
Expression of an antisense Datura stramonium S-adenosylmethionine decarboxylase cDNA in tobacco:...
13
Journal of Plant Physiology 162 (2005) 559—571 Expression of an antisense Datura stramonium S-adenosylmethionine decarboxylase cDNA in tobacco: changes in enzyme activity, putrescine-spermidine ratio, rhizogenic potential, and response to methyl jasmonate Patrizia Torrigiani a, , Sonia Scaramagli a , Vanina Ziosi a , Melinda Mayer b , Stefania Biondi a a Dipartimento di Biologia e.s., University of Bologna, Via Irnerio 42, 40126 Bologna, Italy b Department of Genetics and Microbiology, Institute of Food Research, Colney Park, Norwich, UK Received 21 April 2004; accepted 25 October 2004 Summary S-adenosylmethionine decarboxylase activity (SAMDC; EC 4.1.1.21) leads to spermi- dine and spermine synthesis through specific synthases which use putrescine, spermidine and decarboxylated S-adenosylmethionine as substrates. In order to better understand the regulation of polyamine (PA), namely spermidine and spermine, biosynthesis, a SAMDC cDNA of Datura stramonium was introduced in tobacco (Nicotiana tabacum L. cv. Xanthi) in antisense orientation under the CaMV 35S promoter, by means of Agrobacterium tumefaciens and leaf disc transformation. The effect of the genetic manipulation on PA metabolism, ethylene production and plant morphology was analysed in primary transformants (R0), and in the transgenic progeny (second generation, R1) of self-fertilised primary transformants, relative to empty vector-transformed (pBin19) and wild-type (WT) controls. All were maintained in vitro by micropropagation. Primary transformants, which were confirmed by Southern and northern analyses, efficiently transcribed the antisense SAMDC gene, but SAMDC activity and PA titres did not change. By contrast, in most transgenic R1 shoots, SAMDC activity was remarkably lower than in controls, and the putrescine-to-spermidine ratio was altered, mainly due to increased putrescine, even though putrescine ARTICLE IN PRESS www.elsevier.de/jplph KEYWORDS Antisense transformation; SAMDC; Polyamines; Methyl jasmonate; Rhizogenesis; Tobacco 0176-1617/$ - see front matter & 2005 Elsevier GmbH. All rights reserved. doi:10.1016/j.jplph.2004.10.008 Abbreviations: ADC, arginine decarboxylase; DAO, diamine oxidase; dcSAM, decarboxylated S-adenosylmethionine; ODC, ornithine decarboxylase; MJ, methyl jasmonate, PA; polyamines; PCA, perchloric acid; PCR, polymerase chain reaction; SAM, S-adenosylmethionine; SAMDC, S-adenosylmethionine decarboxylase, WT, wild-type Corresponding author. Tel.: +39051 2091291; fax: +39051 242576. E-mail address: [email protected] (P. Torrigiani).
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Expression of an antisense Datura stramonium S-adenosylmethionine decarboxylase cDNA in tobacco:...

ARTICLE IN PRESS
Journal of Plant Physiology 162 (2005) 559—571
KEYWORDAntisensetransformaSAMDC;PolyamineMethyl jasRhizogeneTobacco
0176-1617/$ - sdoi:10.1016/j.
Abbreviationornithine decaS-adenosylmet�CorrespondE-mail addr
www.elsevier.de/jplph
Expression of an antisense Datura stramoniumS-adenosylmethionine decarboxylase cDNAin tobacco: changes in enzyme activity,putrescine-spermidine ratio, rhizogenic potential,and response to methyl jasmonate
Patrizia Torrigiania,�, Sonia Scaramaglia, Vanina Ziosia,Melinda Mayerb, Stefania Biondia
aDipartimento di Biologia e.s., University of Bologna, Via Irnerio 42, 40126 Bologna, ItalybDepartment of Genetics and Microbiology, Institute of Food Research, Colney Park, Norwich, UK
Received 21 April 2004; accepted 25 October 2004
S
tion;
s;monate;sis;
ee front matter & 200jplph.2004.10.008
s: ADC, arginine decarboxylase; MJ, methylhionine; SAMDC, S-adeing author. Tel.: +39 0ess: patrizia.torrigian
SummaryS-adenosylmethionine decarboxylase activity (SAMDC; EC 4.1.1.21) leads to spermi-dine and spermine synthesis through specific synthases which use putrescine,spermidine and decarboxylated S-adenosylmethionine as substrates. In order tobetter understand the regulation of polyamine (PA), namely spermidine and spermine,biosynthesis, a SAMDC cDNA of Datura stramonium was introduced in tobacco(Nicotiana tabacum L. cv. Xanthi) in antisense orientation under the CaMV 35Spromoter, by means of Agrobacterium tumefaciens and leaf disc transformation. Theeffect of the genetic manipulation on PA metabolism, ethylene production and plantmorphology was analysed in primary transformants (R0), and in the transgenic progeny(second generation, R1) of self-fertilised primary transformants, relative to emptyvector-transformed (pBin19) and wild-type (WT) controls. All were maintained in vitroby micropropagation. Primary transformants, which were confirmed by Southern andnorthern analyses, efficiently transcribed the antisense SAMDC gene, but SAMDCactivity and PA titres did not change. By contrast, in most transgenic R1 shoots, SAMDCactivity was remarkably lower than in controls, and the putrescine-to-spermidineratio was altered, mainly due to increased putrescine, even though putrescine
5 Elsevier GmbH. All rights reserved.
rboxylase; DAO, diamine oxidase; dcSAM, decarboxylated S-adenosylmethionine; ODC,jasmonate, PA; polyamines; PCA, perchloric acid; PCR, polymerase chain reaction; SAM,nosylmethionine decarboxylase, WT, wild-type51 2091291; fax: +39 051 [email protected] (P. Torrigiani).

