
Expression of arginine decarboxylase is induced during early fruit development and in young tissues...
13
Plant Molecular Biology 28: 997-1009, 1995. © 1995 Kluwer Academic Publishers. Printed in Belgium. 997 Expression of arginine decarboxylase is induced during early fruit development and in young tissues of Pisum sativum (L.) Miguel A. Prrez-Amador, Juan Carbonell and Antonio Granell* Departamento de Biologia del Desarrollo de Plantas, Instituto de Biolog[a Molecular y Celular de Plantas, UPVA-CSIC, Camino de Vera 14, 46022-Valencia, Spain (*author for correspondence) Received 31 January 1995; accepted in revised form 30 May 1995 Key words: arginine decarboxylase (ADC), fruit development, pea ovary, Pisum sativum, polyamine, putrescine Abstract A cDNA coding for arginine decarboxylase (ADC, EC 4.1.1.19) has been isolated from a cDNA library of parthenocarpic young fruits of Pisum sativum (L.). The deduced aminoacid sequence is 74?/0, 46?0 and 35?0 identical to ADCs from tomato, oat and Escherichia coli, respectively. When the pea ADC cDNA was put under the control of the galactose inducible yeast promoter CYC1-GAL10 and intro- duced into Saccharomyces cerevisiae, it conferred galactose-regulated expression of the ADC activity. The ADC activity expressed in S. cerevisiae was inhibited 99?0 by ~-DL-difluoromethylarginine (DFMA), a specific inhibitor of ADC activity. No activity was detected in the untransformed S. cerevisiae, nor when it was transformed with an antisense ADC construct. This provides direct evidence that the ADC cDNA from pea encoded a functional, specific ADC activity and that S. cerevisiae is able to process correctly the protein. In the pea plant, gene expression of the ADC is high in young developing tissues like shoot tips, young leaflets and flower buds. Fully expanded leaflets and roots have much lower, but still detectable, levels of the ADC transcript. In the ovary and fruit, they are developmentally regulated, showing high levels of expression during the early stages of fruit growth, which in pea is mainly due to cell expansion. The observed changes in the steady-state levels of ADC mRNA alone, however, can- not account for the differences in ADC activity suggesting that other regulatory mechanisms must be acting. Introduction Arginine decarboxylase (ADC, EC 4.1.1.19) is a key enzyme in the synthesis of polyamines in plants and bacteria. It catalyzes the synthesis of agmatine, an intermediate in the production of putrescine. Putrescine and the polyamines de- rived from it are ubiquitous in all type of organ- isms, including bacteria, yeast, animals and plants [44]. In plants these polycations have been im- The nucleotide sequence data reported will appear in the EMBL, GenBank and DDBJ Nucleotide Sequence Data banks under the accession number Z37540.
Transcript of Expression of arginine decarboxylase is induced during early fruit development and in young tissues...

Plant Molecular Biology 28: 997-1009, 1995. © 1995 Kluwer Academic Publishers. Printed in Belgium. 997
Expression of arginine decarboxylase is induced during early fruit development and in young tissues of Pisum sativum (L.)
Miguel A. Prrez-Amador, Juan Carbonell and Antonio Granell* Departamento de Biologia del Desarrollo de Plantas, Instituto de Biolog[a Molecular y Celular de Plantas, UPVA-CSIC, Camino de Vera 14, 46022-Valencia, Spain (*author for correspondence)
Received 31 January 1995; accepted in revised form 30 May 1995
Key words: arginine decarboxylase (ADC), fruit development, pea ovary, Pisum sativum, polyamine, putrescine
Abstract
A cDNA coding for arginine decarboxylase (ADC, EC 4.1.1.19) has been isolated from a cDNA library of parthenocarpic young fruits of Pisum sativum (L.). The deduced aminoacid sequence is 74?/0, 46?0 and 35?0 identical to ADCs from tomato, oat and Escherichia coli, respectively. When the pea ADC cDNA was put under the control of the galactose inducible yeast promoter CYC1-GAL10 and intro- duced into Saccharomyces cerevisiae, it conferred galactose-regulated expression of the ADC activity. The ADC activity expressed in S. cerevisiae was inhibited 99?0 by ~-DL-difluoromethylarginine (DFMA), a specific inhibitor of ADC activity. No activity was detected in the untransformed S. cerevisiae, nor when it was transformed with an antisense ADC construct. This provides direct evidence that the ADC cDNA from pea encoded a functional, specific ADC activity and that S. cerevisiae is able to process correctly the protein. In the pea plant, gene expression of the ADC is high in young developing tissues like shoot tips, young leaflets and flower buds. Fully expanded leaflets and roots have much lower, but still detectable, levels of the ADC transcript. In the ovary and fruit, they are developmentally regulated, showing high levels of expression during the early stages of fruit growth, which in pea is mainly due to cell expansion. The observed changes in the steady-state levels of ADC mRNA alone, however, can- not account for the differences in ADC activity suggesting that other regulatory mechanisms must be acting.
Introduction
Arginine decarboxylase (ADC, EC 4.1.1.19) is a key enzyme in the synthesis of polyamines in plants and bacteria. It catalyzes the synthesis of
agmatine, an intermediate in the production of putrescine. Putrescine and the polyamines de- rived from it are ubiquitous in all type of organ- isms, including bacteria, yeast, animals and plants [44]. In plants these polycations have been im-
The nucleotide sequence data reported will appear in the EMBL, GenBank and DDBJ Nucleotide Sequence Data banks under the accession number Z37540.

998
plicated in a range of developmental processes, including cell division, embryogenesis, root and flower development and leaf senescence, although their role in these processes remains unclear [ 12].
Putrescine can also be synthesized from orni- thine in a reaction catalyzed by ornithine decar- boxylase (ODC, EC 4.1.1.17). While ADC seems to be restricted to plants and bacteria, ODC is widespread in all living organisms. Both ADC and ODC seem to be functional in higher plants, although the plant seems to use one, the other or both in different situations. Thus, while ODC ap- pears to be associated with actively dividing cells in meristematic and reproductive tissues, ADC would be the primary source of putrescine during cell elongation and in the synthesis of secondary metabolites [44].
Activity of both ODC and ADC have been found to correlate with growth rates during fruit development in mandarin orange [29], tobacco [39], tomato [8, 16, 42] and avocado [48]. In pea, an increase in putrescine content and a tran- sient increase in ADC activity was detected after gibberellic acid (GA3) induction of fruit develop- ment in unpollinated ovaries [6, 32]. This increase in ADC activity was associated to rapid enlarge- ment of mesocarp cells [45]. In untreated unpol- linated ovaries ADC activity decreased and was nearly zero when the ovaries started to senesce. ODC activity was in all stages below detection. These results indicate that the synthesis of polyamines is regulated at the level of ADC in unpollinated ovaries during the two alternative pathways of their development: fruit set and ovary senescence.
Studies on the expression of ADC and other polyamine biosynthetic enzymes in plants are scarce. ADC has been cloned from oat leaf [1] and tomato pericarp [36]. Two genes coding re- spectively for a biosynthetic and a biodegradative form of ADC have also been cloned from E. coli [27, 40]. Biosynthetic ADC from E. coli and plant ADCs share a high degree of sequence similarity, whereas this is not so for E. coli biodegradative ADC. Other genes involved in polyamine meta- bolism have been recently isolated from plants: S-adenosylmethionine synthetase from A rabidop-
sis thaliana [31], carnation petal [24], parsley [21], poplar leaf [10] and tomato seedling [ 11 ], and S-adenosylmethionine decarboxylase (SAMDC) from potato stolon [41, 25]. To the best of our knowledge, ODC, spermidine and spermine synthase from plants remain to be cloned.
In this work we report the isolation, character- ization and expression of a cDNA encoding a functional ADC from pea. The heterologous ex- pression of this cDNA in S. cerevisiae gave high levels of ADC activity in this otherwise ADC- organism. We also present the pattern of ADC expression during ovary development and senes- cence, early stages of fruit development and in actively growing and fully developed tissues of the pea plant.
