
Synthesis, crystal structure, nuclease and in vitro antitumor activities of a new mononuclear...
10
Synthesis, crystal structure, nuclease and in vitro antitumor activities of a new mononuclear copper(II) complex containing a tripodal N 3 O ligand Christiane Fernandes a , Gabrieli L. Parrilha a , Josane A. Lessa a , Laura J.M. Santiago b , Milton M. Kanashiro c , Fabrizio S. Boniolo d , Adailton J. Bortoluzzi e , Ney V. Vugman f , Marcelo H. Herbst f , Adolfo Horn Jr. a, * a Laborato ´rio de Cie ˆncias Quı ´micas, Universidade Estadual do Norte Fluminense, 28013-602, Campos dos Goytacazes – RJ, Brazil b Laborato ´rio de Biodiversidade e Biotecnologia, Universidade Federal do Estado do Rio de Janeiro, 22290-240, Rio de Janeiro – RJ, Brazil c Laborato ´ rio de Biologia do Reconhecer, Universidade Estadual do Norte Fluminense, 28013-602, Campos dos Goytacazes – RJ, Brazil d Laborato ´rio de Biotecnologia, Universidade Estadual do Norte Fluminense, 28013-602, Campos dos Goytacazes – RJ, Brazil e Departamento de Quı ´mica, Universidade Federal de Santa Catarina, 88040-900, Floriano ´ polis – SC, Brazil f Departamento de Fı ´sica dos So ´ lidos, Universidade Federal do Rio de Janeiro, 21945-970, Rio de Janeiro – RJ, Brazil Received 8 October 2005; accepted 17 April 2006 Available online 22 April 2006 Abstract We present here the synthesis, crystal structure, electrochemical behavior, spectroscopic properties (FT-IR, UV–Vis and EPR), nucle- ase and in vitro antitumor activities against human myeloid leukemia cell line of the mononuclear copper complex [Cu(HPCl- NOL)(Cl)]Cl MeOH (1). The reaction of the tetradentate ligand HPClNOL [1-(bis-pyridin-2-ylmethyl-amino)-3-chloropropan-2-ol] and 1 equiv. of [Cu(OH 2 ) 6 ](Cl) 2 , in methanol, resulted in 1, which crystallizes as blue monoclinic crystals. The complex is pentacoordi- nated with a distorted square-pyramidal geometry. The activity of complex 1 toward plasmid DNA and THP-1 carcinogenic cells was investigated. Complex 1 promotes the cleavage of supercoiled DNA (pBlueScript KS + DNA) to nicked circular and linear DNA forms. In addition to the three typical KS + DNA forms, the cleavage resulted in a fourth band, which was visualized above of the nicked cir- cular form. The results reveal that the cleavage mechanism is radical-independent. Furthermore, complex 1 is able to promote cell death of THP-1 cells by apoptosis, as confirmed by fluorescent microscopy, cell morphology and DNA degradation. Ó 2006 Elsevier B.V. All rights reserved. Keywords: Mononuclear copper(II) complex; Nuclease activity; In vitro antitumor activity; Apoptosis; Lewis acidity; THP-1 cell 1. Introduction Copper is considered an essential trace element and its concentration as free metal ion inside cells should be consid- ered lower than 10 15 M (calculated)/10 12 M (observed), since concentrations higher than 10 9 M in the cytoplasm can be poisonous [1]. The free copper concentration in the cytoplasm is regulated by different mechanisms, including pumps, exchangers, and by proteins expression, which use copper in their active sites [2]. The great majority of the cop- per proteins are involved mainly in oxidation/reduction reactions as well as in the dioxygen transport and activation [3–5]. The development of mimic systems for copper metal- loenzymes has provided important compounds which have allowed scientists to understand both the physical–chemi- cal properties of the active site of the copper enzymes as well as the reactivity exhibited by the metalloenzymes, since some of these mimetic compounds show similar activities to that of natural metalloenzymes [6–8]. Actually the mimic compounds have shown importance not only to 0020-1693/$ - see front matter Ó 2006 Elsevier B.V. All rights reserved. doi:10.1016/j.ica.2006.04.007 * Corresponding author. Tel./fax: +55 22 2726 1519. E-mail address: [email protected] (A. Horn Jr.). www.elsevier.com/locate/ica Inorganica Chimica Acta 359 (2006) 3167–3176
Transcript of Synthesis, crystal structure, nuclease and in vitro antitumor activities of a new mononuclear...

www.elsevier.com/locate/ica
Inorganica Chimica Acta 359 (2006) 3167–3176
Synthesis, crystal structure, nuclease and in vitro antitumoractivities of a new mononuclear copper(II) complex containing a
tripodal N3O ligand
Christiane Fernandes a, Gabrieli L. Parrilha a, Josane A. Lessa a, Laura J.M. Santiago b,Milton M. Kanashiro c, Fabrizio S. Boniolo d, Adailton J. Bortoluzzi e, Ney V. Vugman f,
Marcelo H. Herbst f, Adolfo Horn Jr. a,*
a Laboratorio de Ciencias Quımicas, Universidade Estadual do Norte Fluminense, 28013-602, Campos dos Goytacazes – RJ, Brazilb Laboratorio de Biodiversidade e Biotecnologia, Universidade Federal do Estado do Rio de Janeiro, 22290-240, Rio de Janeiro – RJ, Brazil
c Laboratorio de Biologia do Reconhecer, Universidade Estadual do Norte Fluminense, 28013-602, Campos dos Goytacazes – RJ, Brazild Laboratorio de Biotecnologia, Universidade Estadual do Norte Fluminense, 28013-602, Campos dos Goytacazes – RJ, Brazil
e Departamento de Quımica, Universidade Federal de Santa Catarina, 88040-900, Florianopolis – SC, Brazilf Departamento de Fısica dos Solidos, Universidade Federal do Rio de Janeiro, 21945-970, Rio de Janeiro – RJ, Brazil
Received 8 October 2005; accepted 17 April 2006Available online 22 April 2006
Abstract
We present here the synthesis, crystal structure, electrochemical behavior, spectroscopic properties (FT-IR, UV–Vis and EPR), nucle-ase and in vitro antitumor activities against human myeloid leukemia cell line of the mononuclear copper complex [Cu(HPCl-NOL)(Cl)]Cl Æ MeOH (1). The reaction of the tetradentate ligand HPClNOL [1-(bis-pyridin-2-ylmethyl-amino)-3-chloropropan-2-ol]and 1 equiv. of [Cu(OH2)6](Cl)2, in methanol, resulted in 1, which crystallizes as blue monoclinic crystals. The complex is pentacoordi-nated with a distorted square-pyramidal geometry. The activity of complex 1 toward plasmid DNA and THP-1 carcinogenic cells wasinvestigated. Complex 1 promotes the cleavage of supercoiled DNA (pBlueScript KS+ DNA) to nicked circular and linear DNA forms.In addition to the three typical KS+ DNA forms, the cleavage resulted in a fourth band, which was visualized above of the nicked cir-cular form. The results reveal that the cleavage mechanism is radical-independent. Furthermore, complex 1 is able to promote cell deathof THP-1 cells by apoptosis, as confirmed by fluorescent microscopy, cell morphology and DNA degradation.� 2006 Elsevier B.V. All rights reserved.
