Cohorts of arbuscular mycorrhizal fungi (AMF) in Vitis vinifera, a typical Mediterranean fruit crop

1 23
Genetic Resources and CropEvolutionAn International Journal ISSN 0925-9864 Genet Resour Crop EvolDOI 10.1007/s10722-014-0169-3
Chloroplast diversity indicates twoindependent maternal lineages incultivated grapevine (Vitis vinifera L.subsp. vinifera)
Rita Lózsa, Ning Xia, Tamás Deák &György Dénes Bisztray

1 23
Your article is protected by copyright and all
rights are held exclusively by Springer Science
+Business Media Dordrecht. This e-offprint
is for personal use only and shall not be self-
archived in electronic repositories. If you wish
to self-archive your article, please use the
accepted manuscript version for posting on
your own website. You may further deposit
the accepted manuscript version in any
repository, provided it is only made publicly
available 12 months after official publication
or later and provided acknowledgement is
given to the original source of publication
and a link is inserted to the published article
on Springer's website. The link must be
accompanied by the following text: "The final
publication is available at link.springer.com”.

RESEARCH ARTICLE
Chloroplast diversity indicates two independent maternallineages in cultivated grapevine (Vitis vinifera L. subsp.vinifera)
Rita Lozsa • Ning Xia • Tamas Deak •
Gyorgy Denes Bisztray
Received: 7 February 2014 / Accepted: 25 August 2014
� Springer Science+Business Media Dordrecht 2014
Abstract Chloroplast markers are powerful tools for
research into relationships among grapevine (Vitis
vinifera L. subsp. vinifera) cultivars. However, the
high number of regions common to the chloroplast and
mitochondrion genomes described in grapevine could
interfere with correct phylogeny reconstruction. In this
study, we established a unique chloroplast marker
(UCM) set and investigated the chloroplast haplotype
diversity of 17 grapevine cultivars and seven Asian
Vitis species. Sequencing of four UCMs revealed four
haplotype groups in grapevine based on three nucle-
otide substitutions, three simple sequence repeats, and
one 54 nucleotide (nt) deletion in the trnC-petN
region. The constructed molecular-variance parsi-
mony and statistical parsimony networks indicated
that two independent maternal lineages of grapevine
cultivars originated from a core group of Asian Vitis
species. The presence and absence of a newly
discovered 54 nt deletion corresponded to the two
maternal lineages: deletion (D) and non-deletion (ND)
lineages. The D lineage consisted of three haplotypes,
represented by ‘Furmint’, ‘Gouais blanc’, and ‘Sau-
vignon blanc’. Ancient noble cultivars, such as ‘Pinot
noir’, ‘Riesling’, and ‘Afus ali’, belong to a single
haplotype group (ND) in which the deletion was
absent. Our results provide compelling evidence for
the dual maternal origin of grapevine, and suggest the
occurrence of an ancient hybridization event.
Keywords Chloroplast marker � Domestication �Haplotype � Grapevine � Vitis vinifera
Introduction
Grapevine (Vitis vinifera L. subsp. vinifera [syn. V.
vinifera subsp. sativa Hegi]; Robinson et al. 2012) is
among the most ancient domesticated plants and
has a history associated with humankind since the
Neolithicum (Forni 2012). Currently, several thousand
grapevine cultivars exist, exhibiting extreme morpho-
logical and genetic diversity and also a high hetero-
zygosity that has been maintained because of
vegetative propagation since ancient times (This
et al. 2006). Polymorphism of grapevine is so
prominent that a Russian ampelographer, Alexander
Mikhailovich Negrul, established three main groups
(proles) based on the morphology, physiology, and
geographic origin of the cultivars: proles orientalis
(Armenia, Azerbaijan, Central Asia, and Iran), proles
pontica (Balkan Peninsula, Georgia, Hungary, Mol-
dova, Romania, and Turkey), and proles occidentalis
(Western Europe) (Robinson et al. 2012). This
Electronic supplementary material The online version ofthis article (doi:10.1007/s10722-014-0169-3) contains supple-mentary material, which is available to authorized users.
R. Lozsa � N. Xia � T. Deak � G. D. Bisztray (&)
Department of Viticulture, Corvinus University of
Budapest, Villanyi str. 29-43, Budapest 1118, Hungary
e-mail: [email protected]
123
Genet Resour Crop Evol
DOI 10.1007/s10722-014-0169-3
Author's personal copy

grouping is supported to some extent by genetic data
(Myles et al. 2011; Bacilieri et al. 2013).
Some cultivars, such as ‘Gouais blanc’, ‘Pinot
noir’, ‘Riesling’, and ‘Afus ali’, likely have been
preserved through historical periods and some of them
developed high clonal variation owing to human
selection (Myles et al. 2011; Bacilieri et al. 2013). As a
result of long-term vegetative propagation, the genetic
distance between modern grapevine cultivars and the
wild form might represent separation for only a few
generations, unlike herbaceous plants where propaga-
tion is usually via sexually reproduced seeds (Janick
2005).
Derivation of a phylogeny from chloroplast
genome (plastome) data is a popular approach to
reconstruct evolutionary relationships. The methods
that use organellar data for phylogeny reconstruction
are based on the hypothesis that the input data are
haploid and inherited uniparentally. In grapevine,
more than 40 % of the chloroplast genome is also
present in the mitochondrion genome (chondriome) at
least in part owing to lateral gene transfer between the
organelles. Theoretically, this could lead to an inac-
curate phylogeny reconstruction if unique chloroplast
markers (UCMs) are not used, because paralogous
regions evolve independently and could have higher
‘‘ploidity’’ and polymorphy (Goremykin et al. 2009).
Single nucleotide polymorphisms (SNPs), inser-
tions/deletions, and inversions in the chloroplast
genome are potentially informative characters for the
phylogenetic relationships of a given taxa (Shaw et al.
2005), but may be rare between closely related species
or at the infra-specific level because of the plastome’s
low mutation rate. Hypervariable chloroplast regions,
such as simple sequence repeats (cpSSRs), show
higher polymorphism at lower taxonomic levels, but
the risk of homoplasy is higher, especially when only
length polymorphism is examined. In V. vinifera,
cpSSRs are mostly mononucleotide (T or A) repeats
that have been extensively used in phylogenetic and
phylogeographic studies (Imazio et al. 2006; Arroyo-
Garcıa et al. 2006; Grassi et al. 2006; Peros et al. 2011;
Riahi et al. 2011).
In this work, we identified UCMs in silico and used
them to investigate relationships among ancient
grapevine cultivars and with Asian Vitis species that
are the closest relatives of V. vinifera (Zecca et al.
2012). Surprisingly, our results discriminated two
maternal lineages among grapevine cultivars that
originated independently from Asian Vitis species.