Materials and methods
Plant material Growth conditions and treatments
Pea (Pisum sativum (L.) cv. Alaska) plants were grown as previously described [5]. Only the first and/or second flower in each plant were used for experiments. Unpollinated pea ovaries were ob- tained by emasculation of the flowers (petals and stamens were removed) two days before anthesis. Parthenocarpic fruits were obtained by appli- cation of 20 #1 of either 0.33 mM GA3 (Fluka, Buchs Switzerland), 0.44 mM benzyladenine (BA, Fluka) or 0.045 mM 2,4-dichlorophenoxy- acetic acid (2,4-D, Fluka) to unpollinated ova- ries. All the solutions contained 0.1 ~o Tween 80. Flowers were treated on the day of anthesis or two days later. Samples were collected, weighed and immediately frozen in liquid nitrogen and stored at -80 °C.
Construction and screening of a pea young fruit cDNA library. Cloning and sequencing of pEADC311 and pEADC621
Poly(A) + RNA from unpollinated pea ovaries, treated at day two after anthesis with G A 3 and

harvested 12 h later, was purified by guanidinium thioisocyanate/N-lauroylsarcosine extraction and oligo-dT chromatography using the QuickPrep Micro mRNA purification kit (Pharmacia Bio- tech AB, Uppsala, Sweden). cDNA was synthe- sized using the UNI-Zap XR cDNA synthesis kit (Stratagene, La Jolla, CA), size-fractionated by Sephacryl S-400 (Pharmacia) and selected frac- tions cloned into the Eco RI-Xho I sites of the Lambda ZAP II vector (Stratagene). Recombi- nant DNA was packaged in vitro using Gigapack Gold extracts (Stratagene). Two cDNA libraries were obtained and one of them (2G212h-F2) was plate-amplified once.
1 x 10 s pfu from the amplified cDNA library were screened by standard plaque lift methods [37], using as a probe the ADC cDNA from to- mato pericarp [36]. The probe was random prime radiolabeled with [~-32p]dCTP using the Ready to Go DNA labeling kit (Pharmacia) and purified by Sephadex G50 chromatography (Boehringer- Mannheim Biochemica, Mannheim, Germany). Filters were prehybridized at 38 °C for 6-8 h in 50~o formamide, 6x SSPE, 0.5~o SDS, 5x Denhardt's solution and 250 #g/ml denatured herring sperm DNA. Hybridization was carried out overnight at 38 °C with the same solution. Filters were washed twice in 1 x SSC and 0.1~o SDS at room temperature for 30 min and twice in0.1 x SSCand0.1~o SDS at40 °Cfor 15 min, and exposed to X-Omat SX film (Kodak, Ger- many) with an intensifying screen at -80 ° C.
Positive plaques obtained from each screening round were used to make small-scale liquid ly- sates and analyzed by PCR as follows. XLR-Blue MRF' strain of E. coli was infected with positive clones and grown at 37 °C for 10-14 h. 1 #1 of a 1:100 dilution of this culture was used for a PCR assay using vector primers. Total PCR products were analyzed by Southern blot and hybrid- ized with the same probe as used for screening. Three rounds of plaque purification were per- formed. Positive clones were excised in vivo from phagemids in Bluescript I SK using the biologi- cal rescue system from Stratagene. Clones pEADC311 and pEADC621 were selected for further analysis.
999
DNA sequencing was conducted according to the dideoxy chain termination method [38] for double-stranded DNA using the Sequenase ver- sion 2.0 DNA Sequencing kit (U.S. Biochemi- cals). Nested unidirectional deletions, in 5' and 3' directions, were generated by the ExolII/S1 method (Erase-a-base System, Promega). Se- quence analysis and computation were carried out with the Genetics Computer Group (GCG, WI) package or locally with the McVector soft- ware for Macintosh.
Expression of pea ADC cDNA in Saccharomyces cerevisiae: ADC activity
Plasmid pEADC311 was linearized at the 3' end by Xho I digestion and made blunt-ended with Klenow and dNTPs. The linearized plasmid was digested with Dra I and the Dra I/Xho 1-blunt fragment generated was purified by agarose gel electrophoresis and DEAE-cellulose. The pEMBL-yex4 expression vector [7] was linear- ized by Sma I digestion at the multicloning site and ligated with the Dra I/Xho I fragment of pEADC311. Transformation was carried out in E. coli DH5~ strain and recombinants were se- lected by restriction analysis. Sense (pYex4- ADC6) and antisense (pYex4-ADCS) plasmids were purified using Magic Minipreps DNA Purification Columns (Promega).
S. cerevisiae strain ENY.WA-1A (MAT~ ura3-52 leu2-3,112 trp1-285 his3-A1) was used for transformation with pYex4-ADC6 and pYex4- ADCS, as well as with the original plasmid pEMBL-yex4. Yeast transformation was carried out by alkali treatment [18] using 2 #g plasmid DNA. Transformants were selected at 30 °C in minimal medium (0.7~o Yeast Nitrogen Base without amino acids, Difco) plates, supplemented with 2~o glucose (MMGlu) or 4~o galactose (MMGal) as carbon source. In all cases, plates and liquid media were supplemented with histi- dine, leucine and tryptophane at 20, 30 and 20/~g/ml, respectively.
Yeast transformants harboring each of the plasmids pEMBL-yex4, pYex4-ADC6 and

1000
pYex4-ADC8 were grown in liquid MMGlu for 8 h at 30 °C. Cells were collected by centri- fugation at 3500 x g for 5 min, rinsed several times with distilled water, resuspended in liquid MMGIu and MMGal at OD6o o 0.2-0.3 and in- cubated for 6 - 2 0 h at 30 °C. When OD600
reached twice the initial, 10 ml culture was cen- trifuged at 3500 × g for 5 min and the pellet was rinsed once in 100 mM potassium phosphate pH 7.0. After centrifugation, the cells were ho- mogenized by vortexing in 500/zl cold homog- enization buffer (100 mM potassium phosphate pH 7.0, 1 mM PMSF, 7 mM benzamidine, 10 mM 2-mercaptoethanol, 10 #M leupeptin, 150 #M pyridoxal phosphate) with glass beads (0.45 mm diameter) for five periods of 1 min with intervals of 1 min on ice. Crude extracts were clarified by centrifugation at 6500 x g for 5 min at 4 ° C. Supernatant was used for enzyme assays. ADC activity was determined as previously de- scribed [32] by measuring the amount of labeled CO2 liberated from L-[U-14C]Arg (Amersham, 305 mCi/mmol). DFMA (Marion Merrell Dow Research Institute, Cincinnati, OH), an irrevers- ible inhibitor of ADC activity, was used to deter- mine the specificity of the decarboxylation reac- tion. Extracts were preincubated in the dark with 2 mM DFMA for 30 min at room temperature and assayed as described before. Protein was de- termined by the Bradford method [3] using BSA as standard.
Analysis of ADC mRNA levels
RNA was extracted as previously described [ 19]. Plant tissues were ground with liquid nitrogen in a mortar. The powder was homogenized in ex- traction buffer (10 ml/g tissue) containing 5 M guanidinium thioisocyanate, 0.5 ~o N-lauroylsar- cosine, 25 mM sodium citrate pH 7.0, 2 mM EDTA and 0.1 M 2-mercaptoethanol. Total RNA was extracted by phenol/chloroform/ isoamyl alcohol and sodium acetate and lithium chloride precipitation. Poly(A) + RNA was ex- tracted with the QuickPrep Micro mRNA Puri- fication kit (Pharmacia). Denatured RNA was
fractionated in 1.2 ~o agarose-formaldehyde gels and transferred in 10 × SSC onto nylon mem- branes (Hybond-N, Amersham) according to standard methods [37]. Prior to blotting, the gels were stained with ethidium bromide to ensure that equal amounts of RNA were loaded. RNA was cross-linked to Hybond-N membrane by UV irradiation (1.2× 105 J) supplied by UV Stratalinker 1800 (Stratagene). Filters were hy- bridized at 42 °C in 50 ~o formamide hybridiza- tion solution, similar as for cDNA library screen- ing. pEADC311 was radiolabeled by the random primer method as described above. Washes were four times in 1 × SSC and 0.1~o SDS at room temperature for 5 min and twice in 0.1 × SSC and 0.1~o SDS at 55-60 °C for 15 min.
