
Tolerance of pea (Pisum sativum L.) to long-term salt stress is associated with induction of...
10
© 2000 Blackwell Science Ltd 853 Plant, Cell and Environment (2000) 23, 853–862 salinity. Salt stress results in alterations in plant metabolism including reduced water potential, ion imbalance and toxi- city, and reduction of CO 2 assimilation (Bohnert & Jensen 1996). The effects of various environmental stresses in plants are known to be mediated, at least in part, by an enhanced generation of activated oxygen species (AOS; O 2 · - ,H 2 O 2 and ·OH) (Hernández et al. 1993, 1995; Mittler & Zilinskas 1994; Alscher, Donahue & Cramer 1997; Noctor & Foyer 1998). Although a wide range of genetic adaptations to saline conditions has been observed and a number of significant physiological responses have been associated with tolerance, the underlying mechanisms of salt-tolerance in plants are still poorly understood. However, one determinant of salt tolerance could be how well stressed plants deal with the accompanying oxida- tive damage to subcellular compartments (Hernández et al. 1993, 1995; López et al. 1996; Van Camp et al. 1996; Gueta-Dahan et al. 1997). To mitigate and repair damage initiated by AOS, plants have developed a complex antioxidant system. The primary components of this system include carotenoids, ascorbate, glutathione and tocopherols, and enzymes such as superoxide dismutase (SOD, EC 1.15.1.1), catalase (EC 1 · 11 · 1.6), glutathione peroxidase (GPX, EC 1.11.1.9), per- oxidases and the enzymes involved in the ascorbate– glutathione cycle (ASC–GSH cycle; Foyer & Halliwell 1976): ascorbate peroxidase (APX, EC 1.11.1.1), dehy- droascorbate reductase (DHAR, EC 1.8.5.1), monodehy- droascorbate reductase (MDHAR, EC 1.6.5.4) and glutathione reductase (GR, EC 1.6.4.2) (Noctor & Foyer 1998). Many components of this antioxidant defence system can be found in different subcellular compartments (Jiménez et al. 1997, 1998). Several environmental stresses induce the expression and/or levels of antioxidative enzymes and their mRNAs (Edwards et al. 1994; Mittler & Zilinskas 1994; Stevens, Creissen & Mullineaux 1997). For example, the protein level of phospholipid hydroperoxide glutathione peroxi- dase (PHGPX) was shown to be induced in salt-adapted Citrus sinensis callus and also in leaves from plants grown on media containing 0·2 m NaCl (Holland et al. 1993; Gueta- Dahan et al. 1997). However, little is known about the effect of salt stress on the enzymes of the ASC–GSH cycle in ABSTRACT Using two cultivars of Pisum sativum L. with different sen- sitivity to NaCl, the effect of long-term (15 d) NaCl (70 mM) treatments on the activity and expression of the foliar ascorbate–glutathione cycle enzymes, superoxide dismu- tase isozymes and their mRNAs was evaluated and related to their ascorbate and glutathione contents. High-speed supernatant (soluble) fractions, enriched for cytosolic components of the antioxidant system, were used. In this fraction from the NaCl-tolerant variety (cv Granada), the activities of ascorbate peroxidase (APX), glu- tathione reductase (GR), monodehydroascorbate reduc- tase (MDHAR), Mn-superoxide dismutase (Mn-SOD) and dehydroascorbate reductase (DHAR) increased, while CuZn-SOD activity remained constant. In the NaCl- sensitive plants (cv Challis), salinity did not produce sig- nificant changes in APX, MDHAR and GR activities. Only DHAR activity was induced in cv Challis, whereas soluble CuZn-SOD activity decreased by about 35%. Total ascor- bate and glutathione contents decreased in both cultivars, but the decline was greater in NaCl-sensitive plants. This difference between the two cultivars was more pronounced when the transcript levels of some these enzymes were examined. Transcript levels for mitochondrial Mn-SOD, chloroplastic CuZn-SOD and phospholipid hydroperoxide glutathione peroxidase (PHGPX), cytosolic GR and APX were strongly induced in the NaCl-tolerant variety but not in the NaCl-sensitive variety. These data strongly suggest that induction of antioxidant defences is at least one com- ponent of the tolerance mechanism of peas to long-term salt-stress. Key-words: Pisum sativum; antioxidant enzymes; ascor- bate–glutathione cycle; gene expression; oxidative stress; salt stress. INTRODUCTION One of the most important abiotic factors limiting plant productivity is water stress brought about by drought or Correspondence: Dr Francisca Sevilla. Fax: +34 968 396213; e-mail: [email protected] Tolerance of pea (Pisum sativum L.) to long-term salt stress is associated with induction of antioxidant defences J. A. HERNÁNDEZ, 1 A. JIMÉNEZ, 1 P. MULLINEAUX 2 & F. SEVILLA 1 1 Departamento de Nutrición y Fisiología Vegetal, Centro de Edafología y Biología Aplicada del Segura, CSIC, Apartado 4195, E-30100 Murcia, Spain, and 2 John Innes Centre, Norwich Research Park, Colney, Norwich NR4 7UH, UK
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Tolerance of pea (Pisum sativum L.) to long-term salt stress is associated with induction of...

© 2000 Blackwell Science Ltd 853
Plant, Cell and Environment (2000) 23, 853–862
salinity. Salt stress results in alterations in plant metabolismincluding reduced water potential, ion imbalance and toxi-city, and reduction of CO2 assimilation (Bohnert & Jensen1996). The effects of various environmental stresses inplants are known to be mediated, at least in part, by anenhanced generation of activated oxygen species (AOS;O2·-, H2O2 and ·OH) (Hernández et al. 1993, 1995; Mittler& Zilinskas 1994; Alscher, Donahue & Cramer 1997;Noctor & Foyer 1998). Although a wide range of geneticadaptations to saline conditions has been observed and anumber of significant physiological responses have beenassociated with tolerance, the underlying mechanisms of salt-tolerance in plants are still poorly understood.However, one determinant of salt tolerance could be howwell stressed plants deal with the accompanying oxida-tive damage to subcellular compartments (Hernández et al. 1993, 1995; López et al. 1996; Van Camp et al. 1996;Gueta-Dahan et al. 1997).
To mitigate and repair damage initiated by AOS, plantshave developed a complex antioxidant system.The primarycomponents of this system include carotenoids, ascorbate,glutathione and tocopherols, and enzymes such as superoxide dismutase (SOD, EC 1.15.1.1), catalase (EC1·11·1.6), glutathione peroxidase (GPX, EC 1.11.1.9), per-oxidases and the enzymes involved in the ascorbate–glutathione cycle (ASC–GSH cycle; Foyer & Halliwell1976): ascorbate peroxidase (APX, EC 1.11.1.1), dehy-droascorbate reductase (DHAR, EC 1.8.5.1), monodehy-droascorbate reductase (MDHAR, EC 1.6.5.4) andglutathione reductase (GR, EC 1.6.4.2) (Noctor & Foyer1998). Many components of this antioxidant defence systemcan be found in different subcellular compartments(Jiménez et al. 1997, 1998).
Several environmental stresses induce the expressionand/or levels of antioxidative enzymes and their mRNAs(Edwards et al. 1994; Mittler & Zilinskas 1994; Stevens,Creissen & Mullineaux 1997). For example, the proteinlevel of phospholipid hydroperoxide glutathione peroxi-dase (PHGPX) was shown to be induced in salt-adaptedCitrus sinensis callus and also in leaves from plants grownon media containing 0·2 m NaCl (Holland et al. 1993; Gueta-Dahan et al. 1997). However, little is known about the effectof salt stress on the enzymes of the ASC–GSH cycle in
ABSTRACT
Using two cultivars of Pisum sativum L. with different sen-sitivity to NaCl, the effect of long-term (15 d) NaCl (70 mM)treatments on the activity and expression of the foliarascorbate–glutathione cycle enzymes, superoxide dismu-tase isozymes and their mRNAs was evaluated and relatedto their ascorbate and glutathione contents. High-speedsupernatant (soluble) fractions, enriched for cytosolic components of the antioxidant system, were used. In this fraction from the NaCl-tolerant variety (cv Granada),the activities of ascorbate peroxidase (APX), glu-tathione reductase (GR), monodehydroascorbate reduc-tase (MDHAR), Mn-superoxide dismutase (Mn-SOD) anddehydroascorbate reductase (DHAR) increased, whileCuZn-SOD activity remained constant. In the NaCl-sensitive plants (cv Challis), salinity did not produce sig-nificant changes in APX, MDHAR and GR activities. OnlyDHAR activity was induced in cv Challis, whereas solubleCuZn-SOD activity decreased by about 35%. Total ascor-bate and glutathione contents decreased in both cultivars,but the decline was greater in NaCl-sensitive plants. Thisdifference between the two cultivars was more pronouncedwhen the transcript levels of some these enzymes wereexamined. Transcript levels for mitochondrial Mn-SOD,chloroplastic CuZn-SOD and phospholipid hydroperoxideglutathione peroxidase (PHGPX), cytosolic GR and APXwere strongly induced in the NaCl-tolerant variety but notin the NaCl-sensitive variety. These data strongly suggestthat induction of antioxidant defences is at least one com-ponent of the tolerance mechanism of peas to long-termsalt-stress.