ARTICLE IN PRESS
P. Torrigiani et al.560
oxidising activity (diamine oxidase, EC 1.4.3.6) did not change relative to controls.Despite the reduction in SAMDC activity, the production of ethylene, which shares withPAs the common precursor SAM, was not influenced by the foreign gene. Some plantswere transferred to pots and acclimatised in a growth chamber. In these in vivo-grownsecond generation transgenic plants, at the vegetative stage, SAMDC activity wasscarcely reduced, and PA titres did not change. Finally, the rhizogenic potential of invitro-cultured leaf explants excised from antisense plants was significantly diminishedas compared with WTones, and the response to methyl jasmonate, a stress-mimickingcompound, in terms of PA conjugation, was higher and differentially affected intransgenic leaf discs relative to WT ones. The effects of SAMDC manipulation arediscussed in relation to plant generation, culture conditions and response to stress.& 2005 Elsevier GmbH. All rights reserved.
Introduction
Polyamines (PAs) are multivalent cations presentin all living organisms, which regulate chromatinorganisation, transcription and translation (Cohen1998); therefore, they are required in cell divisionand differentiation (Bagni and Torrigiani 1992),including programmed cell death (Serafini-Fracas-sini et al. 2002), and stress responses (Bouchereauet al. 1999). Their synthesis starts from the aminoacids arginine and ornithine, which lead to putres-cine, either indirectly via arginine decarboxylase(ADC), or directly via ornithine decarboxylase(ODC). Methionine is converted to S-adenosylmethionine (SAM), which is primarily a methyldonor and also an ethylene precursor (Ravanel etal. 1998), and then to decarboxylated SAM (dcSAM)via the key enzyme S-adenosylmethionine decar-boxylase (SAMDC, Bennett et al. 2002). DcSAM isthe substrate, with putrescine and spermidine, forsynthases responsible for spermidine and sperminesynthesis, respectively (Hanzawa et al. 2000; Zhanget al. 2003).
PA titres are regulated at the transcriptional,post-transcriptional, translational and post-transla-tional levels (Cohen 1998), and by conjugation tohydroxycinnamic acids (Martin-Tanguy 1985). Thissuggests that, although a rapid PA response isrequired under some physiological or pathologicalconditions, their cellular homeostasis is important.Attempts have been made to alter endogenous PAlevels in various ways. Mutants were the first sourceof plants with ‘‘non-physiological’’ PA levels (Malm-berg and McIndoo 1983; Watson et al. 1998;Hanzawa et al. 2000), but their scarcity hashindered progress in understanding the regulationof their metabolism. Attempts to geneticallymodulate PA levels in plants have involved themanipulation of putrescine biosynthesis with theintroduction in tobacco, carrot and rice of hetero-logous ADC or ODC cDNA in sense orientationresulting in variable increases in the target enzyme
activity and/or product (Kumar and Minocha 1998;Mayer and Michael 2003; Trung-Nghia et al. 2003).With respect to spermidine and spermine, plantsand cultured cells overexpressing the SAMDC genealso display changes in enzyme activity and PAlevels compared with controls (Noh and Minocha1994; Kumar et al. 1996; Rafart Pedros et al. 1999;Quan et al. 2002; Thu-Hang et al. 2002; Waie andRajam 2003).
Less is known about antisense engineering of PAmetabolism. Rice callus lines containing an oat ADCcDNA in antisense orientation showed decreasedADC activity, which was reflected in depletedputrescine and spermidine levels, although thesechanges were reversed in regenerated plants(Capell et al. 2000). Under the CaMV 35S promoteror the tuber-specific patatin promoter, SAMDCantisense potato plants exhibited reduced SAMDCmRNA and activity, and spermidine levels (Kumar etal. 1996; Rafart Pedros et al. 1999).
It is well known that alteration in PA titres is oneof the ways in which plants respond to biotic andabiotic stress (Bouchereau et al. 1999; Walters2003). In fact, rice plants overexpressing ADC orSAMDC and tobacco overexpressing ODC haveexhibited enhanced osmotic stress resistance (Royand Wu 2001, 2002; Kumria and Rajam 2002; Waieand Rajam 2003). Jasmonates are well-knownsignalling molecules in stress responses and elicitorsof defense-related compounds (Creelman and Mullet1997). Among jasmonate-induced genes, there arethose responsible for PA biosynthesis, with theconsequent accumulation of PAs, especially theconjugated ones (Biondi et al. 2001, 2003).