Results
Isolation and sequence analysis of pea ADC cDNA clones
To isolate cDNA clones for pea ADC, two cDNA libraries, 2G2~2h-F2 and 2G212h-F3, were made from poly(A) + RNA extracted from unpollinated pea ovaries 12 h after GA3 treatment. At this stage, ovaries showed a strong induction in ADC activity [32]. Titers for 2G212h-F2 and 2G212h-F3 libraries were 5 x 105 and 1 × 10 6 primary recom- binants, respectively. The average size of the in- serts was 2.2 kb with < 1 ~o ofnon-recombinants. The 2G212h-F2 library was amplified once and screened using as probe the cDNA for the tomato pericarp ADC clone [36]. As a result two clones, pEADC311 of 3.2 kb and pEADC621 of 2.0 kb, were isolated in Bluescript I SK by in vivo exci- sion.
Figure 1 shows the nucleotide and deduced amino acid sequence ofpEADC311. The insert of pEADC311 is 3161 bp long with an open reading frame (ORF) from position 558 to 2741, encod- ing a polypeptide of 728 amino acids with a pre- dicted molecular mass of 78.7 kDa. The 5'- untranslated region (UTR) of this cDNA is 557 bp long, ranging well over the average size of the plant leader sequences [20]. This untrans-
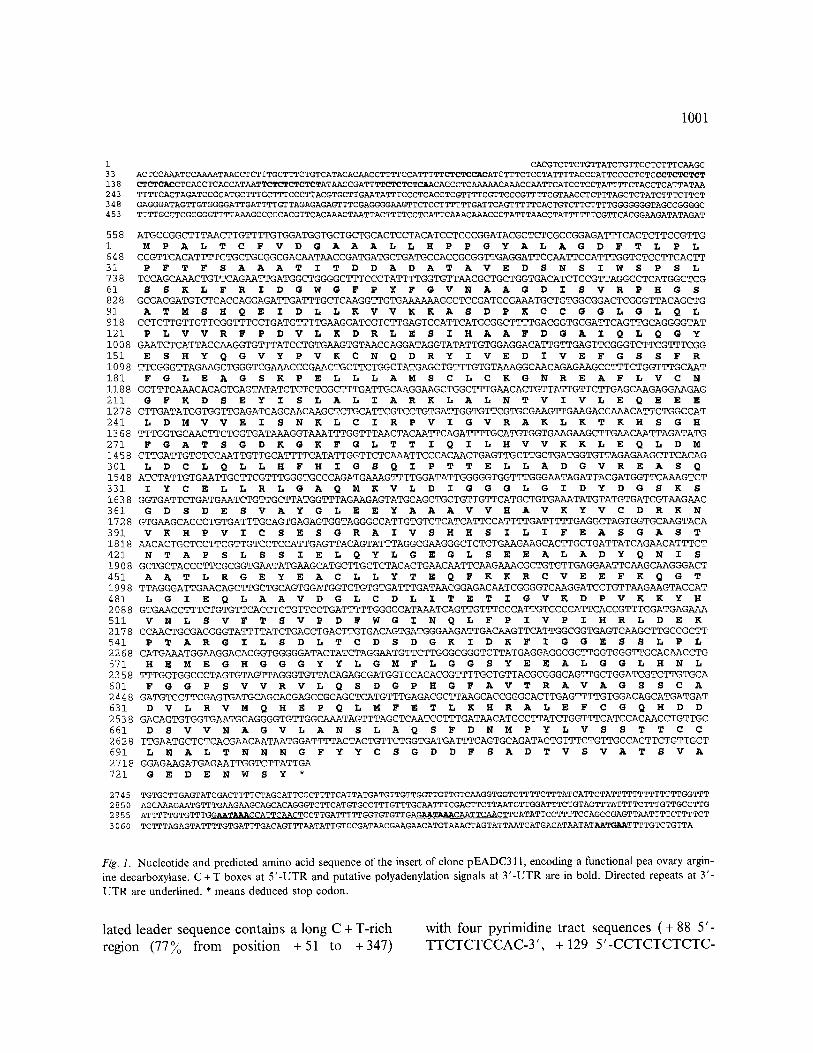
1001
1 CACGTC TTC TGTTATCTGTTCCTC TTTCAAGC 33 ACTCCAAATCCAAAATAACCTCTTTGCTTTCTGTCATACACAAC CTTTTCCATTTTTCTCTCCACATC TTTCTCCTATTTTACCCATTCCCCTCTCCCTCTCTCT 138 CTCTCACC TCAC CTCAC CATAATTCTCTCTCTCTATAACCGATTTTCTCTCTCAACACCCTCAAAAACAAACCAATTCATC CTCCTATTTTCTAC CTCATTATAA 243 TTTTCACTAGATCCGCATGCTTTGCTTTCCCTTACGTGCTTGAATATTTCCCTCACCTCGTTTTCGTTCCCGTTTTC GTAACCTCTTTAGCTCTATCTTTC TTCT 348 GAGGGATAGTTGTGGGGATTGATTTTGTTAGAGAGAGTTTCGAGGGGAAGTTCTCCTTTTTTGATTCAGTTTTTCACTGTCTTCTTTTGGGGGGGTAGC CGGGGC 453 TTTTGCCTCGCGGGTTTTAAAGC CCCCAC GTTCACAAACTAATTACTTTTCCTCATTCAAACAAACCCTATTTAACCTATTTTTTCGTTCACGGAAGATATAGAT
558 ATGCCGGCTTTAACTTGTTTTGTGGATGGTGCTGCTGCACTCCTACATCCTCCCGGATACGCTCTCGCCGGAGATTTCACTCTTCCGTTG 1 M P A L T C F V D G A A A L L H P P G Y A L A G D F T L P L 648 C CGTTCACATTTTCTGCTGCGGCGACAATAACCGATGATGCTGATGC CACCGCGGTTGAGGATTCCAATTCCATTTGGTCTCCTTCACTT 31 P F T F S A A A T I T D D A D A T A V E D S N S I W S P S L 738 TCCAGCAAACTGTTCAGAATTGATGGCTGGGGCTTTCCCTATTTTGGTGTTAACGCTGCTGGTGACATCTCCGTTAGGCCTCATGGCTCG 61 S S K L F R I D G W G F P Y F G V N A A G D I S V R P H G S 828 GCGACGATGTCTCACCAGGAGATTGATTTGCTCAAGGTTGTGAAAAAAGCCTCCGATCCGAAATGCTGTGGCGGACTCGGGTTACAGCTG 91 A T M S H Q E I D L L K V V K K A S D P K C C G G L G L Q L 918 C CTC TTGTTGTTCGGTTTCCTGATGTTTTGAAGGATCGTCTTGAGTC CATTCATGCGGCTTTTGACGGTGCGATTCAGTTGCAGGGGTAT 121 P L V V R F P D V L K D R L E S I H A A F D G A 7 Q L Q G Y 1008 GAATCTCATTAC CAAGGTGTTTATCCTGTGAAGTGTAACCAGGATAGGTATATTGTGGAGGACATTGTTGAGTTCGGGTCTTCGTTTCGG 151 E S H Y Q G V Y P V K C N Q D R Y I V E D i V E F G S S F R 1098 TTCGGGTTAGAAGCTGGGTCGAAAC CC GAACTGCTTCTGGCTATGAGCTGTTTGTGTAAAGGCAACAGAGAAGCCTTTCTGGTTTGCAAT 181 F G L E A G S K P E L L L A M S C L C K G N R E A F L V C N 1188 GGTTTCAAAGACAGTGAGTATATCTCTCTGGCTTTGATTGCAAGGAAG CTGGCTTTGAACACTGTTATTGTTCTTGAGCAAGAGGAAGAG 211 G F K D S E Y I S L A L 7 A R K L A L N T V I V L E Q E E E 1278 CTTGATATGGTGGTTGAGATCAGCAACAAGCTCTGCATTCGTCCTGTGATTGGTGTTCGTGCGAAGTTGAAGACCAAACATTCTGGC CAT 241 L D M V V E I S N K L C I R P V 7 G V R A K L K T K H S G H 1368 TTTGGTGCAACTTCTGGTGATAAAGGTAAATTTGGTTTAACTACAATTCAGATTTTG CATGTGGTGAAGAAGCTTGAACAATTAGATATG 271 F G A T S G D K G K F G L T T I Q I L H V V K K L E Q L D M 1458 CTTGATTGTCTCCAATTGTTGCATTTTCATATTGGTTCTCAAATTCCCACAACTGAGTTGCTTGCTGATGGTGTTAGAGAAGCTTCACAG 301 L D C L Q L L H F H I G S Q I P T T E L L A D G V R E A S Q 1548 ATCTATTGTGAATTGCTTCGTTTGGGTGCC CAGATGAAAGTTTTGGATATTGGGGGTGGTTTGGGAATAGATTAC GATGGTTCAAAGTCT 331 I Y C E L L R L G A Q M K V L D I G G G L G I D Y D G S K S 1638 GGTGATTCTGATGAATCTGTTGCTTATGGTTTAGAAGAGTATGCAGCTGCTGTTGTTCATGCTGTGAAATATGTATGTGATCGTAAGAAC 361 G D S D E S V A Y G L E E Y A A A V V H A V K Y V C D R K N 1728 GTGAAGCAC C CTGTGATTTGCAGTGAGAGTGGTAGGGCCATTGTGTCTCATCATTC CATTTTGATTTTTGAGGCTAGTGGTGCAAGTACA
