
Cloning, functional identification and structural modelling of Vitis vinifera S-adenosylmethionine...
12
Journal of Plant Physiology 164 (2007) 1208—1219 Cloning, functional identification and structural modelling of Vitis vinifera S-adenosylmethionine decarboxylase Annalisa Tassoni a , Marina Franceschetti a , Gianluca Tasco b , Rita Casadio a,b , Nello Bagni a, a Department of Biology e.s. and Interdepartmental Centre for Biotechnology, University of Bologna, Via Irnerio 42, Bologna 40126, Italy b CIRB Biocomputing Group, University of Bologna, Bologna, Italy Received 12 May 2006; accepted 5 July 2006 KEYWORDS Grapevine; Homology modelling; Polyamines; S-adenosylmethio- nine decarboxylase; Vitis vinifera Summary In this paper we report the cloning and full sequencing of S-adenosylmethionine decarboxylase (SAMDC, EC 4.1.1.50) cDNA from Vitis vinifera L. (VV) leaves, an enzyme belonging to the polyamine biosynthetic pathway, which appears to play an important role in the regulation of plant growth and development. The presence of two overlapping ORFs (tiny ORF and small ORF) upstream of the main ORF is reported in the Vitis cDNA. When the Vitis SAMDC cDNA was expressed in yeast without the two upstream ORFs, the resulting activity was about 50 times higher than the activity obtained with the full cDNA. These results demonstrated the strong regulatory activity of the tiny and small ORFs. RT-PCR expression analysis showed evidence of a similar mRNA level in all the tissues tested, with the exception of the petioles. The VV SAMDC was also modelled using its homologues from Solanum tuberosum and Homo sapiens as template. The present work confirmed, for the first time in a woody plant of worldwide economic interest such as grapevine, the presence of a regulatory mechanism of SAMDC, enzyme that has a well-established importance in the modulation of plant growth and development. & 2006 Elsevier GmbH. All rights reserved. Introduction Grapevine plant (Vitis vinifera L. (VV)) is representative of a group of deciduous perennial plants that produce non-climateric fleshy fruits ARTICLE IN PRESS www.elsevier.de/jplph 0176-1617/$ - see front matter & 2006 Elsevier GmbH. All rights reserved. doi:10.1016/j.jplph.2006.07.009 Abbreviations: MGBG, methylglyoxal-bis(guanilhydrazone); ORF, open reading frame; RSMD, root-mean-square deviation; RT-PCR, reverse transcriptase-polymerase chain reaction; SAMDC, S-adenosylmethionine decarboxylase; T a , annealing temperature; uORF, upstream open reading frame. Corresponding author. Tel.: +390512091280; fax: +39 051242576. E-mail address: [email protected] (N. Bagni).
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Cloning, functional identification and structural modelling of Vitis vinifera S-adenosylmethionine...

ARTICLE IN PRESS
Journal of Plant Physiology 164 (2007) 1208—1219
0176-1617/$ - sdoi:10.1016/j.
AbbreviationORF, open readRT-PCR, reversSAMDC, S-adentemperature; u�Correspond
fax: +39 05124E-mail addr
www.elsevier.de/jplph
Cloning, functional identification and structuralmodelling of Vitis vinifera S-adenosylmethioninedecarboxylase
Annalisa Tassonia, Marina Franceschettia, Gianluca Tascob,Rita Casadioa,b, Nello Bagnia,�
aDepartment of Biology e.s. and Interdepartmental Centre for Biotechnology, University of Bologna,Via Irnerio 42, Bologna 40126, ItalybCIRB Biocomputing Group, University of Bologna, Bologna, Italy
Received 12 May 2006; accepted 5 July 2006
KEYWORDSGrapevine;Homology modelling;Polyamines;S-adenosylmethio-nine decarboxylase;Vitis vinifera
ee front matter & 2006jplph.2006.07.009
s: MGBG, methylglyoing frame; RSMD, root-e transcriptase-polymerosylmethionine decarboORF, upstream open reing author. Tel.: +39 0512576.ess: [email protected]
SummaryIn this paper we report the cloning and full sequencing of S-adenosylmethioninedecarboxylase (SAMDC, EC 4.1.1.50) cDNA from Vitis vinifera L. (VV) leaves, anenzyme belonging to the polyamine biosynthetic pathway, which appears to play animportant role in the regulation of plant growth and development. The presence oftwo overlapping ORFs (tiny ORF and small ORF) upstream of the main ORF is reportedin the Vitis cDNA. When the Vitis SAMDC cDNA was expressed in yeast without thetwo upstream ORFs, the resulting activity was about 50 times higher than theactivity obtained with the full cDNA. These results demonstrated the strongregulatory activity of the tiny and small ORFs. RT-PCR expression analysis showedevidence of a similar mRNA level in all the tissues tested, with the exception of thepetioles. The VV SAMDC was also modelled using its homologues from Solanumtuberosum and Homo sapiens as template. The present work confirmed, for the firsttime in a woody plant of worldwide economic interest such as grapevine, thepresence of a regulatory mechanism of SAMDC, enzyme that has a well-establishedimportance in the modulation of plant growth and development.& 2006 Elsevier GmbH. All rights reserved.
Elsevier GmbH. All rights rese
xal-bis(guanilhydrazone);mean-square deviation;ase chain reaction;xylase; Ta, annealingading frame.2091280;
bo.it (N. Bagni).
Introduction
Grapevine plant (Vitis vinifera L. (VV)) isrepresentative of a group of deciduous perennialplants that produce non-climateric fleshy fruits
rved.

ARTICLE IN PRESS
Grapevine S-adenosylmethionine decarboxylase 1209
containing a great variety of secondary metabolitessuch as those responsible for colour, flavour, aromaand defence against pathogens.
Among secondary metabolites, polyamines aresmall, positively charged aliphatic amines found inall living organisms. They are involved in a varietyof processes in plants, including the regulation ofDNA replication, cell division, organ development,fruit ripening, and leaf senescence (Bagni andTassoni, 2001). Their biosynthetic pathway isrelatively well established. Putrescine is produceddirectly from ornithine via ornithine decarboxylase(ODC; EC 4.1.1.17) or indirectly from arginine viaarginine decarboxylase (ADC; EC 4.21.1.19). Sper-midine and spermine are formed, respectively, byaddition to putrescine or spermidine of an amino-propyl moiety that results from the decarbo-xylation of S-adenosylmethionine (SAM) byS-adenosylmethionine decarboxylase (SAMDC;EC 4.1.1.50). SAMDC is considered to be therate-limiting enzyme for the synthesis of spermi-dine and spermine (Kumar et al., 1997; Tiburcioet al., 1997).
Isolation of genomic and cDNA sequences ofSAMDC from several plant systems indicated thatthis enzyme is generally encoded by a smallmultigene family in monocot and dicot plants(Franceschetti et al., 2001). Analysis of rice, maizeand Arabidopsis thaliana SAMDC cDNA speciesrevealed that the monocot enzyme possesses anextended C-terminus relative to dicot and humanenzymes. Interestingly, it was discovered that allexpressed plant SAMDC mRNA 50 leader sequencescontain a highly conserved pair of upstream openreading frames (uORFs) that overlap by one base(Franceschetti et al., 2001). The tiny 50 uORFconsists of two or three codons and the 30 smalluORF encodes 50–54 residues. Sequences of thesmall uORF are conserved between monocot, dicotand gymnosperm SAMDC mRNA species. Such aconserved arrangement is strongly suggestive of aregulatory mechanism. Different research groupshave investigated this regulation, hypothesisingtwo levels of gene expression control. Hu et al.(2005) suggested that the leader sequence couldplay a role at both the transcriptional and post-transcriptional levels. On the other hand, theelimination of the small uORF from A. thalianaSAMDC1 cDNA led to increased relative transla-tional efficiency of SAMDC proenzyme in transgenictobacco plants. The resulting increased activity ofSAMDC caused disruption to polyamine levels andsevere growth and development defects (Hanfreyet al., 2002). In vitro expression demonstrated thatboth tiny and small uORFs are translated and thatthe tiny uORF is required for normal polyamine-
responsive SAMDC mRNA translation, probablyachieved by control of ribosomal recognition ofthe small uORF (Hanfrey et al., 2005).