On the basis of our results, an ancient hybridization
event and dual maternal origin of grapevine are
hypothesized.
Materials and methods
Plant material
We sampled 163 grapevine cultivars originating from
several locations worldwide (Table 1) and seven
Asian Vitis species (Vitis amurensis Rupr., Vitis
armata Diels et Gilg, Vitis coignetiae Pulliat ex
Planch., Vitis flexuosa Thunb., Vitis piasezkii Maxim.,
Vitis romanetii Rom. Caill., and Vitis yeshanensis J.
X. Chen), and selected Muscadinia rotundifolia
Michx. as an outgroup. All accessions were sampled
from the germplasm collection of the Research
Institute for Viticulture and Oenology, University of
Pecs, Hungary, except for V. flexuosa and V. piasezkii
(obtained from the Botanical Garden of Corvinus
University of Budapest, Soroksar, Hungary), ‘Fur-
mint’ (T85 clone from Tokaj-Hetsz}ol}o Ltd, Tokaj,
Hungary), ‘Carina’, ‘Durif’, ‘Syrah’, ‘Dureza’,
‘Mondeuse blanche’, ‘Nebbiolo’, ‘Peloursin’, and
‘Roubinoy de Magaratch’ (from the INRA collection
of Vassal, Montpellier, France), and ‘Roubine’ (Julius
Kuhn Institute, Siebeldingen, Germany).
DNA extraction
Total DNA was extracted from leaf tissue based on the
method applied for Rosa roxburghii Tratt. (Xu et al.
2004) with minor modifications. Fresh or frozen leaf
tissue (50 mg) was homogenized in 1 mL washing
buffer [100 mM Tris–HCl (pH 8.0), 5 mM EDTA
(pH 8.0), 350 mM glucose, 2 % PVP, 0.4 %
b-mercaptoethanol (freshly added)] and kept on ice
for 30 min. Samples were centrifuged at 3,000 g in a
tabletop centrifuge for 10 min and then the superna-
tant was discarded. Next, 600 lL extraction buffer
[100 mM Tris–HCl (pH 8.0), 1.5 M NaCl, 50 mM
EDTA (pH 8.0), 3 % CTAB, 0.4 % b-mercap-
toethanol (freshly added)] supplemented with 1 lL
of 1 mg/mL RNAse was added to the pellet, vortexed,
and incubated at 65 �C for 30 min, then 60 lL of 5 M
potassium-acetate and 600 lL chloroform-isoamylal-
cohol were added. The samples were vortexed
Genet Resour Crop Evol
123
Author's personal copy

Ta
ble
1C
ult
ivar
sca
rry
ing
ND
and
Dh
aplo
typ
eli
nea
ges
clas
sifi
edb
yg
eog
rap
hic
ori
gin
(htt
p:/
/ww
w.v
ivc.
de)
ND
D
E:
Afu
sa
li,
Asy
lk
ara,
Oja
lesh
i,T
soli
ko
vri
W:
Ara
mo
nb
ou
chet
,C
ari
gn
an
bla
nc,
Car
ina,
Du
rif,
Gam
ayb
lan
c,G
old
ries
lin
g,
Gre
nac
he
bla
nc,
Gre
nac
he
no
ir,
Hu
mag
ne,
Mo
nd
euse
bla
nch
e,M
on
deu
sen
oir
,
Pel
ou
rsin
,P
erle
tte,
Per
lrie
slin
g,
Pin
ot
no
ir,
Rie
slin
g,
Rie
slin
gtr
amin
er,
Sy
rah
,
Tei
ntu
rier
fem
elle
,T
ein
turi
erm
ale,
Zw
eig
elt
E:
Ab
bas
i,A
lex
and
rou
li,
Ask
eri,
Bah
tio
ri,
Ber
dak
i,C
hao
uch
rozo
vy
i,D
abo
uk
i,D
ili
kaf
tar,
Do
roi
bel
yi,
Dzh
and
zhal
kar
a,H
alil
i,H
alw
ani,
Hu
ssai
ne,
Iri
kar
a,K
akh
et,
Kam
aly
,K
ara
dzh
idzh
igi,
Kar
ak
alta
k,
Kat
chit
chi,
Kat
tak
urg
an,
Kh
alil
ib
ely
i,K
hal
ili
cher
ny
i,K
har
istv
ala
ko
lkh
uri
,K
hin
do
gn
y,
Kh
uss
ain
eb
ely
i,K
ish
mis
hch
ern
y,
Kis
hm
ish
irty
sar,
Kis
hm
ish
vat
kan
a,N
imra
ng
,P
erzs
iai
mag
vat
lan
,R
ka
tsit
eli,
Su
ltan
ina,
Tag
ob
i,T
aifi
rozo
vy
i,T
avk
ver
ip
atal
ante
uly
,V
asar
ga
bel
aya,
Vas
arg
a
cher
nay
a,Z
aarm
a
W:
Ag
ost
eng
a,A
lig
ote
,A
mig
ne,
An
gel
op
iro
van
o,
Ara
mo
nb
lan
c,A
ram
on
no
ir,
Bla
ufr
ank
isch
,B
uk
ettr
aub
e,C
ab
ern
etfr
an
c,C
ard
inal
,C
arig
nan
no
ir,
Ch
ard
on
nay
bla
nc,
Ch
asse
las
cio
uta
t,C
hen
inb
lan
c,C
insa
ut,
Cla
iret
teb
lan
che,
Cla
iret
tero
se,
Du
reza
,F
oll
eb
lan
che,
Heu
nis
chro
t,H
eun
isch
wei
ss,
Gam
ayn
oir
,G
ou
ais
bla
nc,
Mer
lot,
Mu
scad
elle
,M
usc
ata
Pet
itG
rain
sB
lan
c,M
usc
atH
amb
urg
,N
ebb
iolo
,
Oh
anez
,P
icco
lit,
Piq
uep
ou
lb
lan
c,P
iqu
epo
ul
gri
s,P
iqu
epo
ul
no
ir,
Ras
meo
,R
osa
men
na
di
vac
ca,
Ro
ub
ine,
Sag
ran
tin
o,
Sai
nt
Lau
ren
t,S
au
vig
no
nb
lan
c,S
auv
ign
on
no
ir,
Sav
agn
inb
lan
c,S
chia
va
gro
ssa,
Sem
illo
n,
Sil
van
erg
run
,T
an
na
t,T
ener
on
,
Ter
od
elg
o,
Ter
ret
bla
nc,
Ter
ret
gri
s,T
imp
uri
e,T
ram
iner
gri
s,T
ram
iner
rot,
Tro
llin
ger
rot,
Ug
ni
bla
nc,
Ug
ni
rose
,V
acca
rese
,V
eltl
iner
fru
hro
t,V
eltl
iner
gra
u,
Vel
tlin
erg
run
,
Vel
tlin
erro
t
EE
B:
Bak
ato
rk
ek,
Bak
ato
rp
iro
s,B
akat
or
tud} os
zın} u,
Ba
tuta
nea
gra
,B
ereg
iro
zsas
,
Bra
gh
ina
rosi
e,C
irfa
nd
li,
Cu
ko
rsz} o
l} o,
Cse
rsze
gi
f} usz
eres
,C
sok
asz} o
l} o,
Fu
rmin
t,G
oh
erfe
her
,G
oh
erp
iro
s,G
oh
erv
alto
zo,H
efta
kil
o,Iz
sak
in
agy
szem
} u,Iz
sak
isa
rfeh
er,
Kad
ark
a,K
arp
ati
rizl
ing
,K
ecsk
ecso
cs} u,
Kek
ny
el} u,
Kir
aly
sz} ol
} o,K
ori
nth
usi
pir
os,
Ko
ver
sz} ol
} o,K
ov
