Deep incision in an Aptian carbonate succession indicates major sea-level fall in the Cretaceous
Upload
independentCategory
view
1download
0
Journal of Neurochemistry Raven Press, New York 0 1983 International Society for Neurochemistry
oO22-3042/83/1001-957/$03.50/0
Studies on Subcellular Fractions Which Are Involved in Myelin Membrane Assembly: Isolation from Developing Mouse Brain and Characterization by Enzyme Markers,
Electron Microscopy, and Electrophoresis
Pedro M. Pereyra and Peter E. Braun
Department of Biochemistry, McGill University, Montre‘al, Que‘bec, Canada
Abstract: An extensive scheme for the subcellular frac- tionation of myelinating mouse brain is presented. Sev- eral centrifugation procedures for the separation of mem- branes involved in myelinogenesis are critically ap- praised, and guidelines for selection of centrifugation conditions are given. Characteristics of subcellular frac- tions are presented in the form of electron micrographs; also presented are distribution of RNA and protein; elec- trophoretic profiles of membrane proteins, and verifica- tion of the myelin-specific basic proteins, proteolipid pro- tein, and glycoprotein by the immuno-electroblot tech- nique; and the distribution of eight marker enzyme activities. Myelin-related membranes were found to differ both qualitatively and quantitatively in their complement of myelin-specific proteins. These myelin-containing
fractions appear to represent different stages of myeli- nation that coexist in developing mouse brain. These re- sults provide the fundamental methodologies and back- ground information for kinetic radioisotope analysis of intracellular events in the assembly of myelin presented in a companion article. Key Words: Myelin-Membrane assembly-Myelin proteins-2’,3’-Cyclic nucleotide 3’- phosphodiesterase-Golgi-Transferases-Myelin - asso- ciated glycoprotein. Pereyra P. M. and Braun P. E. Studies on subcellular fractions which are involved in myelin membrane assembly: Isolation from developing mouse brain and characterization by enzyme markers, electron microscopy, and electrophoresis. J . Neurochem. 41, 957-973 (1983).
Myelinogenesis in the developing CNS consti- tutes an experimental system that offers several major advantages for a study of eucaryotic cell membrane assembly. The oligodendroglial cell, like the PNS Schwann cell, produces vast quantities (up to 1 mm2 per cell) of plasma membrane (Norton, 1976), which undergoes molecular modification to become the lamellar myelin sheath. In mice the syn- thesis of this membrane occurs at an exponential rate for approximately a 2-week period, beginning about day 10. Moreover, the protein composition is relatively simple; the well-characterized myelin basic proteins (MBP) and the intrinsic protein lipophilin (proteolipid protein) each account for -30% of the total myelin protein (Braun and Brostoff, 1977). These and other favorable considerations suggested
to us the possibility of (1) defining the intracellular events in the process of oligodendroglial membrane assembly and (2) testing the validity of several as- pects of existing hypothetical assembly mecha- nisms (for reviews, see Rothman and Lenard, 1977; MorrC et al., 1979; Wickner, 1979; Waksman et al., 1980; Sabatini et al., 1982) as they pertain to my- elination.
In an earlier report we described a microsomal fraction (obtained as a “light” fraction by rate sed- imentation in a sucrose gradient) in which the oli- godendroglial-specific MBP accumulated very dif- ferently from that of all other membrane fractions during development (Barbarese et al., 1979). As a working hypothesis, it seemed reasonable that this fraction could contain precursor vesicles involved
Received October 12, 1982; accepted March 15, 1983. Address correspondence and reprint requests to Dr. Peter E.
Braun, Department of Biochemistry, McGill University, 3655 Drummond Street, McIntyre Medical Building Montreal, Quebec H3G 1Y6, Canada.
Abbreviations used: CNP, 2‘,3’-Cyclic nucleotide 3’-phospho-
diesterase; MAG, Myelin-associated glycoprotein; MBP, Myelin basic protein; PLP, Proteolipid protein; RSA, Relative specific activity; SDS-PAGE, Sodium dodecyl sulfate polyacrylamide gel electrophoresis; INT, p-Iodonitrotetrazolium violet; PAPS, Adenosine 3’-phosphate-5‘ phosphosulfate.
957

958 P . M . PEREYRA AND P . E . BRAUN
in the membrane assembly process, as has been de- scribed in preliminary reports (Braun and Pereyra, 1979; Braun et al., 1980a,b; Greenfield et ELL, 1983). Subsequent equilibrium-density centrifugation of this still heterogeneous fraction yielded several micro- soma1 subfractions, of which one appeared at a den- sity lighter than myelin (1.04 g/cc), consisted mostly of smooth vesicles, and contained all the proteins and enzymes typical of myelin. In further studies designed to show the relationship of these vesicles to myelin assembly, and to obtain other intracellular membranes from brains of developing animals, it became clear that the use of non-equilibrium sedi- mentation protocols resulted in myelin-clontaining fractions that include many types of ertdomem- branes, some of which could be associaited with myelination. This fractionation is complicated by the fact that during development the CNS contains axons at various stages of myelination. For ex- ample, at 15 days of age, the mouse brain has struc- tures that are just beginning to myelinate, while other regions have myelin varying from 1 to 5‘ days of apparent age.
The homogenization and fractionation procedure we have devised for our developmental studies pro- vides reproducible fractions of myelinated axons and endomembranes during myelination. In view of the above considerations, the morphological and bio- chemical characteristics, which we report here, lead us to suggest that the myelin-containing fractions are largely composed of myelinated axons and membranes-differing in both size and buoyant density-representing different stages of myelina- tion that coexist in developing brain. Additional support of this interpretation is provided by the metabolic studies described in a companion paper (Pereyra et al., 1983).
MATERIALS AND METHODS All radioisotopes and chemicals for scintillation counting
were purchased from New England Nuclear, Boston. The other biochemicals were products of Sigma Chemical, St. Louis. Rabbit anti-proteolipid protein (bovine) was a gen- erous gift from Drs. W. Macklin and M. Lees, Boston. Rabbit anti-MAG was generously provided by Dr. N. Sternberger, Rochester, NY. Rabbit anti-basic protein, raised against purified bovine 18.5 K MBP, was prepared by a standard procedure (Barbarese et al., 197’7). BEEM embedding capsules were purchased from J. B. EM Ser- vices Inc. Subcellular fractionation
Our fractionation scheme represents the extlended de- velopment of our earlier procedure (Barbarese el. al., 1979) and has been influenced by the detailed discussions of procedures for other tissues as given by DeF’ierre and Dallner (1976), Adelman et al. (1974), and Whittaker (1 969).
Typically, brains from 40 young mice of a specific age were rapidly (20 s ) removed and transferred to cold su- crose (0.25 M) and rinsed clean of adhering blood. A 30%
(wtlwt) homogenate was prepared in fresh 0.25 M sucrose with a glass homogenizer and Teflon pestle with a medium fit (clearance range 0.15 to 0.23 mm).
Ten up-and-down cycles at 800 rpm was a condition rigidly adhered to for all preparations; these and all other procedures were carried out in a cold-room at 4°C. The homogenate was diluted to 10% (wt/wt) with cold 0.25 M sucrose and centrifuged for 1.7 X lo3 g-min (5 min/2000 rpmitype 30) to remove broken cells and nuclear material. The supernatant (So) was then fractionated by differential velocity centrifugation in a type 30 rotor according to the scheme in Fig. 1. Briefly, centrifugation for 2.4 X lo4 g- min (17 min/4000 rpm) produced the pellet PI. Further centrifugation of S1 for 1.05 x lo6 g-min (20 min/27,000 rpm) yielded pellet Pz, with S2 being centrifuged for an- other 1.5 x 107 g-min (200 min/25,000 rpm) to produce pellet P3 and supernate S3.
Pellets P1 and P2 were each further fractionated during the time that S2 was in the centrifuge. They were rehom- ogenized in 0.25 M sucrose (20% vol/vol homogenate) by three up-and-down cycles at 800 rpm, and applied as a 16-1111 sample to a sucrose gradient consisting of 12 m10.8 M sucrose overlaid with 10 ml of 0.5 M sucrose. Rate zonal separations were achieved by centrifuging for 1.5 x lo7 g-min (180 min/25,000 rpm/SW 27). Figure 1 illus- trates the location of resultant fractions. The visualization of bands was aided by positioning a Tensor Minilamp di- rectly above the gradient tube. Fractions were collected with a 10-ml syringe fitted with a 10-cm blunted 19-gauge needle. Sucrose concentrations were determined by the refractive index measurements of each fraction.
The fractionation of pellet P3 differed somewhat from that of PI and P2. It was resuspended in 1 ml of 0.25 M sucrose by gentle agitation with a syringe needle as de- scribed above, followed by three cycles of gentle aspi- ration ending in the transfer to a single tube. The pooled material was diluted to a 10% (voVvo1) homogenate im- mediately prior to the next step.
Density equilibrium separations of particles were ob- tained as illustrated in Fig. 1. The resuspended material of fraction P3 and the bands recovered as P ~ A and P ~ A , were applied as a 14-1111 sample to a gradient consisting of 4 ml of 1.3 M sucrose overlaid with 10 ml of 0.8 M sucrose followed by 10 ml of 0.5 M sucrose. Similarly, 14-ml samples of PIB and PZB were applied to a tube con- taining 10 ml of 1.3 M sucrose overlaid with 14 ml of 0.8 M sucrose. All tubes were centrifuged for 1.44 x lo8 g- min (24 h/23,000 rpm/SW 27). Particle distribution was unchanged upon additional centrifugation and a linear gradient resulted from this operation.
Subfractions of PIC and P ~ c are complex and have not yet been completely characterized; they will be described in future communications. Preliminary studies show that they contain the bulk of the mitochondrial, synaptosomal, Golgi, and endoplasmic reticulum membranes.