Most decarboxylases require pyridoxal phosphateas cofactor. Instead, SAMDC is a pyruvoyl enzyme,and the covalently bound prosthetic group pyruvateis formed in an internal autocatalytic reaction thatcleaves the proenzyme precursor and converts aserine into pyruvate. This reaction occurs in severalorganisms including plants (e.g., potato) (Xionget al., 1997), humans (Stanley et al., 1994) and alsoin the bifunctional SAMDC/ODC enzyme of Plasmo-dium falciparum (Wrenger et al., 2001). Studiesperformed on potato, human, yeast and P. falcipar-um SAMDCs have shown that the cleavage site wascontained in the sequence YVLSESS with theunderlined serine forming the pyruvate (Kashiwagiet al., 1990; Stanley and Pegg, 1991; Xiong et al.,1997; Wrenger et al., 2001). Moreover, the auto-calysis of the proenzyme generates an a subunit,from the C-terminus, and a b subunit from theN-terminus, both taking part to the final folding ofthe active protein. In humans both the enzymeactivity and the proteolityc processing are stimu-lated by putrescine (Stanley et al., 1994), whilein plants (Xiong et al., 1997) and P. falciparum(Wrenger et al., 2001) no stimulation was observed,after putrescine addition. Structural study onsolved SAMDC of human and potato and site-directed mutagenesis (Ekstrom et al., 2001; Ben-nett et al., 2002), as well as the computationalmodelling of homologue P. falciparum enzyme(Wells et al., 2006), confirmed these results.
The present paper reports the full sequencingand expression in yeast of VV SAMDC cDNA. Inaddition to the well-established importance ofSAMDC in the regulation of plant growth anddevelopment, no information is available aboutthe cDNA sequence in this globally important woodycrop. Modelling of the protein three-dimensional(3D) structure reveals interesting features of thisenzyme that account for the lack of putrescinebinding site, confirming, for the first time, this typeof mechanism in a woody plant.
Materials and methods
VV samples
Small young leaves (SL; less than 8 cm of length),big fully expanded leaves (BL; 8–15 cm length),petioles (P) and apical green branches (B) ofgrapevine (VV L.) were collected during thevegetative season from an adult plant (about 30

ARTICLE IN PRESS
A. Tassoni et al.1210
years old) of the Bologna’s Botanical Garden,immediately frozen in liquid nitrogen and storedat �80 1C until used for experiments.
RNA extraction and SAMDC cDNA cloning
For SAMDC cDNA cloning, total RNAwas extractedfrom about 2 g of grape small leaves as described byChang et al. (1993).
SAMDC degenerated primers for both reversetranscriptase-polymerase chain reaction (RT-PCR)and PCR amplifications were designed on conservedregions of aligned plant database sequences (datanot shown). Mixtures of nucleotides in the primersare indicated with the following one-letter code: R:a, g; Y: c, t; M: a, c; K: g, t; S: g, c; W: a, t; H: a, t,c; B: g, t, c; V: g, a, c; D: g, a, t; N: a, g, c, t. Totalextracted RNA (0.5 mg) was used for RT-PCR (Ready-To-Go RT-PCR Beads Kit, Amersham Biosciences,Little Chalfont, UK) according to the manufac-turer’s instructions. The amplification of 1.1 kbSAMDC cDNA fragment was carried out using FOR1(50-CTYGGHTACAGGATTGARGAC-30) and REV1 (50-TCATDGARTARCCRCAGGG-30) primers (25 pmoleach) in a PTC-150 MiniCycler (MJ Research,Waltham, MA, USA) at an annealing temperature(Ta) of 52.5 1C. Aliquots (0.5–1 mL) of these reactionmixtures were used as template for nested PCRamplification (Ready-To-Go PCR Beads Kit,Amersham Biosciences, UK) using FOR2(50-TCMRRTCWKCTGYHTACTCCAAC-30) and REV2(50-GYTCRAAVTCAAARTCRCA-30) degenerated pri-mers (25 pmols each) at a Ta of 50 1C. The obtainedSAMDC cDNA fragment (0.9 kb) was gel extracted,amplified with Pfu DNA polymerase (Promega,Madison, WI, USA), cloned with the PCR-ScriptTM
Amp Cloning Kit (Stratagene, La Jolla, CA, USA) andsequenced. The identity of the fragment wasconfirmed by database searching. The full-lengthSAMDC cDNA was successfully obtained by RT-PCR(Prostar Ultra HF RT-PCR System, Stratagene) usingfor the first strand amplification 0.5 mg of total RNAand a mixture of oligo(dT) and for the secondstrand 25 pmol each of the primers FOR3 (50-CGTGAATGAKMTRATGGARTC-30) and NotI(dT)18(50-AACTGGAAGAATTCGCGGCCGCAGGAA(T)18-30)at a Ta of 52 1C. The 2.1 kb cDNA was amplified byPfu Turbo DNA polymerase (Stratagene, La Jolla,CA, USA) and cloned as described above. The cDNAwas fully sequenced using primers T3, T7 and thenon-degenerated primer FOR5 (50-TTGAAGTTGGCT-GACACC-30) designed on a previously sequencedregion of the 0.9 kb cDNA fragment. The completeVV SAMDC cDNA sequence (Fig. 1) was reported in
the EMBL Nucleotide Sequence database AccessionNo. AJ567368.
Expression in yeast and enzyme activity
For expression in yeast, two SAMDC fragments,FULL and MAIN, were cloned into the pYES2expression vector (Invitrogen, Milan, Italy), down-stream of a galactose-inducible GAL1 promoter.The FULL clone contained the 1.7 kbp full-lengthVitis SAMDC. The MAIN clone consisted of a 1.4 kbpKpnI–NotI fragment containing the main ORF andlacking of the upstream 50 UTR with the tiny andsmall ORFs. It was obtained by PCR amplificationwith FOR6 (50-CCACGGTACCGCTTGACATTGTT-30
from base 268 to 290 of the full-length clone(Fig. 1; KpnI site underlined) and NotI(dT)18primers.
For yeast transformation, the Saccharomycescerevisiae strain 2602 (MATa ura3-52 his6 leu2)(obtained from H. Tabor, National Institutes ofHealth, Bethesda, MD) was used. Yeast cultureswere grown aerobically at 30 1C in minimal YPADmedium. Yeast transformation was performed usinga modified lithium acetate procedure (Elbe, 1992).Three independent transformed colonies for eachyeast strain (FULL, MAIN and empty pYES2) weregrown in 100mL SC medium minus uracile with 2%(w/v) glucose (not induced samples), or lacking ofglucose but with 4% (w/v) galactose (inducedsamples), at 30 1C for 16 h. As a negative control,the wild-type 2602 yeast strain was grown underthe same conditions in 100mL minimal SC mediumplus 0.01M uracile with 2% glucose or with 4%galactose. The cultures were pelleted (1000 g,5min), washed with deionised water and immedi-ately used for enzyme assay.
