
Mega TALs- a rare-cleaving nuclease architecture for therapeutic genome engineering
11
megaTALs: a rare-cleaving nuclease architecture for therapeutic genome engineering Sandrine Boissel 1,2 , Jordan Jarjour 3 , Alexander Astrakhan 3 , Andrew Adey 1,4 , Agne ` s Gouble 5 , Philippe Duchateau 5 , Jay Shendure 4 , Barry L. Stoddard 6 , Michael T. Certo 2 , David Baker 7,8 and Andrew M. Scharenberg 2,9, * 1 Molecular and Cellular Biology Program, University of Washington, Seattle, WA 98195, USA, 2 Center for Immunity and Immunotherapies, Seattle Children’s Research Institute, Seattle, WA 98101, USA, 3 Pregenen, Inc., Seattle, WA 98103, USA, 4 Department of Genome Sciences, University of Washington, Seattle, WA 98195, USA, 5 Cellectis S.A., Paris, 75013, France, 6 Division of Basic Sciences, Fred Hutch Cancer Research Center, Seattle, WA 98109, USA, 7 Department of Biochemistry, University of Washington, Seattle, WA 98195, USA, 8 Howard Hughes Medical Institute, University of Washington, Seattle, WA 98195, USA and 9 Department of Immunology, University of Washington, Seattle, WA 98195, USA Received September 26, 2013; Revised November 4, 2013; Accepted November 5, 2013 ABSTRACT Rare-cleaving endonucleases have emerged as im- portant tools for making targeted genome modifica- tions. While multiple platforms are now available to generate reagents for research applications, each existing platform has significant limitations in one or more of three key properties necessary for thera- peutic application: efficiency of cleavage at the desired target site, specificity of cleavage (i.e. rate of cleavage at ‘off-target’ sites), and efficient/facile means for delivery to desired target cells. Here, we describe the development of a single-chain rare- cleaving nuclease architecture, which we designate ‘megaTAL’, in which the DNA binding region of a transcription activator-like (TAL) effector is used to ‘address’ a site-specific meganuclease adjacent to a single desired genomic target site. This architec- ture allows the generation of extremely active and hyper-specific compact nucleases that are compat- ible with all current viral and nonviral cell delivery methods. INTRODUCTION Targeted genome modifications can be achieved using rare cleaving nucleases, such as zinc-finger nucleases (ZFNs), transcription activator-like effector nucleases (TALENs), meganucleases (mns, also termed homing endonucleases) and clustered, regularly interspaced, short palindromic repeat (CRISPR) RNA-guided nucleases (1–9). These reagents allow precise alterations to be introduced at their DNA recognition sites through cleavage of both DNA strands to yield double-strand breaks (DSBs) that are recognized and processed by the nonhomologous end joining (NHEJ) or homologous recombination (HR) DNA repair pathway (10–12). Repair by NHEJ involves religation of the DNA ends, which, after repeated cleavage, is often accompanied by mutagenesis via the introduction of small insertions or deletions (indels) at the site of the DSB that can result in the disruption of the coding sequence of a gene. In contrast, repair by HR results in seamless modification of the genome by copying an appropriate homologous DNA template at the site of the DNA break. The most demanding application of rare-cleaving nucle- ases is their use for therapeutic genome editing. For these applications, nuclease reagents must be delivered to a desired target cell in sufficient quantity to efficiently generate the desired DNA modification, while maintaining exquisite specificity to avoid compromising the genomic integrity of the cell. While the latest advances in each of the above-mentioned nuclease platforms enable quality nuclease reagents that are suitable for nearly all research and nontherapeutic commercial applications to be generated for little cost (13–20), several considerations still remain that limit their use for human cell therapies. These include a significant potential for off-target cleavage, particularly if expressed at high levels, as has been observed in the therapeutic application of ZFNs for disruption of the CCR5 gene in human T-cells (21–23). Additionally, since the FokI cleavage domain *To whom correspondence should be addressed. Tel: +1 206 987 7314; Fax:+1 206 987 7310; Email: [email protected] Present Address: Michael T. Certo, Department of Systems Biology, Harvard Medical School, Boston, MA 02115, USA. Nucleic Acids Research, 2013, 1–11 doi:10.1093/nar/gkt1224 ß The Author(s) 2013. Published by Oxford University Press. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/3.0/), which permits unrestricted reuse, distribution, and reproduction in any medium, provided the original work is properly cited. Nucleic Acids Research Advance Access published November 26, 2013 by guest on March 5, 2014 http://nar.oxfordjournals.org/ Downloaded from
-
Upload
washington -
Category
Documents
-
view
2 -
download
0
Transcript of Mega TALs- a rare-cleaving nuclease architecture for therapeutic genome engineering

megaTALs a rare-cleaving nuclease architecture fortherapeutic genome engineeringSandrine Boissel12 Jordan Jarjour3 Alexander Astrakhan3 Andrew Adey14
Agnes Gouble5 Philippe Duchateau5 Jay Shendure4 Barry L Stoddard6 Michael
T Certo2 David Baker78 and Andrew M Scharenberg29
1Molecular and Cellular Biology Program University of Washington Seattle WA 98195 USA 2Center forImmunity and Immunotherapies Seattle Childrenrsquos Research Institute Seattle WA 98101 USA 3Pregenen IncSeattle WA 98103 USA 4Department of Genome Sciences University of Washington Seattle WA 98195USA 5Cellectis SA Paris 75013 France 6Division of Basic Sciences Fred Hutch Cancer Research CenterSeattle WA 98109 USA 7Department of Biochemistry University of Washington Seattle WA 98195 USA8Howard Hughes Medical Institute University of Washington Seattle WA 98195 USA and 9Department ofImmunology University of Washington Seattle WA 98195 USA
Received September 26 2013 Revised November 4 2013 Accepted November 5 2013
ABSTRACT
Rare-cleaving endonucleases have emerged as im-portant tools for making targeted genome modifica-tions While multiple platforms are now available togenerate reagents for research applications eachexisting platform has significant limitations in oneor more of three key properties necessary for thera-peutic application efficiency of cleavage at thedesired target site specificity of cleavage (ie rateof cleavage at lsquooff-targetrsquo sites) and efficientfacilemeans for delivery to desired target cells Here wedescribe the development of a single-chain rare-cleaving nuclease architecture which we designatelsquomegaTALrsquo in which the DNA binding region of atranscription activator-like (TAL) effector is used tolsquoaddressrsquo a site-specific meganuclease adjacent toa single desired genomic target site This architec-ture allows the generation of extremely active andhyper-specific compact nucleases that are compat-ible with all current viral and nonviral cell deliverymethods
INTRODUCTION
Targeted genome modifications can be achieved using rarecleaving nucleases such as zinc-finger nucleases (ZFNs)transcription activator-like effector nucleases (TALENs)meganucleases (mns also termed homing endonucleases)and clustered regularly interspaced short palindromic
repeat (CRISPR) RNA-guided nucleases (1ndash9) Thesereagents allow precise alterations to be introduced attheir DNA recognition sites through cleavage of bothDNA strands to yield double-strand breaks (DSBs) thatare recognized and processed by the nonhomologous endjoining (NHEJ) or homologous recombination (HR)DNA repair pathway (10ndash12) Repair by NHEJ involvesreligation of the DNA ends which after repeatedcleavage is often accompanied by mutagenesis via theintroduction of small insertions or deletions (indels) atthe site of the DSB that can result in the disruption ofthe coding sequence of a gene In contrast repair by HRresults in seamless modification of the genome by copyingan appropriate homologous DNA template at the site ofthe DNA breakThe most demanding application of rare-cleaving nucle-
ases is their use for therapeutic genome editing For theseapplications nuclease reagents must be delivered to adesired target cell in sufficient quantity to efficientlygenerate the desired DNA modification while maintainingexquisite specificity to avoid compromising the genomicintegrity of the cell While the latest advances in each ofthe above-mentioned nuclease platforms enable qualitynuclease reagents that are suitable for nearly all researchand nontherapeutic commercial applications to begenerated for little cost (13ndash20) several considerationsstill remain that limit their use for human cell therapiesThese include a significant potential for off-targetcleavage particularly if expressed at high levels as hasbeen observed in the therapeutic application of ZFNsfor disruption of the CCR5 gene in human T-cells(21ndash23) Additionally since the FokI cleavage domain
To whom correspondence should be addressed Tel +1 206 987 7314 Fax +1 206 987 7310 Email andrewmsuweduPresent AddressMichael T Certo Department of Systems Biology Harvard Medical School Boston MA 02115 USA
Nucleic Acids Research 2013 1ndash11doi101093nargkt1224
The Author(s) 2013 Published by Oxford University PressThis is an Open Access article distributed under the terms of the Creative Commons Attribution License (httpcreativecommonsorglicensesby30) whichpermits unrestricted reuse distribution and reproduction in any medium provided the original work is properly cited
Nucleic Acids Research Advance Access published November 26 2013 by guest on M
arch 5 2014httpnaroxfordjournalsorg
Dow
nloaded from
used in ZFNs and TALENs is activated via dimerizationZFN and TALEN technologies require the constructionand delivery of two protein halves each comprising adistinct DNA binding domain fused to FokI for asingle target site This architectural requirement limitsthe potential for ZFN and TALEN delivery intoprimary cells using viral vectors as well as their simultan-eous (ie multiplexed) delivery of two or more nucleases tomodify more than one gene (2425) Although CRISPRsystems have shown exciting promise in developingreagents for genome engineering with easily designedtarget recognition they exhibit high off-target cleavagewhen used with single-guide RNAs thus necessitatingthe use of nicking enzymes and dual guide RNAs pertarget to achieve specificity equivalent to that of theother major platforms (26ndash28) The consequent need todeliver both the Cas9 ORF plus two guides per targetwould appear to pose significant limitations for the useof CRISPR in therapeutic applications requiring viral vec-torization as well as for applications requiring multiplex-ing Finally although architecturally well suited fortherapeutic applications mnsrsquo tightly coupled cleavageand binding activity have limited their potential for re-design toward novel DNA target sites with the majorityof redesign efforts resulting in respecified enzymes withlow target site affinity and consequent low overallactivity (2930)To address the limitations of present nuclease plat-
forms we have developed a hybrid nuclease architecturethat combines the ease of engineerability of a TALeffector (TALE) with the cleavage sequence specificity ofa mn cleavage domain This lsquomegaTALrsquo architecture wasachieved by fusing minimal TAL effector domains to theN-terminus of mns derived from the LAGLIDADGhoming endonuclease family
MATERIALS AND METHODS
megaTAL and mn construct generation
MegaTALs were constructed using the Golden Gateassembly strategy previously described by Cermak et alusing an repeat variable diresidue (RVD) plasmid libraryand destination vector generously provided by the Voytaslab (17) Plasmids were modified to allow the assembly ofTAL effectors 15ndash105 RVDs long The pthX01 destin-ation vector was modified to include a hemagglutinin(HA) tag immediately downstream of the nuclearlocalization signal (NLS) and to yield a N154 C+63TALEN scaffold TAL effectors were built using the fol-lowing RVDs to target each specific nucleotide AmdashNICmdashHD GmdashNN and TmdashNG Following cloning of theTAL effector repeats into the destination vector theprotein linkers (Fok SQLVKS Zn4 VGGS Flex1GGSG or Flex2 GGSGGGSG) and I-AniI or I-OnuImn variants were cloned in place of the FokI nucleasecatalytic domain between the Xba-I and Sal-I restrictionsites All megaTALs except those used to compare linkeractivity were made using the Zn4 protein linker Thecodon diverged 65C-WT megaTAL (SupplementaryFigure S2d) was constructed by gene synthesizing a TAL
effector repeat array (Invitrogen) that was then clonedbetween Age-I and Xho-I sites cloned into the GoldenGate destination vector by site-directed mutagenesisControl constructs expressing mn variants were made bycloning the nuclease downstream of an NLS and HA tagbetween the Sbf-I and Sal-I restriction sites All con-structs except those used for mRNA production encodeBFP-T2A-nuclease for tracking of nuclease expressionduring flow cytometry An example of a full megaTALprotein sequence (L538-Zn4-Y2 I-AniI) as well as thesequence for each mn tested and the RVDs usedfor each synthetic TAL effector are provided inSupplementary Figure S6
Lentiviral production
Lentivirus (LV) and integration-deficient LV (IDLV) weregenerated as previously described (31) Briefly 9 106
cells were transfected in 10-cm dishes with 6mg of thenuclease construct 3 mg of psPAX2 viral packagingplasmid and 15 mg of either pMD2G (for LV) or integra-tion-deficient D64V pMD2G (for IDLV) viral envelopeplasmid using PEI (Polysciences) Two days after transfec-tion viral supernatants were collected and concentratedby centrifugation Viral titers were determined from thepopulation of blue fluorescent protein (BFP) expressingcells 72 h after transducing 293T cells with varyingvolumes of concentrated virus using polybrene
Cell line derivation
HEK293T cell lines were generated harboring either thetraffic light reporter [TLR 21 (32)] or a modified trafficlight reporter (TLR) containing an near-infraredfluorescent protein (iRFP) gene in place of puromycin[epigenetic TLR (33)] Cells were derived as previouslydescribed with slight modifications Briefly HEK293Tcells were transduced with recombinant LV to yield 5ndash10 transduction based on either iRFP expression (epi-genetic TLR) or cell survival (TLR 21) Approximately 5days after transduction cells were sorted for iRFP+mCherry populations (epigenetic TLR) or mCherrypopulations (TLR 21)
Cell sorting and flow cytometry
Cells were analyzed by flow cytometry on the BD LSRIIand sorted on the BD FACS ARIAII Fluorophores weredetected using the following lasers and filters mCherry mdashexcited 561 nm acquired 61020 mTagBFPmdashexcited405 nm acquired 45050 eGFPmdashexcited 488 nmacquired 52550 iRFPmdashexcited 640 nm acquired 73045 Data were analyzed using FlowJo software
TLR assay
The TLR assay was performed as previously describedwith slight modifications Cells harboring the TLR or epi-genetic TLR 21 were plated at 1ndash20 105 cellswell in a24-well dish 24 h before transfection or transductionXtremeGene9 (Roche) was used at 2 mlmg DNA to trans-fect cells with 05 mg of both nuclease and green fluorescentprotein (GFP) donor constructs Transductions were
2 Nucleic Acids Research 2013
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
performed at an MOI of 5ndash10 using polybrene Cells wereharvested 72 h after transfectiontransduction and read onthe flow cytometer Data were obtained from BFP+(TLR21) or iRFP+BFP+populations (epigenetic TLR)
Western blot analysis
Reporter 293T cells were collected 72 h after treatmentand lysed using radioimmunoprecipitation assay (RIPA)lysis buffer with protease inhibitors and then lysates wererun on a QiaShredder column (Qiagen) Protein concen-trations were determined using the BioRad protein assayreagent and 5ndash10 mg of protein was run on a 10 acryl-amide gel alongside Kaleidoscope protein ladder (BioRad)then transferred onto an Immobilo-FL membrane(Millipore) using semidry transfer The membrane wasblocked using Licor blocking buffer before staining withprimary rabbit anti-HA (Cell Signaling Technology) andmouse anti-b-actin monoclonal antibodies and secondaryPE-conjugated anti-mouse (Medial amp BiologicalLaboratories) and GFP-conjugated anti-rabbit(Invitrogen) monoclonal antibodies Membrane fluores-cence was then detected using the Licor Odyssey
Polymerase chain reaction analysis of TAL effector size
Reporter 293T cells were collected 72 h after lentiviraltransduction and genomic DNA was extracted fromthese cells using the Qiagen Blood and Tissue KitNested polymerase chain reaction (PCR) was performedusing two sets of primers to amplify the TAL effectorrepeats from the integrated megaTAL ORFs as well asthe original megaTAL constructs for comparison ThePCR products from the LV-treated samples were clonedinto the pJET vector (Clonejet) and transformed intoDH5a cells For each sample 47 colonies were pickedand subjected to colony PCR to amplify the pJETinsert PCR products were run on a 12 agarose gelfor size determination
In vitro production of synthetic mRNA
We used the T7-Scribe kit (CellScript) for production ofsynthetic mRNA Two different templates were utilizedfor in vitro transcription a linearized plasmid (megaTALand HE) or a PCR product containing a T7 promoter(Trex2) generated with the following primers Trex2Forward GGATCCTAATACGACTCACTATAGGGGCCGCCACCATGTCTGAGCCACCTCGGGCTGAGTrex2 reverse TTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTAGGCTTCGAGGCTTGGACwith T7 promoter underlined The resultant mRNA waspurified with the RNeasy miniprep columns (Qiagen) anda 50 cap was enzymatically added using the vacciniacapping system combined with a Cap 20-O-Methyltransferase enzyme (both from NEB) A poly(A)tailing kit (Life Technologies) was used to add apolyadenylation tail to 50-capped mRNA and mRNAwas purified using the RNeasy kit The cappedpolyadenylated mRNA was eluted in sterile water andstored at 20
Human T-cell culture and transfection
We used the EasySEP Human T-cell enrichment kit(Stemcell technologies) to purify T-cells from human per-ipheral blood mononuclear cells obtained from anonym-ous donors at the Puget Sound Blood Center IsolatedT-cells were stimulated with CD3CD28 activationbeads (Life Technologies) based on manufacturerrsquosinstructions Activated T-cells were cultured in RPMImedia supplemented with 10 fetal bovine serum 1GlutaMAX 55mM bmercaptoethanol 100Uml penicil-lin 100mgml streptomycin 10mM HEPES and 10 ngmlIL-2 (Biolegend) The beads were magnetically removedafter 36 h of incubation and the T-cell resuspended in freshculture media for 6ndash12 h before electroporation (48ndash56 htotal culture time before electroporation) The Neontransfection system (Life Technologies) was used for elec-troporation with the 10 ml electroporation kit used for alltransfections T-cells were washed with PBS and resus-pended in Buffer T at 2 107 cellsml with 2 105 cellsused per transfection Approximately 1ndash15 mg of eachindividual mRNA was added to each sample before elec-troporation A maximum of 25 mg of total mRNA wasused for samples that had more than mRNA species(ie megaTAL with Trex2) Cells were electroporatedwith the following conditions three pulses 1400 Vpulse with 10ms pulse width After electroporationcells were immediately dispersed into prewarmedantibotic-free T-cell media with IL2 and cultured for 4ndash6days with regular media changes before analysisAnalysis of the T-cell receptor expression was performedusing the aCD3-AlexaFluor 488 antibody (Biolegendclone SK7)
High-throughput sequencing and analysis
High-throughput sequencing was performed on cellssorted for either BFP expression (293T cells) or whichwere CD3- (T-cells except control whole population)PCR primers were designed to target 150ndash200 bp oneach side of the putative megaTAL cut site and appendIllumina sequencing primer adaptors Amplicons for allsamples including negative controls were then subjectedto an additional round of amplification to appendlibrary barcodes as well as outermost flowcell sequencesThis setup allowed for library indexing at both thebarcode and amplicon level Final libraries weresequenced on 125 runs of a MiSeq (version 2) usingpaired end 250 bp read chemistry with a 9-bp index readReads for each de-multiplexed library were aligned totheir respective amplicon references using the Phasterread aligner (Phil Green personal communication)owing to its superior ability to accurately align readswith large indel events Each aligned read pair was thenindividually genotyped for the presence of indelsStatistical analysis for indel frequency and skewing(bias toward mn or TALE region) were performed usinga permutation analysis (100 000 iterations) and testingagainst the null hypothesis (negative controls or lack ofskew)
Nucleic Acids Research 2013 3
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
Statistical analysis
Error bars on graphs represent sem P-values werecalculated using Studentrsquos one-tailed unpaired t-testto compare activity of megaTALs with their specific mncounterpart (Plt 005 shown as Plt 0005Plt 00005)
RESULTS
Designing a TAL effectormdashmn fusion proteinarchitecture megaTAL
Based on the recently proposed DNA binding mechanismof TAL effectors in which binding is initiated by a seriesof degenerate TALE repeats that lack sequence specificityfollowed by annealing of the sequence-specific RVD array(34ndash36) we chose to evaluate a TALE-mn fusion architec-ture (schematized in Figure 1a) hereafter designated asthe lsquomegaTALrsquo architecture We hypothesized that thisarchitecture would lsquoaddressrsquo the mn which has intrinsicspecificity for a small set of target sites to a single desiredtarget and thus substantially enhance cleavage at that sitewithout nonspecifically enhancing the activity at relatedlsquononaddressedrsquo targetsTo evaluate the feasibility of this approach we
generated an initial set of megaTALs consisting of a syn-thetic TAL effector [L538 (37)] fused through a shortpeptide linker to the mn I-AniI [hereafter WT (38)] aswell as a catalytically inactive I-AniI mutant [hereafterE148D (20)] The nuclease activity of each megaTALwas tested in parallel with the corresponding stand-aloneI-AniI mn using cell lines containing TLRs with a series oftarget sites possessing varying spacers between the TALeffector and mn target sites (Supplementary Figure S1a)In these assays the WT I-AniI megaTAL exhibited up to18- and 24-fold increases in levels of mutagenic NHEJ(mutNHEJ) and gene targeting respectively relative tothe stand-alone WT I-AniI (Figure 1b) ImportantlymegaTAL activity was found to be spacer dependentwith the maximum increase in activity achieved against atarget containing a 7-bp spacer (assuming a 20-bp mntarget) No discernible differences in activity wereobserved between megaTALs constructed with four differ-ent protein linkers (Supplementary Figure S1b)
Effect of TALE array repeat number on megaTALactivity
Because the FokI catalytic domain (used in traditionalTALENs) has low intrinsic affinity for DNA and lackssequence specificity for DNA cleavage (25) both theaffinity and specificity of a TALEN for an intendedtarget site are entirely dependent on the DNA bindingspecificity of the TALE RVD arrays necessitating RVDarray lengths of 13 TALE repeats to avoid toxicityrelated to nonspecific binding (19) In contrast becausemeganucleases have a high degree of both intrinsicaffinity and specificity for a small group of target sites(67) we hypothesized that megaTALs could be con-structed with much shorter RVD array lengths
To evaluate the relationship between TALE RVD arraylength and megaTAL activity we generated I-AniImegaTALs with truncated versions of the L538 TALeffector ranging from 25 to 165 repeats long ThesemegaTALs were tested against their appropriate targeteach with a 7-bp spacer (Supplementary Figure S2a)alongside the stand-alone WT I-AniI mn using the TLRreporter lines generated above (Figure 2a SupplementaryFigure S2b) All megaTALs were functional and noneexhibited any detectable toxicity and a megaTAL madewith only 35 repeats showed a significant increase in levelsof both gene targeting and mutNHEJ Cleavage activityrose in close correlation with the number of TAL effectorrepeats with megaTALs made by fusion of a 75 RVDTALE with the low affinity WT mn reaching nearmaximal activity We observed an idiosyncratic loss ofefficacy with the 65 RVD TALE which subsequentanalyses demonstrated could be attributed to the inter-action between the final RVD and its target base(Supplementary Figure S2c)
By reducing the number of TAL effector repeats presentin a megaTAL we hypothesized that the DNA sequenceencoding the TALE domain could be sufficiently diverged
(b)
mutNHEJ GTWT mn (90nM)
25
scr 2 3 4 5 6 7 8 9 10 12 160
5
10
15
20
Spacer length (bp)
Fol
dov
erW
Tm
n
(a)
5rsquo- ATGTTTCATTACACCTGCAGCTCAGCTTATGAGGAGGTTTCTCTGTAAA TACAAAGTAATGTGGACGTCGAGTCGAATACTCCTCCAAAGAGACATTT -5rsquo
NΔ154 C+63
L538I-Ani I
Figure 1 DNA spacer requirement for megaTAL fusions(a) Schematic representation of a megaTAL The megaTAL architec-ture involves fusion of a TAL effector with truncated N- andC-terminal domains through a short linker to the N-terminus of amn (I-AniI which displays a KD of approximately 90 nM for itscognate target site) Aligned below the megaTAL schematic is theDNA sequence for the L538-I-AniI megaTAL target with a spacerlength of 7 bp the L538 DNA target underlined in green the I-AniIDNA target underlined in blue and the DNA spacer separating the twooutlined with a black box (b) Increase in levels of mutagenic NHEJ(mutNHEJ) and gene targeting (GT) achieved by I-AniI megaTALfusion Results shown are derived from assays of cleavage activityusing TLR reporter 293T cells treated 72 h with the L538-Zn4-WTmegaTAL (mT) or WT I-AniI mn across targets with different spacerlengths or a lsquoscrambledrsquo (scr) I-AniI target Cleavage activity for L538-WT megaTAL for a given spacer length is normalized to the activity ofthe WT mn level (represented by a dashed line) across all cell lines Forall graphs error bars indicate sem and P-value comparing megaTALswith their appropriate mn counterpart is indicated by asterisk(Plt 005 Plt 0005 Plt 00005)
4 Nucleic Acids Research 2013
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
so as to reduce its repetitiveness and thus allow lentiviralpackaging and delivery of the nuclease transgene withoutrecombination of its open reading frame (ORF) We testedthis by constructing a codon diverged megaTAL with a 65RVD array TAL effector (designated CD for lsquocodondivergedrsquo sequence in Supplementary Figure S2d) andcomparing its cleavage activity to a nondiverged(designated GG for lsquogolden gatersquo) version using bothlentiviral delivery and DNA transfection A 16- and 21-fold difference in mutNHEJ levels was observed inreporter cells treated with an lentiviral or integration-de-ficient lentiviral vector carrying the CD or GG megaTAL
version suggesting a difference in either delivery or ex-pression efficiencies (Figure 2b) Western blot analysis ofthe megaTAL and mn treated cells revealed expression ofthe full-length product in diverged megaTAL treated cellswhereas those that received the nondiverged version ex-pressed protein of different sizes (Supplementary Figure2e) This observation was confirmed using clonal PCRanalysis of the TAL effector RVD array from CD andGG megaTAL treated samples which indicated that 97and 20 of the integrated ORFs from the respective cellpopulations encoded megaTAL proteins with intactTALE arrays (Figure 2c)
Effect of megaTALs on mn target affinity andcleavage activity
The most straightforward explanation for the increasedactivity observed with the I-AniI megaTAL comparedwith the stand-alone mn is that the TALE array effectivelyincreases the local concentration of I-AniI by its adjacentcleavage site We hypothesized that fusing TAL effectorDNA binding domains to mns would thus generally act topotentiate the activity of mns toward targets for whichthey had low affinity but not toward those for whichthey displayed slow cleavage kinetics To test this hypoth-esis we further evaluated the behaviors of (i) a series ofmegaTALs generated from mns with varying affinities forthe I-AniI native target and (ii) a single megaTAL attarget sites for which the mn exhibits differing cleavagekineticsWe generated megaTALs using two additional I-AniI
variants (hereafter F13Y and Y2) with slightly increasedaffinities for the native I-AniI target site (KD=90nMfor I-AniI WT KD=30nM for I-AniI F13YKD=8nM for I-AniI Y2) (38) MegaTALs made withthe moderate (F13Y) and high (Y2) affinity mn variantsalso showed significant increases in activity (Figure 3aSupplementary Figure S1b and c) albeit to a lesserdegree (13-fold increase for F13Y 4-fold for Y2)than observed with the original I-AniI megaTAL Thusthe effect of TALE array addressing correlates with theintrinsic affinity of the mn variant low affinity variantsare boosted to a much greater extent than those with in-trinsically higher affinity consistent with the idea that theTAL effector domain influences nuclease activity byincreasing the local concentration of the mn cleavagedomain at its addressed target site Additionally themoderate affinity mn F13Y reached a plateau of activitywith a shorter (45 RVD) TALE array relative to theTALE array length needed (55ndash75 RVD) to maximizethe activity of the lower affinity WT I-AniI mn(Supplementary Figure S2b)We then further defined the TALE lsquoaddressingrsquo effect by
testing the activity of the high affinity Y2mn and L538-Y2megaTAL against a series of DNA target variants thatharbor single base-pair substitutions (relative to thenative I-AniI target) for which the kcat and KM of theY2 variant have previously been determined (Figure 3b)(39) Nonnative targets were chosen for which Y2 displayseither lsquohigh KMrsquo (Y2 exhibits a similar kcat but reducedaffinity relative to the original target site) or lsquolow kcatrsquo
(a)
(b)
(c)
25 35 45 55 65 75 85 105 1650
10
20
30
40
No of RVD array units
Fol
dov
erW
Tm
n
E148D
WT
mn
CDm
T
GGm
TW
T
CDm
T
GGm
TW
T
CDm
T
GGm
T
0
1
2
3
4
5
Per
cent
mar
ked
cells
(mut
NH
EJ)
pDNA LV IDLV
Treatment Intact TALE ORF (ct) Total countCD megaTAL 28 966 29GG megaTAL 4 211 19
Figure 2 RVD array requirement for megaTAL fusions (a) Level ofcleavage activity (assayed by mutNHEJ shown in red and gene tar-geting shown in green) of megaTALs built with TAL effectors withdifferent number of repeat units Cleavage activity of TALE-Zn4-WTmegaTALs built with a varying number of RVD array units wasnormalized to the activity of the WT I-AniI mn (represented by adashed line) across all cell lines tested (b) Level of cleavage activity(assayed by mutNHEJ) of megaTALs built with either a codondiverged or nondiverged 65 RVD TAL effector array delivered toreporter cells by DNA transfection or lentiviral delivery Both thecodon diverged (CD) and nondiverged (GG) megaTALs were able torescue the activity of the WT mn using plasmid DNA (pDNA) trans-fection lentiviral (LV) or integration-deficient lentiviral (IDLV) trans-duction However delivery using lentiviral vectors resulted insignificantly lower activity in cells treated with the nondivergedversion of the megaTAL (c) Proportion of intact (full length) andrecombined (short) megaTALs integrated into the genome of 293Tcells Either the codon diverged (CD) or nondiverged (GG) 65 RVDWT megaTAL were delivered to cells by lentiviral transduction and theviral integrants were assessed using clonal PCR amplification
Nucleic Acids Research 2013 5
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
(Y2 exhibits a reduced kcat but comparable bindingaffinity compared with the original target site) Cleavageactivity was measured using TLR reporter cells withtargets composed of the L538 binding site and the indi-vidual high KM low kcat or wt I-AniI targets separatedby an 8-bp spacer As expected based on its known
biochemical properties the stand-alone Y2mn showedlow or background levels of activity against all six nonna-tive targets In contrast cleavage activity was considerablyincreased (12ndash38-fold for HR 8ndash27-fold for mutNHEJ) inall high KM targets treated with their corresponding lsquoad-dressedrsquo megaTAL while little or no effect was observed
Data obtained from Thyme et al (2009)
Target site wt base kcat (min-1) KM (nM)
wild-type wt 94 10
-9A G 105 117
-8G A 143 129
-6T G 113 72
-1G T 02 48
+2A C 038 10
+3C T 014 104
Y2 megaTALY2 mn Y2 megaTALY2 mn+2A-8G
wt -9A -8G -6T -1G +2A +3C00
05
10
15
20
25
30
80
Per
cent
mar
ked
cells
high K M low k cat
Target
GT Y2 mnGT Y2 mT
mutNHEJ Y2 mnmutNHEJ Y2 mT
E148D megaTAL WT megaTAL F13Y megaTAL Y2 megaTAL
WT mn (90nM) F13Y mn (50nM) Y2 mn (10nM)
scr 2 3 4 5 6 7 8 9 10 12 16
Spacer length
Fol
dov
erY
2 m
n (m
utN
HE
J)
0
1
2
3
4
5
0 102 103 104 105
0
102
103
104
105
00274
0173
mC
herr
y
GFP
BFP
coun
t
744
0 102 103 104 105
0
102
103
104
105
109
799
BFP
coun
t
694
mC
herr
y
GFP0 102 103 104 105
0
102
103
104
105
00376
00188
BFP
coun
t
747
0 102 103 104 105
0
102
103
104
105
00299
00299
BFP
coun
t
683
(a)
(b)
(c) (d)
Figure 3 megaTAL addressing acts by increasing the effective mn concentration at the addressed target site (a) Level of cleavage activity (assayedby mutNHEJ) of megaTALs built with mns with varying affinities in TLR cell lines containing targets with different DNA spacer lengths The levelof mutNHEJ for each megaTAL tested was normalized to the activity of Y2 (represented by green line) in the same TLR cell line The averageactivity of the WT and F13Y mns across each cell line are represented by blue and orange lines respectively (b) Effect of megaTAL addressing onactivity of Y2 I-AniI toward DNA targets for which it exhibits different biochemical properties Left panel Wild-type and singly substituted I-AniItargets and previously determined kcat and KM values Right panel Mutagenic NHEJ and gene targeting of Y2 and Y2 megaTAL at each targetassayed by TLR Representative flow plots for data from (b) are shown for the Y2mn (left column) and megaTAL (right column) against high KM(c) and low kcat (d) targets Numbers inside the gates give the percent of mCherry and GFP positive cells and the inset graphs show BFP positivecells on which the experiment was gated
6 Nucleic Acids Research 2013
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
for the low kcat targets (0ndash3-fold for GT and mutNHEJ)(Figure 3b and d) However megaTAL activity againstthe low kcat target -1G was found to be significantlyincreased compared with mn treatment suggesting thatin some cases lsquoaddressingrsquo may partially compensate forreduced catalytic activity
Increasing on-target cleavage with megaTALs
One of the major shortcomings of current nuclease plat-forms is their potential for off-target cleavage and thenegative effects that this may have on the efficacy orsafety profile of a putative nuclease-based therapeutic ap-plication To evaluate the specificity properties of themegaTAL architecture we tested whether a megaTALcould consistently distinguish between lsquoaddressedrsquo andlsquononaddressedrsquo genomic target sites all cleaved by acommon mn cleavage domain Near-native sites of theI-AniI target (DNA sequences containing three or fewerbase-pair differences from the native I-AniI recognitionsequence) were previously identified in the humangenome using the UCSC BLAT tool (40) The cleavageactivity of the Y2 I-AniI variant mn was assessed at thesenear-natives targets using both episomal and endogenoustargets (data not shown) and the five sites that displayedthe highest levels of cleavage by Y2 in those assays werechosen for further examination (Figure 4a) SyntheticTAL effectors specific to regions upstream of the +9Tand+5A+8T near-native targets were built and fused tothe WT F13Y and Y2 I-AniI mns These megaTALs weretested alongside the stand-alone mn variants in TLRreporter cell lines for each of the five targets The +9Tand+5A+8T megaTALs showed increased levels of genetargeting at their addressed targets only while having noobserved effect on the activity at the remaining sites(Figure 4b) Notably while the addition of a sequence-specific TAL effector domain to the high-affinity Y2mndid not reduce its cleavage at nonaddressed targets (theY2mn and megaTALs show similar levels of GT at off-target sites) treatment with megaTALs made with thelower affinity WT and F13Y mns resulted in levels ofgene targeting at addressed targets equivalent to thatachieved with the Y2 megaTALs but without detectableoff-target activity
To verify that a similar effect would be observed atthe endogenous near-native loci 293T cells were treatedwith +9T megaTALs +5A+8T megaTALs and mnvariants and the level of mutNHEJ was determined ateach near-native site by high-throughput sequencing(Figure 4c Supplementary Figure S3a and b) Theseresults mirrored those from the reporter assay with theWT and F13Y megaTALs achieving indel rates up to75 at their addressed sites while maintaining near-background activity at nonaddressed sites
Highly effective knockout of the T-cell receptor alphagene in primary T-cells with a megaTAL
To verify that the benefits observed when fusing wild-typemns to a TAL effector would be achievable with anengineered mn and could be captured in a primary celltype in a relevant therapeutic application we designed a
variant of the I-OnuI mn to knockout the T-cell receptoralpha (TCRa) gene Knockout of the TCRa gene inhuman T-cells has been proposed as an approach toimprove the safety of T-cells endowed with chimericantigen receptors or heterologous TCRs by preventingthem from generating graft-versus-host disease (41)Thus knockout of TCRa in human T-cells is among themost translationally relevant applications of rare cleavingnuclease technologyIn vitro characterization of the TCRa mn showed that
its target site affinity and cleavage activity were similar tothat of its parent enzyme Specificity profiling of theTCRa mn and subsequent high-throughput in vitrocleavage analysis led to the identification of a numberof potential off-target sites which were cleaved with com-parable efficiency to the desired target (SupplementaryFigure S4a and e) To determine if generation of aTCRa megaTAL would yield improvements in activityand specificity the TCRa mn was fused to a 105 repeatTALE array designed to bind a DNA sequence upstreamof the TCRamn binding site (Figure 5a) T-cells were thentransfected with mRNA encoding either the TCRamegaTAL or the stand-alone mn In order to increasethe efficiency of gene knockout samples were includedwherein either the mn or the megaTAL were co-trans-fected with mRNA encoding Trex2 a 30 to 50 exonucleasethat has been shown to significantly increase rates ofmutNHEJ at mn cleavage sites (4243) Disruption ofthe TCRa gene was measured by staining for CD3 amulti-protein complex that is associated with the ab T-cell receptor Strikingly while disruption of the TCRagene occurred at minimal rates in cells treated with thestand-alone mn (16) the TCRa megaTAL exhibited a20-fold increase in activity at the addressed target site inthe TCRa gene (Figure 5b representative flow plots areshown in Figure 5c) Co-transfection of each nuclease withTrex2 further increased the rate of disruption yieldingrates of TCRa gene disruption consistently exceeding70Both on- and off-target cleavage rates were analyzed by
