator for the desired length of time.
4.3. Alcian blue staining
This was carried out essentially according to Shum et al.(1993). Here, embryos were fixed and stained simulta-neously in a solution of ethanol:glacial acetic acid (4:1)containing 0.15mg/ml Alcian blue powder. After 24–48 hthe embryos were gradually hydrated through a series ofethanol washes and cleared in 0.1 M potassium hydroxide.They were stored in 25% ethanol and 50% glycerol.
4.4. TUNEL reactions
DNA fragmentation was assayed using terminal transfer-ase (Gavrieli et al., 1992). 6-mm thick tissue sections werecut from paraffin wax blocks using standard techniques.
After xylene washes to remove the wax, and rehydration,TUNEL reactions were performed according to Mu¨ller et al.(1995) using Boehringer (Mannheim, Germany) transferaseenzyme and FlTC-dUTP.
Acknowledgements
We are enormously grateful to Drs. V. Rosen and J. Woz-ney for their assistance in supplying the growth factors usedin this study, and to Professor Cheryll Tickle for advice onbead implantation. We thank Paul Perry for taking the fluor-escent TUNEL pictures, Norman Davidson, Sandy Bruceand Douglas Stuart for their expert assembling of the fig-ures, and Pauline McDonald and Audrey Wardrope forsecretarial assistance.
References
Barlow, A.J., Francis-West, P.H., 1997. Ectopic application of recombi-nant BMP-2 and BMP-4 can change patterning of developing chickfacial primordia. Development 124, 391–398.
Colvin, J.S., Bohne, B.A., Harding, G.W., McEwen, D.G., Ornitz, D.M.,1996. Skeletal overgrowth and deafness in mice lacking fibroblastgrowth factor receptor 3. Nature Genet. 12, 390–397.
Crossley, P.H., Minowada, G., MacArthur, C.A., Martin, G.R., 1996.Roles for FGF8 in the induction, initiation and maintenance of chicklimb development. Cell 84, 127–136.
Deng, C., Wynshaw-Boris, A., Zhou, F., Kuo, A., Leder, P., 1996. Fibro-blast growth factor receptor 3 is a negative regulator of bone growth.Cell 84, 911–921.
Duprez, D., Kostakopoulou, K., Francis-West, P., Tickle, C. and Brickell,P., 1996a. Activation of Fgf-4 and Hox D gene expression by BMP-2expressing cells in the developing chick limb. Development 122, 1821–1828.
Duprez, D., Bell, E.J., Richardson, M.K., Archer, C.W., Wolpert, L.,Brickell, P.M. and Francis-West, P.H., 1996b. Overexpression ofBMP-2 and BMP-4 alters the size and shape of developing skeletalelements in the chick limb. Mech. Dev. 57, 145–157.
Francis, P.H., Richardson, M.K., Brickell, P.M., Tickle, C., 1994. Bonemorphogenetic proteins and a signalling pathway that controls pattern-ing in the developing chick limb. Development 120, 209–218.
Ganan, Y., Macias, D., Duterque-Coquillaud, M., Ros, M.A., Hurle, J.M.,1996. Role of TGFbs and BMPs as signals controlling the position ofthe digits and the areas of interdigital cell death in the developing chicklimb autopod. Development 122, 2349–2357.
Gavrieli, Y., Sherman, Y., Ben-Sasson, S.A., 1992. Identification of pro-grammed cell death in situ via specific labelling of nuclear DNAfragmentation. J. Cell Biol. 119, 493–501.
Graham, A., Francis-West, P., Brickell, P., Lumsden, A., 1994. The sig-nalling molecule BMP4 mediates apoptosis in the rhombencephalicneural crest. Nature 372, 684–686.
Hamburger, V., Hamilton, H.L., 1951. A series of normal stages in thedevelopment of the chick embryo. J. Morph. 88, 49–92.
Hogan, B.L.M., 1996. Bone morphogenic proteins: multifunctional regu-lators of vertebrate development. Genes Dev. 10, 1580–1594.
Hornbruch, A., Wolpert, L., 1970. Cell division in the early growth andmorphogenesis of the chick limb. Nature 226, 764–766.
Kingsley, D.M., Bland, A.E., Grubber, J.M., Marker, P.C., Russell, L.B.,Copeland, N.G., Jenkins, N.A., 1992. The mouseshort ear skeletalmorphogenesis locus is associated with defects in a bone morphogeneticmember of the TGFb superfamily. Cell 71, 399–410.
149R.A. Buckland et al. / Mechanisms of Development 71 (1998) 143–150

Muller, C., Kowenz-Leutz, E., Grieser-Ade, S., Graf, T., Leutz, A., 1995.NF-M (chicken C/EBPb) induces eosinophilic differentiation and apop-tosis in a hematopoietic progenitor cell line. EMBO J. 14, 6127–6135.