SAMDC activity was determined by a radiochemi-cal method as described by Tassoni et al. (2000).The pelleted cell cultures were resuspended in500 mL of SAMDC assay buffer (100mM Tris-HCl (pH7.6), 50 mM EDTA), added with 400mg of acid-washed glass beads (425–600 mm diameter, SIGMA,Milan, Italy), vortexed five times for 30 s each todisrupt the cells, and centrifuged (20,000 g, 30min,4 1C) to remove the cell debris. The supernatantwas removed and brought to 1.5mL with assaybuffer and 0.2mL aliquots were incubated sepa-rately with 7.4 kBq [1-14C]S-adenosylmethionine(specific activity 2.06 GBqmmol�1, Amersham Bios-ciences, Little Chalfont, UK). The activity ofinduced samples was determined in the absenceor in the presence of 200 mM methylglyoxal-bis(guanilhydrazone) (MGBG), an inhibitor of SAMDCenzyme. The 14CO2 evolution was measured for 2 h

ARTICLE IN PRESS
tgcgggggcggggtccccctcgaggtcgacggtatcgataagcttgatatcgaattcctg 60FOR 3 Tiny ORF – Small ORF
cagcccgggggatccgccccgtgaatgagataatggagtctaaacggagcaagaagtcta 120FOR 1
gcagtagtaaatcgttattctatgaagctcctcttggctacagcattgaagacattcgac 180FOR2
cgaacggtggaatcaagaagttcagatctgctgcatactccaactgcgctcgcaaaccat 240FOR6
cctgatattcataagttggctgcctgaccacgtcgccgcttgacattgttaactgttgac 300
gtggtcgtttaattgttacagctgtagtttaaaatccttttcttcttcaacaaccacttt 360cttgtcctctctttgggactgtaatcacttctgttagtcaacatggccttgccagtctca 420
M A L P V S
gcaattggatttgaaggttatgaaaagaggctagagatatcattttctgaaccaggcatc 480A I G F E G Y E K R L E I S F S E P G I
ttttctgatcctgaaggaaggggtctccgatccctgtcgagagcccagttggatgagatt 540F S D P E G R G L R S L S R A Q L D E I
cttgaaccagctgagtgcactatagttggttcactgtcaaatgatattgttgattcctat 600L E P A E C T I V G S L S N D I V D S Y
gtcttgtctgagtctagcctctttgtttacccttacaagatcatcatcaaaacctgtggg 660V L S E S S L F V Y P Y K I I I K T C G
acaacaaagctgcttctttcaatcccacccatcttgaagttggctgacaccctctccctc 720T T K L L L S I P P I L K L A D T L S L
tctgtaagttgtgtgaggtatactcgtgggagcttcaattttcctggtgctcagccatat 780S V S C V R Y T R G S F N F P G A Q P Y
cctcatcgtcacttctcagaagaagtggccgttcttgacagctattttgggaagcttggt 840P H R H F S E E V A V L D S Y F G K L G
tcaggtagcaaggcttatgtgatgggtagttctgacaaatcacagaaatggcatgtgtac 900S G S K A Y V M G S S D K S Q K W H V Y
tctgcttctgcagaagtgaggagcgcctgtgaccctgtttatactctggagatgtgcatg 960S A S A E V R S A C D P V Y T L E M C M
actggtttggacagggagatggcttctgtattctacaaaacccactcaagctcagcagtt 1020T G L D R E M A S V F Y K T H S S S A V
REV 2aaaatgactgacacttctggtataagaaagattcttccagattctgaaatatgtgatttt 1080K M T D T S G I R K I L P D S E I C D F
REV 1 gaatttgatccatgtggttattccatgaatgccattgaaggagctgcaatttccaccatt 1140E F D P C G Y S M N A I E G A A I S T I
catgttacgcctgaagatggttttagttatgctagctttgaaacggtgggatataatcca 1200H V T P E D G F S Y A S F E T V G Y N P
REV 3aaagatgtgaacctgagccatctgattgaaagggtgttgtcttgtttccaaccgaatgag 1260K D V N L S H L I E R V L S C F Q P N E
ttctctgtagcagtgcatgctgacatctcgggtaagttactcgagcgaaattgccttctg 1320F S V A V H A D I S G K L L E R N C L L
gatgtaaagggatactgttgtgaagagagaagcaacgaagaacttggtatgtgtggttcc 1380D V K G Y C C E E R S N E E L G M C G S
atggtataccataggttcatgaagactgaggggttggtgtctcctagatcaattctgaaa 1440M V Y H R F M K T E G L V S P R S I L K
tgctgctggaaagaggaagaagaagaagaaaaggaatagctgaatggtttattttttgtt 1500C C W K E E E E E E K E
tgtcttgtctatggttaaataagccaacagtttgatggcctattaatatgatgcttgtag 1560tcatattatcagccgagatgatgttggatttacttttagaagtctggtgttgagccagaa 1620acttgcagctatgtttgtgcttatgagtccatttaaatatttgataatgttaaacccggc 1680 agtttgagggtaaaaaaaaaaaaaaaaaa 1709
FOR 5
Main ORF
Not I (dT)18
Figure 1. Full nucleotide sequence of Vitis vinifera S-adenosylmethionine decarboxylase cDNA. The different forwardand reverse primers (underlined nucleotides), the presence of the tiny ORF and small ORF (Franceschetti et al., 2001)and the deduced aminoacid sequence of the main ORF are indicated. This sequence was reported in the EMBLNucleotide Sequence database Accession No. AJ567368.
Grapevine S-adenosylmethionine decarboxylase 1211

ARTICLE IN PRESS
A. Tassoni et al.1212
at 37 1C using 1mL of 1M KOH as trappingcompound. The amount of radioactivity was mea-sured with a scintillation counter (Beckman LS6500, Beckman Instruments Inc., Fullerton, CA,USA). Protein content was determined according toLowry et al. (1951). The experiment was repeatedtwice with three independent clones for eachtransformed strain and triplicate samples for eachenzymatic assay.
Southern blot analysis and RT-PCRexpression analysis
For Southern blot analysis about 20 mg of genomicDNA isolated from young grapevine leaves (Michaelet al., 1996) was digested with EcoRV, HindIII andSacI restriction enzymes (Stratagene), run onto0.7% (w/v) agarose gel in 1� TBE buffer (Sambrooket al., 1989) and blotted onto a HybondTM-N nylonmembranes (Amersham Biosciences, UK).
The previously amplified 0.9 kb SAMDC cDNAfragment was labelled with a[32P]dCTP (Ready-To-Go DNA Labelling Beads Kit, Amersham Biosciences,UK) and used as probe for hybridisation. Pre-hybridisation (2 h) and hybridisation (16 h) werecarried out at 42 1C following Sambrook et al.(1989). The membrane was washed at high strin-gency for 30min at 60 1C and exposed to 32PImaging Plate for Fluorescent Image Analyzer(FLA-3000, Fujifilm, Japan). Densitometry of ex-posed films was performed by image analysissoftware TotalLab version 2.01 (Nonlinear DynamicsLtd., Newcastle upon Tyne, UK).
For RT-PCR expression analysis, total RNA wasextracted from 2 g of small young leaves (SL), bigfully expanded leaves (BL), petioles (P) and apicalgreen branches (B) (Chang et al., 1993). Theamount of total RNA used in each SAMDC amplifica-tion reaction was calibrated by RT-PCR (Ready-To-Go RT-PCR Beads Kit, Amersham Biosciences, UK)using primers amplifying the Vitis actin gene(vvACTfor: 50-GTGCTGGATTCTGGTGATGG-30; vvAC-Trev: 50-AAGAGGACTTCTGGACAACGG-30), 20 cyclesat a Ta 55 1C (331 bp fragment). Vitis SAMDC RT-PCRexpression analysis was carried out by using FOR5and REV3 (50-AATCAGATGGCTCGGTTC-30) primers(Fig. 1), 20 cycles at a Ta of 49 1C (516 bp fragment).
Homology modelling of SAMDC
The 3D model of SAMDC from VV was computedby comparison (building by homology) with the 3Dstructures of homologues from Solanum tuberosum(ST; PDB code:1MHM), known with a 0.23 nmresolution (Bennett et al., 2002), and from Homo
sapiens (HS; PDB code:1JL0), X-ray solved at0.15 nm resolution (Ekstrom et al., 2001), respec-tively. Sequence identities between VV/ST and VV/HS were approximately 76% and 36%. The templatesequence of SAMDC from ST structurally covered allthe target chain of VV with the exception of theN-terminus portion, where one of the two glutamicacid residues, included in the enzyme active site,was missing (green box in Fig. 6). This part wascovered and modelled only considering the align-ment with the HS counterpart. Alignment was donewith ClustalW (Thompson et al., 1994) (withBlosum62 and without gap penalty), and manuallychecked. Modelling was performed with the pro-gram Modeller v7.7 (Marti-Renom et al., 2000).For a given alignment, ten model structures werebuilt and were evaluated and validated with thePROCHECK and PROSA2003 suites of programs(Laskowski et al., 1993; Sippl, 1993).