idin
ka,
Ku
jun
dzu
sa,
Mu
scat
kri
msk
y,
Mu
sto
asa
de
Mad
erat
,
Pap
sap
ka,
Ro
ub
ino
yd
eM
agar
atch
,S
gh
igar
da,
Sh
esh
iZ
i,T
sits
ak
apre
i,T
uk
ors
z} ol} o
,
Urm
id
ink
a,V
arn
ensk
ag
imza
,V
oro
sh
arsl
evel
} u,Z
infa
nd
el,
Zold
din
ka,
Zsi
tvai
din
ka
Cu
ltiv
ars
inb
old
ind
icat
eac
cess
ion
sth
ath
ave
bee
nse
qu
ence
dfo
rth
efo
ur
sele
cted
UC
Ms,
wh
erea
sth
ere
mai
nd
ero
fth
ecu
ltiv
ars
wer
ese
qu
ence
do
nly
for
the
trn
C-p
etN
reg
ion
EE
aste
rn(c
ou
ntr
ies
toth
eea
stan
dso
uth
of
Asi
am
ino
r),W
Wes
tern
(fro
mA
ust
ria
toP
ort
ug
al),
EE
BE
aste
rnE
uro
pea
n–
Bal
kan
ian
(Mo
ldo
va,
Uk
rain
e,H
un
gar
y,an
dco
un
trie
so
f
the
Bal
kan
Pen
insu
laex
cep
tT
urk
ey)
Genet Resour Crop Evol
123
Author's personal copy

vigorously and then centrifuged for 10 min at 12,000 g.
The supernatant (600 lL) was aspirated and extracted
again with 600 lL chloroform-isoamylalcohol. The
supernatant (500 lL) was precipitated with 500 lL
isopropanol and 50 lL of 3 M sodium-acetate. DNA
was pelleted and washed twice with 70 % ethanol.
Pellets were resuspended in 50 lL distilled water.
Selection of chloroplast-specific markers
Given the high number of regions common to the
grapevine chloroplast and mitochondrion genomes
(Goremykin et al. 2009), the marker set was selected
only from predicted unique plastome regions. The
complete grapevine chloroplast genome (DQ424856)
(Jansen et al. 2006) and mitochondrion genomes
(NC_012119.1) (Goremykin et al. 2009) were aligned
online with PipMaker Advanced v2009-03-25-01
(Schwartz et al. 2000) and positions of the primer
sites of 17 chloroplast markers previously used in Vitis
were localized in the alignment (Fig. 1). The markers
comprised trnH-psbA, trnL-trnF, trnC-petN (Ren et al.
2011), ccmp5, ccmp3, ccmp10 (Arroyo-Garcıa et al.
2002), ccSSR-23, ccSSR-9 (Arroyo-Garcıa et al.
2006), VVCP14789, VVCP28926, VVCP50403,
VVCP13285, VVCP32585, VVCP67629,
VVCP69871, VVCP121638, and VVCP123308
(Peros et al. 2011). Those markers were predicted to
be UCMs that potentially amplify only one fragment
solely from the plastome. Figure 1 was generated from
the .pip file with Circos v0.64 (Krzywinski et al. 2009)
to visualize the marker positions. Ultimately, four
markers were selected for our study: trnC-petN (Shaw
et al. 2005), VVCP14789, VVCP28926, and
VVCP50403 (Peros et al. 2011) (see Table S1 for
additional details).
DNA amplification
Primers for the cpSSR markers VVCP14789,
VVCP28926, and VVCP50403 were described by
Peros et al. (2011) (see Table S1). Amplification was
performed with the proofreading Phusion High-Fidelity
DNA Polymerase (Thermo Scientific, Waltham, MA,
USA). Following the initial denaturation (98 �C for
30 s), amplification was performed in 35 cycles of
98 �C for 5 s, 53 �C for 10 s, and 72 �C for 10 s, with
final extension for 7 min at 72 �C. The trnC-petN
region was amplified with grapevine-specific primers
based on those used by Shaw et al. (2005) (For:
CAGTTCAAATCCGGGTG; Rev: CCAAGCGAG
ACTTACTATATCC). Polymerase chain reaction
(PCR) was conducted with Taq polymerase (MBI
Fermentas, Vilnius, Lithuania). After 3 min initial
denaturation at 94 �C, 35 cycles were performed with
the following conditions: 30 s at 94 �C, 30 s at 60 �C,
and 1 min at 72 �C, with final extension for 7 min at
72 �C. After PCR, one volume of 20 % polyethylene
glycol 6,000 (PEG; Sigma-Aldrich, Budapest, Hun-
gary) and 2.5 M NaCl (Molar Chemicals, Budapest,
Hungary) was added to the PCR reaction solution for
purification of the amplified fragments from the
primers prior to commercial direct sequencing
(Macrogen, Amsterdam, The Netherlands, and Base-
Clear, Leiden, The Netherlands) (Lundin et al. 2010).
The mixtures were incubated for 20 min at room
temperature and then centrifuged at 20,0009g for
20 min at 4 �C. The pellets were washed twice with
70 % ethanol, dried, and resuspended in 25 lL
distilled water.
Sequence analysis
Sequence assembly and raw alignments were per-
formed using CLC Sequence Viewer 6.7.1. (CLC bio
A/S, Aarhus, Denmark). Alignments were corrected
manually using BioEdit 7.1.3.0. (Hall 1999). Obtained
sequences are available in GenBank (accession num-
bers: KC962565–KC962664).
For character-based sequence analysis a matrix was
constructed, in which indels in complex regions were
treated as single mutational events (one character), but
in mononucleotide repeat regions (SSRs) every gain or
loss of a base was counted as a single mutational event
(stepwise mutation model) (Banfer et al. 2006).