The aim of the present work was to determinewhether downregulation of SAMDC can depletesteady-state levels of higher PAs. To this end, anantisense SAMDC cDNA of Datura stramonium, asolanaceous species close to tobacco, was intro-duced in tobacco under the CaMV 35S promoter.Primary transformants (R0) and the segregatingprogeny in the second generation (R1) were

ARTICLE IN PRESS
Expression of an antisense SAMDC in tobacco 561
analysed, relative to wild type (WT) and emptyvector-containing (pBin19) controls, for (i) SAMDCtranscript levels, (ii) SAMDC activity, and (iii) free,and soluble and insoluble conjugated PA titres. Theprogeny were also analysed for diamine oxidase(DAO) activity, ethylene production and morpholo-gical changes. In addition, the rhizogenic potentialof these plants, which is very sensitive to changesin the endogenous PA pool (Altamura et al. 1991;Couee et al. 2004), was evaluated using leafexplants induced to form roots in the presence ofauxin. Finally, to examine whether antisense plantsexhibited an altered PA metabolism in response to astress-mimicking compound relative to WT ones,leaf discs were incubated in a methyl jasmonate(MJ)-containing solution, and PA accumulation wasinvestigated.
Results showed that, in most of the transgenicprogeny, SAMDC activity and PA levels were stronglyaltered relative to controls, but only in micro-propagated shoots, and that explants from in vivo-grown plants displayed a depressed rhizogenicpotential, and a different response to MJ in termsof PA conjugation.
Table 1. Experimental design
Plant generation Type of analysis
Primary transformantsPlants IV, V and VI Southern blot
SAMDC northern blotSAMDC activityPolyamine titres
Second generationtransformantsIn vitro micropropagatedshoots
PCR
SAMDC activityPolyamine titresDAO activityEthylene productionMorphology
In vivo-grown plants SAMDC activityPolyamine titres
Explants from in vivoplants
Polyamine response to MJ
Rhizogenic potential
Materials and methods
Construction of the antisense vector, planttransformation and regeneration
A 1839-bp D. stramonium SAMDC cDNA (accessionnumber Y07768), sharing a 92% homology with thetobacco SAMDC gene sequence (accession numberAF033100; Franceschetti et al. 2001), was sub-cloned in an antisense orientation between acauliflower mosaic virus (CaMV) 35S RNA promoterwith duplicated enhancer sequences, and the CaMVtermination sequence in the vector pJIT60 (Guer-ineau et al. 1992). The promoter, antisense cDNAand terminator were then excised and ligated intothe KpnI and SalI sites of pBin19 (Bevan 1984). Thisconstruct and an empty pBin19 vector weretransferred into Agrobacterium tumefaciens strainLBA4404 by triparental mating as described byBevan (1984). Conventional leaf disc transforma-tion of Nicotiana tabacum L. cv. Xanthi XHFD8 andselection of plantlets was performed as describedpreviously (Mayer et al. 2001). Rooted primarytransformants (R0) were transferred to soil in agreenhouse. Young leaves from three SAMDC anti-sense (IV, V and VI) and three pBin19 control plantswere harvested one month later, at the vegetativestage, and stored at �80 1C until use for molecularand biochemical analyses.
Seed germination in vitro and plantcultivation
Seed obtained by self-pollination of WT plants,empty vector-transformed plants (pBin19) and theprimary transformants IV, V and VI were surface-sterilised with 5% commercial solution of sodiumhypochloride (6% active chloride) for 10min, rinsedfive times with sterile distilled water and germi-nated in glass jars on MS medium (Murashige andSkoog 1962). The latter was supplemented withkanamycin in order to select for transgenic progeny(R1). Selected seedlings of lines IV, V and VI weresubsequently micropropagated in vitro. Singleshoots were grown in test tubes on hormone-freeMS medium and subcultured every three weeks;they constituted the second generation of in vitromicropropagated shoots.
A number of these micropropagated shoots wereallowed to root; when they had 3–4 leaves andwere about 5-cm tall they were transferred to pots,acclimatised and grown up to flowering in a growthchamber (in vivo plants) under controlled light(Philips SON-Tand HPI-T, 400W, 320 mEm�2 s�1) andtemperature (day/night temperature of 25/19 1C)regimes, and with a 12-h photoperiod. Theyconstituted in vivo-grown second generationplants, and these were used for morphometricanalyses. Internode number, plant height, and leafsize were measured on three WTand pBin19 plants,four plants of line IV and five of line V atthe vegetative and flowering stages. In vitromicropropagated shoots, as well as leaves from

ARTICLE IN PRESS
P. Torrigiani et al.562
in vivo-grown plants at the vegetative stage werecollected and stored at �80 1C until use. Primary(R0) and second generation (R1) transformantswere used for biochemical, molecular and morpho-metric analyses, and leaves from in vivo-grown R1plants were used as source of leaf discs and leafexplants to study the response to MJ and therhizogenic potential according to the followingscheme (Table 1).
Molecular characterisation of transgenicplants
Genomic DNA was extracted and restricted withEcoRI for Southern blot analysis as describedpreviously (Mayer et al. 2001). The presence ofthe pBIN19 vector was confirmed by hybridisationto a 1955-bp PstI fragment encompassing the T-DNAregion (not shown). The transgene in SAMDCantisense R0 plants was identified by hybridisationto a 730-bp probe for the 35S promoter excisedfrom pJIT60 using KpnI and SmaI. Expression of thetransgene was confirmed by northern analysis usingthe D. stramonium SAMDC cDNA as a probe. Threeprimary transformants (plants IV, V, and VI) ex-hibiting the highest level of SAMDC antisense mRNAwere selected for further experiments.