Results and discussion
The full nucleotide sequence of S-adenosyl-methionine decarboxylase cDNA and the deducedamino acid sequence of the main ORF (358 aminoacids) are reported in Fig. 1. Two overlapping smallORFs (named tiny uORF and small uORF, Fig. 1)upstream of the main ORF were also present in theVitis cDNA such as in other plants (Franceschettiet al., 2001). In fact, the Vitis 50 tiny uORF consistsof three codons (Fig. 1 from base 84 to 93), twocodons encoding the amino acids MR and one stopcodon, while the small uORF encodes 50 residues(Fig. 1 from 92 to 245). The deduced amino acidsequence of the Vitis small uORF (MESKRSKK-SSSSKSLFYEAPLGYSIEDIRPNGGIKKFRSAAYSNCARKPS)shows a sequence identity higher than 75% with thesmall uORF of the known plant SAMDCs.
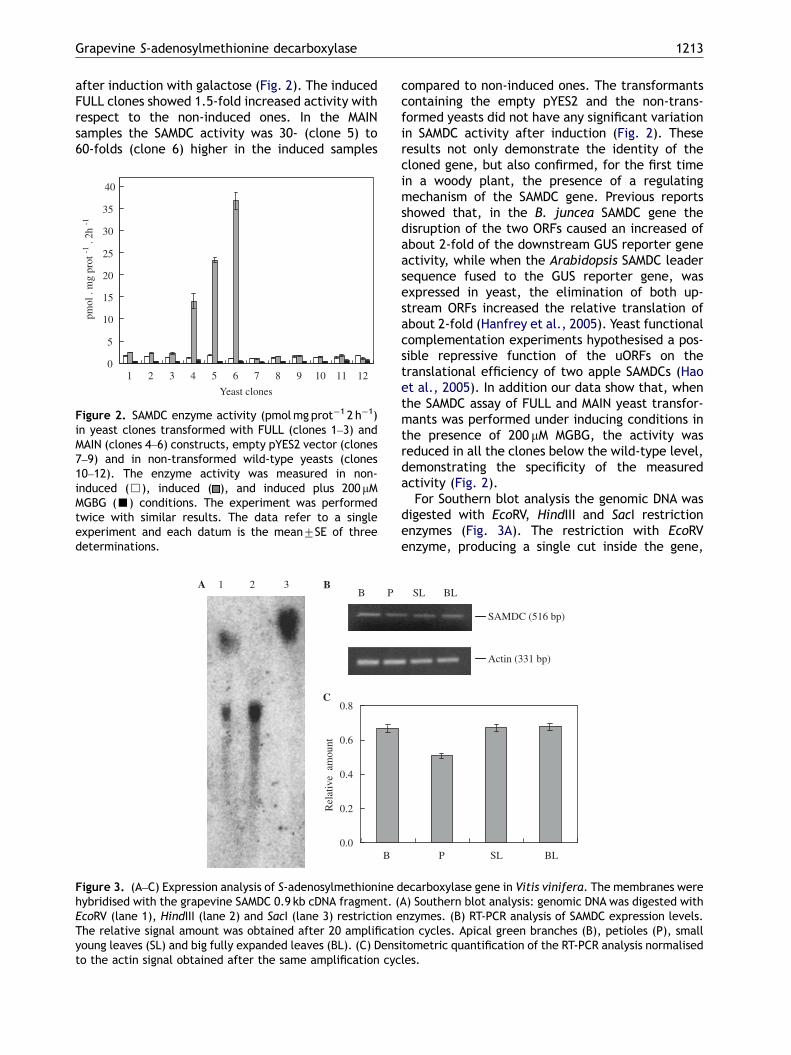
The high sequence identity of these two uORFswith the other cDNA sequences present in thedatabase lead to the hypothesis of the presence ofa gene expression regulation mechanism also inVitis, as proposed for Brassica juncea at thetranscriptional level (Hu et al., 2005) and for A.thaliana at the translational level (Hanfrey et al.,2002, 2005). Two SAMDC fragments, FULL (fulllength cDNA) and MAIN (main ORF without theupstream ORFs) were cloned into the pYES2expression vector, downstream of a galactose-inducible GAL1 promoter. SAMDC activity wasmeasured in three independent transformants ofFULL (clones 1–3), MAIN (clones 4–6), empty pYES2(clones 7–9) and in three non-transformed wild-type yeast colonies (clones 10–12), before and
ARTICLE IN PRESS
Grapevine S-adenosylmethionine decarboxylase 1213
after induction with galactose (Fig. 2). The inducedFULL clones showed 1.5-fold increased activity withrespect to the non-induced ones. In the MAINsamples the SAMDC activity was 30- (clone 5) to60-folds (clone 6) higher in the induced samples
0
5
10
15
20
25
30
35
40
1 2 3 4 5 6 7 8 9 10 11 12
Yeast clones
pmol
. m
g pr
ot -1
. 2h
-1
Figure 2. SAMDC enzyme activity (pmolmg prot�1 2 h�1)in yeast clones transformed with FULL (clones 1–3) andMAIN (clones 4–6) constructs, empty pYES2 vector (clones7–9) and in non-transformed wild-type yeasts (clones10–12). The enzyme activity was measured in non-induced (&), induced ( ), and induced plus 200 mMMGBG (’) conditions. The experiment was performedtwice with similar results. The data refer to a singleexperiment and each datum is the mean7SE of threedeterminations.
1 2 3A
C
B
0.0
0.2
0.4
0.6
0.8
B
B P
Rel
ativ
e a
mou
nt
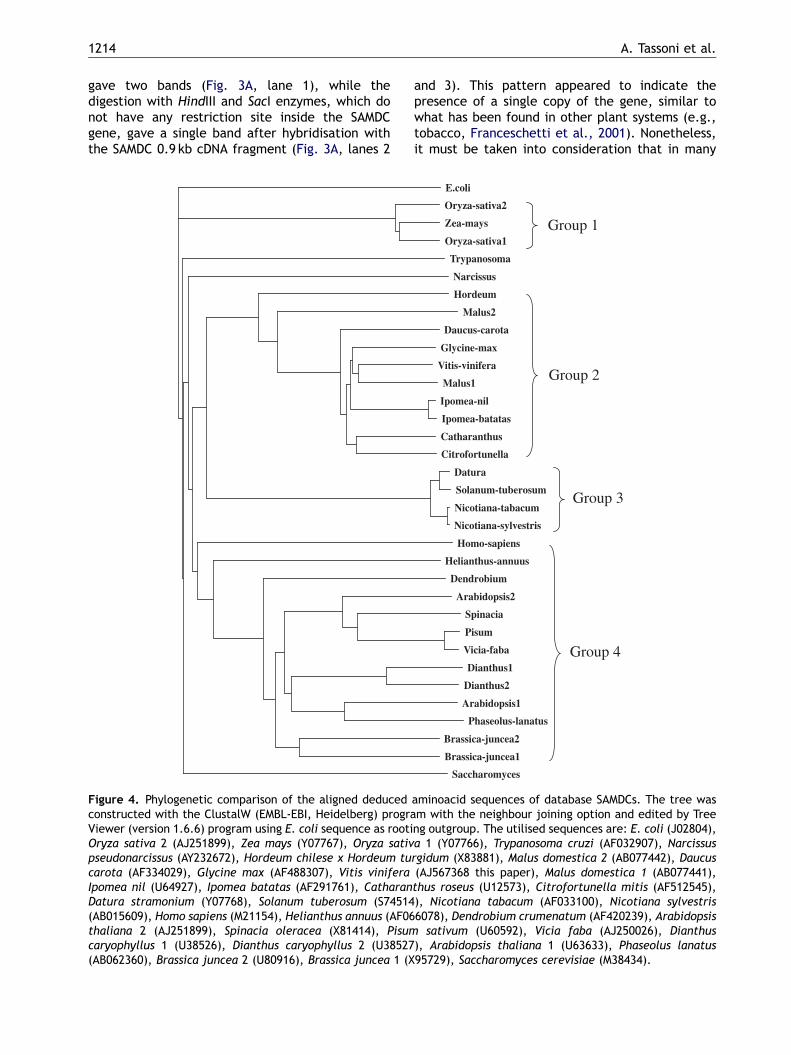
Figure 3. (A–C) Expression analysis of S-adenosylmethionine dhybridised with the grapevine SAMDC 0.9 kb cDNA fragment. (EcoRV (lane 1), HindIII (lane 2) and SacI (lane 3) restriction eThe relative signal amount was obtained after 20 amplificatyoung leaves (SL) and big fully expanded leaves (BL). (C) Densto the actin signal obtained after the same amplification cyc
compared to non-induced ones. The transformantscontaining the empty pYES2 and the non-trans-formed yeasts did not have any significant variationin SAMDC activity after induction (Fig. 2). Theseresults not only demonstrate the identity of thecloned gene, but also confirmed, for the first timein a woody plant, the presence of a regulatingmechanism of the SAMDC gene. Previous reportsshowed that, in the B. juncea SAMDC gene thedisruption of the two ORFs caused an increased ofabout 2-fold of the downstream GUS reporter geneactivity, while when the Arabidopsis SAMDC leadersequence fused to the GUS reporter gene, wasexpressed in yeast, the elimination of both up-stream ORFs increased the relative translation ofabout 2-fold (Hanfrey et al., 2005). Yeast functionalcomplementation experiments hypothesised a pos-sible repressive function of the uORFs on thetranslational efficiency of two apple SAMDCs (Haoet al., 2005). In addition our data show that, whenthe SAMDC assay of FULL and MAIN yeast transfor-mants was performed under inducing conditions inthe presence of 200 mM MGBG, the activity wasreduced in all the clones below the wild-type level,demonstrating the specificity of the measuredactivity (Fig. 2).