Keywords: Mononuclear copper(II) complex; Nuclease activity; In vitro antitumor activity; Apoptosis; Lewis acidity; THP-1 cell
1. Introduction
Copper is considered an essential trace element and itsconcentration as free metal ion inside cells should be consid-ered lower than 10�15 M (calculated)/10�12 M (observed),since concentrations higher than 10�9 M in the cytoplasmcan be poisonous [1]. The free copper concentration in thecytoplasm is regulated by different mechanisms, includingpumps, exchangers, and by proteins expression, which use
0020-1693/$ - see front matter � 2006 Elsevier B.V. All rights reserved.
doi:10.1016/j.ica.2006.04.007
* Corresponding author. Tel./fax: +55 22 2726 1519.E-mail address: [email protected] (A. Horn Jr.).
copper in their active sites [2]. The great majority of the cop-per proteins are involved mainly in oxidation/reductionreactions as well as in the dioxygen transport and activation[3–5].
The development of mimic systems for copper metal-loenzymes has provided important compounds which haveallowed scientists to understand both the physical–chemi-cal properties of the active site of the copper enzymes aswell as the reactivity exhibited by the metalloenzymes, sincesome of these mimetic compounds show similar activitiesto that of natural metalloenzymes [6–8]. Actually themimic compounds have shown importance not only to

3168 C. Fernandes et al. / Inorganica Chimica Acta 359 (2006) 3167–3176
understand the processes of natural biomolecules; theiractions can also be extrapolated to systems other thanthose of natural occurrence. For example, the lack of nat-ural copper nucleases does not invalidate the use of coppercompounds for mechanistic studies, drug design or as bio-technological tools whose actions are based on DNA cleav-age. Really, copper has proved to be a valuable metalcenter in the development of artificial nuclease systems asreported in recent reviews [9–13]. Thus, copper syntheticcompounds have been investigated on their ability to pro-mote nucleic acid cleavage as well as to act as metallophar-maco, receiving special attention to their actions on theDNA [14–16].
Nature has already found ways to hydrolyze DNAwithin 1 second, using metal-containing enzymes such asP1 and EcoRV nucleases [17] reducing the low velocityassociated with the spontaneous nucleic acids cleavage[9]. The main obstacle in DNA hydrolysis is the large neg-ative charge of its polymeric structure that inhibits theattack of nucleophiles on the DNA backbone. Thus,charge neutralization is one of several mechanisms usedby natural nuclease molecules [10]. Therefore, a large num-ber of nucleases have metal ions in their active sites, whichare electrically attracted by the phosphate backbone andcan generate nucleophilic species at mild neutral pH range[18]. There are two main types of nucleases: the exonuc-leases, which hydrolyze only the outermost phosphodiesterbond of their substrate, thus removing one base at a timefrom either 5 0 or the 3 0 terminus, and the endonucleases,which hydrolyze interior phosphodiester bonds, therebycausing rapid changes in the length of their structure [19].
In the specific case of synthetic copper nucleases models,two main processes associated to DNA cleavage have beenestablished: (i) oxidative and (ii) hydrolytic. In the oxida-tive mechanism, the copper specie changes between CuII/CuI, promoting the formation of radical species that canpromote the oxidative break of DNA strands [20–27]. Inthe hydrolytic pathway, the copper atom shows ability togenerate nucleophilic species at almost neutral pH, whichare able to attack the phosphate group of the DNAstrands, promoting their cleavage [9,28–34]. It is importantto point out that all the natural nucleases act through ahydrolytic mechanism [19].
Compounds that present ability in promoting the DNAcleavage near physiological conditions are suitable candi-dates to be investigated both in vitro and in vivo againstcancer cells. The development of cisplatin as one of themost successful drugs for the treatment of testicular andovarian tumours has brought metal-based anticancer drugsinto focus [35]. This and the second generation platinumdrug, carboplatin, have paved the way for screening a num-ber of metal-based drugs for their antineoplastic activities.In addition to its well-characterized effects on the DNA ofproliferating cells, cisplatin is now known to kill cells byapoptosis [36]. Apoptosis is a tightly controlled process inwhich the cell death is executed through the activation ofspecific signaling pathways. Although the biochemical
and molecular factors regulating it are complex and notcompletely defined, important regulatory mechanismsinclude death receptor, caspases, mitochondria andtumor-suppressor genes [37]. Cells that are eliminatedthrough an apoptotic mechanism undergo quick phagocy-tosis process. They do not release their cytosolic content toextracellular space, avoiding, thus, an inflammatory pro-cess [38,39].
Therefore, as part of our investigation into coordinationchemistry and into metal complexes as chemical nucleases[40,41], we report herein in-depth, physical and chemicalcharacterization of a new mononuclear copper(II) complexsynthesized with the ligand 1-(bis-pyridin-2-ylmethyl-amino)-3-chloropropan-2-ol (HPClNOL) [42], its interac-tion with DNA and initial studies of its interaction withTHP-1 human myeloid leukemia cells.
2. Experimental
All the reagents and solvents were commercially avail-able of high purity and used as such. Plasmid pBlueScriptII KS(+) (pB II KS+) was purchased from StratageneCloning Systems. The DNA was transformed into XL1Blue bacterial cell lines by heating shock treatment, andthe transformed bacteria was cultured in medium contain-ing ampicillin 60 lg/mL. The DNA was harvested andpurified according to the Concert Rapid Plasmid MiniprepSystem Kit (Gibco-BRL). The DNA was further purifiedvia ethanol precipitation and dissolved in an appropriatevolume of sterile water. The purity and concentration ofthe DNA were determined by comparison with differentconcentrations of standard Lambda DNA (Pharmacia-Biotech) in agarose gel electrophoresis and UV–Vis spec-troscopy by determining the ratio of absorbance at260 nm and at 280 nm [28]. Solutions of the metal complexand other reagents for strand scission experiments wereprepared fresh daily. All other chemicals were used aspurchased.
The human myeloid leukemia cell line (THP-1)was grown in DMEM-F12 medium (Gibco-BRL)supplemented with 10% fetal calf serum (Gibco-BRL),50 lM 2-mercaptoethanol and 20 lg/mL gentamicin andmaintained in an incubator at 37 �C with humidified envi-ronment and 5% CO2. THP-1 cells were found to be nega-tive for mycoplasma contamination according to Hoechststaining.