In the transgenic progeny (R1), the presence ofthe D. stramonium sequence was confirmed bygenomic polymerase chain reaction (PCR) amplifi-cation. The latter was carried out utilising theReady-To-Go PCR beads kit (Amersham PharmaciaBiotech Italia, Milano, Italy) containing 200 mM eachdNTPs and 1.5 U of Taq DNA polymerase, with 1 mgDNA and 1 mM of each primer, in a total volumeof 25 ml. The forward primer was complementaryto the CaMV 35S promoter sequence (50-GGTGGCTCCTACAAATGCCA-30); the reverse primerwas complementary to the D. stramonium SAMDCtransgene (50-TAGAGATGTGTATGACTG-30). The PCRprogramme included 1 cycle at 94 1C for 3min,1 cycle at 75 1C for 3min, 25 cycles at 94 1C for 15 s,25 cycles at 55 1C for 45 s, 25 cycles at 72 1C for1.5min and a final cycle at 72 1C for 10min. The1-kb product was visualised by agarose gel electro-phoresis.
RNA extraction and northern blots
The presence of the transgene in primarytransformants (plants IV, V and VI) was alsoconfirmed by northern analysis. Total RNA wasextracted from ca. 200 to 300mg fresh weight leafsamples using an RNeasy Plant Mini Kit (Qiagen,
Hilden, Germany) according to the manufacturer’sinstructions. RNA (15 mg per track) was size-fractionated on a 1.2% agarose formaldehyde geland transferred in 10� SSC (20� SSC: 0.3M sodiumcitrate, 3.0M NaCl, pH 7) onto nylon membranes(Hybond-N, Amersham Pharmacia Biotech Italia)overnight according to standard methods(Sambrook et al. 1989). RNA was cross-linked tothe membrane by exposure to UV at 312 nm (VilberLourmat, Marne La Vallee, France) for 4min.
RNA blots were pre-hybridised at 42 1C for 2 h andhybridised at 42 1C for 18–20 h with a (32P)dCTP-labelled PCR fragment (random priming with aRediprime DNA labelling Kit, Amersham PharmaciaBiotech Italia) of D. stramonium SAMDC as de-scribed in Scaramagli et al. (1999). Followinghybridisation, membranes were washed as de-scribed previously, and then exposed to X-ray filmat �80 1C for 24 h with intensifying screen (DuPont,Wilmington, DE, USA). Equal loading of RNA on gelswas verified by reprobing stripped filters with anAntirrhinum majus ubiquitin gene probe.
Enzyme activity assays
The SAMDC activity (EC 4.1.1.21) assay wasperformed by a radiometric method, as previouslydescribed by Biondi et al. (2001). Enzyme extrac-tion procedures were carried out in an ice bath.Micropropagated shoots and leaves were homoge-nised with 5 volumes of 0.1M Tris–HCl buffer, pH7.6, containing 50 mM EDTA. The homogenate wascentrifuged at 20 000g for 30min at 4 1C; 0.2-mlaliquots of the supernatant were incubated with3.7 kBq S-adenosyl-L-[carboxyl-14C]methionine(2.07 TBqmol�1, Amersham Pharmacia Biotech Ita-lia) and 100 mM unlabelled SAM in a final assayvolume of 0.5ml, and the rate of 14CO2 evolutionfrom SAM decarboxylation evaluated. At the end ofa 2-h incubation at 37 1C, the 14CO2 trapped in 2MKOH (150 mM) was counted in a Beckman LS 7800beta counter.
For DAO activity assays (DAO, EC 1.4.3.6), shootswere homogenised on ice with a mortar and pestlein 3 volumes of 100mM potassium phosphate buffer,pH 8, containing 2mM dithiothreitol. After centri-fugation at 20 000g for 30min at 4 1C, supernatantswere used to measure DAO activity by a radiometricmethod based on the production of D1[14C]pyrrolinefrom [1,4-14C]putrescine as previously described(Biondi et al. 2001). The [14C]pyrroline formedduring a 30-min incubation at 37 1C was immedi-ately extracted in 0.5ml toluene; aliquots (100 ml)of the lipophilic phase were withdrawn and added

ARTICLE IN PRESS
Expression of an antisense SAMDC in tobacco 563
to 2ml scintillation liquid (Ultima Gold), and theradioactivity counted in a Beckman scintillationcounter. In both cases, protein content wasmeasured using Bradford’s method (Bradford1976), with bovine serum albumin as the standard.
Experiments were repeated at least once (3–4replicates each). Mean values were compared usinga one-way ANOVA to check for significant differ-ences in enzyme activity levels between transgenicindividuals and WT controls.
HPLC polyamine analysis
Leaves from primary transformants, in vitromicropropagated R1 shoots, and leaves and leafexplants from in vivo-grown R1 plants wereextracted in 10 volumes of 4% perchloric acid(PCA), and centrifuged at 20 000g for 30min at 4 1C.Aliquots (0.3ml) of the supernatant (PCA-solublefraction), and of the resuspended pellet (PCA-insoluble fraction) were subjected to acid hydro-lysis (6 N HCl at 110 1C overnight) in order to releasePAs from their PCA-soluble and -insoluble conju-gates, respectively. Aliquots (0.2ml) of the super-natant, containing free PAs, and of the hydrolysedsupernatant and resuspended pellet were dansy-lated, extracted in toluene and analysed by HPLC(PU-980 Jasco, Tokyo, Japan) on a reverse phaseC18 column (Spherisorb ODS2, 5-mm particle dia-meter, 4.6� 250mm, Waters, Wexford, Ireland)using a programmed acetonitrile:water step gradi-ent (Scaramagli et al. 1995). Eluted peaks weredetected by a spectrofluorometer (821-FP Jasco,excitation 365 nm, emission 510 nm), and theirareas were recorded and integrated relative tothose of standard PAs using the JCL6000 software(Jasco). Experiments were repeated at least once(3 replicates each) and, where appropriate, meanvalues were statistically analysed using a one-wayANOVA to check for significant differences betweentransgenic individuals and WT controls.