All fractions were collected in preweighed vials and then weighed. Measurement of this weight and of the density of the sucrose solution containing the membra- nous particles, as determined by refractive index mea- surement, permits a rapid and reproducible calculation of the volume occupied by a given fraction. This parameter is used to calculate total enzyme activities and protein recovery.
All subfractions were diluted 1:l with cold water and were pelleted as follows: P 2 ~ and P3 subfractions required
J . Neurochem.. Vol. 41, No. 4, 1983

SUBCELLULAR FRACTIONS OF MYELINATING MOUSE BRAIN 959
F: 1.5 x
p2
1.5 x
4 107g- rnin "1 B
C
1.44 x 10' q-min
p3 1.44 x
I--J 10' g - rnin
60 min at 35,000 rpm in the Beckman Ti 75 rotor for complete pelleting; all other subfractions were centri- fuged under the same conditions for 35 min. This opera- tion was performed to separate membrane-bound and trapped proteins from soluble proteins.
Pellets were resuspended so that the final protein con- centration was 2-5 mgiml.
Calculation of sedimentation coefficients Calculation of sedimentation coefficient ranges for par-
ticles pelleted by velocity sedimentation as a function of equivalent radius (req) and the fraction of material left in the supernatant (4) were done by computer, using a pro- gram based on the equations of Rasmussen (1973).
For particles centrifuged in discontinuous sucrose gra- dients in swinging-bucket rotors, the minimum sedimen- tation coefficients at a given particle density (pp) required for a particle to traverse a given gradient step were cal- culated with a computer program based on the equations of McEwen (1967). These values were calculated as a function of the radius difference between the uppermost meniscus and the interface of each gradient, as well as a function of particle density. The sucrose concentration was assumed to be constant throughout each gradient step, a valid assumption for centrifugation times less than 3 h. Particles with densities equal to or less then the sucrose
FIG. 1. Subcellular fractionation p dure for developing mouse brain.
iroce-
density were assumed to be immobile or to move centri- petally.
Determination of protein We used a slightly modified version of the dye-binding
assay of Bradford (1976), with bovine y-globulin as stan- dard, and the BioradB protein assay reagent. Assays were performed with the standard procedure in the range of 10 to 140 pg of protein, or by a semimicro assay in the range of 1 to 14 pg. For the latter, samples were contained in a volume of 0.1 ml to which was added 1.5 ml of Biorad reagent. Standard curves were computed by linear regres- sion analysis.
Determination of RNA We employed the standard procedure of Fleck and
Munro (1962) for the isolation of RNA and its spectro- photometric assay. We used the equation CRNA,~ (pglml) = 11.87E2m - 10.4E3275 to calculate the amount of RNA recovered from each subfraction, assuming that the ab- sorbances of RNA and of polypeptides were additive. Values of RNA represent relative content amongst subfractions and cannot be assumed to be absolute values.
Electron microscopy Subcellular fractions were prepared for electron mi-
croscopy by suspending them in Karnovsky's glutaral-
J . Neurochem., Vol. 41, No. 4, 1983

960 P . M . PEREYRA AND P . E . BRAWN
dehyde fixative for 8 h. The suspended material was then pelleted in BEEM capsules by centrifugation for 60 min at 23,000 rpm in a SW 50.1 rotor fitted with special adapters. This produced a pellet 2-3 mm thick. The su- pernant was removed and the pellet gently rinsed three times with fresh fixative. The pellet was carefully recov- ered by slicing away the capsule. It was then transferred intact to a 3-ml screw-cap vial, and the samples were postfixed with OsO4-potassium ferrocyanide, stained en bloc with uranyl acetate, and dehydrated prior to embed- ding in Vestopal". Representative sections were cut from all regions of the tissue blocks and examined in a Philips 300 electron microscope.
Gel electrophoresis Proteins were separated by sodium dodecyl sulfate
polyacrylamide gel electrophoresis in slab gelis (1.5 mm x 10 cm x 15 cm) made with a linear gradient of 5-25% acrylamide, using the system of Laemmli (1970). Gels were aged overnight and prerun at 70 V for 1 h. Electro- phoresis was carried out at 80 V for 16 h, o r until the cytochrome c marker just reached the bottom of the gel. Gels were stained overnight with Coomassie blue (0.1%) in 25% isopropanol-10% acetic acid, and destained first in 10% isopropanol-10% acetic acid and then 10% acetic acid. They were stored in the hydrated state on a glass plate tightly wrapped in Saranwrap.
Strips of stained gels were scanned spectrophotomet- rically at 555 nm in a Gilford gel scanner.
Immunochemical identification of myelimi proteins An immunochemical assessment of the presence or ab-
sence of specific myelin proteins in subcellulaa fractions was made. Membrane proteins, separated by sodium do- decyl sulfate polyacrylamide gel electrophoresis (SDS- PAGE) as described above, were transferred electro- phoretically from the gels to nitrocellulose sheets (Towbin et al., 1979) and identified by the peroxidase-conjugated antibody technique (Barbarese and Pfeiffer, 198 1). Typi- cally, this procedure permitted an unequivocal identifi- cation on the gel of as little as 50 ng myelin basic proteins, 1 bg proteolipid protein (Macklin et al., 1982)., and 50 ng myelin-associated glycoprotein (Braun et al., 1982).
Enzyme markers Acetylthiocholinesterase. This activity was measured
by the procedure of Klingman et al. (1968). 2',3'-Cyclic nucleotide 3'-phosphohydrolase (CNP; EC
3.1.4.37). Activity was assayed by measuring the con- version of 2',3'-cyclic NADP to 2'-NADP (Sogin, 1976), modified by the addition to the assay mixture 01'2% Triton X-100 and sufficient NaCl to bring the final ionic strength to 0.75, as recommended by Sims et al. (1979)l.
NADH-Cytochrome c reductase. Activity was mea- sured both in the absence and in the presence of anti- mycin A, as recommended by Mahler (1955); the proce- dure of Huang et al. (1979) was used.
Succinate dehydrogenase (EC 1.3.99.1). Activity was measured as described by Huang et al. (1979).
PAPS-galactocerebroside sulfotransfe,rase (EC 2.8.2.11). The assay was an amalgamation of procedures described by Farrell and McKhann (1971) and Siegrist et al. (1977).
Volume used Stock solution in assay
Tris-HC1, pH 7.1, 250 mM ATP, 6.25 mM
NaCl, 385 mM PAP3%, 2.0 FCi/ml in 0.9% NaCl Galactocerebroside, 1 mg/ml in 2%
Enzyme (0.1-0.2 mg protein)
0.2 ml
MgClz, 50 mM
0.1 ml 0.1 ml
0.1 ml Triton X-100 and 0.9% NaCl
All ingredients, in screw-capped tubes, were brought to 37°C and the reaction was initiated by the addition of the PAP3%. The reaction was terminated by the addition of 5 ml chloroform-methanol (2:l) with mixing. A two- phase mixture was obtained by the addition of 1.2 ml 0.74% KCl, with vortexing and centrifuging at room tem- perature. The phases were separated and the lower phase was washed twice with theoretical upper phase (Folch et al., 1957). Samples were transferred to vials, dried, and then incubated at 50°C for 60 min with 0.4 ml Protosol solubilizer. Econofluor scintillation mix was added and [35S]sulfatide was determined by scintillation counting.
N-Acetyl glucosarninyltransferase utilizing endoge- nous acceptors. Activity was measured by the procedure of KO and Raghupathy (1972b). Each assay tube con- tained 0.3 pCi [3H]UDP-N-acetyl glucosamine. ATP (2 mM) was also added to protect the substrate against en- dogenous pyrophosphatase, as suggested by Bretz et al. (1980). The P3 fraction (500 pg total protein per assay) served as the acceptor source. This material was resus- pended in the assay buffer.
Galactosyltransferase utilizing endogenous acceptors. Activity was measured by the procedure of KO and Raghu- pathy ( 1 9 7 2 ~ ) . Each assay tube contained 0.08 FCi [14C]UDP-galactose. ATP (2 mM) and the P3 endogenous acceptor source were also added as for the preceding en- zyme.
RESULTS
Subcellular fractionation: Consideration of sedimentation parameters
Since our approach to the study of myelinogen- esis is to examine early events, it is imperative that intracellular membrane fractions that participate in this process be dissociated from myelin fragments, and that myelin elements be separated as exten- sively as possible into fractions that reflect struc- tural and compositional differences. Thus, we de- veloped a fractionation procedure (Fig. 1) for young myelinating animals based on both size and buoyant density characteristics of particles.
Although there are a number of methods entirely suitable for the isolation of mature myelin from adult animals, they are inadequate for isolating myelin fractions representing various stages of develop- ment at a given moment during myelinogenesis. Thus, a detailed assessment of how particles sepa- rate under several centrifugational procedures seemed appropriate in view of the many different
J . Neurochem.. Val. 41. No. 4 , 1983

SUBCELLULAR FRACTIONS OF MYELINATING MOUSE BRAIN 961
1.10
4 P 1.08
protocols used in research on myelinogenesis and the misinterpretation of results from procedures that fail to separate membranes on the basis of defined sedimentation parameters, but rather produce frac- tions of mixed characteristics. This has been a se- rious problem, making meaningful comparison of results among the various reports in the literature impossible.