For Southern blot analysis the genomic DNA wasdigested with EcoRV, HindIII and SacI restrictionenzymes (Fig. 3A). The restriction with EcoRVenzyme, producing a single cut inside the gene,
P SL BL
SL BL
SAMDC (516 bp)
Actin (331 bp)
ecarboxylase gene in Vitis vinifera. The membranes wereA) Southern blot analysis: genomic DNA was digested withnzymes. (B) RT-PCR analysis of SAMDC expression levels.ion cycles. Apical green branches (B), petioles (P), smallitometric quantification of the RT-PCR analysis normalisedles.
ARTICLE IN PRESS
A. Tassoni et al.1214
gave two bands (Fig. 3A, lane 1), while thedigestion with HindIII and SacI enzymes, which donot have any restriction site inside the SAMDCgene, gave a single band after hybridisation withthe SAMDC 0.9 kb cDNA fragment (Fig. 3A, lanes 2
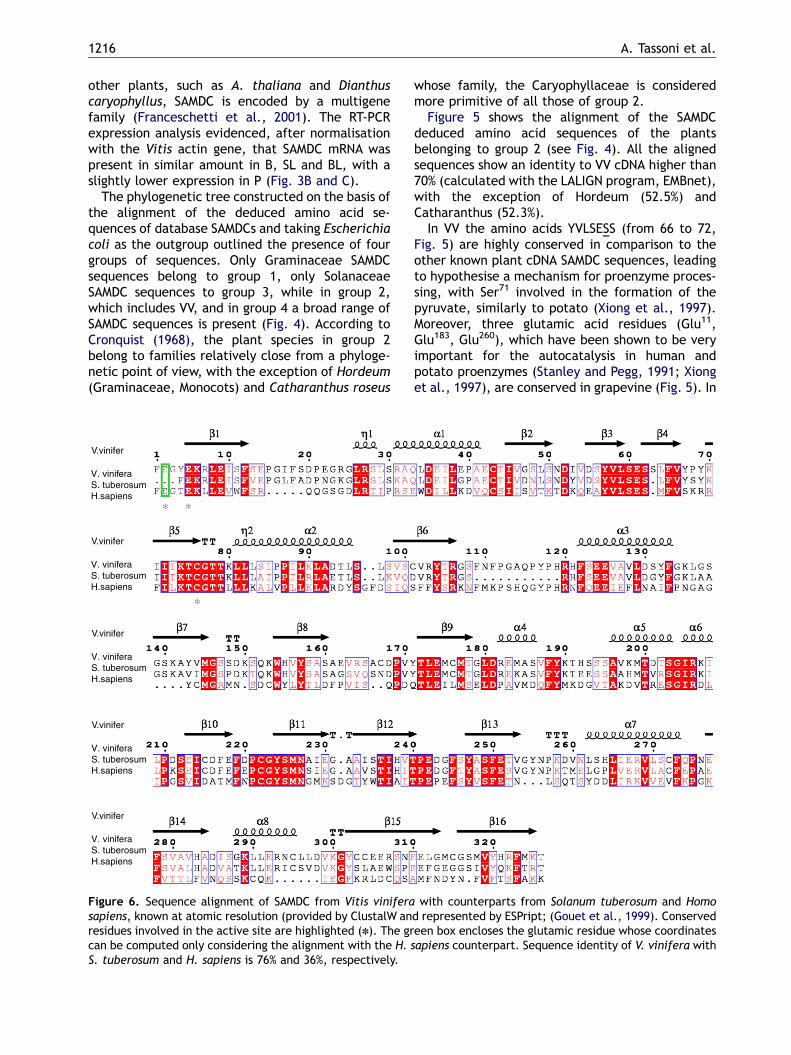
Figure 4. Phylogenetic comparison of the aligned deducedconstructed with the ClustalW (EMBL-EBI, Heidelberg) prograViewer (version 1.6.6) program using E. coli sequence as rootiOryza sativa 2 (AJ251899), Zea mays (Y07767), Oryza sativpseudonarcissus (AY232672), Hordeum chilese x Hordeum tucarota (AF334029), Glycine max (AF488307), Vitis viniferaIpomea nil (U64927), Ipomea batatas (AF291761), CatharanDatura stramonium (Y07768), Solanum tuberosum (S74514(AB015609), Homo sapiens (M21154), Helianthus annuus (AF06thaliana 2 (AJ251899), Spinacia oleracea (X81414), Pisumcaryophyllus 1 (U38526), Dianthus caryophyllus 2 (U38527(AB062360), Brassica juncea 2 (U80916), Brassica juncea 1 (X
and 3). This pattern appeared to indicate thepresence of a single copy of the gene, similar towhat has been found in other plant systems (e.g.,tobacco, Franceschetti et al., 2001). Nonetheless,it must be taken into consideration that in many
E.coli
Oryza-sativa2
Zea-mays
Oryza-sativa1
Trypanosoma
Narcissus
Hordeum
Malus2
Daucus-carota
Glycine-max
Vitis-vinifera
Malus1
Ipomea-nil
Ipomea-batatas
Catharanthus
Citrofortunella
Datura
Solanum-tuberosum
Nicotiana-tabacum
Nicotiana-sylvestris
Homo-sapiens
Helianthus-annuus
Dendrobium
Arabidopsis2
Spinacia
Pisum
Vicia-faba
Dianthus1
Dianthus2
Arabidopsis1
Phaseolus-lanatus
Brassica-juncea2
Brassica-juncea1
Saccharomyces
Group 1
Group 2
Group 3
Group 4
aminoacid sequences of database SAMDCs. The tree wasm with the neighbour joining option and edited by Treeng outgroup. The utilised sequences are: E. coli (J02804),a 1 (Y07766), Trypanosoma cruzi (AF032907), Narcissusrgidum (X83881), Malus domestica 2 (AB077442), Daucus(AJ567368 this paper), Malus domestica 1 (AB077441),thus roseus (U12573), Citrofortunella mitis (AF512545),), Nicotiana tabacum (AF033100), Nicotiana sylvestris6078), Dendrobium crumenatum (AF420239), Arabidopsissativum (U60592), Vicia faba (AJ250026), Dianthus
), Arabidopsis thaliana 1 (U63633), Phaseolus lanatus95729), Saccharomyces cerevisiae (M38434).