Ethylene measurement
Test tubes in which one week-old micropropa-gated shoots were growing were sealed with air-tight serum caps. After 18 h, 10-ml gas sampleswere withdrawn from the headspace and injectedinto a Dani DS 86.01 (Dani, Milano, Italy) packed-gas chromatograph equipped with a Poropak Qcolumn and a flame ionisation detector, as pre-viously described (Biondi et al. 1998). The carriergas was nitrogen, at a flow rate of 16mlmin�1.
Rhizogenesis from leaf explants
Mature leaves from three WT and four antisense(line V, plants C, E, F and G) in vivo-grown plantswere used as source of explants, which weresterilised with a diluted (10%) commercial solutionof sodium hypochloride, rinsed three times, andthen cultured on a root-inducing MS medium(Bellincampi et al. 1996) containing 2% sucrose,0.8% agar and 0.6 mM indoleacetic acid (IAA, Sigma-Aldrich, Milano, Italy). Leaf explants (2� 1 cm),bearing the mid-vein, were cultured (6 per Petridish) in the dark at 24 1C for 15 d. The time courseof rhizogenesis was evaluated macroscopically at 9,12 and 15 d in culture on 120–180 explants. Atculture end, the mean number of roots per explantand root length (total number of roots 500–800)were measured, and data statistically treated withthe one-way ANOVA test to separate WT fromtransgenic samples.
MJ treatment
Leaf discs (1-cm diameter) were excised, using acork borer, from fully expanded leaves of WT andline V transgenic in vivo-grown plants, and floatedin Petri dishes on 20ml of 10mM potassiumphosphate buffer (pH 6) containing or not (controls)10 mM MJ (Serva Electrophoresis, Heidelberg,Germany), as previously described (Biondi et al.2003). Since MJ was dissolved in ethanol, controlscontained the same concentration of ethanolpresent in MJ treatments. After 48 h of incubationat 2271 1C in the light (50 mmolm�2 s�1) with a 16/8 h light/dark photoperiod, leaf discs were col-lected, weighed, frozen in liquid nitrogen, andstored at �80 1C until further use.
Results
Primary transformants
The presence and expression of the D. stramo-nium SAMDC sequence in primary transformantswas demonstrated by Southern and northernanalyses. The 35S sequence was present in genomicDNA restricted with EcoRI from SAMDC antisenseplants (IV, V and VI), but absent from pBin19controls (Fig. 1A). The SAMDC probe cross-hybri-dised to tobacco SAMDC mRNA (Fig. 1B,C), butSAMDC antisense plants IV, V and VI also contained alarger message corresponding to the D. stramoniumsequence which was absent in RNA from pBin19controls. SAMDC activity, as well as free, and

ARTICLE IN PRESS
Figure 1. Southern analysis of the D. stramonium SAMDCgene (A) and SAMDC northern analysis (B, B0) in leavesfrom antisense SAMDC primary transformants (IV, V andVI) and empty vector-transformed (pBin19) tobaccoplants. Fifteen mg DNA and RNA were loaded per well.The Southern blot was hybridised to the 35S fragment (A)as described in Methods, and the northern to the D.stramonium SAMDC cDNA (B) or to the Antirrhinum majusubiquitin probe as a loading control (C).
Figure 2. PCR analysis for the presence of the D.stramonium SAMDC gene in antisense micropropagatedtobacco shoots of line IV, line V and line VI. Lettersrepresent different individuals of each line.* ¼ molecular-weight marker, + ¼ positive control,� ¼ negative control.
P. Torrigiani et al.564
soluble- and insoluble conjugated PAs exhibited nosignificant differences in SAMDC antisense primarytransformants relative to pBin19 plants (data notshown).