Key-words: Pisum sativum; antioxidant enzymes; ascor-bate–glutathione cycle; gene expression; oxidative stress;salt stress.
INTRODUCTION
One of the most important abiotic factors limiting plantproductivity is water stress brought about by drought or
Correspondence: Dr Francisca Sevilla. Fax: +34 968 396213;e-mail: [email protected]
Tolerance of pea (Pisum sativum L.) to long-term salt stress is associated with induction of antioxidant defences
J. A. HERNÁNDEZ,1 A. JIMÉNEZ,1 P. MULLINEAUX2 & F. SEVILLA1
1Departamento de Nutrición y Fisiología Vegetal, Centro de Edafología y Biología Aplicada del Segura, CSIC, Apartado4195, E-30100 Murcia, Spain, and 2John Innes Centre, Norwich Research Park, Colney, Norwich NR4 7UH, UK

plants, and there is not enough information about theexpression of the different SOD isozymes in plants grownunder salt-stress situations. In previous studies with pea(Pisum sativum L.), we demonstrated that the metabolismof chloroplasts and mitochondria under NaCl stressfavoured the formation of O2·- radicals and H2O2 in twocultivars of differing NaCl sensitivity, and that tolerantplants, but not sensitive ones, responded to NaCl stress withincreased mitochondrial Mn-SOD and chloroplastic CuZn-SOD and ascorbate peroxidase activities (Hernández et al.1993, 1995). The effects of salt stress on the cytosol com-partment of pea leaf cells have not been studied, and yet itis this compartment that might be the most important in theplant’s response to salt-stress-associated oxidative stress,such as has been described for the response to paraquat andto SO2 exposure (Alscher, Donahue & Cramer 1997).
In this paper, we report a more detailed study of theeffect of long-term salt stress on NaCl-tolerant and NaCl-sensitive pea cultivars. We report here that there are cleardifferences between the salt-responsive genotypes in termsof the activity of antioxidant enzymes and low molecularweight antioxidants in cytosol-enriched fractions and thelevels of their mRNAs, which might indicate a significantrole for antioxidant defences in conferring NaCl tolerancein pea plants.
MATERIALS AND METHODS
Growth of plants in salt-stress conditions
Pea (Pisum sativum L.) seeds were surface-sterilized(ethanol (96% v/v) for 3 min and sodium hypochlorite(10% v/v) for 5 min), germinated and grown in vermiculite.Vigorous seedlings were selected for hydroponic culture ina growth chamber (ASL, Madrid, Spain). First, plants werecultivated in aerated distilled water for 7 d (Hernándezet al. 1993). Then, plants were transplanted to aeratedoptimum nutrient solution for another 7 d. The growthchamber was set at 24/18 °C, 80% relative humidity and200 mmol m-2 s-1 of light intensity with a 16 h photoperiod.After 7 d, 70 mm NaCl were added to the nutrient solutionand plant leaves were sampled at 0, 2, 4, 8 and 24 h and 15 dafter the NaCl was added.
Enzyme extraction and assays
All operations were performed at 0–4 °C. For total extracts,leaves (1 g) were homogenized with a mortar and pestle in2 mL of ice-cold 50 mm K-phosphate buffer pH 7·8, 0·1 mmEDTA containing 5 mm cysteine, 1% w/v PVP, 0·1 mmPMSF and 0·2% v/v Triton X-100. For APX activity,20 mm ascorbate was added. The homogenate was cen-trifuged at 14 000 g for 20 min and the supernatant fractionwas filtered through a column containing 1 mL of Sepha-dex G-50 equilibrated with the same buffer used for thehomogenization.
Soluble fractions were prepared by homogenizing 2 gfresh leaf material with a mortar and pestle with 8 mL of a
854 J. A. Hernández et al.
grinding medium containing 0·35 m mannitol, 30 mm MOPSbuffer (pH 7·5), 4 mm l-cysteine, 1 mm EDTA and 0·2% w/vBSA (Hernández et al. 1995). For APX activity, 20 mmascorbate was added. The homogenate was filtered throughtwo layers of cheesecloth and centrifuged at 2200 g for 30 sto pellet the chloroplast fraction. The supernatant was cen-trifuged at 12 000 g to discard mitochondria and peroxi-somes. Then, the 12 000 g supernatant was centrifuged for20 min at 82 000 g. The supernatant obtained was partiallypurified in columns containing Sephadex G-50 equilibratedwith the same buffer used for homogenization, and wasconsidered as the soluble fraction and used for variousassays.
Marker enzymes were used to measure the contamina-tion of the soluble fractions on both control and NaCl-treated plants. CuZn-SOD II activity, estimated by PAGE,according to Hernández et al. (1999), and glucose-6-phosphate dehydrogenase (G6PDH), assayed as describedby Löhr & Waller (1974), were used as chloroplasticmarkers. Phosphoenolpyruvate carboxylase (PEPC) wastaken as the cytosolic marker (Winter et al. 1982). Mito-chondrial contamination was analysed as the contributiondue to cytochrome c oxidase and Mn-SOD, whereas cata-lase activity was used as the peroxisomal marker (Jiménezet al. 1997). Triose phosphate isomerase (TPI) activity,present in chloroplasts (40%) and cytosol (60%) (Schar-renberger et al. 1985) was also assayed (Feierabend 1975).
APX, DHAR, MDHAR, GR and SOD were assayedaccording to Jiménez et al. (1997). The specific activityvalues for SOD,APX, DHAR, MDHAR and GR in solublefractions, of each cultivar under control and NaCl-stressconditions, were corrected for cross-contamination usingmarker enzymes. These corrections were performed usingthe marker activity giving the highest contribution in the82 000 g supernatant fractions, which under our experi-mental conditions was due to the chloroplastic CuZn-SOD II activity (see below). Therefore, this would give themost conservative valuations for activities in the super-natant fractions. Protein was estimated according to Bradford (1976).Ascorbate and dehydroascorbate, reducedand oxidized glutathione were determined by HPLC as pre-viously described (Jiménez et al. 1997).
For the separation of SOD isozymes, non-denaturingPAGE was performed on 10% w/v acrylamide gels using aBioRad mini-protean II dual slab cell (Hernández et al.1999).
Stomatal conductance measurements
Stomatal conductance was determined on fully expandedintact leaves with a portable porometer (model Licor 1600,Li-Cor Inc., Lincoln, Nebraska, USA), using six plants ineach experiment.
RNA extraction and Northern blots
Leaves were ground in liquid nitrogen and RNA prepara-tion was performed by a phenol/LiCl procedure according
© 2000 Blackwell Science Ltd, Plant, Cell and Environment, 23, 853–862

to Creissen & Mullineaux (1995). Poly(A+) mRNA was isolated from total RNA by oligo(dT) cellulose affinitychromatography (Sambrook, Fricsch & Maniatis 1989). Pealeaves (30 g) from both cultivars grown in the absence(control) and in the presence of 70 mm NaCl were used.Poly(A+) mRNA (up to 5 mg) was separated on 1·5% (w/v)agarose gels after formaldehyde/formamide denaturationin MOPS buffer, and transferred to nylon membranes(Boehringer Mannheim) according to Sambrook et al.(1989). Membranes were pre-hybridized and hybridized at65 °C in a 0·9 m NaCl, 10% dextran sulphate and 1% SDSsolution containing 50 mg mL-1 salmon sperm DNA. Pre-hybridization was carried out for at least 6 h and hybridiza-tion was carried out overnight. 32P-labelled DNA probeswere prepared by the method of Feinberg & Vogelstein(1983). Membranes were washed once in 2¥ SSC, 0·1% SDSat room temperature for 15 min and twice in 1¥ SSC, 1%SDS at 65 °C for 15 min. Filters were autoradiographedusing Fuji X-ray film with an intensifying screen at –80 °C.Signal intensities were estimated from autoradiogramsusing a Shimadzu CS-9000 densitometer.