sequencing of the TCRa locus and putative off-target sites(identified based on in vitro cleavage profiling) in FACSsorted CD3- populations of nucleaseTrex2 treated cells(Figure 5d Supplementary Figure S5andashc) For each treat-ment modification rates at the TCRa locus were expectedto be 50 signifying knockout of the gene at theexpressing allele in conjunction with some additionalrate of modification of the nonexpressing allele InmegaTAL treated cells we observed 704 modificationsuggesting a substantial rate (ca 1 in 5 cells) of modifica-tion of the nonexpressed allele In contrast when weexpressed the megaTAL with Trex2 we observed thatonly 520 of the alleles carried indels and even lowerrates were obtained from the mn-alone treated cells Weattribute the differences in modification rates observed inthe presence and absence of Trex2 to lowered expressionof the nuclease in cells co-transfected with the exonucle-ase The lower modification rates observed in the lowabundance CD3- mn alone treated cells are attributableto contamination of the sorted populations with a largerproportion of background unstained cells The
Nucleic Acids Research 2013 7
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
sequencing analysis also demonstrated that off-targetcleavage in cells treated with the megaTAL alone is ex-tremely low with only one of 22 potential sites exhibitinga rate of cleavage above background in the presence ofTrex2 (OT23) Moreover the megaTAL produced lowerlevels of mutNHEJ at several putative off-target loci thanthe stand-alone mn (ie OT18) mdash this result may reflectstatistical variation in the deep sequencing results or al-ternatively could be accounted for by reduced expressionof the megaTAL nuclease compared with its shorter mncounterpart
DISCUSSION
The megaTAL nuclease architecture described hereaddresses several key limitations of existing nuclease
platforms for therapeutic applications as it enables thegeneration of compact single chain nucleases that are ex-tremely active and specific
MegaTALs exhibit dramatic increases in cleavageactivity compared with their isolated mn cleavage headsan effect that appears to be an outcome of the auxiliaryTAL effector DNA-binding domain docking (lsquoaddress-ingrsquo) the mn adjacent to its target and effectivelyincreasing the rate of enzymendashsubstrate complex forma-tion Furthermore the mn-mediated cleavage producesDNA ends with 30 overhangs that are efficiently processedby the Trex2 exonuclease thus allowing the co-expressionof a megaTAL and Trex2 to achieve unprecedentedly hightargeted modification rates in primary cells of gt70
MegaTALs offer an extreme level of target specificitydue to the combined DNA binding and cleavage
Name Genomic sequence Location
+9T TGTCATGTAAATAACTGTACTGTATGTTGAGGAGGTTTCTCTGTATA Ch9 (-)63127729-63127775
+5A+8T TCTTCTGCTGTTTCATATACCTCTATGTGAGGAGGTTTCTCAGTTAG Ch15(-)69553515-69553561
+2T+9G CATAGGTGACTTTTACTCTCATGATAGTGAGGAGGTTTTTCTGTAGA Ch12(+)84384994-84385040
+1C+5A GTCTTTGAATAAGTCACAATGCTTCTTTGAGGAGGTTCCTCAGTAAA Ch7 (+)28151155-28151201
-2A+1G ATGATG GAAATGTTACACTGAGCTCT TGAGGAGGATGCTCTGTAAA Ch1 (+)223534899-223534945
mut
NH
EJ
by
seq
()
+9T +5A+8T +1C+5A -2A+1G0
20
40
60
80
100
+2T+9G locus harbors an extra mutation in 293T cells data not shown
WT +9T WT MegaTAL +5A+8T WT MegaTALF13Y +9T F13Y MegaTAL +5A+8T F13Y MegaTALY2 +9T Y2 MegaTAL +5A+8T Y2 MegaTAL
TLR cell line
+9T +5A+8T +2T+9G +1C+5A -2A+1G0
1
2
3
Per
cent
GF
P+
cells
(a)
(b)
(c)
Figure 4 megaTAL addressing selectively increases on-target cleavage (a) I-AniI human genomic near-native sequences and their location in thegenome Synthetic TAL effectors were built against the+9T and+5A+8T sequences underlined in green and the I-AniI near-natives sequences areunderlined in blue (b) Level of gene targeting in TLR cells with endogenous human I-AniI near-native targets using lsquoaddressedrsquo+9T and+5A+8TmegaTALs and lsquounaddressedrsquo I-AniI mns mutNHEJ was not measured via TLR due to the presence of a stop codon in some target sites thatprevented flow cytometry readout of these events (c) Mutation rates at endogenous targets using lsquoaddressedrsquo I-AniI megaTALs and lsquounadressedrsquomns The level of mutNHEJ at the endogenous I-AniI near-native targets in 293T cells after 72 h of treatment with mn and megaTAL variants wasdetermined based on MiSeq sequencing results
8 Nucleic Acids Research 2013
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
properties of the protein architecture The activity ofmegaTALs is reliant on recognition of the tandem DNAtargets of the TALE array and the mn respectively ef-fectively extending the length of the target site andreducing the likelihood that more than one target sitewould be present in the human genome Similarly as
low affinity mn cleavage heads are not able to efficientlyaccess target sites to which they are not lsquoaddressedrsquo by theTALE array the potential for cleavage at nonaddressedmn recognition sites can be markedly reduced This is re-flected in the megaTAL used for TCRa gene knockout inprimary human T-cells which achieved 70 knockout
Control (-) TCRα mn TCRα mn + Trex2 TCRα mT + Trex2TCRα MegaTAL
Ch14 (+)23016490-23016529
100
mn megaTAL
0
20
40
60
80
CD
3- c
ells
()
- +- +
BFP-
--Trex2
Nuclease
5rsquo- TAAATCCAGTGACAAGTCTGTCTGCCTATTCACCGATTTT ATTTAGGTCACTGTTCAGACAGACGGATAAGTGGCTAAAA-5rsquo
NΔ154 C+63 TCRα mn
TCRα RVD array
283
0 50K 100K 150K 200K
0102
103
104
105
0866
0 50K 100K 150K 200K
519
0 50K 100K 150K 200K
368
0 50K 100K 150K 200K
756
0 50K 100K 150K 200K
CD
3
FSC
384
704
520
50 1000 0125 025 05 075 1 5 10 25NA indel by seq
Putative off target sitesTCRα
mn
mT
Trex2
+
+
-
-
003 0 0 0 0
006 0
0 05
0 1 2
0 14
0 03 0 0
004
0 02
0 08 0
002 0
0 01
0 11 0
OT 1 2 3 4 5 6 7 8 9 10 11 13 14 15 16 17 18 20 21 22 23 24
(a)
(c)
(d)
(b)
Figure 5 megaTAL targeting TCRa achieves extremely high gene disruption with no detectable off-target cleavage in human primary T-cells(a) Schematic of TCRa megaTAL formed by fusing a synthetic TAL effector built toward the TCRa sequence underlined in green to a designedTCRa mn (target underlined in blue) (b) TCR knockout achieved by parental TCRa mn and TCRa megaTAL in human primary T-cells Humanprimary T-cells were transfected with 10 micrograms of RNA encoding the indicated constructs with or without 10 micrograms of mRNA encodingthe Trex2 exonuclease TCRa gene disruption was assessed by measuring the percentage of cells that transition to being CD3- at 5 days followingtransfection Mutation rates at the endogenous TCRa gene were also assessed by Miseq sequencing of amplicons around the cleavage site andshowed indel rates gt50 suggesting that there was a preference for cleavage and disruption of the expressed TCRa allele particularly when themegaTAL is co-expressed with Trex2 (c) Representative flow cytometry data from transfection of human primary T-cells with TCRa targeting mn ormegaTALs+ Trex2 (d) Heat map showing mutation rates at the endogenous TCRa and putative off-target loci in CD3- primary T-cells treatedwith the TCRa mn or megaTAL+ Trex2 Map values were calculated for each locus by subtracting the background rate of indel formation in thecontrol population from that obtained for each nuclease treatment by Miseq sequencing with the heat map color scale indicated by the key (bottom)Sequencing results across all samples are shown (top) as well as an enlarged view with indel rates at putative off-target (OT) sites in cells treated withthe TCRa megaTAL alone (middle) Values calculated as 0 are given as lsquo0rsquo Sequencing results were not obtained for putative off-targets OT12 andOT19 and are therefore not shown on the map other samples for which sequencing results were not obtained are indicated as lsquoNArsquo
Nucleic Acids Research 2013 9
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
while above background off-target cleavage was limited toa single site (eg Figure 5d and Supplementary Figure 5)As the single off-target cleavage site identified is in anoncoding region of the genome this enzyme is of suffi-cient quality at its present state of development for clinicaluse However a further advantage of the megaTAL archi-tecture is the potential to further lsquotunersquo the mn cleavagehead to specifically avoid off-target cleavage sitesThus we anticipate that iterative refinement ofmegaTALs cleavage heads will allow the eventual identi-fication of megaTALs that couple high efficiency withperfect on target genomic specificityThe megaTAL architecture also possesses several
properties that facilitate efficient delivery for therapeuticapplications MegaTALs can be expressed with a single2 kb ORF smaller than two ZFN halves [22 kb total(11 kb per ZFN half targeting a 9-bp target)] and signifi-cantly shorter than a single TALEN half [gt50 kb total(25 kb per TALEN half targeting a 13-bp target)] orCas9 nuclease (41 kb not including guide RNA) ThusmegaTALs are compatible with efficient vectorization as aDNA mRNA or with any widely used viral vector systemThe capacity to use a TAL effector domain with as few as55 repeats also facilitates codon divergence of the TALeffector repeats to allow packaging into retroviral vectorswhere recombination among TAL effector RVDs duringreverse transcription can be problematic (25) Finally theindependent function of megaTALs due to their singlechain design avoids the possibility of inappropriateheterodimerization that may occur with enzymes basedon dimerization-dependent cleavage domains such asFokI an important advantage for applications requiringmultiplex genetic modifications
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Onlineincluding [44]
ACKNOWLEDGEMENTS
We would like to thank members of the NorthwestGenome Engineering Consortium (NGEC) (httpngec-seattleorg) for their many insightful discussions
FUNDING
National Institutes of Health [RL1CA133832UL1DE019582 R01-HL075453 PL1-HL092557 RL1-HL092553 RL1-HL92554 and U19-AI96111] SeattleChildrenrsquos Center for Immunity and ImmunotherapiesFunding for open access charge NIH [U19-AI96111]
Conflict of interest statement AMS is an employee ofCellectis therapeutics and receives salary and equity com-pensation He is also founder member of the board of dir-ectors and equity holder in Pregenen Inc JJ is an employeeof Pregenen Inc and receives salary and equitycompensation AA is an employee of Pregenen Inc andreceives salary and equity compensation PD is an
employee of Cellectis therapeutics and receives salary andequity compensation AG is an employee of Cellectistherapeutics and receives salary and equity compensation
REFERENCES
1 BakerM (2012) Gene-editing nucleases Nat Methods 9 23ndash262 KimYG ChaJ and ChandrasegaranS (1996) Hybrid
restriction enzymes zinc finger fusions to Fok I cleavage domainProc Natl Acad Sci USA 93 1156ndash1160
3 KlugA (2010) The discovery of zinc fingers and theirapplications in gene regulation and genome manipulation AnnRev Biochem 79 213ndash231
4 ChristianM CermakT DoyleEL SchmidtC ZhangFHummelA BogdanoveAJ and VoytasDF (2010) TargetingDNA double-strand breaks with TAL effector nucleases Genetics186 757ndash761
5 LiT HuangS JiangWZ WrightD SpaldingMHWeeksDP and YangB (2011) TAL nucleases (TALNs) hybridproteins composed of TAL effectors and FokI DNA-cleavagedomain Nucleic Acids Res 39 359ndash372
6 PaquesF and DuchateauP (2007) Meganucleases and DNAdouble-strand break-induced recombination perspectives for genetherapy Curr Gene Ther 7 49ndash66
7 StoddardBL (2011) Homing endonucleases from microbialgenetic invaders to reagents for targeted DNA modificationStructure 19 7ndash15
8 JinekM ChylinskiK FonfaraI HauerM DoudnaJA andCharpentierE (2012) A programmable dual-RNAndashguided DNAendonuclease in adaptive bacterial immunity Science 337816ndash821
9 MaliP YangL EsveltKM AachJ GuellM DiCarloJENorvilleJE and ChurchGM (2013) RNA-guided humangenome engineering via Cas9 Science 339 823ndash826
10 McMahonMA RahdarM and PorteusM (2012) Gene editingnot just for translation anymore Nat Methods 9 28ndash31
11 LieberMR (2010) The mechanism of double-strand DNA breakrepair by the nonhomologous DNA end-joining pathway AnnRev Biochem 79 181ndash211
12 ShrivastavM DE HaroLP and NickoloffJA (2008)Regulation of DNA double-strand break repair pathway choiceCell Res 18 134ndash147
13 Method of the Year 2011 Nat Methods 9 114 MandellJG and BarbasCF (2006) Zinc finger tools custom
DNA-binding domains for transcription factors and nucleasesNucleic Acids Res 34 W516ndashW523
15 MaederML Thibodeau-BegannyS OsiakA WrightDAAnthonyRM EichtingerM JiangT FoleyJE WinfreyRJTownsendJA et al (2008) Rapid lsquoopen-sourcersquo engineering ofcustomized zinc-finger nucleases for highly efficient genemodification Mol Cell 31 294ndash301
16 SanderJD DahlborgEJ GoodwinMJ CadeL ZhangFCifuentesD CurtinSJ BlackburnJS Thibodeau-BegannySQiY et al (2011) Selection-free zinc-finger-nuclease engineeringby context-dependent assembly (CoDA) Nat Methods 8 67ndash69
17 CermakT DoyleEL ChristianM WangL ZhangYSchmidtC BallerJA SomiaNV BogdanoveAJ andVoytasDF (2011) Efficient design and assembly of customTALEN and other TAL effector-based constructs for DNAtargeting Nucleic Acids Res 39 e82ndashe82
18 ZhangF CongL LodatoS KosuriS ChurchGM andArlottaP (2011) Efficient construction of sequence-specific TALeffectors for modulating mammalian transcription NatBiotechnol 29 149ndash153
19 ReyonD TsaiSQ KhayterC FodenJA SanderJD andJoungJK (2012) FLASH assembly of TALENs for high-throughput genome editing Nat Biotechnol 30 460ndash465
20 JarjourJ West-FoyleH CertoMT HubertCG DoyleLGetzMM StoddardBL and ScharenbergAM (2009) High-resolution profiling of homing endonuclease binding and catalyticspecificity using yeast surface display Nucleic Acids Res 376871ndash6880
10 Nucleic Acids Research 2013
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
21 CornuTI Thibodeau-BegannyS GuhlE AlwinSEichtingerM JoungJK JoungJK and CathomenT (2008)DNA-binding specificity is a major determinant of the activityand toxicity of zinc-finger nucleases Mol Ther 16 352ndash358
22 GabrielR LombardoA ArensA MillerJC GenovesePKaeppelC NowrouziA BartholomaeCC WangJFriedmanG et al (2011) An unbiased genome-wide analysis ofzinc-finger nuclease specificity Nat Biotechnol 29 816ndash823
23 PattanayakV RamirezCL JoungJK and LiuDR (2011)Revealing off-target cleavage specificities of zinc-finger nucleasesby in vitro selection Nat Methods 8 765ndash770
24 SolluC ParsK CornuTI Thibodeau-BegannySMaederML JoungJK HeilbronnR and CathomenT (2010)Autonomous zinc-finger nuclease pairs for targeted chromosomaldeletion Nucleic Acids Res 38 8269ndash8276
25 HolkersM MaggioI LiuJ JanssenJM MiselliFMussolinoC RecchiaA CathomenT and GoncalvesMA(2012) Differential integrity of TALE nuclease genes followingadenoviral and lentiviral vector gene transfer into human cellsNucleic Acids Res 41 e63
26 FuY FodenJA KhayterC MaederML ReyonDJoungJK and SanderJD (2013) High-frequency off-targetmutagenesis induced by CRISPR-Cas nucleases in human cellsNat Biotechnol 31 822ndash826
27 CradickTJ FineEJ AnticoCJ and BaoG (2013) CRISPRCas9 systems targeting b-globin and CCR5 genes have substantialoff-target activity Nucleic Acids Res 41 9584ndash9592
28 RanFA HsuPD LinCY GootenbergJS KonermannSTrevinoAE ScottDA InoueA MatobaS ZhangY et al(2013) Double nicking by RNA-guided CRISPR Cas9 forenhanced genome editing specificity Cell 154 1380ndash1389
29 AshworthJ TaylorGK HavranekJJ QuadriSAStoddardBL and BakerD (2010) Computationalreprogramming of homing endonuclease specificity at multipleadjacent base pairs Nucleic Acids Res 38 5601ndash5608
30 TakeuchiR LambertAR MakAN JacobyK DicksonRJGloorGB ScharenbergAM EdgellDR and StoddardBL(2011) Tapping natural reservoirs of homing endonucleases fortargeted gene modification Proc Natl Acad Sci USA 10813077ndash13082
31 SatherBD RyuBY StirlingBV GaribovM KernsHMHumblet-BaronS AstrakhanA and RawlingsDJ (2011)Development of B-lineage predominant lentiviral vectors for usein genetic therapies for b cell disorders Mol Ther 19 515ndash525
32 CertoMT RyuBY AnnisJE GaribovM JarjourJRawlingsDJ and ScharenbergAM (2011) Tracking genomeengineering outcome at individual DNA breakpoints NatMethods 8 671ndash676
33 KuharR GwiazdaKS HumbertO MandtT PangalloJBraultM KhanI MaizelsN RawlingsDJ ScharenbergAMet al (2013) Novel fluorescent genome editing reporters formonitoring DNA repair pathway utilization at endonuclease-induced breaks Nucleic Acids Res (epub ahead of print October10 2013)
34 BochJ ScholzeH SchornackS LandgrafA HahnS KaySLahayeT NickstadtA and BonasU (2009) Breaking the codeof DNA binding specificity of TAL-type III effectors Science326 1509ndash1512
35 MoscouMJ and BogdanoveAJ (2009) A simple cipher governsDNA recognition by TAL effectors Science 326 1501ndash1501
36 GaoH WuX ChaiJ and HanZ (2012) Crystal structure of aTALE protein reveals an extended N-terminal DNA bindingregion Cell Res 22 1716ndash1720
37 MillerJC TanS QiaoG BarlowKA WangJ XiaDFMengX PaschonDE LeungE HinkleySJ et al (2011) ATALE nuclease architecture for efficient genome editing NatBiotechnol 29 143ndash148
38 TakeuchiR CertoM CapraraMG ScharenbergAM andStoddardBL (2009) Optimization of in vivo activity of abifunctional homing endonuclease and maturase reversesevolutionary degradation Nucleic Acids Res 37 877ndash890
39 ThymeSB JarjourJ TakeuchiR HavranekJJ AshworthJScharenbergAM StoddardBL and BakerD (2009)Exploitation of binding energy for catalysis and design Nature461 1300ndash1304
40 KentWJ (2002) BLATmdashthe BLAST-like alignment tool GenomeRes 12 656ndash664
41 TorikaiH ReikA LiuPQ ZhouY ZhangL MaitiSHulsH MillerJC KebriaeiP RabinovitchB et al (2012) Afoundation for universal T-cell based immunotherapy T cellsengineered to express a CD19-specific chimeric-antigen-receptorand eliminate expression of endogenous TCR Blood 1195697ndash5705
42 CertoMT GwiazdaKS KuharR SatherB CuringaGMandtT BraultM LambertAR BaxterSK JacobyK et al(2012) Coupling endonucleases with DNA end-processing enzymesto drive gene disruption Nat Methods 9 973ndash975
43 DelacoteF PerezC GuyotV DuhamelM RochonCOllivierN MacmasterR SilvaGH PaquesF DaboussiFet al (2013) High frequency targeted mutagenesis usingengineered endonucleases and DNA-end processing enzymesPLoS One 8 e53217
44 BaxterS LambertAR KuharR JarjourJ KulshinaNParmeggianiF DanaherP GanoJ BakerD StoddardBLet al (2012) Engineering domain fusion chimeras from I-OnuIfamily LAGLIDADG homing endonucleases Nucleic Acids Res40 7985ndash8000
Nucleic Acids Research 2013 11
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from

used in ZFNs and TALENs is activated via dimerizationZFN and TALEN technologies require the constructionand delivery of two protein halves each comprising adistinct DNA binding domain fused to FokI for asingle target site This architectural requirement limitsthe potential for ZFN and TALEN delivery intoprimary cells using viral vectors as well as their simultan-eous (ie multiplexed) delivery of two or more nucleases tomodify more than one gene (2425) Although CRISPRsystems have shown exciting promise in developingreagents for genome engineering with easily designedtarget recognition they exhibit high off-target cleavagewhen used with single-guide RNAs thus necessitatingthe use of nicking enzymes and dual guide RNAs pertarget to achieve specificity equivalent to that of theother major platforms (26ndash28) The consequent need todeliver both the Cas9 ORF plus two guides per targetwould appear to pose significant limitations for the useof CRISPR in therapeutic applications requiring viral vec-torization as well as for applications requiring multiplex-ing Finally although architecturally well suited fortherapeutic applications mnsrsquo tightly coupled cleavageand binding activity have limited their potential for re-design toward novel DNA target sites with the majorityof redesign efforts resulting in respecified enzymes withlow target site affinity and consequent low overallactivity (2930)To address the limitations of present nuclease plat-
forms we have developed a hybrid nuclease architecturethat combines the ease of engineerability of a TALeffector (TALE) with the cleavage sequence specificity ofa mn cleavage domain This lsquomegaTALrsquo architecture wasachieved by fusing minimal TAL effector domains to theN-terminus of mns derived from the LAGLIDADGhoming endonuclease family
MATERIALS AND METHODS
megaTAL and mn construct generation
MegaTALs were constructed using the Golden Gateassembly strategy previously described by Cermak et alusing an repeat variable diresidue (RVD) plasmid libraryand destination vector generously provided by the Voytaslab (17) Plasmids were modified to allow the assembly ofTAL effectors 15ndash105 RVDs long The pthX01 destin-ation vector was modified to include a hemagglutinin(HA) tag immediately downstream of the nuclearlocalization signal (NLS) and to yield a N154 C+63TALEN scaffold TAL effectors were built using the fol-lowing RVDs to target each specific nucleotide AmdashNICmdashHD GmdashNN and TmdashNG Following cloning of theTAL effector repeats into the destination vector theprotein linkers (Fok SQLVKS Zn4 VGGS Flex1GGSG or Flex2 GGSGGGSG) and I-AniI or I-OnuImn variants were cloned in place of the FokI nucleasecatalytic domain between the Xba-I and Sal-I restrictionsites All megaTALs except those used to compare linkeractivity were made using the Zn4 protein linker Thecodon diverged 65C-WT megaTAL (SupplementaryFigure S2d) was constructed by gene synthesizing a TAL
effector repeat array (Invitrogen) that was then clonedbetween Age-I and Xho-I sites cloned into the GoldenGate destination vector by site-directed mutagenesisControl constructs expressing mn variants were made bycloning the nuclease downstream of an NLS and HA tagbetween the Sbf-I and Sal-I restriction sites All con-structs except those used for mRNA production encodeBFP-T2A-nuclease for tracking of nuclease expressionduring flow cytometry An example of a full megaTALprotein sequence (L538-Zn4-Y2 I-AniI) as well as thesequence for each mn tested and the RVDs usedfor each synthetic TAL effector are provided inSupplementary Figure S6
Lentiviral production
Lentivirus (LV) and integration-deficient LV (IDLV) weregenerated as previously described (31) Briefly 9 106
cells were transfected in 10-cm dishes with 6mg of thenuclease construct 3 mg of psPAX2 viral packagingplasmid and 15 mg of either pMD2G (for LV) or integra-tion-deficient D64V pMD2G (for IDLV) viral envelopeplasmid using PEI (Polysciences) Two days after transfec-tion viral supernatants were collected and concentratedby centrifugation Viral titers were determined from thepopulation of blue fluorescent protein (BFP) expressingcells 72 h after transducing 293T cells with varyingvolumes of concentrated virus using polybrene
Cell line derivation
HEK293T cell lines were generated harboring either thetraffic light reporter [TLR 21 (32)] or a modified trafficlight reporter (TLR) containing an near-infraredfluorescent protein (iRFP) gene in place of puromycin[epigenetic TLR (33)] Cells were derived as previouslydescribed with slight modifications Briefly HEK293Tcells were transduced with recombinant LV to yield 5ndash10 transduction based on either iRFP expression (epi-genetic TLR) or cell survival (TLR 21) Approximately 5days after transduction cells were sorted for iRFP+mCherry populations (epigenetic TLR) or mCherrypopulations (TLR 21)
Cell sorting and flow cytometry
Cells were analyzed by flow cytometry on the BD LSRIIand sorted on the BD FACS ARIAII Fluorophores weredetected using the following lasers and filters mCherry mdashexcited 561 nm acquired 61020 mTagBFPmdashexcited405 nm acquired 45050 eGFPmdashexcited 488 nmacquired 52550 iRFPmdashexcited 640 nm acquired 73045 Data were analyzed using FlowJo software
TLR assay
The TLR assay was performed as previously describedwith slight modifications Cells harboring the TLR or epi-genetic TLR 21 were plated at 1ndash20 105 cellswell in a24-well dish 24 h before transfection or transductionXtremeGene9 (Roche) was used at 2 mlmg DNA to trans-fect cells with 05 mg of both nuclease and green fluorescentprotein (GFP) donor constructs Transductions were
2 Nucleic Acids Research 2013
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
performed at an MOI of 5ndash10 using polybrene Cells wereharvested 72 h after transfectiontransduction and read onthe flow cytometer Data were obtained from BFP+(TLR21) or iRFP+BFP+populations (epigenetic TLR)
Western blot analysis
Reporter 293T cells were collected 72 h after treatmentand lysed using radioimmunoprecipitation assay (RIPA)lysis buffer with protease inhibitors and then lysates wererun on a QiaShredder column (Qiagen) Protein concen-trations were determined using the BioRad protein assayreagent and 5ndash10 mg of protein was run on a 10 acryl-amide gel alongside Kaleidoscope protein ladder (BioRad)then transferred onto an Immobilo-FL membrane(Millipore) using semidry transfer The membrane wasblocked using Licor blocking buffer before staining withprimary rabbit anti-HA (Cell Signaling Technology) andmouse anti-b-actin monoclonal antibodies and secondaryPE-conjugated anti-mouse (Medial amp BiologicalLaboratories) and GFP-conjugated anti-rabbit(Invitrogen) monoclonal antibodies Membrane fluores-cence was then detected using the Licor Odyssey
Polymerase chain reaction analysis of TAL effector size
Reporter 293T cells were collected 72 h after lentiviraltransduction and genomic DNA was extracted fromthese cells using the Qiagen Blood and Tissue KitNested polymerase chain reaction (PCR) was performedusing two sets of primers to amplify the TAL effectorrepeats from the integrated megaTAL ORFs as well asthe original megaTAL constructs for comparison ThePCR products from the LV-treated samples were clonedinto the pJET vector (Clonejet) and transformed intoDH5a cells For each sample 47 colonies were pickedand subjected to colony PCR to amplify the pJETinsert PCR products were run on a 12 agarose gelfor size determination
In vitro production of synthetic mRNA
We used the T7-Scribe kit (CellScript) for production ofsynthetic mRNA Two different templates were utilizedfor in vitro transcription a linearized plasmid (megaTALand HE) or a PCR product containing a T7 promoter(Trex2) generated with the following primers Trex2Forward GGATCCTAATACGACTCACTATAGGGGCCGCCACCATGTCTGAGCCACCTCGGGCTGAGTrex2 reverse TTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTAGGCTTCGAGGCTTGGACwith T7 promoter underlined The resultant mRNA waspurified with the RNeasy miniprep columns (Qiagen) anda 50 cap was enzymatically added using the vacciniacapping system combined with a Cap 20-O-Methyltransferase enzyme (both from NEB) A poly(A)tailing kit (Life Technologies) was used to add apolyadenylation tail to 50-capped mRNA and mRNAwas purified using the RNeasy kit The cappedpolyadenylated mRNA was eluted in sterile water andstored at 20
Human T-cell culture and transfection
We used the EasySEP Human T-cell enrichment kit(Stemcell technologies) to purify T-cells from human per-ipheral blood mononuclear cells obtained from anonym-ous donors at the Puget Sound Blood Center IsolatedT-cells were stimulated with CD3CD28 activationbeads (Life Technologies) based on manufacturerrsquosinstructions Activated T-cells were cultured in RPMImedia supplemented with 10 fetal bovine serum 1GlutaMAX 55mM bmercaptoethanol 100Uml penicil-lin 100mgml streptomycin 10mM HEPES and 10 ngmlIL-2 (Biolegend) The beads were magnetically removedafter 36 h of incubation and the T-cell resuspended in freshculture media for 6ndash12 h before electroporation (48ndash56 htotal culture time before electroporation) The Neontransfection system (Life Technologies) was used for elec-troporation with the 10 ml electroporation kit used for alltransfections T-cells were washed with PBS and resus-pended in Buffer T at 2 107 cellsml with 2 105 cellsused per transfection Approximately 1ndash15 mg of eachindividual mRNA was added to each sample before elec-troporation A maximum of 25 mg of total mRNA wasused for samples that had more than mRNA species(ie megaTAL with Trex2) Cells were electroporatedwith the following conditions three pulses 1400 Vpulse with 10ms pulse width After electroporationcells were immediately dispersed into prewarmedantibotic-free T-cell media with IL2 and cultured for 4ndash6days with regular media changes before analysisAnalysis of the T-cell receptor expression was performedusing the aCD3-AlexaFluor 488 antibody (Biolegendclone SK7)
High-throughput sequencing and analysis
High-throughput sequencing was performed on cellssorted for either BFP expression (293T cells) or whichwere CD3- (T-cells except control whole population)PCR primers were designed to target 150ndash200 bp oneach side of the putative megaTAL cut site and appendIllumina sequencing primer adaptors Amplicons for allsamples including negative controls were then subjectedto an additional round of amplification to appendlibrary barcodes as well as outermost flowcell sequencesThis setup allowed for library indexing at both thebarcode and amplicon level Final libraries weresequenced on 125 runs of a MiSeq (version 2) usingpaired end 250 bp read chemistry with a 9-bp index readReads for each de-multiplexed library were aligned totheir respective amplicon references using the Phasterread aligner (Phil Green personal communication)owing to its superior ability to accurately align readswith large indel events Each aligned read pair was thenindividually genotyped for the presence of indelsStatistical analysis for indel frequency and skewing(bias toward mn or TALE region) were performed usinga permutation analysis (100 000 iterations) and testingagainst the null hypothesis (negative controls or lack ofskew)
Nucleic Acids Research 2013 3
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
Statistical analysis
Error bars on graphs represent sem P-values werecalculated using Studentrsquos one-tailed unpaired t-testto compare activity of megaTALs with their specific mncounterpart (Plt 005 shown as Plt 0005Plt 00005)
RESULTS
Designing a TAL effectormdashmn fusion proteinarchitecture megaTAL
Based on the recently proposed DNA binding mechanismof TAL effectors in which binding is initiated by a seriesof degenerate TALE repeats that lack sequence specificityfollowed by annealing of the sequence-specific RVD array(34ndash36) we chose to evaluate a TALE-mn fusion architec-ture (schematized in Figure 1a) hereafter designated asthe lsquomegaTALrsquo architecture We hypothesized that thisarchitecture would lsquoaddressrsquo the mn which has intrinsicspecificity for a small set of target sites to a single desiredtarget and thus substantially enhance cleavage at that sitewithout nonspecifically enhancing the activity at relatedlsquononaddressedrsquo targetsTo evaluate the feasibility of this approach we
generated an initial set of megaTALs consisting of a syn-thetic TAL effector [L538 (37)] fused through a shortpeptide linker to the mn I-AniI [hereafter WT (38)] aswell as a catalytically inactive I-AniI mutant [hereafterE148D (20)] The nuclease activity of each megaTALwas tested in parallel with the corresponding stand-aloneI-AniI mn using cell lines containing TLRs with a series oftarget sites possessing varying spacers between the TALeffector and mn target sites (Supplementary Figure S1a)In these assays the WT I-AniI megaTAL exhibited up to18- and 24-fold increases in levels of mutagenic NHEJ(mutNHEJ) and gene targeting respectively relative tothe stand-alone WT I-AniI (Figure 1b) ImportantlymegaTAL activity was found to be spacer dependentwith the maximum increase in activity achieved against atarget containing a 7-bp spacer (assuming a 20-bp mntarget) No discernible differences in activity wereobserved between megaTALs constructed with four differ-ent protein linkers (Supplementary Figure S1b)
Effect of TALE array repeat number on megaTALactivity
Because the FokI catalytic domain (used in traditionalTALENs) has low intrinsic affinity for DNA and lackssequence specificity for DNA cleavage (25) both theaffinity and specificity of a TALEN for an intendedtarget site are entirely dependent on the DNA bindingspecificity of the TALE RVD arrays necessitating RVDarray lengths of 13 TALE repeats to avoid toxicityrelated to nonspecific binding (19) In contrast becausemeganucleases have a high degree of both intrinsicaffinity and specificity for a small group of target sites(67) we hypothesized that megaTALs could be con-structed with much shorter RVD array lengths
To evaluate the relationship between TALE RVD arraylength and megaTAL activity we generated I-AniImegaTALs with truncated versions of the L538 TALeffector ranging from 25 to 165 repeats long ThesemegaTALs were tested against their appropriate targeteach with a 7-bp spacer (Supplementary Figure S2a)alongside the stand-alone WT I-AniI mn using the TLRreporter lines generated above (Figure 2a SupplementaryFigure S2b) All megaTALs were functional and noneexhibited any detectable toxicity and a megaTAL madewith only 35 repeats showed a significant increase in levelsof both gene targeting and mutNHEJ Cleavage activityrose in close correlation with the number of TAL effectorrepeats with megaTALs made by fusion of a 75 RVDTALE with the low affinity WT mn reaching nearmaximal activity We observed an idiosyncratic loss ofefficacy with the 65 RVD TALE which subsequentanalyses demonstrated could be attributed to the inter-action between the final RVD and its target base(Supplementary Figure S2c)
By reducing the number of TAL effector repeats presentin a megaTAL we hypothesized that the DNA sequenceencoding the TALE domain could be sufficiently diverged
(b)
mutNHEJ GTWT mn (90nM)
25
scr 2 3 4 5 6 7 8 9 10 12 160
5
10
15
20
Spacer length (bp)
Fol
dov
erW
Tm
n
(a)
5rsquo- ATGTTTCATTACACCTGCAGCTCAGCTTATGAGGAGGTTTCTCTGTAAA TACAAAGTAATGTGGACGTCGAGTCGAATACTCCTCCAAAGAGACATTT -5rsquo
NΔ154 C+63
L538I-Ani I
Figure 1 DNA spacer requirement for megaTAL fusions(a) Schematic representation of a megaTAL The megaTAL architec-ture involves fusion of a TAL effector with truncated N- andC-terminal domains through a short linker to the N-terminus of amn (I-AniI which displays a KD of approximately 90 nM for itscognate target site) Aligned below the megaTAL schematic is theDNA sequence for the L538-I-AniI megaTAL target with a spacerlength of 7 bp the L538 DNA target underlined in green the I-AniIDNA target underlined in blue and the DNA spacer separating the twooutlined with a black box (b) Increase in levels of mutagenic NHEJ(mutNHEJ) and gene targeting (GT) achieved by I-AniI megaTALfusion Results shown are derived from assays of cleavage activityusing TLR reporter 293T cells treated 72 h with the L538-Zn4-WTmegaTAL (mT) or WT I-AniI mn across targets with different spacerlengths or a lsquoscrambledrsquo (scr) I-AniI target Cleavage activity for L538-WT megaTAL for a given spacer length is normalized to the activity ofthe WT mn level (represented by a dashed line) across all cell lines Forall graphs error bars indicate sem and P-value comparing megaTALswith their appropriate mn counterpart is indicated by asterisk(Plt 005 Plt 0005 Plt 00005)
4 Nucleic Acids Research 2013
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
so as to reduce its repetitiveness and thus allow lentiviralpackaging and delivery of the nuclease transgene withoutrecombination of its open reading frame (ORF) We testedthis by constructing a codon diverged megaTAL with a 65RVD array TAL effector (designated CD for lsquocodondivergedrsquo sequence in Supplementary Figure S2d) andcomparing its cleavage activity to a nondiverged(designated GG for lsquogolden gatersquo) version using bothlentiviral delivery and DNA transfection A 16- and 21-fold difference in mutNHEJ levels was observed inreporter cells treated with an lentiviral or integration-de-ficient lentiviral vector carrying the CD or GG megaTAL
version suggesting a difference in either delivery or ex-pression efficiencies (Figure 2b) Western blot analysis ofthe megaTAL and mn treated cells revealed expression ofthe full-length product in diverged megaTAL treated cellswhereas those that received the nondiverged version ex-pressed protein of different sizes (Supplementary Figure2e) This observation was confirmed using clonal PCRanalysis of the TAL effector RVD array from CD andGG megaTAL treated samples which indicated that 97and 20 of the integrated ORFs from the respective cellpopulations encoded megaTAL proteins with intactTALE arrays (Figure 2c)
Effect of megaTALs on mn target affinity andcleavage activity
The most straightforward explanation for the increasedactivity observed with the I-AniI megaTAL comparedwith the stand-alone mn is that the TALE array effectivelyincreases the local concentration of I-AniI by its adjacentcleavage site We hypothesized that fusing TAL effectorDNA binding domains to mns would thus generally act topotentiate the activity of mns toward targets for whichthey had low affinity but not toward those for whichthey displayed slow cleavage kinetics To test this hypoth-esis we further evaluated the behaviors of (i) a series ofmegaTALs generated from mns with varying affinities forthe I-AniI native target and (ii) a single megaTAL attarget sites for which the mn exhibits differing cleavagekineticsWe generated megaTALs using two additional I-AniI
variants (hereafter F13Y and Y2) with slightly increasedaffinities for the native I-AniI target site (KD=90nMfor I-AniI WT KD=30nM for I-AniI F13YKD=8nM for I-AniI Y2) (38) MegaTALs made withthe moderate (F13Y) and high (Y2) affinity mn variantsalso showed significant increases in activity (Figure 3aSupplementary Figure S1b and c) albeit to a lesserdegree (13-fold increase for F13Y 4-fold for Y2)than observed with the original I-AniI megaTAL Thusthe effect of TALE array addressing correlates with theintrinsic affinity of the mn variant low affinity variantsare boosted to a much greater extent than those with in-trinsically higher affinity consistent with the idea that theTAL effector domain influences nuclease activity byincreasing the local concentration of the mn cleavagedomain at its addressed target site Additionally themoderate affinity mn F13Y reached a plateau of activitywith a shorter (45 RVD) TALE array relative to theTALE array length needed (55ndash75 RVD) to maximizethe activity of the lower affinity WT I-AniI mn(Supplementary Figure S2b)We then further defined the TALE lsquoaddressingrsquo effect by
testing the activity of the high affinity Y2mn and L538-Y2megaTAL against a series of DNA target variants thatharbor single base-pair substitutions (relative to thenative I-AniI target) for which the kcat and KM of theY2 variant have previously been determined (Figure 3b)(39) Nonnative targets were chosen for which Y2 displayseither lsquohigh KMrsquo (Y2 exhibits a similar kcat but reducedaffinity relative to the original target site) or lsquolow kcatrsquo
(a)
(b)
(c)
25 35 45 55 65 75 85 105 1650
10
20
30
40
No of RVD array units
Fol
dov
erW
Tm
n
E148D
WT
mn
CDm
T
GGm
TW
T
CDm
T
GGm
TW
T
CDm
T
GGm
T
0
1
2
3
4
5
Per
cent
mar
ked
cells
(mut
NH
EJ)
pDNA LV IDLV