391 V K H P V I C S E S G R A I V S H H S I L I F E A S G A S T 1818 AACACTGCTC CTTCGTTGTCCTCCATTGAGTTACAGTATTTAGGCGAAGGGCTCTCTGAAGAAGCACTTGCTGATTATCAGAACATTT•T 421 N T A P S L S S x E L Q Y L G E G L S E E A L A D Y Q N I S 1908 GCTGCTACC C TTCGC GGTGAATATGAAGCATGCTTGCTCTACACTGAACAATTCAAGAAACGCTGTGTTGAGGAATTCAAGCAAGGGACT 451 A A T L R G E Y E A C L L Y T E Q F K K R C V E E F K Q G T 1998 TTAGGGATTGAACAGC TTGCTGCAGTGGATGGTCTGTGTGATTTGATAACGGAGACAATCGGGGTCAAGGATCC TGTTAAGAAGTACCAT 481 L G I E Q L A A V D G L C D L I T E T i G V K D P V K K Y H 2088 GTGAAC CTTTCTGTGTTCACCTCTGTTC CTGATTTTTGGGGCATAAATCAGTTGTTTCC CATTGTCCC CATTCACCGTTTGGATGAGAAA 511 V N L S V F T S V P D F W G I N Q L F P I V P X H R L D E K 2178 C CAACTGCGAGGGGTATTTTATCTGAC CTGACTTGTGACAGTGATGGGAAGATTGACAAGTTCATTGGCGGTGAGTCAAGCTTGCCGCTT 541 P T A R G 7 L S D L T C D S D G K I D K F I G G E S S L P L 2268 CATGAAATGGAAGGACACGGTGGGGGATACTAT•TAGGAATGTTCTTGGG•GGGTCTTATGAGGAGGCGCTTGGTGGGTq?GCACAAC•TG 571 H E M E G H G G G Y Y L G M F L G G S Y E E A L G G L H N L 2358 TTTGGTGGC C CTAGTGTAGTTAGGGTGTTACAGAGCGATGGTCCACACGGTTTTGCTGTTACGCGGGCAGTTGCTGGATCGTCTTGTGCA 601 F G G P S V V R V L Q S D G P H G F A V T R A V A G S S C A 2448 GATGTC CTTCGAGTGATGCAGCACGAGC CGCAGC TCATGTTTGAGACGCTTAAGCACCGGGCACTTGAGTTTTGTGGACAGCATGATGAT 631 D V L R V M Q H E P Q L M F E T L K H R A L E F C G Q H D D 2538 GACAGTGTGGTGAATGCAGGGGTGTTGGCAAATAGTTTAGCTCAATC C~]?TGATAACATGCC TTATCTGGTTTCATCCACAAC CTGTTGC 661 D S V V N A G V L A N S L A Q S F D N M P Y L V S S T T C C 2628 TTGAATG CTCTCACGAACAATAATGGATTTTACTACTGTTCTGGTGATGATTTCAGTGCAGATACTGTTTCTGTTGC CACTTCTGTTGC T 691 L N A L T N N N G F Y Y C S G D D F S A D T V S V A T S V A
2718 GGAGAAGATGAGAATTGGTCTTATTGA 721 G E D E N W S Y *
2745 TGTGCTTGAGTATCGACTTTTCTAGCATTC CCTTTTCATTATGATGTTGTTGGTTGTTGTCAAGGTGGTCTTTTCTTTATCATTCTATTTTTTTTTTTTTGGTTT 2850 AGCAAAGAATGTTTGAAGAAGCAGCACAGGGTCTTCATGTGCCTTTGTTTGCAATTT C GACTTCTTAATGTTGGATTTCT GTAGTTTATTTTCTTTGTTGCCTTG 2955 ATTTTTGTGTTTGGAATAAAC CATTCAACTCCTTGATTTTTGGTGTGTTGAGAKTAAACAATTCAACTTCATATTC CTTTTCCAGCCGAGTTAATTTTCTTTTCT 3060 TCTTTAGAGTATTTTGTGATTTGACAGTTTAATATTGTCCGATAACGAAGAACATGTAAACTAGTATTAATCATGACATAATATAATGAATTTTGTCTGTTA
Fig. 1. Nucleotide and predicted amino acid sequence of the insert of clone pEADC311, encoding a functional pea ovary argin- ine decarboxylase. C + T boxes at 5 ' -UTR and putative polyadenylation signals at 3 ' -UTR are in bold. Directed repeats at 3 '- UTR are underlined. * means deduced stop codon.
lated leader sequence contains a long C + T-rich region (77~o from position +51 to +347)
with four pyrimidine tract sequences (+ 88 5'- TTCTCTCCAC-3', + 129 5'-CCTCTCTCTC-

1002
TCTCAC-3' , + 160 5'-TTCTCTCTCTCT-3' and + 182 5' -TTCTCTCTCA-3' ) related with those previously described in the leader sequences of spinach genes for thylakoid proteins (CT-leader box, CT-LB) [2]. There is also an ATG at posi- tion 259 that gives a 24 bp long ORF. The 3'- UTR of the pea ADC cDNA is 420 bp long and contains two mammalian consensus polyadeny- lation signals at positions 2969 and 3144 respec- tively (AATAAA, typed bold in Fig. 1), although their positions (154 and 192 bp upstream of the polyadenylation site), and other evidence [17] suggest that these sequences may not be func- tional in this gene. An AATAAA-like motif (AATGAA) is located at position 3144 ( - 17 from the polyadenylation site), which is identical to that found in the zein 24 kDa gene and in the pea rbcS (site 2) gene [ 17]. The 3 '-UTR also contains an almost perfect 17 bp direct repeat sequence, around the two canonical polyadenylation sig- nals, and a G+T-r ich region from position 2789 to 2804. Sequence analysis confirmed that pEADC621 was a partial clone of the described cDNA and therefore the study was restricted to the longest one.