2.1. Synthesis of [Cu(HPClNOL)Cl]Cl Æ CH3OH (1)
The ligand HPClNOL (1-(bis-pyridin-2-ylmethyl-amino)-3-chloropropan-2-ol) was prepared through the reactionbetween BMPA (bis-(2-pyridylmethyl) amine) and epichlo-rohydrin, as reported previously [42]. The complex[Cu(HPClNOL)Cl]Cl Æ CH3OH (1) was obtained by the reac-tion of a methanolic solution (10 mL) of HPClNOL(1 mmol, 0.29 g) and a methanolic solution of [Cu(OH2)6]Cl2(1 mmol, 0.2 g), resulting in a greenish-blue solution. After

Table 1Crystal data and structure refinement for complex 1
Empirical formula C16H22Cl3CuN3O2
Formula weight 458.26Temperature (K) 193(2)Wavelength (A) 0.71069Crystal system, space group monoclinic, C2/cUnit cell dimensions
a (A) 20.531(3)b (A) 7.940(1)c (A) 25.494(5)b (�) 113.01(1)
Volume (A3) 3825.3(11)Z, calculated density (g/cm3) 8, 1.591Absorption coefficient (mm�1) 1.576F(000) 1880Crystal size (mm) 0.40 · 0.33 · 0.20h Range for data collection (�) 2.79–25.07Index ranges 0 6 h 6 24, 0 6 k 6 9,
�30 6 l 6 27Reflections collected/unique [Rint] 3494/3395 [0.0776]Absorption correction w-scanMaximum and minimum transmission 0.9481 and 0.7498Refinement method full-matrix least-
squares on F2
Data/restraints/parameters 3395/0/238Goodness-of-fit on F2 1.067Final R indices [I > 2r(I)] R1 = 0.0470, wR2 = 0.1299R indices (all data) R1 = 0.0584, wR2 = 0.1368Extinction coefficient 0.0003(3)Largest difference in peak and hole (e A�3) 0.970 and �0.839
Table 2Selected bond lengths (A) and angles (�) for complex 1
Cu1–N12 1.994(3)Cu1–N22 2.003(3)Cu1–N1 2.048(3)Cu1–Cl1 2.246(1)Cu1–O1 2.386(3)
N12–Cu1–N22 162.89(14)N12–Cu1–N1 83.04(13)N22–Cu1–N1 81.55(12)N12–Cu1–Cl1 98.25(11)N22–Cu1–Cl1 96.97(9)N1–Cu1–Cl1 177.99(8)N12–Cu1–O1 86.04(11)N22–Cu1–O1 98.61(11)N1–Cu1–O1 80.33(10)Cl1–Cu1–O1 101.28(7)C2–O1–Cu1 103.48(19)
C. Fernandes et al. / Inorganica Chimica Acta 359 (2006) 3167–3176 3169
one week, blue crystals were formed, which were filtered off.Yield: 67%. Anal. Calc. for CuC16H22N3O2Cl3: C, 41.6; H,4.7, N, 9.3. Found: C, 41.9; H, 4.8; N, 9.2%. KM (CH3CN):137 X�1 cm2 mol�1 (1:1 electrolyte type) [43].
2.2. Physical measurements
Elemental analysis was performed on a Perkin Elmer2400 CHN analyzer. Infrared spectrum was recorded inCsI disks on a Nicolet-Magna IR 760 instrument and theelectronic absorption spectra (200–1100 nm range) wererecorded in CH3CN with a Shimadzu 1601 PC UV–Visspectrophotometer. The EPR studies were carried outusing a Bruker ESP 380E/FT-CW spectrometer operatingin X-Band (9.5 GHz) and 100 kHz frequency modulation.Cyclic voltammograms (CVs) were carried out with anAutolab PGSTAT 10 potentiostat/galvanostat in acetoni-trile containing 0.1 M tetrabutylammonium perchlorate(TBAClO4) as supporting electrolyte under argon atmo-sphere at room temperature. The electrochemical cellemployed was a standard three-electrode configuration: aglassy carbon working electrode, a platinum-wire auxiliaryelectrode and a commercial Ag/AgCl electrode immersedin a salt bridge containing 0.1 M TBAClO4. The formalpotential of the ferrocenium/ferrocene couple was 0.426 Vversus the reference electrode Ag/AgCl, being establishedas 0.400 V versus NHE [44]. The electrical conductivityof 1 · 10�3 M solution of 1 in acetonitrile was measuredwith a Biocristal conductometer.
2.3. Crystal structure determination
From the crystalline sample of 1, a prismatic blue crystalwas selected for the crystallographic analysis. The intensitydata was collected with an Enraf–Nonius CAD4 diffrac-tometer, at room temperature, with graphite-monochro-mated Mo Ka radiation. The unit cell parameters weredetermined on the setting angles of 25 centered reflections.All data were corrected for Lorentz and polarization effects[45]. Empirical absorption correction based on the azi-muthal scans of seven appropriate reflections was alsoapplied over all collected intensities with PLATON program[46,47]. The structure was solved by direct methods andrefined by full-matrix least-squares methods using SIR97[48] and SHELXL97 [49] programs, respectively. All non-hydrogen atoms were refined anisotropically. The hydro-gen atom of the coordinated alcohol group was localizedfrom Fourier map and treated with riding model, whereasH atom of the methanol solvent was not found. H atomsattached to carbon atoms were placed at their idealizedpositions, with C–H distances and Ueq values taken fromthe default settings of the refinement program. The figuresof the molecular structures were produced with the ORTEP3program [50]. Further crystal structure and refinement datafor complex 1 are summarized in Table 1. Bond distancesand angles relevant to the copper coordination sphere arelisted in Table 2.
2.4. Nuclease activity of [Cu(HPClNOL)Cl]Cl Æ CH3OH
(1)
2.4.1. Anaerobic cleavage of pB II KS+ DNA by complex 1The reaction conditions followed the procedure
described previously [40,41]. Solutions containing 1 lg pBII KS+ DNA were incubated with 100, 200, 500, 1000and 2000 lM of complex 1 solutions, at 37 �C, for 24 h,in Tris–HCl buffer pH 7.7 (according to the Sigma Catalog(p. 1039, 1999) the buffer solution is pH 8.0 at 25 �C and

3170 C. Fernandes et al. / Inorganica Chimica Acta 359 (2006) 3167–3176
becomes pH 7.7 at 37 �C). For the control samples thecomplex 1 was omitted. The DNA pellets were resuspendedin sterile water and analyzed by gel electrophoresis.
2.4.2. Linearization of pB II KS+ DNA with EcoRI
endonucleaseThe pB II KS+ DNA was digested by EcoRI endonucle-
ase. The reaction was performed with 1 lg pB II KS+ DNAand 5U EcoRI (Promega) in digestion buffer consisting of500 mM Tris (pH 7.71 at 37 �C), 100 mM MgCl2, 50 mMNaCl, 25 mM spermidine (3HCl) for 4 h. The reactionwas stopped by incubation at 75 �C for 10 min and DNAwas stored at 20 �C until the experiments with the complex1.
2.4.3. DNA religation
The pB II KS+ DNA religation reactions were per-formed with 1.3 lL (100 ng) of pB II KS+ DNA and1.0 lL (1U) of T4 DNA ligase (Promega), in a total volumeof 20 lL of ligation buffer consisting of 50 mM Tris–HCl(pH 7.6), 10 mM MgCl2, 1 mM ATP, 1 mM DTT, 5%(w/v) polyethylene glycol-8000, at 4 �C for 24 h. Religationwas stopped by incubation at 85 �C for 10 min and theDNA was analyzed by gel electrophoresis.
2.4.4. Product analysis and quantification
The extension of cleavage was determined via densito-metric analysis of ethidium bromide-stained agarose gels.Electrophoreses were performed by using a Horizon11.14 horizontal gel electrophoresis apparatus from LifeTechnologies and the gels were then digitalized on a ImageMaster VDS-Pharmacia Biotech. The relative amounts ofdifferent forms of pB II KS+ DNA were determined byusing the Gel Perfect program [51]. In all cases, back-ground fluorescence was determined by quantification ofthe lane in the gel containing no DNA. The relativeamounts of the different forms of DNA were determinedby dividing the fluorescence intensity of a particular bandby the sum of the fluorescence intensities for each bandin that lane. A correction factor of 1.42 was used toaccount for the decreased ability of ethidium bromide tointercalate into supercoiled plasmid DNA [28,41].