In preliminary experiments we determined the overlap of particle size that can be expected for a given set of velocity sedimentation conditions (data not shown), since the success of subsequent sepa- rations in discontinuous gradients under nonequi- librium conditions depends on the degree of size overlap of a given fraction. In our first gradient (Fig. 1) small particles originating from plasma mem- branes, endoplasmic reticulum, Golgi elements, etc., were retained in the upper two or three layers of the gradient, even though their density is higher than that of sucrose. Water shock followed by velocity centrifugational collection of particles is no im- provement, since small particles still remain inter- mixed with other membranes of interest. A specific example of the inadequacy of nonequilibrium sedi- mentation procedures to separate complex mixtures is the presence of large quantities of ribosomes and ribonucleoprotein particles, which fill the perikarya of oligodendroglia during active myelination (Peters et al., 1976). We have observed that these small free ribosomal elements (S < 80) are present in most fractions, especially those that contain myelin ob- tained by rate zonal centrifugation on discontinuous sucrose gradients. They can be removed by centri- fuging these fractions to their equilibrium densities, or they can be maintained in an aggregated state with the aid of low-salt buffers containing 2-4 mM Mg2+ as recommended for the isolation of endo- plasmic reticulum and other intracellular mem- branes (AdeIman et al., 1974).
In the case of very large myelin or myelinated axon fragments (Sw,20 = 104-105) the careful choice of a sedimentation rate can render isopycnic cen- trifugation unnecessary. For example, the small free ribosomal contamination (4% on a RNA basis) of PIA can be eliminated by simple rate sedimentation under appropriate conditions. However, the isola- tion of smaller myelin and membrane elements (S,,~O = lo2-lo4) requires centrifugation to density equi- librium in order to achieve adequate separations, i.e., subfractionation.
Figure 2 is an attempt to obtain a summary per- spective of these subfraction characteristics in terms of a two-dimensional representation of their par- ticle-size range and particle density. This has broad biochemical implications in the context of size and density differences that occur amongst organelles or membranes of a given type in the developing brain. Myelin itself undergoes considerable change
Pz AY - %BY -
- G A S - P 2 e a PI B a
Minimum particle size pelleted after a given time (min)
Rotor Ti 75: 35Krpm
-
1.04 - &L L 1
in morphology and in its relationship to the axon during development (see Discussion).
Morphological assessment by electron microscopy Four distinct myelin and myelinated axon frac-
tions, PIAa, P ~ A ~ , P I B ~ , P ~ B ~ (Table I), were consis- tently obtained from 15-day-old mice (Fig. 3). Most of the axonal fragments in the light subfractions ( P I A ~ , P Z A ~ ) contain aggregated filaments, identifi- able axolemma, and compact myelin. The axonal fragments in the denser subfractions ( P I B ~ , P ~ B ~ ) contain nonaggregated filaments and well-pre- served mitochondria in the axoplasm, and the myelin has fewer lamellae.
The heterogenous subfractions, P l ~ p and P~BP, also contain myelinated axons with well-defined and nonaggregated axoplasmic filaments and, in many cases, axoplasmic mitochondria. These axonal ele- ments frequently have cytoplasm entrapped mes- axonally and in intramyelinic pockets. Many large vesicles and membrane fragments are also present, but fewer lamellar myelin fragments are observed.
The microsomal fraction P3a1 contains all the typ- ical myelin proteins, but its density (1.043) is lower than that of myelin-containing fractions. It appears transiently during early stages of myelinogenesis and has a characteristic vesiculated electron micro- scopic morphology (Fig. 4a).
Another interesting microsomal fraction, P3a, also contains typical myelin proteins, but the SDS-PAGE profiles are nevertheless quantitatively and quali- tatively different from those of P3a, (as presented
J . Neurochem., Vol. 41, N o . 4, 1983
962 P . M . PEREYRA AND P . E. BRAUN
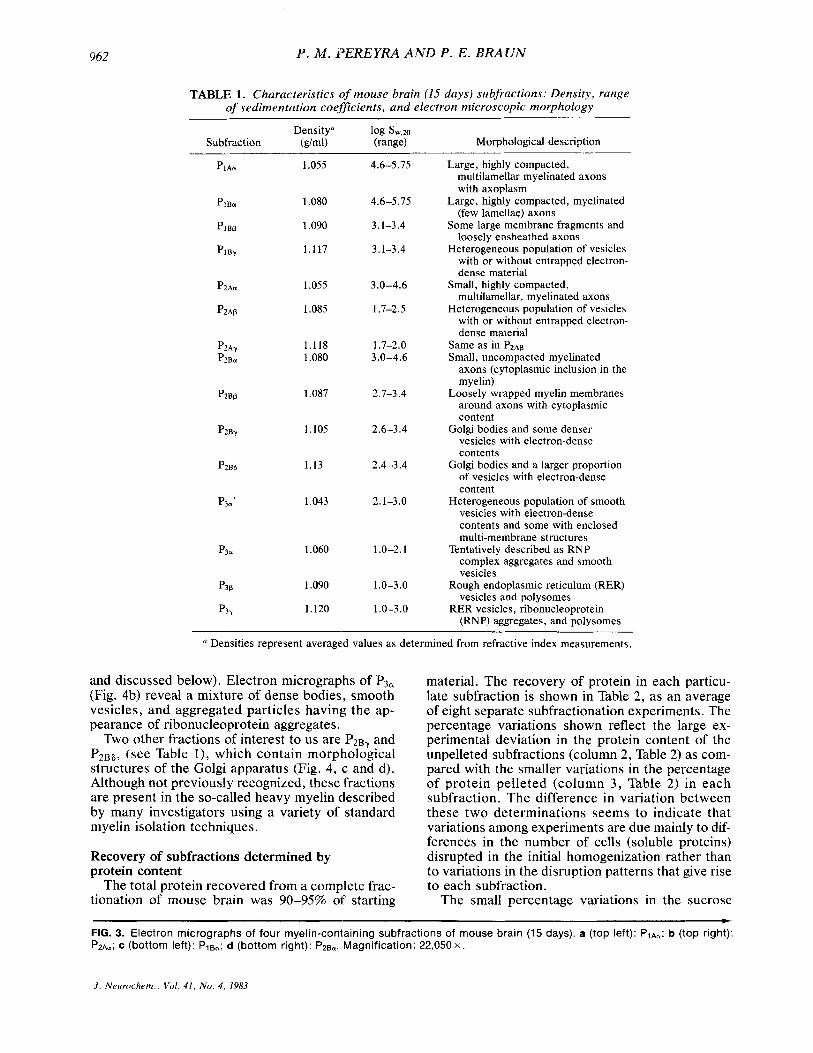
TABLE 1. Characteristics ,of mouse brain (15 days) subfractions: Density, range of sedimentation coeficients, and electron microscopic morphology
Densitya 1% sw,2a Subfraction (dml) (range) Morphological description
1.055
1.080
1.090
1.117
1.055
1.085
1.118 1.080
1.087
1.105
1.13
1.043
1.060
1.090
1.120
4.6-5.75
4.6-5.75
3.1-3.4
3.1-3.4
3.0-4.6
1.7-2.5
1.7-2.0 3.0-4.6
2.7-3.4
2.6-3.4
2.4-3.4
2.1-3.0
1 .o-2.1
1.0-3.0
1 .O-3 .O
Large, highly compacted, multilamellar myelinated axons with axoplasm
Large, highly compacted, myelinated (few lamellae) axons
Some large membrane fragments and loosely ensheathed axons
Heterogeneous population of vesicles with or without entrapped electron- dense material
Small, highly compacted, multilamellar, myelinated axons
Heterogeneous population of vesicles with or without entrapped electron- dense material
Same as in P2Ap Small, uncompacted myelinated
axons (cytoplasmic inclusion in the myelin)
Loosely wrapped myelin membranes around axons with cytoplasmic content
Golgi bodies and some denser vesicles with electron-dense contents
Golgi bodies and a larger proportion of vesicles with electron-dense content
Heterogeneous population of smooth vesicles with electron-dense contents and some with enclosed multi-membrane structures
Tentatively described as RNP complex aggregates and smooth vesicles
Rough endoplasmic reticulum (RER) vesicles and polysomes
RER vesicles, ribonucleoprotein (RNP) aggregates, and polysomes
a Densities represent averaged values as determined from refractive index measurements.
and discussed below). Electron micrographs of P3a (Fig. 4b) reveal a mixture of dense bodies, smooth vesicles, and aggregated particles having the ap- pearance of ribonucleoprotein aggregates.
Two other fractions of interest to us are P2By and P 2 B 8 , (see Table I ) , which contain morplhological structures of the Golgi apparatus (Fig. 4, c and d). Although not previously recognized, these fractions are present in the so-called heavy myelin described by many investigators using a variety of standard myelin isolation techniques.