The cDNA probes used were those encoding mitochon-drial Mn-SOD (Wong-Vega, Burke & Allen 1991), cytoso-lic APX (Santos et al. 1996), cytosolic GR (Stevens et al.1997) and PHGPX (Mullineaux et al. 1998) from pea leaves,stromatic APX from spinach (Ishikawa et al. 1995), andcDNA probes encoding cytosolic and chloroplastic CuZn-SOD from Pinus sylvestris were obtained from Dr S.Karpinski (Karpinski et al. 1992).
RESULTS
In previous work, by using different growth parameters(fresh and dry weight of leaves and leaf area) of plants in70 mm NaCl-containing grown medium, two pea cultivarswere designated as NaCl-tolerant (cv Granada) and NaCl-sensitive (cv Challis), respectively (Hernández et al. 1993,1995).These parameters have now been supplemented withstomatal conductance data which confirm the designationof NaCl-tolerant and NaCl-sensitive genotypes. After 15 dof NaCl treatment, stomatal conductance was reduced by 13% in the NaCl-tolerant cultivar compared with adecrease of approximately 88% in the sensitive cultivar(Fig. 1).
The short-term response to NaCl treatment (up to 24 hexposure) of the total activities of antioxidant enzymes and the concentrations and reduced/oxidized ratios of glutathione and ascorbate, in foliar cell-free extracts or in the soluble fraction, revealed no statistically significantresponses or differences between the NaCl-tolerant andNaCl-sensitive cultivars (data not shown). In contrast, long-term (15 d) exposure to 70 mm NaCl did reveal responseswhen measured in total leaf extracts (data not shown), andmore so when determined from soluble fractions (Tables 3and 4).
Cross-contamination of the soluble fraction by other sub-cellular organelles was estimated, using marker enzymes,in both control and NaCl-treated plants. When pea leaves
Salt tolerance and antioxidant defences in pea 855
were subjected to differential centrifugation, the activity ofthe marker enzymes indicated a relatively clean separationof cytosolic fraction (82 000 g supernatant) from organellefractions (chloroplasts, mitochondria, and peroxisomes).In general, the percentage contamination values were quitesimilar in both treatments (control and NaCl-treatedplants) (Tables 1 and 2). In NaCl-tolerant plants, the per-centage contaminations of the soluble fractions (82 000 gsupernatant) by the chloroplastic CuZn-SOD II were 28and 21·4% (Tables 1 and 2) in control and NaCl-treatedplants, respectively. In NaCl-sensitive plants, CuZn-SOD IIcontamination values increased slightly in soluble fractionsof NaCl-stressed plants in comparison to their values incontrol plants (23·75 and 27·7%, respectively) (Tables 1 and2). This different pattern agrees with the chloroplastintegrity results previously found in both cultivars underNaCl-stress conditions (Hernández et al. 1995).
Cytosolic PEPC showed up to 200% recovered activityin the 82 000 g supernatant fractions from control plants,showing a good correlation with the corresponding fraction(Table 1). Similar PEPC recovery values were obtained onNaCl-treated tolerant plants, and these activity values wereslightly lower in NaCl-treated sensitive plants (Table 2).
G6PDH and TPI activities were found at varying degreesin the 82 000 g supernatant from control plants, up to23–30% and 68–80%, respectively (Table 1). In NaCl-treated plants, the activity values of both enzymes did notshow significant changes in relation to control plants, being28–30% and 68–78%, respectively (Table 2). It has beenreported that TPI is located in the chloroplasts (about 40%)and in the cytosol (about 60%) (Scharrenberger et al. 1985),whereas the bulk of G6PDH has been identified in chloro-plasts (80%), and is also present in the cytosol (10%) andperoxisomes (10%) (Corpas et al. 1998).
© 2000 Blackwell Science Ltd, Plant, Cell and Environment, 23, 853–862
Figure 1. Leaf stomatal conductance (mmol m–2 s–1) of Pisumsativum plants after a growth period of 15 d in nutrient solutionscontaining 0 or 70 mm NaCl. Each histogram is the mean of sixmeasurements (± SE). Differences from control values weresignificant at P < 0·001 (c) according to the Duncan’s multiplerange test.
856 J. A. Hernández et al.
© 2000 Blackwell Science Ltd, Plant, Cell and Environment, 23, 853–862
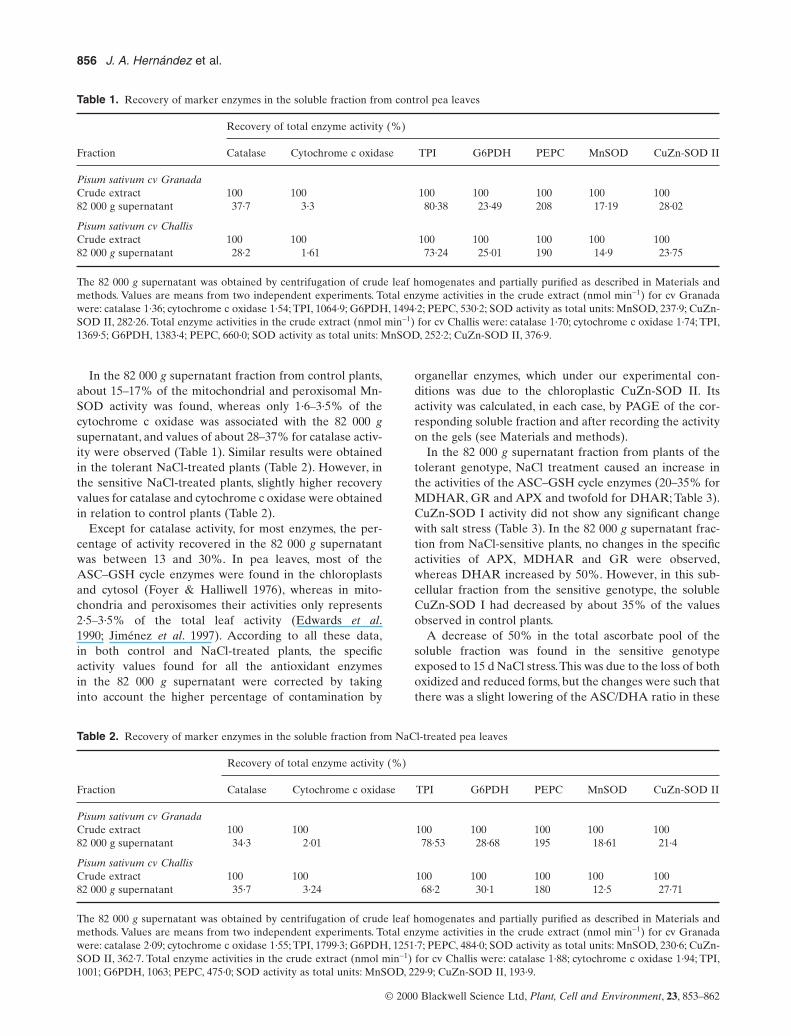
Table 1. Recovery of marker enzymes in the soluble fraction from control pea leaves
Recovery of total enzyme activity (%)
Fraction Catalase Cytochrome c oxidase TPI G6PDH PEPC MnSOD CuZn-SOD II
Pisum sativum cv GranadaCrude extract 100 100 100 100 100 100 10082 000 g supernatant 37·7 3·3 80·38 23·49 208 17·19 28·02
Pisum sativum cv ChallisCrude extract 100 100 100 100 100 100 10082 000 g supernatant 28·2 1·61 73·24 25·01 190 14·9 23·75
The 82 000 g supernatant was obtained by centrifugation of crude leaf homogenates and partially purified as described in Materials andmethods. Values are means from two independent experiments. Total enzyme activities in the crude extract (nmol min-1) for cv Granadawere: catalase 1·36; cytochrome c oxidase 1·54; TPI, 1064·9; G6PDH, 1494·2; PEPC, 530·2; SOD activity as total units: MnSOD, 237·9; CuZn-SOD II, 282·26. Total enzyme activities in the crude extract (nmol min-1) for cv Challis were: catalase 1·70; cytochrome c oxidase 1·74; TPI,1369·5; G6PDH, 1383·4; PEPC, 660·0; SOD activity as total units: MnSOD, 252·2; CuZn-SOD II, 376·9.