Second generation transformants
Enzyme activities, polyamine titres and ethyleneproduction in micropropagated shoots
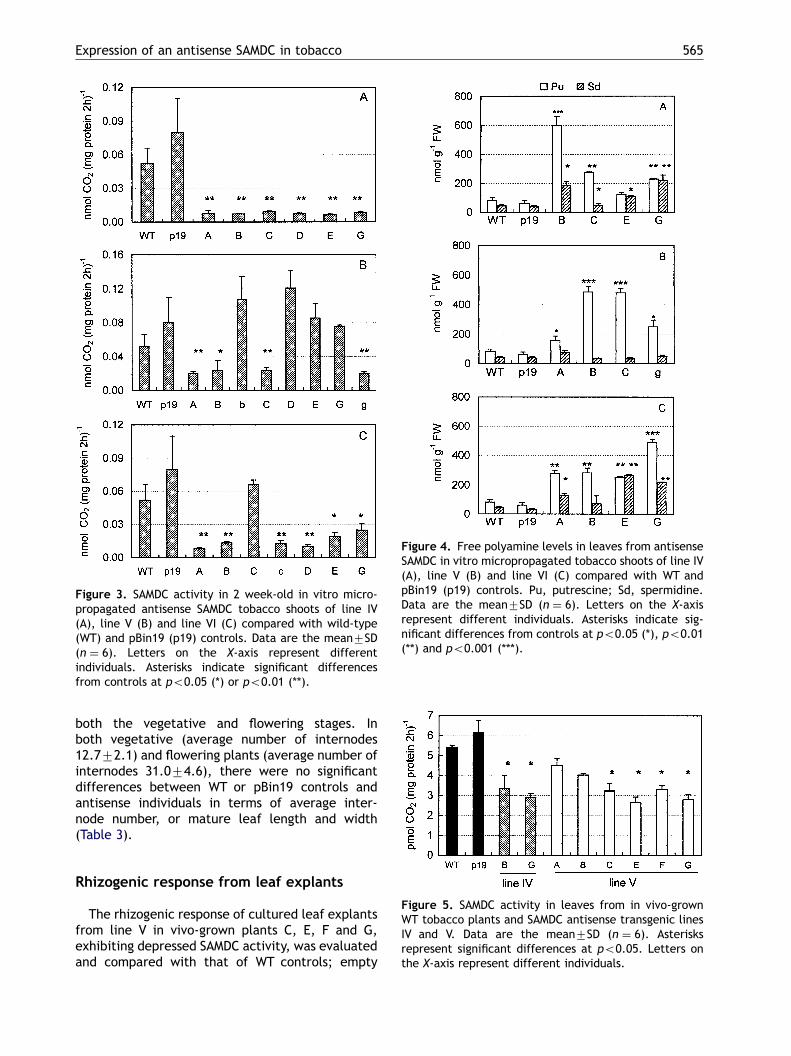
The presence of the foreign gene in all theindividuals selected was confirmed by PCR analysisusing specific primers for the D. stramoniumsequence (Fig. 2). In the antisense micropropa-gated shoots, SAMDC activity was significantly(po0.01 or 0.05) lower in all line IV (Fig. 3A), in50% of line V (Fig. 3B), and in 86% of line VIindividuals (Fig. 3C) compared with that of WT andpBin19 controls (both the average of three plants),which were not significantly different from eachother.
Four antisense genotypes per line exhibitingdecreased SAMDC activity were analysed for PAlevels. They showed an alteration in the freeputrescine-to-spermidine ratio relative to WT andpBin19 controls (which did not differ significantlyfrom each other), mainly due to elevation ofputrescine concentration. In fact, putrescinetitres were enhanced in all transgenics up to6-fold relative to controls, while spermidinedid not increase, or increased to a lesser extent
(Fig. 4A,B,C). In contrast to the controls, the ratiobetween the two amines in the individuals of thethree different lines ranged from 1 to as much as10. Spermine was below the level of detection, andno soluble or insoluble conjugates were detectablein this juvenile physiological stage.
DAO activity, which could be responsible forchanges in putrescine titres, remained essentiallythe same in WT (0.2270.05 nmolmg�1 protein30min�1) and pBin19 (0.2370.08 nmolmg�1 pro-tein 30min�1) controls, and in antisense genotypesof the three lines (mean value 0.26 nmolmg�1
protein 30min�1).In addition, mean ethylene production from
shoots of the three SAMDC antisense lines was notsignificantly different (17.073.8 pl g�1 h�1) fromthat of WT and pBin19 controls (18.673.3 and16.273.25 pl g�1 h�1, respectively).
SAMDC activity, polyamine titres, andmorphology of in vivo-grown plants
In mature leaves of in vivo-grown plants,although SAMDC activity was modestly but signifi-cantly (po0.05) downregulated in the 2 line IVindividuals, and in 4 out of 6 line V individuals(Fig. 5), free, and soluble and insoluble-conjugatedPA titres (average values for each line) displayed nosignificant differences compared with controls(Table 2).
Morphometric analyses were performed on threeuntransformed (WT) and three empty vector-transformed (pBin19) controls, and on the above-cited antisense tobacco plants of lines IV and V at
ARTICLE IN PRESS
Figure 3. SAMDC activity in 2 week-old in vitro micro-propagated antisense SAMDC tobacco shoots of line IV(A), line V (B) and line VI (C) compared with wild-type(WT) and pBin19 (p19) controls. Data are the mean7SD(n ¼ 6). Letters on the X-axis represent differentindividuals. Asterisks indicate significant differencesfrom controls at po0.05 (*) or po0.01 (**).
Figure 4. Free polyamine levels in leaves from antisenseSAMDC in vitro micropropagated tobacco shoots of line IV(A), line V (B) and line VI (C) compared with WT andpBin19 (p19) controls. Pu, putrescine; Sd, spermidine.Data are the mean7SD (n ¼ 6). Letters on the X-axisrepresent different individuals. Asterisks indicate sig-nificant differences from controls at po0.05 (*), po0.01(**) and po0.001 (***).
Expression of an antisense SAMDC in tobacco 565
both the vegetative and flowering stages. Inboth vegetative (average number of internodes12.772.1) and flowering plants (average number ofinternodes 31.074.6), there were no significantdifferences between WT or pBin19 controls andantisense individuals in terms of average inter-node number, or mature leaf length and width(Table 3).