Figure 2 shows the alignments of the deduced pea ADC protein sequence with ADC protein sequences from tomato pericarp (Leargdeca- mod) [36], oat leaf(Asspel) [ 1 ], and biosynthetic ADC from E. coli (Ecspea) [27]. The tomato se- quence has been modified by addition of a G at position 1531. This modification extends the pre- viously published ORF by moving the corre- sponding C-terminal from position 1530 to 1845 of the nucleotide sequence. After this modifica- tion, the tomato sequence shows a stop codon at the same position that pea ADC. The N-terminal region has also been extended by translation of the 5 '-UTR reported by the authors. The modi- fied tomato ADC protein is 113 aminoacids longer with extended homology with pea ADC in that region. The pea ADC protein is 74~o, 46Yo and 35Yo identical and 8670, 65~o and 60Yo similar to those from tomato, oat and E. coli, respectively. From the alignment presented in Fig. 2, it is ob- served that ADC protein sequences from plants are highly conserved in two regions, from posi-
tions 115 to 418 (6470 identity) and from 509 to 609 (71 70 identity) of the pea ADC sequence. There are also some conserved residues and changes in ADC and ODC sequences from eucaryotes (asterisks and periods, respectively in Fig. 2).
Expression of ADC cDNA in yeast
A Xho I-Dra I DNA fragment of pEADC311 in- cluding the coding region was subcloned into the S. cerevisiae expression vector pEMBL-yex4 under the control of the galactose-inducible pro- moter CYC1-GAL10 (Fig. 3). Most of the 5'- UTR of the pea clone was eliminated by Dra I digestion at site 490 to avoid ATG at position 259 and secondary structure domains, which are known to impair the expression of heterologous genes in yeast. Sense (pYex4-ADC6) and anti- sense ADC (pYex4-ADC8) plasmids were ob- tained and S. cerevisiae strain ENY.WA-1A was transformed with them and with the original pEMBL-yex4. Table 1 shows that very high levels of ADC activity were detected in pYex4-ADC6 yeast transformants under inducing conditions (galactose), but only a low level of activity was detected under repressing conditions (glucose). No activity was detected in extracts from cells containing antisense or the parent plasmid in MMGlu or MMGal. To test if the activity mea- sured was due to ADC and not to an unspecific decarboxylase, crude extracts from pYex4-ADC6
Table 1. ADC activity in extracts from Saccharomyces cerevi- siae transformants bearing either pea ADC coding region in sense (pYex4-ADC6) and antisense (pYex4-ADC8 or ex- pression vector pEMBL-yex4.
Cons~uct ADC activity, nmol/h per mg protein
glucose glucose galactose galactose + DFMA + DFMA
pYex4-ADC6 5 + 0 <1 2711+27 <1 pYex4-ADC8 < 2 nt < 2 nt pEMBL-yex4 < 2 nt < 2 nt
nt, not tested. Mean _+ SE.

1003
Adcpea Leargdeca-Mod
Asspel Ecspea
Adcpea Leargdeca-Mod
Asspel Ecspea
Adcpea Leargdeca-Mod
Asspel Ecspea
Adcpea Leargdeca-Mod
Asspel Ecspea
Adcpea Leargdeca-Mod
Asspel Ecspea
Adcpea Leargdeca-Mod
Asspel Ecspea
Adcpea Learqdeca-Mod
Asspel Ecspea
Adcpea Leargdeca-Mod
Asspel Ecspea
MPALTCFVDG AAALLHPPGY ALAGDFTLPL PFTFSAAATI
..................... MSDDMSMGL PSSAGEHGVL
WSPSLSSKLF RIDG~FP AA DIS~ RPHGSATMSH
..ASKMLRTY NIAW~NN~Y D~EL~HIS~ ..CPDPDVPE
TDDADAT~AVE DSNSI 55 ............... 0 ............... 0 RSMQEVAMSS QE... 31
QEI~LLKVVK KASDP ii0 O
Q E I ~ V L S V I E Q A T S A 54 A R V ~ B A Q L V . . . . . . 76
. . . . " i ~ c c G i ~ I ~ . ~v ~ I ~ m n ~ I ° ~ I ~ I c o ~.5
KTREA~ ALF QH~R~I NA ~KR~RESY~ N G D~F L ~ ~ 151
. . . . . . . . . . . . ._.~ ~n~l~u i | / ~n K D~QmmM~H~ ~Y D Y Y S 163
. . "iiii , , ! ~ V ~ L L I s v i ~ i o m a ~ H ~ i ~ H ~ a s R ~ ~ 4 i A ~ G ~ v x l ~ ~ ~ E ~ S ~ ~ x ~ ~IGE~MGHK VY L~Z~KM S~ IAZ~LDEAER ~R~R S QG~K !~:Q~ S G 237
e e • • ~ , w ~ • • ,e * w
i ~ m ~ ~ ~ ~ H ~ G " E TT DESG L D Q 219 K PA E ~ E ~ KALNK W IVFKAA SD 273
~ E ~ S ~ A A T ~ Q L~E T~ R E A G R ~ S ~ ANIRDI~T~ SAR 292
. . . . . . . . . . . . . . . ' " | S V ~A G~K F I I I S K~S N B V ~ C V Q L 272 e v v ~ ~ , ~ T ~ L N e i v • ~G S ~ M I ~ A z ~ ~ 32S
F~V~H . . ~VNIQCF~V~ V T R I ~ . Q ~ C ~ N ~ N ~ N N I ~W~G 344
iiPi . . . . .
x m ~ . i I V ~ I ~ F VS . . . . . STSHVSTQPS sGEI 322 HBVP i M A Y~_Y~M I__~L LS ..... IPEPKDDE . . . . . . . 371 N~LP~ V T A~T V~~ S NIIGVERNEY TVPTAPAEDA PEA~ 399
Adcpea Y L G ~ G L S I . E A L A D Y Q N ~ A Leargd .... Mod S L V,T L N, . D A RAD Y,N,A A AV RG E Y D T . . . . . . . . C LI
Asspel . . D~A T T~..Q LHGRIHD~ S KLQ PTGLSM ........ S. S Ecspea SMW~TWQ~MH EPGTR RE
Adcpea Leargdeca-Mod
Asspel Ecspea
Adcpea Leargdeca-Mod
Asspel Ecspea
A~LR~Y~ . . . . . . . . CLL y~EmFI~CV IE~IQ 47~ y S D I L m ~ C V mQ~mD 36S HAVHI~ HGI ~MY~L 41~
WLHDSQMDLH DIHIGYSSGI FSL~ERAWAE QLYLS 45~
ITLGIEQLAA DGLCDLITE TIGVKDPVKK V ~ F ~ N ~ IV 533 SLDIEQLAA VDSICDWVSK AIGVADPVRT liF~s ~ IV 423 K ............. KLSKS VTTDAHTIYN M i Y B Q H _ ~ MM 456
MY MCHEVQKQLD pQNRAHRPII DELQERMADK ~ Q ~ M ~ A ~ D ~ VL 509
I - " " I H ~ m G I m S m L i G S S ~ i G S G D ~ R 475 V S H K A T H V H V i R D T E . . T ~ L D ~ K L m m . 507 L ~ ~ IAvmLmxi D i O ~ X A + T M P M Y D P ~ P P ~ 56~
. . . . . . . . ! i# i , i Adcpea G S E 'GL LQSDG PHG~A~TRAV A SS
Leargdeca-Mod G E GL MQSDS PHS~ABTRSV P PSC V R AM~FE 530 Asspel i T Q S N K ~ VGTGN GGA~NBEAAL L STTEIELIG TVSYD 562 Ecspea V Q ~ N M ~ g T iFVFPD G.SVE~ELSD EmDTV~M~Q YV~LD 616
Adcpea ~QLMFETL~H RALE.FCGQH DDDSVVNAGV LANSLAQSFD Leargdeca-Mod ~ELMFETLRH RAEESLEQGE GEGEGVAFGS LTSSLAQSFH
Asspel ....... V__Q DISSVIEERA RENKV..WEM VEKLVESGLH Ecspea ~KTLLTQFRD QVKKTDLDAE LQQQ ...... FLEEFEAGLY GYT
VSS ..... TT 688 SSCCF TAEAT 585 ADYKP PPMA* 607 EDE* ...... 658
Adcpea CCLNALTNNN GFYYCSGDDF SADTVSVATS VAGEDENWSY 728 Leargdeca-Mod ANANTNTNNG GYYYYSEDN .......... A AAEEDEIWSY 615
607 Asspel ........................................ 658 Ecspea ........................................
Fig. 2. Comparison of protein sequence of pea ovary ADC (Adcpea) with those from tomato pericarp modified as explained in the text (Leargdeea-mod) [36], oat leaf (Asspel) [1] and Escherichia coli (Ecspea) [27]. Conserved residues in all sequences are outlined in black and conserved changes are light shaded. * and • mean conserved residues and changes in ADC and ODC proteins
from eukaryotes, respectively.