Treatment Intact TALE ORF (ct) Total countCD megaTAL 28 966 29GG megaTAL 4 211 19
Figure 2 RVD array requirement for megaTAL fusions (a) Level ofcleavage activity (assayed by mutNHEJ shown in red and gene tar-geting shown in green) of megaTALs built with TAL effectors withdifferent number of repeat units Cleavage activity of TALE-Zn4-WTmegaTALs built with a varying number of RVD array units wasnormalized to the activity of the WT I-AniI mn (represented by adashed line) across all cell lines tested (b) Level of cleavage activity(assayed by mutNHEJ) of megaTALs built with either a codondiverged or nondiverged 65 RVD TAL effector array delivered toreporter cells by DNA transfection or lentiviral delivery Both thecodon diverged (CD) and nondiverged (GG) megaTALs were able torescue the activity of the WT mn using plasmid DNA (pDNA) trans-fection lentiviral (LV) or integration-deficient lentiviral (IDLV) trans-duction However delivery using lentiviral vectors resulted insignificantly lower activity in cells treated with the nondivergedversion of the megaTAL (c) Proportion of intact (full length) andrecombined (short) megaTALs integrated into the genome of 293Tcells Either the codon diverged (CD) or nondiverged (GG) 65 RVDWT megaTAL were delivered to cells by lentiviral transduction and theviral integrants were assessed using clonal PCR amplification
Nucleic Acids Research 2013 5
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
(Y2 exhibits a reduced kcat but comparable bindingaffinity compared with the original target site) Cleavageactivity was measured using TLR reporter cells withtargets composed of the L538 binding site and the indi-vidual high KM low kcat or wt I-AniI targets separatedby an 8-bp spacer As expected based on its known
biochemical properties the stand-alone Y2mn showedlow or background levels of activity against all six nonna-tive targets In contrast cleavage activity was considerablyincreased (12ndash38-fold for HR 8ndash27-fold for mutNHEJ) inall high KM targets treated with their corresponding lsquoad-dressedrsquo megaTAL while little or no effect was observed
Data obtained from Thyme et al (2009)
Target site wt base kcat (min-1) KM (nM)
wild-type wt 94 10
-9A G 105 117
-8G A 143 129
-6T G 113 72
-1G T 02 48
+2A C 038 10
+3C T 014 104
Y2 megaTALY2 mn Y2 megaTALY2 mn+2A-8G
wt -9A -8G -6T -1G +2A +3C00
05
10
15
20
25
30
80
Per
cent
mar
ked
cells
high K M low k cat
Target
GT Y2 mnGT Y2 mT
mutNHEJ Y2 mnmutNHEJ Y2 mT
E148D megaTAL WT megaTAL F13Y megaTAL Y2 megaTAL
WT mn (90nM) F13Y mn (50nM) Y2 mn (10nM)
scr 2 3 4 5 6 7 8 9 10 12 16
Spacer length
Fol
dov
erY
2 m
n (m
utN
HE
J)
0
1
2
3
4
5
0 102 103 104 105
0
102
103
104
105
00274
0173
mC
herr
y
GFP
BFP
coun
t
744
0 102 103 104 105
0
102
103
104
105
109
799
BFP
coun
t
694
mC
herr
y
GFP0 102 103 104 105
0
102
103
104
105
00376
00188
BFP
coun
t
747
0 102 103 104 105
0
102
103
104
105
00299
00299
BFP
coun
t
683
(a)
(b)
(c) (d)
Figure 3 megaTAL addressing acts by increasing the effective mn concentration at the addressed target site (a) Level of cleavage activity (assayedby mutNHEJ) of megaTALs built with mns with varying affinities in TLR cell lines containing targets with different DNA spacer lengths The levelof mutNHEJ for each megaTAL tested was normalized to the activity of Y2 (represented by green line) in the same TLR cell line The averageactivity of the WT and F13Y mns across each cell line are represented by blue and orange lines respectively (b) Effect of megaTAL addressing onactivity of Y2 I-AniI toward DNA targets for which it exhibits different biochemical properties Left panel Wild-type and singly substituted I-AniItargets and previously determined kcat and KM values Right panel Mutagenic NHEJ and gene targeting of Y2 and Y2 megaTAL at each targetassayed by TLR Representative flow plots for data from (b) are shown for the Y2mn (left column) and megaTAL (right column) against high KM(c) and low kcat (d) targets Numbers inside the gates give the percent of mCherry and GFP positive cells and the inset graphs show BFP positivecells on which the experiment was gated
6 Nucleic Acids Research 2013
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
for the low kcat targets (0ndash3-fold for GT and mutNHEJ)(Figure 3b and d) However megaTAL activity againstthe low kcat target -1G was found to be significantlyincreased compared with mn treatment suggesting thatin some cases lsquoaddressingrsquo may partially compensate forreduced catalytic activity
Increasing on-target cleavage with megaTALs
One of the major shortcomings of current nuclease plat-forms is their potential for off-target cleavage and thenegative effects that this may have on the efficacy orsafety profile of a putative nuclease-based therapeutic ap-plication To evaluate the specificity properties of themegaTAL architecture we tested whether a megaTALcould consistently distinguish between lsquoaddressedrsquo andlsquononaddressedrsquo genomic target sites all cleaved by acommon mn cleavage domain Near-native sites of theI-AniI target (DNA sequences containing three or fewerbase-pair differences from the native I-AniI recognitionsequence) were previously identified in the humangenome using the UCSC BLAT tool (40) The cleavageactivity of the Y2 I-AniI variant mn was assessed at thesenear-natives targets using both episomal and endogenoustargets (data not shown) and the five sites that displayedthe highest levels of cleavage by Y2 in those assays werechosen for further examination (Figure 4a) SyntheticTAL effectors specific to regions upstream of the +9Tand+5A+8T near-native targets were built and fused tothe WT F13Y and Y2 I-AniI mns These megaTALs weretested alongside the stand-alone mn variants in TLRreporter cell lines for each of the five targets The +9Tand+5A+8T megaTALs showed increased levels of genetargeting at their addressed targets only while having noobserved effect on the activity at the remaining sites(Figure 4b) Notably while the addition of a sequence-specific TAL effector domain to the high-affinity Y2mndid not reduce its cleavage at nonaddressed targets (theY2mn and megaTALs show similar levels of GT at off-target sites) treatment with megaTALs made with thelower affinity WT and F13Y mns resulted in levels ofgene targeting at addressed targets equivalent to thatachieved with the Y2 megaTALs but without detectableoff-target activity
To verify that a similar effect would be observed atthe endogenous near-native loci 293T cells were treatedwith +9T megaTALs +5A+8T megaTALs and mnvariants and the level of mutNHEJ was determined ateach near-native site by high-throughput sequencing(Figure 4c Supplementary Figure S3a and b) Theseresults mirrored those from the reporter assay with theWT and F13Y megaTALs achieving indel rates up to75 at their addressed sites while maintaining near-background activity at nonaddressed sites
Highly effective knockout of the T-cell receptor alphagene in primary T-cells with a megaTAL
To verify that the benefits observed when fusing wild-typemns to a TAL effector would be achievable with anengineered mn and could be captured in a primary celltype in a relevant therapeutic application we designed a
variant of the I-OnuI mn to knockout the T-cell receptoralpha (TCRa) gene Knockout of the TCRa gene inhuman T-cells has been proposed as an approach toimprove the safety of T-cells endowed with chimericantigen receptors or heterologous TCRs by preventingthem from generating graft-versus-host disease (41)Thus knockout of TCRa in human T-cells is among themost translationally relevant applications of rare cleavingnuclease technologyIn vitro characterization of the TCRa mn showed that
its target site affinity and cleavage activity were similar tothat of its parent enzyme Specificity profiling of theTCRa mn and subsequent high-throughput in vitrocleavage analysis led to the identification of a numberof potential off-target sites which were cleaved with com-parable efficiency to the desired target (SupplementaryFigure S4a and e) To determine if generation of aTCRa megaTAL would yield improvements in activityand specificity the TCRa mn was fused to a 105 repeatTALE array designed to bind a DNA sequence upstreamof the TCRamn binding site (Figure 5a) T-cells were thentransfected with mRNA encoding either the TCRamegaTAL or the stand-alone mn In order to increasethe efficiency of gene knockout samples were includedwherein either the mn or the megaTAL were co-trans-fected with mRNA encoding Trex2 a 30 to 50 exonucleasethat has been shown to significantly increase rates ofmutNHEJ at mn cleavage sites (4243) Disruption ofthe TCRa gene was measured by staining for CD3 amulti-protein complex that is associated with the ab T-cell receptor Strikingly while disruption of the TCRagene occurred at minimal rates in cells treated with thestand-alone mn (16) the TCRa megaTAL exhibited a20-fold increase in activity at the addressed target site inthe TCRa gene (Figure 5b representative flow plots areshown in Figure 5c) Co-transfection of each nuclease withTrex2 further increased the rate of disruption yieldingrates of TCRa gene disruption consistently exceeding70Both on- and off-target cleavage rates were analyzed by
sequencing of the TCRa locus and putative off-target sites(identified based on in vitro cleavage profiling) in FACSsorted CD3- populations of nucleaseTrex2 treated cells(Figure 5d Supplementary Figure S5andashc) For each treat-ment modification rates at the TCRa locus were expectedto be 50 signifying knockout of the gene at theexpressing allele in conjunction with some additionalrate of modification of the nonexpressing allele InmegaTAL treated cells we observed 704 modificationsuggesting a substantial rate (ca 1 in 5 cells) of modifica-tion of the nonexpressed allele In contrast when weexpressed the megaTAL with Trex2 we observed thatonly 520 of the alleles carried indels and even lowerrates were obtained from the mn-alone treated cells Weattribute the differences in modification rates observed inthe presence and absence of Trex2 to lowered expressionof the nuclease in cells co-transfected with the exonucle-ase The lower modification rates observed in the lowabundance CD3- mn alone treated cells are attributableto contamination of the sorted populations with a largerproportion of background unstained cells The
Nucleic Acids Research 2013 7
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
sequencing analysis also demonstrated that off-targetcleavage in cells treated with the megaTAL alone is ex-tremely low with only one of 22 potential sites exhibitinga rate of cleavage above background in the presence ofTrex2 (OT23) Moreover the megaTAL produced lowerlevels of mutNHEJ at several putative off-target loci thanthe stand-alone mn (ie OT18) mdash this result may reflectstatistical variation in the deep sequencing results or al-ternatively could be accounted for by reduced expressionof the megaTAL nuclease compared with its shorter mncounterpart
DISCUSSION
The megaTAL nuclease architecture described hereaddresses several key limitations of existing nuclease
platforms for therapeutic applications as it enables thegeneration of compact single chain nucleases that are ex-tremely active and specific
MegaTALs exhibit dramatic increases in cleavageactivity compared with their isolated mn cleavage headsan effect that appears to be an outcome of the auxiliaryTAL effector DNA-binding domain docking (lsquoaddress-ingrsquo) the mn adjacent to its target and effectivelyincreasing the rate of enzymendashsubstrate complex forma-tion Furthermore the mn-mediated cleavage producesDNA ends with 30 overhangs that are efficiently processedby the Trex2 exonuclease thus allowing the co-expressionof a megaTAL and Trex2 to achieve unprecedentedly hightargeted modification rates in primary cells of gt70
MegaTALs offer an extreme level of target specificitydue to the combined DNA binding and cleavage
Name Genomic sequence Location
+9T TGTCATGTAAATAACTGTACTGTATGTTGAGGAGGTTTCTCTGTATA Ch9 (-)63127729-63127775
+5A+8T TCTTCTGCTGTTTCATATACCTCTATGTGAGGAGGTTTCTCAGTTAG Ch15(-)69553515-69553561
+2T+9G CATAGGTGACTTTTACTCTCATGATAGTGAGGAGGTTTTTCTGTAGA Ch12(+)84384994-84385040
+1C+5A GTCTTTGAATAAGTCACAATGCTTCTTTGAGGAGGTTCCTCAGTAAA Ch7 (+)28151155-28151201
-2A+1G ATGATG GAAATGTTACACTGAGCTCT TGAGGAGGATGCTCTGTAAA Ch1 (+)223534899-223534945
mut
NH
EJ
by
seq
()
+9T +5A+8T +1C+5A -2A+1G0
20
40
60
80
100
+2T+9G locus harbors an extra mutation in 293T cells data not shown
WT +9T WT MegaTAL +5A+8T WT MegaTALF13Y +9T F13Y MegaTAL +5A+8T F13Y MegaTALY2 +9T Y2 MegaTAL +5A+8T Y2 MegaTAL
TLR cell line
+9T +5A+8T +2T+9G +1C+5A -2A+1G0
1
2
3
Per
cent
GF
P+
cells
(a)
(b)
(c)
Figure 4 megaTAL addressing selectively increases on-target cleavage (a) I-AniI human genomic near-native sequences and their location in thegenome Synthetic TAL effectors were built against the+9T and+5A+8T sequences underlined in green and the I-AniI near-natives sequences areunderlined in blue (b) Level of gene targeting in TLR cells with endogenous human I-AniI near-native targets using lsquoaddressedrsquo+9T and+5A+8TmegaTALs and lsquounaddressedrsquo I-AniI mns mutNHEJ was not measured via TLR due to the presence of a stop codon in some target sites thatprevented flow cytometry readout of these events (c) Mutation rates at endogenous targets using lsquoaddressedrsquo I-AniI megaTALs and lsquounadressedrsquomns The level of mutNHEJ at the endogenous I-AniI near-native targets in 293T cells after 72 h of treatment with mn and megaTAL variants wasdetermined based on MiSeq sequencing results
8 Nucleic Acids Research 2013
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
properties of the protein architecture The activity ofmegaTALs is reliant on recognition of the tandem DNAtargets of the TALE array and the mn respectively ef-fectively extending the length of the target site andreducing the likelihood that more than one target sitewould be present in the human genome Similarly as
low affinity mn cleavage heads are not able to efficientlyaccess target sites to which they are not lsquoaddressedrsquo by theTALE array the potential for cleavage at nonaddressedmn recognition sites can be markedly reduced This is re-flected in the megaTAL used for TCRa gene knockout inprimary human T-cells which achieved 70 knockout
Control (-) TCRα mn TCRα mn + Trex2 TCRα mT + Trex2TCRα MegaTAL
Ch14 (+)23016490-23016529
100
mn megaTAL
0
20
40
60
80
CD
3- c
ells
()
- +- +
BFP-
--Trex2
Nuclease
5rsquo- TAAATCCAGTGACAAGTCTGTCTGCCTATTCACCGATTTT ATTTAGGTCACTGTTCAGACAGACGGATAAGTGGCTAAAA-5rsquo
NΔ154 C+63 TCRα mn
TCRα RVD array
283
0 50K 100K 150K 200K
0102
103
104
105
0866
0 50K 100K 150K 200K
519
0 50K 100K 150K 200K
368
0 50K 100K 150K 200K
756
0 50K 100K 150K 200K
CD
3
FSC
384
704
520
50 1000 0125 025 05 075 1 5 10 25NA indel by seq
Putative off target sitesTCRα
mn
mT
Trex2
+
+
-
-
003 0 0 0 0
006 0
0 05
0 1 2
0 14
0 03 0 0
004
0 02
0 08 0
002 0
0 01
0 11 0
OT 1 2 3 4 5 6 7 8 9 10 11 13 14 15 16 17 18 20 21 22 23 24
(a)
(c)
(d)
(b)
Figure 5 megaTAL targeting TCRa achieves extremely high gene disruption with no detectable off-target cleavage in human primary T-cells(a) Schematic of TCRa megaTAL formed by fusing a synthetic TAL effector built toward the TCRa sequence underlined in green to a designedTCRa mn (target underlined in blue) (b) TCR knockout achieved by parental TCRa mn and TCRa megaTAL in human primary T-cells Humanprimary T-cells were transfected with 10 micrograms of RNA encoding the indicated constructs with or without 10 micrograms of mRNA encodingthe Trex2 exonuclease TCRa gene disruption was assessed by measuring the percentage of cells that transition to being CD3- at 5 days followingtransfection Mutation rates at the endogenous TCRa gene were also assessed by Miseq sequencing of amplicons around the cleavage site andshowed indel rates gt50 suggesting that there was a preference for cleavage and disruption of the expressed TCRa allele particularly when themegaTAL is co-expressed with Trex2 (c) Representative flow cytometry data from transfection of human primary T-cells with TCRa targeting mn ormegaTALs+ Trex2 (d) Heat map showing mutation rates at the endogenous TCRa and putative off-target loci in CD3- primary T-cells treatedwith the TCRa mn or megaTAL+ Trex2 Map values were calculated for each locus by subtracting the background rate of indel formation in thecontrol population from that obtained for each nuclease treatment by Miseq sequencing with the heat map color scale indicated by the key (bottom)Sequencing results across all samples are shown (top) as well as an enlarged view with indel rates at putative off-target (OT) sites in cells treated withthe TCRa megaTAL alone (middle) Values calculated as 0 are given as lsquo0rsquo Sequencing results were not obtained for putative off-targets OT12 andOT19 and are therefore not shown on the map other samples for which sequencing results were not obtained are indicated as lsquoNArsquo
Nucleic Acids Research 2013 9
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
while above background off-target cleavage was limited toa single site (eg Figure 5d and Supplementary Figure 5)As the single off-target cleavage site identified is in anoncoding region of the genome this enzyme is of suffi-cient quality at its present state of development for clinicaluse However a further advantage of the megaTAL archi-tecture is the potential to further lsquotunersquo the mn cleavagehead to specifically avoid off-target cleavage sitesThus we anticipate that iterative refinement ofmegaTALs cleavage heads will allow the eventual identi-fication of megaTALs that couple high efficiency withperfect on target genomic specificityThe megaTAL architecture also possesses several
properties that facilitate efficient delivery for therapeuticapplications MegaTALs can be expressed with a single2 kb ORF smaller than two ZFN halves [22 kb total(11 kb per ZFN half targeting a 9-bp target)] and signifi-cantly shorter than a single TALEN half [gt50 kb total(25 kb per TALEN half targeting a 13-bp target)] orCas9 nuclease (41 kb not including guide RNA) ThusmegaTALs are compatible with efficient vectorization as aDNA mRNA or with any widely used viral vector systemThe capacity to use a TAL effector domain with as few as55 repeats also facilitates codon divergence of the TALeffector repeats to allow packaging into retroviral vectorswhere recombination among TAL effector RVDs duringreverse transcription can be problematic (25) Finally theindependent function of megaTALs due to their singlechain design avoids the possibility of inappropriateheterodimerization that may occur with enzymes basedon dimerization-dependent cleavage domains such asFokI an important advantage for applications requiringmultiplex genetic modifications
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Onlineincluding [44]
ACKNOWLEDGEMENTS
We would like to thank members of the NorthwestGenome Engineering Consortium (NGEC) (httpngec-seattleorg) for their many insightful discussions
FUNDING
National Institutes of Health [RL1CA133832UL1DE019582 R01-HL075453 PL1-HL092557 RL1-HL092553 RL1-HL92554 and U19-AI96111] SeattleChildrenrsquos Center for Immunity and ImmunotherapiesFunding for open access charge NIH [U19-AI96111]
Conflict of interest statement AMS is an employee ofCellectis therapeutics and receives salary and equity com-pensation He is also founder member of the board of dir-ectors and equity holder in Pregenen Inc JJ is an employeeof Pregenen Inc and receives salary and equitycompensation AA is an employee of Pregenen Inc andreceives salary and equity compensation PD is an
employee of Cellectis therapeutics and receives salary andequity compensation AG is an employee of Cellectistherapeutics and receives salary and equity compensation
REFERENCES
1 BakerM (2012) Gene-editing nucleases Nat Methods 9 23ndash262 KimYG ChaJ and ChandrasegaranS (1996) Hybrid
restriction enzymes zinc finger fusions to Fok I cleavage domainProc Natl Acad Sci USA 93 1156ndash1160
3 KlugA (2010) The discovery of zinc fingers and theirapplications in gene regulation and genome manipulation AnnRev Biochem 79 213ndash231
4 ChristianM CermakT DoyleEL SchmidtC ZhangFHummelA BogdanoveAJ and VoytasDF (2010) TargetingDNA double-strand breaks with TAL effector nucleases Genetics186 757ndash761
5 LiT HuangS JiangWZ WrightD SpaldingMHWeeksDP and YangB (2011) TAL nucleases (TALNs) hybridproteins composed of TAL effectors and FokI DNA-cleavagedomain Nucleic Acids Res 39 359ndash372
6 PaquesF and DuchateauP (2007) Meganucleases and DNAdouble-strand break-induced recombination perspectives for genetherapy Curr Gene Ther 7 49ndash66
7 StoddardBL (2011) Homing endonucleases from microbialgenetic invaders to reagents for targeted DNA modificationStructure 19 7ndash15
8 JinekM ChylinskiK FonfaraI HauerM DoudnaJA andCharpentierE (2012) A programmable dual-RNAndashguided DNAendonuclease in adaptive bacterial immunity Science 337816ndash821
9 MaliP YangL EsveltKM AachJ GuellM DiCarloJENorvilleJE and ChurchGM (2013) RNA-guided humangenome engineering via Cas9 Science 339 823ndash826
10 McMahonMA RahdarM and PorteusM (2012) Gene editingnot just for translation anymore Nat Methods 9 28ndash31
11 LieberMR (2010) The mechanism of double-strand DNA breakrepair by the nonhomologous DNA end-joining pathway AnnRev Biochem 79 181ndash211
12 ShrivastavM DE HaroLP and NickoloffJA (2008)Regulation of DNA double-strand break repair pathway choiceCell Res 18 134ndash147
13 Method of the Year 2011 Nat Methods 9 114 MandellJG and BarbasCF (2006) Zinc finger tools custom
DNA-binding domains for transcription factors and nucleasesNucleic Acids Res 34 W516ndashW523
15 MaederML Thibodeau-BegannyS OsiakA WrightDAAnthonyRM EichtingerM JiangT FoleyJE WinfreyRJTownsendJA et al (2008) Rapid lsquoopen-sourcersquo engineering ofcustomized zinc-finger nucleases for highly efficient genemodification Mol Cell 31 294ndash301
16 SanderJD DahlborgEJ GoodwinMJ CadeL ZhangFCifuentesD CurtinSJ BlackburnJS Thibodeau-BegannySQiY et al (2011) Selection-free zinc-finger-nuclease engineeringby context-dependent assembly (CoDA) Nat Methods 8 67ndash69
17 CermakT DoyleEL ChristianM WangL ZhangYSchmidtC BallerJA SomiaNV BogdanoveAJ andVoytasDF (2011) Efficient design and assembly of customTALEN and other TAL effector-based constructs for DNAtargeting Nucleic Acids Res 39 e82ndashe82
18 ZhangF CongL LodatoS KosuriS ChurchGM andArlottaP (2011) Efficient construction of sequence-specific TALeffectors for modulating mammalian transcription NatBiotechnol 29 149ndash153
19 ReyonD TsaiSQ KhayterC FodenJA SanderJD andJoungJK (2012) FLASH assembly of TALENs for high-throughput genome editing Nat Biotechnol 30 460ndash465
20 JarjourJ West-FoyleH CertoMT HubertCG DoyleLGetzMM StoddardBL and ScharenbergAM (2009) High-resolution profiling of homing endonuclease binding and catalyticspecificity using yeast surface display Nucleic Acids Res 376871ndash6880
10 Nucleic Acids Research 2013
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
21 CornuTI Thibodeau-BegannyS GuhlE AlwinSEichtingerM JoungJK JoungJK and CathomenT (2008)DNA-binding specificity is a major determinant of the activityand toxicity of zinc-finger nucleases Mol Ther 16 352ndash358
22 GabrielR LombardoA ArensA MillerJC GenovesePKaeppelC NowrouziA BartholomaeCC WangJFriedmanG et al (2011) An unbiased genome-wide analysis ofzinc-finger nuclease specificity Nat Biotechnol 29 816ndash823
23 PattanayakV RamirezCL JoungJK and LiuDR (2011)Revealing off-target cleavage specificities of zinc-finger nucleasesby in vitro selection Nat Methods 8 765ndash770
24 SolluC ParsK CornuTI Thibodeau-BegannySMaederML JoungJK HeilbronnR and CathomenT (2010)Autonomous zinc-finger nuclease pairs for targeted chromosomaldeletion Nucleic Acids Res 38 8269ndash8276
25 HolkersM MaggioI LiuJ JanssenJM MiselliFMussolinoC RecchiaA CathomenT and GoncalvesMA(2012) Differential integrity of TALE nuclease genes followingadenoviral and lentiviral vector gene transfer into human cellsNucleic Acids Res 41 e63
26 FuY FodenJA KhayterC MaederML ReyonDJoungJK and SanderJD (2013) High-frequency off-targetmutagenesis induced by CRISPR-Cas nucleases in human cellsNat Biotechnol 31 822ndash826
27 CradickTJ FineEJ AnticoCJ and BaoG (2013) CRISPRCas9 systems targeting b-globin and CCR5 genes have substantialoff-target activity Nucleic Acids Res 41 9584ndash9592
28 RanFA HsuPD LinCY GootenbergJS KonermannSTrevinoAE ScottDA InoueA MatobaS ZhangY et al(2013) Double nicking by RNA-guided CRISPR Cas9 forenhanced genome editing specificity Cell 154 1380ndash1389
29 AshworthJ TaylorGK HavranekJJ QuadriSAStoddardBL and BakerD (2010) Computationalreprogramming of homing endonuclease specificity at multipleadjacent base pairs Nucleic Acids Res 38 5601ndash5608
30 TakeuchiR LambertAR MakAN JacobyK DicksonRJGloorGB ScharenbergAM EdgellDR and StoddardBL(2011) Tapping natural reservoirs of homing endonucleases fortargeted gene modification Proc Natl Acad Sci USA 10813077ndash13082
31 SatherBD RyuBY StirlingBV GaribovM KernsHMHumblet-BaronS AstrakhanA and RawlingsDJ (2011)Development of B-lineage predominant lentiviral vectors for usein genetic therapies for b cell disorders Mol Ther 19 515ndash525
32 CertoMT RyuBY AnnisJE GaribovM JarjourJRawlingsDJ and ScharenbergAM (2011) Tracking genomeengineering outcome at individual DNA breakpoints NatMethods 8 671ndash676
33 KuharR GwiazdaKS HumbertO MandtT PangalloJBraultM KhanI MaizelsN RawlingsDJ ScharenbergAMet al (2013) Novel fluorescent genome editing reporters formonitoring DNA repair pathway utilization at endonuclease-induced breaks Nucleic Acids Res (epub ahead of print October10 2013)
34 BochJ ScholzeH SchornackS LandgrafA HahnS KaySLahayeT NickstadtA and BonasU (2009) Breaking the codeof DNA binding specificity of TAL-type III effectors Science326 1509ndash1512
35 MoscouMJ and BogdanoveAJ (2009) A simple cipher governsDNA recognition by TAL effectors Science 326 1501ndash1501
36 GaoH WuX ChaiJ and HanZ (2012) Crystal structure of aTALE protein reveals an extended N-terminal DNA bindingregion Cell Res 22 1716ndash1720
37 MillerJC TanS QiaoG BarlowKA WangJ XiaDFMengX PaschonDE LeungE HinkleySJ et al (2011) ATALE nuclease architecture for efficient genome editing NatBiotechnol 29 143ndash148
38 TakeuchiR CertoM CapraraMG ScharenbergAM andStoddardBL (2009) Optimization of in vivo activity of abifunctional homing endonuclease and maturase reversesevolutionary degradation Nucleic Acids Res 37 877ndash890
39 ThymeSB JarjourJ TakeuchiR HavranekJJ AshworthJScharenbergAM StoddardBL and BakerD (2009)Exploitation of binding energy for catalysis and design Nature461 1300ndash1304
40 KentWJ (2002) BLATmdashthe BLAST-like alignment tool GenomeRes 12 656ndash664
41 TorikaiH ReikA LiuPQ ZhouY ZhangL MaitiSHulsH MillerJC KebriaeiP RabinovitchB et al (2012) Afoundation for universal T-cell based immunotherapy T cellsengineered to express a CD19-specific chimeric-antigen-receptorand eliminate expression of endogenous TCR Blood 1195697ndash5705
42 CertoMT GwiazdaKS KuharR SatherB CuringaGMandtT BraultM LambertAR BaxterSK JacobyK et al(2012) Coupling endonucleases with DNA end-processing enzymesto drive gene disruption Nat Methods 9 973ndash975
43 DelacoteF PerezC GuyotV DuhamelM RochonCOllivierN MacmasterR SilvaGH PaquesF DaboussiFet al (2013) High frequency targeted mutagenesis usingengineered endonucleases and DNA-end processing enzymesPLoS One 8 e53217
44 BaxterS LambertAR KuharR JarjourJ KulshinaNParmeggianiF DanaherP GanoJ BakerD StoddardBLet al (2012) Engineering domain fusion chimeras from I-OnuIfamily LAGLIDADG homing endonucleases Nucleic Acids Res40 7985ndash8000
Nucleic Acids Research 2013 11
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from

performed at an MOI of 5ndash10 using polybrene Cells wereharvested 72 h after transfectiontransduction and read onthe flow cytometer Data were obtained from BFP+(TLR21) or iRFP+BFP+populations (epigenetic TLR)
Western blot analysis
Reporter 293T cells were collected 72 h after treatmentand lysed using radioimmunoprecipitation assay (RIPA)lysis buffer with protease inhibitors and then lysates wererun on a QiaShredder column (Qiagen) Protein concen-trations were determined using the BioRad protein assayreagent and 5ndash10 mg of protein was run on a 10 acryl-amide gel alongside Kaleidoscope protein ladder (BioRad)then transferred onto an Immobilo-FL membrane(Millipore) using semidry transfer The membrane wasblocked using Licor blocking buffer before staining withprimary rabbit anti-HA (Cell Signaling Technology) andmouse anti-b-actin monoclonal antibodies and secondaryPE-conjugated anti-mouse (Medial amp BiologicalLaboratories) and GFP-conjugated anti-rabbit(Invitrogen) monoclonal antibodies Membrane fluores-cence was then detected using the Licor Odyssey
Polymerase chain reaction analysis of TAL effector size
Reporter 293T cells were collected 72 h after lentiviraltransduction and genomic DNA was extracted fromthese cells using the Qiagen Blood and Tissue KitNested polymerase chain reaction (PCR) was performedusing two sets of primers to amplify the TAL effectorrepeats from the integrated megaTAL ORFs as well asthe original megaTAL constructs for comparison ThePCR products from the LV-treated samples were clonedinto the pJET vector (Clonejet) and transformed intoDH5a cells For each sample 47 colonies were pickedand subjected to colony PCR to amplify the pJETinsert PCR products were run on a 12 agarose gelfor size determination
In vitro production of synthetic mRNA
We used the T7-Scribe kit (CellScript) for production ofsynthetic mRNA Two different templates were utilizedfor in vitro transcription a linearized plasmid (megaTALand HE) or a PCR product containing a T7 promoter(Trex2) generated with the following primers Trex2Forward GGATCCTAATACGACTCACTATAGGGGCCGCCACCATGTCTGAGCCACCTCGGGCTGAGTrex2 reverse TTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTAGGCTTCGAGGCTTGGACwith T7 promoter underlined The resultant mRNA waspurified with the RNeasy miniprep columns (Qiagen) anda 50 cap was enzymatically added using the vacciniacapping system combined with a Cap 20-O-Methyltransferase enzyme (both from NEB) A poly(A)tailing kit (Life Technologies) was used to add apolyadenylation tail to 50-capped mRNA and mRNAwas purified using the RNeasy kit The cappedpolyadenylated mRNA was eluted in sterile water andstored at 20
Human T-cell culture and transfection
We used the EasySEP Human T-cell enrichment kit(Stemcell technologies) to purify T-cells from human per-ipheral blood mononuclear cells obtained from anonym-ous donors at the Puget Sound Blood Center IsolatedT-cells were stimulated with CD3CD28 activationbeads (Life Technologies) based on manufacturerrsquosinstructions Activated T-cells were cultured in RPMImedia supplemented with 10 fetal bovine serum 1GlutaMAX 55mM bmercaptoethanol 100Uml penicil-lin 100mgml streptomycin 10mM HEPES and 10 ngmlIL-2 (Biolegend) The beads were magnetically removedafter 36 h of incubation and the T-cell resuspended in freshculture media for 6ndash12 h before electroporation (48ndash56 htotal culture time before electroporation) The Neontransfection system (Life Technologies) was used for elec-troporation with the 10 ml electroporation kit used for alltransfections T-cells were washed with PBS and resus-pended in Buffer T at 2 107 cellsml with 2 105 cellsused per transfection Approximately 1ndash15 mg of eachindividual mRNA was added to each sample before elec-troporation A maximum of 25 mg of total mRNA wasused for samples that had more than mRNA species(ie megaTAL with Trex2) Cells were electroporatedwith the following conditions three pulses 1400 Vpulse with 10ms pulse width After electroporationcells were immediately dispersed into prewarmedantibotic-free T-cell media with IL2 and cultured for 4ndash6days with regular media changes before analysisAnalysis of the T-cell receptor expression was performedusing the aCD3-AlexaFluor 488 antibody (Biolegendclone SK7)
High-throughput sequencing and analysis
High-throughput sequencing was performed on cellssorted for either BFP expression (293T cells) or whichwere CD3- (T-cells except control whole population)PCR primers were designed to target 150ndash200 bp oneach side of the putative megaTAL cut site and appendIllumina sequencing primer adaptors Amplicons for allsamples including negative controls were then subjectedto an additional round of amplification to appendlibrary barcodes as well as outermost flowcell sequencesThis setup allowed for library indexing at both thebarcode and amplicon level Final libraries weresequenced on 125 runs of a MiSeq (version 2) usingpaired end 250 bp read chemistry with a 9-bp index readReads for each de-multiplexed library were aligned totheir respective amplicon references using the Phasterread aligner (Phil Green personal communication)owing to its superior ability to accurately align readswith large indel events Each aligned read pair was thenindividually genotyped for the presence of indelsStatistical analysis for indel frequency and skewing(bias toward mn or TALE region) were performed usinga permutation analysis (100 000 iterations) and testingagainst the null hypothesis (negative controls or lack ofskew)
Nucleic Acids Research 2013 3
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
Statistical analysis
Error bars on graphs represent sem P-values werecalculated using Studentrsquos one-tailed unpaired t-testto compare activity of megaTALs with their specific mncounterpart (Plt 005 shown as Plt 0005Plt 00005)
RESULTS
Designing a TAL effectormdashmn fusion proteinarchitecture megaTAL
Based on the recently proposed DNA binding mechanismof TAL effectors in which binding is initiated by a seriesof degenerate TALE repeats that lack sequence specificityfollowed by annealing of the sequence-specific RVD array(34ndash36) we chose to evaluate a TALE-mn fusion architec-ture (schematized in Figure 1a) hereafter designated asthe lsquomegaTALrsquo architecture We hypothesized that thisarchitecture would lsquoaddressrsquo the mn which has intrinsicspecificity for a small set of target sites to a single desiredtarget and thus substantially enhance cleavage at that sitewithout nonspecifically enhancing the activity at relatedlsquononaddressedrsquo targetsTo evaluate the feasibility of this approach we
generated an initial set of megaTALs consisting of a syn-thetic TAL effector [L538 (37)] fused through a shortpeptide linker to the mn I-AniI [hereafter WT (38)] aswell as a catalytically inactive I-AniI mutant [hereafterE148D (20)] The nuclease activity of each megaTALwas tested in parallel with the corresponding stand-aloneI-AniI mn using cell lines containing TLRs with a series oftarget sites possessing varying spacers between the TALeffector and mn target sites (Supplementary Figure S1a)In these assays the WT I-AniI megaTAL exhibited up to18- and 24-fold increases in levels of mutagenic NHEJ(mutNHEJ) and gene targeting respectively relative tothe stand-alone WT I-AniI (Figure 1b) ImportantlymegaTAL activity was found to be spacer dependentwith the maximum increase in activity achieved against atarget containing a 7-bp spacer (assuming a 20-bp mntarget) No discernible differences in activity wereobserved between megaTALs constructed with four differ-ent protein linkers (Supplementary Figure S1b)
Effect of TALE array repeat number on megaTALactivity
Because the FokI catalytic domain (used in traditionalTALENs) has low intrinsic affinity for DNA and lackssequence specificity for DNA cleavage (25) both theaffinity and specificity of a TALEN for an intendedtarget site are entirely dependent on the DNA bindingspecificity of the TALE RVD arrays necessitating RVDarray lengths of 13 TALE repeats to avoid toxicityrelated to nonspecific binding (19) In contrast becausemeganucleases have a high degree of both intrinsicaffinity and specificity for a small group of target sites(67) we hypothesized that megaTALs could be con-structed with much shorter RVD array lengths
To evaluate the relationship between TALE RVD arraylength and megaTAL activity we generated I-AniImegaTALs with truncated versions of the L538 TALeffector ranging from 25 to 165 repeats long ThesemegaTALs were tested against their appropriate targeteach with a 7-bp spacer (Supplementary Figure S2a)alongside the stand-alone WT I-AniI mn using the TLRreporter lines generated above (Figure 2a SupplementaryFigure S2b) All megaTALs were functional and noneexhibited any detectable toxicity and a megaTAL madewith only 35 repeats showed a significant increase in levelsof both gene targeting and mutNHEJ Cleavage activityrose in close correlation with the number of TAL effectorrepeats with megaTALs made by fusion of a 75 RVDTALE with the low affinity WT mn reaching nearmaximal activity We observed an idiosyncratic loss ofefficacy with the 65 RVD TALE which subsequentanalyses demonstrated could be attributed to the inter-action between the final RVD and its target base(Supplementary Figure S2c)
By reducing the number of TAL effector repeats presentin a megaTAL we hypothesized that the DNA sequenceencoding the TALE domain could be sufficiently diverged
(b)
mutNHEJ GTWT mn (90nM)
25
scr 2 3 4 5 6 7 8 9 10 12 160
5
10
15
20
Spacer length (bp)
Fol
dov
erW
Tm
n
(a)
5rsquo- ATGTTTCATTACACCTGCAGCTCAGCTTATGAGGAGGTTTCTCTGTAAA TACAAAGTAATGTGGACGTCGAGTCGAATACTCCTCCAAAGAGACATTT -5rsquo
NΔ154 C+63
L538I-Ani I
Figure 1 DNA spacer requirement for megaTAL fusions(a) Schematic representation of a megaTAL The megaTAL architec-ture involves fusion of a TAL effector with truncated N- andC-terminal domains through a short linker to the N-terminus of amn (I-AniI which displays a KD of approximately 90 nM for itscognate target site) Aligned below the megaTAL schematic is theDNA sequence for the L538-I-AniI megaTAL target with a spacerlength of 7 bp the L538 DNA target underlined in green the I-AniIDNA target underlined in blue and the DNA spacer separating the twooutlined with a black box (b) Increase in levels of mutagenic NHEJ(mutNHEJ) and gene targeting (GT) achieved by I-AniI megaTALfusion Results shown are derived from assays of cleavage activityusing TLR reporter 293T cells treated 72 h with the L538-Zn4-WTmegaTAL (mT) or WT I-AniI mn across targets with different spacerlengths or a lsquoscrambledrsquo (scr) I-AniI target Cleavage activity for L538-WT megaTAL for a given spacer length is normalized to the activity ofthe WT mn level (represented by a dashed line) across all cell lines Forall graphs error bars indicate sem and P-value comparing megaTALswith their appropriate mn counterpart is indicated by asterisk(Plt 005 Plt 0005 Plt 00005)
4 Nucleic Acids Research 2013
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
so as to reduce its repetitiveness and thus allow lentiviralpackaging and delivery of the nuclease transgene withoutrecombination of its open reading frame (ORF) We testedthis by constructing a codon diverged megaTAL with a 65RVD array TAL effector (designated CD for lsquocodondivergedrsquo sequence in Supplementary Figure S2d) andcomparing its cleavage activity to a nondiverged(designated GG for lsquogolden gatersquo) version using bothlentiviral delivery and DNA transfection A 16- and 21-fold difference in mutNHEJ levels was observed inreporter cells treated with an lentiviral or integration-de-ficient lentiviral vector carrying the CD or GG megaTAL