Although at higher taxonomic levels hypervariable
regions are excluded from analyses (Zecca et al.
2012), at the subspecies level, inclusion of
mononucleotide repeats is commonly accepted
(Arroyo-Garcıa et al. 2006; Imazio et al. 2006). This
allowed resolution of two haplotype groups, but did
not alter the network’s topology obtained from only
SNP and indel data (data not shown).
A minimum spanning network based on molecular-
variance parsimony was constructed with Arlequin 3.5
(Excoffier et al. 2005). A statistical parsimony
network was constructed with TCS 1.21 (Clement
et al. 2000). The same data matrix was used in both
Genet Resour Crop Evol
123
Author's personal copy

analyses. Gaps were treated as a fifth character and the
connection length was set to 20 steps. The obtained
haplotype networks were redrawn manually.
For phylogenetic analysis the optimal evolutionary
model was estimated with jModelTest v2.1.5 (Guin-
don and Gascuel 2003; Darriba et al. 2012) after
exclusion of indels from the alignments. All decision
theory methods suggested the same substitution model
for all loci. Phylogenetic congruence of the chloroplast
loci was tested and verified using Concaterpillar
v1.7.2 (Leigh et al. 2008) and therefore the concate-
nated sequence set was used for subsequent analyses.
Bayesian analysis was carried out with Beast v1.8.0
(Bouckaert et al. 2014). Data were partitioned into two
sets. First, all indels were excluded leaving one partition
of sequences containing only SNP variations. The
second partition contained indel (insertions, deletions,
and SSRs) data transcoded to a binary matrix. Indels in
complex regions were treated as single mutational
events (one character), but in mononucleotide repeat
regions (SSRs) every gain or loss of a base was counted
as a single mutational event (stepwise mutation model)
(Banfer et al. 2006). Bayesian inference analysis was
carried out using the F81 substitution model for the SNP
data and a simple stochastic Dollo model for binary
(indel) data. Bayesian inference analysis was carried out
in three independent runs each of 10,000,000 genera-
tions with trees sampled at an interval of 1,000
generations. The final target tree with posterior proba-
bility (PP) values for the clades was calculated with
TreeAnnotator v1.8.1 (included with Beast) with 10 %
of the trees defined as burn-in, and was visualized with
FigTree (http://tree.bio.ed.ac.uk/software/figtree/).
Maximum parsimony (MP) analysis was carried out
using the Phylip v3.69 software package (Felsenstein
1989), using the ‘seqboot’ program to generate 1,000
bootstrapped input data matrices and ‘dnapenny’ for MP
analysis. A 50 % majority rule consensus tree was cal-
culated from the generated MP trees using the ‘con-
sense’ program of the Phylip package.
Detection of deletion in the trnC-petN region
A 54 nt deletion was identified in sequences of the
trnC-petN region in numerous cultivars (Table 1)
(accession numbers: KJ857084–KJ857228). This
ccSSR-23 §
trnH
-psb
A #
trnL-trnF # ccmp10 @
Fig. 1 Circular
representation of
homologous regions
between grapevine plastome
and chondriome, and
localization of commonly
used markers on the
chloroplast genome. The
organelle genomes are
depicted on the periphery of
the figure, homologous
regions are connected with
light gray links. Nucleotide
positions are given for every
10 and 200 kb for the
plastome and chondriome,
respectively. Note that the
sizes of the genomes are not
proportional. Location of
the 17 widely used markers
are indicated on the
plastome. IR Inverted repeat
region, SSC small single
copy region, LSC large
single copy region. @
(Arroyo-Garcıa et al. 2002),
§ (Arroyo-Garcıa et al.
2006), * (Peros et al. 2011),
# (Ren et al. 2011)
Genet Resour Crop Evol
123
Author's personal copy

deletion was large enough for it to be screened by
agarose gel electrophoresis. The PCR fragments
obtained with the trnC-petN primers were resolved on
a 2 % agarose gel in 1 9 TBE at 0.8 V/cm. Gels were
stained with ethidium bromide and size differences
among bands were detected visually in relation to
appropriate control samples.
Results
Selection of UCMs
In this study, we focused on the chloroplast diversity
of grapevine with consideration of the high amount
of genetic content in common between the organellar
genomes. We predicted regions that are specific to
the plastome by aligning the available reference
genomes of the chloroplast from ‘Syrah’ (Jansen
et al. 2006) and the mitochondrion from ‘Pinot noir’
(Goremykin et al. 2009) (Fig. 1). We examined
whether 17 markers (Peros et al. 2011; Ren et al.
2011) were located in a unique chloroplast segment
(Fig. 1). If a region surrounding the marker was
transferred to the chondriome, we also verified if the
transfer occurred in one piece, potentially allowing
amplification of the same region from both
organelles. If the transfer occurred in several pieces
the PCR reaction could be hindered.
Eight markers were predicted to lie in chloroplast-
specific regions in one copy (trnH-psbA, VVCP14789,
ccmp5, VVCP28926, trnC-petN, ccSSR-9,
VVCP50403, and trnL-trnF). An additional two
markers were located on the border of a unique and
a non-unique region (ccmp3 and ccSSR-23), therefore
a single PCR product could be obtained in these cases.
These markers were specified as UCMs.
The ccmp10 marker is located in the inverted repeat
(IR) region, thus two loci could be amplified from the
chloroplast (Fig. 1). Therefore, this marker was
excluded from the UCMs.
The remaining six markers (VVCP13285,
VVCP32585, VVCP67629, VVCP69871, VVCP121638,
and VVCP123308) were transferred to the mitochon-
drion genome in one piece. Therefore, these markers
were also excluded from further analysis.
Based on the in silico predictions we chose four
UCMs for detailed investigation: one that was used
successfully in Vitis (trnC-petN) (Ren et al. 2011;
Liu et al. 2013) and three hypervariable cpSSR
markers that were specifically designed for Vitis
(VVCP14789 [atpF-atpH], VVCP28926 [rpoB-trnC],
and VVCP50403 [trnT-trnL]) (Peros et al. 2011).
Establishing haplotype groups
We chose 17 widely cultivated and/or ancient cultivars
with diverse geographic origins (Table 1, bold acces-
sions), which are ancestors of many other cultivars
(http://www.vivc.de), and seven Asian Vitis species
and M. rotundifolia as an outgroup for investigation of
the four selected UCMs (trnC-petN, VVCP14789,
VVCP28926, and VVCP50403). We sequenced in
total 1,680 nt of the four noncoding intergenic spacers
of the plastome for the 17 representative accessions.