1004
TGA @ l
Smal Xhol digestion Smal digestion Klenow treatment
Dial ] m I
t I DtaI-Xhol fragment elution /
~ Ligetion /
Selection of transformants J
pYex4-ADC6 sense
Gene expression of pea ADC. Changes in unpolli- nated ovaries and tissue specificity
The pattern of ADC gene expression in the ovary and during early fruit development as well as its tissue specificity were analyzed by northern blot. In all cases a unique product of ca. 3.2 kb was detected (Figs. 4, 5 and 6).
Figure 4A shows the time course of ADC m R N A levels, ADC specific activity and ovary weight during development of untreated and GA3-treated unpollinated pea ovaries. In un- treated unpollinated ovaries, the ADC mRNA was detected throughout the different stages of ovary growth (DO - and D 1 - ), although its ex-
Fig. 3. Subcloning of a Dra I-Xho I fragment of pEADC311 in sense (pYex4-ADC6) and antisense (pYex4-ADC8) into the yeast expression vector pEMBL-yex4. Black box in pEMBL- yex4 and pYex4 plasmids means CYC1-GAL10 promoter. See Materials and methods for details.
transformants were assayed in the presence of DFMA, an irreversible specific inhibitor of ADC activity. The decarboxylase activity on arginine was inhibited ca. 99.9~o. A loss in total protein content, after the beginning of the exponential phase of growth, was observed in a culture of yeast transformed with pYex4-ADC6 and grown in MMGal. The cause of this drop in protein might be either depletion of the arginine pool re- quired for protein synthesis, or a possible toxic effect of the product of the ADC activity (agma- tine) when the ADC activity reaches its highest level.
Fig. 4. A. ADC mRNA levels, ADC activity (nmol/h per ovary) and ovary weight (rag per ovary) of unpollinated pea ovaries treated with GA 3 ( + ) or untreated ( - ) and collected at day 0 (DO), 1 (D1), 2 (D2), 3 (D3) and 4 (D4) after anthesis. Treatment was carried out at day of anthesis. 40/~g of total RNA were loaded per well. B. ADC mRNA level in ovaries treated with GA 3 ( + ) at day two after anthesis and collected at 0, 1, 3, 6, 9 and 12 hours, or untreated ( - ) at 12 h after treatment. 2/~g of poly(A) + RNA were loaded per well.

1005
pression decreased steadily down from D1 - to the lowest level 4 days after anthesis (D4 - ). ADC activity however decreased from the day of an- thesis to two days later and remained constant during the senescence of the ovary ( D 3 - 4 - ). This remaining activity was not inhibite.d_~y ..... DFMA [32]. In its fate, the ovary increased in weight up to day 2-3 after anthesis, and then decreased during senescence. GA 3 treatment pre- vented ovary senescence and induced partheno- carpic fruit growth. Treatment of the ovaries with GA3 at the day of anthesis induced a parallel increase in both ADC mRNA and ADC activity levels. This increase was transient, with a maxi- mum two days later (D2 + ). Levels of ADC mRNA at day four after anthesis revealed no difference in treated (D4 + ) and untreated (D4 -.) ovaries, but the level of ADC activity in GA 3- treated ovaries was 3-fold that of untreated ova- ries. As reported before, fruit weight increased steadily after treatment, with a maximum at the eighth day after the GA3 treatment [14]. An ex- periment was designed to see how fast ovaries respond to GA3 treatment by inducing ADC mRNA. When ovaries were treated with GA3 at day 2 after anthesis, the increase in ADC mRNA levels were already detected 12 h after treatment (Fig. 4B).
The pattern of ADC mRNA expression was also studied during early fruit growth in natural pollinated ovaries (Fig. 5A). A transient increase in ADC transcript, that peaks three days after treatment, was observed. This pattern is similar to that obtained for GA3-treated unpollinated ovaries with differences in timing. Small differ- ences in ADC mRNA levels between GA3-treated and self-pollinated ovaries could be explained by differences in the growth curve. Thus, although GA 3 treatment induces fruits that are identical in shape, size and structure to those obtained by pollination [47], the increase in ovary weight as promoted by GA 3 treatment at day 0 was about 1 day faster than that promoted by pollination (see weights and ADC mRNA levels in Figs. 4A and 5A).
To determine if the increase in ADC mRNA levels was specific of the GA3 treatment, ovaries
Fig. 5. Effect of pollination and treatment with different plant growth substances on the accumulation of ADC mRNA. A. ADC mRNA level and ovary weight after self-pollination of pea ovaries. Ovaries were collected at day 0) P0, 1 (P1), 2 (P2), 3 (P3) and 4 (P4) after anthesis. 40 #g of total RNA were loaded per well. B. ADC mRNA level and ovary weight after treatment of unpollinated pea ovaries with GA 3 (G), 2,4-D (D) and BA (B). Treatment was at day two after anthesis. Ovaries were collected at 0, 12 and 24 h after treatment. 30 #g of total RNA were loaded per well.
at day two after anthesis were treated with other plant growth substances (PGS), BA and 2,4-D, that are able to induce also parthenocarpic fruit development [14]. These PGS treatments have an effect on ADC mRNA levels similar to that achieved by GA3 (Fig. 5B). As shown in the fig- ure, 24 h after the treatments, there were no marked differences in ADC mRNA levels and weights between ovaries treated with each of these hormones.
Figure 6 shows ADC mRNA levels in different organs and tissues of the adult pea plant. Expres- sion of ADC was higher in young and develop- ing tissues such as shoot tips, young leaflets (0.5- 1.0 cm long) and flower buds (two days before

1006
Fig. 6. Tissue specificity of ADC mRNA. ADC mRNA ac- cumulation in different adult pea plant tissues and organs: shoot tip (ST), young leaflet (YL, 0.5-1.0 cm long), adult fully expanded leaflet (AL, 3.0-3.5 cm long), flower bud (F, two days before anthesis), stem (S) and root (R). 40 #g of total RNA were loaded per well.
anthesis). Low levels of ADC transcript were de- tected in mature organs such as fully expanded leaflets and roots. In fully elongated stems the ADC m R N A levels were between those of young and adult leaves.
Discussion
This paper describes the cloning and expression of a cDNA, isolated from a pea young fruit li- brary, and provides evidence that this cDNA en- codes a functional ADC. Northern analysis indi- cate that the expression o fADC gene is regulated, at least in part, at the level of m R N A accumula- tion during ovary growth and early fruit develop- ment. Previously, we reported that ADC is the enzyme responsible for the putrescine biosynthe- sis in pea ovaries and in young fruits [32].
Sequences encoding ADC have been conserved throughout evolution. The sequence of pea ovary ADC is highly identical to that from tomato peri- carp [36], and very similar to that from oat leaf [ 1 ] and the biosynthetic enzyme from E. coli [27 ]. Even though they are different in length, the four proteins present some conserved identical regions (highlighted in Fig. 2). Some of these regions are even similar to those present in other decarboxy- lases, mainly in ODC from eucaryotes. More- over, the sequence L D I G G G L G I at position 345-353 in the pea sequence, previously de- scribed as a putative substrate-binding site [27], is found in all ODC and ADC proteins reported
from eucaryotes. All these proteins also conserve the residues described as functional binding sites for pyridoxal phosphate and the specific inhibitor of ODC activity, ~-DL-difluoromethylornithine (lysine at position 161 and cysteine at 552 of the pea sequence, respectively) [34]. The conserva- tion of these residues, and the sequences around them, could mean that the catalytic mechanism of ODC and ADC proteins from eucaryotes would be similar. A second ADC gene has been reported from E. coli where it is induced in acidic enriched medium containing arginine [28, 40]. The de- duced ADC protein is similar in size to pea ADC, but has only 16 ~o identity and 40 ~o overall ami- noacid sequence similarity, and furthermore, it does not contain the conserved regions cited above.