Table 2. Recovery of marker enzymes in the soluble fraction from NaCl-treated pea leaves
Recovery of total enzyme activity (%)
Fraction Catalase Cytochrome c oxidase TPI G6PDH PEPC MnSOD CuZn-SOD II
Pisum sativum cv GranadaCrude extract 100 100 100 100 100 100 10082 000 g supernatant 34·3 2·01 78·53 28·68 195 18·61 21·4
Pisum sativum cv ChallisCrude extract 100 100 100 100 100 100 10082 000 g supernatant 35·7 3·24 68·2 30·1 180 12·5 27·71
The 82 000 g supernatant was obtained by centrifugation of crude leaf homogenates and partially purified as described in Materials andmethods. Values are means from two independent experiments. Total enzyme activities in the crude extract (nmol min-1) for cv Granadawere: catalase 2·09; cytochrome c oxidase 1·55; TPI, 1799·3; G6PDH, 1251·7; PEPC, 484·0; SOD activity as total units: MnSOD, 230·6; CuZn-SOD II, 362·7. Total enzyme activities in the crude extract (nmol min-1) for cv Challis were: catalase 1·88; cytochrome c oxidase 1·94; TPI,1001; G6PDH, 1063; PEPC, 475·0; SOD activity as total units: MnSOD, 229·9; CuZn-SOD II, 193·9.
In the 82 000 g supernatant fraction from control plants,about 15–17% of the mitochondrial and peroxisomal Mn-SOD activity was found, whereas only 1·6–3·5% of thecytochrome c oxidase was associated with the 82 000 gsupernatant, and values of about 28–37% for catalase activ-ity were observed (Table 1). Similar results were obtainedin the tolerant NaCl-treated plants (Table 2). However, inthe sensitive NaCl-treated plants, slightly higher recoveryvalues for catalase and cytochrome c oxidase were obtainedin relation to control plants (Table 2).
Except for catalase activity, for most enzymes, the per-centage of activity recovered in the 82 000 g supernatantwas between 13 and 30%. In pea leaves, most of theASC–GSH cycle enzymes were found in the chloroplastsand cytosol (Foyer & Halliwell 1976), whereas in mito-chondria and peroxisomes their activities only represents2·5–3·5% of the total leaf activity (Edwards et al.1990; Jiménez et al. 1997). According to all these data,in both control and NaCl-treated plants, the specific activity values found for all the antioxidant enzymes in the 82 000 g supernatant were corrected by taking into account the higher percentage of contamination by
organellar enzymes, which under our experimental con-ditions was due to the chloroplastic CuZn-SOD II. Its activity was calculated, in each case, by PAGE of the cor-responding soluble fraction and after recording the activityon the gels (see Materials and methods).
In the 82 000 g supernatant fraction from plants of thetolerant genotype, NaCl treatment caused an increase inthe activities of the ASC–GSH cycle enzymes (20–35% forMDHAR, GR and APX and twofold for DHAR; Table 3).CuZn-SOD I activity did not show any significant changewith salt stress (Table 3). In the 82 000 g supernatant frac-tion from NaCl-sensitive plants, no changes in the specificactivities of APX, MDHAR and GR were observed,whereas DHAR increased by 50%. However, in this sub-cellular fraction from the sensitive genotype, the solubleCuZn-SOD I had decreased by about 35% of the valuesobserved in control plants.
A decrease of 50% in the total ascorbate pool of thesoluble fraction was found in the sensitive genotypeexposed to 15 d NaCl stress.This was due to the loss of bothoxidized and reduced forms, but the changes were such thatthere was a slight lowering of the ASC/DHA ratio in these
plants. In NaCl-tolerant plants, both ASC and DHA levelsalso fell, but less so and there was no change in theASC/DHA ratio (Table 4).
In the soluble fraction of NaCl-treated sensitive plants,the glutathione pool decreased by 60% compared to thelevel in the controls (Table 4). This fall was due to thedecline of both oxidized (GSSG) and reduced (GSH) formsof glutathione. In the tolerant cultivar, the soluble GSSGcontent fell to almost zero after salt treatment, althoughmore than 50% of the initial soluble GSH remained(Table 4). Thus, in contrast to ascorbate, there were few differences observed in the salt-induced changes in glu-tathione concentrations between sensitive and tolerantgenotypes.
Northern blot analysis of the steady-state levels ofmRNAs encoding the above antioxidant enzymes showedthat tolerant plants responded to long-term NaCl treatmentby increasing mitochondrial Mn-SOD (threefold), cytosolicAPX (threefold), cytosolic GR (fivefold) and chloroplasticCuZn-SOD II (fivefold) transcript levels (Fig. 2).The levelsof some mRNAs that were screened did not respond to salt stress in either cultivar, for example stromatic APX andcytosolic CuZn-SOD (Fig. 2) and chloroplastic GR (data not shown). In the case of cytosolic CuZn-SOD, itstranscript level even decreased in the NaCl-treated geno-type (Fig. 2). Similarly, PHGPX mRNA levels increased
Salt tolerance and antioxidant defences in pea 857
considerably (threefold) in the leaves of tolerant plants butnot in sensitive plants (Fig. 2).
DISCUSSION
The study reported here was an extension of previousstudies aimed at understanding the relationship betweentolerance to salt stress and oxidative stress.A first approachis to observe which component enzymes of the antioxidantdefences of the plant are regulated in their expression. Theimportance of components of the antioxidant defences canbe further assessed using the approach of comparing culti-vars differentially responsive to long-term NaCl stress.
The effect of salinity on stomatal conductance has been shown to be much more dramatic in NaCl-sensitivethan in NaCl-tolerant plants. The data for the NaCl-sensitive genotype (Fig. 1) are in agreement with stomatalconductance data from salt-stressed spinach and beanplants (Robinson, Dowton & Millhouse 1983; Brugnoli &Lauteri 1991). The increase in AOS production observed inchloroplasts from NaCl-sensitive plants in relation tochloroplasts from salt-tolerant ones (Hernández et al. 1995)may result from stomatal closure causing a decrease in theCO2 concentration inside the chloroplasts. This, in turn,might cause a decrease in the concentration of NADP+
available to accept electrons from photosystems I/II and
© 2000 Blackwell Science Ltd, Plant, Cell and Environment, 23, 853–862
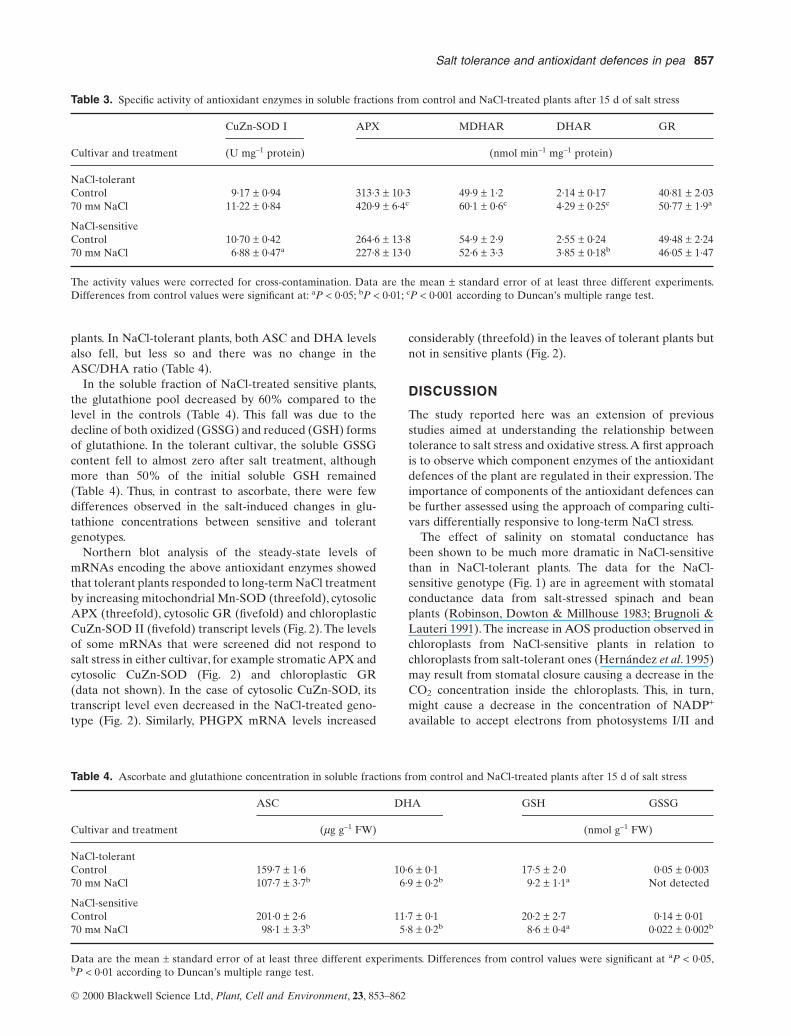
Table 3. Specific activity of antioxidant enzymes in soluble fractions from control and NaCl-treated plants after 15 d of salt stress
CuZn-SOD I APX MDHAR DHAR GR
Cultivar and treatment (U mg–1 protein) (nmol min–1 mg–1 protein)
NaCl-tolerantControl 9·17 ± 0·94 313·3 ± 10·3 49·9 ± 1·2 2·14 ± 0·17 40·81 ± 2·0370 mm NaCl 11·22 ± 0·84 420·9 ± 6·4c 60·1 ± 0·6c 4·29 ± 0·25c 50·77 ± 1·9a
NaCl-sensitiveControl 10·70 ± 0·42 264·6 ± 13·8 54·9 ± 2·9 2·55 ± 0·24 49·48 ± 2·2470 mm NaCl 6·88 ± 0·47a 227·8 ± 13·0 52·6 ± 3·3 3·85 ± 0·18b 46·05 ± 1·47
The activity values were corrected for cross-contamination. Data are the mean ± standard error of at least three different experiments.Differences from control values were significant at: aP < 0·05; bP < 0·01; cP < 0·001 according to Duncan’s multiple range test.