We detected several polymorphisms in V. vinifera
[three SNPs, three SSRs (mononucleotide repeats),
and a 54 nt deletion] and additional SNPs in the other
species (Fig. 2). Four haplotype groups were resolved
in V. vinifera, and an additional four groups among
the Vitis spp. and M. rotundifolia. Five species (V.
amurensis, V. armata, V. flexuosa, V. piasezkii, and V.
yeshanensis) were indistinguishable from each other
with this marker set.
A minimum spanning network was constructed for
the haplotype groups with molecular-variance parsi-
mony using the software Arlequin. Most Asian Vitis
species formed a core group, except for V. romanetii
and V. coignetiae, which each differed in one SNP
from the central haplotype (Fig. 2a). Surprisingly, the
four haplogroups of V. vinifera were descended from
this core group in two independent lineages. Because
the two lineages were easily distinguished by the pre-
sence or absence of a 54 nt deletion in the trnC-petN
region, we designated the deletion-containing haplo-
groups as D1, D2, and D3, and the group lacking the
deletion as non-deletion (ND) (Fig. 2).
Eastern (countries to the east and south of Asia
Minor) cultivars (‘Afus ali’ and ‘Tsolikovri’) and
Western (countries from Austria to Portugal) cultivars
(‘Carignan blanc’, ‘Pinot’, ‘Riesling’, and ‘Syrah’)
were classified in the ND group (Table 1, cf.
Figure 2). Eastern European–Balkanian (EEB; Mol-
dova, Ukraine, Hungary, and countries of the Balkan
Peninsula except Turkey) cultivars were absent from
the ND group.
In the deletion (D) lineage, the haplotype group D1
differed from the core group in a 54 nt deletion in the
Genet Resour Crop Evol
123
Author's personal copy
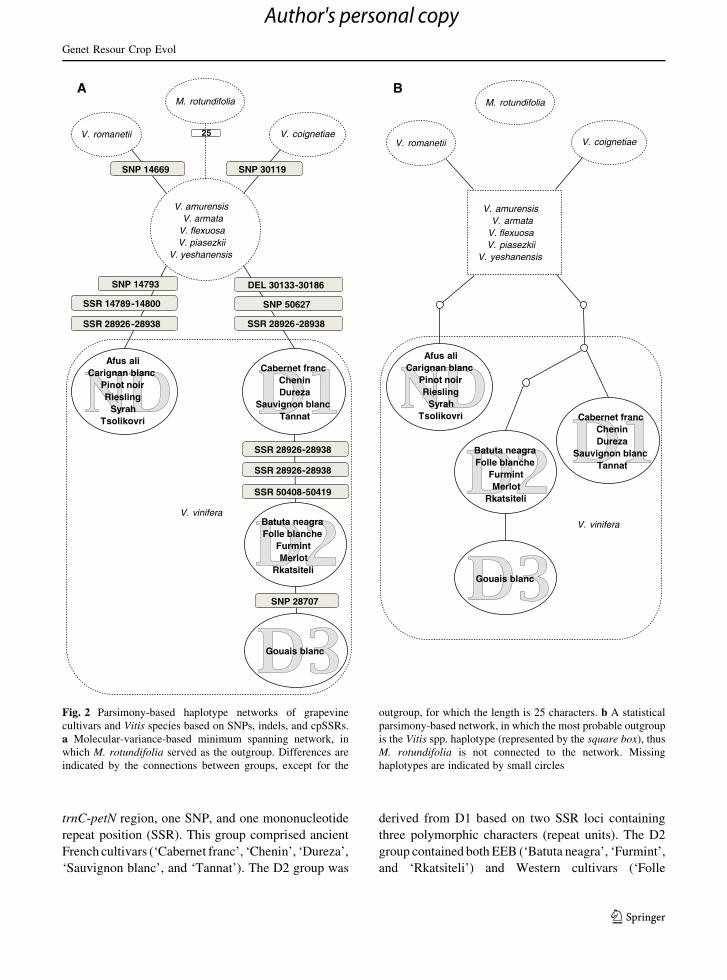
trnC-petN region, one SNP, and one mononucleotide
repeat position (SSR). This group comprised ancient
French cultivars (‘Cabernet franc’, ‘Chenin’, ‘Dureza’,
‘Sauvignon blanc’, and ‘Tannat’). The D2 group was
derived from D1 based on two SSR loci containing
three polymorphic characters (repeat units). The D2
group contained both EEB (‘Batuta neagra’, ‘Furmint’,
and ‘Rkatsiteli’) and Western cultivars (‘Folle
V. amurensis V. armata
V. flexuosa V. piasezkii
V. yeshanensis
Afus ali Carignan blanc
Pinot noir Riesling
Syrah Tsolikovri
V. coignetiae V. romanetii
M. rotundifolia
Batuta neagra Folle blanche
Furmint Merlot
Rkatsiteli
Gouais blanc
Cabernet franc Chenin Dureza
Sauvignon blanc Tannat
V. vinifera
Afus ali Carignan blanc
Pinot noir Riesling
Syrah Tsolikovri
Cabernet franc Chenin Dureza
Sauvignon blanc Tannat
Batuta neagra Folle blanche
Furmint Merlot
Rkatsiteli
Gouais blanc
M. rotundifolia
V. romanetii V. coignetiae
V. amurensis V. armata
V. flexuosa V. piasezkii
V. yeshanensis
SNP 14669 SNP 30119
SNP 14793
SNP 50627
DEL 30133-30186
SSR 14789-14800
SSR 28926-28938 SSR 28926-28938
SSR 28926-28938
SSR 28926-28938
SSR 50408-50419
SNP 28707
V. vinifera
25
A B
Fig. 2 Parsimony-based haplotype networks of grapevine
cultivars and Vitis species based on SNPs, indels, and cpSSRs.
a Molecular-variance-based minimum spanning network, in
which M. rotundifolia served as the outgroup. Differences are
indicated by the connections between groups, except for the
outgroup, for which the length is 25 characters. b A statistical
parsimony-based network, in which the most probable outgroup
is the Vitis spp. haplotype (represented by the square box), thus
M. rotundifolia is not connected to the network. Missing
haplotypes are indicated by small circles
Genet Resour Crop Evol
123
Author's personal copy

blanche’ and ‘Merlot’). Containing the haplotype most
distinct from the core group, the D3 group differed
from the D2 group in one SNP and was represented by
‘Gouais blanc’, an ancient French cultivar.
A statistical parsimony network was generated
using the software TCS (Fig. 2b). The network
contained no loops, thus indicating the absence of
homoplasy. Based on this analysis, the Asian Vitis core
group was the most probable outgroup, thus M. rotun-
difolia was not connected in the network. The ND and
D lineages were well separated and originated inde-
pendently from the Asian core group, thus supporting
the results of the molecular-variance parsimony ana-
lysis. The D lineage included three missing haplotypes
and was not linear, as the D1 group was separated from
D2 and D3. The ND lineage contained only one
missing haplotype. The ND haplotype group was
closer than D1 or D2 to the core group.