2.4.5. Activity of 1 toward a human myeloid leukemia cell
line (THP-1)
THP-1 cells were seeded in a 96 well plate at a concen-tration of 1.5 · 104 cells per well, treated with 50, 100and 200 lM of complex 1 and apoptotic or necrotic cellswere scored at 12 and 24 h. Apoptosis evaluation was per-formed using fluorescent microscopy (Zeiss Axioplanmicroscope) with the cells stained by a solution containing5 lg/mL acridine orange and 25 lg/mL ethidium bromide.At least 200 cells in a randomly selected microscopic fieldwere counted for each experimental condition. The per-centage of apoptotic or necrotic cells was calculated as:total number of cells with apoptotic or necrotic morphol-ogy · 100/total number of cells counted.
DNA fragmentation assay was employed to evaluateinternucleossomal degradation of cellular DNA [52].Briefly, 2 · 106 cells were washed in PBS, lysed in 10 mMTris pH 7.5, 2 mM EDTA, 1% Triton X-100 for 20 min.on ice and then treated with 20 lg/mL RNAse A at37 �C for 1 h and 0.5 mg/mL of Proteinase K for an addi-tional hour. The DNA was precipitated in ethanol, centri-fuged for 20 min at 11000g, dissolved in loading buffer(10 mM Tris, pH 7.5, 2 mM EDTA, 3% Ficoll 400, 0.1%bromophenol blue-xylene cyanole), electrophoresed on a1.2% agarose gel for 1 h at 85 V and stained with ethidiumbromide.
3. Results and discussion
3.1. Description of the structure of
[Cu(HPClNOL)Cl]Cl Æ CH3OH (1)
The molecular structure of 1 is composed of a mononu-clear [Cu(HPClNOL)Cl]+ cation, one chloride anion and amethanol molecule as crystallization solvent. The crystalstructure of the cation is presented in Fig. 1 and selectedbond lengths and bond angles are presented in Table 2.
In 1, HPClNOL acts as a tetradentate chelating ligandthrough the pyridinic nitrogen atoms (N12 and N22), ami-nic nitrogen atom (N1) and the oxygen atom from the alco-hol (O1); furthermore, one chloride anion is coordinated tothe copper center, resulting in a pentacoordinated complex.Recently, the synthesis and chemical properties of a similarcopper complex [Cu(HPClNOL)(CH3CN)](ClO4)2 (2) wasreported [42]. Complex 2 was obtained by the reaction ofthe same ligand with [Cu(H2O)6](ClO4)2 instead of[Cu(H2O)6](Cl)2. The difference is that 2 presents an aceto-nitrile molecule coordinated to the copper ion, while in 1
the same position is occupied by a chloride anion. All theother coordination positions are equal for both.
The respective equatorial bond lengths are: Cu1–N1 = 2.048(3), Cu1–N12 = 1.994(3), Cu1–N22 = 2.003(3),Cu1–Cl1 = 2.246(1) A. The axial position is occupied bythe oxygen atom from the HPClNOL alcohol moiety,which is protonated, as indicated by the large bond lengthCu1–O1 = 2.386(3) A. The bonds formed between thenitrogen atoms and the copper ion are slightly longer in 1
than in 2. The main difference is observed in the Cu–Oalcohol bond length (2.340(4) in 2). This difference indi-cates a change in the Lewis acidity of the copper centerdue to the chloride coordination, which is a better donorspecie than the acetonitrile molecule. We can also compare1 and 2 with the analogue compound CuL02 ðL02 ¼N -ð2-hydroxyethylÞbisðbenzimidazolylmethylÞamineÞ [53],which shows a similar coordination environment aroundthe copper center, but with benzimidazolyl groups insteadof pyridyl groups. The Cu–Oalcohol distance of 2.509(6) Ain CuL02 is much greater than those observed for 1(2.386(3) A) and 2 (2.340(4) A). This may be reflectinga higher basicity of the benzimidazolyl groups (Cu–Nbenzimidazolyl bond lengths average = 1.949 A in CuL02)

Fig. 1. A perspective view of molecular structure of 1 with atomic labeling. The ellipsoids are shown with a 50% probability level.
C. Fernandes et al. / Inorganica Chimica Acta 359 (2006) 3167–3176 3171
when compared with the pyridyl groups (Cu–Npyridyl bondlengths average = 1.999 A in 1). Interestingly, the Cu–Clbond lengths in 1 (2.246(1) A) and in CuL02(2.2346(20) A) are similar, showing that the axial positionis the most sensitive to changes in the Lewis acidity ofthe copper center. Thus, the crystallographic analysisallows us to infer that the copper center in 1 is less acidthan in 2 which is in turn less acid than in CuL02. Thesquare-pyramidal geometry is confirmed by the structuralindex parameter s = 0.25 (this parameter is 1 for a perfecttrigonal bipyramidal geometry and 0 for perfect square-pyramidal geometry) [54].
Although the C2 carbon atom is chiral, both enantio-meric forms are present in the unit cell, confirming thepresence of a racemic mixture.
3.2. Spectroscopic and electrochemical studies
The electronic spectrum of 1, in CH3CN, displays bandsat 257 (10.3 · 103 dm3 mol�1 cm�1), 281 (3.2 ·103 dm3 mol�1 cm�1) and 727 nm (57 dm3 mol�1 cm�1).The band at higher energy is attributed to an intra-ligandcharger transfer process and the band at 281 nm can beaccounted as a ligand–metal charge transfer (LMCTCl! Cu2+). The band at lower energy is attributed to d–d transition. The d–d transition appears at a lower energythan that previously reported for compound 2 (621 nm)[42] and as in 2, the presence of only one well-definedd–d absorption indicates that the pentacoordinated struc-ture observed in the solid state is not preserved, possiblydue to the coordination of a solvent molecule to the coppercenter, resulting in tetragonally distorted octahedral geom-etry. Comparing the positions of the d–d absorptions forcomplexes 1 and 2, it is also possible to infer that the cop-per center in 1 is less acid than in 2. Considering the crystalfield theory, a more acid center should attract more theligands, resulting in a more accentuated electrostatic inter-action and therefore a higher splitting between the t2g and
eg orbitals (considering Oh symmetry), which would resultin d–d transition at higher energy. The molecular orbitalapproach is also in agreement with the observed results,since the coordination environment for complex 1 presentsr and p donor groups while in the complex 2 there are onlyr donor groups. Thus, in 1 the bonding combination of thepp(Cl�)–dp(t2gCu) orbitals results in an increase of theenergy of the t�2g orbital. The net effect is the reduction inthe ligand field splitting and therefore a d–d transition atlower energy is observed for complex 1.
Complex 1 was also investigated by electron paramag-netic resonance (EPR). In the solid state and in frozen ace-tonitrile solution (77 K), only a wide anisotropical signalwith a typical profile of a copper(II) center with tetragonalaxially distorted coordination was observed. Only whenTBAClO4 was added to the acetonitrile solution was it pos-sible to see the hyperfine lines resulting from the coupling ofthe unpaired electron with the nuclear spin of the copperion. The values of the principal components of the hyperfineand the g-tensors were obtained by computational simula-tions: gi = 2.236(1); g^ = 2.042(1); Ai = 174 G; A^ = 10 G.Complex 1 shows hyperfine coupling constants lower thanthose exhibited by complex 2 (Ai = 185 G; A^ = 15 G)[42]. The shape of the EPR lines and the values of gi andg^ (gi > g^ > 2.00) obtained at low temperature indicate thatthe unpaired electron is located in the dx2�y2 orbital [55].