Recovery of subfractions determined by protein content
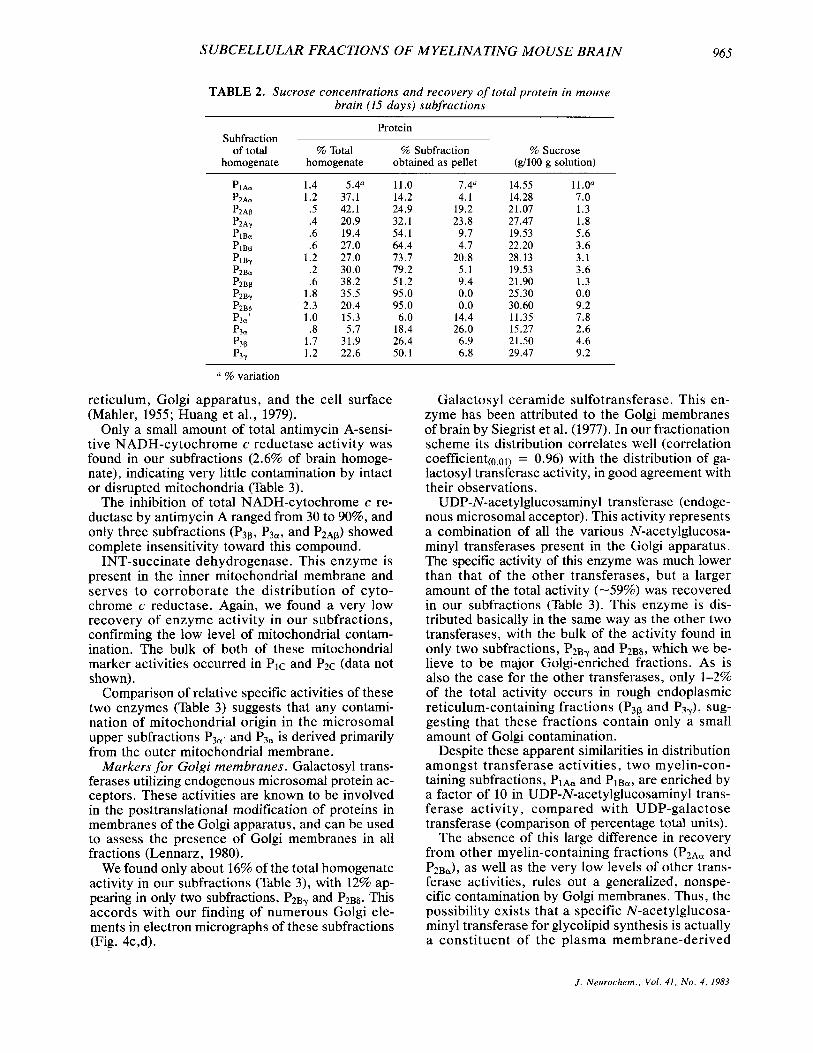
The total protein recovered from a complete frac- tionation of mouse brain was 90-95% of starting
material. The recovery of protein in each particu- late subfraction is shown in Table 2, as an average of eight separate subfractionation experiments. The percentage variations shown reflect the large ex- perimental deviation in the protein content of the unpelleted subfractions (column 2, Table 2) as com- pared with the smaller variations in the percentage of protein pelleted (column 3 , Table 2) in each subfraction. The difference in variation between these two determinations seems to indicate that variations among experiments are due mainly to dif- ferences in the number of cells (soluble proteins) disrupted in the initial homogenization rather than to variations in the disruption patterns that give rise to each subfraction.
The small percentage variations in the sucrose b
FIG. 3. Electron micrographs of four myelin-containing subfractions of mouse brain (15 days). a (top left): PIAcr; b (top right): P z A ~ ; c (bottom left): Pie,; d (bottom right): P2Bu. Magnification: 2 2 , 0 5 0 ~ .
J . Neurochem., Vol . 41, No. 4, 1983

SUBCELL ULAR FRACTIONS OF MYELINATING MOUSE BRAIN 963
J . Neiirochem., Vol. 41, No. 4, 1983

964 P. M . PEREYRA AND P . E. BRAUN
FIG. 4. Electron micrographs of four microsomal subfractions of mouse brain (15 days). a (top left): P3-,; b (top right): P3-; c (bottom left): P z B ~ ; d (bottom right): P2~6. Magnification: 3 5 , 7 7 0 ~ .
concentration of each subfraction (colurnm 3, Table 2) suggest that it is possible to achieve considerable uniformity and reproducibility in determining boundaries of bands and in collecting them.
Enzyme markers and the distribution of RNA Our choice of enzyme markers was based not only
on the need to identify subfractions, but also in- cluded the objective of establishing some of their metabolic interrelationships. The distribution of en- zyme activities is summarized in Table 3.
Mitochondria1 markers. Electron micrographs of
some of our subfractions which contain my elinated axons ( P I A ~ , P ~ A ~ , PIBa , P ~ B J reveal the presence of a few axoplasmic mitochondria. In addition, small vesiculated inner and outer mitochondrial mem- brane fragments can be expected to co-isolate with microsomal subfractions. We assessed this by mea- suring the activity of two enzymes:
NADH-cytochrome c reductase. The antimycin A-sensitive form of this enzyme from brain has been reported to be localized in the outer mitochondrial membrane, and the insensitive form appears to be distributed among membranes of the endoplasmic
J . Neurochem., Vol. 41, N o . 4, 1983
SUBCELLULAR FRACTIONS OF MYELINATING MOUSE BRAIN 965
TABLE 2. Sucrose concentrations and recovery of total protein in mouse brain (15 days) subfractions
Protein Subfraction
of total % Total % Subfraction % Sucrose homogenate homogenate obtained as pellet (g/lOO g solution)
PIAa 1.4 5.4" 11.0 7.4" 14.55 11.0a P2Aa 1.2 37.1 14.2 4.1 14.28 7.0 P2Ap .5 42.1 24.9 19.2 21.07 1.3 P2Ay .4 20.9 32.1 23.8 27.47 1.8 PlBu .6 19.4 54.1 9.7 19.53 5.6 PlBP .6 27.0 64.4 4.7 22.20 3.6 PIBy 1.2 27.0 73.7 20.8 28.13 3.1 P2Ba .2 30.0 79.2 5 .1 19.53 3.6 P2BP .6 38.2 51.2 9.4 21.90 1.3
P2B6 2.3 20.4 95.0 0.0 30.60 9.2 P3=' 1.0 15.3 6.0 14.4 11.35 7.8 p3a .8 5.7 18.4 26.0 15.27 2.6
1.7 31.9 26.4 6.9 21.50 4.6 P3Y 1.2 22.6 50.1 6.8 29.47 9.2
P2By 1.8 35.5 95.0 0.0 25.30 0.0
P3P
% variation
reticulum, Golgi apparatus, and the cell surface (Mahler, 1955; Huang et al., 1979).
Only a small amount of total antimycin A-sensi- tive NADH-cytochrome c reductase activity was found in our subfractions (2.6% of brain homoge- nate), indicating very little contamination by intact or disrupted mitochondria (Table 3).
The inhibition of total NADH-cytochrome c re- ductase by antimycin A ranged from 30 to 90%, and only three subfractions (P3p, P3a, and P2Ap) showed complete insensitivity toward this compound.
INT-succinate dehydrogenase. This enzyme is present in the inner mitochondrial membrane and serves to corroborate the distribution of cyto- chrome c reductase. Again, we found a very low recovery of enzyme activity in our subfractions, confirming the low level of mitochondrial contam- ination. The bulk of both of these mitochondrial marker activities occurred in PlC and P2c (data not shown).
Comparison of relative specific activities of these two enzymes (Table 3) suggests that any contami- nation of mitochondrial origin in the microsomal upper subfractions P3af and P301 is derived primarily from the outer mitochondrial membrane.
Markers for Golgi membranes. Galactosyl trans- ferases utilizing endogenous microsomal protein ac- ceptors. These activities are known to be involved in the posttranslational modification of proteins in membranes of the Golgi apparatus, and can be used to assess the presence of Golgi membranes in all fractions (Lennarz, 1980).
We found only about 16% of the total homogenate activity in our subfractions (Table 3), with 12% ap- pearing in only two subfractions, P2By and P2B& This accords with our finding of numerous Golgi ele- ments in electron micrographs of these subfractions (Fig. 4c,d).
Galactosyl ceramide sulfotransferase. This en- zyme has been attributed to the Golgi membranes of brain by Siegrist et al. (1977). In our fractionation scheme its distribution correlates well (correlation coefficient(o.01) = 0.96) with the distribution of ga- lactosyl transferase activity, in good agreement with their observations.
UDP-N-acetylglucosaminyl transferase (endoge- nous microsomal acceptor). This activity represents a combination of all the various N-acetylglucosa- minyl transferases present in the Golgi apparatus. The specific activity of this enzyme was much lower than that of the other transferases, but a larger amount of the total activity (-59%) was recovered in our subfractions (Table 3). This enzyme is dis- tributed basically in the same way as the other two transferases, with the bulk of the activity found in only two subfractions, P ~ B ? and P2B6, which we be- lieve to be major Golgi-enriched fractions. As is also the case for the other transferases, only 1-2% of the total activity occurs in rough endoplasmic reticulum-containing fractions (P3p and P3?), sug- gesting that these fractions contain only a small amount of Golgi contamination.
Despite these apparent similarities in distribution amongst transferase activities, two myelin-con- taining subfractions, P ~ A ~ and P I ~ a , are enriched by a factor of 10 in UDP-N-acetylglucosaminyl trans- ferase activity, compared with UDP-galactose transferase (comparison of percentage total units).
The absence of this large difference in recovery from other myelin-containing fractions ( P 2 ~ a and P2Ba), as well as the very low levels of other trans- ferase activities, rules out a generalized, nonspe- cific contamination by Golgi membranes. Thus, the possibility exists that a specific N-acetylglucosa- minyl transferase for glycolipid synthesis is actually a constituent of the plasma membrane-derived
J . Neurochem., Vol. 41, N o . 4, 1983

966 P . M . PEREYRA AND P . E. BRAUN
TABLE 3 . Characterization ~ f m o u s e brain (15 days) subfractions by their content of marker enzyme activity and of RNA
Mitochondria1 + Succinate-INT UDP-NAGluA UDP-Gal. NADH Cyt. c Red. reductase transferase transferase
Subfraction RSA % T U RSA % T U RSA % T U RSA % T U
PIAa p2Aa 1.4 PZAP 0.0 p2Ay .3 PlBu PlBB PlBy P2Bu .2 p2Bp .3 P2By . I
p3u .7 P3P 0.0 P3Y 0.0
H Ribosomal total pellets
P2BS P3a’ 3.0
,1 0.0
.2
. I
.2 1.3
.2
.2 0.0 0.0
.8
.3
.04 0.0
. 1
.3 0.0 0.0
. I
. 1
.2
. I
.2
.2
.03
.01
.04
.01 0.0
.02
.2 0.0 0.0
.04
. I
.4 0.0
. 1
.2 0.0
1.3 1 .o .2 .1 2.4 .4 .2 .1
.5 . I .5 . I 1.2 .2 .8 . I 5.4 1.9 .9 . 3 2.6 .8 1 . 1 .3 2.4 1.2 1.1 .5 4.0 .5 1.9 .2 2.3 .6 2.7 .7
27.8 15.6 5.0 2.8 21.7 35.3 5.8 9.5
2.0 . 1 .8 . 1 0.0 0.0 .8 .04 2.6 .9 .8 .3
.5 . 3 1.2 .7
2.3 1.2 58.7 15.9
structure in PIAa and PIBu. In fact, the methodology of the assay does not preclude the possibility that we have observed here the incorporation of isotope into oligosaccharides and gangliosides. In this re- gard, some gangliosides are known to contain N- acetylglucosamine instead of N-acetylgalactosa- mine (Lennarz, 1980). Furthermore, we also cannot exclude the possibility of artifact due to nonspecific binding to proteins and lipids of isotopic contami- nants that might be present in the commercially la- beled substrate.