version suggesting a difference in either delivery or ex-pression efficiencies (Figure 2b) Western blot analysis ofthe megaTAL and mn treated cells revealed expression ofthe full-length product in diverged megaTAL treated cellswhereas those that received the nondiverged version ex-pressed protein of different sizes (Supplementary Figure2e) This observation was confirmed using clonal PCRanalysis of the TAL effector RVD array from CD andGG megaTAL treated samples which indicated that 97and 20 of the integrated ORFs from the respective cellpopulations encoded megaTAL proteins with intactTALE arrays (Figure 2c)
Effect of megaTALs on mn target affinity andcleavage activity
The most straightforward explanation for the increasedactivity observed with the I-AniI megaTAL comparedwith the stand-alone mn is that the TALE array effectivelyincreases the local concentration of I-AniI by its adjacentcleavage site We hypothesized that fusing TAL effectorDNA binding domains to mns would thus generally act topotentiate the activity of mns toward targets for whichthey had low affinity but not toward those for whichthey displayed slow cleavage kinetics To test this hypoth-esis we further evaluated the behaviors of (i) a series ofmegaTALs generated from mns with varying affinities forthe I-AniI native target and (ii) a single megaTAL attarget sites for which the mn exhibits differing cleavagekineticsWe generated megaTALs using two additional I-AniI
variants (hereafter F13Y and Y2) with slightly increasedaffinities for the native I-AniI target site (KD=90nMfor I-AniI WT KD=30nM for I-AniI F13YKD=8nM for I-AniI Y2) (38) MegaTALs made withthe moderate (F13Y) and high (Y2) affinity mn variantsalso showed significant increases in activity (Figure 3aSupplementary Figure S1b and c) albeit to a lesserdegree (13-fold increase for F13Y 4-fold for Y2)than observed with the original I-AniI megaTAL Thusthe effect of TALE array addressing correlates with theintrinsic affinity of the mn variant low affinity variantsare boosted to a much greater extent than those with in-trinsically higher affinity consistent with the idea that theTAL effector domain influences nuclease activity byincreasing the local concentration of the mn cleavagedomain at its addressed target site Additionally themoderate affinity mn F13Y reached a plateau of activitywith a shorter (45 RVD) TALE array relative to theTALE array length needed (55ndash75 RVD) to maximizethe activity of the lower affinity WT I-AniI mn(Supplementary Figure S2b)We then further defined the TALE lsquoaddressingrsquo effect by
testing the activity of the high affinity Y2mn and L538-Y2megaTAL against a series of DNA target variants thatharbor single base-pair substitutions (relative to thenative I-AniI target) for which the kcat and KM of theY2 variant have previously been determined (Figure 3b)(39) Nonnative targets were chosen for which Y2 displayseither lsquohigh KMrsquo (Y2 exhibits a similar kcat but reducedaffinity relative to the original target site) or lsquolow kcatrsquo
(a)
(b)
(c)
25 35 45 55 65 75 85 105 1650
10
20
30
40
No of RVD array units
Fol
dov
erW
Tm
n
E148D
WT
mn
CDm
T
GGm
TW
T
CDm
T
GGm
TW
T
CDm
T
GGm
T
0
1
2
3
4
5
Per
cent
mar
ked
cells
(mut
NH
EJ)
pDNA LV IDLV
Treatment Intact TALE ORF (ct) Total countCD megaTAL 28 966 29GG megaTAL 4 211 19
Figure 2 RVD array requirement for megaTAL fusions (a) Level ofcleavage activity (assayed by mutNHEJ shown in red and gene tar-geting shown in green) of megaTALs built with TAL effectors withdifferent number of repeat units Cleavage activity of TALE-Zn4-WTmegaTALs built with a varying number of RVD array units wasnormalized to the activity of the WT I-AniI mn (represented by adashed line) across all cell lines tested (b) Level of cleavage activity(assayed by mutNHEJ) of megaTALs built with either a codondiverged or nondiverged 65 RVD TAL effector array delivered toreporter cells by DNA transfection or lentiviral delivery Both thecodon diverged (CD) and nondiverged (GG) megaTALs were able torescue the activity of the WT mn using plasmid DNA (pDNA) trans-fection lentiviral (LV) or integration-deficient lentiviral (IDLV) trans-duction However delivery using lentiviral vectors resulted insignificantly lower activity in cells treated with the nondivergedversion of the megaTAL (c) Proportion of intact (full length) andrecombined (short) megaTALs integrated into the genome of 293Tcells Either the codon diverged (CD) or nondiverged (GG) 65 RVDWT megaTAL were delivered to cells by lentiviral transduction and theviral integrants were assessed using clonal PCR amplification
Nucleic Acids Research 2013 5
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
(Y2 exhibits a reduced kcat but comparable bindingaffinity compared with the original target site) Cleavageactivity was measured using TLR reporter cells withtargets composed of the L538 binding site and the indi-vidual high KM low kcat or wt I-AniI targets separatedby an 8-bp spacer As expected based on its known
biochemical properties the stand-alone Y2mn showedlow or background levels of activity against all six nonna-tive targets In contrast cleavage activity was considerablyincreased (12ndash38-fold for HR 8ndash27-fold for mutNHEJ) inall high KM targets treated with their corresponding lsquoad-dressedrsquo megaTAL while little or no effect was observed
Data obtained from Thyme et al (2009)
Target site wt base kcat (min-1) KM (nM)
wild-type wt 94 10
-9A G 105 117
-8G A 143 129
-6T G 113 72
-1G T 02 48
+2A C 038 10
+3C T 014 104
Y2 megaTALY2 mn Y2 megaTALY2 mn+2A-8G
wt -9A -8G -6T -1G +2A +3C00
05
10
15
20
25
30
80
Per
cent
mar
ked
cells
high K M low k cat
Target
GT Y2 mnGT Y2 mT
mutNHEJ Y2 mnmutNHEJ Y2 mT
E148D megaTAL WT megaTAL F13Y megaTAL Y2 megaTAL
WT mn (90nM) F13Y mn (50nM) Y2 mn (10nM)
scr 2 3 4 5 6 7 8 9 10 12 16
Spacer length
Fol
dov
erY
2 m
n (m
utN
HE
J)
0
1
2
3
4
5
0 102 103 104 105
0
102
103
104
105
00274
0173
mC
herr
y
GFP
BFP
coun
t
744
0 102 103 104 105
0
102
103
104
105
109
799
BFP
coun
t
694
mC
herr
y
GFP0 102 103 104 105
0
102
103
104
105
00376
00188
BFP
coun
t
747
0 102 103 104 105
0
102
103
104
105
00299
00299
BFP
coun
t
683
(a)
(b)
(c) (d)
Figure 3 megaTAL addressing acts by increasing the effective mn concentration at the addressed target site (a) Level of cleavage activity (assayedby mutNHEJ) of megaTALs built with mns with varying affinities in TLR cell lines containing targets with different DNA spacer lengths The levelof mutNHEJ for each megaTAL tested was normalized to the activity of Y2 (represented by green line) in the same TLR cell line The averageactivity of the WT and F13Y mns across each cell line are represented by blue and orange lines respectively (b) Effect of megaTAL addressing onactivity of Y2 I-AniI toward DNA targets for which it exhibits different biochemical properties Left panel Wild-type and singly substituted I-AniItargets and previously determined kcat and KM values Right panel Mutagenic NHEJ and gene targeting of Y2 and Y2 megaTAL at each targetassayed by TLR Representative flow plots for data from (b) are shown for the Y2mn (left column) and megaTAL (right column) against high KM(c) and low kcat (d) targets Numbers inside the gates give the percent of mCherry and GFP positive cells and the inset graphs show BFP positivecells on which the experiment was gated
6 Nucleic Acids Research 2013
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
for the low kcat targets (0ndash3-fold for GT and mutNHEJ)(Figure 3b and d) However megaTAL activity againstthe low kcat target -1G was found to be significantlyincreased compared with mn treatment suggesting thatin some cases lsquoaddressingrsquo may partially compensate forreduced catalytic activity
Increasing on-target cleavage with megaTALs
One of the major shortcomings of current nuclease plat-forms is their potential for off-target cleavage and thenegative effects that this may have on the efficacy orsafety profile of a putative nuclease-based therapeutic ap-plication To evaluate the specificity properties of themegaTAL architecture we tested whether a megaTALcould consistently distinguish between lsquoaddressedrsquo andlsquononaddressedrsquo genomic target sites all cleaved by acommon mn cleavage domain Near-native sites of theI-AniI target (DNA sequences containing three or fewerbase-pair differences from the native I-AniI recognitionsequence) were previously identified in the humangenome using the UCSC BLAT tool (40) The cleavageactivity of the Y2 I-AniI variant mn was assessed at thesenear-natives targets using both episomal and endogenoustargets (data not shown) and the five sites that displayedthe highest levels of cleavage by Y2 in those assays werechosen for further examination (Figure 4a) SyntheticTAL effectors specific to regions upstream of the +9Tand+5A+8T near-native targets were built and fused tothe WT F13Y and Y2 I-AniI mns These megaTALs weretested alongside the stand-alone mn variants in TLRreporter cell lines for each of the five targets The +9Tand+5A+8T megaTALs showed increased levels of genetargeting at their addressed targets only while having noobserved effect on the activity at the remaining sites(Figure 4b) Notably while the addition of a sequence-specific TAL effector domain to the high-affinity Y2mndid not reduce its cleavage at nonaddressed targets (theY2mn and megaTALs show similar levels of GT at off-target sites) treatment with megaTALs made with thelower affinity WT and F13Y mns resulted in levels ofgene targeting at addressed targets equivalent to thatachieved with the Y2 megaTALs but without detectableoff-target activity
To verify that a similar effect would be observed atthe endogenous near-native loci 293T cells were treatedwith +9T megaTALs +5A+8T megaTALs and mnvariants and the level of mutNHEJ was determined ateach near-native site by high-throughput sequencing(Figure 4c Supplementary Figure S3a and b) Theseresults mirrored those from the reporter assay with theWT and F13Y megaTALs achieving indel rates up to75 at their addressed sites while maintaining near-background activity at nonaddressed sites
Highly effective knockout of the T-cell receptor alphagene in primary T-cells with a megaTAL
To verify that the benefits observed when fusing wild-typemns to a TAL effector would be achievable with anengineered mn and could be captured in a primary celltype in a relevant therapeutic application we designed a
variant of the I-OnuI mn to knockout the T-cell receptoralpha (TCRa) gene Knockout of the TCRa gene inhuman T-cells has been proposed as an approach toimprove the safety of T-cells endowed with chimericantigen receptors or heterologous TCRs by preventingthem from generating graft-versus-host disease (41)Thus knockout of TCRa in human T-cells is among themost translationally relevant applications of rare cleavingnuclease technologyIn vitro characterization of the TCRa mn showed that
its target site affinity and cleavage activity were similar tothat of its parent enzyme Specificity profiling of theTCRa mn and subsequent high-throughput in vitrocleavage analysis led to the identification of a numberof potential off-target sites which were cleaved with com-parable efficiency to the desired target (SupplementaryFigure S4a and e) To determine if generation of aTCRa megaTAL would yield improvements in activityand specificity the TCRa mn was fused to a 105 repeatTALE array designed to bind a DNA sequence upstreamof the TCRamn binding site (Figure 5a) T-cells were thentransfected with mRNA encoding either the TCRamegaTAL or the stand-alone mn In order to increasethe efficiency of gene knockout samples were includedwherein either the mn or the megaTAL were co-trans-fected with mRNA encoding Trex2 a 30 to 50 exonucleasethat has been shown to significantly increase rates ofmutNHEJ at mn cleavage sites (4243) Disruption ofthe TCRa gene was measured by staining for CD3 amulti-protein complex that is associated with the ab T-cell receptor Strikingly while disruption of the TCRagene occurred at minimal rates in cells treated with thestand-alone mn (16) the TCRa megaTAL exhibited a20-fold increase in activity at the addressed target site inthe TCRa gene (Figure 5b representative flow plots areshown in Figure 5c) Co-transfection of each nuclease withTrex2 further increased the rate of disruption yieldingrates of TCRa gene disruption consistently exceeding70Both on- and off-target cleavage rates were analyzed by
sequencing of the TCRa locus and putative off-target sites(identified based on in vitro cleavage profiling) in FACSsorted CD3- populations of nucleaseTrex2 treated cells(Figure 5d Supplementary Figure S5andashc) For each treat-ment modification rates at the TCRa locus were expectedto be 50 signifying knockout of the gene at theexpressing allele in conjunction with some additionalrate of modification of the nonexpressing allele InmegaTAL treated cells we observed 704 modificationsuggesting a substantial rate (ca 1 in 5 cells) of modifica-tion of the nonexpressed allele In contrast when weexpressed the megaTAL with Trex2 we observed thatonly 520 of the alleles carried indels and even lowerrates were obtained from the mn-alone treated cells Weattribute the differences in modification rates observed inthe presence and absence of Trex2 to lowered expressionof the nuclease in cells co-transfected with the exonucle-ase The lower modification rates observed in the lowabundance CD3- mn alone treated cells are attributableto contamination of the sorted populations with a largerproportion of background unstained cells The
Nucleic Acids Research 2013 7
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
sequencing analysis also demonstrated that off-targetcleavage in cells treated with the megaTAL alone is ex-tremely low with only one of 22 potential sites exhibitinga rate of cleavage above background in the presence ofTrex2 (OT23) Moreover the megaTAL produced lowerlevels of mutNHEJ at several putative off-target loci thanthe stand-alone mn (ie OT18) mdash this result may reflectstatistical variation in the deep sequencing results or al-ternatively could be accounted for by reduced expressionof the megaTAL nuclease compared with its shorter mncounterpart
DISCUSSION
The megaTAL nuclease architecture described hereaddresses several key limitations of existing nuclease
platforms for therapeutic applications as it enables thegeneration of compact single chain nucleases that are ex-tremely active and specific
MegaTALs exhibit dramatic increases in cleavageactivity compared with their isolated mn cleavage headsan effect that appears to be an outcome of the auxiliaryTAL effector DNA-binding domain docking (lsquoaddress-ingrsquo) the mn adjacent to its target and effectivelyincreasing the rate of enzymendashsubstrate complex forma-tion Furthermore the mn-mediated cleavage producesDNA ends with 30 overhangs that are efficiently processedby the Trex2 exonuclease thus allowing the co-expressionof a megaTAL and Trex2 to achieve unprecedentedly hightargeted modification rates in primary cells of gt70
MegaTALs offer an extreme level of target specificitydue to the combined DNA binding and cleavage
Name Genomic sequence Location
+9T TGTCATGTAAATAACTGTACTGTATGTTGAGGAGGTTTCTCTGTATA Ch9 (-)63127729-63127775
+5A+8T TCTTCTGCTGTTTCATATACCTCTATGTGAGGAGGTTTCTCAGTTAG Ch15(-)69553515-69553561
+2T+9G CATAGGTGACTTTTACTCTCATGATAGTGAGGAGGTTTTTCTGTAGA Ch12(+)84384994-84385040
+1C+5A GTCTTTGAATAAGTCACAATGCTTCTTTGAGGAGGTTCCTCAGTAAA Ch7 (+)28151155-28151201
-2A+1G ATGATG GAAATGTTACACTGAGCTCT TGAGGAGGATGCTCTGTAAA Ch1 (+)223534899-223534945
mut
NH
EJ
by
seq
()
+9T +5A+8T +1C+5A -2A+1G0
20
40
60
80
100
+2T+9G locus harbors an extra mutation in 293T cells data not shown
WT +9T WT MegaTAL +5A+8T WT MegaTALF13Y +9T F13Y MegaTAL +5A+8T F13Y MegaTALY2 +9T Y2 MegaTAL +5A+8T Y2 MegaTAL
TLR cell line
+9T +5A+8T +2T+9G +1C+5A -2A+1G0
1
2
3
Per
cent
GF
P+
cells
(a)
(b)
(c)
Figure 4 megaTAL addressing selectively increases on-target cleavage (a) I-AniI human genomic near-native sequences and their location in thegenome Synthetic TAL effectors were built against the+9T and+5A+8T sequences underlined in green and the I-AniI near-natives sequences areunderlined in blue (b) Level of gene targeting in TLR cells with endogenous human I-AniI near-native targets using lsquoaddressedrsquo+9T and+5A+8TmegaTALs and lsquounaddressedrsquo I-AniI mns mutNHEJ was not measured via TLR due to the presence of a stop codon in some target sites thatprevented flow cytometry readout of these events (c) Mutation rates at endogenous targets using lsquoaddressedrsquo I-AniI megaTALs and lsquounadressedrsquomns The level of mutNHEJ at the endogenous I-AniI near-native targets in 293T cells after 72 h of treatment with mn and megaTAL variants wasdetermined based on MiSeq sequencing results
8 Nucleic Acids Research 2013
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
properties of the protein architecture The activity ofmegaTALs is reliant on recognition of the tandem DNAtargets of the TALE array and the mn respectively ef-fectively extending the length of the target site andreducing the likelihood that more than one target sitewould be present in the human genome Similarly as
low affinity mn cleavage heads are not able to efficientlyaccess target sites to which they are not lsquoaddressedrsquo by theTALE array the potential for cleavage at nonaddressedmn recognition sites can be markedly reduced This is re-flected in the megaTAL used for TCRa gene knockout inprimary human T-cells which achieved 70 knockout
Control (-) TCRα mn TCRα mn + Trex2 TCRα mT + Trex2TCRα MegaTAL
Ch14 (+)23016490-23016529
100
mn megaTAL
0
20
40
60
80
CD
3- c
ells
()
- +- +
BFP-
--Trex2
Nuclease
5rsquo- TAAATCCAGTGACAAGTCTGTCTGCCTATTCACCGATTTT ATTTAGGTCACTGTTCAGACAGACGGATAAGTGGCTAAAA-5rsquo
NΔ154 C+63 TCRα mn
TCRα RVD array
283
0 50K 100K 150K 200K
0102
103
104
105
0866
0 50K 100K 150K 200K
519
0 50K 100K 150K 200K
368
0 50K 100K 150K 200K
756
0 50K 100K 150K 200K
CD
3
FSC
384
704
520
50 1000 0125 025 05 075 1 5 10 25NA indel by seq
Putative off target sitesTCRα
mn
mT
Trex2
+
+
-
-
003 0 0 0 0
006 0
0 05
0 1 2
0 14
0 03 0 0
004
0 02
0 08 0
002 0
0 01
0 11 0
OT 1 2 3 4 5 6 7 8 9 10 11 13 14 15 16 17 18 20 21 22 23 24
(a)
(c)
(d)
(b)
Figure 5 megaTAL targeting TCRa achieves extremely high gene disruption with no detectable off-target cleavage in human primary T-cells(a) Schematic of TCRa megaTAL formed by fusing a synthetic TAL effector built toward the TCRa sequence underlined in green to a designedTCRa mn (target underlined in blue) (b) TCR knockout achieved by parental TCRa mn and TCRa megaTAL in human primary T-cells Humanprimary T-cells were transfected with 10 micrograms of RNA encoding the indicated constructs with or without 10 micrograms of mRNA encodingthe Trex2 exonuclease TCRa gene disruption was assessed by measuring the percentage of cells that transition to being CD3- at 5 days followingtransfection Mutation rates at the endogenous TCRa gene were also assessed by Miseq sequencing of amplicons around the cleavage site andshowed indel rates gt50 suggesting that there was a preference for cleavage and disruption of the expressed TCRa allele particularly when themegaTAL is co-expressed with Trex2 (c) Representative flow cytometry data from transfection of human primary T-cells with TCRa targeting mn ormegaTALs+ Trex2 (d) Heat map showing mutation rates at the endogenous TCRa and putative off-target loci in CD3- primary T-cells treatedwith the TCRa mn or megaTAL+ Trex2 Map values were calculated for each locus by subtracting the background rate of indel formation in thecontrol population from that obtained for each nuclease treatment by Miseq sequencing with the heat map color scale indicated by the key (bottom)Sequencing results across all samples are shown (top) as well as an enlarged view with indel rates at putative off-target (OT) sites in cells treated withthe TCRa megaTAL alone (middle) Values calculated as 0 are given as lsquo0rsquo Sequencing results were not obtained for putative off-targets OT12 andOT19 and are therefore not shown on the map other samples for which sequencing results were not obtained are indicated as lsquoNArsquo
Nucleic Acids Research 2013 9
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
while above background off-target cleavage was limited toa single site (eg Figure 5d and Supplementary Figure 5)As the single off-target cleavage site identified is in anoncoding region of the genome this enzyme is of suffi-cient quality at its present state of development for clinicaluse However a further advantage of the megaTAL archi-tecture is the potential to further lsquotunersquo the mn cleavagehead to specifically avoid off-target cleavage sitesThus we anticipate that iterative refinement ofmegaTALs cleavage heads will allow the eventual identi-fication of megaTALs that couple high efficiency withperfect on target genomic specificityThe megaTAL architecture also possesses several
properties that facilitate efficient delivery for therapeuticapplications MegaTALs can be expressed with a single2 kb ORF smaller than two ZFN halves [22 kb total(11 kb per ZFN half targeting a 9-bp target)] and signifi-cantly shorter than a single TALEN half [gt50 kb total(25 kb per TALEN half targeting a 13-bp target)] orCas9 nuclease (41 kb not including guide RNA) ThusmegaTALs are compatible with efficient vectorization as aDNA mRNA or with any widely used viral vector systemThe capacity to use a TAL effector domain with as few as55 repeats also facilitates codon divergence of the TALeffector repeats to allow packaging into retroviral vectorswhere recombination among TAL effector RVDs duringreverse transcription can be problematic (25) Finally theindependent function of megaTALs due to their singlechain design avoids the possibility of inappropriateheterodimerization that may occur with enzymes basedon dimerization-dependent cleavage domains such asFokI an important advantage for applications requiringmultiplex genetic modifications
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Onlineincluding [44]
ACKNOWLEDGEMENTS
We would like to thank members of the NorthwestGenome Engineering Consortium (NGEC) (httpngec-seattleorg) for their many insightful discussions
FUNDING
National Institutes of Health [RL1CA133832UL1DE019582 R01-HL075453 PL1-HL092557 RL1-HL092553 RL1-HL92554 and U19-AI96111] SeattleChildrenrsquos Center for Immunity and ImmunotherapiesFunding for open access charge NIH [U19-AI96111]
Conflict of interest statement AMS is an employee ofCellectis therapeutics and receives salary and equity com-pensation He is also founder member of the board of dir-ectors and equity holder in Pregenen Inc JJ is an employeeof Pregenen Inc and receives salary and equitycompensation AA is an employee of Pregenen Inc andreceives salary and equity compensation PD is an
employee of Cellectis therapeutics and receives salary andequity compensation AG is an employee of Cellectistherapeutics and receives salary and equity compensation
REFERENCES
1 BakerM (2012) Gene-editing nucleases Nat Methods 9 23ndash262 KimYG ChaJ and ChandrasegaranS (1996) Hybrid
restriction enzymes zinc finger fusions to Fok I cleavage domainProc Natl Acad Sci USA 93 1156ndash1160
3 KlugA (2010) The discovery of zinc fingers and theirapplications in gene regulation and genome manipulation AnnRev Biochem 79 213ndash231
4 ChristianM CermakT DoyleEL SchmidtC ZhangFHummelA BogdanoveAJ and VoytasDF (2010) TargetingDNA double-strand breaks with TAL effector nucleases Genetics186 757ndash761
5 LiT HuangS JiangWZ WrightD SpaldingMHWeeksDP and YangB (2011) TAL nucleases (TALNs) hybridproteins composed of TAL effectors and FokI DNA-cleavagedomain Nucleic Acids Res 39 359ndash372
6 PaquesF and DuchateauP (2007) Meganucleases and DNAdouble-strand break-induced recombination perspectives for genetherapy Curr Gene Ther 7 49ndash66
7 StoddardBL (2011) Homing endonucleases from microbialgenetic invaders to reagents for targeted DNA modificationStructure 19 7ndash15
8 JinekM ChylinskiK FonfaraI HauerM DoudnaJA andCharpentierE (2012) A programmable dual-RNAndashguided DNAendonuclease in adaptive bacterial immunity Science 337816ndash821
9 MaliP YangL EsveltKM AachJ GuellM DiCarloJENorvilleJE and ChurchGM (2013) RNA-guided humangenome engineering via Cas9 Science 339 823ndash826
10 McMahonMA RahdarM and PorteusM (2012) Gene editingnot just for translation anymore Nat Methods 9 28ndash31
11 LieberMR (2010) The mechanism of double-strand DNA breakrepair by the nonhomologous DNA end-joining pathway AnnRev Biochem 79 181ndash211
12 ShrivastavM DE HaroLP and NickoloffJA (2008)Regulation of DNA double-strand break repair pathway choiceCell Res 18 134ndash147
13 Method of the Year 2011 Nat Methods 9 114 MandellJG and BarbasCF (2006) Zinc finger tools custom
DNA-binding domains for transcription factors and nucleasesNucleic Acids Res 34 W516ndashW523
15 MaederML Thibodeau-BegannyS OsiakA WrightDAAnthonyRM EichtingerM JiangT FoleyJE WinfreyRJTownsendJA et al (2008) Rapid lsquoopen-sourcersquo engineering ofcustomized zinc-finger nucleases for highly efficient genemodification Mol Cell 31 294ndash301
16 SanderJD DahlborgEJ GoodwinMJ CadeL ZhangFCifuentesD CurtinSJ BlackburnJS Thibodeau-BegannySQiY et al (2011) Selection-free zinc-finger-nuclease engineeringby context-dependent assembly (CoDA) Nat Methods 8 67ndash69
17 CermakT DoyleEL ChristianM WangL ZhangYSchmidtC BallerJA SomiaNV BogdanoveAJ andVoytasDF (2011) Efficient design and assembly of customTALEN and other TAL effector-based constructs for DNAtargeting Nucleic Acids Res 39 e82ndashe82
18 ZhangF CongL LodatoS KosuriS ChurchGM andArlottaP (2011) Efficient construction of sequence-specific TALeffectors for modulating mammalian transcription NatBiotechnol 29 149ndash153
19 ReyonD TsaiSQ KhayterC FodenJA SanderJD andJoungJK (2012) FLASH assembly of TALENs for high-throughput genome editing Nat Biotechnol 30 460ndash465
20 JarjourJ West-FoyleH CertoMT HubertCG DoyleLGetzMM StoddardBL and ScharenbergAM (2009) High-resolution profiling of homing endonuclease binding and catalyticspecificity using yeast surface display Nucleic Acids Res 376871ndash6880
10 Nucleic Acids Research 2013
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
21 CornuTI Thibodeau-BegannyS GuhlE AlwinSEichtingerM JoungJK JoungJK and CathomenT (2008)DNA-binding specificity is a major determinant of the activityand toxicity of zinc-finger nucleases Mol Ther 16 352ndash358
22 GabrielR LombardoA ArensA MillerJC GenovesePKaeppelC NowrouziA BartholomaeCC WangJFriedmanG et al (2011) An unbiased genome-wide analysis ofzinc-finger nuclease specificity Nat Biotechnol 29 816ndash823
23 PattanayakV RamirezCL JoungJK and LiuDR (2011)Revealing off-target cleavage specificities of zinc-finger nucleasesby in vitro selection Nat Methods 8 765ndash770
24 SolluC ParsK CornuTI Thibodeau-BegannySMaederML JoungJK HeilbronnR and CathomenT (2010)Autonomous zinc-finger nuclease pairs for targeted chromosomaldeletion Nucleic Acids Res 38 8269ndash8276
25 HolkersM MaggioI LiuJ JanssenJM MiselliFMussolinoC RecchiaA CathomenT and GoncalvesMA(2012) Differential integrity of TALE nuclease genes followingadenoviral and lentiviral vector gene transfer into human cellsNucleic Acids Res 41 e63
26 FuY FodenJA KhayterC MaederML ReyonDJoungJK and SanderJD (2013) High-frequency off-targetmutagenesis induced by CRISPR-Cas nucleases in human cellsNat Biotechnol 31 822ndash826
27 CradickTJ FineEJ AnticoCJ and BaoG (2013) CRISPRCas9 systems targeting b-globin and CCR5 genes have substantialoff-target activity Nucleic Acids Res 41 9584ndash9592
28 RanFA HsuPD LinCY GootenbergJS KonermannSTrevinoAE ScottDA InoueA MatobaS ZhangY et al(2013) Double nicking by RNA-guided CRISPR Cas9 forenhanced genome editing specificity Cell 154 1380ndash1389
29 AshworthJ TaylorGK HavranekJJ QuadriSAStoddardBL and BakerD (2010) Computationalreprogramming of homing endonuclease specificity at multipleadjacent base pairs Nucleic Acids Res 38 5601ndash5608
30 TakeuchiR LambertAR MakAN JacobyK DicksonRJGloorGB ScharenbergAM EdgellDR and StoddardBL(2011) Tapping natural reservoirs of homing endonucleases fortargeted gene modification Proc Natl Acad Sci USA 10813077ndash13082
31 SatherBD RyuBY StirlingBV GaribovM KernsHMHumblet-BaronS AstrakhanA and RawlingsDJ (2011)Development of B-lineage predominant lentiviral vectors for usein genetic therapies for b cell disorders Mol Ther 19 515ndash525
32 CertoMT RyuBY AnnisJE GaribovM JarjourJRawlingsDJ and ScharenbergAM (2011) Tracking genomeengineering outcome at individual DNA breakpoints NatMethods 8 671ndash676
33 KuharR GwiazdaKS HumbertO MandtT PangalloJBraultM KhanI MaizelsN RawlingsDJ ScharenbergAMet al (2013) Novel fluorescent genome editing reporters formonitoring DNA repair pathway utilization at endonuclease-induced breaks Nucleic Acids Res (epub ahead of print October10 2013)
34 BochJ ScholzeH SchornackS LandgrafA HahnS KaySLahayeT NickstadtA and BonasU (2009) Breaking the codeof DNA binding specificity of TAL-type III effectors Science326 1509ndash1512
35 MoscouMJ and BogdanoveAJ (2009) A simple cipher governsDNA recognition by TAL effectors Science 326 1501ndash1501
36 GaoH WuX ChaiJ and HanZ (2012) Crystal structure of aTALE protein reveals an extended N-terminal DNA bindingregion Cell Res 22 1716ndash1720
37 MillerJC TanS QiaoG BarlowKA WangJ XiaDFMengX PaschonDE LeungE HinkleySJ et al (2011) ATALE nuclease architecture for efficient genome editing NatBiotechnol 29 143ndash148
38 TakeuchiR CertoM CapraraMG ScharenbergAM andStoddardBL (2009) Optimization of in vivo activity of abifunctional homing endonuclease and maturase reversesevolutionary degradation Nucleic Acids Res 37 877ndash890
39 ThymeSB JarjourJ TakeuchiR HavranekJJ AshworthJScharenbergAM StoddardBL and BakerD (2009)Exploitation of binding energy for catalysis and design Nature461 1300ndash1304
40 KentWJ (2002) BLATmdashthe BLAST-like alignment tool GenomeRes 12 656ndash664
41 TorikaiH ReikA LiuPQ ZhouY ZhangL MaitiSHulsH MillerJC KebriaeiP RabinovitchB et al (2012) Afoundation for universal T-cell based immunotherapy T cellsengineered to express a CD19-specific chimeric-antigen-receptorand eliminate expression of endogenous TCR Blood 1195697ndash5705
42 CertoMT GwiazdaKS KuharR SatherB CuringaGMandtT BraultM LambertAR BaxterSK JacobyK et al(2012) Coupling endonucleases with DNA end-processing enzymesto drive gene disruption Nat Methods 9 973ndash975
43 DelacoteF PerezC GuyotV DuhamelM RochonCOllivierN MacmasterR SilvaGH PaquesF DaboussiFet al (2013) High frequency targeted mutagenesis usingengineered endonucleases and DNA-end processing enzymesPLoS One 8 e53217
44 BaxterS LambertAR KuharR JarjourJ KulshinaNParmeggianiF DanaherP GanoJ BakerD StoddardBLet al (2012) Engineering domain fusion chimeras from I-OnuIfamily LAGLIDADG homing endonucleases Nucleic Acids Res40 7985ndash8000
Nucleic Acids Research 2013 11
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
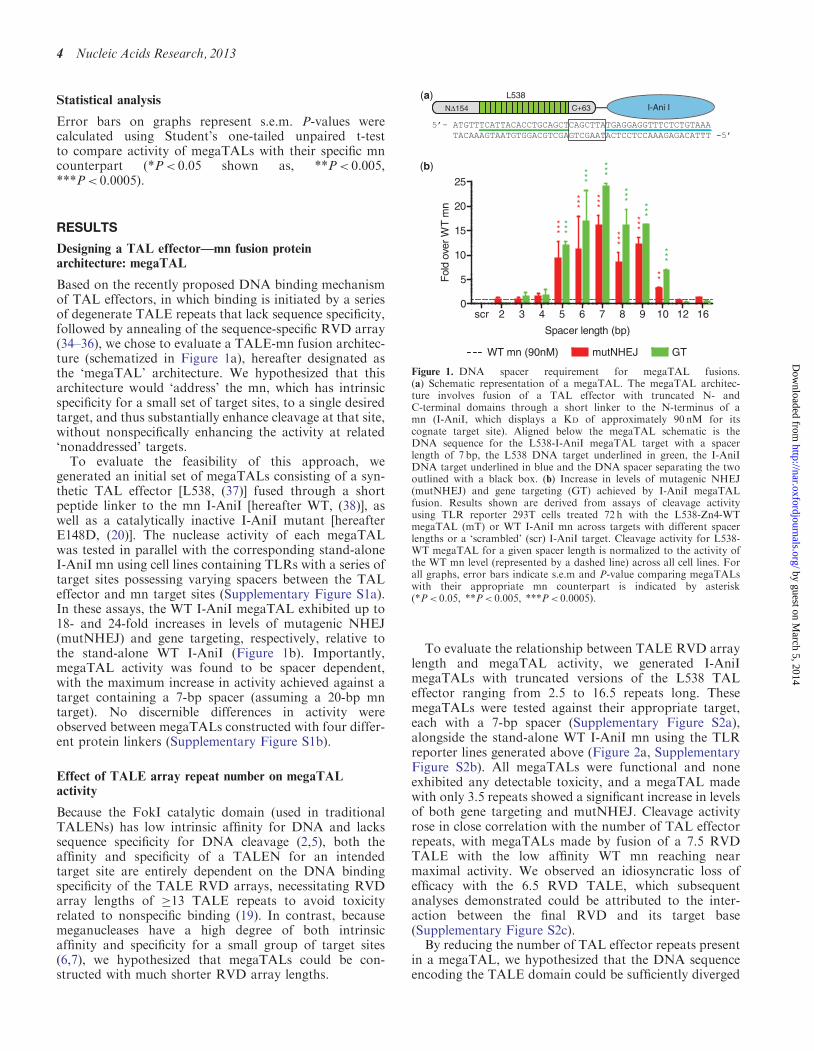
Statistical analysis
Error bars on graphs represent sem P-values werecalculated using Studentrsquos one-tailed unpaired t-testto compare activity of megaTALs with their specific mncounterpart (Plt 005 shown as Plt 0005Plt 00005)
RESULTS
Designing a TAL effectormdashmn fusion proteinarchitecture megaTAL
Based on the recently proposed DNA binding mechanismof TAL effectors in which binding is initiated by a seriesof degenerate TALE repeats that lack sequence specificityfollowed by annealing of the sequence-specific RVD array(34ndash36) we chose to evaluate a TALE-mn fusion architec-ture (schematized in Figure 1a) hereafter designated asthe lsquomegaTALrsquo architecture We hypothesized that thisarchitecture would lsquoaddressrsquo the mn which has intrinsicspecificity for a small set of target sites to a single desiredtarget and thus substantially enhance cleavage at that sitewithout nonspecifically enhancing the activity at relatedlsquononaddressedrsquo targetsTo evaluate the feasibility of this approach we
generated an initial set of megaTALs consisting of a syn-thetic TAL effector [L538 (37)] fused through a shortpeptide linker to the mn I-AniI [hereafter WT (38)] aswell as a catalytically inactive I-AniI mutant [hereafterE148D (20)] The nuclease activity of each megaTALwas tested in parallel with the corresponding stand-aloneI-AniI mn using cell lines containing TLRs with a series oftarget sites possessing varying spacers between the TALeffector and mn target sites (Supplementary Figure S1a)In these assays the WT I-AniI megaTAL exhibited up to18- and 24-fold increases in levels of mutagenic NHEJ(mutNHEJ) and gene targeting respectively relative tothe stand-alone WT I-AniI (Figure 1b) ImportantlymegaTAL activity was found to be spacer dependentwith the maximum increase in activity achieved against atarget containing a 7-bp spacer (assuming a 20-bp mntarget) No discernible differences in activity wereobserved between megaTALs constructed with four differ-ent protein linkers (Supplementary Figure S1b)
Effect of TALE array repeat number on megaTALactivity
Because the FokI catalytic domain (used in traditionalTALENs) has low intrinsic affinity for DNA and lackssequence specificity for DNA cleavage (25) both theaffinity and specificity of a TALEN for an intendedtarget site are entirely dependent on the DNA bindingspecificity of the TALE RVD arrays necessitating RVDarray lengths of 13 TALE repeats to avoid toxicityrelated to nonspecific binding (19) In contrast becausemeganucleases have a high degree of both intrinsicaffinity and specificity for a small group of target sites(67) we hypothesized that megaTALs could be con-structed with much shorter RVD array lengths
To evaluate the relationship between TALE RVD arraylength and megaTAL activity we generated I-AniImegaTALs with truncated versions of the L538 TALeffector ranging from 25 to 165 repeats long ThesemegaTALs were tested against their appropriate targeteach with a 7-bp spacer (Supplementary Figure S2a)alongside the stand-alone WT I-AniI mn using the TLRreporter lines generated above (Figure 2a SupplementaryFigure S2b) All megaTALs were functional and noneexhibited any detectable toxicity and a megaTAL madewith only 35 repeats showed a significant increase in levelsof both gene targeting and mutNHEJ Cleavage activityrose in close correlation with the number of TAL effectorrepeats with megaTALs made by fusion of a 75 RVDTALE with the low affinity WT mn reaching nearmaximal activity We observed an idiosyncratic loss ofefficacy with the 65 RVD TALE which subsequentanalyses demonstrated could be attributed to the inter-action between the final RVD and its target base(Supplementary Figure S2c)
By reducing the number of TAL effector repeats presentin a megaTAL we hypothesized that the DNA sequenceencoding the TALE domain could be sufficiently diverged
(b)
mutNHEJ GTWT mn (90nM)
25
scr 2 3 4 5 6 7 8 9 10 12 160
5
10
15
20
Spacer length (bp)
Fol
dov
erW
Tm
n
(a)
5rsquo- ATGTTTCATTACACCTGCAGCTCAGCTTATGAGGAGGTTTCTCTGTAAA TACAAAGTAATGTGGACGTCGAGTCGAATACTCCTCCAAAGAGACATTT -5rsquo
NΔ154 C+63
L538I-Ani I
Figure 1 DNA spacer requirement for megaTAL fusions(a) Schematic representation of a megaTAL The megaTAL architec-ture involves fusion of a TAL effector with truncated N- andC-terminal domains through a short linker to the N-terminus of amn (I-AniI which displays a KD of approximately 90 nM for itscognate target site) Aligned below the megaTAL schematic is theDNA sequence for the L538-I-AniI megaTAL target with a spacerlength of 7 bp the L538 DNA target underlined in green the I-AniIDNA target underlined in blue and the DNA spacer separating the twooutlined with a black box (b) Increase in levels of mutagenic NHEJ(mutNHEJ) and gene targeting (GT) achieved by I-AniI megaTALfusion Results shown are derived from assays of cleavage activityusing TLR reporter 293T cells treated 72 h with the L538-Zn4-WTmegaTAL (mT) or WT I-AniI mn across targets with different spacerlengths or a lsquoscrambledrsquo (scr) I-AniI target Cleavage activity for L538-WT megaTAL for a given spacer length is normalized to the activity ofthe WT mn level (represented by a dashed line) across all cell lines Forall graphs error bars indicate sem and P-value comparing megaTALswith their appropriate mn counterpart is indicated by asterisk(Plt 005 Plt 0005 Plt 00005)
4 Nucleic Acids Research 2013
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
so as to reduce its repetitiveness and thus allow lentiviralpackaging and delivery of the nuclease transgene withoutrecombination of its open reading frame (ORF) We testedthis by constructing a codon diverged megaTAL with a 65RVD array TAL effector (designated CD for lsquocodondivergedrsquo sequence in Supplementary Figure S2d) andcomparing its cleavage activity to a nondiverged(designated GG for lsquogolden gatersquo) version using bothlentiviral delivery and DNA transfection A 16- and 21-fold difference in mutNHEJ levels was observed inreporter cells treated with an lentiviral or integration-de-ficient lentiviral vector carrying the CD or GG megaTAL
version suggesting a difference in either delivery or ex-pression efficiencies (Figure 2b) Western blot analysis ofthe megaTAL and mn treated cells revealed expression ofthe full-length product in diverged megaTAL treated cellswhereas those that received the nondiverged version ex-pressed protein of different sizes (Supplementary Figure2e) This observation was confirmed using clonal PCRanalysis of the TAL effector RVD array from CD andGG megaTAL treated samples which indicated that 97and 20 of the integrated ORFs from the respective cellpopulations encoded megaTAL proteins with intactTALE arrays (Figure 2c)
Effect of megaTALs on mn target affinity andcleavage activity
The most straightforward explanation for the increasedactivity observed with the I-AniI megaTAL comparedwith the stand-alone mn is that the TALE array effectivelyincreases the local concentration of I-AniI by its adjacentcleavage site We hypothesized that fusing TAL effectorDNA binding domains to mns would thus generally act topotentiate the activity of mns toward targets for whichthey had low affinity but not toward those for whichthey displayed slow cleavage kinetics To test this hypoth-esis we further evaluated the behaviors of (i) a series ofmegaTALs generated from mns with varying affinities forthe I-AniI native target and (ii) a single megaTAL attarget sites for which the mn exhibits differing cleavagekineticsWe generated megaTALs using two additional I-AniI
variants (hereafter F13Y and Y2) with slightly increasedaffinities for the native I-AniI target site (KD=90nMfor I-AniI WT KD=30nM for I-AniI F13YKD=8nM for I-AniI Y2) (38) MegaTALs made withthe moderate (F13Y) and high (Y2) affinity mn variantsalso showed significant increases in activity (Figure 3aSupplementary Figure S1b and c) albeit to a lesserdegree (13-fold increase for F13Y 4-fold for Y2)than observed with the original I-AniI megaTAL Thusthe effect of TALE array addressing correlates with theintrinsic affinity of the mn variant low affinity variantsare boosted to a much greater extent than those with in-trinsically higher affinity consistent with the idea that theTAL effector domain influences nuclease activity byincreasing the local concentration of the mn cleavagedomain at its addressed target site Additionally themoderate affinity mn F13Y reached a plateau of activitywith a shorter (45 RVD) TALE array relative to theTALE array length needed (55ndash75 RVD) to maximizethe activity of the lower affinity WT I-AniI mn(Supplementary Figure S2b)We then further defined the TALE lsquoaddressingrsquo effect by