Figure 5. SAMDC activity in leaves from in vivo-grownWT tobacco plants and SAMDC antisense transgenic linesIV and V. Data are the mean7SD (n ¼ 6). Asterisksrepresent significant differences at po0.05. Letters onthe X-axis represent different individuals.
Rhizogenic response from leaf explants
The rhizogenic response of cultured leaf explantsfrom line V in vivo-grown plants C, E, F and G,exhibiting depressed SAMDC activity, was evaluatedand compared with that of WT controls; empty

ARTICLE IN PRESS
Table 2. Free, PCA-soluble and PCA-insoluble conjugated polyamine levels (nmol g�1 FW) in leaves from in vivo-grown tobacco plants of WT, pBin19 and transgenic lines IV and V
Free Soluble conjugated Insoluble conjugated
Pu Sd Sm Pu Sd Sm Pu Sd Sm
WT 130721 200743 26712 3717125 4267150 2875 104723 63712 3.170.8pBin 19 172772 210735 19711 251794 3517106 38715 86716 5578 4.070.5Line IV 199792 254735 26711 350770 3617120 22714 110725 5079 5.371.2Line V 120725 207752 1773 4127118 375764 3178 95712 70713 3.570.7
Data are the mean7SD (n ¼ 6–10) for each line. Pu, putrescine; Sd, spermidine; Sm, spermine.
Table 3. Morphometric analyses (length and width of mature leaves) of in vivo-grown vegetative and flowering WT,pBin19 and SAMDC antisense tobacco plants of lines IV and V. Data are the mean7SD (n ¼ 30–100) for each line
Flowering Vegetative
Leaf length Leaf width Leaf length Leaf width
WT 17.3671.76 11.6671.21 26.3371.76 14.7771.12pBin19 16.3371.73 9.7370.77 29.1773.22 17.7572.38Line IV 19.5871.72 12.1971.40 32.2373.02 17.5871.27Line V 17.7972.40 9.9271.61 26.5673.04 14.4171.78
P. Torrigiani et al.566
vector-transformed controls were not checkedbecause their PA content was the same as in WT.The time course of root formation showed that thepercentage of explants with macroscopic rootsincreased with time (5–10% on day 9, 60% on day12 and 100% on day 15), but without significantdifferences between transgenic and WT genotypes(Fig. 6A,B). At culture end, however, a differencein rhizogenic response between control and anti-sense plants occurred. In fact, the mean number ofroots per explant was 20% lower (po0.05) intransgenic plants relative to controls (Fig. 6C) andthe mean root length (Fig. 6D) was 26% lower(po0.001) in line V than in WT explants.
Figure 6. Rhizogenesis from leaf explants of WT (A) andline V (B) in vivo-grown vegetative plants cultured for15 d on a root-inducing medium containing 0.6mM IAA.Mean number of roots per explant (C) and mean rootlength (D) at culture end. Asterisks represent differencesat po0.001 (***) and po0.05 (*). Bar ¼ 1 cm:
Response of leaf discs to MJ
Free, and soluble and insoluble-conjugated PAtitres were measured in leaf discs from WTand lineV plants treated or not with 10 mM MJ. Again,pBin19 controls were excluded from these experi-ments because they did not display significantdifferences in PA titres compared with WT. FreePA composition showed a prevalence of putrescinein WTand even more so in line V untreated explants(putrescine-to-spermidine ratio 3.8 and 7.1, re-spectively). Upon MJ treatment, free PA titresdecreased by about 50% in WT, but remainedunchanged in line V discs (Fig. 7A).
Upon treatment with MJ, soluble and insolubleconjugated putrescine and spermidine accumula-tion in transgenic explants was differentiallyaffected relative to WT ones (Fig. 7B,C). Thus,soluble conjugated putrescine increased approxi-mately three times in WTand 6 times in line V discs(Fig. 7B). On the contrary, soluble conjugatedspermidine accumulated more in WT (5-fold) thanin transgenic (3.4-fold) explants. Equally, insoluble

ARTICLE IN PRESS
Figure 7. Free (A), PCA-soluble conjugated (B) and PCA-insoluble conjugated (C) polyamine content in leaf discsof WT and line V tobacco plants incubated with 10 mM MJfor 48 h. Pu, putrescine; Sd, spermidine, Sm, spermine.
Expression of an antisense SAMDC in tobacco 567
conjugated putrescine increased 4.4-fold andspermidine 17.5-fold in MJ-treated WT discs, whilein line V ones putrescine increased 6.4-fold andspermidine only 6.8-fold (Fig. 7C).
Discussion
In the present paper, an antisense SAMDC cDNAfrom D. stramonium was expressed in tobacco;primary transformants (R0) and their transgenicprogeny (R1), belonging to three different lines,were compared with WT and empty vector-trans-formed (pBin19) controls in terms of PA metabo-lism, morphogenic potential and response to thestress-mimicking compound MJ.