Acetylthiocholinesterase. This activity actually represents two enzymes: acetylcholinesterase, found mainly in neuronal surface membranes, and pseu- docholinesterase, which appears to be localized mainly in glial membranes and microsornal mem- branes in general. We were able to recovier a max- imum of 28% of total acetylthiocholinesterase ac- tivity in our subfractions (Table 3); the remainder of the activity appeared in fractions not shown here. The highest relative specific activities were located in the p or y subfractions, with the exception of P ~ A ~ and P3ac. This is what we expected, given the possible appearance of dense, synaptosome-de- rived contaminants in the p and y regions of the gradient. In fact, these specific activities dlo parallel the density distribution one expects of synlaptosom- ally derived elements containing acetylcholines- terase activity. The high relative specific activity value for the light fraction P2Au is puzzling, and we offer no immediate explanation, except to indicate that the total units of this activity are very low. In P3a, we cannot rule out some contamination by syn- aptic vesicles.
RNA determination. In the absence of salt buffers we expected a substantial quantity of free poly- somes, ribosomes, and RNA in some of our subfractions, especially since glial cells and their processes in the developing brain are filled with these structures. PIA was found to contain up to 2% of the total RNA, consistent with the visual aggre- gates of free ribosomes interspersed with the large myelin fragments seen in electron micrographs. Centrifugational pelleting of these membranes by velocity sedimentation or separation by equilibrium density centrifugation removed about 90% of the RNA, congruent with the visual disappearance of ribosomal particles.
Fractionation of microsomal material by the pro- cedure of Barbarese et al. (1979) resulted in the ap- pearance of most of the RNA as ribonucleoprotein aggregates and ribosomes in the uppermost layers (P3A and P3B) of the step gradient (Table 4, A), frac- tions which contain MBPs and other myelin con- stituents during development. Centrifugation of particles in P3 to their equilibrium density produced four distinct bands of membranous material and a pellet containing ribosomes (Table 4, B). The small amount of RNA in the uppermost myelin-related subfraction (P3,,) represents free RNA (Sw,20 = 50) rather than ribosomes. Note that there has been a redistribution of free and membrane-associated RNA from the “light” myelin-related subfractions P3A and P3B into the rough endoplasmic reticulum-enriched subfractions P3, and P3y, by subjecting the same initial discontinuous gradient to isopycnic condi- tions. Table 3 lists the content of RNA in subfrac- tions obtained by our current procedure (Fig. 1).
J . Neurochem., Vol. 41, N o . 4, 1983

SUBCELLULAR FRACTIONS OF MYELfNATING MOUSE BRAIN
TABLE &Continued
PAP3% Acetyl-thio transferase cholinesterase CNP RNA
Subfraction RSA % T U RSA % T U RSA % T U %
CLg Total
. 1 . I
.7 . I
.03 0.0
.8 .I
.7 .2 1.3 .4 1.5 .7 2.4 .3 4.6 1.2
11.8 6.6 9.0 14.6
.3 .02
.3 .01 1.2 .4 2.4 1.5
3.5 .6 8.5 .2 9.6 .3 6.9 1.5
2.3 2.6
1.9 .4 6.8 1.7 2.8 10.8
21.1 .7 2.8 .3
10.2 4.9 3.9 4.3
15.9 2.5 17.4 3 .1 9.3 2.8 3.3 2.0
14.6 5.1 13.8 1.8 4.7 2.3
18.7 2.2 8.5 2.2 2.8 1.6 1.2 2.0
14.3 1 .o 1.3 . 1 4.6 1.6 1.2 .8
4.9 .2 7.4 .2 1.4 .04 4.0 .1 6.1 .2 4.1 . 1 2.1 .06 3.8 . 1 5.4 .2
20.1 .6
9.0 .3 64.4 2.0
226.8 7.0 146.5 4.5
z 26.3 28.3 31.2
Ribosomal total pellets 635.3 19.5
RSA relative specific activity (with respect to homogenate). % TU is % total units of enzyme activity.
Thus, we can identify P3p and P3? as subfractions containing rough endoplasmic reticulum, and show that only small amounts of RNA are associated with other lighter subfractions.
Myelin markers Our objective was not only to show the distri-
bution of typical myelin components amongst the various subfractions, but also to determine the way in which these fractions could be grouped according to common characteristics. In this way we hope to achieve a more comprehensive understanding of metabolically interrelated parameters in myelin as- sembly.
2 I , 3 -Cyclic nu cl e o tide 3 ' -p h osp ho h y dro las e (CNP). Although the function of this enzyme re- mains an enigma, its structural association with the myelin sheath and oligodendroglial plasma mem- brane is firmly established (Braun and Brostoff, 1977). However, its presence in endomembranes of myelinating cells, as well as in membranes of non- neural tissues, has also been reported (Dreiling et al., 1981; Weissbarth et al., 1981) and suggests a broader function than heretofore recognized.
Table 3 shows that only 31% of total units are recovered in our subfractions; the remainder of the activity was found in the postmicrosomal superna- tant and in PIC and P2c. The enzymatic activity is distributed very similarly amongst most of the subfractions (average of 2.1 * 0.3%), except in PZAa, PZAp, and P I B ~ , in which it varies from 2.8 to 5.1%, or in some microsomal subfractions, in which ac- tivity accounts for 0.6-1% of total units.
967
Distribution of CNP on the basis of relative spe- cific activity, however, is quite different. Subfrac- tions which contain typical morphological charac- teristics of myelin (PIAa, PIBp, P2Aor, PI&, P2Bu) are enriched 13- to 18.7-fold over total homogenate. Subfractions P ~ A ~ and P2Bp have relative specific activities (RSA) about 9, and a third category (PIBY, P ~ A ? , P ~ B ~ , P2B6) is enriched 1.2- to 5-fold over the homogenate. The microsomally derived subfrac- tions (P3uf, P3u, P3@, P3?) also fall into the above three categories, according to their RSA of CNP.
TABLE 4. Comparison oJ'RNA content among microsomal fractions of mouse brain (15 days) obtained
by discontinuous gradient velocity sedimentation (A) and equilibrium density sedimentation (B)
Subfraction CLg R"4
A. 3 h/25,000 rpm/SW 27 p3 top P3A p3B P3BiC p3c P3cm P3D
Z = 880.
B. 24 h/25,000 rprnlSW 27 P3=' p3a P3P P3Y P3 pellet
H = 876.
32.40 406.60 365.60
15.60 25.40
1.20 34.00
8.90 64.40
226.80 146.50 430.00
J . Nrurochem., Vol. 41, N o . 4, 1983
968
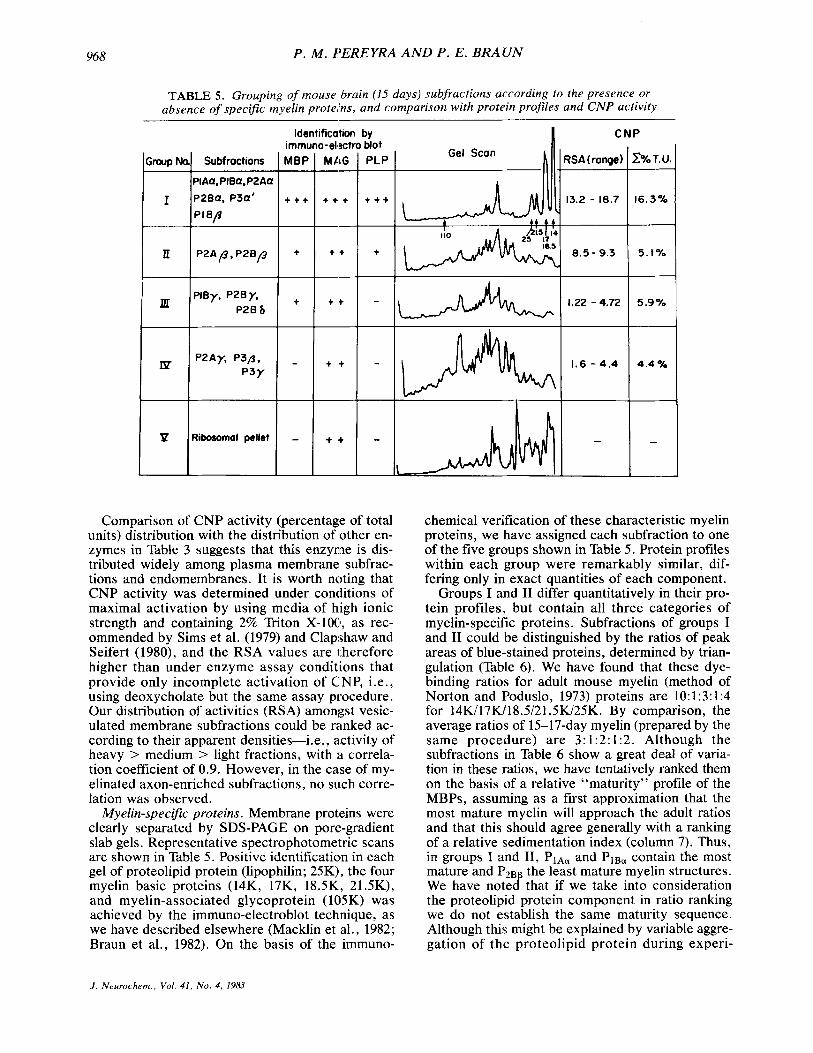
G m p k
1
II
P . M . I’EREYRA A N D P . E . BRAUN
Subfractions MBP MAG - PIAQ. PIBQ. P2AQ
P2BQ. P3Q’ + + + + + + PI B p
P2Af ,P2Bp + + +
-
-
TABLE 5 . Grouping of mouse brain (15 days) subfractions according fo the presence or absence of specific myelin protehs, and comparison with protein profiles and CNP activity
Identification bv tiot PLP -
t + +
+
Comparison of CNP activity (percentage of total units) distribution with the distribution of other en- zymes in Table 3 suggests that this enzyme is dis- tributed widely among plasma membrane subfrac- tions and endomembranes. It is worth noting that CNP activity was determined under conditions of maximal activation by using media of high ionic strength and containing 2% Triton X-100, as rec- ommended by Sims et al. (1979) and Clapshaw and Seifert (1980), and the RSA values are therefore higher than under enzyme assay conditions that provide only incomplete activation of CNP, i.e., using deoxycholate but the same assay procedure. Our distribution of activities (RSA) amongst vesic- ulated membrane subfractions could be ranked ac- cording to their apparent densities-i.e., activity of heavy > medium > light fractions, with i i correla- tion coefficient of 0.9. However, in the case of my- elinated axon-enriched subfractions, no such corre- lation was observed.