testing the activity of the high affinity Y2mn and L538-Y2megaTAL against a series of DNA target variants thatharbor single base-pair substitutions (relative to thenative I-AniI target) for which the kcat and KM of theY2 variant have previously been determined (Figure 3b)(39) Nonnative targets were chosen for which Y2 displayseither lsquohigh KMrsquo (Y2 exhibits a similar kcat but reducedaffinity relative to the original target site) or lsquolow kcatrsquo
(a)
(b)
(c)
25 35 45 55 65 75 85 105 1650
10
20
30
40
No of RVD array units
Fol
dov
erW
Tm
n
E148D
WT
mn
CDm
T
GGm
TW
T
CDm
T
GGm
TW
T
CDm
T
GGm
T
0
1
2
3
4
5
Per
cent
mar
ked
cells
(mut
NH
EJ)
pDNA LV IDLV
Treatment Intact TALE ORF (ct) Total countCD megaTAL 28 966 29GG megaTAL 4 211 19
Figure 2 RVD array requirement for megaTAL fusions (a) Level ofcleavage activity (assayed by mutNHEJ shown in red and gene tar-geting shown in green) of megaTALs built with TAL effectors withdifferent number of repeat units Cleavage activity of TALE-Zn4-WTmegaTALs built with a varying number of RVD array units wasnormalized to the activity of the WT I-AniI mn (represented by adashed line) across all cell lines tested (b) Level of cleavage activity(assayed by mutNHEJ) of megaTALs built with either a codondiverged or nondiverged 65 RVD TAL effector array delivered toreporter cells by DNA transfection or lentiviral delivery Both thecodon diverged (CD) and nondiverged (GG) megaTALs were able torescue the activity of the WT mn using plasmid DNA (pDNA) trans-fection lentiviral (LV) or integration-deficient lentiviral (IDLV) trans-duction However delivery using lentiviral vectors resulted insignificantly lower activity in cells treated with the nondivergedversion of the megaTAL (c) Proportion of intact (full length) andrecombined (short) megaTALs integrated into the genome of 293Tcells Either the codon diverged (CD) or nondiverged (GG) 65 RVDWT megaTAL were delivered to cells by lentiviral transduction and theviral integrants were assessed using clonal PCR amplification
Nucleic Acids Research 2013 5
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
(Y2 exhibits a reduced kcat but comparable bindingaffinity compared with the original target site) Cleavageactivity was measured using TLR reporter cells withtargets composed of the L538 binding site and the indi-vidual high KM low kcat or wt I-AniI targets separatedby an 8-bp spacer As expected based on its known
biochemical properties the stand-alone Y2mn showedlow or background levels of activity against all six nonna-tive targets In contrast cleavage activity was considerablyincreased (12ndash38-fold for HR 8ndash27-fold for mutNHEJ) inall high KM targets treated with their corresponding lsquoad-dressedrsquo megaTAL while little or no effect was observed
Data obtained from Thyme et al (2009)
Target site wt base kcat (min-1) KM (nM)
wild-type wt 94 10
-9A G 105 117
-8G A 143 129
-6T G 113 72
-1G T 02 48
+2A C 038 10
+3C T 014 104
Y2 megaTALY2 mn Y2 megaTALY2 mn+2A-8G
wt -9A -8G -6T -1G +2A +3C00
05
10
15
20
25
30
80
Per
cent
mar
ked
cells
high K M low k cat
Target
GT Y2 mnGT Y2 mT
mutNHEJ Y2 mnmutNHEJ Y2 mT
E148D megaTAL WT megaTAL F13Y megaTAL Y2 megaTAL
WT mn (90nM) F13Y mn (50nM) Y2 mn (10nM)
scr 2 3 4 5 6 7 8 9 10 12 16
Spacer length
Fol
dov
erY
2 m
n (m
utN
HE
J)
0
1
2
3
4
5
0 102 103 104 105
0
102
103
104
105
00274
0173
mC
herr
y
GFP
BFP
coun
t
744
0 102 103 104 105
0
102
103
104
105
109
799
BFP
coun
t
694
mC
herr
y
GFP0 102 103 104 105
0
102
103
104
105
00376
00188
BFP
coun
t
747
0 102 103 104 105
0
102
103
104
105
00299
00299
BFP
coun
t
683
(a)
(b)
(c) (d)
Figure 3 megaTAL addressing acts by increasing the effective mn concentration at the addressed target site (a) Level of cleavage activity (assayedby mutNHEJ) of megaTALs built with mns with varying affinities in TLR cell lines containing targets with different DNA spacer lengths The levelof mutNHEJ for each megaTAL tested was normalized to the activity of Y2 (represented by green line) in the same TLR cell line The averageactivity of the WT and F13Y mns across each cell line are represented by blue and orange lines respectively (b) Effect of megaTAL addressing onactivity of Y2 I-AniI toward DNA targets for which it exhibits different biochemical properties Left panel Wild-type and singly substituted I-AniItargets and previously determined kcat and KM values Right panel Mutagenic NHEJ and gene targeting of Y2 and Y2 megaTAL at each targetassayed by TLR Representative flow plots for data from (b) are shown for the Y2mn (left column) and megaTAL (right column) against high KM(c) and low kcat (d) targets Numbers inside the gates give the percent of mCherry and GFP positive cells and the inset graphs show BFP positivecells on which the experiment was gated
6 Nucleic Acids Research 2013
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
for the low kcat targets (0ndash3-fold for GT and mutNHEJ)(Figure 3b and d) However megaTAL activity againstthe low kcat target -1G was found to be significantlyincreased compared with mn treatment suggesting thatin some cases lsquoaddressingrsquo may partially compensate forreduced catalytic activity
Increasing on-target cleavage with megaTALs
One of the major shortcomings of current nuclease plat-forms is their potential for off-target cleavage and thenegative effects that this may have on the efficacy orsafety profile of a putative nuclease-based therapeutic ap-plication To evaluate the specificity properties of themegaTAL architecture we tested whether a megaTALcould consistently distinguish between lsquoaddressedrsquo andlsquononaddressedrsquo genomic target sites all cleaved by acommon mn cleavage domain Near-native sites of theI-AniI target (DNA sequences containing three or fewerbase-pair differences from the native I-AniI recognitionsequence) were previously identified in the humangenome using the UCSC BLAT tool (40) The cleavageactivity of the Y2 I-AniI variant mn was assessed at thesenear-natives targets using both episomal and endogenoustargets (data not shown) and the five sites that displayedthe highest levels of cleavage by Y2 in those assays werechosen for further examination (Figure 4a) SyntheticTAL effectors specific to regions upstream of the +9Tand+5A+8T near-native targets were built and fused tothe WT F13Y and Y2 I-AniI mns These megaTALs weretested alongside the stand-alone mn variants in TLRreporter cell lines for each of the five targets The +9Tand+5A+8T megaTALs showed increased levels of genetargeting at their addressed targets only while having noobserved effect on the activity at the remaining sites(Figure 4b) Notably while the addition of a sequence-specific TAL effector domain to the high-affinity Y2mndid not reduce its cleavage at nonaddressed targets (theY2mn and megaTALs show similar levels of GT at off-target sites) treatment with megaTALs made with thelower affinity WT and F13Y mns resulted in levels ofgene targeting at addressed targets equivalent to thatachieved with the Y2 megaTALs but without detectableoff-target activity
To verify that a similar effect would be observed atthe endogenous near-native loci 293T cells were treatedwith +9T megaTALs +5A+8T megaTALs and mnvariants and the level of mutNHEJ was determined ateach near-native site by high-throughput sequencing(Figure 4c Supplementary Figure S3a and b) Theseresults mirrored those from the reporter assay with theWT and F13Y megaTALs achieving indel rates up to75 at their addressed sites while maintaining near-background activity at nonaddressed sites
Highly effective knockout of the T-cell receptor alphagene in primary T-cells with a megaTAL
To verify that the benefits observed when fusing wild-typemns to a TAL effector would be achievable with anengineered mn and could be captured in a primary celltype in a relevant therapeutic application we designed a
variant of the I-OnuI mn to knockout the T-cell receptoralpha (TCRa) gene Knockout of the TCRa gene inhuman T-cells has been proposed as an approach toimprove the safety of T-cells endowed with chimericantigen receptors or heterologous TCRs by preventingthem from generating graft-versus-host disease (41)Thus knockout of TCRa in human T-cells is among themost translationally relevant applications of rare cleavingnuclease technologyIn vitro characterization of the TCRa mn showed that
its target site affinity and cleavage activity were similar tothat of its parent enzyme Specificity profiling of theTCRa mn and subsequent high-throughput in vitrocleavage analysis led to the identification of a numberof potential off-target sites which were cleaved with com-parable efficiency to the desired target (SupplementaryFigure S4a and e) To determine if generation of aTCRa megaTAL would yield improvements in activityand specificity the TCRa mn was fused to a 105 repeatTALE array designed to bind a DNA sequence upstreamof the TCRamn binding site (Figure 5a) T-cells were thentransfected with mRNA encoding either the TCRamegaTAL or the stand-alone mn In order to increasethe efficiency of gene knockout samples were includedwherein either the mn or the megaTAL were co-trans-fected with mRNA encoding Trex2 a 30 to 50 exonucleasethat has been shown to significantly increase rates ofmutNHEJ at mn cleavage sites (4243) Disruption ofthe TCRa gene was measured by staining for CD3 amulti-protein complex that is associated with the ab T-cell receptor Strikingly while disruption of the TCRagene occurred at minimal rates in cells treated with thestand-alone mn (16) the TCRa megaTAL exhibited a20-fold increase in activity at the addressed target site inthe TCRa gene (Figure 5b representative flow plots areshown in Figure 5c) Co-transfection of each nuclease withTrex2 further increased the rate of disruption yieldingrates of TCRa gene disruption consistently exceeding70Both on- and off-target cleavage rates were analyzed by
sequencing of the TCRa locus and putative off-target sites(identified based on in vitro cleavage profiling) in FACSsorted CD3- populations of nucleaseTrex2 treated cells(Figure 5d Supplementary Figure S5andashc) For each treat-ment modification rates at the TCRa locus were expectedto be 50 signifying knockout of the gene at theexpressing allele in conjunction with some additionalrate of modification of the nonexpressing allele InmegaTAL treated cells we observed 704 modificationsuggesting a substantial rate (ca 1 in 5 cells) of modifica-tion of the nonexpressed allele In contrast when weexpressed the megaTAL with Trex2 we observed thatonly 520 of the alleles carried indels and even lowerrates were obtained from the mn-alone treated cells Weattribute the differences in modification rates observed inthe presence and absence of Trex2 to lowered expressionof the nuclease in cells co-transfected with the exonucle-ase The lower modification rates observed in the lowabundance CD3- mn alone treated cells are attributableto contamination of the sorted populations with a largerproportion of background unstained cells The
Nucleic Acids Research 2013 7
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
sequencing analysis also demonstrated that off-targetcleavage in cells treated with the megaTAL alone is ex-tremely low with only one of 22 potential sites exhibitinga rate of cleavage above background in the presence ofTrex2 (OT23) Moreover the megaTAL produced lowerlevels of mutNHEJ at several putative off-target loci thanthe stand-alone mn (ie OT18) mdash this result may reflectstatistical variation in the deep sequencing results or al-ternatively could be accounted for by reduced expressionof the megaTAL nuclease compared with its shorter mncounterpart
DISCUSSION
The megaTAL nuclease architecture described hereaddresses several key limitations of existing nuclease
platforms for therapeutic applications as it enables thegeneration of compact single chain nucleases that are ex-tremely active and specific
MegaTALs exhibit dramatic increases in cleavageactivity compared with their isolated mn cleavage headsan effect that appears to be an outcome of the auxiliaryTAL effector DNA-binding domain docking (lsquoaddress-ingrsquo) the mn adjacent to its target and effectivelyincreasing the rate of enzymendashsubstrate complex forma-tion Furthermore the mn-mediated cleavage producesDNA ends with 30 overhangs that are efficiently processedby the Trex2 exonuclease thus allowing the co-expressionof a megaTAL and Trex2 to achieve unprecedentedly hightargeted modification rates in primary cells of gt70
MegaTALs offer an extreme level of target specificitydue to the combined DNA binding and cleavage
Name Genomic sequence Location
+9T TGTCATGTAAATAACTGTACTGTATGTTGAGGAGGTTTCTCTGTATA Ch9 (-)63127729-63127775
+5A+8T TCTTCTGCTGTTTCATATACCTCTATGTGAGGAGGTTTCTCAGTTAG Ch15(-)69553515-69553561
+2T+9G CATAGGTGACTTTTACTCTCATGATAGTGAGGAGGTTTTTCTGTAGA Ch12(+)84384994-84385040
+1C+5A GTCTTTGAATAAGTCACAATGCTTCTTTGAGGAGGTTCCTCAGTAAA Ch7 (+)28151155-28151201
-2A+1G ATGATG GAAATGTTACACTGAGCTCT TGAGGAGGATGCTCTGTAAA Ch1 (+)223534899-223534945
mut
NH
EJ
by
seq
()
+9T +5A+8T +1C+5A -2A+1G0
20
40
60
80
100
+2T+9G locus harbors an extra mutation in 293T cells data not shown
WT +9T WT MegaTAL +5A+8T WT MegaTALF13Y +9T F13Y MegaTAL +5A+8T F13Y MegaTALY2 +9T Y2 MegaTAL +5A+8T Y2 MegaTAL
TLR cell line
+9T +5A+8T +2T+9G +1C+5A -2A+1G0
1
2
3
Per
cent
GF
P+
cells
(a)
(b)
(c)
Figure 4 megaTAL addressing selectively increases on-target cleavage (a) I-AniI human genomic near-native sequences and their location in thegenome Synthetic TAL effectors were built against the+9T and+5A+8T sequences underlined in green and the I-AniI near-natives sequences areunderlined in blue (b) Level of gene targeting in TLR cells with endogenous human I-AniI near-native targets using lsquoaddressedrsquo+9T and+5A+8TmegaTALs and lsquounaddressedrsquo I-AniI mns mutNHEJ was not measured via TLR due to the presence of a stop codon in some target sites thatprevented flow cytometry readout of these events (c) Mutation rates at endogenous targets using lsquoaddressedrsquo I-AniI megaTALs and lsquounadressedrsquomns The level of mutNHEJ at the endogenous I-AniI near-native targets in 293T cells after 72 h of treatment with mn and megaTAL variants wasdetermined based on MiSeq sequencing results
8 Nucleic Acids Research 2013
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
properties of the protein architecture The activity ofmegaTALs is reliant on recognition of the tandem DNAtargets of the TALE array and the mn respectively ef-fectively extending the length of the target site andreducing the likelihood that more than one target sitewould be present in the human genome Similarly as
low affinity mn cleavage heads are not able to efficientlyaccess target sites to which they are not lsquoaddressedrsquo by theTALE array the potential for cleavage at nonaddressedmn recognition sites can be markedly reduced This is re-flected in the megaTAL used for TCRa gene knockout inprimary human T-cells which achieved 70 knockout
Control (-) TCRα mn TCRα mn + Trex2 TCRα mT + Trex2TCRα MegaTAL
Ch14 (+)23016490-23016529
100
mn megaTAL
0
20
40
60
80
CD
3- c
ells
()
- +- +
BFP-
--Trex2
Nuclease
5rsquo- TAAATCCAGTGACAAGTCTGTCTGCCTATTCACCGATTTT ATTTAGGTCACTGTTCAGACAGACGGATAAGTGGCTAAAA-5rsquo
NΔ154 C+63 TCRα mn
TCRα RVD array
283
0 50K 100K 150K 200K
0102
103
104
105
0866
0 50K 100K 150K 200K
519
0 50K 100K 150K 200K
368
0 50K 100K 150K 200K
756
0 50K 100K 150K 200K
CD
3
FSC
384
704
520
50 1000 0125 025 05 075 1 5 10 25NA indel by seq
Putative off target sitesTCRα
mn
mT
Trex2
+
+
-
-
003 0 0 0 0
006 0
0 05
0 1 2
0 14
0 03 0 0
004
0 02
0 08 0
002 0
0 01
0 11 0
OT 1 2 3 4 5 6 7 8 9 10 11 13 14 15 16 17 18 20 21 22 23 24
(a)
(c)
(d)
(b)
Figure 5 megaTAL targeting TCRa achieves extremely high gene disruption with no detectable off-target cleavage in human primary T-cells(a) Schematic of TCRa megaTAL formed by fusing a synthetic TAL effector built toward the TCRa sequence underlined in green to a designedTCRa mn (target underlined in blue) (b) TCR knockout achieved by parental TCRa mn and TCRa megaTAL in human primary T-cells Humanprimary T-cells were transfected with 10 micrograms of RNA encoding the indicated constructs with or without 10 micrograms of mRNA encodingthe Trex2 exonuclease TCRa gene disruption was assessed by measuring the percentage of cells that transition to being CD3- at 5 days followingtransfection Mutation rates at the endogenous TCRa gene were also assessed by Miseq sequencing of amplicons around the cleavage site andshowed indel rates gt50 suggesting that there was a preference for cleavage and disruption of the expressed TCRa allele particularly when themegaTAL is co-expressed with Trex2 (c) Representative flow cytometry data from transfection of human primary T-cells with TCRa targeting mn ormegaTALs+ Trex2 (d) Heat map showing mutation rates at the endogenous TCRa and putative off-target loci in CD3- primary T-cells treatedwith the TCRa mn or megaTAL+ Trex2 Map values were calculated for each locus by subtracting the background rate of indel formation in thecontrol population from that obtained for each nuclease treatment by Miseq sequencing with the heat map color scale indicated by the key (bottom)Sequencing results across all samples are shown (top) as well as an enlarged view with indel rates at putative off-target (OT) sites in cells treated withthe TCRa megaTAL alone (middle) Values calculated as 0 are given as lsquo0rsquo Sequencing results were not obtained for putative off-targets OT12 andOT19 and are therefore not shown on the map other samples for which sequencing results were not obtained are indicated as lsquoNArsquo
Nucleic Acids Research 2013 9
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
while above background off-target cleavage was limited toa single site (eg Figure 5d and Supplementary Figure 5)As the single off-target cleavage site identified is in anoncoding region of the genome this enzyme is of suffi-cient quality at its present state of development for clinicaluse However a further advantage of the megaTAL archi-tecture is the potential to further lsquotunersquo the mn cleavagehead to specifically avoid off-target cleavage sitesThus we anticipate that iterative refinement ofmegaTALs cleavage heads will allow the eventual identi-fication of megaTALs that couple high efficiency withperfect on target genomic specificityThe megaTAL architecture also possesses several
properties that facilitate efficient delivery for therapeuticapplications MegaTALs can be expressed with a single2 kb ORF smaller than two ZFN halves [22 kb total(11 kb per ZFN half targeting a 9-bp target)] and signifi-cantly shorter than a single TALEN half [gt50 kb total(25 kb per TALEN half targeting a 13-bp target)] orCas9 nuclease (41 kb not including guide RNA) ThusmegaTALs are compatible with efficient vectorization as aDNA mRNA or with any widely used viral vector systemThe capacity to use a TAL effector domain with as few as55 repeats also facilitates codon divergence of the TALeffector repeats to allow packaging into retroviral vectorswhere recombination among TAL effector RVDs duringreverse transcription can be problematic (25) Finally theindependent function of megaTALs due to their singlechain design avoids the possibility of inappropriateheterodimerization that may occur with enzymes basedon dimerization-dependent cleavage domains such asFokI an important advantage for applications requiringmultiplex genetic modifications
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Onlineincluding [44]
ACKNOWLEDGEMENTS
We would like to thank members of the NorthwestGenome Engineering Consortium (NGEC) (httpngec-seattleorg) for their many insightful discussions
FUNDING
National Institutes of Health [RL1CA133832UL1DE019582 R01-HL075453 PL1-HL092557 RL1-HL092553 RL1-HL92554 and U19-AI96111] SeattleChildrenrsquos Center for Immunity and ImmunotherapiesFunding for open access charge NIH [U19-AI96111]
Conflict of interest statement AMS is an employee ofCellectis therapeutics and receives salary and equity com-pensation He is also founder member of the board of dir-ectors and equity holder in Pregenen Inc JJ is an employeeof Pregenen Inc and receives salary and equitycompensation AA is an employee of Pregenen Inc andreceives salary and equity compensation PD is an
employee of Cellectis therapeutics and receives salary andequity compensation AG is an employee of Cellectistherapeutics and receives salary and equity compensation
REFERENCES
1 BakerM (2012) Gene-editing nucleases Nat Methods 9 23ndash262 KimYG ChaJ and ChandrasegaranS (1996) Hybrid
restriction enzymes zinc finger fusions to Fok I cleavage domainProc Natl Acad Sci USA 93 1156ndash1160
3 KlugA (2010) The discovery of zinc fingers and theirapplications in gene regulation and genome manipulation AnnRev Biochem 79 213ndash231
4 ChristianM CermakT DoyleEL SchmidtC ZhangFHummelA BogdanoveAJ and VoytasDF (2010) TargetingDNA double-strand breaks with TAL effector nucleases Genetics186 757ndash761
5 LiT HuangS JiangWZ WrightD SpaldingMHWeeksDP and YangB (2011) TAL nucleases (TALNs) hybridproteins composed of TAL effectors and FokI DNA-cleavagedomain Nucleic Acids Res 39 359ndash372
6 PaquesF and DuchateauP (2007) Meganucleases and DNAdouble-strand break-induced recombination perspectives for genetherapy Curr Gene Ther 7 49ndash66
7 StoddardBL (2011) Homing endonucleases from microbialgenetic invaders to reagents for targeted DNA modificationStructure 19 7ndash15
8 JinekM ChylinskiK FonfaraI HauerM DoudnaJA andCharpentierE (2012) A programmable dual-RNAndashguided DNAendonuclease in adaptive bacterial immunity Science 337816ndash821
9 MaliP YangL EsveltKM AachJ GuellM DiCarloJENorvilleJE and ChurchGM (2013) RNA-guided humangenome engineering via Cas9 Science 339 823ndash826
10 McMahonMA RahdarM and PorteusM (2012) Gene editingnot just for translation anymore Nat Methods 9 28ndash31
11 LieberMR (2010) The mechanism of double-strand DNA breakrepair by the nonhomologous DNA end-joining pathway AnnRev Biochem 79 181ndash211
12 ShrivastavM DE HaroLP and NickoloffJA (2008)Regulation of DNA double-strand break repair pathway choiceCell Res 18 134ndash147
13 Method of the Year 2011 Nat Methods 9 114 MandellJG and BarbasCF (2006) Zinc finger tools custom
DNA-binding domains for transcription factors and nucleasesNucleic Acids Res 34 W516ndashW523
15 MaederML Thibodeau-BegannyS OsiakA WrightDAAnthonyRM EichtingerM JiangT FoleyJE WinfreyRJTownsendJA et al (2008) Rapid lsquoopen-sourcersquo engineering ofcustomized zinc-finger nucleases for highly efficient genemodification Mol Cell 31 294ndash301
16 SanderJD DahlborgEJ GoodwinMJ CadeL ZhangFCifuentesD CurtinSJ BlackburnJS Thibodeau-BegannySQiY et al (2011) Selection-free zinc-finger-nuclease engineeringby context-dependent assembly (CoDA) Nat Methods 8 67ndash69
17 CermakT DoyleEL ChristianM WangL ZhangYSchmidtC BallerJA SomiaNV BogdanoveAJ andVoytasDF (2011) Efficient design and assembly of customTALEN and other TAL effector-based constructs for DNAtargeting Nucleic Acids Res 39 e82ndashe82
18 ZhangF CongL LodatoS KosuriS ChurchGM andArlottaP (2011) Efficient construction of sequence-specific TALeffectors for modulating mammalian transcription NatBiotechnol 29 149ndash153
19 ReyonD TsaiSQ KhayterC FodenJA SanderJD andJoungJK (2012) FLASH assembly of TALENs for high-throughput genome editing Nat Biotechnol 30 460ndash465
20 JarjourJ West-FoyleH CertoMT HubertCG DoyleLGetzMM StoddardBL and ScharenbergAM (2009) High-resolution profiling of homing endonuclease binding and catalyticspecificity using yeast surface display Nucleic Acids Res 376871ndash6880
10 Nucleic Acids Research 2013
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
21 CornuTI Thibodeau-BegannyS GuhlE AlwinSEichtingerM JoungJK JoungJK and CathomenT (2008)DNA-binding specificity is a major determinant of the activityand toxicity of zinc-finger nucleases Mol Ther 16 352ndash358
22 GabrielR LombardoA ArensA MillerJC GenovesePKaeppelC NowrouziA BartholomaeCC WangJFriedmanG et al (2011) An unbiased genome-wide analysis ofzinc-finger nuclease specificity Nat Biotechnol 29 816ndash823
23 PattanayakV RamirezCL JoungJK and LiuDR (2011)Revealing off-target cleavage specificities of zinc-finger nucleasesby in vitro selection Nat Methods 8 765ndash770
24 SolluC ParsK CornuTI Thibodeau-BegannySMaederML JoungJK HeilbronnR and CathomenT (2010)Autonomous zinc-finger nuclease pairs for targeted chromosomaldeletion Nucleic Acids Res 38 8269ndash8276
25 HolkersM MaggioI LiuJ JanssenJM MiselliFMussolinoC RecchiaA CathomenT and GoncalvesMA(2012) Differential integrity of TALE nuclease genes followingadenoviral and lentiviral vector gene transfer into human cellsNucleic Acids Res 41 e63
26 FuY FodenJA KhayterC MaederML ReyonDJoungJK and SanderJD (2013) High-frequency off-targetmutagenesis induced by CRISPR-Cas nucleases in human cellsNat Biotechnol 31 822ndash826
27 CradickTJ FineEJ AnticoCJ and BaoG (2013) CRISPRCas9 systems targeting b-globin and CCR5 genes have substantialoff-target activity Nucleic Acids Res 41 9584ndash9592
28 RanFA HsuPD LinCY GootenbergJS KonermannSTrevinoAE ScottDA InoueA MatobaS ZhangY et al(2013) Double nicking by RNA-guided CRISPR Cas9 forenhanced genome editing specificity Cell 154 1380ndash1389
29 AshworthJ TaylorGK HavranekJJ QuadriSAStoddardBL and BakerD (2010) Computationalreprogramming of homing endonuclease specificity at multipleadjacent base pairs Nucleic Acids Res 38 5601ndash5608
30 TakeuchiR LambertAR MakAN JacobyK DicksonRJGloorGB ScharenbergAM EdgellDR and StoddardBL(2011) Tapping natural reservoirs of homing endonucleases fortargeted gene modification Proc Natl Acad Sci USA 10813077ndash13082
31 SatherBD RyuBY StirlingBV GaribovM KernsHMHumblet-BaronS AstrakhanA and RawlingsDJ (2011)Development of B-lineage predominant lentiviral vectors for usein genetic therapies for b cell disorders Mol Ther 19 515ndash525
32 CertoMT RyuBY AnnisJE GaribovM JarjourJRawlingsDJ and ScharenbergAM (2011) Tracking genomeengineering outcome at individual DNA breakpoints NatMethods 8 671ndash676
33 KuharR GwiazdaKS HumbertO MandtT PangalloJBraultM KhanI MaizelsN RawlingsDJ ScharenbergAMet al (2013) Novel fluorescent genome editing reporters formonitoring DNA repair pathway utilization at endonuclease-induced breaks Nucleic Acids Res (epub ahead of print October10 2013)
34 BochJ ScholzeH SchornackS LandgrafA HahnS KaySLahayeT NickstadtA and BonasU (2009) Breaking the codeof DNA binding specificity of TAL-type III effectors Science326 1509ndash1512
35 MoscouMJ and BogdanoveAJ (2009) A simple cipher governsDNA recognition by TAL effectors Science 326 1501ndash1501
36 GaoH WuX ChaiJ and HanZ (2012) Crystal structure of aTALE protein reveals an extended N-terminal DNA bindingregion Cell Res 22 1716ndash1720
37 MillerJC TanS QiaoG BarlowKA WangJ XiaDFMengX PaschonDE LeungE HinkleySJ et al (2011) ATALE nuclease architecture for efficient genome editing NatBiotechnol 29 143ndash148
38 TakeuchiR CertoM CapraraMG ScharenbergAM andStoddardBL (2009) Optimization of in vivo activity of abifunctional homing endonuclease and maturase reversesevolutionary degradation Nucleic Acids Res 37 877ndash890
39 ThymeSB JarjourJ TakeuchiR HavranekJJ AshworthJScharenbergAM StoddardBL and BakerD (2009)Exploitation of binding energy for catalysis and design Nature461 1300ndash1304
40 KentWJ (2002) BLATmdashthe BLAST-like alignment tool GenomeRes 12 656ndash664
41 TorikaiH ReikA LiuPQ ZhouY ZhangL MaitiSHulsH MillerJC KebriaeiP RabinovitchB et al (2012) Afoundation for universal T-cell based immunotherapy T cellsengineered to express a CD19-specific chimeric-antigen-receptorand eliminate expression of endogenous TCR Blood 1195697ndash5705
42 CertoMT GwiazdaKS KuharR SatherB CuringaGMandtT BraultM LambertAR BaxterSK JacobyK et al(2012) Coupling endonucleases with DNA end-processing enzymesto drive gene disruption Nat Methods 9 973ndash975
43 DelacoteF PerezC GuyotV DuhamelM RochonCOllivierN MacmasterR SilvaGH PaquesF DaboussiFet al (2013) High frequency targeted mutagenesis usingengineered endonucleases and DNA-end processing enzymesPLoS One 8 e53217
44 BaxterS LambertAR KuharR JarjourJ KulshinaNParmeggianiF DanaherP GanoJ BakerD StoddardBLet al (2012) Engineering domain fusion chimeras from I-OnuIfamily LAGLIDADG homing endonucleases Nucleic Acids Res40 7985ndash8000
Nucleic Acids Research 2013 11
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from

so as to reduce its repetitiveness and thus allow lentiviralpackaging and delivery of the nuclease transgene withoutrecombination of its open reading frame (ORF) We testedthis by constructing a codon diverged megaTAL with a 65RVD array TAL effector (designated CD for lsquocodondivergedrsquo sequence in Supplementary Figure S2d) andcomparing its cleavage activity to a nondiverged(designated GG for lsquogolden gatersquo) version using bothlentiviral delivery and DNA transfection A 16- and 21-fold difference in mutNHEJ levels was observed inreporter cells treated with an lentiviral or integration-de-ficient lentiviral vector carrying the CD or GG megaTAL
version suggesting a difference in either delivery or ex-pression efficiencies (Figure 2b) Western blot analysis ofthe megaTAL and mn treated cells revealed expression ofthe full-length product in diverged megaTAL treated cellswhereas those that received the nondiverged version ex-pressed protein of different sizes (Supplementary Figure2e) This observation was confirmed using clonal PCRanalysis of the TAL effector RVD array from CD andGG megaTAL treated samples which indicated that 97and 20 of the integrated ORFs from the respective cellpopulations encoded megaTAL proteins with intactTALE arrays (Figure 2c)
Effect of megaTALs on mn target affinity andcleavage activity
The most straightforward explanation for the increasedactivity observed with the I-AniI megaTAL comparedwith the stand-alone mn is that the TALE array effectivelyincreases the local concentration of I-AniI by its adjacentcleavage site We hypothesized that fusing TAL effectorDNA binding domains to mns would thus generally act topotentiate the activity of mns toward targets for whichthey had low affinity but not toward those for whichthey displayed slow cleavage kinetics To test this hypoth-esis we further evaluated the behaviors of (i) a series ofmegaTALs generated from mns with varying affinities forthe I-AniI native target and (ii) a single megaTAL attarget sites for which the mn exhibits differing cleavagekineticsWe generated megaTALs using two additional I-AniI
variants (hereafter F13Y and Y2) with slightly increasedaffinities for the native I-AniI target site (KD=90nMfor I-AniI WT KD=30nM for I-AniI F13YKD=8nM for I-AniI Y2) (38) MegaTALs made withthe moderate (F13Y) and high (Y2) affinity mn variantsalso showed significant increases in activity (Figure 3aSupplementary Figure S1b and c) albeit to a lesserdegree (13-fold increase for F13Y 4-fold for Y2)than observed with the original I-AniI megaTAL Thusthe effect of TALE array addressing correlates with theintrinsic affinity of the mn variant low affinity variantsare boosted to a much greater extent than those with in-trinsically higher affinity consistent with the idea that theTAL effector domain influences nuclease activity byincreasing the local concentration of the mn cleavagedomain at its addressed target site Additionally themoderate affinity mn F13Y reached a plateau of activitywith a shorter (45 RVD) TALE array relative to theTALE array length needed (55ndash75 RVD) to maximizethe activity of the lower affinity WT I-AniI mn(Supplementary Figure S2b)We then further defined the TALE lsquoaddressingrsquo effect by
testing the activity of the high affinity Y2mn and L538-Y2megaTAL against a series of DNA target variants thatharbor single base-pair substitutions (relative to thenative I-AniI target) for which the kcat and KM of theY2 variant have previously been determined (Figure 3b)(39) Nonnative targets were chosen for which Y2 displayseither lsquohigh KMrsquo (Y2 exhibits a similar kcat but reducedaffinity relative to the original target site) or lsquolow kcatrsquo
(a)
(b)
(c)
25 35 45 55 65 75 85 105 1650
10
20
30
40
No of RVD array units
Fol
dov
erW
Tm
n
E148D
WT
mn
CDm
T
GGm
TW
T
CDm
T
GGm
TW
T
CDm
T
GGm
T
0
1
2
3
4
5
Per
cent
mar
ked
cells
(mut
NH
EJ)
pDNA LV IDLV
Treatment Intact TALE ORF (ct) Total countCD megaTAL 28 966 29GG megaTAL 4 211 19
Figure 2 RVD array requirement for megaTAL fusions (a) Level ofcleavage activity (assayed by mutNHEJ shown in red and gene tar-geting shown in green) of megaTALs built with TAL effectors withdifferent number of repeat units Cleavage activity of TALE-Zn4-WTmegaTALs built with a varying number of RVD array units wasnormalized to the activity of the WT I-AniI mn (represented by adashed line) across all cell lines tested (b) Level of cleavage activity(assayed by mutNHEJ) of megaTALs built with either a codondiverged or nondiverged 65 RVD TAL effector array delivered toreporter cells by DNA transfection or lentiviral delivery Both thecodon diverged (CD) and nondiverged (GG) megaTALs were able torescue the activity of the WT mn using plasmid DNA (pDNA) trans-fection lentiviral (LV) or integration-deficient lentiviral (IDLV) trans-duction However delivery using lentiviral vectors resulted insignificantly lower activity in cells treated with the nondivergedversion of the megaTAL (c) Proportion of intact (full length) andrecombined (short) megaTALs integrated into the genome of 293Tcells Either the codon diverged (CD) or nondiverged (GG) 65 RVDWT megaTAL were delivered to cells by lentiviral transduction and theviral integrants were assessed using clonal PCR amplification
Nucleic Acids Research 2013 5
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
(Y2 exhibits a reduced kcat but comparable bindingaffinity compared with the original target site) Cleavageactivity was measured using TLR reporter cells withtargets composed of the L538 binding site and the indi-vidual high KM low kcat or wt I-AniI targets separatedby an 8-bp spacer As expected based on its known
biochemical properties the stand-alone Y2mn showedlow or background levels of activity against all six nonna-tive targets In contrast cleavage activity was considerablyincreased (12ndash38-fold for HR 8ndash27-fold for mutNHEJ) inall high KM targets treated with their corresponding lsquoad-dressedrsquo megaTAL while little or no effect was observed
Data obtained from Thyme et al (2009)
Target site wt base kcat (min-1) KM (nM)
wild-type wt 94 10
-9A G 105 117
-8G A 143 129
-6T G 113 72
-1G T 02 48
+2A C 038 10
+3C T 014 104
Y2 megaTALY2 mn Y2 megaTALY2 mn+2A-8G
wt -9A -8G -6T -1G +2A +3C00
05
10
15
20
25
30
80
Per
cent
mar
ked
cells
high K M low k cat
Target
GT Y2 mnGT Y2 mT
mutNHEJ Y2 mnmutNHEJ Y2 mT
E148D megaTAL WT megaTAL F13Y megaTAL Y2 megaTAL
WT mn (90nM) F13Y mn (50nM) Y2 mn (10nM)
scr 2 3 4 5 6 7 8 9 10 12 16
Spacer length
Fol
dov
erY
2 m
n (m
utN
HE
J)
0
1
2
3
4
5
0 102 103 104 105
0
102
103
104
105
00274
0173
mC
herr
y
GFP
BFP
coun
t
744
0 102 103 104 105
0
102
103
104
105
109
799
BFP
coun
t
694
mC
herr
y
GFP0 102 103 104 105
0
102
103
104
105
00376
00188
BFP
coun
t
747
0 102 103 104 105
0
102
103
104
105
00299
00299
BFP
coun
t
683
(a)
(b)
(c) (d)
Figure 3 megaTAL addressing acts by increasing the effective mn concentration at the addressed target site (a) Level of cleavage activity (assayedby mutNHEJ) of megaTALs built with mns with varying affinities in TLR cell lines containing targets with different DNA spacer lengths The levelof mutNHEJ for each megaTAL tested was normalized to the activity of Y2 (represented by green line) in the same TLR cell line The averageactivity of the WT and F13Y mns across each cell line are represented by blue and orange lines respectively (b) Effect of megaTAL addressing onactivity of Y2 I-AniI toward DNA targets for which it exhibits different biochemical properties Left panel Wild-type and singly substituted I-AniItargets and previously determined kcat and KM values Right panel Mutagenic NHEJ and gene targeting of Y2 and Y2 megaTAL at each targetassayed by TLR Representative flow plots for data from (b) are shown for the Y2mn (left column) and megaTAL (right column) against high KM(c) and low kcat (d) targets Numbers inside the gates give the percent of mCherry and GFP positive cells and the inset graphs show BFP positivecells on which the experiment was gated
6 Nucleic Acids Research 2013
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
for the low kcat targets (0ndash3-fold for GT and mutNHEJ)(Figure 3b and d) However megaTAL activity againstthe low kcat target -1G was found to be significantlyincreased compared with mn treatment suggesting thatin some cases lsquoaddressingrsquo may partially compensate forreduced catalytic activity
Increasing on-target cleavage with megaTALs
One of the major shortcomings of current nuclease plat-forms is their potential for off-target cleavage and thenegative effects that this may have on the efficacy orsafety profile of a putative nuclease-based therapeutic ap-plication To evaluate the specificity properties of themegaTAL architecture we tested whether a megaTALcould consistently distinguish between lsquoaddressedrsquo andlsquononaddressedrsquo genomic target sites all cleaved by acommon mn cleavage domain Near-native sites of theI-AniI target (DNA sequences containing three or fewerbase-pair differences from the native I-AniI recognitionsequence) were previously identified in the humangenome using the UCSC BLAT tool (40) The cleavageactivity of the Y2 I-AniI variant mn was assessed at thesenear-natives targets using both episomal and endogenoustargets (data not shown) and the five sites that displayedthe highest levels of cleavage by Y2 in those assays werechosen for further examination (Figure 4a) SyntheticTAL effectors specific to regions upstream of the +9Tand+5A+8T near-native targets were built and fused tothe WT F13Y and Y2 I-AniI mns These megaTALs weretested alongside the stand-alone mn variants in TLRreporter cell lines for each of the five targets The +9Tand+5A+8T megaTALs showed increased levels of genetargeting at their addressed targets only while having noobserved effect on the activity at the remaining sites(Figure 4b) Notably while the addition of a sequence-specific TAL effector domain to the high-affinity Y2mndid not reduce its cleavage at nonaddressed targets (theY2mn and megaTALs show similar levels of GT at off-target sites) treatment with megaTALs made with thelower affinity WT and F13Y mns resulted in levels ofgene targeting at addressed targets equivalent to thatachieved with the Y2 megaTALs but without detectableoff-target activity
To verify that a similar effect would be observed atthe endogenous near-native loci 293T cells were treatedwith +9T megaTALs +5A+8T megaTALs and mnvariants and the level of mutNHEJ was determined ateach near-native site by high-throughput sequencing(Figure 4c Supplementary Figure S3a and b) Theseresults mirrored those from the reporter assay with theWT and F13Y megaTALs achieving indel rates up to75 at their addressed sites while maintaining near-background activity at nonaddressed sites