Phylogenetic analysis
We conducted Bayesian inference and MP analyses on
the eight haplotype groups and generated a Bayesian
tree with PP values complemented with bootstrap (BS)
values (Fig. 3). Indels were deleted from the align-
ments, coded as a binary character for the Bayesian
inference analysis and inserted as a separate partition
in the sequence data set for the analyses.
Consistent with the parsimony networks, the D and
ND lineages of V. vinifera were resolved in the
Bayesian tree. The highest PP value (0.99) was
obtained for the D group, which did not contain the
ND haplotype. D1 was separated from D2 and D3
(both PP = 0.75). This result was in accordance with
the statistical parsimony network (Fig. 2b), in which
the same topology was observed. All Asian Vitis
species were grouped (PP = 0.88). The ND haplotype
grouped with the Asian Vitis clade, although with little
support (PP = 0.51). In the MP analysis all BS values
were less than 95 %.
A novel easy-to-use deletion marker
The 54 nt deletion (from nt 30,133 to 30,186) in
the trnC-petN plastome region resulted in a size
difference between PCR products and enabled
screening of samples by agarose gel electrophoresis.
We screened 163 cultivars including ancient Eastern,
EEB, and Western accessions for the absence or
presence of the deletion (Table 1). Approximately
85 % of the samples yielded a short fragment length.
Sequencing of the fragments for all 163 accessions
(Table 1) proved that all of the short fragments
contained the same deletion, thus excluding possible
homoplasy.
Eastern and Western cultivars were present in both
groups [see ‘Asyl kara’ (ND), ‘Sultanina’ (D), ‘Tein-
turier’ (ND), and ‘Traminer’ (D)], whereas EEB
cultivars (e.g., ‘Heftakilo’, ‘Kovidinka’, ‘Papsapka’,
‘Tsitsa kaprei’, and ‘Varnenska gimza’) were present
exclusively in the D group. Interestingly, some
cultivars that belonged to the same conculta (colour
variants of the same cultivar; e.g., ‘Gamay blanc’ and
‘Gamay noir’, ‘Carignan blanc’, and ‘Carignan noir’)
were placed in different lineages.
Discussion
The literature on grapevine cultivar classification into
chloroplast haplotype groups is often controversial. It
is a common phenomenon that different studies
classify the same cultivar into different haplotype
groups, even if the same marker set was used (e.g.,
Arroyo-Garcıa et al. 2006 vs Peros et al. 2011). Biases
in plant collections, sampling, data handling, and
homoplasy are among the many possible reasons
behind this phenomenon. Additional complications
could arise when usage of non-unique (multilocus)
markers results in non-haploid data, given that the
plastome data are treated a priori as haploid in
M. rotundifolia
V. coignetiae
V. romanetii
V. amurensis
ND
D3
D2
D1
0,75
0,99
0,79
0,51
0,88
0,35
87
51
100
<50
Fig. 3 Bayesian tree of the eight haplotypes with PP comple-
mented with bootstrap (BS) values from MP analysis where
applicable. PP values are between 0 and 1, BS values are
between 1 and 100
Genet Resour Crop Evol
123
Author's personal copy

experimental procedures and bioinformatic analysis
pipelines. In grapevine, the high amount of genetic
content common to the chloroplast and mitochondrion
genomes raises the possibility of such a problem
(Goremykin et al. 2009). Hence, we chose UCMs to
investigate chloroplast diversity of grapevine cultivars
and their relationship with Vitis species.
We checked in silico 17 plastid markers previously
used in Vitis and found that some of the markers
indeed were located in regions that have been
transferred to the chondriome, and in addition one
marker (ccmp10) comprised two copies in the plas-
tome. Thus, PCR products for these markers likely
come from at least two loci. For example, the ccmp10
marker, which is located in the rps19–rp12 intergenic
spacer in the IR region, is highly polymorphic
(Arroyo-Garcıa et al. 2002) possibly owing to its
duplication. Interestingly, this marker was originally
designed (Weising and Gardner 1999) based on the
chloroplast genome of Nicotiana tabacum L. (Shino-
zaki et al. 1986), where this marker is on the border of
the large single copy and IR regions, potentially
yielding a single PCR product. Based on our predic-
tions, we recommend using non-UCMs with reserva-
tions or omitting them from grapevine studies.
We sequenced in total 1,680 nt of four noncoding
chloroplast intergenic spacers for eight Vitis species
and 17 grapevine cultivars. The cultivars were chosen
to represent ancient grapevines from several
geographic regions. For example, ‘Pinot noir’,
‘Rkatsiteli’, ‘Gouais blanc’, and ‘Cabernet franc’ are
all very ancient and are parents of numerous important
cultivars (Robinson et al. 2012).
Grapevine is commonly considered to have been
domesticated from Vitis vinifera L. subsp. sylvestris
(C.C.Gmel.) Hegi, which we did not analysed. This is
a highly diverse taxon that comprises morphologically
and genetically distinct geographic populations and its
taxonomic status remains uncertain (Ekhvaia and
Akhalkatsi 2010; Myles et al. 2011; Zecca et al. 2012).
We identified four haplogroups among V. vinifera
cultivars that could be distinguished by the most
polymorphic VVCP28926 marker alone. This marker
lies in the rpoB-trnC region, which is highly informa-
tive in angiosperms, even among closely related
species (Shaw et al. 2005). The four grapevine
haplogroups belong to two maternal lineages, desig-
nated ND and D. The ND group was uniform with
regard to the markers used and was identical to the
‘Syrah’ reference genome sequence (Jansen et al. 2006).
In contrast, the D group comprised three haplotypes: D1,
D2, and D3. D3, which harbored one additional SNP,
was represented by ‘Gouais blanc’ only.
According to character-based parsimony analyses,
the ND and D maternal lineages are related indepen-
dently to a central group of Asian Vitis species (Fig. 2),
which could indicate an ancient hybridization event
prior to domestication. Interspecific hybridization has
played an important role in the history of some
domesticated plant species, such as rice, apple, and
potato (Coart and Van Glabeke 2006; Takahashi et al.
2008; Gavrilenko et al. 2013). The high heterozygosity
of grapevine (Myles et al. 2011; Bacilieri et al. 2013)
and the finding that both maternal lineages are
represented among ancient cultivars from the center
of domestication (‘Rkatsiteli’ and ‘Tsolikouri’) also
support the hybridization theory. Given that we
identified more subgroups in the D lineage relative to
the ND lineage, despite the limited number of cultivars
analyzed, it is possible that the primary genetic pool
carried the deletion and that the ND haplotype
originated from a single introgression event. Lineage
D could have originated from V. v. subsp. sylvestris or
an unsampled population of an ancestral Vitis species
or species complex, not necessarily an extant popula-
tion. Another, but less likely, explanation is that
chloroplast capture (Stegemann et al. 2012), when
plastids are transmitted from one species to another
during grafting, could conceivably occur naturally
between sympatric species, although this phenomenon
has not been described in Vitis.