Myelin-specific proteins. Membrane protieins were clearly separated by SDS-PAGE on poregradient slab gels. Representative spectrophotometric scans are shown in Table 5. Positive identification in each gel of proteolipid protein (lipophilin; 25K), the four myelin basic proteins (14K, 17K, 18.5K, 21.5K), and myelin-associated glycoprotein (1015K) was achieved by the immuno-electroblot technique, as we have described elsewhere (Macklin et al., 1982; Braun et al., 1982). On the basis of the immuno-
I CNP
?SA (range)
13.2 - 18.7
8.5 - 9.3
1.22 -4.72
1.6 - 4.4
-
?Yo T. U.
16.3%
5 . I %
5.9%
4.4 ?!a
-
chemical verification of these characteristic myelin proteins, we have assigned each subfraction to one of the five groups shown in Table 5. Protein profiles within each group were remarkably similar, dif- fering only in exact quantities of each component.
Groups I and I1 differ quantitatively in their pro- tein profiles, but contain all three categories of myelin-specific proteins. Subfractions of groups I and I1 could be distinguished by the ratios of peak areas of blue-stained proteins, determined by trian- gulation (Table 6). We have found that these dye- binding ratios for adult mouse myelin (method of Norton and Poduslo, 1973) proteins are 10: 1 :3: 1 :4 for 14K/17K/18.5/21.5K/25K. By comparison, the average ratios of 15-17-day myelin (prepared by the same procedure) are 3:1:2:1:2. Although the subfractions in Table 6 show a great deal of varia- tion in these ratios, we have tentatively ranked them on the basis of a relative “maturity” profile of the MBPs, assuming as a first approximation that the most mature myelin will approach the adult ratios and that this should agree generally with a ranking of a relative sedimentation index (column 7). Thus, in groups I and 11, P ~ A ~ and PIBa contain the most mature and PZB the least mature myelin structures. We have notecf that if we take into consideration the proteolipid protein component in ratio ranking we do not establish the same maturity sequence. Although this might be explained by variable aggre- gation of the proteolipid protein during experi-
J . Neurochem.. Vol. 41, No. 4, 1983

SUBCELL ULAR FRACTIONS OF M YELINATING MOUSE BRAIN 969
TABLE 6 . Ranking of Group Z and ZZ subfractions according to their enrichment in 14K MBP and 18.5K MBP and comparison with their particle index (+)
Subfraction 14K MBP 17K MBP 18.5K MBP 21.5K MBP PLP 4)
PlAcl 6 2 3 1 3 5.28 1 3 1 2.5 5.28 plea 4
p2Aa 3 1 2 1 1 3.94 P2Ba 3 1 2 1 1 3.94 PlBP 3 1 2 1 2 3.43 p2Bp 1 2 1 1 3.5 3.25 P3e’ 2 1 2 1 2 2.75
mental manipulations, our metabolic studies (Per- eyra et al., 1983) suggest that a more likely reason resides in the relative proportions of proteolipid protein to MBPs message expressed at different stages of development.
Group I11 is characterized by the absence of de- tectable proteolipid protein, the presence of (rela- tively) very small amounts of basic proteins, and a moderate amount of myelin-associated glycopro- tein. Group IV subfractions have no detectable pro- teolipid protein or basic proteins, but have an amount of myelin-associated glycoprotein compa- rable to that in authentic myelin-containing subfrac- tions. Group V is the ribosomal pellet, which has no detectable myelin proteins except for several components of about 50K and 70K, which react with the antiserum to myelin-associated glycoprotein. At this point we cannot be certain if these proteins are degradative fragments of myelin-associated glyco- protein, its incompletely synthesized precursors, or artifacts.
At this point, such a ranking scheme is valid only to the degree that a specific component can be clearly identified. Comparisons of the content of any specific component are analogous to compari- sons of relative specific activities in the case of en- zymes.
DISCUSSION An integral aspect of our approach to the cellular
mechanisms of myelinogenesis was to develop a methodology that would allow us to distinguish the various endo- and exomembrane fractions involved in this process. The methodology we describe here is not necessarily the best to achieve purity of any given fraction, but rather is a compromise permit- ting the acquisition of the greatest number of iden- tifiable myelin-related fractions without regard to purity. Thus, we sought to achieve a gentle, repro- ducible homogenization and fractionation protocol for developing mouse brain, which would result in fractions that retain as many internodal axonal seg- ments, organelles, and endomembranes as possible in a simple procedure. For this reason we avoided the more extreme conditions of hypoosmotic shock, which are usually used to obtain the more heavily
lamellated myelin elements from adult brain, free of other endomembrane and axoplasmic compo- nents.
We have paid careful attention to the physical principles that govern separation of particles by ve- locity and buoyant density sedimentation in order to assign to each of our subfractions operational parameters, which we use as independent variables in the correlation of quantitative and qualitative dif- ferences among subfractions. These parameters are (a) the average buoyant density, determined by re- fractive index measurement of the sucrose gradient; (b) the calculated range of sedimentation rates, ex- pressed as the logarithm of the Svedberg unit (log SW,2o; see Materials and Methods); and (c) the vector sum of these parameters, which we express as a Particle Index. In general, our experience shows that although the buoyant density is a good param- eter for distinguishing various membrane types, such as plasma membrane, Golgi membranes, and en- doplasmic reticulum, the sedimentation index or the Particle Index is better suited to describe more com- plex elements, such as myelinated axons.
Having taken into consideration the limitations and pitfalls of this approach with respect to marker distribution, we have arrived at a subfractionation scheme that yields physically and biochemically de- fined membrane fractions from myelinating brain. These can be grouped on the basis of their specific myelin protein content (Table 5 ) as determined by the immuno-electroblot technique.
We found that the four groups of membrane-con- taining subfractions (Table 5) fell into the following general categories: group I, myelinated axons and associated elements; group 11, amorphous plasma membranes; group 111, Golgi membranes; Group IV, endoplasmic reticulum (rough and smooth). In making these assignments, we were careful not to overinterpret data from marker enzyme determi- nations; thus, some overlap exists.
Subfractions in groups I and I1 can be ranked according to their enrichment in the 14K and 18.5K MBPs (Table 6); this ranking correlates closely with the Particle Index (+). We currently interpret this correlation as showing that the developmental char- acteristics of the myelin-axon unit are related more
J . Neurochem., Vol. 41, No. 4, 1983

970 P . M . PEREYRA AND P . E. BRAUN
TABLE 7 . Correlation matrixa of marker enzyme distribution among subfractions
UDP-NAG UDP-NAG UDP-Gal Sulfo Sulfo AcTChE Trans (RSA) Trans (TU) Trans (RSA) Trans (RSA) Trans (TU) W A )
UDP-NAG Trans (RSA) 1 .oo .85 .90 .94 .84 - .22 UDP-NAG naris (TU) 1 .oo .88 .I9 .9Y - .25 UDP-Gal Trans (RSA) 1 .oo 9 6 .91 - .30 Sulfo Trans (RSA) 1 .oo .82 - .31 Sulfo Trans (TU) 1 .oo - .28 AcTChE (RSA) 1 .oo AcTChE (TU) CNP (RSA) CNP (TU) RNA
(96) UDP-Gal Trans (TU)
closely to the observed size distribution of myeli- nated axon fragments than to their buoyant densi- ties.