The pea ADC clone encoded a functional ADC protein. The strongest evidence of the nature of the cDNA clone from pea was obtained by its expression in S. cerevisiae. The ADC activity of the transformed S. cerevisiae with the pea se- quence under inducing conditions and its specific inhibition by D F M A provides the first sound evi- dence that the product of the pea ADC cDNA is an active ADC enzyme. Previously, Malmberg et al. [26] showed that in vitro translated oat ADC was inactive. Only after adding a purified oat pro- tein fraction was it possible to obtain ADC ac- tivity [26]. Evidence was given that activation of the in vitro translated protein by the oat extract involved proteolytic processing. Our results indi- cate that, whatever the factor is, S. cerevisiae is able to adequately process the ADC protein to give an active enzyme. Expression of heterolo- gous eukaryotic genes in yeast ensures a correct protein conformation, proteolytic processing of pre- or proproteins and their glycosylation [43]. Furthermore, a number of plant genes have been successfully expressed in yeast. A cDNA encod- ing an ethylene-forming enzyme, isolated from to- mato fruit, was identified by its expression in yeast [ 15]. More recently, a SAMDC cDNA from po- tato was demonstrated by its ability to comple- ment a yeast null mutant [25].
A different role for ODC and ADC in growing plant tissues has been proposed. ODC would be

1007
the enzyme responsible for putrescine biosynthe- sis in tissues undergoing active cell division while ADC would be in charge in non-dividing tissues or in response to environmental stresses [44]. Studies on the expression and localization of these genes should corroborate this hypothesis. Parthenocarpic fruit development in pea is mainly due to cell expansion (enlargement of the meso- carp cells and differentiation of the endocarp cells) [45, 46]. Furthermore, in these fruits, ADC is the key enzyme involved in putrescine biosynthesis as ODC activity is below detection [32]. In pea, the expression of ADC gene is developmentally regulated in reproductive and other actively grow- ing tissues. The levels of ADC transcript were correlated with ADC activity during early stages of fruit development. Pea ADC is expressed at high levels in other rapid growing tissues like young leaflets, where cell division and expansion is occurring. The expression of pea ADC gene decreases during the last step of leaf develop- ment. The higher level is detected when cell en- largement is the main process implicated in leaf expansion, although some cell division may still occur [9]. By contrast, the adult stem does not show any change in length, but shows relatively high levels of ADC mRNA. This could be ex- plained by secondary growth of the xylem/phloem system. In the tomato, ADC transcripts were de- tected at the highest levels in the corpus and leaf primordium of the apical meristem in the shoot apex [13], but also in the pericarp during the breaker stage of fruit ripening [35], when most of the expansion and division are not longer occur- ring. Further studies using immunolocalization and in situ hybridization techniques are therefore needed to define the cellular localization of ADC expression in tissues undergoing simultaneous cell division and expansion.
The levels of ADC transcript accumulation alone can not always account for the differences observed at the level of ADC activity (see Fig. 4A lanes 1, 2 and 4, lines 8 and 9). Similarly, in to- mato, ADC mRNA levels do not correlate with ADC activity during ripening of Rutgers-alc and Rutgers fruits [35, 36]. ADC activity in alc fruits rises during the ripening process, whereas in nor-
mal Rutgers fruits a decrease in ADC activity occurs at and following the breaker stage. In con- trast, both normal and alc fruits show a similar pattern in ADC gene expression with a maximum at the breaker stage. All this suggests that other points of regulation at the level of mRNA stabil- ity, transcription and translation efficiency or posttranslational processing, may be involved in the production of active ADC in pea and tomato.
Transcription and translation efficiency can be affected by sequences within the mRNA in sev- eral ways. The length of most leader sequences in eukaryotic mRNAs range from 40 to 80 bp, being the extremely long leaders which are rare and usually confined to viral messengers [22, 23]. Most of leader sequences in plant genes range from 9 to 193 bp [20]. The pea ADC has a 5'- UTR of 557 bp. Some examples of long leader sequences have been reported in plants: SAMDC clone from potato (over 500 bp) [25], and 563 bp for the cysteine synthase from watermelon [30]. The later and the pea ADC leader sequences show a pyrimidine tract and cryptic ATGs similar to those present in the long leader sequences of pi- cornaviruses [33]. Whether CT boxes present in the pea ADC sequence modify transcriptional efficiency as reported for the genes of some thy- lakoid proteins [2], or the sequences around ATG may affect translational start recognition [ 22, 23 ] remains to be investigated.
Finally, ADC activity may be regulated post- translationally as was reported for the oat enzyme [1]. In oat, a posttranslational cleavage of the ADC precursor by a not yet characterized pro- cessing enzyme seems to be a necessary step to obtain active ADC [26]. The phylogenetically distant biosynthetic ADC from E. coli is synthe- sized as a propeptide that is cleaved, loosing a transit peptide, to be translocated to the inner periplasmic space [4, 27].
Acknowledgements
We wish to thank Dr R. Rastogi and Dr S.J. Rothstein for the tomato ADC clone and the Marion Merrell Dow Research Institute for the

1008
gift of DFMA. We are also grateful to Dr P Sanz and Miss F Rfindez-Gil for their help in the work with Saecharomyces and for critically reading the manuscript and Mr R Martinez-Pardo and Mr A Villar for their help in the greenhouse. This work was supported by grant PB92-0018-C02- 01. M.A.P.-A. received a fellowship from Direc- cirn General de Investigacirn Cientifica y T~c- nica (Spain).
References
1. Bell E, Malmberg RL: Analysis of a cDNA encoding arginine decarboxylase from oat reveals similarity to the Escherichia coli arginine decarboxylase and evidence of protein processing. Mol Gen Genet 224:431-436 (1990).
2. Bolle C, Sopory S, Lttbberstedt Th, Herrmann RG, OemOller R: Segments encoding 5'-untranslated leaders of genes for thylakoid proteins contain cis-elements es- sential for transcription. Plant J 6:513-523 (1994).
3. Bradford MM: A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72: 248-254 (1976).
4. Buch JK, Boyle SM: Biosynthetic arginine decarboxylase in Escherichia coli is synthesized as a precursor and located in the cell envelope. J Bact 163:522-527 (1985).
5. Carbonell J, Garcia-Martinez JL: Ribulose-l,5-bisphos- phate carboxylase and fruit set or degeneration of unpol- linated ovaries of Pisum sativum. Planta 164:534-539 (1985).
6. Carbonell J, Navarro JL: Correlation of spermine levels with ovary senescence and fruit set and development in Pisum sativum L. Planta 178:482-487 (1989).
7. Cesareni G, Murray AA: Plasmid-veetors carrying the replication origin of filamentous single-stranded phages. In: Stlow JK (Ed), Genetic Engineering vol. 9, pp. 135- 154. Plenum Press, New York (1987).
8. Cohen E, Arad S(M), Heimer YM, Mizrahi Y: Partici- pation of ornithine decarboxylase in early stages of to- mato fruit development. Plant Physio170:540-543 (1982).
9. Dale JE: The control of leaf expansion. Annu Rev Plant Physiol Plant Mol Biol 39:267-295 (1988).
10. Doorsselaere JV, Gielen J, Van Montagu M, Inz6 D: A cDNA encoding S-adenosyl-L-methionine synthetase from poplar. Plant Physiol 102:1365-1366 (1993).