The phylogenetic trees obtained by Bayesian
inference and MP analyses were in agreement with
the statistical parsimony network in terms of the D
clade, for which the same topology was observed
(Figs. 2b, 3). The ND group was consistently sepa-
rated from the D clade, although its relatedness to the
other Vitis species is uncertain on account of the low
statistical support. However, it must be borne in mind
that the applicability of our marker set is limited for
evolutionary model-based phylogenetic assessment
because it is mostly based on hypervariable regions
that evolve faster than other regions, and furthermore
it resolves V. vinifera but not the other species, leaving
the latter conclusions somewhat speculative.
In the trnC-petN region, we identified a 54 nt
deletion for the first time in V. vinifera. This deletion
was absent in the investigated Vitis species and in
Genet Resour Crop Evol
123
Author's personal copy

other Vitis species investigated in previous studies
(Ren et al. 2011). This region tends to contain
relatively large deletions (Shaw et al. 2005), and the
deletions are more than twice as rare in angiosperms as
nucleotide substitutions (Shaw et al. 2007). The
identified deletion can be easily screened by agarose
gel electrophoresis. We tested the utility of the trnC-
petN marker by screening 163 grapevine cultivars for
the deletion and classified them into the D or ND
lineages. We excluded possible homoplasy by
sequencing this region in all accessions. We found
that only 15 % of the sampled cultivars belonged to
the ND group, whereas 85 % contained the deletion.
The 54 nt deletion in the trnC-petN region is
suitable for assessment of certain pedigrees or of
relationships between cultivars. For example, the ND
group contains several related cultivars, e.g., ‘Pinot
noir’, ‘Mondeause blanche’, ‘Syrah’, ‘Durif’, and
‘Carina’ belong to the same family; ‘Goldriesling’,
‘Perlriesling’, and ‘Rieslingtraminer’ are offspring of
‘Riesling’. We also observed some interesting
pedigrees by screening for the deletion: ‘Gamay noir’
and ‘Gamay blanc’ are siblings (‘Gouais blanc’ 9
‘Pinot’) (Robinson et al. 2012), but they were classi-
fied to different lineages in the present study. This
could mean that the mother was ‘Gouais blanc’ for
‘Gamay noir’ and ‘Pinot’ for ‘Gamay blanc’. ‘Zwei-
gelt’ is known to have been raised from the cross of
‘Blaufrankisch’ and ‘St Laurent’ (Maletic et al. 1999);
however, both parents were classified in the D group,
whereas ‘Zweigelt’ carries an ND haplotype. ‘Gouais
blanc’ (haplotype D3) is proven to be a parent of
‘Riesling’ (Robinson et al. 2012); our results show that
it could be the paternal parent of ‘Riesling’ as the two
cultivars belong to different chloroplast lineages. This
finding is particularly interesting, because ‘Gouais’—
on the basis of a single chloroplast SNP— is proven to
be the maternal parent of several important varieties
(Hunt et al. 2010), which we found to be classified in
the D lineage (‘Chardonnay’, ‘Gamay noir’, and
‘Aligote’) (Table 1). Although these results could be
the outcome of errors in germplasm collections or
sampling, they could also provide important informa-
tion on sometimes incorrect pedigrees.
To determine the history of domesticated plants is a
difficult task, and current findings usually complicate
existing models and result in additional uncertainties.
Our work is the first that examines the plastome
diversity with consideration of inter-organellar gene
transfer and indicates possible ambiguities in chloro-
plast marker studies in grapevine. The results provide
convincing evidence for the existence of two inde-
pendent maternal lineages in grapevine and suggest
the occurrence of an ancient hybridization event.
Acknowledgments Funding was provided by the Corvinus
University of Budapest and by the New Szechenyi Plan of the
Hungarian Government (project KTIA_AIK_12-1-2013-0001).
We thank Endre Sebestyen, Anita Lozsa, and Peter Bodor for
their help and useful comments and Pal Kozma, Sandrine Lalet,
Adam Guthermut, and Erika Maul for providing materials.
References
Arroyo-Garcıa R, Lefort F, De Andres MT, Ibanez J, Borrego J,
Jouve N, Cabello F, Martınez-Zapater JM (2002) Chloro-
plast microsatellite polymorphisms in Vitis species.
Genome 45:1142–1149
Arroyo-Garcıa R, Ruız-Garcıa L, Bolling L et al. (2006) Mul-
tiple origins of cultivated grapevine (Vitis vinifera L. ssp.
sativa) based on chloroplast DNA polymorphisms. Mol
Ecol 15:3707–3714
Bacilieri R, Lacombe T, Le Cunff L, Di Vecchi-Staraz M,
Laucou V, Genna B, Peros JP, This P, Boursiquot JM
(2013) Genetic structure in cultivated grapevines is linked
to geography and human selection. BMC Plant Biol 13:25
Banfer G, Moog U, Fiala B, Mohamed M, Weising K, Blattner
FR (2006) A chloroplast genealogy of myrmecophytic
Macaranga species (Euphorbiaceae) in Southeast Asia
reveals hybridization, vicariance and long-distance dis-
persals. Mol Ecol 15:4409–4424
Bouckaert R, Heled J, Kuhnert D, Vaughan TG, Wu C-H, Xie D,
Suchard MA, Rambaut A, Drummond AJ (2014) BEAST2:
a software platform for Bayesian evolutionary analysis.
PLoS Comput Biol 10:e1003537
Clement M, Posada D, Crandall KA (2000) TCS: a computer
programto estimate gene genealogies. Mol Ecol 9:1657–1659
Coart E, Van Glabeke S (2006) Chloroplast diversity in the
genus Malus: new insights into the relationship between
the European wild apple (Malus sylvestris L. Mill.) and the
domesticated apple (Malus domestica Borkh.). Mol Ecol
15:2171–2182
Darriba D, Taboada GL, Doallo R, Posada D (2012) jModelTest
2: more models, new heuristics and parallel computing. Nat
Methods 9:772
De Mattia F, Imazio S, Grassi F (2008) Study of genetic rela-
tionships between wild and domesticated grapevine
distributed from middle east regions to European countries.