ARTICLE IN PRESS
Vitis-vinifera 1 MALPVSAIGFEGYEKRLEISFSEPGIFSDPEGRGLRSLSRAQLDEILEPAECTIVGSLSNDIVDSYVDaucus-carota 1 MSSEVSAIGFEGFEKRLEISFFEPSFFADPEGKGLRVLSKNQLDEFLGPAECTIVASLSNEHVDSYVMalus1 1 MAMEGSAIGFEGYEKRLEIAFFEPSIFLDPEGRGLRSLSKAQIDEFLDQAECTIVSSLSNDDVDSYVIpomea-nil 1 MALSTSAIGFEGYEKRLEISFFEAGIFSDPEGRGLRALSKEQLDKVLKPAECTIVSSLSNNEVDSYVIpomea-batatas 1 MALSTSAIGFEGYENRLEISFFEAGIFSDPEGRGLRALSKEQLDKVLKPAECTIVSSLSNNEVDSYVGlycine-max 1 MAMAVSAIGFEGFEKRLEISFFQPGLFADPEGRGLRALTKSQLGEILTPAACTIVSSLKNDNVDSYVMalus2 1 MAVPVSAIGFEGYEKRLEVSFFEPGLFADPKGMGLRSLSKAQIDEILTPAECTIVSSLSNDDLDSYVCitrofortunella 1 MALPVSAIGFEGYEKRLEVSFFEPGVFADPGGRGLRSLSKHQLDEILKPAECTIVSSLSNEYLDSYVCatharanthus 1 MALPASAIGFEGYEKRLEISFFESSFFADPDGKGLRALNKSQIDEILEPAECTIVDSLSNQYLDSYVHordeum 1 MAAPVSAIGFEGYEKRLEITFSEASIFADPHGRGLRALSRAQIDSVLDLARCTIVSELSNKDFDSYV
Vitis-vinifera 68 LSESSLFVYPYKIIIKTCGTTKLLLSIPPILKLADTLSLSVSCVRYTRGSFNFPGAQPYPHRHFSEDaucus-carota 68 LSESSLFVYAYKIIIKTCGTTKLLKSIPPILKLADSLSLTVRSVRYTRGCFIFPGAQSYPHRSFSEMalus1 68 LSESSLFIHPYKMIIKTCGTTKLLRVIPTVLQLAETISLAVRSVKYTRGSFIFPGAQTYPHRSFSEIpomea-nil 68 LSESSLFVYPYKIIIKTCGTTKLLLSIPPILELADGLSLKVKSVKYTRGSFNFPEVQPYPHRNFSEIpomea-batatas 68 LSESSLFVYPYKIIIKTCGTTKLLLSIPPILELADGLSLKVKSVKYTRGSFNFPEVQPYPHRNFSEGlycine-max 68 LSESSLFVYAYKIIIKTCGTTKLLLAIPPILKFAEMLSLNVKSVNYTRGSFIFPSAQPYPHRNFSEMalus2 68 LSESSLFVYPYKVIIKTCGTTKLLRSIPAILKLADSLSLAVKSVRYSRGSFIFPGAHPSPHRSFSECitrofortunella 68 LSESSLFVYPYKVIIKTCGTTKLLLSIPAILKLAESLSLSVRSVRFTRGSFIFAGAQPFPHRSFSECatharanthus 68 LSESSLFVYPYKIIIKTCGTTKLLLSIPAILKLAESLSLSVRNVKYTRGSFIFPGAQSFPHRSFSEHordeum 68 LSESSLFIYSQKIVIKTCGTTMLLLTIPRILELAEELCMPLAAVKYSRGMFIFPGAQPAPHRSFSE
Vitis-vinifera 134 EVAVLDSYFGKLGSGSKAYVMGSSDKS-QKWHVYSASAEVRSACD------PVYTLEMCMTGLDREDaucus-carota 134 EVSVLDNYFGKLGSGSKAYIMGGSDKQ-QKWHVYSACAASTRTLD------PVYTMEMCMTSLNRDMalus1 134 EVSVLDSYFGKLGSGSRAYVMGRSDGP-QKWHVYSATAESASFVE------PVYTLEMCMTGLDRGIpomea-nil 134 EVAILDGYFGKLGTGSKAYVMGGAGKQ-QQWHVYSASAESAENTF------PIYTLEMCMTGLDKKIpomea-batatas 134 EVAILDSYFGKLSTGSKAYVMGGAGKQ-QQWHVYSASAESAENTF------PIYTLEMCMTGLDKKGlycine-max 134 EVAILDGYFGKLGAGSNAYILGGQDKA-QNWHVYSASADSVTQCD------NVYTLEMCMTGLDREMalus2 134 EVAVLDGHFGKLGLASKAYVMGSPDKT-QKWHIYSASAELASLLWGSRQSGPTYTLEMCMTGLDRKCitrofortunella 134 EVAVLDGHFGKFGMDSTAFVMGSPDNT-KRWHVYSASAEAGSHVN------LVYTLEMCMTGLDRKCatharanthus 134 EVELLDNYFGKLGLESNAFIMGNPDQP-QKWHVYSASVGSEQSSD------PTYTLEMCMTGLDREHordeum 134 EVDVLNRYFGHLNSGGNAYVIGDPAKPGQKWHIYYATEQPEQ---------PMVTLEMCMTGLDKT
Vitis-vinifera 193 MASVFYKTHS----SSAVKMTDTSGIRKILPDSEICDFEFDPCGYSMNAIEGAAISTIHVTPEDGFDaucus-carota 193 KASVFYKTNS----SSATSVTDNSGLRDILPNSRICDFEFDPCGYSMNAVEGPAVSTIHITPEDGFMalus1 193 KASVFYKSES----SSATTMTNESGIRKILPDSDICDFEFDPCGYSMNSIEGAAISTIHVTPEDGFIpomea-nil 193 SASVFFKTQS----SSAAVMTDASGIRKILPGSDICDFDFEPCGYSMNAVEGGTISTIHVTPEDGFIpomea-batatas 193 SASVFFKTQS----SSAAVMTDASGIRKILPGSEICDFDFEPCGYSMNAVEGGAISPIHVTPEDGFGlycine-max 193 KAQVFYKEQS----ASAAMMTVNSGIRKILPDSEICDFDFEPCGYSMNSVEGAAVSTIHVTPEDGFMalus2 199 RASVFYKSDA----SSAAGMTVESGIRNILPQSDICDFEFEPCGYSMNSIEGNAVSTIHVTPEDGFCitrofortunella 193 RASVFYKTNE----SSAAMMTEDSGIRKILPKSEICDFEFDPCGYSMNSIEGDAVSTIHVTPEDGFCatharanthus 193 KASVFYKSES----SSAALMTTRSGIRKILPDSEICDFEFDPCGYSMNSIEEAAISTIHVTPEDGFHordeum 191 KASVFFKTHADGHVSCAKEMTKLSGISDIIPEMEVCDFDFEPCGYSMNAINGSAFSTIHVTPEDGF
Vitis-vinifera 255 SYASFETVGYNPKDVNLSHLIERVLSCFQPNEFSVAVHA-DISGKLLERNCLLDVKGYCCEERSNEDaucus-carota 255 SYSSFEAVGYDPKSVNLSDLVARVLNCFQPGEFSIALQA-DIASELLEKTSSVHVKGYRVEEKTCEMalus1 255 SYASFETAGYDLKDVNLNQLVERVLVCFQPKEFSIAVHA-NVVSKSLQQHSLVDLKGYCREESSHEIpomea-nil 255 SYASFEAMGYDFKDVNLDALIQRVLSCFQPAEFSVALHC-DSIGEKLDSVFELDVKGYACGERSYEIpomea-batatas 255 SYASFEAMGYDFKDVNLDALIQRVLSCFQPAEFSVALHC-DSIGEKLDSVFKLDVKGYACGERSYEGlycine-max 255 SYASFETVGYDFKAVNLNEMVQRVLACFLPTEFSVAVHV-DGASKSFEQTCFLDVKGYCREERSHEMalus2 261 SYASFETVGYDFKDVNLTQLLYRVLDCFKPAEFSIALHTTSTAGEDLDAKCPLDLKGYCCGGSSYECitrofortunella 255 SYASFEAVGYDFEVVKLTSLLERVLACFKPAEFSVALHS-DIVGDEHGDTFTLDFKGYSCGEKIYECatharanthus 255 SYASFEAAGYDLKAQNLGMMIERVLACFQPSEFSVAVHC-DVTCKSLEQICSLELKEYSLDEKINEHordeum 257 SYASYEVQGMDASALAYGDIVKRVLRCFGPSEFSVAVTIFGGRGHAATWGKKLDAEAYDCNNVVEQ
Vitis-vinifera 320 ELGMCGSMVYHRFMKT----EGLVSPRSILKC--------------------CWKEEEEEEKE---Daucus-carota 320 ELGMDGSIVYQKFVKTT---ERCESPRSVLKC--------------------CWKEEEKEEKEYQ-Malus1 320 ELGLGGAIVYQRFLKT----ERCGSPRSTLKG--------------------CWREEE-EEEDY--Ipomea-nil 320 ALGKGGSIMYCGFTST----GSCGSPRSTLLC--------------------CWSENEDQEGEKKHIpomea-batatas 320 GLNKGGSIMYCGFTST----GSCGSPRSTLLC--------------------CWSENEDEEGEKKHGlycine-max 320 GLGMGGSVVYQKFGKT----SDCGSPRSTLK---------------------CWNEEDEEE-----Malus2 327 GLGLGGAVMYHSFVKDD---SGSQSPRSILKC--------------------CWSEDEKDEEVEEICitrofortunella 320 ELGNNGSLIYYSFSRTD---C-STSPRSILKC--------------------CWSEDEKDEEVEGKCatharanthus 320 ELGLGGSIIYKKFLRID---A-CGSPRSILKC--------------------CWKEDESEEE----Hordeum 323 ELPCGGVLIYQSFAANEELAVSAGSPRSVFHCFENVESGHPLVKEGKLANLLAWRAEEESLEEGTG
Figure 5. Alignment of deduced aminoacid sequences of main ORF of SAMDC cDNAs of plants belonging to the group 2indicated in Fig. 3. The alignment was performed with ClustalW program (EMBL-EBI, Heildelberg) and edited with theBoxShade program (version 3.21, EMBnet, Switzerland). Conserved glutamic acid residues necessary for the stimulationof proenzyme processing of the human SAMDC are indicated by filled circles. Location of the proenzyme-processing siteis indicated by the filled triangle. The open square indicates the Vitis valine residue substituting the aspartic acidresidue present in the human enzyme and necessary for the stimulation of enzyme processing (Xiong et al., 1997).