Cyclic voltammetry and conductimetry were employedin the study of the electrochemical behaviour of 1. Con-ductimetry measurement in acetonitrile suggests that 1 isa 1:1 electrolyte type [43], indicating that there is a chlorideanion coordinated to the copper center when 1 is dissolvedin acetonitrile. The cyclic voltammetry (Fig. 2) shows onlyone quasi-reversible redox process at E1/2 = �0.172 V ver-sus NHE (DEpa–pc = 105 mV; DEpa–pc Fc/Fc+ = 85 mV)
attributed to Cu2+/Cu1+ redox couple. The process at0.426 V refers to internal standard Fc/Fc+. As it was pos-sible to investigate the Lewis acidity of the copper centersin 1 and 2 by comparing the molecular structure and the
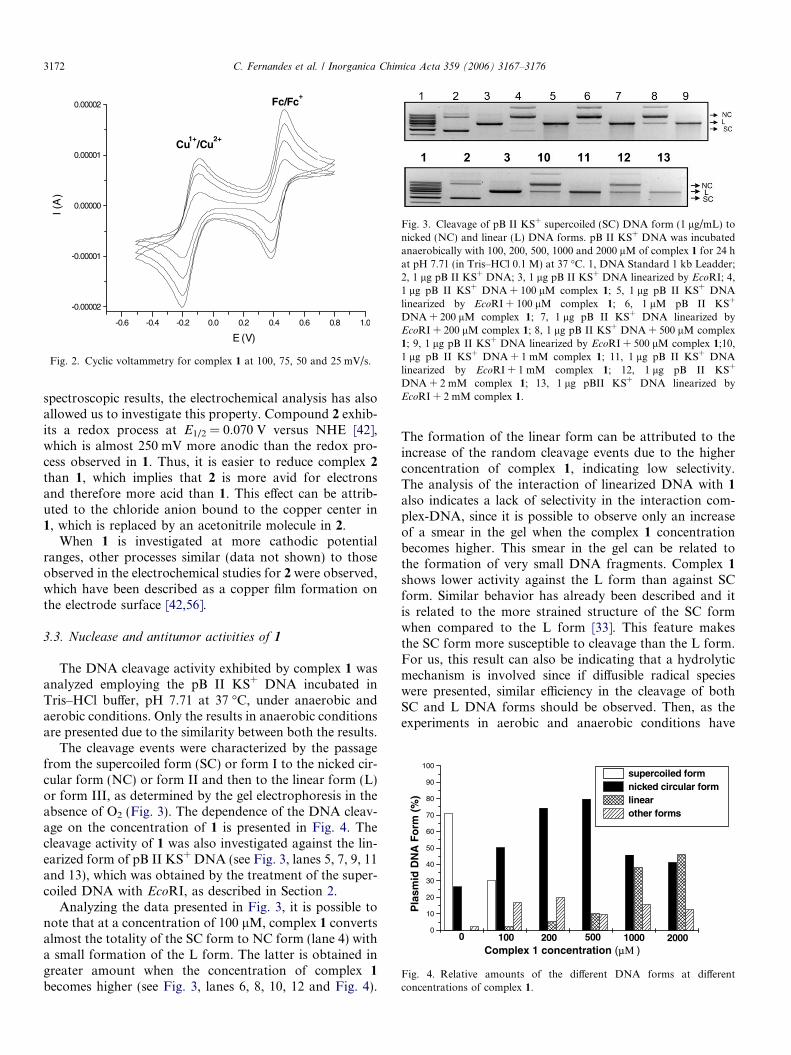
Fig. 3. Cleavage of pB II KS+ supercoiled (SC) DNA form (1 lg/mL) tonicked (NC) and linear (L) DNA forms. pB II KS+ DNA was incubatedanaerobically with 100, 200, 500, 1000 and 2000 lM of complex 1 for 24 hat pH 7.71 (in Tris–HCl 0.1 M) at 37 �C. 1, DNA Standard 1 kb Leadder;2, 1 lg pB II KS+ DNA; 3, 1 lg pB II KS+ DNA linearized by EcoRI; 4,1 lg pB II KS+ DNA + 100 lM complex 1; 5, 1 lg pB II KS+ DNAlinearized by EcoRI + 100 lM complex 1; 6, 1 lM pB II KS+
DNA + 200 lM complex 1; 7, 1 lg pB II KS+ DNA linearized byEcoRI + 200 lM complex 1; 8, 1 lg pB II KS+ DNA + 500 lM complex1; 9, 1 lg pB II KS+ DNA linearized by EcoRI + 500 lM complex 1;10,1 lg pB II KS+ DNA + 1 mM complex 1; 11, 1 lg pB II KS+ DNAlinearized by EcoRI + 1 mM complex 1; 12, 1 lg pB II KS+
DNA + 2 mM complex 1; 13, 1 lg pBII KS+ DNA linearized byEcoRI + 2 mM complex 1.
-0.6 -0.4 -0.2 0.0 0.2 0.4 0.6 0.8 1.0
-0.00002
-0.00001
0.00000
0.00001
0.00002
Cu1+
/Cu2+
Fc/Fc+
I (A
)
E (V)
Fig. 2. Cyclic voltammetry for complex 1 at 100, 75, 50 and 25 mV/s.
0
10
20
30
40
50
60
70
80
90
100
200010005002001000
Pla
smid
DN
A F
orm
(%
)
Complex 1 concentration (μΜ )
supercoiled formnicked circular formlinearother forms
Fig. 4. Relative amounts of the different DNA forms at differentconcentrations of complex 1.
3172 C. Fernandes et al. / Inorganica Chimica Acta 359 (2006) 3167–3176
spectroscopic results, the electrochemical analysis has alsoallowed us to investigate this property. Compound 2 exhib-its a redox process at E1/2 = 0.070 V versus NHE [42],which is almost 250 mV more anodic than the redox pro-cess observed in 1. Thus, it is easier to reduce complex 2
than 1, which implies that 2 is more avid for electronsand therefore more acid than 1. This effect can be attrib-uted to the chloride anion bound to the copper center in1, which is replaced by an acetonitrile molecule in 2.
When 1 is investigated at more cathodic potentialranges, other processes similar (data not shown) to thoseobserved in the electrochemical studies for 2 were observed,which have been described as a copper film formation onthe electrode surface [42,56].
3.3. Nuclease and antitumor activities of 1
The DNA cleavage activity exhibited by complex 1 wasanalyzed employing the pB II KS+ DNA incubated inTris–HCl buffer, pH 7.71 at 37 �C, under anaerobic andaerobic conditions. Only the results in anaerobic conditionsare presented due to the similarity between both the results.
The cleavage events were characterized by the passagefrom the supercoiled form (SC) or form I to the nicked cir-cular form (NC) or form II and then to the linear form (L)or form III, as determined by the gel electrophoresis in theabsence of O2 (Fig. 3). The dependence of the DNA cleav-age on the concentration of 1 is presented in Fig. 4. Thecleavage activity of 1 was also investigated against the lin-earized form of pB II KS+ DNA (see Fig. 3, lanes 5, 7, 9, 11and 13), which was obtained by the treatment of the super-coiled DNA with EcoRI, as described in Section 2.
Analyzing the data presented in Fig. 3, it is possible tonote that at a concentration of 100 lM, complex 1 convertsalmost the totality of the SC form to NC form (lane 4) witha small formation of the L form. The latter is obtained ingreater amount when the concentration of complex 1
becomes higher (see Fig. 3, lanes 6, 8, 10, 12 and Fig. 4).