11. Espartero J, Pintor-Toro JA, Pardo JM: Differential ac- cumulation of S-adenosylmethionine synthetase tran- scripts in response to salt stress. Plant Mol Biol 25: 217- 227 (1994).
12. Evans PT, Malmberg RL: Do polyamines have roles in plant development? Annu Rev Plant Physiol Plant Mol Biol 40:235-269 (1989).
13. Fleming AJ, Mandel T, Roth I, Kuhlemeier C: The pat- terns of gene expression in the tomato shoot apical mer- istem. Plant Cell 5:297-309 (1993).
14. Garcia-Martinez JL, Carbonell J: Fruit-set of unpolli- nated ovaries of Pisum sativum L. Influence of plant- growth regulators. Planta 147:451-456 (1980).
15. Hamilton AJ, Bouzayen M, Grierson D: Identification of a tomato gene for the ethylene-forming enzyme by ex- pression in yeast. Proc Natl Acad Sci USA 88: 7434- 7437 (1991).
16. Heimer YM, Mizrahi Y, Bachrach U: Ornithine deear- boxylase activity in rapidly proliferating plant cells. FEB S Lett 104:146-148 (1979).
17. Hunt AG: Messenger RNA 3' end formation in plants. Annu Rev Plant Physiol Plant Mol Bio145:47-60 (1994).
18. Ito H, Jukuda K, Murata K, Kimura A: Transformation of intact yeast cells treated with alkali cations. J Bact 153: 163-168 (1983).
19. Jones JDG, Dunsmuir P, Bedbrook J: High levels of expression of introduced chimeric gene in regenerated transformed plants. EMBO J 4:2411-2414 (1985).
20. Joshi CP: An inspection of the domain between putative TATA box and translation start in 79 plant genes. Nucl Acids Res 15:6643-6653 (1987).
21. Kawalleck P, Plesch G, Hahlbrock K, Somssich IE: In- duction by fungal elicitor of S-adenosyl-L-methionine synthetase and S-adenosyl-L-homocysteine hydrolase mRNAs in cultured cells and leaves of Petroselinum crispum. Proc Natl Acad Sci USA 89:4713-4717 (1992).
22. Kozak M: Comparison of initiation of protein synthesis in procaryotes, eucaryotes, and organelles. Microbiol Rev 47:1-45 (1983).
23. Kozak M: Context effects and inefficient initiation at non- AUG codons in eucaryotic cell-free translation systems. Mol Cell Biol 9:5073-5080 (1989).
24. Larsen PB, Woodson WR: Cloning and nucleotide se- quence of a S-adenosylmethionine synthetase eDNA from carnation. Plant Physiol 96:997-999 (1991).
25. Mad Arif SA, Taylor MA, George LA, Butler AR, Burch LR, Davies HV, Stark MJR, Kumar A: Characterization of the S-adenosylmethionine decarboxylase (SAMDC) gene from potato. Plant Mol Biol 26:327-338 (1994).
26. Malmberg RL, Cellino ML: Arginine decarboxylase of oats is activated by enzymatic cleavage into two polypep- tides. J Biol Chem 269:2703-2706 (1994).
27. Moore RC, Boyle SM: Nucleotide sequence and analy- sis of the speA gene encoding biosynthetic arginine de- carboxylase in Escheriehia coli. J Bact 172:4631-4640 (1990).
28. Morris DR, Boeker EA: Biosynthetic and biodegradative ornithine and arginine decarboxylases from Eseherichia coll. Meth Enzymol 94:125-134 (1983).
29. Nathan R, Altman A, Monselise SP: Changes in activity of polyamine biosynthetic enzymes and in polyamine con- tents in developing fruit tissues of 'Murcott' mandarin. Sci Hort 22:359-364 (1984).

30. Noji M, Murakoshi Y, Saito K: Molecular cloning of a cysteine synthase cDNA from Citrullus vulgaris (water- melon) by genetic complementation in an Escherichia coli Cys- auxotroph. Mol Gen Genet 244:57-66 (1994).
31. Peleman J, Boerjan W, Engler G, Seurinck J, Botterman J, Alliotte T, Van Montagu M, Inz6 D: Strong cellular preference in the expression of a housekeeping gene of Arabidopsis thaliana encoding S-adenosylmethionine syn- thetase. Plant Cell 1:81-93 (1989).
32. P6rez-Amador MA, Carbonell J: Arginine decarboxylase and putrescine oxidase in Pisum sativum L. Changes dur- ing ovary senescence and early stages of fruit develop- ment. Plant Physiol 107:865-872 (1995).
33. Pilipenko EV, Gmyl AP, Maslova SV, Svitkin YV, Sinyakov AN, Agol VI: Prokaryotic-like cis elements in the cap-independent internal initiation of translation on picornavirus RNA. Cell 68:119-131 (1992).
34. Poulin R, Lu L, Ackermann B, Bey P, Pegg AE: Mecha- nism of the irreversible inactivation of mouse ornithine decarboxylase by :~-difluoromethylornithine. J Biol Chem 267:150-158 (1992).
35. Rastogi R, Davies P J: Polyamine metabolism in ripening tomato fruit. II. Polyamine metabolism and synthesis in relation to enhanced putrescine content and storage life of alc tomato fruit. Plant Physiol 95:41-45 (1991).
36. Rastogi R, Dulson J, Rothstein SJ: Cloning of tomato (Lycopersicon esculentum Mill.) arginine decarboxylase gene and its expression during fruit ripening. Plant Physiol 103:829-834 (1993).
37. Sambrook J, Fritsch EF, Maniatis T: Molecular Cloning: A Laboratory Manual, 2nd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY (1989).
38. Sanger F, Nicklen S, Coulson AR: DNA sequencing with chain terminating inhibitors. Proc Natl Acad Sci USA 74: 5463-5467 (1977).
39. Slocum RD, Galston AW: Changes in polyamine biosyn- thesis associated with postfertilization growth and devel- opment in tobacco ovary tissues. Plant Physiol 79: 336- 343 (1985).
1009
40. Stim KP, Bennett GN: Nucleotide sequence of the adi gene, which encodes the biodegradative acid-induced arginine decarboxylase of Escherichia coll. J Bact 175: 1221-1234 (1993).
41. Taylor MA, Mad Arif SA, Kumar A, Davies HV, Scobie LA, Pearce SR, Flavell AJ: Expression and sequence analysis of cDNAs induced during the early stages of tuberisation in different organs of the potato plant (Solanum tuberosum L.). Plant Mol Biol 20:641-651 (1992).
42. Teitel DC, Cohen E, Arad S(M), Birnbaum E, Mizrahi Y: The possible involvement of polyamines in the devel- opment of tomato fruits in vitro. Plant Growth Regul 3: 309-317 (1985).
43. Tekamp-Olson P, Valenzuela P: Gene expression and engineering in yeast and other fungi. Curr Opin Biotech- nol 1:28-35 (1990).
44. Tiburcio AF, Kaur-Sawhney R, Galston AW: Polyamine metabolism. In: Miflin BJ (Ed) Intermediary Nitrogen Metabolism. The Biochemistry of Plants, vol. 16, pp. 283-325. Academic Press, New York (1990).
45. Vercher Y, Carbonell J: Changes in the structure of ovary tissues and in the ultrastructure of mesocarp cells during ovary senescence or fruit development induced by plant growth substances in Pisum sativum. Physiol Plant 81: 518-526 (1991).
46. Vercher Y, Molowny A, Carbonell J: Gibberellic acid effects on the ultrastructure of endocarp cells of unpolli- nated ovaries ofPisum sativum. Physiol Plant 71:302-308 (1987).
47. Vercher Y, Molowny A, Lrpez C, Garcia-Martinez JL, Carbonell J: Structural changes in the ovary of Pisum sativum L. induced by pollination and gibberellic acid. Plant Sci Lett 36:87-91 (1994).
48. Winer L, Vinkler C, Apelbaum A: Partial purification and characterization of arginine decarboxylase from avocado fruit, a thermostable enzyme. Plant Physiol 76:233-237 (1984).




