Highly effective knockout of the T-cell receptor alphagene in primary T-cells with a megaTAL
To verify that the benefits observed when fusing wild-typemns to a TAL effector would be achievable with anengineered mn and could be captured in a primary celltype in a relevant therapeutic application we designed a
variant of the I-OnuI mn to knockout the T-cell receptoralpha (TCRa) gene Knockout of the TCRa gene inhuman T-cells has been proposed as an approach toimprove the safety of T-cells endowed with chimericantigen receptors or heterologous TCRs by preventingthem from generating graft-versus-host disease (41)Thus knockout of TCRa in human T-cells is among themost translationally relevant applications of rare cleavingnuclease technologyIn vitro characterization of the TCRa mn showed that
its target site affinity and cleavage activity were similar tothat of its parent enzyme Specificity profiling of theTCRa mn and subsequent high-throughput in vitrocleavage analysis led to the identification of a numberof potential off-target sites which were cleaved with com-parable efficiency to the desired target (SupplementaryFigure S4a and e) To determine if generation of aTCRa megaTAL would yield improvements in activityand specificity the TCRa mn was fused to a 105 repeatTALE array designed to bind a DNA sequence upstreamof the TCRamn binding site (Figure 5a) T-cells were thentransfected with mRNA encoding either the TCRamegaTAL or the stand-alone mn In order to increasethe efficiency of gene knockout samples were includedwherein either the mn or the megaTAL were co-trans-fected with mRNA encoding Trex2 a 30 to 50 exonucleasethat has been shown to significantly increase rates ofmutNHEJ at mn cleavage sites (4243) Disruption ofthe TCRa gene was measured by staining for CD3 amulti-protein complex that is associated with the ab T-cell receptor Strikingly while disruption of the TCRagene occurred at minimal rates in cells treated with thestand-alone mn (16) the TCRa megaTAL exhibited a20-fold increase in activity at the addressed target site inthe TCRa gene (Figure 5b representative flow plots areshown in Figure 5c) Co-transfection of each nuclease withTrex2 further increased the rate of disruption yieldingrates of TCRa gene disruption consistently exceeding70Both on- and off-target cleavage rates were analyzed by
sequencing of the TCRa locus and putative off-target sites(identified based on in vitro cleavage profiling) in FACSsorted CD3- populations of nucleaseTrex2 treated cells(Figure 5d Supplementary Figure S5andashc) For each treat-ment modification rates at the TCRa locus were expectedto be 50 signifying knockout of the gene at theexpressing allele in conjunction with some additionalrate of modification of the nonexpressing allele InmegaTAL treated cells we observed 704 modificationsuggesting a substantial rate (ca 1 in 5 cells) of modifica-tion of the nonexpressed allele In contrast when weexpressed the megaTAL with Trex2 we observed thatonly 520 of the alleles carried indels and even lowerrates were obtained from the mn-alone treated cells Weattribute the differences in modification rates observed inthe presence and absence of Trex2 to lowered expressionof the nuclease in cells co-transfected with the exonucle-ase The lower modification rates observed in the lowabundance CD3- mn alone treated cells are attributableto contamination of the sorted populations with a largerproportion of background unstained cells The
Nucleic Acids Research 2013 7
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
sequencing analysis also demonstrated that off-targetcleavage in cells treated with the megaTAL alone is ex-tremely low with only one of 22 potential sites exhibitinga rate of cleavage above background in the presence ofTrex2 (OT23) Moreover the megaTAL produced lowerlevels of mutNHEJ at several putative off-target loci thanthe stand-alone mn (ie OT18) mdash this result may reflectstatistical variation in the deep sequencing results or al-ternatively could be accounted for by reduced expressionof the megaTAL nuclease compared with its shorter mncounterpart
DISCUSSION
The megaTAL nuclease architecture described hereaddresses several key limitations of existing nuclease
platforms for therapeutic applications as it enables thegeneration of compact single chain nucleases that are ex-tremely active and specific
MegaTALs exhibit dramatic increases in cleavageactivity compared with their isolated mn cleavage headsan effect that appears to be an outcome of the auxiliaryTAL effector DNA-binding domain docking (lsquoaddress-ingrsquo) the mn adjacent to its target and effectivelyincreasing the rate of enzymendashsubstrate complex forma-tion Furthermore the mn-mediated cleavage producesDNA ends with 30 overhangs that are efficiently processedby the Trex2 exonuclease thus allowing the co-expressionof a megaTAL and Trex2 to achieve unprecedentedly hightargeted modification rates in primary cells of gt70
MegaTALs offer an extreme level of target specificitydue to the combined DNA binding and cleavage
Name Genomic sequence Location
+9T TGTCATGTAAATAACTGTACTGTATGTTGAGGAGGTTTCTCTGTATA Ch9 (-)63127729-63127775
+5A+8T TCTTCTGCTGTTTCATATACCTCTATGTGAGGAGGTTTCTCAGTTAG Ch15(-)69553515-69553561
+2T+9G CATAGGTGACTTTTACTCTCATGATAGTGAGGAGGTTTTTCTGTAGA Ch12(+)84384994-84385040
+1C+5A GTCTTTGAATAAGTCACAATGCTTCTTTGAGGAGGTTCCTCAGTAAA Ch7 (+)28151155-28151201
-2A+1G ATGATG GAAATGTTACACTGAGCTCT TGAGGAGGATGCTCTGTAAA Ch1 (+)223534899-223534945
mut
NH
EJ
by
seq
()
+9T +5A+8T +1C+5A -2A+1G0
20
40
60
80
100
+2T+9G locus harbors an extra mutation in 293T cells data not shown
WT +9T WT MegaTAL +5A+8T WT MegaTALF13Y +9T F13Y MegaTAL +5A+8T F13Y MegaTALY2 +9T Y2 MegaTAL +5A+8T Y2 MegaTAL
TLR cell line
+9T +5A+8T +2T+9G +1C+5A -2A+1G0
1
2
3
Per
cent
GF
P+
cells
(a)
(b)
(c)
Figure 4 megaTAL addressing selectively increases on-target cleavage (a) I-AniI human genomic near-native sequences and their location in thegenome Synthetic TAL effectors were built against the+9T and+5A+8T sequences underlined in green and the I-AniI near-natives sequences areunderlined in blue (b) Level of gene targeting in TLR cells with endogenous human I-AniI near-native targets using lsquoaddressedrsquo+9T and+5A+8TmegaTALs and lsquounaddressedrsquo I-AniI mns mutNHEJ was not measured via TLR due to the presence of a stop codon in some target sites thatprevented flow cytometry readout of these events (c) Mutation rates at endogenous targets using lsquoaddressedrsquo I-AniI megaTALs and lsquounadressedrsquomns The level of mutNHEJ at the endogenous I-AniI near-native targets in 293T cells after 72 h of treatment with mn and megaTAL variants wasdetermined based on MiSeq sequencing results
8 Nucleic Acids Research 2013
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
properties of the protein architecture The activity ofmegaTALs is reliant on recognition of the tandem DNAtargets of the TALE array and the mn respectively ef-fectively extending the length of the target site andreducing the likelihood that more than one target sitewould be present in the human genome Similarly as
low affinity mn cleavage heads are not able to efficientlyaccess target sites to which they are not lsquoaddressedrsquo by theTALE array the potential for cleavage at nonaddressedmn recognition sites can be markedly reduced This is re-flected in the megaTAL used for TCRa gene knockout inprimary human T-cells which achieved 70 knockout
Control (-) TCRα mn TCRα mn + Trex2 TCRα mT + Trex2TCRα MegaTAL
Ch14 (+)23016490-23016529
100
mn megaTAL
0
20
40
60
80
CD
3- c
ells
()
- +- +
BFP-
--Trex2
Nuclease
5rsquo- TAAATCCAGTGACAAGTCTGTCTGCCTATTCACCGATTTT ATTTAGGTCACTGTTCAGACAGACGGATAAGTGGCTAAAA-5rsquo
NΔ154 C+63 TCRα mn
TCRα RVD array
283
0 50K 100K 150K 200K
0102
103
104
105
0866
0 50K 100K 150K 200K
519
0 50K 100K 150K 200K
368
0 50K 100K 150K 200K
756
0 50K 100K 150K 200K
CD
3
FSC
384
704
520
50 1000 0125 025 05 075 1 5 10 25NA indel by seq
Putative off target sitesTCRα
mn
mT
Trex2
+
+
-
-
003 0 0 0 0
006 0
0 05
0 1 2
0 14
0 03 0 0
004
0 02
0 08 0
002 0
0 01
0 11 0
OT 1 2 3 4 5 6 7 8 9 10 11 13 14 15 16 17 18 20 21 22 23 24
(a)
(c)
(d)
(b)
Figure 5 megaTAL targeting TCRa achieves extremely high gene disruption with no detectable off-target cleavage in human primary T-cells(a) Schematic of TCRa megaTAL formed by fusing a synthetic TAL effector built toward the TCRa sequence underlined in green to a designedTCRa mn (target underlined in blue) (b) TCR knockout achieved by parental TCRa mn and TCRa megaTAL in human primary T-cells Humanprimary T-cells were transfected with 10 micrograms of RNA encoding the indicated constructs with or without 10 micrograms of mRNA encodingthe Trex2 exonuclease TCRa gene disruption was assessed by measuring the percentage of cells that transition to being CD3- at 5 days followingtransfection Mutation rates at the endogenous TCRa gene were also assessed by Miseq sequencing of amplicons around the cleavage site andshowed indel rates gt50 suggesting that there was a preference for cleavage and disruption of the expressed TCRa allele particularly when themegaTAL is co-expressed with Trex2 (c) Representative flow cytometry data from transfection of human primary T-cells with TCRa targeting mn ormegaTALs+ Trex2 (d) Heat map showing mutation rates at the endogenous TCRa and putative off-target loci in CD3- primary T-cells treatedwith the TCRa mn or megaTAL+ Trex2 Map values were calculated for each locus by subtracting the background rate of indel formation in thecontrol population from that obtained for each nuclease treatment by Miseq sequencing with the heat map color scale indicated by the key (bottom)Sequencing results across all samples are shown (top) as well as an enlarged view with indel rates at putative off-target (OT) sites in cells treated withthe TCRa megaTAL alone (middle) Values calculated as 0 are given as lsquo0rsquo Sequencing results were not obtained for putative off-targets OT12 andOT19 and are therefore not shown on the map other samples for which sequencing results were not obtained are indicated as lsquoNArsquo
Nucleic Acids Research 2013 9
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
while above background off-target cleavage was limited toa single site (eg Figure 5d and Supplementary Figure 5)As the single off-target cleavage site identified is in anoncoding region of the genome this enzyme is of suffi-cient quality at its present state of development for clinicaluse However a further advantage of the megaTAL archi-tecture is the potential to further lsquotunersquo the mn cleavagehead to specifically avoid off-target cleavage sitesThus we anticipate that iterative refinement ofmegaTALs cleavage heads will allow the eventual identi-fication of megaTALs that couple high efficiency withperfect on target genomic specificityThe megaTAL architecture also possesses several
properties that facilitate efficient delivery for therapeuticapplications MegaTALs can be expressed with a single2 kb ORF smaller than two ZFN halves [22 kb total(11 kb per ZFN half targeting a 9-bp target)] and signifi-cantly shorter than a single TALEN half [gt50 kb total(25 kb per TALEN half targeting a 13-bp target)] orCas9 nuclease (41 kb not including guide RNA) ThusmegaTALs are compatible with efficient vectorization as aDNA mRNA or with any widely used viral vector systemThe capacity to use a TAL effector domain with as few as55 repeats also facilitates codon divergence of the TALeffector repeats to allow packaging into retroviral vectorswhere recombination among TAL effector RVDs duringreverse transcription can be problematic (25) Finally theindependent function of megaTALs due to their singlechain design avoids the possibility of inappropriateheterodimerization that may occur with enzymes basedon dimerization-dependent cleavage domains such asFokI an important advantage for applications requiringmultiplex genetic modifications
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Onlineincluding [44]
ACKNOWLEDGEMENTS
We would like to thank members of the NorthwestGenome Engineering Consortium (NGEC) (httpngec-seattleorg) for their many insightful discussions
FUNDING
National Institutes of Health [RL1CA133832UL1DE019582 R01-HL075453 PL1-HL092557 RL1-HL092553 RL1-HL92554 and U19-AI96111] SeattleChildrenrsquos Center for Immunity and ImmunotherapiesFunding for open access charge NIH [U19-AI96111]
Conflict of interest statement AMS is an employee ofCellectis therapeutics and receives salary and equity com-pensation He is also founder member of the board of dir-ectors and equity holder in Pregenen Inc JJ is an employeeof Pregenen Inc and receives salary and equitycompensation AA is an employee of Pregenen Inc andreceives salary and equity compensation PD is an
employee of Cellectis therapeutics and receives salary andequity compensation AG is an employee of Cellectistherapeutics and receives salary and equity compensation
REFERENCES
1 BakerM (2012) Gene-editing nucleases Nat Methods 9 23ndash262 KimYG ChaJ and ChandrasegaranS (1996) Hybrid
restriction enzymes zinc finger fusions to Fok I cleavage domainProc Natl Acad Sci USA 93 1156ndash1160
3 KlugA (2010) The discovery of zinc fingers and theirapplications in gene regulation and genome manipulation AnnRev Biochem 79 213ndash231
4 ChristianM CermakT DoyleEL SchmidtC ZhangFHummelA BogdanoveAJ and VoytasDF (2010) TargetingDNA double-strand breaks with TAL effector nucleases Genetics186 757ndash761
5 LiT HuangS JiangWZ WrightD SpaldingMHWeeksDP and YangB (2011) TAL nucleases (TALNs) hybridproteins composed of TAL effectors and FokI DNA-cleavagedomain Nucleic Acids Res 39 359ndash372
6 PaquesF and DuchateauP (2007) Meganucleases and DNAdouble-strand break-induced recombination perspectives for genetherapy Curr Gene Ther 7 49ndash66
7 StoddardBL (2011) Homing endonucleases from microbialgenetic invaders to reagents for targeted DNA modificationStructure 19 7ndash15
8 JinekM ChylinskiK FonfaraI HauerM DoudnaJA andCharpentierE (2012) A programmable dual-RNAndashguided DNAendonuclease in adaptive bacterial immunity Science 337816ndash821
9 MaliP YangL EsveltKM AachJ GuellM DiCarloJENorvilleJE and ChurchGM (2013) RNA-guided humangenome engineering via Cas9 Science 339 823ndash826
10 McMahonMA RahdarM and PorteusM (2012) Gene editingnot just for translation anymore Nat Methods 9 28ndash31
11 LieberMR (2010) The mechanism of double-strand DNA breakrepair by the nonhomologous DNA end-joining pathway AnnRev Biochem 79 181ndash211
12 ShrivastavM DE HaroLP and NickoloffJA (2008)Regulation of DNA double-strand break repair pathway choiceCell Res 18 134ndash147
13 Method of the Year 2011 Nat Methods 9 114 MandellJG and BarbasCF (2006) Zinc finger tools custom
DNA-binding domains for transcription factors and nucleasesNucleic Acids Res 34 W516ndashW523
15 MaederML Thibodeau-BegannyS OsiakA WrightDAAnthonyRM EichtingerM JiangT FoleyJE WinfreyRJTownsendJA et al (2008) Rapid lsquoopen-sourcersquo engineering ofcustomized zinc-finger nucleases for highly efficient genemodification Mol Cell 31 294ndash301
16 SanderJD DahlborgEJ GoodwinMJ CadeL ZhangFCifuentesD CurtinSJ BlackburnJS Thibodeau-BegannySQiY et al (2011) Selection-free zinc-finger-nuclease engineeringby context-dependent assembly (CoDA) Nat Methods 8 67ndash69
17 CermakT DoyleEL ChristianM WangL ZhangYSchmidtC BallerJA SomiaNV BogdanoveAJ andVoytasDF (2011) Efficient design and assembly of customTALEN and other TAL effector-based constructs for DNAtargeting Nucleic Acids Res 39 e82ndashe82
18 ZhangF CongL LodatoS KosuriS ChurchGM andArlottaP (2011) Efficient construction of sequence-specific TALeffectors for modulating mammalian transcription NatBiotechnol 29 149ndash153
19 ReyonD TsaiSQ KhayterC FodenJA SanderJD andJoungJK (2012) FLASH assembly of TALENs for high-throughput genome editing Nat Biotechnol 30 460ndash465
20 JarjourJ West-FoyleH CertoMT HubertCG DoyleLGetzMM StoddardBL and ScharenbergAM (2009) High-resolution profiling of homing endonuclease binding and catalyticspecificity using yeast surface display Nucleic Acids Res 376871ndash6880
10 Nucleic Acids Research 2013
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
21 CornuTI Thibodeau-BegannyS GuhlE AlwinSEichtingerM JoungJK JoungJK and CathomenT (2008)DNA-binding specificity is a major determinant of the activityand toxicity of zinc-finger nucleases Mol Ther 16 352ndash358
22 GabrielR LombardoA ArensA MillerJC GenovesePKaeppelC NowrouziA BartholomaeCC WangJFriedmanG et al (2011) An unbiased genome-wide analysis ofzinc-finger nuclease specificity Nat Biotechnol 29 816ndash823
23 PattanayakV RamirezCL JoungJK and LiuDR (2011)Revealing off-target cleavage specificities of zinc-finger nucleasesby in vitro selection Nat Methods 8 765ndash770
24 SolluC ParsK CornuTI Thibodeau-BegannySMaederML JoungJK HeilbronnR and CathomenT (2010)Autonomous zinc-finger nuclease pairs for targeted chromosomaldeletion Nucleic Acids Res 38 8269ndash8276
25 HolkersM MaggioI LiuJ JanssenJM MiselliFMussolinoC RecchiaA CathomenT and GoncalvesMA(2012) Differential integrity of TALE nuclease genes followingadenoviral and lentiviral vector gene transfer into human cellsNucleic Acids Res 41 e63
26 FuY FodenJA KhayterC MaederML ReyonDJoungJK and SanderJD (2013) High-frequency off-targetmutagenesis induced by CRISPR-Cas nucleases in human cellsNat Biotechnol 31 822ndash826
27 CradickTJ FineEJ AnticoCJ and BaoG (2013) CRISPRCas9 systems targeting b-globin and CCR5 genes have substantialoff-target activity Nucleic Acids Res 41 9584ndash9592
28 RanFA HsuPD LinCY GootenbergJS KonermannSTrevinoAE ScottDA InoueA MatobaS ZhangY et al(2013) Double nicking by RNA-guided CRISPR Cas9 forenhanced genome editing specificity Cell 154 1380ndash1389
29 AshworthJ TaylorGK HavranekJJ QuadriSAStoddardBL and BakerD (2010) Computationalreprogramming of homing endonuclease specificity at multipleadjacent base pairs Nucleic Acids Res 38 5601ndash5608
30 TakeuchiR LambertAR MakAN JacobyK DicksonRJGloorGB ScharenbergAM EdgellDR and StoddardBL(2011) Tapping natural reservoirs of homing endonucleases fortargeted gene modification Proc Natl Acad Sci USA 10813077ndash13082
31 SatherBD RyuBY StirlingBV GaribovM KernsHMHumblet-BaronS AstrakhanA and RawlingsDJ (2011)Development of B-lineage predominant lentiviral vectors for usein genetic therapies for b cell disorders Mol Ther 19 515ndash525
32 CertoMT RyuBY AnnisJE GaribovM JarjourJRawlingsDJ and ScharenbergAM (2011) Tracking genomeengineering outcome at individual DNA breakpoints NatMethods 8 671ndash676
33 KuharR GwiazdaKS HumbertO MandtT PangalloJBraultM KhanI MaizelsN RawlingsDJ ScharenbergAMet al (2013) Novel fluorescent genome editing reporters formonitoring DNA repair pathway utilization at endonuclease-induced breaks Nucleic Acids Res (epub ahead of print October10 2013)
34 BochJ ScholzeH SchornackS LandgrafA HahnS KaySLahayeT NickstadtA and BonasU (2009) Breaking the codeof DNA binding specificity of TAL-type III effectors Science326 1509ndash1512
35 MoscouMJ and BogdanoveAJ (2009) A simple cipher governsDNA recognition by TAL effectors Science 326 1501ndash1501
36 GaoH WuX ChaiJ and HanZ (2012) Crystal structure of aTALE protein reveals an extended N-terminal DNA bindingregion Cell Res 22 1716ndash1720
37 MillerJC TanS QiaoG BarlowKA WangJ XiaDFMengX PaschonDE LeungE HinkleySJ et al (2011) ATALE nuclease architecture for efficient genome editing NatBiotechnol 29 143ndash148
38 TakeuchiR CertoM CapraraMG ScharenbergAM andStoddardBL (2009) Optimization of in vivo activity of abifunctional homing endonuclease and maturase reversesevolutionary degradation Nucleic Acids Res 37 877ndash890
39 ThymeSB JarjourJ TakeuchiR HavranekJJ AshworthJScharenbergAM StoddardBL and BakerD (2009)Exploitation of binding energy for catalysis and design Nature461 1300ndash1304
40 KentWJ (2002) BLATmdashthe BLAST-like alignment tool GenomeRes 12 656ndash664
41 TorikaiH ReikA LiuPQ ZhouY ZhangL MaitiSHulsH MillerJC KebriaeiP RabinovitchB et al (2012) Afoundation for universal T-cell based immunotherapy T cellsengineered to express a CD19-specific chimeric-antigen-receptorand eliminate expression of endogenous TCR Blood 1195697ndash5705
42 CertoMT GwiazdaKS KuharR SatherB CuringaGMandtT BraultM LambertAR BaxterSK JacobyK et al(2012) Coupling endonucleases with DNA end-processing enzymesto drive gene disruption Nat Methods 9 973ndash975
43 DelacoteF PerezC GuyotV DuhamelM RochonCOllivierN MacmasterR SilvaGH PaquesF DaboussiFet al (2013) High frequency targeted mutagenesis usingengineered endonucleases and DNA-end processing enzymesPLoS One 8 e53217
44 BaxterS LambertAR KuharR JarjourJ KulshinaNParmeggianiF DanaherP GanoJ BakerD StoddardBLet al (2012) Engineering domain fusion chimeras from I-OnuIfamily LAGLIDADG homing endonucleases Nucleic Acids Res40 7985ndash8000
Nucleic Acids Research 2013 11
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from

(Y2 exhibits a reduced kcat but comparable bindingaffinity compared with the original target site) Cleavageactivity was measured using TLR reporter cells withtargets composed of the L538 binding site and the indi-vidual high KM low kcat or wt I-AniI targets separatedby an 8-bp spacer As expected based on its known
biochemical properties the stand-alone Y2mn showedlow or background levels of activity against all six nonna-tive targets In contrast cleavage activity was considerablyincreased (12ndash38-fold for HR 8ndash27-fold for mutNHEJ) inall high KM targets treated with their corresponding lsquoad-dressedrsquo megaTAL while little or no effect was observed
Data obtained from Thyme et al (2009)
Target site wt base kcat (min-1) KM (nM)
wild-type wt 94 10
-9A G 105 117
-8G A 143 129
-6T G 113 72
-1G T 02 48
+2A C 038 10
+3C T 014 104
Y2 megaTALY2 mn Y2 megaTALY2 mn+2A-8G
wt -9A -8G -6T -1G +2A +3C00
05
10
15
20
25
30
80
Per
cent
mar
ked
cells
high K M low k cat
Target
GT Y2 mnGT Y2 mT
mutNHEJ Y2 mnmutNHEJ Y2 mT
E148D megaTAL WT megaTAL F13Y megaTAL Y2 megaTAL
WT mn (90nM) F13Y mn (50nM) Y2 mn (10nM)
scr 2 3 4 5 6 7 8 9 10 12 16
Spacer length
Fol
dov
erY
2 m
n (m
utN
HE
J)
0
1
2
3
4
5
0 102 103 104 105
0
102
103
104
105
00274
0173
mC
herr
y
GFP
BFP
coun
t
744
0 102 103 104 105
0
102
103
104
105
109
799
BFP
coun
t
694
mC
herr
y
GFP0 102 103 104 105
0
102
103
104
105
00376
00188
BFP
coun
t
747
0 102 103 104 105
0
102
103
104
105
00299
00299
BFP
coun
t
683
(a)
(b)
(c) (d)
Figure 3 megaTAL addressing acts by increasing the effective mn concentration at the addressed target site (a) Level of cleavage activity (assayedby mutNHEJ) of megaTALs built with mns with varying affinities in TLR cell lines containing targets with different DNA spacer lengths The levelof mutNHEJ for each megaTAL tested was normalized to the activity of Y2 (represented by green line) in the same TLR cell line The averageactivity of the WT and F13Y mns across each cell line are represented by blue and orange lines respectively (b) Effect of megaTAL addressing onactivity of Y2 I-AniI toward DNA targets for which it exhibits different biochemical properties Left panel Wild-type and singly substituted I-AniItargets and previously determined kcat and KM values Right panel Mutagenic NHEJ and gene targeting of Y2 and Y2 megaTAL at each targetassayed by TLR Representative flow plots for data from (b) are shown for the Y2mn (left column) and megaTAL (right column) against high KM(c) and low kcat (d) targets Numbers inside the gates give the percent of mCherry and GFP positive cells and the inset graphs show BFP positivecells on which the experiment was gated
6 Nucleic Acids Research 2013
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
for the low kcat targets (0ndash3-fold for GT and mutNHEJ)(Figure 3b and d) However megaTAL activity againstthe low kcat target -1G was found to be significantlyincreased compared with mn treatment suggesting thatin some cases lsquoaddressingrsquo may partially compensate forreduced catalytic activity
Increasing on-target cleavage with megaTALs
One of the major shortcomings of current nuclease plat-forms is their potential for off-target cleavage and thenegative effects that this may have on the efficacy orsafety profile of a putative nuclease-based therapeutic ap-plication To evaluate the specificity properties of themegaTAL architecture we tested whether a megaTALcould consistently distinguish between lsquoaddressedrsquo andlsquononaddressedrsquo genomic target sites all cleaved by acommon mn cleavage domain Near-native sites of theI-AniI target (DNA sequences containing three or fewerbase-pair differences from the native I-AniI recognitionsequence) were previously identified in the humangenome using the UCSC BLAT tool (40) The cleavageactivity of the Y2 I-AniI variant mn was assessed at thesenear-natives targets using both episomal and endogenoustargets (data not shown) and the five sites that displayedthe highest levels of cleavage by Y2 in those assays werechosen for further examination (Figure 4a) SyntheticTAL effectors specific to regions upstream of the +9Tand+5A+8T near-native targets were built and fused tothe WT F13Y and Y2 I-AniI mns These megaTALs weretested alongside the stand-alone mn variants in TLRreporter cell lines for each of the five targets The +9Tand+5A+8T megaTALs showed increased levels of genetargeting at their addressed targets only while having noobserved effect on the activity at the remaining sites(Figure 4b) Notably while the addition of a sequence-specific TAL effector domain to the high-affinity Y2mndid not reduce its cleavage at nonaddressed targets (theY2mn and megaTALs show similar levels of GT at off-target sites) treatment with megaTALs made with thelower affinity WT and F13Y mns resulted in levels ofgene targeting at addressed targets equivalent to thatachieved with the Y2 megaTALs but without detectableoff-target activity
To verify that a similar effect would be observed atthe endogenous near-native loci 293T cells were treatedwith +9T megaTALs +5A+8T megaTALs and mnvariants and the level of mutNHEJ was determined ateach near-native site by high-throughput sequencing(Figure 4c Supplementary Figure S3a and b) Theseresults mirrored those from the reporter assay with theWT and F13Y megaTALs achieving indel rates up to75 at their addressed sites while maintaining near-background activity at nonaddressed sites
Highly effective knockout of the T-cell receptor alphagene in primary T-cells with a megaTAL
To verify that the benefits observed when fusing wild-typemns to a TAL effector would be achievable with anengineered mn and could be captured in a primary celltype in a relevant therapeutic application we designed a
variant of the I-OnuI mn to knockout the T-cell receptoralpha (TCRa) gene Knockout of the TCRa gene inhuman T-cells has been proposed as an approach toimprove the safety of T-cells endowed with chimericantigen receptors or heterologous TCRs by preventingthem from generating graft-versus-host disease (41)Thus knockout of TCRa in human T-cells is among themost translationally relevant applications of rare cleavingnuclease technologyIn vitro characterization of the TCRa mn showed that
its target site affinity and cleavage activity were similar tothat of its parent enzyme Specificity profiling of theTCRa mn and subsequent high-throughput in vitrocleavage analysis led to the identification of a numberof potential off-target sites which were cleaved with com-parable efficiency to the desired target (SupplementaryFigure S4a and e) To determine if generation of aTCRa megaTAL would yield improvements in activityand specificity the TCRa mn was fused to a 105 repeatTALE array designed to bind a DNA sequence upstreamof the TCRamn binding site (Figure 5a) T-cells were thentransfected with mRNA encoding either the TCRamegaTAL or the stand-alone mn In order to increasethe efficiency of gene knockout samples were includedwherein either the mn or the megaTAL were co-trans-fected with mRNA encoding Trex2 a 30 to 50 exonucleasethat has been shown to significantly increase rates ofmutNHEJ at mn cleavage sites (4243) Disruption ofthe TCRa gene was measured by staining for CD3 amulti-protein complex that is associated with the ab T-cell receptor Strikingly while disruption of the TCRagene occurred at minimal rates in cells treated with thestand-alone mn (16) the TCRa megaTAL exhibited a20-fold increase in activity at the addressed target site inthe TCRa gene (Figure 5b representative flow plots areshown in Figure 5c) Co-transfection of each nuclease withTrex2 further increased the rate of disruption yieldingrates of TCRa gene disruption consistently exceeding70Both on- and off-target cleavage rates were analyzed by
sequencing of the TCRa locus and putative off-target sites(identified based on in vitro cleavage profiling) in FACSsorted CD3- populations of nucleaseTrex2 treated cells(Figure 5d Supplementary Figure S5andashc) For each treat-ment modification rates at the TCRa locus were expectedto be 50 signifying knockout of the gene at theexpressing allele in conjunction with some additionalrate of modification of the nonexpressing allele InmegaTAL treated cells we observed 704 modificationsuggesting a substantial rate (ca 1 in 5 cells) of modifica-tion of the nonexpressed allele In contrast when weexpressed the megaTAL with Trex2 we observed thatonly 520 of the alleles carried indels and even lowerrates were obtained from the mn-alone treated cells Weattribute the differences in modification rates observed inthe presence and absence of Trex2 to lowered expressionof the nuclease in cells co-transfected with the exonucle-ase The lower modification rates observed in the lowabundance CD3- mn alone treated cells are attributableto contamination of the sorted populations with a largerproportion of background unstained cells The
Nucleic Acids Research 2013 7
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
sequencing analysis also demonstrated that off-targetcleavage in cells treated with the megaTAL alone is ex-tremely low with only one of 22 potential sites exhibitinga rate of cleavage above background in the presence ofTrex2 (OT23) Moreover the megaTAL produced lowerlevels of mutNHEJ at several putative off-target loci thanthe stand-alone mn (ie OT18) mdash this result may reflectstatistical variation in the deep sequencing results or al-ternatively could be accounted for by reduced expressionof the megaTAL nuclease compared with its shorter mncounterpart
DISCUSSION
The megaTAL nuclease architecture described hereaddresses several key limitations of existing nuclease
platforms for therapeutic applications as it enables thegeneration of compact single chain nucleases that are ex-tremely active and specific
MegaTALs exhibit dramatic increases in cleavageactivity compared with their isolated mn cleavage headsan effect that appears to be an outcome of the auxiliaryTAL effector DNA-binding domain docking (lsquoaddress-ingrsquo) the mn adjacent to its target and effectivelyincreasing the rate of enzymendashsubstrate complex forma-tion Furthermore the mn-mediated cleavage producesDNA ends with 30 overhangs that are efficiently processedby the Trex2 exonuclease thus allowing the co-expressionof a megaTAL and Trex2 to achieve unprecedentedly hightargeted modification rates in primary cells of gt70
MegaTALs offer an extreme level of target specificitydue to the combined DNA binding and cleavage
Name Genomic sequence Location
+9T TGTCATGTAAATAACTGTACTGTATGTTGAGGAGGTTTCTCTGTATA Ch9 (-)63127729-63127775
+5A+8T TCTTCTGCTGTTTCATATACCTCTATGTGAGGAGGTTTCTCAGTTAG Ch15(-)69553515-69553561
+2T+9G CATAGGTGACTTTTACTCTCATGATAGTGAGGAGGTTTTTCTGTAGA Ch12(+)84384994-84385040
+1C+5A GTCTTTGAATAAGTCACAATGCTTCTTTGAGGAGGTTCCTCAGTAAA Ch7 (+)28151155-28151201
-2A+1G ATGATG GAAATGTTACACTGAGCTCT TGAGGAGGATGCTCTGTAAA Ch1 (+)223534899-223534945
mut
NH
EJ
by
seq
()
+9T +5A+8T +1C+5A -2A+1G0
20
40
60
80
100
+2T+9G locus harbors an extra mutation in 293T cells data not shown
WT +9T WT MegaTAL +5A+8T WT MegaTALF13Y +9T F13Y MegaTAL +5A+8T F13Y MegaTALY2 +9T Y2 MegaTAL +5A+8T Y2 MegaTAL
TLR cell line
+9T +5A+8T +2T+9G +1C+5A -2A+1G0
1
2
3
Per
cent
GF
P+
cells
(a)
(b)
(c)
Figure 4 megaTAL addressing selectively increases on-target cleavage (a) I-AniI human genomic near-native sequences and their location in thegenome Synthetic TAL effectors were built against the+9T and+5A+8T sequences underlined in green and the I-AniI near-natives sequences areunderlined in blue (b) Level of gene targeting in TLR cells with endogenous human I-AniI near-native targets using lsquoaddressedrsquo+9T and+5A+8TmegaTALs and lsquounaddressedrsquo I-AniI mns mutNHEJ was not measured via TLR due to the presence of a stop codon in some target sites thatprevented flow cytometry readout of these events (c) Mutation rates at endogenous targets using lsquoaddressedrsquo I-AniI megaTALs and lsquounadressedrsquomns The level of mutNHEJ at the endogenous I-AniI near-native targets in 293T cells after 72 h of treatment with mn and megaTAL variants wasdetermined based on MiSeq sequencing results
8 Nucleic Acids Research 2013
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
properties of the protein architecture The activity ofmegaTALs is reliant on recognition of the tandem DNAtargets of the TALE array and the mn respectively ef-fectively extending the length of the target site andreducing the likelihood that more than one target sitewould be present in the human genome Similarly as
low affinity mn cleavage heads are not able to efficientlyaccess target sites to which they are not lsquoaddressedrsquo by theTALE array the potential for cleavage at nonaddressedmn recognition sites can be markedly reduced This is re-flected in the megaTAL used for TCRa gene knockout inprimary human T-cells which achieved 70 knockout
Control (-) TCRα mn TCRα mn + Trex2 TCRα mT + Trex2TCRα MegaTAL
Ch14 (+)23016490-23016529
100
mn megaTAL
0
20
40
60
80
CD
3- c
ells
()
- +- +
BFP-
--Trex2
Nuclease
5rsquo- TAAATCCAGTGACAAGTCTGTCTGCCTATTCACCGATTTT ATTTAGGTCACTGTTCAGACAGACGGATAAGTGGCTAAAA-5rsquo
NΔ154 C+63 TCRα mn
TCRα RVD array
283
0 50K 100K 150K 200K
0102
103
104
105
0866
0 50K 100K 150K 200K
519
0 50K 100K 150K 200K
368
0 50K 100K 150K 200K
756
0 50K 100K 150K 200K
CD
3
FSC
384
704
520
50 1000 0125 025 05 075 1 5 10 25NA indel by seq
Putative off target sitesTCRα
mn
mT
Trex2
+
+
-
-
003 0 0 0 0
006 0
0 05
0 1 2
0 14
0 03 0 0
004
0 02
0 08 0
002 0
0 01
0 11 0
OT 1 2 3 4 5 6 7 8 9 10 11 13 14 15 16 17 18 20 21 22 23 24
(a)
(c)
(d)
(b)
Figure 5 megaTAL targeting TCRa achieves extremely high gene disruption with no detectable off-target cleavage in human primary T-cells(a) Schematic of TCRa megaTAL formed by fusing a synthetic TAL effector built toward the TCRa sequence underlined in green to a designedTCRa mn (target underlined in blue) (b) TCR knockout achieved by parental TCRa mn and TCRa megaTAL in human primary T-cells Humanprimary T-cells were transfected with 10 micrograms of RNA encoding the indicated constructs with or without 10 micrograms of mRNA encodingthe Trex2 exonuclease TCRa gene disruption was assessed by measuring the percentage of cells that transition to being CD3- at 5 days followingtransfection Mutation rates at the endogenous TCRa gene were also assessed by Miseq sequencing of amplicons around the cleavage site andshowed indel rates gt50 suggesting that there was a preference for cleavage and disruption of the expressed TCRa allele particularly when themegaTAL is co-expressed with Trex2 (c) Representative flow cytometry data from transfection of human primary T-cells with TCRa targeting mn ormegaTALs+ Trex2 (d) Heat map showing mutation rates at the endogenous TCRa and putative off-target loci in CD3- primary T-cells treatedwith the TCRa mn or megaTAL+ Trex2 Map values were calculated for each locus by subtracting the background rate of indel formation in thecontrol population from that obtained for each nuclease treatment by Miseq sequencing with the heat map color scale indicated by the key (bottom)Sequencing results across all samples are shown (top) as well as an enlarged view with indel rates at putative off-target (OT) sites in cells treated withthe TCRa megaTAL alone (middle) Values calculated as 0 are given as lsquo0rsquo Sequencing results were not obtained for putative off-targets OT12 andOT19 and are therefore not shown on the map other samples for which sequencing results were not obtained are indicated as lsquoNArsquo
Nucleic Acids Research 2013 9
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
while above background off-target cleavage was limited toa single site (eg Figure 5d and Supplementary Figure 5)As the single off-target cleavage site identified is in anoncoding region of the genome this enzyme is of suffi-cient quality at its present state of development for clinicaluse However a further advantage of the megaTAL archi-tecture is the potential to further lsquotunersquo the mn cleavagehead to specifically avoid off-target cleavage sitesThus we anticipate that iterative refinement ofmegaTALs cleavage heads will allow the eventual identi-fication of megaTALs that couple high efficiency withperfect on target genomic specificityThe megaTAL architecture also possesses several
properties that facilitate efficient delivery for therapeuticapplications MegaTALs can be expressed with a single2 kb ORF smaller than two ZFN halves [22 kb total(11 kb per ZFN half targeting a 9-bp target)] and signifi-cantly shorter than a single TALEN half [gt50 kb total(25 kb per TALEN half targeting a 13-bp target)] orCas9 nuclease (41 kb not including guide RNA) ThusmegaTALs are compatible with efficient vectorization as aDNA mRNA or with any widely used viral vector systemThe capacity to use a TAL effector domain with as few as55 repeats also facilitates codon divergence of the TALeffector repeats to allow packaging into retroviral vectorswhere recombination among TAL effector RVDs duringreverse transcription can be problematic (25) Finally theindependent function of megaTALs due to their singlechain design avoids the possibility of inappropriateheterodimerization that may occur with enzymes basedon dimerization-dependent cleavage domains such asFokI an important advantage for applications requiringmultiplex genetic modifications
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Onlineincluding [44]
ACKNOWLEDGEMENTS
We would like to thank members of the NorthwestGenome Engineering Consortium (NGEC) (httpngec-seattleorg) for their many insightful discussions
FUNDING
National Institutes of Health [RL1CA133832UL1DE019582 R01-HL075453 PL1-HL092557 RL1-HL092553 RL1-HL92554 and U19-AI96111] SeattleChildrenrsquos Center for Immunity and ImmunotherapiesFunding for open access charge NIH [U19-AI96111]
Conflict of interest statement AMS is an employee ofCellectis therapeutics and receives salary and equity com-pensation He is also founder member of the board of dir-ectors and equity holder in Pregenen Inc JJ is an employeeof Pregenen Inc and receives salary and equitycompensation AA is an employee of Pregenen Inc andreceives salary and equity compensation PD is an