The formation of the linear form can be attributed to theincrease of the random cleavage events due to the higherconcentration of complex 1, indicating low selectivity.The analysis of the interaction of linearized DNA with 1
also indicates a lack of selectivity in the interaction com-plex-DNA, since it is possible to observe only an increaseof a smear in the gel when the complex 1 concentrationbecomes higher. This smear in the gel can be related tothe formation of very small DNA fragments. Complex 1
shows lower activity against the L form than against SCform. Similar behavior has already been described and itis related to the more strained structure of the SC formwhen compared to the L form [33]. This feature makesthe SC form more susceptible to cleavage than the L form.For us, this result can also be indicating that a hydrolyticmechanism is involved since if diffusible radical specieswere presented, similar efficiency in the cleavage of bothSC and L DNA forms should be observed. Then, as theexperiments in aerobic and anaerobic conditions have

C. Fernandes et al. / Inorganica Chimica Acta 359 (2006) 3167–3176 3173
shown similar results and the activities against the SC andthe L forms have been significantly different, the resultspresented here indicate that the cleavage of DNA by 1 isradical-independent.
Seeking more insights into the mechanism of cleavageexhibited by complex 1, religation assays were carriedout. It is worth pointing out that only DNA cleaved by ahydrolytic mechanism can be religated after treatment withthe ligase enzyme.
The religation assay of the DNA previously treated with1 (Fig. 5, lane 6) did not produce any additional visibleDNA forms (for comparison, see lanes 4 and 6). On theother hand, the L DNA form, obtained by digestion ofthe SC form by EcoRI and incubated in the presence ofcomplex 1, showed partial transformation to the NC formwhen it was incubated with T4 ligase (for comparison, seelane 5 and 7). This result could be indicating that the mech-anism involved in this process does not follow a hydrolyticpathway. However, some studies carried out with othercomplexes have also failed to show DNA religation,although the hydrolytic mechanism were proposed[34,41]. The accepted mechanism for the hydrolytic DNAcleavage promoted by metal complexes is the substitutionof a labile molecule, water for example, by the phosphategroup of the DNA backbone (Lewis activation) followedby an intramolecular nucleophilic attack [9–13]. Accordingto this mechanism, Neves and co-workers, supported bystructural and kinetic data, have shown that the coppercomplex [Cu2(Hbtppnol)(l-CH3COO)](ClO4)2 (H2btpp-nol = N-(2-hydroxybenzyl)-N,N 0,N 0-tris(2-pyridylmethyl)-1,3-diaminopropan-2-ol) can promote only the stoichiome-tric hydrolysis of the bis(2,4-dinitrophenyl)phosphate(DNA mimic), promoting the cleavage of only one 2,4-dinitrophenyl group, while the 2,4-dinitrophenylphosphategroup remains bonded to the copper centers after thehydrolysis process, forming a phosphate bridge [34]. Theyhave used this result to explain why the ligase enzyme couldnot religate the DNA cleaved by the complex [Cu2(Hbtpp-nol)(l-CH3COO)](ClO4)2, inferring that the DNA phos-phate group remained bound to the complex after theDNA hydrolysis process. Thus, the phosphate group isnot free to be accessed by the ligase enzyme.
Therefore, based on the above results and due to thestructural features (presence of labile ligands), our pro-posal is that complex 1 promotes the hydrolytic cleavageof the DNA phosphate ester groups, which remain coordi-
Fig. 5. Analysis of the capacity of T4 DNA ligase to religate DNA cleavedby complex 1. 1, DNA Standard 1 kb Leadder; 2, pB II KS+ DNA; 3, pBII KS+ DNA linearized by EcoRI; 4, pB II KS+ DNA + complex 1; 5, pBII KS+ DNA linearized by EcoRI + complex 1; 6, pB II KS+
DNA + complex 1 + T4 ligase; 7, pB II KS+ DNA linearized byEcoRI + complex 1 + T4 ligase.
nated to the copper(II) center, preventing the DNA religa-tion. The strong interaction between 1 and the phosphategroups would prevent the action of the ligase enzyme onthe DNA strands and explains why it was not possible toperform the DNA religation even though the cleavageshould have followed a hydrolytic mechanism.
As can be seen by comparing lanes 4, 6, 8 and 10(Fig. 3), the cleavage process under investigation resultedin a fourth band that can be seen on the top of the gel. Thisband is quantified in Fig. 4, being identified as ‘‘otherforms’’ (in this classification we have also included a bandof very small intensity located near the NC band, which iseasiest observed in lane 2). The main feature of the fourthband is that it shows smaller displacement in the gel aga-rose when compared with the other bands. This featureindicates a stronger interaction between this form and thegel. The migration of the bands is related to the size andform of the DNA [57]. Therefore, this band may be attrib-uted to the formation of a more voluminous form thanthose well-documented SC, NC, and L forms. Based onthis result, we can propose that: (a) complex 1 is promotingthe cleavage of the SC form to the NC form, resulting inthe formation of two types of NC forms, one of them witha more relaxed structure; (b) complex 1 is presenting astrong interaction with the DNA (electrostatic, for exam-ple), resulting in the neutralization of the negative chargesof the DNA. This neutralization would result in changes inits migration velocity through the gel; (c) complex 1 is pro-moting an agglutination of two or more DNA molecules,resulting in a larger specie which would show more diffi-culty to pass through the porous gel. So, the band locatedon the top of the gel could be related to this agglutinationprocess. This proposal is supported by the results obtainedby Onoa and Marcos, who have reported the formation ofdimeric DNA structure when SC pBR 322 DNA wasrelaxed by Calf Thymus DNA topoisomerase I. Thisdimeric form showed the smallest displacement in the aga-rose gel electrophoresis [58].
Since complex 1 induces conformational changes in theDNA, and DNA replication is a condition for cell prolif-eration, we have carried out in vitro assays to evaluate thecytotoxic and antiproliferative effects performed by com-plex 1 on a culture of human myeloid leukemia cells line(THP-1). The cells were incubated with different complex1 concentrations (50, 100, 200 lM) and the cell deathinduction was evaluated after 12 and 24 h. In order toevaluate the mechanism of cell death, the cells werestained with acridine orange and ethidium bromide. Onlythe apoptotic cells are permeable to acridine orange whilenecrotic cells are permeable to ethidium bromide [59].Thus, is possible to evaluate the mechanism of cell death(apoptotic or necrotic) by fluorescent microscopy. Cellmorphology and DNA degradation studies were also usedto confirm the apoptotic or necrotic mechanism. Apopto-sis is a form of self-regulated cell death that differs fromnecrosis with characteristic morphological changes thatinclude membrane bledding, chromatin condensation

Fig. 6. Induction of apoptosis by complex 1. THP-1 cells were cultivated with different complex 1 concentrations and apoptosis was evaluated. (A)photomicrography of apoptotic cells treated with 200 lM of complex 1 during 24 h. Arrow indicates the typical apoptotic cell with fragmented nucleus.Arrowhead shows an initial apoptosis phase, a cell with picnotic nucleus. (B) Control cells. (C) Agarose gel electrophoresis of DNA from cells treated withcomplex 1 after 24 h: 1, complex 1 200 lM; 2, complex 1 100 lM; 3, complex 1 50 lM; 4, control cells. (D) Apoptotic cells were counted after 12 h ofincubation. (E) Evaluation of apoptotic cells after 24 h. AP, apoptotic cells; NE, necrotic cells; NO, normal cells.