Grapevine S-adenosylmethionine decarboxylase 1215
ARTICLE IN PRESS
A. Tassoni et al.1216
other plants, such as A. thaliana and Dianthuscaryophyllus, SAMDC is encoded by a multigenefamily (Franceschetti et al., 2001). The RT-PCRexpression analysis evidenced, after normalisationwith the Vitis actin gene, that SAMDC mRNA waspresent in similar amount in B, SL and BL, with aslightly lower expression in P (Fig. 3B and C).
The phylogenetic tree constructed on the basis ofthe alignment of the deduced amino acid se-quences of database SAMDCs and taking Escherichiacoli as the outgroup outlined the presence of fourgroups of sequences. Only Graminaceae SAMDCsequences belong to group 1, only SolanaceaeSAMDC sequences to group 3, while in group 2,which includes VV, and in group 4 a broad range ofSAMDC sequences is present (Fig. 4). According toCronquist (1968), the plant species in group 2belong to families relatively close from a phyloge-netic point of view, with the exception of Hordeum(Graminaceae, Monocots) and Catharanthus roseus
* *
*
V.vinifer
V. viniferaS. tuberosumH.sapiens
V.vinifer
V. viniferaS. tuberosumH.sapiens
V.vinifer
V. viniferaS. tuberosumH.sapiens
V.vinifer
V. viniferaS. tuberosumH.sapiens
V.vinifer
V. viniferaS. tuberosumH.sapiens
Figure 6. Sequence alignment of SAMDC from Vitis viniferasapiens, known at atomic resolution (provided by ClustalW anresidues involved in the active site are highlighted (n). The grcan be computed only considering the alignment with the H. sS. tuberosum and H. sapiens is 76% and 36%, respectively.
whose family, the Caryophyllaceae is consideredmore primitive of all those of group 2.
Figure 5 shows the alignment of the SAMDCdeduced amino acid sequences of the plantsbelonging to group 2 (see Fig. 4). All the alignedsequences show an identity to VV cDNA higher than70% (calculated with the LALIGN program, EMBnet),with the exception of Hordeum (52.5%) andCatharanthus (52.3%).
In VV the amino acids YVLSESS (from 66 to 72,Fig. 5) are highly conserved in comparison to theother known plant cDNA SAMDC sequences, leadingto hypothesise a mechanism for proenzyme proces-sing, with Ser71 involved in the formation of thepyruvate, similarly to potato (Xiong et al., 1997).Moreover, three glutamic acid residues (Glu11,Glu183, Glu260), which have been shown to be veryimportant for the autocatalysis in human andpotato proenzymes (Stanley and Pegg, 1991; Xionget al., 1997), are conserved in grapevine (Fig. 5). In
with counterparts from Solanum tuberosum and Homod represented by ESPript; (Gouet et al., 1999). Conservedeen box encloses the glutamic residue whose coordinatesapiens counterpart. Sequence identity of V. vinifera with

ARTICLE IN PRESS
Grapevine S-adenosylmethionine decarboxylase 1217
fact, processing and activity of the mammalian andyeast SAMDCs is stimulated by the presence ofputrescine, which interacts with the three con-served Glu residues together with a fourth residue(Asp174 in human SAMDC) activating the autocata-lytic reaction (Stanley et al., 1994). However, plantSAMDC proenzyme processing has been reportednot to be activated by putrescine (Xiong et al.,1997) due to the absence of this fourth residue.Confirming this particular feature in plants, VVenzyme lacks of the Asp residue that is substituted,as in potato, by a valine (Val179) (Fig. 5).
The SAMDC protein chain of VV can be alignedwith the two structures of the enzyme counterpartspresently known at atomic resolution: that from STand that from HS. The result of the alignment isshown in Fig. 6. VV sequence identity with ST(475%) is higher than with HS (435%) and for thisreason the ST structure is the natural templatewhen homology building of VV is considered.However, as shown in Fig. 6, the structure of the
NC
V.vinifera
S.tuberosum
H.sapiens
A
B
Figure 7. (A–C) 3D model of the SAMDC from V. vinifera ((A) Cartoon representation provided by YASARA moleculasuperposition of SAMDC from V. vinifera (blue, VV), S. tuberesidues involved in the active site of the three structures (V
N-terminal portion of ST sequence, is lacking (notsolved at atomic resolution) of one of the twoglutamic acid residues that take part together witha cysteine to the enzyme active site (green box inFig. 6). This residue is present in the HS counter-part and therefore, in order to model the activesite of VV, we included in the set of putativetemplates also the HS structure.
The VV SAMDC computed model is shown inFig. 7A. The first nine residues of the VV chaincannot be modelled due to lack of the correspond-ing 3D coordinates in the templates. The root-mean-square deviation (RSMD) calculated valueconfirmed that the VV structure is rather similarto those of the original templates (VV/ST:RSMD ¼ 0.3 (A; VV/HS: RSMD ¼ 1.9 (A, Fig. 7B) andsimilarly belongs to the alpha and beta class,according to the SCOP classification (Murzinet al., 1995). In Fig. 7C the compute active site ishighlighted and compared both to that of ST andthat of HS. Noticeably, only the presence of HS
N C
C76
E2
E5
C
VV), computed by homology modelling (Modeller v7.7).r visualisation tool (http://www.yasara.org); (B) therosum (green, ST) and H. sapiens (red, HS) and (C) theV/ST: RSMD ¼ 0.3 (A; VV/HS: RSMD ¼ 1.9 (A).

ARTICLE IN PRESS
S113
F111
F285
D174
T176
E256 S254
A243E11
L13
K59
R103
H283
V170
T172 E251
S249
H238
K74
E5
E9
T105R7
Figure 8. Comparison of the Vitis vinifera (blue) and Homo sapiens (red) buried charged site of SAMDC. The Vitisvinifera site (model shown in Fig. 7B) is represented together with the same site of Solanum tuberosum (green). Thetwo sequences are endowed with the same residues in the site.
A. Tassoni et al.1218
counterpart allows the calculation of the enzymeactive site in VV.