Under specific conditions of homogenization, the separation of cellular organelles and nuclei by sed- imentation techniques has been well documented. The literature in the area of myelin research shows that it is widely assumed that the relationship of myelin to the axon is completely disrupteld by ho- mogenization, and that a continuous range of par- ticle sizes is generated, rather than several popu- lations of discrete size ranges. However, even in mature brain, the disruption of myelinated axons will depend on the stability of the myelin-myelin and myelin-axon interactions, given a constant shear force. From this argument, it follows that one may expect to obtain at least two discrete size popula- tions, given an appropriate sedimentation protocol. One of these will be composed of a mixture of small particles, which arise from large, structuirally un- stable myelinated segments, or from small structur- ally stable or unstable myelinated internodes. The other population of particles will contain large par- ticles, whose initial structural stability allows them to survive as relatively large myelin membrane ag- gregates or even as myelinated axon segments. Fur- thermore, in the case of water-shocked1 myelin subfractions, the buoyancy of the myeliin mem- branes will depend on the degree of integrity of as- sociation among the disrupted myelin lamellae. On the other hand, in the case of non-water-shocked myelinated axon fragments, their structural integ-
continued
rity and size will determine their overall entrapped axoplasmic volume, and the extent of myelin la- mellation will determine their impermeability to the surrounding sucrose medium, conditions which in turn determine the buoyancy of the particle (Sarma and Sitaraman, 1982). Our own electron micro- scopic analysis of myelin-containing fractions sup- ports this interpretation.
For the moment, we are led to suggest that the size contribution to sedimentation behavior of my- elinated axonal segments is more closely related to the extent of myelin maturity than is the buoyant density contribution. Further investigation is needed to obtain data supporting (or contradicting) this novel interpretation.
The microsomal subfraction P3a,, which we have also assigned to Group I because of its shared prop- erties with other subfractions in this category, ac- tually consists of two vesicle populations: one con- sisting of large, trilamellar structures containing myelin proteins in the same relative proportions as P ~ A ~ and P ~ B ~ , the other consisting of smaller, uni- lamellar vesicles, which may or may not contain electron-dense material and have myelin protein proportions intermediate between those of P2Bp and P ~ B ~ . These vesicular subfractions appear only during the most active phase of myelination.
The subfractions in group I11 represent only about one-third of all the Golgi-associated membrane ele- ments; most of these are found in PIC and P2c. How- ever, they are characterized by a relatively high en- richment in three transferases which are known to
J . Neurochem., Vol. 41, N o . 4 , 1983

SUBCELLULAR FRACTIONS OF MYELINATING MOUSE BRAIN 971
TABLE 7.--Continued
UDP-NAG Trans (RSA) .85 - .32 .01 - .24 - . 1 1 .16
Trans (TU) .87 - .40 - .03 - .21 - .07 .98 UDP-Gal Trans (RSA) .78 - .42 - .12 - .22 - .09 .85 Sulfo Bans (RSA) .85 - .42 -.13 - .21 - .03 .73 Sulfo Trans (TU) .91 - .45 - .08 -.16 .02 .99 AcTChE ( R W - .22 .16 - .02 .13 .04 - .27 AcTChE (TU) 1 .oo - .51 - .22 .06 .34 .91 CNP ( R W 1.00 .51 .41 .49 - .41 CNP (TU) 1 .oo - .57 .40 - .05 RNA ((*.g/mg) 1 .oo .85 - .18 RNA (%I 1 .oo .02 UDP-Gal Trans (TU) 1 .oo a Stepwise regression analysis with the Regression Analysis Pac of the HP-85 computer (tolerance = 0.01).
UDP-NAG
be Golgi markers. All three enzymes show a high degree of correlation among these subfractions in both their total unit and specific activity distribu- tions (Table 7). We believe that this argues for a common subcellular origin of these membrane ele- ments, even though they originate from several cell types. From the high correlation between acetyl- cholinesterase activity and the transferase activi- ties, in terms of their total activity distribution, we further conclude that group I11 subfractions contain a significant amount of neuronal or synaptosomal membrane. The presence of myelin-associated gly- coprotein (MAG) in these subfractions can be ex- plained on the basis that the Golgi apparatus con- stitutes the logical processing site for this glycopro- tein on its path to myelin; but we cannot exclude the possibility of its presence in small amounts of oligodendroglial plasma membrane that remain as- sociated with the neuronal membrane. Discrimina- tion between these possibilities must await more highly quantitative measurements of MAG.
Group IV subfractions represent only a very small fraction of the endoplasmic reticulum, most of which is found as large elements in the fractions Plc and P2c. The RNA in this group is mainly of the free type, with good correlation of specific enrichment and total unit distribution.
These subfractions also contain two antigenic components, of molecular weights -50,000 and -70,000, which react with our antiserum to MAG. We do not yet know if these represent MAG pre- cursors or degradation fragments.
Since CNP is widely used as a marker for myelin, we carried out an extensive survey of this enzyme and found activity in all of our membranous subfractions, with variable behavior toward acti- vating conditions and variable ratios of soluble to membrane-bound forms of this enzyme. Other in- vestigators in the myelin field have also noted this, but have not dealt with the broad implications of the general distribution of CNP; instead, the ten- dency has been to assume that this indicates that subcellular fractions are contaminated by myelin. More recently, several reports have appeared showing the presence of CNP activity in tissues other than brain, including muscle, spleen, thymus, and liver. The enzyme is enriched in the nuclear and microsomal fractions of liver, but is also found in other liver membranes (Weissbarth et al., 1981). Dreiling et al. (1981) have studied mitochondrial- associated CNP activity, and Muller et al. (1981) have reported the immunochemical detection of CNP in cytoplasm of a neuronal cell line.
Although the use of “marker” enzyme distribu- tion, as conceived by deDuve et al. (1964), has been fundamental to establishing enrichment of fractions in certain structures, recent studies by Howell et al. (1978), Borgese and Meldolsi (1980), Meldolsi et al. (1980), and Borgese et al. (1980) show that deDuve’s concept is not strictly applicable under all circumstances. They point out that varying cryp- ticity of a given enzyme, its variable sensitivity to inhibition, sensitivity to oxidizing conditions, and multifocal insertion of the protein in membrane as-
J . Neurochem., Vol. 41, N o . 4, 1983

9 72 P . M . PEREYRA AND P . E. BRAUN
sembly processes make it difficult to assign that en- zyme to a specific membranous compartment and may explain apparently ambiguous data with re- spect to its precise localization. A case in point is NADH cytochrome c reductase, long accepted to be a marker of the endoplasmic reticulum, but found in many subcellular fractions enriched in other membranous elements. For many of the reasons cited above, this enzyme is now known to occur in different forms in the Golgi apparatus and plasma membrane-enriched fractions. Our own studies (Table 3 ) show that this enzyme in the developing mouse brain is also found in a variety of mem- branes, in addition to the endoplasmic reticulum. No defined correlation was found between the en- richment of the enzyme (RSA) and the distribution of its activity (% TU) (see Table 7).
In view of the above considerations, we now hold CNP to be distributed in a multifocal fashion, and are further investigating this problem, as well as the role that this enzyme might play in the regulatory machinery of oligodendroglia.
The following observations should be empha- sized:
1. Proteolipid protein is localized to only a re- stricted group of membranes or structures which possess a typical spectrum of myelin proteins; these membranes range in density from 1.04 to II .08 gfml. Absence of detectable amounts of this protein from Golgi-enriched fractions is a potentially important observation, given that the MAG is clearly present, as well as low levels of MBPs. We are attempting to provide independent corroboration of these im- munochemical data, which are reproducible and ap- parently reliable.
2. MBPs are present in three membrane catego- ries, but appear most prominently in only thle myelin- containing compartments. Even though immuno- chemical staining for these antigens on nitrocellu- lose electroblots of the gels is not a strictly quantitative procedure, a relative assessment of amount of protein is, nevertheless, possible. In- deed, examination of the relevant gel scans in Table 5 offers a visual confirmation of diminished MBP bands in the protein profile.
3 . The distribution of MAG is a more general phenomenon, encompassing all membrane frac- tions, as well as the ribosome-containing pellet.
4. Quantitative differences in protein profiles (gel scans) are observed within each major group of subfractions, and qualitative differences are evident when comparing the major groups with each other.
5. CNP, when maximally activated, 11s widely distributed among brain subfractions. We interpret this as multifocal distribution, and not its myelin contamination.
Although more highly definitive structural and functional descriptions of myelin-related mem- branes will only be derived from purified subfrac-
tions, the approach we describe here, based on size and buoyant density considerations, provides a complex overview of protein and enzyme distribu- tions and their interrelationships, with respect to an early stage of myelination. We wish to emphasize that our present, as well as future, experimental designs are not-will not-be constrained by the details of the subfractionation protocol presented here, but rather will employ the strategy embodied in it. A combination of fractionation refinement techniques, protein isolation, immunochemical pro- cedures, and studies of metabolic dynamics by iso- tope methodologies should provide us with deeper insights into the biogenic sequence of events in which these membranes are involved. A companion paper (Pereyra et al., 1983) addresses the kinetic isotope approach.
Acknowledgments: We are grateful to Elsa Horvath and all members of this laboratory for their patience, cheerfulness, and technical assistance. We also greatly appreciate the expert secretarial assistance of Diana Ia- senza. This work was supported by the Medical Research Council of Canada, and by a fellowship of the Multiple Sclerosis Society of Canada to P.M.P.
REFERENCES Adelman M. R., Blobel G., and Sabatini D. D. (1974) Nonde-
structive separation of rat liver rough microsomes into ri- bosomal and membranous compartments, in Methods in En- zymology, Vol. 31 (Fleisher s. and Packer L., eds), pp. 201- 225. Academic Press, New York.
Barbarese E. and Pfeiffer S. E. (1981) Developmental regulation of basic protein in dispersed cultures. Proc. Natl. Acad. Sci.
Barbarese E. , Braun P. E., and Carson J. H. (1977) Identifica- tion of prelarge and presmall basic proteins in mouse myelin and their structural relationship to large and small basic pro- tein. Proc. Natl. Acad. Sci. USA 74, 3360-3364.