Rend Lincei 19:223–240
Ekhvaia J, Akhalkatsi M (2010) Morphological variation and
relationships of Georgian populations of Vitis vinifera L.
subsp. sylvestris (C.C. Gmel.) Hegi. Flora 205:608–617
Excoffier L, Laval G, Schneider S (2005) Arlequin ver. 3.0: an
integrated software package for population genetics data
analysis. Evol Bioinform Online 1:47–50
Genet Resour Crop Evol
123
Author's personal copy

Felsenstein J (1989) PHYLIP-Phylogeny inference package
(version 3.2). Cladistics 5:164–166
Forni G (2012) The origin of ‘‘old world’’ viticulture. In:
Maghradze D, L Rustioni, J Turok, A Scienza, O Failla
(eds.) Caucasus and northern Black Sea region ampelog-
raphy. Vitis, special issue, JKI - Julius Kuhn-Institut,
pp 27–38
Gavrilenko T, Antonova O, Shuvalova A, Krylova E, Alpatyeva
N, Spooner DM, Novikova L (2013) Genetic diversity and
origin of cultivated potatoes based on plastid microsatellite
polymorphism. Genet Resour Crop Evol 60:1997–2015
Goremykin VV, Salamini F, Velasco R, Viola R (2009) Mito-
chondrial DNA of Vitis vinifera and the issue of rampant
horizontal gene transfer. Mol Biol Evol 26:99–110
Grassi F, Labra M, Imazio S, Rubio RO, Failla O, Scienza A,
Sala F (2006) Phylogeographical structure and conserva-
tion genetics of wild grapevine. Conserv Genet 7:837–845
Guindon S, Gascuel O (2003) A simple, fast and accurate
method to estimate large phylogenies by maximum-like-
lihood. Syst Biol 52:696–704
Hall T (1999) BioEdit: a user-friendly biological sequence
alignment editor and analysis program for Windows 95/98/
NT. Nucl Acids Symp Ser 41:95–98
Hunt HV, Lawes MC, Bower MA, Haeger JW, Howe CJ (2010)
A banned variety was the mother of several major wine
grapes. Biol Lett 6:367–369
Imazio S, Labra M, Grassi F, Scienza A, Failla O (2006)
Chloroplast microsatellites to Investigate the origin of
grapevine. Genet Resour Crop Evol 53:1003–1011
Janick J (2005) The origins of fruits, fruit growing, and fruit
breeding. Plant Breed Rev 25:255–321
Jansen RK, Kaittanis C, Saski C, Lee SB, Tomkins J, Alverson
AJ, Daniell H (2006) Phylogenetic analyses of Vitis
(Vitaceae) based on complete chloroplast genome sequen-
ces: effects of taxon sampling and phylogenetic methods on
resolving relationships among rosids. BMC Evol Biol 6:32
Krzywinski M, Schein J, Birol I, Connors J, Gascoyne R,
Horsman D, Jones SJ, Marra MA (2009) Circos: an infor-
mation aesthetic for comparative genomics. Genome Res
19:1639–1645
Leigh JW, Susko E, Baumgartner M, Roger AJ (2008) Testing
congruence in phylogenomic analysis. Syst Biol
57:104–115
Liu XQ, Ickert-Bond SM, Chen LQ, Wen J (2013) Molecular
phylogeny of Cissus L. of Vitaceae (the grape family) and
evolution of its pantropical intercontinental disjunctions.
Mol Phylogenet Evol 66:43–53
Lundin S, Stranneheim H, Pettersson E (2010) Increased
throughput by parallelization of library preparation for
massive sequencing. PLoS One 5:e10029
Maletic E, Sefc K, Steinkellner H, Konti J, Pejic I (1999)
Genetic characterization of Croatian grapevine cultivars
and detection of synonymous cultivars in neighboring
regions. Vitis 38:79–83
Myles S, Boyko AR, Owens CL et al (2011) Genetic structure
and domestication history of the grape. Proc Natl Acad Sci
USA 108:3530–3535
Peros JP, Berger G, Portemont A, Boursiquot JM, Lacombe T
(2011) Genetic variation and biogeography of the disjunct
Vitis subg. Vitis (Vitaceae). J Biogeogr 38:471–486
Ren H, Lu L, Soejima A, Luke Q, Zhang D, Chen Z, Wen J
(2011) Phylogenetic analysis of the grape family (Vita-
ceae) based on the noncoding plastid trnC-petN, trnH-psbA
and trnL-F sequences. Taxon 60:629–637
Riahi L, Zoghlami N, Laucou V, Mliki A, This P (2011) Use of
chloroplast microsatellite markers as a tool to elucidate
polymorphism, classification and origin of Tunisian
grapevines. Sci Hortic 130:781–786
Robinson J, Harding J, Vouillamoz J (2012) Wine Grapes.
HarperCollins, USA
Schwartz S, Zhang Z, Frazer KA, Smit A, Riemer C, Bouck J,
Gibbs R, Hardison R, Miller W (2000) PipMaker—a web
server for aligning two genomic DNA sequences. Genome
Res 10:577–586
Shaw J, Lickey E, Beck J (2005) The tortoise and the hare II:
relative utility of 21 noncoding chloroplast DNA sequences
for phylogenetic analysis. J Bot 92:142–166
Shaw J, Lickey EB, Schilling EE, Small RL (2007) Comparison
of whole chloroplast genome sequences to choose non-
coding regions for phylogenetic studies in angiosperms: the
tortoise and the hare III. Am J Bot 94:275–288
Shinozaki K, Ohme M, Tanaka M et al. (1986) The complete
nucleotide sequence of the tobacco chloroplast genome: its
gene organization and expression. Eur Mol Biol Org J
5:2043–2049
Stegemann S, Keuthe M, Greiner S, Bock R (2012) Horizontal
transfer of chloroplast genomes between plant species.
Proc Natl Acad Sci USA 109:2434–2438
Takahashi H, Sato Y, Nakamura I (2008) Evolutionary analysis
of two plastid DNA sequences in cultivated and wild spe-
cies of Oryza. Breed Sci 58:225–233
This P, Lacombe T, Thomas MR (2006) Historical origins and
genetic diversity of wine grapes. Trends Genet 22:511–519
Weising K, Gardner RC (1999) A set of conserved PCR primers
for the analysis of simple sequence repeat polymorphisms
in chloroplast genomes of dicotyledonous angiosperms.
Genome 42:9–19
Xu Q, Wen X, Deng X (2004) A simple protocol for isolating
genomic DNA from chestnut rose (Rosa roxburghii Tratt.)
for RFLP and PCR analyses. Plant Mol Biol Rep
22:301–302
Zecca G, Abbott JR, Sun WB, Spada A, Sala F, Grassi F (2012)
The timing and the mode of evolution of wild grapes
(Vitis). Mol Phylogenet Evol 62:736–747
Genet Resour Crop Evol
123
Author's personal copy
Copyright © 2022 FDOKUMEN