Table 4. Ascorbate and glutathione concentration in soluble fractions from control and NaCl-treated plants after 15 d of salt stress
ASC DHA GSH GSSG
Cultivar and treatment (mg g–1 FW) (nmol g–1 FW)
NaCl-tolerantControl 159·7 ± 1·6 10·6 ± 0·1 17·5 ± 2·0 0·05 ± 0·00370 mm NaCl 107·7 ± 3·7b 6·9 ± 0·2b 9·2 ± 1·1a Not detected
NaCl-sensitiveControl 201·0 ± 2·6 11·7 ± 0·1 20·2 ± 2·7 0·14 ± 0·0170 mm NaCl 98·1 ± 3·3b 5·8 ± 0·2b 8·6 ± 0·4a 0·022 ± 0·002b
Data are the mean ± standard error of at least three different experiments. Differences from control values were significant at aP < 0·05,bP < 0·01 according to Duncan’s multiple range test.

858 J. A. Hernández et al.
thus initiate O2 reduction with the concomitant generationof AOS (Halliwell 1982).
Data on cytosolic components of the antioxidantenzymes obtained through differential centrifugation show considerable agreement (De Gara, de Pinto &Arrigoni 1997; Gardeström & Edwards 1983; Madamanchiet al. 1992).
Because most of the antioxidative enzymes measured inthis work (SOD and ASC–GSH cycle enzymes) are locatedmainly in the chloroplasts and the cytosol (Foyer & Halliwell 1976; Madamanchi et al. 1992), our major concernwas the chloroplastic contamination in the 82 000 g super-natant fractions. For that reason, we used, for the activitycorrections, the higher percentage of contamination found,which, under our experimental conditions was due, as mentioned above, to the chloroplastic CuZn-SOD II. Thisenzyme is a good marker since its localization has beenclearly demonstrated (Palma, Sandalio & del Río 1986;Hernández et al. 1995, 1999). Results on chloroplasticCuZn-SOD II activity indicated that the degree of chloro-plast contamination in the 82 000 g supernatant fractionsranged from 21 to 28%. It is important to note that in pea(cv Challis and cv Granada) chloroplasts, only CuZn-SOD II is present, and no Fe-SOD was found (Hernándezet al. 1995). These results contrast with those we havedescribed in a different pea cultivar (P. sativum cv Puget)in which two different SOD activity bands, Fe-SOD andCuZn-SOD II, representing 12–15% and 85–88%, respec-tively, of the total SOD activity, were identified (Hernández et al. 1999). In general, the amount of Fe-SODactivity in higher plants and more specifically in chloro-plasts, is very small, compared to that of CuZn-SOD whichis the predominant isozyme (Salin & Bridges 1982; Kurepaet al. 1997).
Little CuZn-SOD activity has been found in thylakoidmembranes (Ogawa et al. 1985) as the bulk of CuZn-SODis localized in the stroma. Thus, we assume that the relativeabundance of chloroplastic CuZn-SOD II is a good markerfor estimating chloroplast breakage.
G6PDH has been extensively used for a long time as acytosolic marker (Federico & Angelini 1986; Polle et al.1990; Ros Barceló 1998; Vanacker et al. 1998a,b). In oppo-sition to the use of G6PDH as a cytosolic marker, a recentpaper by Corpas et al. (1998) reported the presence of different G6PDH isozymes in pea leaves using differenttechniques (differential centrifugations, native PAGE,SDS–PAGE,Western blot analysis and electron microscopyimmunocytochemistry), and they demonstrated that, atleast in pea leaves, the enzyme is located mainly in chloro-plasts (80%), although it is also present in cytosol (10%)and peroxisomes (10%). So, this enzyme could be useful tocompare the data with those measured using chloroplasticCuZn-SOD II and to estimate the chloroplast contamina-tion in the soluble fractions. In fact, a percentage of G6PDHin the soluble fraction greater than 10% could be duemainly to the major chloroplastic isozyme. On the otherhand, the data for chloroplastic contamination in soluble
© 2000 Blackwell Science Ltd, Plant, Cell and Environment, 23, 853–862
Figure 2. Northern blot hybridization analysis of mRNA levelsof some antioxidant scavenging enzymes in poly(A+) RNA.Poly(A+) RNA (5 mg per lane) was isolated from leaves frompea plants (cv Granada and cv Challis) grown in the absence (C)or in the presence of 70 mm NaCl (S) for 15 d. The numericaldata above each lane represent the percentage of enzymaticactivity in the 82 000 g supernatant fractions in leaves of the twocultivars with and without salt treatment (see Table 3). For thespecific activity of mitochondrial Mn-SOD, chloroplastic CuZn-SOD and stromatic APX, see Hernández et al. (1993, 1995).These values are shown here to enable comparison of enzymeactivities with transcript levels for the same enzyme.

fractions calculated according to G6PDH activity werelower than those obtained using the chloroplastic markerCuZn-SOD II in both control and salt-treated plants.
Less than 3% of the cytochrome c oxidase activity was inthe 82 000 g supernatant preparations, indicating a lowdegree of contamination by mitochondrial membrane fractions. Cross-contamination by the mitochondrial andperoxisomal Mn-SOD activity was higher (about 18%).However, taken together, this suggests a low breakage of mitochondria. Cross-contamination by peroxisomal catalase was higher than that by mitochondrial enzymes(Tables 1 and 2). However, since in pea leaf peroxisomes asin mitochondria there is only a small amount (2·5–3·5%) ofthe ASC–GSH cycle enzymes (Edwards et al. 1990; Jiménezet al. 1997), the cross-contamination of the 82 000 g super-natant by peroxisomal components cannot be accountedfor the antioxidative enzyme activities detected in it, sincetheir highest contribution to soluble activities would beabout 1·1%.
Values of about 200% recovered activity of the cytosolicPEPC were found in the 82 000 g supernatant fractionsafter differential centrifugation (Tables 1 and 2), indicatinga high degree of correlation between the supernatant frac-tions and the cytosolic compartment (Winter et al. 1982;Stitt et al. 1989; Schinkel, Streller & Winsgle 1998). Takentogether, these results show that the purity of the 82 000 gsupernatant fractions is about 70–80%, which we considersufficient and more suitable than crude extracts to make ameaningful studies of the soluble antioxidant enzymes.There are other reports in which similar supernatant frac-tions were taken as the cytosolic fraction (Blinda et al.1997).
In the soluble fractions from the leaves of NaCl-tolerantpea plants, long-term exposure (15 d) to NaCl produced a significant increase in all of the antioxidant enzyme activities, whereas in salt-stressed sensitive plants, onlyDHAR activity increased, while SOD activity significantlydecreased (Table 3). These data should be contrasted withtotal leaf extracts which do not show a difference inresponse to salt stress between the two genotypes.Thus, thecytosolic compartment may be important in antioxidantresponses to NaCl. The antioxidant enzymes present in thiscompartment may contribute to the protection of othersubcellular compartments in a way similar to the responseof cytosol antioxidant defences to photoinhibitory light stress in Arabidopsis (Karpinski et al. 1997). Further-more, in both of these pea cultivars, NaCl stress causes an increase in O2·- generation in mitochondria togetherwith an increase in the H2O2 contents in chloroplasts(Hernández et al. 1993, 1995), and an increased leakage ofH2O2 into the cytosol from mitochondria and peroxisomeswas also suggested (Hernández et al. 1993; Corpas et al.1993). Thus, the cytosolic compartment may have to scav-enge AOS from all other subcellular compartments undercertain stress conditions. This could explain the increase inantioxidant enzyme activities in the soluble fractions in theNaCl-tolerant genotype.