Barbarese E., Carson J. H. , and Braun P. E. (1979) Subcellular distribution and structural polymorphism of myelin basic protein in normal and Jimpy mouse brain. J . Neurochem. 32, 1437-1446.
Borgese N. and Meldolesi J. (1980) Localization and biosynthesis of NADH-cytochrome bs reductase, an integral membrane protein, in rat liver cells. I. Distribution of the enzyme ac- tivity in microsomes, mitochondria and Golgi complex. J . Cell Biol. 85, 501-515.
Borgese N., Pietrini G., and Meldolesi J. (1980) Localization and biosynthesis of NADH-cytochrome bs reductase, an integral membrane protein, in rat liver cells. 111. Evidence for the independent insertion and turnover of the enzyme in various subcellular compartments. .I. Cell Biol. 86, 38-45.
Bradford M. (1976) A rapid sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248.
Braun P. E. and Brostoff S. (1977) Proteins of Myelin, in Myelin (Morel1 P., ed), pp. 201-231. Plenum Press, New York.
Braun P. E. and Pereyra P. M. (1979) Characteristics of myelin precursor vesicles. Trans. Am. Soc. Neurochem. 10, 215.
Braun P. E., Pereyra P. M., and Greenfield S. (1980~) Mecha- nisms of assembly of myelin in mice: A new approach to the problem, in INSERM Symposium No . 14 (Braumann N., ed), pp. 413-421. Elsevier, North Holland.
Braun P. E., Pereyra P. M., and Greenfield S. (1980b) Myelin organization and development: A biochemical perspective,
USA 78, 1953-1957.
.I. Neurochem., Vol. 41, No. 4, 1983

SUBCELL ULAR FRACTIONS OF MYELINATING MOUSE BRAIN 973
in Myelin: Chemistry and Biology, pp. 1-17. Alan R. Liss, New York.
Braun P. E., Frail D. E., and Latov N. (1982) Myelin-associated glycoprotein is the antigen for a monoclonal IgM in poly- neuropathy. J. Neurochem. 39, 1261-1265.
Bretz R., Bretz H., and Palade G. E. (1980) Distribution of ter- minal glycosyltransferase in hepatic Golgi fractions. J . Cell B i d . 84, 87-101.
Clapshaw P. A. and Seifert W. (1980) Effects of detergents, pro- teins, and lipids on 2‘:3‘-cyclic-nucleotide 3’-phosphodies- terase activity. J. Neurochem. 35, 164-169.
deDuve C. (1964) Principles of tissue fractionation. J. Theor. Biol. 6, 33-59.
DePierre J. and Dallner G. (1976) Isolation, subfractionation and characterization of the endoplasmic reticulum, in Biochem- ical Analysis of Membranes (Maddy A. H., ed), pp. 79-131. John Wiley, New York.
Dreiling C. E., Schilling R. J., and Reitz R. C. (1981) 2’,3’-Cyclic nucleotide 3’-phosphohydrolase in rat liver mitochondria1 membranes. Biochim. Biophys. Acta 640, 114-120.
Farrell D. F. and McKhann G. M. (1971) Characterization on cerebroside sulfotransferase from rat brain. J . Biol. Chem.
Fleck A. and Munro H. N. (1962) The precision of UV absorp- tion measurements in the Schmidt-Thannhauser procedure for nucleic acid estimation. Biochim. Biophys. Acta 55,
Folch J., Lees M., and Stanley G. H. S. (1957) A simple method for the isolation and purification of total lipids from animal tissues. J . Biol. Chem. 226, 497-509.
Greenfield S., Braun P. E., Gantt G., and Hogan E. L. (1983) Routes of assembly of myelin proteins, in Comparative Pa- thobiology (Chang T., ed), Plenum Press (in press).
Howell K. E., Ito A., and Palade G. E. (1978) Endoplasmic re- ticulum marker enzymes in Golgi fractions-What does it mean? J . Cell Biol. 79, 581-589.
Huang C. M., Goldenberg H., Frantz C., MorrC D. J., Keenan T. W., and Crane F. L. (1979) Comparison of NADH-linked cytochrome c reductases of endoplasmic reticulum, Golgi apparatus and plasma membrane. Int. J. Biochem. 10,
Klingman G. J., Klingman J. D., and Poliszczuk A. (1%8) Acetyl- and pseudocholinesterase activities in sympathetic ganglia of rats. J. Neurochem. 15, 1121-1130.
KO G. K. W. and Raghupathy E. (1972~) Microsomal galacto- syltransferase utilizing endogenous and exogenous protein acceptors. Biochim. Biophys. Acta 244, 396-409.
KO G. K. W. and Raghupathy E. (1972b) Microsomal galacto- saminyltransferase utilizing endogenous and exogenous pro- tein acceptors. Biochim. Biophys. Acta 264, 129-143.
Laemmli U. K. (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227,
Lennarz W. J. (1980) The Biochemistry of Glycoproteins and Proteoglycans, pp. 1-373. Plenum Press, New York.
Macklin W. B., Braun P. E., and Lees M. B. (1982) Electroblot analysis of the myelin proteolipid protein. J. Neurosci. Res. 7, 1-10.
Mahler H. R. (1955) DPNH cytochrome c reductase, in Methods in Enzymology, Vol. 2 (Colowick S. P. and Kaplan N. O., eds), pp. 688-693. Academic Press, New York.
McEwen C. R. (1967) Tables for estimating sedimentation through linear concentration gradients of sucrose solutions. Anal. Biochem. 20, 114-149.
Meier P. J., Spycher M. A., and Meyer U. A. (1981) Isolation and characterization of rough endoplasmic reticulum asso-
246, 4694-4702.
571-583.
723 -73 1.
680-685.
ciated with mitochondria from normal rat liver. Biochim. Biophys. Acta 646, 283-297.
Meldolesi J., Corte G., Pietrini G., and Borgese N. (1980) Lo- calization and biosynthesis of NADH-cytochrome bs reduc- tase, an integral membrane protein, in rat liver cells. 11. Evidence that a single enzyme accounts for the activity in its various subcellular locations. J . Cell Biol. 85, 516-526.
Morrt D. J., Kartenbeck J., and Franke W. W. (1979) Membrane flow and interconversions among endomembranes. Biochim. Biophys. Acta 559, 71-152.
Miiller H. W., Clapshaw P. A., and Seifert W. (1981) Intracellular localization of 2’,3’-cyclic nucleotide 3’-phosphodiesterase in a neuronal cell line as examined by immunofluorescence and cell fractionation. J . Neurochem. 37, 947-955.
Norton W. T. (1976) Formation, structure and biochemistry of myelin, in Basic Neurochemistry, 2nd ed., pp. 74-99. Little, Brown and Co., Boston.
Norton W. T. and Poduslo S. E. (1973) Myelination in the rat brain: Method of myelin isolation. J. Neurochem. 21, 749.
Pereyra P. M., Braun P. E., Greenfield S., and Hogan E. L. (1983) Studies on subcellular fractions which are involved in myelin assembly: Labeling of myelin proteins by a double radioiso- tope approach indicates developmental relationships. J . Neurochem. 41, 974-988.
Peters A., Palay S. L., and Webster H. deF. (1976) The Fine Structure of the Nervous System: Neurons and Supporting Cells. pp. 1-406, Saunders, Philadelphia.
Rasmussen H. N. (1973) Nomogram for calculation of centrif- ugal data. Eur. J . Biochem. 34, 502-505.
Rothman J. E. and Lenard J. (1977) Membrane asymmetry. Sci- ence 195, 743-753.
Sabatini D. D., Kreibich G., Morimoto T., and Adesnik M. (1982) Mechanisms for the incorporation of proteins in membranes and organelles. J. Cell B i d . 92, 1-22.
Sarma M. K. J. and Sitaraman V. (1982) Myelosomes: The os- motically sensitive myelin particles. Biochem. Biophys. Res. Commun. 105, 362-369.
Siegrist H. P., Jutzi H., Steck A. J., Burkart T., Wiesmann U., and Herschkowitz N. (1977) Age-dependent modulation of 3’-phosphoadenosine 5’-phosphosulfate-galactosylceramide sulfotransferase by lipids extracted from the microsomal membranes. Biochim. Biophys. Acta 489, 58-63.
Sims N. R., Horvath L. B., and Carnegie P. R. (1979) Detergent activation and solubilization of 2‘,3’-cyclic nucleotide 3‘- phosphodiesterase from isolated myelin and Cg cells. Biochem. J . 181, 367-375.
Sogin D. D. (1976) 2’,3’-Cyclic NADP as a substrate for 2’,3’- cyclic nucleotide 3’-phosphohydrolase. J. Neurochem. 27,
Towbin H., Staehlin T., and Gordon J. (1979) Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellu- lose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA 76, 4350-4354.
Waksman A., Hubert P., Creme1 G., Rendon A., and Burmn C. (1980) Translocation of proteins through biological mem- branes. A critical review. Biochim. Biophys. Acta 604, 249- 296.
Weissbarth S., Maker H. S., Raes I., Brannan T. S., Lapin E. P., and Lehrer G. M. (1981) The activity of 2‘,3‘-cyclic nucleo- tide 3’-phosphodiesterase in rat tissues. J . Neurochem. 37,
Whittaker V. P. (1969) The synaptosome, in Handbook of Neu- rochemistry, Vol. 2 (Lajtha A., ed), pp. 327-364. Plenum Press, New York.
Wickner W. (1979) The assembly of proteins into biological mem- branes: The membrane trigger hypothesis. Annu. Rev . Biochem. 48, 23-45.
1333-1337.
677-680.
J . Neurochem., Vol. 41, N o . 4, 1983
Copyright © 2022 FDOKUMEN