employee of Cellectis therapeutics and receives salary andequity compensation AG is an employee of Cellectistherapeutics and receives salary and equity compensation
REFERENCES
1 BakerM (2012) Gene-editing nucleases Nat Methods 9 23ndash262 KimYG ChaJ and ChandrasegaranS (1996) Hybrid
restriction enzymes zinc finger fusions to Fok I cleavage domainProc Natl Acad Sci USA 93 1156ndash1160
3 KlugA (2010) The discovery of zinc fingers and theirapplications in gene regulation and genome manipulation AnnRev Biochem 79 213ndash231
4 ChristianM CermakT DoyleEL SchmidtC ZhangFHummelA BogdanoveAJ and VoytasDF (2010) TargetingDNA double-strand breaks with TAL effector nucleases Genetics186 757ndash761
5 LiT HuangS JiangWZ WrightD SpaldingMHWeeksDP and YangB (2011) TAL nucleases (TALNs) hybridproteins composed of TAL effectors and FokI DNA-cleavagedomain Nucleic Acids Res 39 359ndash372
6 PaquesF and DuchateauP (2007) Meganucleases and DNAdouble-strand break-induced recombination perspectives for genetherapy Curr Gene Ther 7 49ndash66
7 StoddardBL (2011) Homing endonucleases from microbialgenetic invaders to reagents for targeted DNA modificationStructure 19 7ndash15
8 JinekM ChylinskiK FonfaraI HauerM DoudnaJA andCharpentierE (2012) A programmable dual-RNAndashguided DNAendonuclease in adaptive bacterial immunity Science 337816ndash821
9 MaliP YangL EsveltKM AachJ GuellM DiCarloJENorvilleJE and ChurchGM (2013) RNA-guided humangenome engineering via Cas9 Science 339 823ndash826
10 McMahonMA RahdarM and PorteusM (2012) Gene editingnot just for translation anymore Nat Methods 9 28ndash31
11 LieberMR (2010) The mechanism of double-strand DNA breakrepair by the nonhomologous DNA end-joining pathway AnnRev Biochem 79 181ndash211
12 ShrivastavM DE HaroLP and NickoloffJA (2008)Regulation of DNA double-strand break repair pathway choiceCell Res 18 134ndash147
13 Method of the Year 2011 Nat Methods 9 114 MandellJG and BarbasCF (2006) Zinc finger tools custom
DNA-binding domains for transcription factors and nucleasesNucleic Acids Res 34 W516ndashW523
15 MaederML Thibodeau-BegannyS OsiakA WrightDAAnthonyRM EichtingerM JiangT FoleyJE WinfreyRJTownsendJA et al (2008) Rapid lsquoopen-sourcersquo engineering ofcustomized zinc-finger nucleases for highly efficient genemodification Mol Cell 31 294ndash301
16 SanderJD DahlborgEJ GoodwinMJ CadeL ZhangFCifuentesD CurtinSJ BlackburnJS Thibodeau-BegannySQiY et al (2011) Selection-free zinc-finger-nuclease engineeringby context-dependent assembly (CoDA) Nat Methods 8 67ndash69
17 CermakT DoyleEL ChristianM WangL ZhangYSchmidtC BallerJA SomiaNV BogdanoveAJ andVoytasDF (2011) Efficient design and assembly of customTALEN and other TAL effector-based constructs for DNAtargeting Nucleic Acids Res 39 e82ndashe82
18 ZhangF CongL LodatoS KosuriS ChurchGM andArlottaP (2011) Efficient construction of sequence-specific TALeffectors for modulating mammalian transcription NatBiotechnol 29 149ndash153
19 ReyonD TsaiSQ KhayterC FodenJA SanderJD andJoungJK (2012) FLASH assembly of TALENs for high-throughput genome editing Nat Biotechnol 30 460ndash465
20 JarjourJ West-FoyleH CertoMT HubertCG DoyleLGetzMM StoddardBL and ScharenbergAM (2009) High-resolution profiling of homing endonuclease binding and catalyticspecificity using yeast surface display Nucleic Acids Res 376871ndash6880
10 Nucleic Acids Research 2013
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
21 CornuTI Thibodeau-BegannyS GuhlE AlwinSEichtingerM JoungJK JoungJK and CathomenT (2008)DNA-binding specificity is a major determinant of the activityand toxicity of zinc-finger nucleases Mol Ther 16 352ndash358
22 GabrielR LombardoA ArensA MillerJC GenovesePKaeppelC NowrouziA BartholomaeCC WangJFriedmanG et al (2011) An unbiased genome-wide analysis ofzinc-finger nuclease specificity Nat Biotechnol 29 816ndash823
23 PattanayakV RamirezCL JoungJK and LiuDR (2011)Revealing off-target cleavage specificities of zinc-finger nucleasesby in vitro selection Nat Methods 8 765ndash770
24 SolluC ParsK CornuTI Thibodeau-BegannySMaederML JoungJK HeilbronnR and CathomenT (2010)Autonomous zinc-finger nuclease pairs for targeted chromosomaldeletion Nucleic Acids Res 38 8269ndash8276
25 HolkersM MaggioI LiuJ JanssenJM MiselliFMussolinoC RecchiaA CathomenT and GoncalvesMA(2012) Differential integrity of TALE nuclease genes followingadenoviral and lentiviral vector gene transfer into human cellsNucleic Acids Res 41 e63
26 FuY FodenJA KhayterC MaederML ReyonDJoungJK and SanderJD (2013) High-frequency off-targetmutagenesis induced by CRISPR-Cas nucleases in human cellsNat Biotechnol 31 822ndash826
27 CradickTJ FineEJ AnticoCJ and BaoG (2013) CRISPRCas9 systems targeting b-globin and CCR5 genes have substantialoff-target activity Nucleic Acids Res 41 9584ndash9592
28 RanFA HsuPD LinCY GootenbergJS KonermannSTrevinoAE ScottDA InoueA MatobaS ZhangY et al(2013) Double nicking by RNA-guided CRISPR Cas9 forenhanced genome editing specificity Cell 154 1380ndash1389
29 AshworthJ TaylorGK HavranekJJ QuadriSAStoddardBL and BakerD (2010) Computationalreprogramming of homing endonuclease specificity at multipleadjacent base pairs Nucleic Acids Res 38 5601ndash5608
30 TakeuchiR LambertAR MakAN JacobyK DicksonRJGloorGB ScharenbergAM EdgellDR and StoddardBL(2011) Tapping natural reservoirs of homing endonucleases fortargeted gene modification Proc Natl Acad Sci USA 10813077ndash13082
31 SatherBD RyuBY StirlingBV GaribovM KernsHMHumblet-BaronS AstrakhanA and RawlingsDJ (2011)Development of B-lineage predominant lentiviral vectors for usein genetic therapies for b cell disorders Mol Ther 19 515ndash525
32 CertoMT RyuBY AnnisJE GaribovM JarjourJRawlingsDJ and ScharenbergAM (2011) Tracking genomeengineering outcome at individual DNA breakpoints NatMethods 8 671ndash676
33 KuharR GwiazdaKS HumbertO MandtT PangalloJBraultM KhanI MaizelsN RawlingsDJ ScharenbergAMet al (2013) Novel fluorescent genome editing reporters formonitoring DNA repair pathway utilization at endonuclease-induced breaks Nucleic Acids Res (epub ahead of print October10 2013)
34 BochJ ScholzeH SchornackS LandgrafA HahnS KaySLahayeT NickstadtA and BonasU (2009) Breaking the codeof DNA binding specificity of TAL-type III effectors Science326 1509ndash1512
35 MoscouMJ and BogdanoveAJ (2009) A simple cipher governsDNA recognition by TAL effectors Science 326 1501ndash1501
36 GaoH WuX ChaiJ and HanZ (2012) Crystal structure of aTALE protein reveals an extended N-terminal DNA bindingregion Cell Res 22 1716ndash1720
37 MillerJC TanS QiaoG BarlowKA WangJ XiaDFMengX PaschonDE LeungE HinkleySJ et al (2011) ATALE nuclease architecture for efficient genome editing NatBiotechnol 29 143ndash148
38 TakeuchiR CertoM CapraraMG ScharenbergAM andStoddardBL (2009) Optimization of in vivo activity of abifunctional homing endonuclease and maturase reversesevolutionary degradation Nucleic Acids Res 37 877ndash890
39 ThymeSB JarjourJ TakeuchiR HavranekJJ AshworthJScharenbergAM StoddardBL and BakerD (2009)Exploitation of binding energy for catalysis and design Nature461 1300ndash1304
40 KentWJ (2002) BLATmdashthe BLAST-like alignment tool GenomeRes 12 656ndash664
41 TorikaiH ReikA LiuPQ ZhouY ZhangL MaitiSHulsH MillerJC KebriaeiP RabinovitchB et al (2012) Afoundation for universal T-cell based immunotherapy T cellsengineered to express a CD19-specific chimeric-antigen-receptorand eliminate expression of endogenous TCR Blood 1195697ndash5705
42 CertoMT GwiazdaKS KuharR SatherB CuringaGMandtT BraultM LambertAR BaxterSK JacobyK et al(2012) Coupling endonucleases with DNA end-processing enzymesto drive gene disruption Nat Methods 9 973ndash975
43 DelacoteF PerezC GuyotV DuhamelM RochonCOllivierN MacmasterR SilvaGH PaquesF DaboussiFet al (2013) High frequency targeted mutagenesis usingengineered endonucleases and DNA-end processing enzymesPLoS One 8 e53217
44 BaxterS LambertAR KuharR JarjourJ KulshinaNParmeggianiF DanaherP GanoJ BakerD StoddardBLet al (2012) Engineering domain fusion chimeras from I-OnuIfamily LAGLIDADG homing endonucleases Nucleic Acids Res40 7985ndash8000
Nucleic Acids Research 2013 11
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from

for the low kcat targets (0ndash3-fold for GT and mutNHEJ)(Figure 3b and d) However megaTAL activity againstthe low kcat target -1G was found to be significantlyincreased compared with mn treatment suggesting thatin some cases lsquoaddressingrsquo may partially compensate forreduced catalytic activity
Increasing on-target cleavage with megaTALs
One of the major shortcomings of current nuclease plat-forms is their potential for off-target cleavage and thenegative effects that this may have on the efficacy orsafety profile of a putative nuclease-based therapeutic ap-plication To evaluate the specificity properties of themegaTAL architecture we tested whether a megaTALcould consistently distinguish between lsquoaddressedrsquo andlsquononaddressedrsquo genomic target sites all cleaved by acommon mn cleavage domain Near-native sites of theI-AniI target (DNA sequences containing three or fewerbase-pair differences from the native I-AniI recognitionsequence) were previously identified in the humangenome using the UCSC BLAT tool (40) The cleavageactivity of the Y2 I-AniI variant mn was assessed at thesenear-natives targets using both episomal and endogenoustargets (data not shown) and the five sites that displayedthe highest levels of cleavage by Y2 in those assays werechosen for further examination (Figure 4a) SyntheticTAL effectors specific to regions upstream of the +9Tand+5A+8T near-native targets were built and fused tothe WT F13Y and Y2 I-AniI mns These megaTALs weretested alongside the stand-alone mn variants in TLRreporter cell lines for each of the five targets The +9Tand+5A+8T megaTALs showed increased levels of genetargeting at their addressed targets only while having noobserved effect on the activity at the remaining sites(Figure 4b) Notably while the addition of a sequence-specific TAL effector domain to the high-affinity Y2mndid not reduce its cleavage at nonaddressed targets (theY2mn and megaTALs show similar levels of GT at off-target sites) treatment with megaTALs made with thelower affinity WT and F13Y mns resulted in levels ofgene targeting at addressed targets equivalent to thatachieved with the Y2 megaTALs but without detectableoff-target activity
To verify that a similar effect would be observed atthe endogenous near-native loci 293T cells were treatedwith +9T megaTALs +5A+8T megaTALs and mnvariants and the level of mutNHEJ was determined ateach near-native site by high-throughput sequencing(Figure 4c Supplementary Figure S3a and b) Theseresults mirrored those from the reporter assay with theWT and F13Y megaTALs achieving indel rates up to75 at their addressed sites while maintaining near-background activity at nonaddressed sites
Highly effective knockout of the T-cell receptor alphagene in primary T-cells with a megaTAL
To verify that the benefits observed when fusing wild-typemns to a TAL effector would be achievable with anengineered mn and could be captured in a primary celltype in a relevant therapeutic application we designed a
variant of the I-OnuI mn to knockout the T-cell receptoralpha (TCRa) gene Knockout of the TCRa gene inhuman T-cells has been proposed as an approach toimprove the safety of T-cells endowed with chimericantigen receptors or heterologous TCRs by preventingthem from generating graft-versus-host disease (41)Thus knockout of TCRa in human T-cells is among themost translationally relevant applications of rare cleavingnuclease technologyIn vitro characterization of the TCRa mn showed that
its target site affinity and cleavage activity were similar tothat of its parent enzyme Specificity profiling of theTCRa mn and subsequent high-throughput in vitrocleavage analysis led to the identification of a numberof potential off-target sites which were cleaved with com-parable efficiency to the desired target (SupplementaryFigure S4a and e) To determine if generation of aTCRa megaTAL would yield improvements in activityand specificity the TCRa mn was fused to a 105 repeatTALE array designed to bind a DNA sequence upstreamof the TCRamn binding site (Figure 5a) T-cells were thentransfected with mRNA encoding either the TCRamegaTAL or the stand-alone mn In order to increasethe efficiency of gene knockout samples were includedwherein either the mn or the megaTAL were co-trans-fected with mRNA encoding Trex2 a 30 to 50 exonucleasethat has been shown to significantly increase rates ofmutNHEJ at mn cleavage sites (4243) Disruption ofthe TCRa gene was measured by staining for CD3 amulti-protein complex that is associated with the ab T-cell receptor Strikingly while disruption of the TCRagene occurred at minimal rates in cells treated with thestand-alone mn (16) the TCRa megaTAL exhibited a20-fold increase in activity at the addressed target site inthe TCRa gene (Figure 5b representative flow plots areshown in Figure 5c) Co-transfection of each nuclease withTrex2 further increased the rate of disruption yieldingrates of TCRa gene disruption consistently exceeding70Both on- and off-target cleavage rates were analyzed by
sequencing of the TCRa locus and putative off-target sites(identified based on in vitro cleavage profiling) in FACSsorted CD3- populations of nucleaseTrex2 treated cells(Figure 5d Supplementary Figure S5andashc) For each treat-ment modification rates at the TCRa locus were expectedto be 50 signifying knockout of the gene at theexpressing allele in conjunction with some additionalrate of modification of the nonexpressing allele InmegaTAL treated cells we observed 704 modificationsuggesting a substantial rate (ca 1 in 5 cells) of modifica-tion of the nonexpressed allele In contrast when weexpressed the megaTAL with Trex2 we observed thatonly 520 of the alleles carried indels and even lowerrates were obtained from the mn-alone treated cells Weattribute the differences in modification rates observed inthe presence and absence of Trex2 to lowered expressionof the nuclease in cells co-transfected with the exonucle-ase The lower modification rates observed in the lowabundance CD3- mn alone treated cells are attributableto contamination of the sorted populations with a largerproportion of background unstained cells The
Nucleic Acids Research 2013 7
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
sequencing analysis also demonstrated that off-targetcleavage in cells treated with the megaTAL alone is ex-tremely low with only one of 22 potential sites exhibitinga rate of cleavage above background in the presence ofTrex2 (OT23) Moreover the megaTAL produced lowerlevels of mutNHEJ at several putative off-target loci thanthe stand-alone mn (ie OT18) mdash this result may reflectstatistical variation in the deep sequencing results or al-ternatively could be accounted for by reduced expressionof the megaTAL nuclease compared with its shorter mncounterpart
DISCUSSION
The megaTAL nuclease architecture described hereaddresses several key limitations of existing nuclease
platforms for therapeutic applications as it enables thegeneration of compact single chain nucleases that are ex-tremely active and specific
MegaTALs exhibit dramatic increases in cleavageactivity compared with their isolated mn cleavage headsan effect that appears to be an outcome of the auxiliaryTAL effector DNA-binding domain docking (lsquoaddress-ingrsquo) the mn adjacent to its target and effectivelyincreasing the rate of enzymendashsubstrate complex forma-tion Furthermore the mn-mediated cleavage producesDNA ends with 30 overhangs that are efficiently processedby the Trex2 exonuclease thus allowing the co-expressionof a megaTAL and Trex2 to achieve unprecedentedly hightargeted modification rates in primary cells of gt70
MegaTALs offer an extreme level of target specificitydue to the combined DNA binding and cleavage
Name Genomic sequence Location
+9T TGTCATGTAAATAACTGTACTGTATGTTGAGGAGGTTTCTCTGTATA Ch9 (-)63127729-63127775
+5A+8T TCTTCTGCTGTTTCATATACCTCTATGTGAGGAGGTTTCTCAGTTAG Ch15(-)69553515-69553561
+2T+9G CATAGGTGACTTTTACTCTCATGATAGTGAGGAGGTTTTTCTGTAGA Ch12(+)84384994-84385040
+1C+5A GTCTTTGAATAAGTCACAATGCTTCTTTGAGGAGGTTCCTCAGTAAA Ch7 (+)28151155-28151201
-2A+1G ATGATG GAAATGTTACACTGAGCTCT TGAGGAGGATGCTCTGTAAA Ch1 (+)223534899-223534945
mut
NH
EJ
by
seq
()
+9T +5A+8T +1C+5A -2A+1G0
20
40
60
80
100
+2T+9G locus harbors an extra mutation in 293T cells data not shown
WT +9T WT MegaTAL +5A+8T WT MegaTALF13Y +9T F13Y MegaTAL +5A+8T F13Y MegaTALY2 +9T Y2 MegaTAL +5A+8T Y2 MegaTAL
TLR cell line
+9T +5A+8T +2T+9G +1C+5A -2A+1G0
1
2
3
Per
cent
GF
P+
cells
(a)
(b)
(c)
Figure 4 megaTAL addressing selectively increases on-target cleavage (a) I-AniI human genomic near-native sequences and their location in thegenome Synthetic TAL effectors were built against the+9T and+5A+8T sequences underlined in green and the I-AniI near-natives sequences areunderlined in blue (b) Level of gene targeting in TLR cells with endogenous human I-AniI near-native targets using lsquoaddressedrsquo+9T and+5A+8TmegaTALs and lsquounaddressedrsquo I-AniI mns mutNHEJ was not measured via TLR due to the presence of a stop codon in some target sites thatprevented flow cytometry readout of these events (c) Mutation rates at endogenous targets using lsquoaddressedrsquo I-AniI megaTALs and lsquounadressedrsquomns The level of mutNHEJ at the endogenous I-AniI near-native targets in 293T cells after 72 h of treatment with mn and megaTAL variants wasdetermined based on MiSeq sequencing results
8 Nucleic Acids Research 2013
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
properties of the protein architecture The activity ofmegaTALs is reliant on recognition of the tandem DNAtargets of the TALE array and the mn respectively ef-fectively extending the length of the target site andreducing the likelihood that more than one target sitewould be present in the human genome Similarly as
low affinity mn cleavage heads are not able to efficientlyaccess target sites to which they are not lsquoaddressedrsquo by theTALE array the potential for cleavage at nonaddressedmn recognition sites can be markedly reduced This is re-flected in the megaTAL used for TCRa gene knockout inprimary human T-cells which achieved 70 knockout
Control (-) TCRα mn TCRα mn + Trex2 TCRα mT + Trex2TCRα MegaTAL
Ch14 (+)23016490-23016529
100
mn megaTAL
0
20
40
60
80
CD
3- c
ells
()
- +- +
BFP-
--Trex2
Nuclease
5rsquo- TAAATCCAGTGACAAGTCTGTCTGCCTATTCACCGATTTT ATTTAGGTCACTGTTCAGACAGACGGATAAGTGGCTAAAA-5rsquo
NΔ154 C+63 TCRα mn
TCRα RVD array
283
0 50K 100K 150K 200K
0102
103
104
105
0866
0 50K 100K 150K 200K
519
0 50K 100K 150K 200K
368
0 50K 100K 150K 200K
756
0 50K 100K 150K 200K
CD
3
FSC
384
704
520
50 1000 0125 025 05 075 1 5 10 25NA indel by seq
Putative off target sitesTCRα
mn
mT
Trex2
+
+
-
-
003 0 0 0 0
006 0
0 05
0 1 2
0 14
0 03 0 0
004
0 02
0 08 0
002 0
0 01
0 11 0
OT 1 2 3 4 5 6 7 8 9 10 11 13 14 15 16 17 18 20 21 22 23 24
(a)
(c)
(d)
(b)
Figure 5 megaTAL targeting TCRa achieves extremely high gene disruption with no detectable off-target cleavage in human primary T-cells(a) Schematic of TCRa megaTAL formed by fusing a synthetic TAL effector built toward the TCRa sequence underlined in green to a designedTCRa mn (target underlined in blue) (b) TCR knockout achieved by parental TCRa mn and TCRa megaTAL in human primary T-cells Humanprimary T-cells were transfected with 10 micrograms of RNA encoding the indicated constructs with or without 10 micrograms of mRNA encodingthe Trex2 exonuclease TCRa gene disruption was assessed by measuring the percentage of cells that transition to being CD3- at 5 days followingtransfection Mutation rates at the endogenous TCRa gene were also assessed by Miseq sequencing of amplicons around the cleavage site andshowed indel rates gt50 suggesting that there was a preference for cleavage and disruption of the expressed TCRa allele particularly when themegaTAL is co-expressed with Trex2 (c) Representative flow cytometry data from transfection of human primary T-cells with TCRa targeting mn ormegaTALs+ Trex2 (d) Heat map showing mutation rates at the endogenous TCRa and putative off-target loci in CD3- primary T-cells treatedwith the TCRa mn or megaTAL+ Trex2 Map values were calculated for each locus by subtracting the background rate of indel formation in thecontrol population from that obtained for each nuclease treatment by Miseq sequencing with the heat map color scale indicated by the key (bottom)Sequencing results across all samples are shown (top) as well as an enlarged view with indel rates at putative off-target (OT) sites in cells treated withthe TCRa megaTAL alone (middle) Values calculated as 0 are given as lsquo0rsquo Sequencing results were not obtained for putative off-targets OT12 andOT19 and are therefore not shown on the map other samples for which sequencing results were not obtained are indicated as lsquoNArsquo
Nucleic Acids Research 2013 9
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
while above background off-target cleavage was limited toa single site (eg Figure 5d and Supplementary Figure 5)As the single off-target cleavage site identified is in anoncoding region of the genome this enzyme is of suffi-cient quality at its present state of development for clinicaluse However a further advantage of the megaTAL archi-tecture is the potential to further lsquotunersquo the mn cleavagehead to specifically avoid off-target cleavage sitesThus we anticipate that iterative refinement ofmegaTALs cleavage heads will allow the eventual identi-fication of megaTALs that couple high efficiency withperfect on target genomic specificityThe megaTAL architecture also possesses several
properties that facilitate efficient delivery for therapeuticapplications MegaTALs can be expressed with a single2 kb ORF smaller than two ZFN halves [22 kb total(11 kb per ZFN half targeting a 9-bp target)] and signifi-cantly shorter than a single TALEN half [gt50 kb total(25 kb per TALEN half targeting a 13-bp target)] orCas9 nuclease (41 kb not including guide RNA) ThusmegaTALs are compatible with efficient vectorization as aDNA mRNA or with any widely used viral vector systemThe capacity to use a TAL effector domain with as few as55 repeats also facilitates codon divergence of the TALeffector repeats to allow packaging into retroviral vectorswhere recombination among TAL effector RVDs duringreverse transcription can be problematic (25) Finally theindependent function of megaTALs due to their singlechain design avoids the possibility of inappropriateheterodimerization that may occur with enzymes basedon dimerization-dependent cleavage domains such asFokI an important advantage for applications requiringmultiplex genetic modifications
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Onlineincluding [44]
ACKNOWLEDGEMENTS
We would like to thank members of the NorthwestGenome Engineering Consortium (NGEC) (httpngec-seattleorg) for their many insightful discussions
FUNDING
National Institutes of Health [RL1CA133832UL1DE019582 R01-HL075453 PL1-HL092557 RL1-HL092553 RL1-HL92554 and U19-AI96111] SeattleChildrenrsquos Center for Immunity and ImmunotherapiesFunding for open access charge NIH [U19-AI96111]
Conflict of interest statement AMS is an employee ofCellectis therapeutics and receives salary and equity com-pensation He is also founder member of the board of dir-ectors and equity holder in Pregenen Inc JJ is an employeeof Pregenen Inc and receives salary and equitycompensation AA is an employee of Pregenen Inc andreceives salary and equity compensation PD is an
employee of Cellectis therapeutics and receives salary andequity compensation AG is an employee of Cellectistherapeutics and receives salary and equity compensation
REFERENCES
1 BakerM (2012) Gene-editing nucleases Nat Methods 9 23ndash262 KimYG ChaJ and ChandrasegaranS (1996) Hybrid
restriction enzymes zinc finger fusions to Fok I cleavage domainProc Natl Acad Sci USA 93 1156ndash1160
3 KlugA (2010) The discovery of zinc fingers and theirapplications in gene regulation and genome manipulation AnnRev Biochem 79 213ndash231
4 ChristianM CermakT DoyleEL SchmidtC ZhangFHummelA BogdanoveAJ and VoytasDF (2010) TargetingDNA double-strand breaks with TAL effector nucleases Genetics186 757ndash761
5 LiT HuangS JiangWZ WrightD SpaldingMHWeeksDP and YangB (2011) TAL nucleases (TALNs) hybridproteins composed of TAL effectors and FokI DNA-cleavagedomain Nucleic Acids Res 39 359ndash372
6 PaquesF and DuchateauP (2007) Meganucleases and DNAdouble-strand break-induced recombination perspectives for genetherapy Curr Gene Ther 7 49ndash66
7 StoddardBL (2011) Homing endonucleases from microbialgenetic invaders to reagents for targeted DNA modificationStructure 19 7ndash15
8 JinekM ChylinskiK FonfaraI HauerM DoudnaJA andCharpentierE (2012) A programmable dual-RNAndashguided DNAendonuclease in adaptive bacterial immunity Science 337816ndash821
9 MaliP YangL EsveltKM AachJ GuellM DiCarloJENorvilleJE and ChurchGM (2013) RNA-guided humangenome engineering via Cas9 Science 339 823ndash826
10 McMahonMA RahdarM and PorteusM (2012) Gene editingnot just for translation anymore Nat Methods 9 28ndash31
11 LieberMR (2010) The mechanism of double-strand DNA breakrepair by the nonhomologous DNA end-joining pathway AnnRev Biochem 79 181ndash211
12 ShrivastavM DE HaroLP and NickoloffJA (2008)Regulation of DNA double-strand break repair pathway choiceCell Res 18 134ndash147
13 Method of the Year 2011 Nat Methods 9 114 MandellJG and BarbasCF (2006) Zinc finger tools custom
DNA-binding domains for transcription factors and nucleasesNucleic Acids Res 34 W516ndashW523
15 MaederML Thibodeau-BegannyS OsiakA WrightDAAnthonyRM EichtingerM JiangT FoleyJE WinfreyRJTownsendJA et al (2008) Rapid lsquoopen-sourcersquo engineering ofcustomized zinc-finger nucleases for highly efficient genemodification Mol Cell 31 294ndash301
16 SanderJD DahlborgEJ GoodwinMJ CadeL ZhangFCifuentesD CurtinSJ BlackburnJS Thibodeau-BegannySQiY et al (2011) Selection-free zinc-finger-nuclease engineeringby context-dependent assembly (CoDA) Nat Methods 8 67ndash69
17 CermakT DoyleEL ChristianM WangL ZhangYSchmidtC BallerJA SomiaNV BogdanoveAJ andVoytasDF (2011) Efficient design and assembly of customTALEN and other TAL effector-based constructs for DNAtargeting Nucleic Acids Res 39 e82ndashe82
18 ZhangF CongL LodatoS KosuriS ChurchGM andArlottaP (2011) Efficient construction of sequence-specific TALeffectors for modulating mammalian transcription NatBiotechnol 29 149ndash153
19 ReyonD TsaiSQ KhayterC FodenJA SanderJD andJoungJK (2012) FLASH assembly of TALENs for high-throughput genome editing Nat Biotechnol 30 460ndash465
20 JarjourJ West-FoyleH CertoMT HubertCG DoyleLGetzMM StoddardBL and ScharenbergAM (2009) High-resolution profiling of homing endonuclease binding and catalyticspecificity using yeast surface display Nucleic Acids Res 376871ndash6880
10 Nucleic Acids Research 2013
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
21 CornuTI Thibodeau-BegannyS GuhlE AlwinSEichtingerM JoungJK JoungJK and CathomenT (2008)DNA-binding specificity is a major determinant of the activityand toxicity of zinc-finger nucleases Mol Ther 16 352ndash358
22 GabrielR LombardoA ArensA MillerJC GenovesePKaeppelC NowrouziA BartholomaeCC WangJFriedmanG et al (2011) An unbiased genome-wide analysis ofzinc-finger nuclease specificity Nat Biotechnol 29 816ndash823
23 PattanayakV RamirezCL JoungJK and LiuDR (2011)Revealing off-target cleavage specificities of zinc-finger nucleasesby in vitro selection Nat Methods 8 765ndash770
24 SolluC ParsK CornuTI Thibodeau-BegannySMaederML JoungJK HeilbronnR and CathomenT (2010)Autonomous zinc-finger nuclease pairs for targeted chromosomaldeletion Nucleic Acids Res 38 8269ndash8276
25 HolkersM MaggioI LiuJ JanssenJM MiselliFMussolinoC RecchiaA CathomenT and GoncalvesMA(2012) Differential integrity of TALE nuclease genes followingadenoviral and lentiviral vector gene transfer into human cellsNucleic Acids Res 41 e63
26 FuY FodenJA KhayterC MaederML ReyonDJoungJK and SanderJD (2013) High-frequency off-targetmutagenesis induced by CRISPR-Cas nucleases in human cellsNat Biotechnol 31 822ndash826
27 CradickTJ FineEJ AnticoCJ and BaoG (2013) CRISPRCas9 systems targeting b-globin and CCR5 genes have substantialoff-target activity Nucleic Acids Res 41 9584ndash9592
28 RanFA HsuPD LinCY GootenbergJS KonermannSTrevinoAE ScottDA InoueA MatobaS ZhangY et al(2013) Double nicking by RNA-guided CRISPR Cas9 forenhanced genome editing specificity Cell 154 1380ndash1389
29 AshworthJ TaylorGK HavranekJJ QuadriSAStoddardBL and BakerD (2010) Computationalreprogramming of homing endonuclease specificity at multipleadjacent base pairs Nucleic Acids Res 38 5601ndash5608
30 TakeuchiR LambertAR MakAN JacobyK DicksonRJGloorGB ScharenbergAM EdgellDR and StoddardBL(2011) Tapping natural reservoirs of homing endonucleases fortargeted gene modification Proc Natl Acad Sci USA 10813077ndash13082
31 SatherBD RyuBY StirlingBV GaribovM KernsHMHumblet-BaronS AstrakhanA and RawlingsDJ (2011)Development of B-lineage predominant lentiviral vectors for usein genetic therapies for b cell disorders Mol Ther 19 515ndash525
32 CertoMT RyuBY AnnisJE GaribovM JarjourJRawlingsDJ and ScharenbergAM (2011) Tracking genomeengineering outcome at individual DNA breakpoints NatMethods 8 671ndash676
33 KuharR GwiazdaKS HumbertO MandtT PangalloJBraultM KhanI MaizelsN RawlingsDJ ScharenbergAMet al (2013) Novel fluorescent genome editing reporters formonitoring DNA repair pathway utilization at endonuclease-induced breaks Nucleic Acids Res (epub ahead of print October10 2013)
34 BochJ ScholzeH SchornackS LandgrafA HahnS KaySLahayeT NickstadtA and BonasU (2009) Breaking the codeof DNA binding specificity of TAL-type III effectors Science326 1509ndash1512
35 MoscouMJ and BogdanoveAJ (2009) A simple cipher governsDNA recognition by TAL effectors Science 326 1501ndash1501
36 GaoH WuX ChaiJ and HanZ (2012) Crystal structure of aTALE protein reveals an extended N-terminal DNA bindingregion Cell Res 22 1716ndash1720
37 MillerJC TanS QiaoG BarlowKA WangJ XiaDFMengX PaschonDE LeungE HinkleySJ et al (2011) ATALE nuclease architecture for efficient genome editing NatBiotechnol 29 143ndash148
38 TakeuchiR CertoM CapraraMG ScharenbergAM andStoddardBL (2009) Optimization of in vivo activity of abifunctional homing endonuclease and maturase reversesevolutionary degradation Nucleic Acids Res 37 877ndash890
39 ThymeSB JarjourJ TakeuchiR HavranekJJ AshworthJScharenbergAM StoddardBL and BakerD (2009)Exploitation of binding energy for catalysis and design Nature461 1300ndash1304
40 KentWJ (2002) BLATmdashthe BLAST-like alignment tool GenomeRes 12 656ndash664
41 TorikaiH ReikA LiuPQ ZhouY ZhangL MaitiSHulsH MillerJC KebriaeiP RabinovitchB et al (2012) Afoundation for universal T-cell based immunotherapy T cellsengineered to express a CD19-specific chimeric-antigen-receptorand eliminate expression of endogenous TCR Blood 1195697ndash5705
42 CertoMT GwiazdaKS KuharR SatherB CuringaGMandtT BraultM LambertAR BaxterSK JacobyK et al(2012) Coupling endonucleases with DNA end-processing enzymesto drive gene disruption Nat Methods 9 973ndash975
43 DelacoteF PerezC GuyotV DuhamelM RochonCOllivierN MacmasterR SilvaGH PaquesF DaboussiFet al (2013) High frequency targeted mutagenesis usingengineered endonucleases and DNA-end processing enzymesPLoS One 8 e53217
44 BaxterS LambertAR KuharR JarjourJ KulshinaNParmeggianiF DanaherP GanoJ BakerD StoddardBLet al (2012) Engineering domain fusion chimeras from I-OnuIfamily LAGLIDADG homing endonucleases Nucleic Acids Res40 7985ndash8000
Nucleic Acids Research 2013 11
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from

sequencing analysis also demonstrated that off-targetcleavage in cells treated with the megaTAL alone is ex-tremely low with only one of 22 potential sites exhibitinga rate of cleavage above background in the presence ofTrex2 (OT23) Moreover the megaTAL produced lowerlevels of mutNHEJ at several putative off-target loci thanthe stand-alone mn (ie OT18) mdash this result may reflectstatistical variation in the deep sequencing results or al-ternatively could be accounted for by reduced expressionof the megaTAL nuclease compared with its shorter mncounterpart
DISCUSSION
The megaTAL nuclease architecture described hereaddresses several key limitations of existing nuclease
platforms for therapeutic applications as it enables thegeneration of compact single chain nucleases that are ex-tremely active and specific
MegaTALs exhibit dramatic increases in cleavageactivity compared with their isolated mn cleavage headsan effect that appears to be an outcome of the auxiliaryTAL effector DNA-binding domain docking (lsquoaddress-ingrsquo) the mn adjacent to its target and effectivelyincreasing the rate of enzymendashsubstrate complex forma-tion Furthermore the mn-mediated cleavage producesDNA ends with 30 overhangs that are efficiently processedby the Trex2 exonuclease thus allowing the co-expressionof a megaTAL and Trex2 to achieve unprecedentedly hightargeted modification rates in primary cells of gt70
MegaTALs offer an extreme level of target specificitydue to the combined DNA binding and cleavage
Name Genomic sequence Location
+9T TGTCATGTAAATAACTGTACTGTATGTTGAGGAGGTTTCTCTGTATA Ch9 (-)63127729-63127775
+5A+8T TCTTCTGCTGTTTCATATACCTCTATGTGAGGAGGTTTCTCAGTTAG Ch15(-)69553515-69553561
+2T+9G CATAGGTGACTTTTACTCTCATGATAGTGAGGAGGTTTTTCTGTAGA Ch12(+)84384994-84385040
+1C+5A GTCTTTGAATAAGTCACAATGCTTCTTTGAGGAGGTTCCTCAGTAAA Ch7 (+)28151155-28151201
-2A+1G ATGATG GAAATGTTACACTGAGCTCT TGAGGAGGATGCTCTGTAAA Ch1 (+)223534899-223534945
mut
NH
EJ
by
seq
()
+9T +5A+8T +1C+5A -2A+1G0
20
40
60
80
100
+2T+9G locus harbors an extra mutation in 293T cells data not shown
WT +9T WT MegaTAL +5A+8T WT MegaTALF13Y +9T F13Y MegaTAL +5A+8T F13Y MegaTALY2 +9T Y2 MegaTAL +5A+8T Y2 MegaTAL
TLR cell line
+9T +5A+8T +2T+9G +1C+5A -2A+1G0
1
2
3
Per
cent
GF
P+
cells
(a)
(b)
(c)
Figure 4 megaTAL addressing selectively increases on-target cleavage (a) I-AniI human genomic near-native sequences and their location in thegenome Synthetic TAL effectors were built against the+9T and+5A+8T sequences underlined in green and the I-AniI near-natives sequences areunderlined in blue (b) Level of gene targeting in TLR cells with endogenous human I-AniI near-native targets using lsquoaddressedrsquo+9T and+5A+8TmegaTALs and lsquounaddressedrsquo I-AniI mns mutNHEJ was not measured via TLR due to the presence of a stop codon in some target sites thatprevented flow cytometry readout of these events (c) Mutation rates at endogenous targets using lsquoaddressedrsquo I-AniI megaTALs and lsquounadressedrsquomns The level of mutNHEJ at the endogenous I-AniI near-native targets in 293T cells after 72 h of treatment with mn and megaTAL variants wasdetermined based on MiSeq sequencing results
8 Nucleic Acids Research 2013
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
properties of the protein architecture The activity ofmegaTALs is reliant on recognition of the tandem DNAtargets of the TALE array and the mn respectively ef-fectively extending the length of the target site andreducing the likelihood that more than one target sitewould be present in the human genome Similarly as
low affinity mn cleavage heads are not able to efficientlyaccess target sites to which they are not lsquoaddressedrsquo by theTALE array the potential for cleavage at nonaddressedmn recognition sites can be markedly reduced This is re-flected in the megaTAL used for TCRa gene knockout inprimary human T-cells which achieved 70 knockout
Control (-) TCRα mn TCRα mn + Trex2 TCRα mT + Trex2TCRα MegaTAL
Ch14 (+)23016490-23016529
100
mn megaTAL
0
20
40
60
80
CD
3- c
ells
()
- +- +
BFP-
--Trex2
Nuclease
5rsquo- TAAATCCAGTGACAAGTCTGTCTGCCTATTCACCGATTTT ATTTAGGTCACTGTTCAGACAGACGGATAAGTGGCTAAAA-5rsquo
NΔ154 C+63 TCRα mn
TCRα RVD array
283
0 50K 100K 150K 200K
0102
103
104
105
0866
0 50K 100K 150K 200K
519
0 50K 100K 150K 200K
368
0 50K 100K 150K 200K
756
0 50K 100K 150K 200K
CD
3
FSC
384
704
520
50 1000 0125 025 05 075 1 5 10 25NA indel by seq
Putative off target sitesTCRα
mn
mT
Trex2
+
+
-
-
003 0 0 0 0
006 0
0 05
0 1 2
0 14
0 03 0 0
004
0 02
0 08 0
002 0
0 01
0 11 0
OT 1 2 3 4 5 6 7 8 9 10 11 13 14 15 16 17 18 20 21 22 23 24
(a)
(c)
(d)
(b)
Figure 5 megaTAL targeting TCRa achieves extremely high gene disruption with no detectable off-target cleavage in human primary T-cells(a) Schematic of TCRa megaTAL formed by fusing a synthetic TAL effector built toward the TCRa sequence underlined in green to a designedTCRa mn (target underlined in blue) (b) TCR knockout achieved by parental TCRa mn and TCRa megaTAL in human primary T-cells Humanprimary T-cells were transfected with 10 micrograms of RNA encoding the indicated constructs with or without 10 micrograms of mRNA encodingthe Trex2 exonuclease TCRa gene disruption was assessed by measuring the percentage of cells that transition to being CD3- at 5 days followingtransfection Mutation rates at the endogenous TCRa gene were also assessed by Miseq sequencing of amplicons around the cleavage site andshowed indel rates gt50 suggesting that there was a preference for cleavage and disruption of the expressed TCRa allele particularly when themegaTAL is co-expressed with Trex2 (c) Representative flow cytometry data from transfection of human primary T-cells with TCRa targeting mn ormegaTALs+ Trex2 (d) Heat map showing mutation rates at the endogenous TCRa and putative off-target loci in CD3- primary T-cells treatedwith the TCRa mn or megaTAL+ Trex2 Map values were calculated for each locus by subtracting the background rate of indel formation in thecontrol population from that obtained for each nuclease treatment by Miseq sequencing with the heat map color scale indicated by the key (bottom)Sequencing results across all samples are shown (top) as well as an enlarged view with indel rates at putative off-target (OT) sites in cells treated withthe TCRa megaTAL alone (middle) Values calculated as 0 are given as lsquo0rsquo Sequencing results were not obtained for putative off-targets OT12 andOT19 and are therefore not shown on the map other samples for which sequencing results were not obtained are indicated as lsquoNArsquo
Nucleic Acids Research 2013 9
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
while above background off-target cleavage was limited toa single site (eg Figure 5d and Supplementary Figure 5)As the single off-target cleavage site identified is in anoncoding region of the genome this enzyme is of suffi-cient quality at its present state of development for clinicaluse However a further advantage of the megaTAL archi-tecture is the potential to further lsquotunersquo the mn cleavagehead to specifically avoid off-target cleavage sitesThus we anticipate that iterative refinement ofmegaTALs cleavage heads will allow the eventual identi-fication of megaTALs that couple high efficiency withperfect on target genomic specificityThe megaTAL architecture also possesses several