Salt tolerance and antioxidant defences in pea 859
The up-regulation of DHAR activity in both NaCl-sensitive and NaCl-tolerant genotypes rather thanMDHAR activity (Table 3), suggests that, in pea subjectedto these stress conditions, ASC is regenerated via glu-tathione. This is in contrast to the situation described byMorán et al. (1994) who proposed that MDHAR was thekey detoxifying enzyme under drought stress conditions. Itmay be that DHAR activity could participate in ASCregeneration under conditions of severe stress whenMDHAR activity is limited by the availability of NADH(Asada & Takahashi 1987). Our data support this view.
In tobacco plants, salinity led to a two- to threefoldincrease in the activity of Fe-SOD, cytosolic and chloro-plastic Cu-Zn-SOD, APX, DHAR and GR activities (VanCamp et al. 1996). The increase in cytosolic DHAR activityhas also been observed in sorghum and sunflower plantsgrown under drought conditions (Zhang & Kirkham 1996).It was suggested that the increase in DHAR activity isinduced when cellular ascorbate contents are significantlydecreased (Arrigoni 1994). This increase in DHAR activitymay be required to sustain cycling of oxidized ascorbatewhen the flux through the ASC–GSH cycle is increased,as may occur in transgenic poplar plants with enhancedexpression of Fe-SOD (Arisi et al. 1998). It should be notedthat, neither in the tolerant nor in the sensitive plants didDHA accumulation take place (Table 4), which could havebeen due to the induction of soluble DHAR activity undersalt-stress conditions (Table 3). In NaCl-tolerant plants, theincrease in APX activity, which could contribute to ascor-bate oxidation, was paralleled by an increased capacity forascorbate regeneration via MDHAR, DHAR and GRactivities (Table 3). Nevertheless, the total soluble ascor-bate pool decreased and this was mainly due to the loss ofthe reduced form. In NaCl-stressed plants, total chloro-plastic ascorbate contents may be higher than in controlplants (Hernández et al. 1995; Meneguzzo et al. 1998). Thiscould suggest that an enhanced import by chloroplasts ofascorbate from the cytosol could be induced under saltstress conditions which could explain the loss of ascorbatein those soluble fractions.
Like ascorbate, the glutathione content of the solublefraction also decreased in NaCl-treated plants from bothcultivars (Table 4). This was despite the increase in GRactivity in NaCl-tolerant plants and the maintenance of thisactivity in sensitive ones. The loss of GSH was not attrib-utable to its oxidation to GSSG in spite of the increase inDHAR activity in both cultivars. The decline in glutathionecontents may be wholly or partly due to depressed rates of GSH synthesis, increased rates of degradation and/orGSH transport to other cell compartments or plant organs(Schneider, Martini & Rennenberg 1992; Jamai et al. 1996;Herschbach et al. 1998). Glutathione can be synthesized inboth the chloroplastic and cytosolic compartments (Noctoret al. 1998), but its degradation may be confined to thecytosol, and under some stress conditions oxidation of GSHis accompanied by net glutathione degradation (Noctor &Foyer 1998).Therefore, the loss of glutathione in the soluble
© 2000 Blackwell Science Ltd, Plant, Cell and Environment, 23, 853–862

fraction (Table 3) may be a consequence of export to otherlocations in the cell or the plant.
The increased export of H2O2 from mitochondria,chloroplasts and peroxisomes to the cytosol under salt-stress conditions (Hernández et al. 1993, 1995; Corpas et al.1993), could also be responsible for the decreased GSHlevels observed in the soluble fractions from both pea cultivars.
Salt stress is known to result in extensive lipid peroxida-tion (Hernández et al. 1995; Gosset et al. 1996), and there-fore GSH could be used for the conjugation of toxicmembrane lipid peroxidation products, in reactions catal-ysed by glutathione-S-transferase (GST; Pickett & Lu 1989;Marrs 1996). This could have contributed to the observeddecrease in glutathione contents in salt-stressed plants ofboth genotypes (Table 4). The recent report that salt toler-ance in transgenic tobacco seedlings can be enhanced byover-expressing a GST cDNA adds support to this sugges-tion (Roxas et al. 1997).
Interestingly, only in the NaCl-tolerant variety was therea concerted increase in the levels of transcript that en-code for some of the enzymes of antioxidant metabolism(Fig. 2). Collectively, these data and those concerning theincrease in activities of the enzymes and the changes inantioxidants of the soluble fraction (Tables 3 and 4), pointto an important role for the cytosol antioxidant defences asone determinant of salt tolerance in peas. However, stress-mediated changes in the abundance of a particular tran-script do not always correlate with corresponding changesin antioxidant protein levels and/or enzyme activities(Edwards et al. 1994; Mittler & Zilinskas 1994; Donahueet al. 1997; Mullineaux & Creissen 1997; Stevens et al. 1997).In keeping with this, in the long-term experiments, weobserved that the NaCl-mediated increases in mitochon-drial Mn-SOD, cytosolic CuZn-SOD, cytosolic APX,cytosolic GR and chloroplastic CuZn-SOD mRNA levels were much greater (Fig. 2) than the increases in the activi-ties of their corresponding enzymes (Table 3). It has beensuggested that changes in the levels of particular isoformsof such enzymes rather than changes in their total activitymay be more important (Edwards et al. 1994; Stevens et al.1997).
The PHGPX mRNA level was also significantly induced,but only in the NaCl-tolerant variety (Fig. 2). This couldexplain the low level of lipid peroxidation previouslydescribed in chloroplasts from salt-tolerant plants (Hernández et al. 1995), since these enzymes reduce lipidhydroperoxides to their corresponding alcohol using GSHas the electron donor (Ursini et al. 1995), and this class ofpea PHGPX has been shown to be located in the chloro-plast stroma (Mullineaux et al. 1998). PHGPX mRNAincreases in tissues of several plant species undergoingstress, including salt stress in suspension cells, heavy metalpoisoning, and infection by viral or bacterial pathogens(Holland et al. 1993; Criqui et al. 1992; Levine et al. 1994;Gueta-Dahan et al. 1997). Thus, the increase in the level ofPHGPX mRNA in several species subjected to a range of stresses suggests that common responses occur at the
860 J. A. Hernández et al.
cellular level, most likely mediated by oxidative damage tomacromolecules.
The increase in transcript levels shown in Fig. 2 could bedue to their de novo synthesis or to their decreased degra-dation. mRNA stability may be an important factor in reg-ulation of the response to salt stress, as shown in pea plantssubjected to drought and chilling stress (Mittler & Zilinskas1994; Stevens et al. 1997). Regulation of mRNA stability isan important point of control of expression of genes in stressresponses (Medhy & Brodl 1998). In pea plants recoveringfrom drought, although very high levels of APX transcriptswere present in the cell, these transcripts did not associatewith the polysome fraction and therefore did not participatein protein synthesis (Mittler & Zilinskas 1994). Further-more, in many plants, abiotic stress can affect translationalprocesses. For example, in carrot protoplasts, heat shock dis-rupts the function of both 5¢ cap and 3¢ poly(A) tail struc-tures, and in animals it has been shown that heat shockaffects the initiation of translation (Medhy & Brodl 1998).
In the NaCl-sensitive cultivar, levels of mitochondrialMnSOD and cytosolic CuZn-SOD transcript were notaffected by salt stress, but activities of mitochondrialMnSOD (Hernández et al. 1993) and cytosolic CuZn-SODdecreased by 35%, suggesting an inhibition of theseenzymes by NaCl as previously described in mesophyll protoplasts (Hernández, del Río & Sevilla 1994).
Stromal APX mRNAs remained at constant levels in bothpea cultivars. In previous work, we have reported that in iso-lated chloroplasts from tolerant plants NaCl stress pro-duced an increase in APX activity (Hernández et al. 1995).Similar results were described for cytosolic APX fromradish plants, suggesting that post-translational processesappear to play an important role in APX expression (Lópezet al. 1996), although it should be noted that cytosolic APXin pea did respond to salt stress in the tolerant cultivar(Fig. 2). These differences between increases or not inenzyme activities and mRNA levels again emphasize thatcontrol of the expression of plant antioxidant defences iscomplex and operates at a several levels.