3174 C. Fernandes et al. / Inorganica Chimica Acta 359 (2006) 3167–3176
and the formation of apoptotic bodies [60]. A typicalmorphology of apoptosis, such as cell shrinkage, chroma-tin condensation and nuclear fragmentation was observedwhen THP-1 cells were treated with complex 1 (compareFig. 6A and B). The analysis of the DNA isolated fromthe cells has revealed that DNA fragmentation wasinvolved in cell death (Fig. 6C), confirming the apoptoticmechanism [61].
Fig. 6D and E show that complex 1 induces apoptosis inTHP-1 cells, showing dependence on time and complex 1
concentration. As can be seen, 33% of apoptotic cells wereobserved in THP-1 cell culture treated with a 50 lM com-plex 1 concentration after 12 h of incubation, as confirmedby acridine orange staining fluorescence. The highest level(75%) was reached after 24 h of treatment with a 200 lMcompound 1 concentration. Necrotic cells remained extre-mely low, less than 5%, until 24 h of culture as presumedby ethidium bromide staining, due to the permeability ofthe necrotic cell membrane to this stain. Higher levels ofsecondary necrotic cells were observed after 48 h of treat-ment, reaching the maximum of 46% when the cells weretreated with 200 lM of complex 1 (data not shown). Thepresence of secondary necrotic cells is expected, since apop-totic cells, in vitro, undergo secondary necrosis as an indi-cation of the end of the process.
One of the possible mechanisms of apoptosis induced bycomplex 1 could be associated with DNA damage, sincecomplex 1 promotes the cleavage of plasmid DNA(Fig. 3). This proposal is in agreement with Liang and
co-workers results, who have proposed that the antitumoractivity of the copper complex [CuL]2+ (L = 3-hydroxyl-1,5,8-triazacyclicamine) may be resulted from its abilityto damage DNA in the cell cycle [16]. However, severalintracellular processes like activation of caspases and/ordisruption of normal mitochondrial function could beinvolved in the DNA degradation [61].
4. Conclusion
In this work, we have shown the synthesis andcharacterization of a mononuclear copper(II) complex[Cu(HPClNOL)Cl]Cl Æ MeOH (1). This compound exhibitsa square-pyramidal geometry. The presence of a free coor-dination position and labile ligands (chloride ion, alcoholgroup) prompt us to investigate its ability in promotingthe DNA cleavage. The DNA cleavage activity of complex1 was confirmed in both anaerobic and aerobic media, indi-cating that the cleavage mechanism is probably oxygen-independent. The distinct activity of 1 against the L andSC DNA forms also indicates that non-diffusible radicalspecies were involved in the cleavage mechanism. Theresults have shown that the DNA cleavage should followa hydrolytic pathway. Interestingly, complex 1 promotesthe death of THP-1, a human leukemia cells in vitro, induc-ing cell death by apoptosis. New investigations are beingcarried out aiming to understand more about the apoptoticmechanism as well as to evaluate the activity of complex 1in vivo.

C. Fernandes et al. / Inorganica Chimica Acta 359 (2006) 3167–3176 3175
Acknowledgements
The authors are grateful to CCDC by the free ofcharge License of Access to the following programs: The
CSD System – The Cambridge Structural Database: aquarter of a million crystal structure and rising. F.H.Aleen, Acta Cryst. B58 (2002) 380–388. ConQuest –New Software for searching the Cambridge StructuralDatabase and visualizing crystal structures. I.J. Bruno,J.C. Cole, P.R. Edington, M. Kessler, C.F. Macrae, P.McCabe, J. Pearson, R. Taylor, Acta Cryst. B58 (2002),389–397. IsoStar – IsoStar: A Library of Informationabout Nonbonded Interactions. I.J. Bruno, J.C. Cole,J.P.M. Lommerse, R.S. Rowland, R. Taylor, M. Verdonl,Jounal of Computer Aided Molecular Design 11-6 (1997)525–537. A.H. Jr. and C.F. are grateful to CNPq and FA-PERJ. A.J.B. is grateful to financial support receivedfrom FINEP and CNPq. N.V.V. is grateful to CNPq byresearch fellowship. A.H. Jr. and C.F. acknowledge thesupport of Instituto de Quımica-Universidade Federaldo Rio de Janeiro.
Appendix A. Supplementary material
The crystallographic data (atomic coordinates andequivalent isotropic displacement parameters, calculatedhydrogen atom parameters, anisotropic thermal parame-ters and bond lengths and angles) have been deposited atthe Cambridge Crystallographic Data Center (depositionnumber CCDC 281283). Copies of this information maybe obtained free of charge from: CCDC, 12 Union Road,Cambridge, CB2 1EZ, UK (fax: +44 1223 366 033, e-mail:[email protected] or on the web www: http://www.ccdc.cam.ac.uk).
References
[1] R.J.P. Williams, J.J.R. Frausto da Silva, Coord. Chem. Rev. 200–202(2000) 247.
[2] R.J.P. Williams, Coord. Chem. Rev. 216–217 (2001) 583.[3] J.J.R. Frausto da Silva, R.J.P. Williams, The Biological Chemistry of
the Elements: The Inorganic Chemistry of Live, 2nd ed., OxfordUniversity Press, Oxford, UK, 2001.
[4] W. Kaim, B. Schwederski, Bioinorganic Chemistry: InorganicElements in the Chemistry of Life: An Introduction and Guide,Wiley, Chichester, England, 1994.
[5] E.I. Solomon, U.M. Sundaram, T.E. Machonkin, Chem. Rev. 96(1996) 2563.
[6] E.A. Lewis, W.B. Tolman, Chem. Rev. 104 (2004) 1047.[7] R. Than, A.A. Feldmann, B. Krebs, Coord. Chem. Rev. 182 (1999)
211.[8] K. Selmeczi, M. Reglier, M. Giorgi, G. Speier, Coord. Chem. Rev.
245 (2003) 191.[9] E.L. Hegg, J.N. Burstyn, Coord. Chem. Rev. 173 (1998) 133.
[10] R. Kramer, Coord. Chem. Rev. 182 (1999) 243.[11] A. Sreedhara, J.A. Cowan, J. Biol. Inorg. Chem. (2001) 337.[12] J.A. Cowan, Curr. Opin. Chem. Biol. (2001) 634.[13] C. Liu, M. Wang, T. Zhang, H. Sun, Coord. Chem. Rev. 248 (2004)
147.[14] M. Navarro, E.J. Cisneros-Fajardo, M. Fernadez-Mestre, D. Arri-
eche, E. Marchan, J. Inorg. Biochem. 97 (2003) 364.
[15] M.S. Melvin, K.E. Wooton, C.C. Rich, G.R. Saluta, G.L.Kucera, N. Lindquist, R.A. Manderville, J. Inorg. Biochem. 87(2001) 129.
[16] F. Liang, C. Wu, H. Lin, T. Li, D. Gao, Z. Li, J. Wei, C. Zheng, M.Sun, Bioorg. Med. Chem. Lett. 13 (2003) 2469.