properties that facilitate efficient delivery for therapeuticapplications MegaTALs can be expressed with a single2 kb ORF smaller than two ZFN halves [22 kb total(11 kb per ZFN half targeting a 9-bp target)] and signifi-cantly shorter than a single TALEN half [gt50 kb total(25 kb per TALEN half targeting a 13-bp target)] orCas9 nuclease (41 kb not including guide RNA) ThusmegaTALs are compatible with efficient vectorization as aDNA mRNA or with any widely used viral vector systemThe capacity to use a TAL effector domain with as few as55 repeats also facilitates codon divergence of the TALeffector repeats to allow packaging into retroviral vectorswhere recombination among TAL effector RVDs duringreverse transcription can be problematic (25) Finally theindependent function of megaTALs due to their singlechain design avoids the possibility of inappropriateheterodimerization that may occur with enzymes basedon dimerization-dependent cleavage domains such asFokI an important advantage for applications requiringmultiplex genetic modifications
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Onlineincluding [44]
ACKNOWLEDGEMENTS
We would like to thank members of the NorthwestGenome Engineering Consortium (NGEC) (httpngec-seattleorg) for their many insightful discussions
FUNDING
National Institutes of Health [RL1CA133832UL1DE019582 R01-HL075453 PL1-HL092557 RL1-HL092553 RL1-HL92554 and U19-AI96111] SeattleChildrenrsquos Center for Immunity and ImmunotherapiesFunding for open access charge NIH [U19-AI96111]
Conflict of interest statement AMS is an employee ofCellectis therapeutics and receives salary and equity com-pensation He is also founder member of the board of dir-ectors and equity holder in Pregenen Inc JJ is an employeeof Pregenen Inc and receives salary and equitycompensation AA is an employee of Pregenen Inc andreceives salary and equity compensation PD is an
employee of Cellectis therapeutics and receives salary andequity compensation AG is an employee of Cellectistherapeutics and receives salary and equity compensation
REFERENCES
1 BakerM (2012) Gene-editing nucleases Nat Methods 9 23ndash262 KimYG ChaJ and ChandrasegaranS (1996) Hybrid
restriction enzymes zinc finger fusions to Fok I cleavage domainProc Natl Acad Sci USA 93 1156ndash1160
3 KlugA (2010) The discovery of zinc fingers and theirapplications in gene regulation and genome manipulation AnnRev Biochem 79 213ndash231
4 ChristianM CermakT DoyleEL SchmidtC ZhangFHummelA BogdanoveAJ and VoytasDF (2010) TargetingDNA double-strand breaks with TAL effector nucleases Genetics186 757ndash761
5 LiT HuangS JiangWZ WrightD SpaldingMHWeeksDP and YangB (2011) TAL nucleases (TALNs) hybridproteins composed of TAL effectors and FokI DNA-cleavagedomain Nucleic Acids Res 39 359ndash372
6 PaquesF and DuchateauP (2007) Meganucleases and DNAdouble-strand break-induced recombination perspectives for genetherapy Curr Gene Ther 7 49ndash66
7 StoddardBL (2011) Homing endonucleases from microbialgenetic invaders to reagents for targeted DNA modificationStructure 19 7ndash15
8 JinekM ChylinskiK FonfaraI HauerM DoudnaJA andCharpentierE (2012) A programmable dual-RNAndashguided DNAendonuclease in adaptive bacterial immunity Science 337816ndash821
9 MaliP YangL EsveltKM AachJ GuellM DiCarloJENorvilleJE and ChurchGM (2013) RNA-guided humangenome engineering via Cas9 Science 339 823ndash826
10 McMahonMA RahdarM and PorteusM (2012) Gene editingnot just for translation anymore Nat Methods 9 28ndash31
11 LieberMR (2010) The mechanism of double-strand DNA breakrepair by the nonhomologous DNA end-joining pathway AnnRev Biochem 79 181ndash211
12 ShrivastavM DE HaroLP and NickoloffJA (2008)Regulation of DNA double-strand break repair pathway choiceCell Res 18 134ndash147
13 Method of the Year 2011 Nat Methods 9 114 MandellJG and BarbasCF (2006) Zinc finger tools custom
DNA-binding domains for transcription factors and nucleasesNucleic Acids Res 34 W516ndashW523
15 MaederML Thibodeau-BegannyS OsiakA WrightDAAnthonyRM EichtingerM JiangT FoleyJE WinfreyRJTownsendJA et al (2008) Rapid lsquoopen-sourcersquo engineering ofcustomized zinc-finger nucleases for highly efficient genemodification Mol Cell 31 294ndash301
16 SanderJD DahlborgEJ GoodwinMJ CadeL ZhangFCifuentesD CurtinSJ BlackburnJS Thibodeau-BegannySQiY et al (2011) Selection-free zinc-finger-nuclease engineeringby context-dependent assembly (CoDA) Nat Methods 8 67ndash69
17 CermakT DoyleEL ChristianM WangL ZhangYSchmidtC BallerJA SomiaNV BogdanoveAJ andVoytasDF (2011) Efficient design and assembly of customTALEN and other TAL effector-based constructs for DNAtargeting Nucleic Acids Res 39 e82ndashe82
18 ZhangF CongL LodatoS KosuriS ChurchGM andArlottaP (2011) Efficient construction of sequence-specific TALeffectors for modulating mammalian transcription NatBiotechnol 29 149ndash153
19 ReyonD TsaiSQ KhayterC FodenJA SanderJD andJoungJK (2012) FLASH assembly of TALENs for high-throughput genome editing Nat Biotechnol 30 460ndash465
20 JarjourJ West-FoyleH CertoMT HubertCG DoyleLGetzMM StoddardBL and ScharenbergAM (2009) High-resolution profiling of homing endonuclease binding and catalyticspecificity using yeast surface display Nucleic Acids Res 376871ndash6880
10 Nucleic Acids Research 2013
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
21 CornuTI Thibodeau-BegannyS GuhlE AlwinSEichtingerM JoungJK JoungJK and CathomenT (2008)DNA-binding specificity is a major determinant of the activityand toxicity of zinc-finger nucleases Mol Ther 16 352ndash358
22 GabrielR LombardoA ArensA MillerJC GenovesePKaeppelC NowrouziA BartholomaeCC WangJFriedmanG et al (2011) An unbiased genome-wide analysis ofzinc-finger nuclease specificity Nat Biotechnol 29 816ndash823
23 PattanayakV RamirezCL JoungJK and LiuDR (2011)Revealing off-target cleavage specificities of zinc-finger nucleasesby in vitro selection Nat Methods 8 765ndash770
24 SolluC ParsK CornuTI Thibodeau-BegannySMaederML JoungJK HeilbronnR and CathomenT (2010)Autonomous zinc-finger nuclease pairs for targeted chromosomaldeletion Nucleic Acids Res 38 8269ndash8276
25 HolkersM MaggioI LiuJ JanssenJM MiselliFMussolinoC RecchiaA CathomenT and GoncalvesMA(2012) Differential integrity of TALE nuclease genes followingadenoviral and lentiviral vector gene transfer into human cellsNucleic Acids Res 41 e63
26 FuY FodenJA KhayterC MaederML ReyonDJoungJK and SanderJD (2013) High-frequency off-targetmutagenesis induced by CRISPR-Cas nucleases in human cellsNat Biotechnol 31 822ndash826
27 CradickTJ FineEJ AnticoCJ and BaoG (2013) CRISPRCas9 systems targeting b-globin and CCR5 genes have substantialoff-target activity Nucleic Acids Res 41 9584ndash9592
28 RanFA HsuPD LinCY GootenbergJS KonermannSTrevinoAE ScottDA InoueA MatobaS ZhangY et al(2013) Double nicking by RNA-guided CRISPR Cas9 forenhanced genome editing specificity Cell 154 1380ndash1389
29 AshworthJ TaylorGK HavranekJJ QuadriSAStoddardBL and BakerD (2010) Computationalreprogramming of homing endonuclease specificity at multipleadjacent base pairs Nucleic Acids Res 38 5601ndash5608
30 TakeuchiR LambertAR MakAN JacobyK DicksonRJGloorGB ScharenbergAM EdgellDR and StoddardBL(2011) Tapping natural reservoirs of homing endonucleases fortargeted gene modification Proc Natl Acad Sci USA 10813077ndash13082
31 SatherBD RyuBY StirlingBV GaribovM KernsHMHumblet-BaronS AstrakhanA and RawlingsDJ (2011)Development of B-lineage predominant lentiviral vectors for usein genetic therapies for b cell disorders Mol Ther 19 515ndash525
32 CertoMT RyuBY AnnisJE GaribovM JarjourJRawlingsDJ and ScharenbergAM (2011) Tracking genomeengineering outcome at individual DNA breakpoints NatMethods 8 671ndash676
33 KuharR GwiazdaKS HumbertO MandtT PangalloJBraultM KhanI MaizelsN RawlingsDJ ScharenbergAMet al (2013) Novel fluorescent genome editing reporters formonitoring DNA repair pathway utilization at endonuclease-induced breaks Nucleic Acids Res (epub ahead of print October10 2013)
34 BochJ ScholzeH SchornackS LandgrafA HahnS KaySLahayeT NickstadtA and BonasU (2009) Breaking the codeof DNA binding specificity of TAL-type III effectors Science326 1509ndash1512
35 MoscouMJ and BogdanoveAJ (2009) A simple cipher governsDNA recognition by TAL effectors Science 326 1501ndash1501
36 GaoH WuX ChaiJ and HanZ (2012) Crystal structure of aTALE protein reveals an extended N-terminal DNA bindingregion Cell Res 22 1716ndash1720
37 MillerJC TanS QiaoG BarlowKA WangJ XiaDFMengX PaschonDE LeungE HinkleySJ et al (2011) ATALE nuclease architecture for efficient genome editing NatBiotechnol 29 143ndash148
38 TakeuchiR CertoM CapraraMG ScharenbergAM andStoddardBL (2009) Optimization of in vivo activity of abifunctional homing endonuclease and maturase reversesevolutionary degradation Nucleic Acids Res 37 877ndash890
39 ThymeSB JarjourJ TakeuchiR HavranekJJ AshworthJScharenbergAM StoddardBL and BakerD (2009)Exploitation of binding energy for catalysis and design Nature461 1300ndash1304
40 KentWJ (2002) BLATmdashthe BLAST-like alignment tool GenomeRes 12 656ndash664
41 TorikaiH ReikA LiuPQ ZhouY ZhangL MaitiSHulsH MillerJC KebriaeiP RabinovitchB et al (2012) Afoundation for universal T-cell based immunotherapy T cellsengineered to express a CD19-specific chimeric-antigen-receptorand eliminate expression of endogenous TCR Blood 1195697ndash5705
42 CertoMT GwiazdaKS KuharR SatherB CuringaGMandtT BraultM LambertAR BaxterSK JacobyK et al(2012) Coupling endonucleases with DNA end-processing enzymesto drive gene disruption Nat Methods 9 973ndash975
43 DelacoteF PerezC GuyotV DuhamelM RochonCOllivierN MacmasterR SilvaGH PaquesF DaboussiFet al (2013) High frequency targeted mutagenesis usingengineered endonucleases and DNA-end processing enzymesPLoS One 8 e53217
44 BaxterS LambertAR KuharR JarjourJ KulshinaNParmeggianiF DanaherP GanoJ BakerD StoddardBLet al (2012) Engineering domain fusion chimeras from I-OnuIfamily LAGLIDADG homing endonucleases Nucleic Acids Res40 7985ndash8000
Nucleic Acids Research 2013 11
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
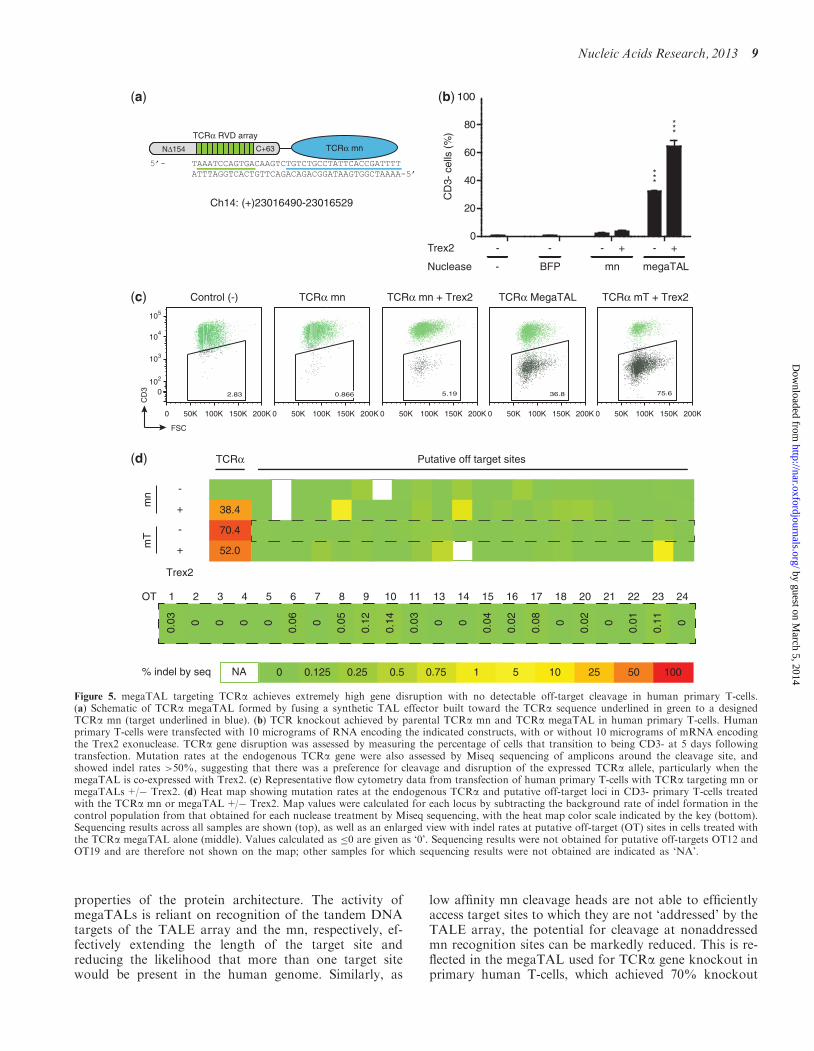
properties of the protein architecture The activity ofmegaTALs is reliant on recognition of the tandem DNAtargets of the TALE array and the mn respectively ef-fectively extending the length of the target site andreducing the likelihood that more than one target sitewould be present in the human genome Similarly as
low affinity mn cleavage heads are not able to efficientlyaccess target sites to which they are not lsquoaddressedrsquo by theTALE array the potential for cleavage at nonaddressedmn recognition sites can be markedly reduced This is re-flected in the megaTAL used for TCRa gene knockout inprimary human T-cells which achieved 70 knockout
Control (-) TCRα mn TCRα mn + Trex2 TCRα mT + Trex2TCRα MegaTAL
Ch14 (+)23016490-23016529
100
mn megaTAL
0
20
40
60
80
CD
3- c
ells
()
- +- +
BFP-
--Trex2
Nuclease
5rsquo- TAAATCCAGTGACAAGTCTGTCTGCCTATTCACCGATTTT ATTTAGGTCACTGTTCAGACAGACGGATAAGTGGCTAAAA-5rsquo
NΔ154 C+63 TCRα mn
TCRα RVD array
283
0 50K 100K 150K 200K
0102
103
104
105
0866
0 50K 100K 150K 200K
519
0 50K 100K 150K 200K
368
0 50K 100K 150K 200K
756
0 50K 100K 150K 200K
CD
3
FSC
384
704
520
50 1000 0125 025 05 075 1 5 10 25NA indel by seq
Putative off target sitesTCRα
mn
mT
Trex2
+
+
-
-
003 0 0 0 0
006 0
0 05
0 1 2
0 14
0 03 0 0
004
0 02
0 08 0
002 0
0 01
0 11 0
OT 1 2 3 4 5 6 7 8 9 10 11 13 14 15 16 17 18 20 21 22 23 24
(a)
(c)
(d)
(b)
Figure 5 megaTAL targeting TCRa achieves extremely high gene disruption with no detectable off-target cleavage in human primary T-cells(a) Schematic of TCRa megaTAL formed by fusing a synthetic TAL effector built toward the TCRa sequence underlined in green to a designedTCRa mn (target underlined in blue) (b) TCR knockout achieved by parental TCRa mn and TCRa megaTAL in human primary T-cells Humanprimary T-cells were transfected with 10 micrograms of RNA encoding the indicated constructs with or without 10 micrograms of mRNA encodingthe Trex2 exonuclease TCRa gene disruption was assessed by measuring the percentage of cells that transition to being CD3- at 5 days followingtransfection Mutation rates at the endogenous TCRa gene were also assessed by Miseq sequencing of amplicons around the cleavage site andshowed indel rates gt50 suggesting that there was a preference for cleavage and disruption of the expressed TCRa allele particularly when themegaTAL is co-expressed with Trex2 (c) Representative flow cytometry data from transfection of human primary T-cells with TCRa targeting mn ormegaTALs+ Trex2 (d) Heat map showing mutation rates at the endogenous TCRa and putative off-target loci in CD3- primary T-cells treatedwith the TCRa mn or megaTAL+ Trex2 Map values were calculated for each locus by subtracting the background rate of indel formation in thecontrol population from that obtained for each nuclease treatment by Miseq sequencing with the heat map color scale indicated by the key (bottom)Sequencing results across all samples are shown (top) as well as an enlarged view with indel rates at putative off-target (OT) sites in cells treated withthe TCRa megaTAL alone (middle) Values calculated as 0 are given as lsquo0rsquo Sequencing results were not obtained for putative off-targets OT12 andOT19 and are therefore not shown on the map other samples for which sequencing results were not obtained are indicated as lsquoNArsquo
Nucleic Acids Research 2013 9
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
while above background off-target cleavage was limited toa single site (eg Figure 5d and Supplementary Figure 5)As the single off-target cleavage site identified is in anoncoding region of the genome this enzyme is of suffi-cient quality at its present state of development for clinicaluse However a further advantage of the megaTAL archi-tecture is the potential to further lsquotunersquo the mn cleavagehead to specifically avoid off-target cleavage sitesThus we anticipate that iterative refinement ofmegaTALs cleavage heads will allow the eventual identi-fication of megaTALs that couple high efficiency withperfect on target genomic specificityThe megaTAL architecture also possesses several
properties that facilitate efficient delivery for therapeuticapplications MegaTALs can be expressed with a single2 kb ORF smaller than two ZFN halves [22 kb total(11 kb per ZFN half targeting a 9-bp target)] and signifi-cantly shorter than a single TALEN half [gt50 kb total(25 kb per TALEN half targeting a 13-bp target)] orCas9 nuclease (41 kb not including guide RNA) ThusmegaTALs are compatible with efficient vectorization as aDNA mRNA or with any widely used viral vector systemThe capacity to use a TAL effector domain with as few as55 repeats also facilitates codon divergence of the TALeffector repeats to allow packaging into retroviral vectorswhere recombination among TAL effector RVDs duringreverse transcription can be problematic (25) Finally theindependent function of megaTALs due to their singlechain design avoids the possibility of inappropriateheterodimerization that may occur with enzymes basedon dimerization-dependent cleavage domains such asFokI an important advantage for applications requiringmultiplex genetic modifications
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Onlineincluding [44]
ACKNOWLEDGEMENTS
We would like to thank members of the NorthwestGenome Engineering Consortium (NGEC) (httpngec-seattleorg) for their many insightful discussions
FUNDING
National Institutes of Health [RL1CA133832UL1DE019582 R01-HL075453 PL1-HL092557 RL1-HL092553 RL1-HL92554 and U19-AI96111] SeattleChildrenrsquos Center for Immunity and ImmunotherapiesFunding for open access charge NIH [U19-AI96111]
Conflict of interest statement AMS is an employee ofCellectis therapeutics and receives salary and equity com-pensation He is also founder member of the board of dir-ectors and equity holder in Pregenen Inc JJ is an employeeof Pregenen Inc and receives salary and equitycompensation AA is an employee of Pregenen Inc andreceives salary and equity compensation PD is an
employee of Cellectis therapeutics and receives salary andequity compensation AG is an employee of Cellectistherapeutics and receives salary and equity compensation
REFERENCES
1 BakerM (2012) Gene-editing nucleases Nat Methods 9 23ndash262 KimYG ChaJ and ChandrasegaranS (1996) Hybrid
restriction enzymes zinc finger fusions to Fok I cleavage domainProc Natl Acad Sci USA 93 1156ndash1160
3 KlugA (2010) The discovery of zinc fingers and theirapplications in gene regulation and genome manipulation AnnRev Biochem 79 213ndash231
4 ChristianM CermakT DoyleEL SchmidtC ZhangFHummelA BogdanoveAJ and VoytasDF (2010) TargetingDNA double-strand breaks with TAL effector nucleases Genetics186 757ndash761
5 LiT HuangS JiangWZ WrightD SpaldingMHWeeksDP and YangB (2011) TAL nucleases (TALNs) hybridproteins composed of TAL effectors and FokI DNA-cleavagedomain Nucleic Acids Res 39 359ndash372
6 PaquesF and DuchateauP (2007) Meganucleases and DNAdouble-strand break-induced recombination perspectives for genetherapy Curr Gene Ther 7 49ndash66
7 StoddardBL (2011) Homing endonucleases from microbialgenetic invaders to reagents for targeted DNA modificationStructure 19 7ndash15
8 JinekM ChylinskiK FonfaraI HauerM DoudnaJA andCharpentierE (2012) A programmable dual-RNAndashguided DNAendonuclease in adaptive bacterial immunity Science 337816ndash821
9 MaliP YangL EsveltKM AachJ GuellM DiCarloJENorvilleJE and ChurchGM (2013) RNA-guided humangenome engineering via Cas9 Science 339 823ndash826
10 McMahonMA RahdarM and PorteusM (2012) Gene editingnot just for translation anymore Nat Methods 9 28ndash31
11 LieberMR (2010) The mechanism of double-strand DNA breakrepair by the nonhomologous DNA end-joining pathway AnnRev Biochem 79 181ndash211
12 ShrivastavM DE HaroLP and NickoloffJA (2008)Regulation of DNA double-strand break repair pathway choiceCell Res 18 134ndash147
13 Method of the Year 2011 Nat Methods 9 114 MandellJG and BarbasCF (2006) Zinc finger tools custom
DNA-binding domains for transcription factors and nucleasesNucleic Acids Res 34 W516ndashW523
15 MaederML Thibodeau-BegannyS OsiakA WrightDAAnthonyRM EichtingerM JiangT FoleyJE WinfreyRJTownsendJA et al (2008) Rapid lsquoopen-sourcersquo engineering ofcustomized zinc-finger nucleases for highly efficient genemodification Mol Cell 31 294ndash301
16 SanderJD DahlborgEJ GoodwinMJ CadeL ZhangFCifuentesD CurtinSJ BlackburnJS Thibodeau-BegannySQiY et al (2011) Selection-free zinc-finger-nuclease engineeringby context-dependent assembly (CoDA) Nat Methods 8 67ndash69
17 CermakT DoyleEL ChristianM WangL ZhangYSchmidtC BallerJA SomiaNV BogdanoveAJ andVoytasDF (2011) Efficient design and assembly of customTALEN and other TAL effector-based constructs for DNAtargeting Nucleic Acids Res 39 e82ndashe82
18 ZhangF CongL LodatoS KosuriS ChurchGM andArlottaP (2011) Efficient construction of sequence-specific TALeffectors for modulating mammalian transcription NatBiotechnol 29 149ndash153
19 ReyonD TsaiSQ KhayterC FodenJA SanderJD andJoungJK (2012) FLASH assembly of TALENs for high-throughput genome editing Nat Biotechnol 30 460ndash465
20 JarjourJ West-FoyleH CertoMT HubertCG DoyleLGetzMM StoddardBL and ScharenbergAM (2009) High-resolution profiling of homing endonuclease binding and catalyticspecificity using yeast surface display Nucleic Acids Res 376871ndash6880
10 Nucleic Acids Research 2013
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
21 CornuTI Thibodeau-BegannyS GuhlE AlwinSEichtingerM JoungJK JoungJK and CathomenT (2008)DNA-binding specificity is a major determinant of the activityand toxicity of zinc-finger nucleases Mol Ther 16 352ndash358
22 GabrielR LombardoA ArensA MillerJC GenovesePKaeppelC NowrouziA BartholomaeCC WangJFriedmanG et al (2011) An unbiased genome-wide analysis ofzinc-finger nuclease specificity Nat Biotechnol 29 816ndash823
23 PattanayakV RamirezCL JoungJK and LiuDR (2011)Revealing off-target cleavage specificities of zinc-finger nucleasesby in vitro selection Nat Methods 8 765ndash770
24 SolluC ParsK CornuTI Thibodeau-BegannySMaederML JoungJK HeilbronnR and CathomenT (2010)Autonomous zinc-finger nuclease pairs for targeted chromosomaldeletion Nucleic Acids Res 38 8269ndash8276
25 HolkersM MaggioI LiuJ JanssenJM MiselliFMussolinoC RecchiaA CathomenT and GoncalvesMA(2012) Differential integrity of TALE nuclease genes followingadenoviral and lentiviral vector gene transfer into human cellsNucleic Acids Res 41 e63
26 FuY FodenJA KhayterC MaederML ReyonDJoungJK and SanderJD (2013) High-frequency off-targetmutagenesis induced by CRISPR-Cas nucleases in human cellsNat Biotechnol 31 822ndash826
27 CradickTJ FineEJ AnticoCJ and BaoG (2013) CRISPRCas9 systems targeting b-globin and CCR5 genes have substantialoff-target activity Nucleic Acids Res 41 9584ndash9592
28 RanFA HsuPD LinCY GootenbergJS KonermannSTrevinoAE ScottDA InoueA MatobaS ZhangY et al(2013) Double nicking by RNA-guided CRISPR Cas9 forenhanced genome editing specificity Cell 154 1380ndash1389
29 AshworthJ TaylorGK HavranekJJ QuadriSAStoddardBL and BakerD (2010) Computationalreprogramming of homing endonuclease specificity at multipleadjacent base pairs Nucleic Acids Res 38 5601ndash5608
30 TakeuchiR LambertAR MakAN JacobyK DicksonRJGloorGB ScharenbergAM EdgellDR and StoddardBL(2011) Tapping natural reservoirs of homing endonucleases fortargeted gene modification Proc Natl Acad Sci USA 10813077ndash13082
31 SatherBD RyuBY StirlingBV GaribovM KernsHMHumblet-BaronS AstrakhanA and RawlingsDJ (2011)Development of B-lineage predominant lentiviral vectors for usein genetic therapies for b cell disorders Mol Ther 19 515ndash525
32 CertoMT RyuBY AnnisJE GaribovM JarjourJRawlingsDJ and ScharenbergAM (2011) Tracking genomeengineering outcome at individual DNA breakpoints NatMethods 8 671ndash676
33 KuharR GwiazdaKS HumbertO MandtT PangalloJBraultM KhanI MaizelsN RawlingsDJ ScharenbergAMet al (2013) Novel fluorescent genome editing reporters formonitoring DNA repair pathway utilization at endonuclease-induced breaks Nucleic Acids Res (epub ahead of print October10 2013)
34 BochJ ScholzeH SchornackS LandgrafA HahnS KaySLahayeT NickstadtA and BonasU (2009) Breaking the codeof DNA binding specificity of TAL-type III effectors Science326 1509ndash1512
35 MoscouMJ and BogdanoveAJ (2009) A simple cipher governsDNA recognition by TAL effectors Science 326 1501ndash1501
36 GaoH WuX ChaiJ and HanZ (2012) Crystal structure of aTALE protein reveals an extended N-terminal DNA bindingregion Cell Res 22 1716ndash1720
37 MillerJC TanS QiaoG BarlowKA WangJ XiaDFMengX PaschonDE LeungE HinkleySJ et al (2011) ATALE nuclease architecture for efficient genome editing NatBiotechnol 29 143ndash148
38 TakeuchiR CertoM CapraraMG ScharenbergAM andStoddardBL (2009) Optimization of in vivo activity of abifunctional homing endonuclease and maturase reversesevolutionary degradation Nucleic Acids Res 37 877ndash890
39 ThymeSB JarjourJ TakeuchiR HavranekJJ AshworthJScharenbergAM StoddardBL and BakerD (2009)Exploitation of binding energy for catalysis and design Nature461 1300ndash1304
40 KentWJ (2002) BLATmdashthe BLAST-like alignment tool GenomeRes 12 656ndash664
41 TorikaiH ReikA LiuPQ ZhouY ZhangL MaitiSHulsH MillerJC KebriaeiP RabinovitchB et al (2012) Afoundation for universal T-cell based immunotherapy T cellsengineered to express a CD19-specific chimeric-antigen-receptorand eliminate expression of endogenous TCR Blood 1195697ndash5705
42 CertoMT GwiazdaKS KuharR SatherB CuringaGMandtT BraultM LambertAR BaxterSK JacobyK et al(2012) Coupling endonucleases with DNA end-processing enzymesto drive gene disruption Nat Methods 9 973ndash975
43 DelacoteF PerezC GuyotV DuhamelM RochonCOllivierN MacmasterR SilvaGH PaquesF DaboussiFet al (2013) High frequency targeted mutagenesis usingengineered endonucleases and DNA-end processing enzymesPLoS One 8 e53217
44 BaxterS LambertAR KuharR JarjourJ KulshinaNParmeggianiF DanaherP GanoJ BakerD StoddardBLet al (2012) Engineering domain fusion chimeras from I-OnuIfamily LAGLIDADG homing endonucleases Nucleic Acids Res40 7985ndash8000
Nucleic Acids Research 2013 11
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from

while above background off-target cleavage was limited toa single site (eg Figure 5d and Supplementary Figure 5)As the single off-target cleavage site identified is in anoncoding region of the genome this enzyme is of suffi-cient quality at its present state of development for clinicaluse However a further advantage of the megaTAL archi-tecture is the potential to further lsquotunersquo the mn cleavagehead to specifically avoid off-target cleavage sitesThus we anticipate that iterative refinement ofmegaTALs cleavage heads will allow the eventual identi-fication of megaTALs that couple high efficiency withperfect on target genomic specificityThe megaTAL architecture also possesses several
properties that facilitate efficient delivery for therapeuticapplications MegaTALs can be expressed with a single2 kb ORF smaller than two ZFN halves [22 kb total(11 kb per ZFN half targeting a 9-bp target)] and signifi-cantly shorter than a single TALEN half [gt50 kb total(25 kb per TALEN half targeting a 13-bp target)] orCas9 nuclease (41 kb not including guide RNA) ThusmegaTALs are compatible with efficient vectorization as aDNA mRNA or with any widely used viral vector systemThe capacity to use a TAL effector domain with as few as55 repeats also facilitates codon divergence of the TALeffector repeats to allow packaging into retroviral vectorswhere recombination among TAL effector RVDs duringreverse transcription can be problematic (25) Finally theindependent function of megaTALs due to their singlechain design avoids the possibility of inappropriateheterodimerization that may occur with enzymes basedon dimerization-dependent cleavage domains such asFokI an important advantage for applications requiringmultiplex genetic modifications
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Onlineincluding [44]
ACKNOWLEDGEMENTS
We would like to thank members of the NorthwestGenome Engineering Consortium (NGEC) (httpngec-seattleorg) for their many insightful discussions
FUNDING
National Institutes of Health [RL1CA133832UL1DE019582 R01-HL075453 PL1-HL092557 RL1-HL092553 RL1-HL92554 and U19-AI96111] SeattleChildrenrsquos Center for Immunity and ImmunotherapiesFunding for open access charge NIH [U19-AI96111]
Conflict of interest statement AMS is an employee ofCellectis therapeutics and receives salary and equity com-pensation He is also founder member of the board of dir-ectors and equity holder in Pregenen Inc JJ is an employeeof Pregenen Inc and receives salary and equitycompensation AA is an employee of Pregenen Inc andreceives salary and equity compensation PD is an
employee of Cellectis therapeutics and receives salary andequity compensation AG is an employee of Cellectistherapeutics and receives salary and equity compensation
REFERENCES
1 BakerM (2012) Gene-editing nucleases Nat Methods 9 23ndash262 KimYG ChaJ and ChandrasegaranS (1996) Hybrid
restriction enzymes zinc finger fusions to Fok I cleavage domainProc Natl Acad Sci USA 93 1156ndash1160
3 KlugA (2010) The discovery of zinc fingers and theirapplications in gene regulation and genome manipulation AnnRev Biochem 79 213ndash231
4 ChristianM CermakT DoyleEL SchmidtC ZhangFHummelA BogdanoveAJ and VoytasDF (2010) TargetingDNA double-strand breaks with TAL effector nucleases Genetics186 757ndash761
5 LiT HuangS JiangWZ WrightD SpaldingMHWeeksDP and YangB (2011) TAL nucleases (TALNs) hybridproteins composed of TAL effectors and FokI DNA-cleavagedomain Nucleic Acids Res 39 359ndash372
6 PaquesF and DuchateauP (2007) Meganucleases and DNAdouble-strand break-induced recombination perspectives for genetherapy Curr Gene Ther 7 49ndash66
7 StoddardBL (2011) Homing endonucleases from microbialgenetic invaders to reagents for targeted DNA modificationStructure 19 7ndash15
8 JinekM ChylinskiK FonfaraI HauerM DoudnaJA andCharpentierE (2012) A programmable dual-RNAndashguided DNAendonuclease in adaptive bacterial immunity Science 337816ndash821
9 MaliP YangL EsveltKM AachJ GuellM DiCarloJENorvilleJE and ChurchGM (2013) RNA-guided humangenome engineering via Cas9 Science 339 823ndash826
10 McMahonMA RahdarM and PorteusM (2012) Gene editingnot just for translation anymore Nat Methods 9 28ndash31
11 LieberMR (2010) The mechanism of double-strand DNA breakrepair by the nonhomologous DNA end-joining pathway AnnRev Biochem 79 181ndash211
12 ShrivastavM DE HaroLP and NickoloffJA (2008)Regulation of DNA double-strand break repair pathway choiceCell Res 18 134ndash147
13 Method of the Year 2011 Nat Methods 9 114 MandellJG and BarbasCF (2006) Zinc finger tools custom
DNA-binding domains for transcription factors and nucleasesNucleic Acids Res 34 W516ndashW523
15 MaederML Thibodeau-BegannyS OsiakA WrightDAAnthonyRM EichtingerM JiangT FoleyJE WinfreyRJTownsendJA et al (2008) Rapid lsquoopen-sourcersquo engineering ofcustomized zinc-finger nucleases for highly efficient genemodification Mol Cell 31 294ndash301
16 SanderJD DahlborgEJ GoodwinMJ CadeL ZhangFCifuentesD CurtinSJ BlackburnJS Thibodeau-BegannySQiY et al (2011) Selection-free zinc-finger-nuclease engineeringby context-dependent assembly (CoDA) Nat Methods 8 67ndash69
17 CermakT DoyleEL ChristianM WangL ZhangYSchmidtC BallerJA SomiaNV BogdanoveAJ andVoytasDF (2011) Efficient design and assembly of customTALEN and other TAL effector-based constructs for DNAtargeting Nucleic Acids Res 39 e82ndashe82
18 ZhangF CongL LodatoS KosuriS ChurchGM andArlottaP (2011) Efficient construction of sequence-specific TALeffectors for modulating mammalian transcription NatBiotechnol 29 149ndash153
19 ReyonD TsaiSQ KhayterC FodenJA SanderJD andJoungJK (2012) FLASH assembly of TALENs for high-throughput genome editing Nat Biotechnol 30 460ndash465
20 JarjourJ West-FoyleH CertoMT HubertCG DoyleLGetzMM StoddardBL and ScharenbergAM (2009) High-resolution profiling of homing endonuclease binding and catalyticspecificity using yeast surface display Nucleic Acids Res 376871ndash6880
10 Nucleic Acids Research 2013
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from
21 CornuTI Thibodeau-BegannyS GuhlE AlwinSEichtingerM JoungJK JoungJK and CathomenT (2008)DNA-binding specificity is a major determinant of the activityand toxicity of zinc-finger nucleases Mol Ther 16 352ndash358
22 GabrielR LombardoA ArensA MillerJC GenovesePKaeppelC NowrouziA BartholomaeCC WangJFriedmanG et al (2011) An unbiased genome-wide analysis ofzinc-finger nuclease specificity Nat Biotechnol 29 816ndash823
23 PattanayakV RamirezCL JoungJK and LiuDR (2011)Revealing off-target cleavage specificities of zinc-finger nucleasesby in vitro selection Nat Methods 8 765ndash770
24 SolluC ParsK CornuTI Thibodeau-BegannySMaederML JoungJK HeilbronnR and CathomenT (2010)Autonomous zinc-finger nuclease pairs for targeted chromosomaldeletion Nucleic Acids Res 38 8269ndash8276
25 HolkersM MaggioI LiuJ JanssenJM MiselliFMussolinoC RecchiaA CathomenT and GoncalvesMA(2012) Differential integrity of TALE nuclease genes followingadenoviral and lentiviral vector gene transfer into human cellsNucleic Acids Res 41 e63
26 FuY FodenJA KhayterC MaederML ReyonDJoungJK and SanderJD (2013) High-frequency off-targetmutagenesis induced by CRISPR-Cas nucleases in human cellsNat Biotechnol 31 822ndash826
27 CradickTJ FineEJ AnticoCJ and BaoG (2013) CRISPRCas9 systems targeting b-globin and CCR5 genes have substantialoff-target activity Nucleic Acids Res 41 9584ndash9592
28 RanFA HsuPD LinCY GootenbergJS KonermannSTrevinoAE ScottDA InoueA MatobaS ZhangY et al(2013) Double nicking by RNA-guided CRISPR Cas9 forenhanced genome editing specificity Cell 154 1380ndash1389
29 AshworthJ TaylorGK HavranekJJ QuadriSAStoddardBL and BakerD (2010) Computationalreprogramming of homing endonuclease specificity at multipleadjacent base pairs Nucleic Acids Res 38 5601ndash5608
30 TakeuchiR LambertAR MakAN JacobyK DicksonRJGloorGB ScharenbergAM EdgellDR and StoddardBL(2011) Tapping natural reservoirs of homing endonucleases fortargeted gene modification Proc Natl Acad Sci USA 10813077ndash13082
31 SatherBD RyuBY StirlingBV GaribovM KernsHMHumblet-BaronS AstrakhanA and RawlingsDJ (2011)Development of B-lineage predominant lentiviral vectors for usein genetic therapies for b cell disorders Mol Ther 19 515ndash525
32 CertoMT RyuBY AnnisJE GaribovM JarjourJRawlingsDJ and ScharenbergAM (2011) Tracking genomeengineering outcome at individual DNA breakpoints NatMethods 8 671ndash676
33 KuharR GwiazdaKS HumbertO MandtT PangalloJBraultM KhanI MaizelsN RawlingsDJ ScharenbergAMet al (2013) Novel fluorescent genome editing reporters formonitoring DNA repair pathway utilization at endonuclease-induced breaks Nucleic Acids Res (epub ahead of print October10 2013)
34 BochJ ScholzeH SchornackS LandgrafA HahnS KaySLahayeT NickstadtA and BonasU (2009) Breaking the codeof DNA binding specificity of TAL-type III effectors Science326 1509ndash1512
35 MoscouMJ and BogdanoveAJ (2009) A simple cipher governsDNA recognition by TAL effectors Science 326 1501ndash1501
36 GaoH WuX ChaiJ and HanZ (2012) Crystal structure of aTALE protein reveals an extended N-terminal DNA bindingregion Cell Res 22 1716ndash1720
37 MillerJC TanS QiaoG BarlowKA WangJ XiaDFMengX PaschonDE LeungE HinkleySJ et al (2011) ATALE nuclease architecture for efficient genome editing NatBiotechnol 29 143ndash148
38 TakeuchiR CertoM CapraraMG ScharenbergAM andStoddardBL (2009) Optimization of in vivo activity of abifunctional homing endonuclease and maturase reversesevolutionary degradation Nucleic Acids Res 37 877ndash890
39 ThymeSB JarjourJ TakeuchiR HavranekJJ AshworthJScharenbergAM StoddardBL and BakerD (2009)Exploitation of binding energy for catalysis and design Nature461 1300ndash1304
40 KentWJ (2002) BLATmdashthe BLAST-like alignment tool GenomeRes 12 656ndash664
41 TorikaiH ReikA LiuPQ ZhouY ZhangL MaitiSHulsH MillerJC KebriaeiP RabinovitchB et al (2012) Afoundation for universal T-cell based immunotherapy T cellsengineered to express a CD19-specific chimeric-antigen-receptorand eliminate expression of endogenous TCR Blood 1195697ndash5705
42 CertoMT GwiazdaKS KuharR SatherB CuringaGMandtT BraultM LambertAR BaxterSK JacobyK et al(2012) Coupling endonucleases with DNA end-processing enzymesto drive gene disruption Nat Methods 9 973ndash975
43 DelacoteF PerezC GuyotV DuhamelM RochonCOllivierN MacmasterR SilvaGH PaquesF DaboussiFet al (2013) High frequency targeted mutagenesis usingengineered endonucleases and DNA-end processing enzymesPLoS One 8 e53217
44 BaxterS LambertAR KuharR JarjourJ KulshinaNParmeggianiF DanaherP GanoJ BakerD StoddardBLet al (2012) Engineering domain fusion chimeras from I-OnuIfamily LAGLIDADG homing endonucleases Nucleic Acids Res40 7985ndash8000
Nucleic Acids Research 2013 11
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from

21 CornuTI Thibodeau-BegannyS GuhlE AlwinSEichtingerM JoungJK JoungJK and CathomenT (2008)DNA-binding specificity is a major determinant of the activityand toxicity of zinc-finger nucleases Mol Ther 16 352ndash358
22 GabrielR LombardoA ArensA MillerJC GenovesePKaeppelC NowrouziA BartholomaeCC WangJFriedmanG et al (2011) An unbiased genome-wide analysis ofzinc-finger nuclease specificity Nat Biotechnol 29 816ndash823
23 PattanayakV RamirezCL JoungJK and LiuDR (2011)Revealing off-target cleavage specificities of zinc-finger nucleasesby in vitro selection Nat Methods 8 765ndash770
24 SolluC ParsK CornuTI Thibodeau-BegannySMaederML JoungJK HeilbronnR and CathomenT (2010)Autonomous zinc-finger nuclease pairs for targeted chromosomaldeletion Nucleic Acids Res 38 8269ndash8276
25 HolkersM MaggioI LiuJ JanssenJM MiselliFMussolinoC RecchiaA CathomenT and GoncalvesMA(2012) Differential integrity of TALE nuclease genes followingadenoviral and lentiviral vector gene transfer into human cellsNucleic Acids Res 41 e63
26 FuY FodenJA KhayterC MaederML ReyonDJoungJK and SanderJD (2013) High-frequency off-targetmutagenesis induced by CRISPR-Cas nucleases in human cellsNat Biotechnol 31 822ndash826
27 CradickTJ FineEJ AnticoCJ and BaoG (2013) CRISPRCas9 systems targeting b-globin and CCR5 genes have substantialoff-target activity Nucleic Acids Res 41 9584ndash9592
28 RanFA HsuPD LinCY GootenbergJS KonermannSTrevinoAE ScottDA InoueA MatobaS ZhangY et al(2013) Double nicking by RNA-guided CRISPR Cas9 forenhanced genome editing specificity Cell 154 1380ndash1389
29 AshworthJ TaylorGK HavranekJJ QuadriSAStoddardBL and BakerD (2010) Computationalreprogramming of homing endonuclease specificity at multipleadjacent base pairs Nucleic Acids Res 38 5601ndash5608
30 TakeuchiR LambertAR MakAN JacobyK DicksonRJGloorGB ScharenbergAM EdgellDR and StoddardBL(2011) Tapping natural reservoirs of homing endonucleases fortargeted gene modification Proc Natl Acad Sci USA 10813077ndash13082
31 SatherBD RyuBY StirlingBV GaribovM KernsHMHumblet-BaronS AstrakhanA and RawlingsDJ (2011)Development of B-lineage predominant lentiviral vectors for usein genetic therapies for b cell disorders Mol Ther 19 515ndash525
32 CertoMT RyuBY AnnisJE GaribovM JarjourJRawlingsDJ and ScharenbergAM (2011) Tracking genomeengineering outcome at individual DNA breakpoints NatMethods 8 671ndash676
33 KuharR GwiazdaKS HumbertO MandtT PangalloJBraultM KhanI MaizelsN RawlingsDJ ScharenbergAMet al (2013) Novel fluorescent genome editing reporters formonitoring DNA repair pathway utilization at endonuclease-induced breaks Nucleic Acids Res (epub ahead of print October10 2013)
34 BochJ ScholzeH SchornackS LandgrafA HahnS KaySLahayeT NickstadtA and BonasU (2009) Breaking the codeof DNA binding specificity of TAL-type III effectors Science326 1509ndash1512
35 MoscouMJ and BogdanoveAJ (2009) A simple cipher governsDNA recognition by TAL effectors Science 326 1501ndash1501
36 GaoH WuX ChaiJ and HanZ (2012) Crystal structure of aTALE protein reveals an extended N-terminal DNA bindingregion Cell Res 22 1716ndash1720
37 MillerJC TanS QiaoG BarlowKA WangJ XiaDFMengX PaschonDE LeungE HinkleySJ et al (2011) ATALE nuclease architecture for efficient genome editing NatBiotechnol 29 143ndash148
38 TakeuchiR CertoM CapraraMG ScharenbergAM andStoddardBL (2009) Optimization of in vivo activity of abifunctional homing endonuclease and maturase reversesevolutionary degradation Nucleic Acids Res 37 877ndash890
39 ThymeSB JarjourJ TakeuchiR HavranekJJ AshworthJScharenbergAM StoddardBL and BakerD (2009)Exploitation of binding energy for catalysis and design Nature461 1300ndash1304
40 KentWJ (2002) BLATmdashthe BLAST-like alignment tool GenomeRes 12 656ndash664
41 TorikaiH ReikA LiuPQ ZhouY ZhangL MaitiSHulsH MillerJC KebriaeiP RabinovitchB et al (2012) Afoundation for universal T-cell based immunotherapy T cellsengineered to express a CD19-specific chimeric-antigen-receptorand eliminate expression of endogenous TCR Blood 1195697ndash5705
42 CertoMT GwiazdaKS KuharR SatherB CuringaGMandtT BraultM LambertAR BaxterSK JacobyK et al(2012) Coupling endonucleases with DNA end-processing enzymesto drive gene disruption Nat Methods 9 973ndash975
43 DelacoteF PerezC GuyotV DuhamelM RochonCOllivierN MacmasterR SilvaGH PaquesF DaboussiFet al (2013) High frequency targeted mutagenesis usingengineered endonucleases and DNA-end processing enzymesPLoS One 8 e53217
44 BaxterS LambertAR KuharR JarjourJ KulshinaNParmeggianiF DanaherP GanoJ BakerD StoddardBLet al (2012) Engineering domain fusion chimeras from I-OnuIfamily LAGLIDADG homing endonucleases Nucleic Acids Res40 7985ndash8000
Nucleic Acids Research 2013 11
by guest on March 5 2014
httpnaroxfordjournalsorgD
ownloaded from




