ACKNOWLEDGEMENTS
This work was supported by grant PB95-0004 from theDGES (Spain). We would like to acknowledge the Depart-ment of Microbiology and Genetics of the University ofMurcia for the facilities to carry out Northern blot analy-sis. P.M. acknowledges the support of the Biotechnologyand Biological Sciences Research Council through theCore Strategic Grant to the John Innes Centre. P.M. and F.S.acknowledge financial support from the British Council,and J.A.H. acknowledges financial support from The RoyalSociety.
REFERENCES
Alscher R.G., Donahue J.L. & Cramer C.L. (1997) Reactive oxygenspecies and antioxidants: relationships in green cells. Physiolo-gia Plantarum 100, 224–223.
© 2000 Blackwell Science Ltd, Plant, Cell and Environment, 23, 853–862

Arisi A.C.M., Cornic G., Jouanin L. & Foyer C.F. (1998) Over-expression of iron superoxide dismutase in transformed poplarmodifies the regulation of photosynthesis at low CO2 partialpressures or following exposure to the prooxidant herbicidemethyl viologen. Plant Physiology 117, 565–574.
Arrigoni O. (1994) Ascorbate system in plant development.Journal of Bioenergetic and Biomembranes 26, 407–419.
Asada K. & Takahashi M. (1987) Production and scavenging ofactive oxygen in photosynthesis. In: Photoinhibition (eds D.J. Kyle, C.B. Osmond & C.J. Arntzen), pp. 227–287. ElsevierScience Publishers, Amsterdam.
Blinda A., Koch B., Ramanjulu S. & Dietz K.J. (1997) De novo syn-thesis and accumulation of apoplastic proteins in leaves of heavymetal-exposed barley seedlings. Plant, Cell and Environment 20,969–981.
Bohnert H.J. & Jensen R.G. (1996) Metabolic engineering forincreased salt tolerance. The next step. Australian Journal ofPlant Physiology 23, 661–667.
Brugnoli E. & Lauteri M. (1991) Effects of salinity on stonatal con-ductance, photosynthetic capacity, and carbon isotope discrimi-nation of NaCl-tolerant (Gossypium hirsutum L.) and saltsensitive (Phaseolus vulgaris L.) C3 non-halophytes. Plant Physiology 95, 628–635.
Corpas F.J., Gómez M., Hernández J.A. & del Río L.A. (1993)Metabolism of activated oxygen in peroxisomes from two Pisumsativum L. cultivars with different sensitivity to sodium chloride.Journal of Plant Physiology 141, 160–165.
Corpas F.J., Barroso J.B., Sandalio L.M., Distefano S., Palma J.M.,Lupiañez J.A. & del Río L.A. (1998) A dehydrogenase-mediatedrecycling system of NADPH in plant peroxisomes. BiochemicalJournal 330, 777–784.
Creissen G.P. & Mullineaux P.M. (1995) Cloning and characterisa-tion of glutathione reductase cDNAs and identification of twogenes encoding the tobacco enzyme. Planta 197, 422–425.
Criqui M.C., Jamet E., Parmentier Y., Marbach J., Durr A. & FleckJ. (1992) Isolation and characterization of a plant cDNA showinghomology to animal glutathione peroxidases. Plant MolecularBiology 18, 623–627.
De Gara L., de Pinto M.C. & Arrigoni O. (1997) Ascorbate syn-thesis and ascorbate peroxidase activity during the early stage ofwheat germination. Physiologia Plantarum 100, 894–900.
Donahue J.L., Okpodu C.M., Cramer C.L., Grabau E.A. & AslcherR.G. (1997) Responses of antioxidant to paraquat in pea leaves.Relationships to resistance. Plant Physiology 113, 249–247.
Edwards E.A., Rawsthone S. & Mullineaux P.M. (1990) Subcellu-lar distribution of multiple forms of glutathione reductase inleaves of pea (Pisum sativum L.). Planta 180, 278–284.
Edwards E.A., Enard C., Creissen G.P. & Mullineaux P.M. (1994)Synthesis and properties of glutathione reductase in stressedpeas. Planta 192, 137–143.
Federico R. & Angelini R. (1986) Occurrence of diamine oxidasein the apoplast of pea epicotyls. Planta 167, 300–302.
Feierabend J. (1975) Developmental studies on microbodies inwheat leaves. III. On the photocontrol of microbody develop-ment. Planta 123, 63–77.
Feinberg A.P. & Vogelstein B. (1983) A technique for radiolabellingDNA-restriction endonuclease fragments to high specific activ-ity. Analytical Biochemistry 132, 6–13.
Foyer C.H. & Halliwell B. (1976) Presence of glutathione and glu-tathione reductase in chloroplasts: a proposed role in ascorbicacid metabolism. Planta 133, 21–25.
Gardeström P. & Edwards G.E. (1983) Isolation of mitochondriafrom leaf tissue of Panicum miliaceum, a NAD-malic enzymetype C4 plant. Plant Physiology 71, 24–29.
Gosset D.R., Banks S.W., Millhollon E.P. & Lucas M.C. (1996)Antioxidant response to NaCl stress in a control and an NaCl-
Salt tolerance and antioxidant defences in pea 861
tolerant cotton cell line grown in the presence of paraquat,buthionine sulfoximine, and exogenous glutathione. Plant Physioogy 112, 803–809.
Gueta-Dahan Y.,Yaniv Z., Zilinskas B.A. & Ben-Hayyim G. (1997)Salt and oxidative stress: similar and specific responses and theirrelation to salt tolerance in Citrus. Planta 203, 460–469.
Halliwell B. (1982) The toxic effects of oxygen on plant tissues. In:Superoxide Dismutase,Vol 1 (ed. L.W. Oberley), pp. 89–124, CRCPress Inc., Boca Raton, Florida.
Hernández J.A., Corpas F.J., Gómez M., del Río L.A. & Sevilla F.(1993) Salt-induced oxidative stress mediated by activatedoxygen species in pea leaf mitochondria. Physiologia Plantarum89, 103–110.
Hernández J.A., del Río L.A. & Sevilla F. (1994) Salt stress-inducedchanges in superoxide dismutase isozymes in leaves and meso-phyll protoplasts from Vigna unguiculata L. Walp New Phytolo-gist 126, 37–44.
Hernández J.A., Olmos E., Corpas F.J., Sevilla F. & del Río L.A.(1995) Salt-induced oxidative stress in chloroplast of pea plants.Plant Science 105, 151–167.
Hernández J.A., Campillo A., Jiménez A., Alarcón J.J. & Sevilla F.(1999) Response of antioxidant systems and leaf water relationsto NaCl stress in pea plants. New Phytologist 141, 241–251.
Herschbach C., Jouanin L. & Rennenberg H. (1998) Overex-pression of gamma-glutamylcysteine synthetase, but not of glutathione synthetase, elevates glutathione allocation in thephloem of transgenic poplar trees. Plant and Cell Physiology 39,447–451.
Holland D., Ben-Hayyim G., Faltin Z., Camoin L., Strosberg A.D.& Eshdat Y. (1993) Molecular characterization of salt-stress-associated protein in citrus: protein and cDNA sequence homol-ogy to mammalian glutathione peroxidases. Plant MolecularBiology 21, 923–927.
Ishikawa T., Sakai K., Takeda T. & Shigeoka S. (1995) Cloning andexpression of cDNA encoding a new type of ascorbate peroxi-dase from spinach. FEBS Letters 367, 28–32.
Jamai A., Tommasini R., Martinoia E. & Delrot S. (1996) Charac-terization of glutathione uptake in broad bean leaf protoplasts.Plant Physiology 111, 1145–1152.
Jiménez A., Hernández J.A., del Río L.A. & Sevilla F. (1997) Evidence for the presence of the ascorbate–glutathione cycle inmitochondria and peroxisomes of pea leaves. Plant Physiology117, 275–284.
Jiménez A., Hernández J.A., Pastori G.M., del Río L.A. & SevillaF. (1998) The role of the ascorbate–glutathione cycle of mito-chondria and peroxisomes in the senescence of pea leaves. PlantPhysiology 118, 1327–1335.
Karpinski S., Wingsle G., Olsson O. & Hälgren J.E. (1992) Charac-terization of cDNAs encoding Cu,Zn-superoxide dismutases inScots pine. Plant Molecular Biology 18, 545–555.
Karpinski S., Escobar C., Karpinska B., Creissen G. & MullineuaxP.M. (1997) Photosynthetic electron transport regulates theexpression of cytosolic ascorbate peroxidase genes in Arabidop-sis during excess light stress. Plant Cell 9, 627–640.