Our model shows that the buried charge site isvery similar to that of potato but different fromthat reported for the human species (Fig. 8).Ekstrom et al. (2001) reported that in the humanstructure this site is the putrescine binding site,due to its excess of negative charges. On thecontrary, in the potato enzyme, substitutionof Leu13/Arg18 (human/potato), Phe111/Arg114,Asp174/Val181 and Phe285/His294 hamper putrescinebinding (Bennett et al., 2002). The high level oflocal similarity between VV and potato in theburied charge site (Fig. 8), justifies the lack ofputrescine binding in the VV enzyme. In fact, thesubstitutions Leu13/Arg7 (human, Vitis), Phe111/Arg103, Asp174/Val170 and Phe285/His283 alter thenet charge in the VV when compared to the humanstructure (Stanley et al., 1994). This is sufficient tohamper putrescine binding and therefore, contra-rily to what shown in humans (Stanley and Pegg,1991), the Vitis SAMDC activity, as well as theenzyme processing, does not require the presenceof putrescine. This hypothesis was confirmed byexperimental data demonstrating the possibility tomeasure SAMDC activity without the addition ofexogenous putrescine both when the Vitis SAMDCwas expressed in yeast (Fig. 2) or when the activitywas measured in different grape extracts (Bertoldiet al., 2004). Similar results were also obtainedin many other plant systems among which areA. thaliana and tobacco (Tassoni et al., 2000;Franceschetti et al., 2004).
Acknowledgements
We wish to thank Prof. Giovanni Cristofolini(Department of Biology e.s., University of Bologna)
for his valuable help in discussing the phylogeneticaspects of the manuscript, Prof. Peter Davies(Cornell University, Ithaca, USA) for critical editingof the manuscript and Dr. Colin Hanfrey (Instituteof Food Research, Norwich, UK) for providing theyeast strain. This research was supported by the ex60% funds from MURST (Ministry of University,Science and Technological Research) of Italy toNello Bagni. G. Tasco is the recipient of a MURSTfellowship for the Functional Genomics Project.
References
Bagni N, Tassoni A. Biosynthesis, oxidation and conjuga-tion of aliphatic polyamines in higher plants. AminoAcids 2001;20:301–17.
Bennett EM, Ekstrom JL, Pegg AE, Ealick SE. MonomericS-adenosylmethionine decarboxylase from plants pro-vides an alternative to putrescine stimulation. Bio-chemistry 2002;41:14509–17.
Bertoldi D, Tassoni A, Martinelli L, Bagni N. Polyaminesand somatic embryogenesis in two Vitis viniferacultivars. Physiol Plant 2004;120:657–66.
Chang S, Puryear J, Cairney J. A simple and efficientmethod for isolating RNA from pine trees. Plant MolRep 1993;11:113–6.
Cronquist A. The evolution and classification of floweringplants. London, Edinburgh: Houghton Mifflin; 1968.
Ekstrom JL, Tolbert WD, Xiong H, Pegg AE, EalickSE. Structure of a human S-adenosylmethioninedecarboxylase self-processing ester intermediate andmechanism of putrescine stimulation of processing asrevealed by the H243A mutant. Biochemistry 2001;40:9495–504.
Elbe R. A simple and efficient procedure for transforma-tion of yeasts. Biotechniques 1992;13:18–22.
Franceschetti M, Hanfrey C, Scaramagli S, Torrigiani P,Bagni N, Burtin D, et al. Characterization of monocotand dicot plant S-adenosyl-l-methionine decarboxy-lase gene families including identification in the mRNA

ARTICLE IN PRESS
Grapevine S-adenosylmethionine decarboxylase 1219
of a highly conserved pair of upstream overlappingopen reading frames. Biochem J 2001;353:403–9.
Franceschetti M, Fornale S, Tassoni A, Zuccherelli K,Mayer MJ, Bagni N. Effects of spermidine synthaseoverexpression on polyamine biosynthetic pathway intobacco plants. J Plant Physiol 2004;161:989–1001.
Gouet P, Courcelle E, Stuart D, Metoz F. ESPript: multiplesequence alignments in PostScript. Bioinformatics1999;15:305–8.
Hanfrey C, Franceschetti M, Mayer MJ, Illingworth C,Michael AJ. Abrogation of upstream open readingframe-mediated translational control of a plantS-adenosylmethionine decarboxylase results in poly-amine disruption and growth perturbations. J BiolChem 2002;277:44131–9.
Hanfrey C, Elliott KA, Franceschetti M, Mayer MJ,Illingworth C, Michael AJ. A dual upstream openreading frame-based autoregulatory circuit control-ling polyamine-responsive translation. J Biol Chem2005;280:39229–37.
Hao YJ, Zhang Z, Kitashiba H, Honda C, Ubi B, Kita M,Moriguchi T. Molecular cloning and functional char-acterization of two apple S-adenosylmethionine de-carboxylase genes and their different expression infruit development, cell growth and stress response.Gene 2005;350:41–50.
Hu WW, Gong H, Pua EC. The pivotal roles of the plantS-adenosylmethionine decarboxylase 50 untranslatedleader sequence in regulation of gene expression atthe transcriptional and posttranscriptional levels.Plant Physiol 2005;138:276–86.
Kashiwagi K, Taneja SK, Liu TY, Tabor CW, TaborH. Spermidine biosynthesis in Saccharomyces cerevi-siae. Biosynthesis and processing of a proenzyme formof S-adenosylmethionine decarboxylase. J Biol Chem1990;265:22321–8.
Kumar A, Altabella T, Taylor M, Tiburcio AF. Recentadvances in polyamine research. Trends Plant Sci1997;2:124–30.
Laskowski R, MacArthur M, Moss D, Thornton J. PRO-CHECK: a program to check the stereochemical qualityof protein structures. J Appl Crystallogr 1993;26:283–91.
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Proteinmeasurement with the Folin phenol reagent. J BiolChem 1951;193:265–75.
Marti-Renom M, Stuart A, Fiser A, Sanchez R, Melo F, SaliA. Comparative protein structure modelling of genes
and genomes. Ann Rev Biophys Biomol Struct 2000;29:291–325.
Michael AJ, Furze JM, Rhodes MJ, Burtin D. Molecularcloning and functional identification of a plantornithine decarboxylase cDNA. Biochem J 1996;314:241–8.
Murzin A, Brenner S, Hubbard T, Chothia C. SCOP: astructural classification of proteins database for theinvestigation of sequences and structures. J Mol Biol1995;247:536–40.
Sambrook J, Fritsch EF, Maniatis T. Molecular cloning: alaboratory manual. Cold Spring Harbor, NY: Cold SpringHarbor Laboratory Press; 1989.
Sippl M. Recognition of errors in three-dimensionalstructures of proteins. Proteins 1993;17:355–62.
Stanley BA, Pegg AE. Amino acid residues necessary forputrescine stimulation of human S-adenosylmethio-nine decarboxylase proenzyme processing and cataly-tic activity. J Biol Chem 1991;266:18502–6.
Stanley BA, Shantz LM, Pegg AE. Expression of mamma-lian S-adenosylmethionine decarboxylase in E. coli.Determination of sites for putrescine activation ofactivity and processing. J Biol Chem 1994;269:7901–7.
Tassoni A, van-Buuren M, Franceschetti M, Fornale S,Bagni N. Polyamine content and metabolism inArabidopsis thaliana and effect of spermidine onplant development. Plant Physiol Biochem 2000;38:383–93.
Thompson J, Higgins D, Gibson T. CLUSTAL W: improvingthe sensitivity of progressive multiple sequencethrough sequence weighting, position-specific gappenalties and weight matrix choice. Nucleic AcidsRes 1994;22:4673–80.
Tiburcio AF, Altabella T, Borrell A, Masgrau C. Polyaminemetabolism and its regulation. Physiol Plant 1997;100:664–74.
Wells G, Birkholtz L, Joubert F, Walter R, Louw A. Novelproperties of malarian S-adenosylmethionine decar-boxylase as revealed by structural modeling. J MolGraph Model 2006;24:307–18.
Wrenger C, Luersen K, Krause T, Muller S, Walter RD. ThePlasmodium falciparum bifunctional ornithine decar-boxylase, S-adenosyl-L-methionine decarboxylase, en-ables a well balanced polyamine synthesis withoutdomain-domain interaction. J Biol Chem 2001;276:29651–6.
Xiong H, Stanley BA, Tekwani BL, Pegg AE. Processing ofmammalian and plant S-adenosylmethionine decar-boxylase proenzymes. J Biol Chem 1997;272:28342–8.




