[17] D. Kostrewa, F.K. Winkler, Biochemistry 34 (1995) 683.[18] J.A. Cowan, Chem. Rev. 98 (1998) 1067.[19] S.M. Hecht, Bioorganic Chemistry: Nucleics Acids, Oxford Univer-
sity Press, New York, 1996.[20] D.R. Mcmillin, K.M. McNett, Chem. Rev. 98 (1998) 1201.[21] X. Wang, H. Chao, H. Li, X. Hong, L. Ji, X. Li, J. Inorg. Biochem.
98 (2004) 423.[22] M. Gonzalez-Alvarez, G. Alzuet, J. Borras, M. Pitie, B. Meunier, J.
Biol. Inorg. Chem. 8 (2003) 644.[23] S. Dhar, D. Senapati, P.K. Das, P. Chattopadhyay, M. Nethaji,
A.R. Chakravarty, J. Am. Chem. Soc. 125 (2003) 12188.[24] A.M. Thomas, M. Nethaji, S. Mahadevan, A.R. Chakravarty, J.
Inorg. Biochem. 94 (2003) 171.[25] A.M. Thomas, M. Nethaji, A.R. Chakravarty, J. Inorg. Biochem. 98
(2004) 1087.[26] K.H. Reddy, P.S. Reddy, P.R. Babu, J. Inorg. Biochem. 77 (1999)
169.[27] T. Hirohama, H. Arii, M. Chikira, J. Inorg. Biochem. 98 (2004) 1778.[28] E.L. Hegg, J.N. Burstyn, Inorg. Chem. 35 (1996) 7474.[29] G.N. De Iuliis, G.A. Lawrance, S. Fieuw-Makaroff, Inorg. Chem.
Commun. 3 (2000) 307.[30] L.M. Rossi, A. Neves, R. Horner, H. Terenzi, B. Szpoganicz, J.
Sugai, Inorg. Chim. Acta 337 (2002) 366.[31] K.M. Deck, T.A. Tseng, J.N. Burstyn, Inorg. Chem. 41 (2002) 669.[32] M. Scarpellini, A. Neves, R. Horner, A.J. Bortoluzzi, B. Szpoganicz,
C. Zucco, R.A. Nome Silva, V. Drago, A.S. Mangrich, W.A. Ortiz,W.A.C. Passos, M.C.B. de Oliveira, H. Terenzi, Inorg. Chem. 42(2003) 8353.
[33] S.I. Kirin, C.M. Happel, S. Hrubanova, T. Weyhermuller, C. Klein,N. Metzler-Nolte, Dalton Trans. (2004) 1201.
[34] L.M. Rossi, A. Neves, A.J. Bortoluzzi, R. Horner, B. Szpoganicz, H.Terenzi, A.S. Mangrich, E. Pereira-Maia, E.E. Castellano, W. Haase,Inorg. Chim. Acta 358 (2005) 1807.
[35] P.S. Fricker, Metal compound in cancer therapy, in: The Role ofMetals in Cancer Therapy, Chapman and Hall, London, 1994.
[36] A. De Vizcaya-Ruiz, A. Rivero-Muller, L. Ruiz-Ramirez, G.E.N.Kass, L.R. Kelland, R.M. Orr, M. Dobrota, Toxicol. In Vitro 14(2000) 1.
[37] M.D. Pulido, A.R. Parrish, Mutat. Res. 533 (2003) 227.[38] J.V. Kidd, Annu. Rev. Physiol. 60 (1998) 533.[39] K.L. King, J.A. Cidlowski, Annu. Rev. Physiol. 60 (1998) 601.[40] A. Horn Jr., I. Vencato, A.J. Bortoluzzi, R. Horner, R.N. Nome
Silva, B. Szpoganicz, V. Drago, H. Terenzi, M.C.B. de Oliveira, R.Werner, W. Haase, A. Neves, Inorg. Chim. Acta 358 (2005) 339.
[41] A. Neves, H. Terenzi, R. Horner, A. Horn Jr., B. Szpoganicz, J.Sugai, Inorg. Chem. Commun. 4 (2001) 388.
[42] A. Horn Jr., C. Fernandes, A.J. Bortoluzzi, N.V. Vugman, M.H.Herbst, J. Mol. Struct. 749 (2005) 96.
[43] W.J. Geary, Coord. Chem. Rev. 7 (1971) 81.[44] R.R. Gagne, C.A. Koval, G.C. Lisensky, Inorg. Chem. 19 (1980)
2854.[45] A.L. Spek, HELENA; CAD-4 Data Reduction Program, Univ. of
Utrecht, The Netherlands, 1996.[46] A.L. Spek, Acta Crystallogr., Sect. A 36 (1990) C34.[47] A.C.T. North, D.C. Phillips, F.S. Mathews, Acta. Crystallogr., Sect.
A 24 (1968) 351.[48] A. Altomare, M.C. Burla, M. Camalli, G.L. Cascarano, C. Giaco-
vazzo, A. Guagliardi, A.G.G. Moliterni, G. Polidori, R. Spagna, J.Appl. Cryst. 32 (1999) 115.
[49] G.M. Sheldrick, SHELXL97, Program for the Refinement of CrystalStructures, University of Gottingen, Gottingen, Germany, 1997.
[50] L.J.J. Farrugia, Appl. Crystallogr. 30 (1997) 565.[51] S. Bozzo, C. Retamal, Arch. Biol. Med. Exp. 24 (1991) R-181.

3176 C. Fernandes et al. / Inorganica Chimica Acta 359 (2006) 3167–3176
[52] E.B. Lasunskaia, I.I. Fridlianskaia, Z. Darieva, M.S.R. da Silva,M.M. Kanashiro, B.A. Margulis, Biotechnol. Bioeng. 81 (2003)496.
[53] M.J. Young, D. Wahnon, R.C. Hynes, J. Chin, J. Am. Chem. Soc.117 (1995) 9441.
[54] A.W. Addison, T.N. Rao, J. Reedijk, J. van Rijn, G.C. Verschoor, J.Chem. Soc., Dalton. Trans. (1984) 1349.
[55] A. Abragam, B. Bleaney, Electron Paramagnetic Resonance ofTransition Ions, Clarendon Press, Oxford, 1970.
[56] S.M.M. Romanowski, A.S. Mangrich, A. Neves, Quim. Nova 24(2001) 592.
[57] L.-Z. Li, C. Zhao, T. Xu, H.-W. Ji, Y.-H. Yu, G.-Q. Guo, H. Chao,J. Inorg. Biochem. 99 (2005) 1076.
[58] G.B. Onoa, V. Moreno, Int. J. Pharm. 245 (2002) 55.[59] J.E. Coligan, A.M. Kruisbeek, D.H. Margulies, E.M. Shevach, W.
Strober, In vitro assay for muose B and T cell functionCurrentProtocols of Immunology, vol. 1, Suppl. 16, Greene PublishingAssociates and Wiley-Interscience, 1995.
[60] M.B. Ferrari, S. Capacchi, G. Pelosi, G. Reffo, P. Tarasconi, R.Albertini, S. Pinelli, P. Lunghi, Inorg. Chim. Acta 286 (1999)134.
[61] N.N. Blatt, G.D. Glick, Bioorg. Med. Chem. 9 (2001) 1371.









![The formation, transport properties and microstructure of 45° [001] grain boundaries induced by epitaxy modification in YBa 2Cu 3O 7− x thin films](https://static.fdokumen.com/doc/165x107/6315b7dbc32ab5e46f0d6fce/the-formation-transport-properties-and-microstructure-of-45-001-grain-boundaries.jpg)