Kurepa J., Hèrouart D., Van Montagu M. & Inzè D. (1997) Differ-ential expression of Cu,Zn- and Fe-superoxide dismutase genesof tobacco during development, oxidative stress, and hormonaltreatments. Plant and Cell Physiology 38, 463–470.
Levine A., Tenhaken R., Dixon R. & Lamb C. (1994) H2O2 fromthe oxidative burst orchestrates the plant hypersensitive diseaseresistance response. Cell 79, 583–593.
Löhr G.W. & Waller H.D. (1974) Glucose-6-phosphate dehydroge-nase. In: Methods of Enzymatic Analysis, Vol 2 (ed. H.U.Bergmeyer), pp. 636–643, Academic Press, New York.
López F., Vansuyt G., Casse-Delbart F. & Fourcroy P. (1996) Ascor-bate peroxidase activity, not the mRNA level, is enhanced in salt-
© 2000 Blackwell Science Ltd, Plant, Cell and Environment, 23, 853–862

stressed Raphanus sativus plants. Physiologia Plantarum 97,13–20.
Madamanchi N.R., Anderson J.V., Alscher R.G., Cramer C.L.& Hess J.L. (1992) Purification of multiple forms of glutathionereductase from pea (Pisum sativum) seedling and enzyme levels in ozone-fumigated pea leaves. Plant Physiology 100,138–145.
Marrs K.A. (1996) The functions and regulation of glutathione-S-transferase in plants. Annual Review of Plant Physiology andPlant Molecular Biology 47, 127–158.
Medhy M.C. & Brodl M.R. (1998) The role of stress in regulationmRNA stability. In: A Look Beyond Transcription: MechanismsDetermining mRNA Stability and Translation in Plants (eds J. Bailey-Serres & R. Gallie), pp. 54–63, American Society ofPlant Physiologists, Rockville, Maryland.
Meneguzzo S., Sgherri C.L.M., Navari-Izzo F. & Izzo R. (1998)Stromal and thylakoid-bound ascorbate peroxidases in NaCltreated wheat. Physiologia Plantarum 104, 735–740.
Mittler R. & Zilinskas B.A. (1994) Regulation of pea cytosolicascorbate peroxidase and other antioxidant enzymes during theprogression of drought stress and following recovery fromdrought. Plant Journal 5, 397–405.
Morán J.F., Becana M., Iturbe-Ormaetxe I., Freschilla S., KlucasR.V. & Aparicio-Tejo P. (1994) Drought induces oxidative stressin pea plants. Planta 194, 346–352.
Mullineaux P.M. & Creissen G. (1997) Glutathione reductase: reg-ulation and role in oxidative stress. In: Oxidative Stress and theMolecular Biology of Antioxidant Defenses (ed. J.G. Scandalios),pp. 667–713, Cold Spring Harbor Laboratory Press, Cold SpringHarbor, New York.
Mullineaux P.M., Karpinski S., Jiménez A., Cleary S., Robinson C.& Creissen G. (1998) Plastid-targeted glutathione peroxidaseprovides evidence for an ascorbate-independent glutathioneredox cycle in plant chloroplast. Plant Journal 13, 375–379.
Noctor G. & Foyer C. (1998) Ascorbate and glutathione: keepingactive oxygen under control. Annual Review of Plant Physiologyand Plant Molecular Biology 49, 249–279.
Noctor G., Arisi A.C.M., Jouanin L. & Foyer C.H. (1998) Manipu-lation of glutathione and amino acid biosynthesis in chloroplasts.Plant Physiology 118, 471–482.
Ogawa K., Kanematsu S., Takabe K. & Asada K. (1985) Attach-ment of Cu,Zn-superoxide dismutase to thylakoid membranesat the site of superoxide generation (PSI) in spinach chloroplast:detection by immuno-gold labeling after rapid freezing and sub-stitution method. Plant and Cell Physiology 36, 565–573.
Palma J.M., Sandalio L.M. & del Río L.A. (1986) Manganesesuperoxide dismutase and higher plant chloroplasts: a reap-praisal of controverted cellular localization. Journal of PlantPhysiology 125, 417–429.
Picket C.G. & Lu A.Y.H. (1989) Glutathione-S-transferase: genestructure, regulation and biological function. Annual Review ofBiochemistry 58, 743–764.
Polle A., Chakrabarti K., Schümann W. & Rennenberg H. (1990)Composition and properties of hydrogen peroxide decomposingsystems in extracellular and total extracts from needles ofNorway spruce (Picea abies L., Karst.). Plant Physiology 94,312–319.
Robinson S.P., Dowton J.S. & Millhouse J.A. (1983) Photosynthe-sis and ion content of leaves and isolated chloroplasts of salt-stressed spinach. Plant Physiology 73, 238–242.
862 J. A. Hernández et al.
Ros Barceló A. (1998) The generation of H2O2 in the xylem ofZinnia elegans is mediated by an NADPH-oxidase-like enzyme.Planta 207, 207–216.
Roxas V.P., Smith R.K., Allen E.R. & Allen R.D. (1997) Over-expression of glutathione-S-transferase/glutathione peroxidaseenhances the growth of transgenic tobacco seedlings duringstress. Nature Biotechnology 15, 968–991.
Salin M.L. & Bridges S.M. (1982) Isolation and characterization ofan iron-containing superoxide dismutase from water lily, Nupharluteum. Plant Physiology 69, 161–165.
Sambrook J., Fricsch T. & Maniatis F. (1989) Molecular Cloning: ALaboratory Manual, 2nd edn. Cold Spring Harbor LaboratoryPress, Cold Spring Harbor, New York.
Santos M., Gousseau H., Lister C., Foyer C., Creissen G. &Mullineaux P.M. (1996) Cytosolic ascorbate peroxidase fromArabidopsis thaliana L. is encoded by a small multigene family.Planta 198, 64–69.
Schinkel H., Streller S. & Winsgle G. (1998) Multiple forms ex-tracellular superoxide dismutase in needles, stem tissues andseedlings of Scots pine. Journal of Experimental Botany 49,931–936.
Schnarrenberger C., Flechner A. & Martin W. (1985) Enzymaticevidence for a complete oxidative pentose phosphate pathwayin the cytosol of spinach leaves. Plant Physiology 108, 609–614.
Schneider A., Martini N. & Rennenberg H. (1992) Reduced glutathione (GSH) transport into cultured tobacco cells. PlantPhysiology and Biochemistry 30, 29–38.
Stevens R., Creissen G. & Mullineaux P.M. (1997) Cloning andcharacterization of a cytosolic glutathione reductase cDNA frompea (Pisum sativum L.) and its expression in response to stress.Plant Molecular Biology 35, 641–654.
Stitt M., Lilley R., McC., Gerhardt R. & Heldt H.W. (1989) Metabo-lite levels in specific cells and subcellular compartments of plantleaves. Methods in Enzymology 174, 518–552.
Ursini F., Miaorino M., Brigelius-Flohé R., Aumann K.D., RoveriA., Schomburg D. & Flohé L. (1995) Diversity of glutathioneperoxidases. Methods in Enzymology 252, 38–53.
Van Camp W., Capiau K., Van Montagu M., Inzè D. & Slooten L.(1996) Enhacement of oxidative stress tolerance in transgenictobacco plants overproducing Fe-superoxide dismutase inchloroplasts. Plant Physiology 112, 1703–1714.
Vanacker H., Carver T.L.W. & Foyer C.H. (1998a) Pathogen-induced changes in the antioxidant status of the apoplast inbarley leaves. Plant Physiology 117, 1103–1114.
Vanacker H., Harbinson J., Carver T.L.W. & Foyer C.H. (1998b)Antioxidant defences of the apoplast. Protoplasma 205, 129–140.
Winter K., Foster J.C., Edwards G.E. & Holtum J.A.M. (1982)Intracellular localization of enzymes of carbon metabolism inMesembryanthemum crystallinum exhibiting C3 photosyntheticcharacteristics or performing crassulacean acid metabolism.Plant Physiology 69, 300–307.
Wong-Vega L., Burke J.J. & Allen R.D. (1991) Isolation andsequence of a cDNA that encodes pea manganese superoxidedismutase. Plant Molecular Biology 17, 1271–1274.
Zhang J. & Kirkham M.B. (1996) Enzymatic responses of the ascorbate-glutathione cycle to drought in sorgum and sun-flower plants. Plant Science 113, 139–147.
Received 11 February 2000; received in revised form 10 March 2000;accepted for publication 10 March 2000
© 2000 Blackwell Science Ltd, Plant, Cell and Environment, 23, 853–862




