Munaim, S.I., Klagsbrun, M., Toole, B.P., 1988. Developmental changesin fibroblast growth factor in the chicken embryo limb bud. Proc. Natl.Acad. Sci. USA 85, 8091–8093.
Neubuser, A., Peters, H., Balling, R., Martin, G.R., 1997. Antagonisticinteractions between FGF and BMP signalling pathways: a mechanismfor positioning the sites of tooth formation. Cell 90, 247–255.
Niswander, L., Tickle, C., Vogel, A., Booth, I., Martin, G.R., 1993. FGF-4replaces the apical ectodermal ridge and directs outgrowth and pattern-ing of the limb. Cell 75, 579–587.
Niswander, L., Martin, G.R., 1993. FGF-4 and BMP-2 have oppositeeffects on limb growth. Nature 361, 68–71.
Ohuchi, H., Nakagawa, T., Yamamoto, A., Araga, A., Ohata, T., Ishimaru,Y., Yoshioka, H. Kuwana, T., Nohno, T. Yamasaki, M., Itoh, N., Noji,S., 1997. The mesenchymal factor, FGF10, initiates and maintains theoutgrowth of the chick limb bud through interaction with FGF8, anapical ectodermal factor. Development 124, 2235–2244.
Orr-Urtreger, A., Givol, D., Yayon, A., Yarden, Y., Lonai, P., 1991.Developmental expression of two murine fibroblast growth factor recep-tors,fig andbek. Development 113, 1419–1434.
Peters, K., Werner, S., Chen, G., Williams, L.T., 1992. Two FGF receptorgenes are differentially expressed in epithelial and mensenchymal tis-sues during limb formation and organogenesis in the mouse.Development 114, 233–243.
Peters, K., Ornitz, D., Werner, S., Williams, L., 1993. Unique expressionpattern of the FGF receptor 3 gene during mouse organogenesis. Dev.Biol. 155, 423–430.
Rowe, D.A., Cairns, J.M., Fallon, J.F., 1982. Spatial and temporal patternsof cell death in limb bud mesoderm after apical ectodermal ridgeremoval. Dev. Biol. 93, 83–91.
Saunders, J.W., 1948. The proximo-distal sequence of origin of parts of thechick wing and the role of the ectoderm. J. Exp. Biol. 108, 363–403.
Shiang, R., Thompson, L.M., Zhu, Y.-Z., Church, D.M., Fielder, T.J.,Bocian, M., Winokur, S.T., Wasmuth, J.J., 1994. Mutations in the trans-membrane domain of FGFR3 cause the most common genetic form ofdwarfism, achondroplasia. Cell 78, 335–342.
Shum, L., Sakakura, Y., Bringas, P., Luo, W., Snead, M.L., Mayo, M.,Crohln, C., Millar, S. Werb, Z., Buckley, S., Hall, F.L., Warburton, D.Slavkin, H.C., 1993. EGF abrogation-inducedfusilli-form dysmorpho-genesis of Meckel’s cartilage during embryonic mouse mandibular mor-phogenesis in vitro. Development 118, 903–917.
Storm, E.E., Huynh, T.V., Copeland, N.G., Jenkins, N.A., Kingsley, D.M.,Lee, S.J., 1994. Limb alterations in brachpodism mice due to mutationsin a new member of the TGFb-superfamily. Nature 368, 639–642.
Summerbell, D., Lewis, J.H., Wolpert, L., 1973. Positional information inchick limb morphogenesis. Nature 244, 492–496.
Tickle, C., Summerbell, D., Wolpert, L., 1975. Positional signalling andspecification of digits in chick limb morphogenesis. Nature 254, 199–202.
Urist, M.R., 1965. Bone: Formation by autoinduction. Science 150, 893–899.
Winnier, G., Blessing, M., Labosky, P.A., Hogan, B.L.M., 1995. Bonemorphogenetic protein-4 is required for mesoderm formation and pat-terning in the mouse. Genes Dev. 9, 2105–2116.
Wozney, J.M., Rosen, V., Celeste, A.J., Mitsock, L.M., Whitters, M.J.,Kriz, R.W., Hewick, R.M., Wang, E.A., 1988. Novel regulators of boneformation: molecular clones and activities. Science 242, 1528–1534.
Yokouchi, Y., Sakiyama, J.I., Kameda, T., Iba, H., Suzuki, A., Ueno, N.,Kuroiwa, A., 1996. BMP-2/-4 mediate programmed cell death inchicken limb buds. Development 122, 3725–3734.
Zou, H., Niswander, L., 1996. Requirement for BMP signalling in inter-digital apoptosis and scale formation. Science 272, 738–741.
Zou, H., Wieseg, Z.R., Massagub, J., Niswander, L., 1997. Distinct roles oftype I bone morphogenic protein receptors in the formation and differ-entiation of cartilage. Genes Dev. 11, 2191–2203.
150 R.A. Buckland et al. / Mechanisms of Development 71 (1998) 143–150
Copyright © 2022 FDOKUMEN




