In leaves of primary transformants, transcriptionof the foreign gene had occurred but SAMDCactivity was unchanged relative to controls. SinceSAMDC is regulated at multiple levels, includingpost-translational cleavage (Xiong et al. 1997;Bennett et al. 2002; Hanfrey et al. 2002), it isplausible that enzyme activity does not parallel
transcript accumulation; alternatively, the anti-sense message may have not been correctlytranslated (Lewin 2000). By contrast, in potatoprimary transformants, constitutive overexpressionof an antisense SAMDC construct led to a 70–90%reduction in SAMDC activity, and to a variabledecline in PA levels with a range of alteredphenotypes (Kumar et al. 1996). Similarly, underthe tuber-specific patatin promoter, some down-regulation of SAMDC activity, and free and con-jugated PA levels occurred but without phenotypicchanges in tubers (Rafart Pedros et al. 1999). In afew cases, other PA biosynthetic genes, such asODC and ADC, were engineered in antisenseorientation. In tobacco transformed with an anti-sense D. stramonium ODC cDNA, no changes in ODCactivity or PA levels were observed in R0 or R1generations (Mayer and Michael 2003). In contrast,previous studies with rice have shown that thoughantisense ADC transgenic callus lines exhibited asignificant reduction of ADC (and ODC) activity, anda decrease in putrescine and spermidine titres,regenerated plants (R0) displayed no variations inPA levels while the progeny (R1) did (Capell et al.2000; Trung-Nghia et al. 2003). Also based onresults obtained with sense transformation (Bassieet al. 2000), the authors underline the fact that,besides promoter strength, the effects of transfor-mation may depend upon developmental stage,i.e., callus, regenerating tissue or shoots.
In accordance with this hypothesis, present datashow that, differently from R0, in second genera-tion micropropagated transformants strong down-regulation of SAMDC activity occurred in themajority of individuals of each line. This, however,did not correlate with depletion of free spermidinelevels (which in fact sometimes increased), andled, instead, to the build-up of probably unutilisedputrescine, resulting in a strongly increased (up to10) putrescine-to-spermidine ratio. Lack of corre-lation between SAMDC activity and spermidinelevels is presumably due to the fact that spermidinesynthase, and not SAMDC, which only furnishes thedcSAM substrate, is directly involved in spermidinesynthesis. In tobacco transformed with a truncatedSAMDC cDNA of Arabidopsis, exhibiting 7-fold up-regulated SAMDC activity, spermidine levels wereunchanged despite dcSAM accumulation up to 400-fold relative to controls (Hanfrey et al. 2002).Although little is known about its regulation inplants, spermidine synthase activity seems todepend, at least in physiological conditions, upondcSAM availability, for which it has a higher affinitythan for putrescine (Yoon et al. 2000). However, intobacco overexpressing spermidine synthase, norelationship was detected between SAMDC and

ARTICLE IN PRESS
P. Torrigiani et al.568
spermidine synthase transcript levels or enzymeactivities suggesting independent regulatory me-chanisms for the two genes (Franceschetti et al.2004). On the other hand, in plants, SAMDC activityis not putrescine-stimulated as in animal cells(Xiong et al. 1997; Bennett et al. 2002); therefore,putrescine accumulation may be compatible withor may be the consequence of the observedrepression of SAMDC activity in micropropagatedshoots. Finally, low levels of SAMDC activity arenormally required in physiological conditions forspermidine synthesis.
Lack of correlation between the extent ofenzyme activity and product accumulation has alsobeen reported for another PA biosynthetic enzyme;in fact, overexpression of a D. stramonium senseODC cDNA in tobacco led to 25-fold increasedenzyme activity but only 2.5-fold putrescineaccumulation (Mayer and Michael 2003). Theauthors suggest that PA homeostatic mechanismsefficiently accommodate changes in enzyme activ-ity since PA biosynthetic control is invested atmultiple independent steps. The fact that enzymeactivity represents single time-point measurementsof a rate of synthesis, while PA levels are the resultof several independent metabolic pathways shouldalso not be overlooked.
Although information is available on plantsexpressing sense constructs (Noury et al. 2000;Thu-Hang et al. 2002; Mayer and Michael 2003), asregards antisense transformation information con-cerning the R1 generation is limited, and practi-cally absent with respect to micropropagatedplants. In vitro-cultured plants are exposed to avariety of stresses, including osmotic, chemical andwound stress. In the present work, substantialSAMDC downregulation was observed only in micro-propagated R1 shoots. This suggests that in vitroculture-induced stress possibly induced the expres-sion of the foreign gene, at least in terms of SAMDCactivity, which otherwise (R0 and in vivo-grown R1plants) was partially or totally counteracted byhomeostatic adjustments. Paradoxically, this led tospermidine accumulation rather than depletion.One possible explanation for this may be that sincerepression of SAMDC activity in such a juvenilestage caused abundant putrescine accumulation,the latter was metabolised to the higher PA. Thepossible regulation of spermidine levels through analtered biosynthesis or utilisation of spermine ispossible, but cannot be evaluated in micropropa-gated tobacco shoots, because spermine is absentor below detection levels.
In general, little is known about the metabolicpathways upstream or downstream of the engi-neered genes (Trung-Nghia et al. 2003; Mayer and
Michael 2003). The increase in putrescine titrespresently observed in transgenic micropropagatedshoots suggests that an upstream metabolic path-way, i.e. that of DAO, may have been altered.However, the differences observed in putrescineoxidising activity remained within the range ofgenotypic variability. Equally, despite the observedreduction in SAMDC activity, ethylene productionwas unaffected in micropropagated shoots, sug-gesting that the two pathways are either notcompetitive and/or that SAM was not limitingunder these conditions (Ravanel et al. 1998; Quanet al. 2002).
Once the micropropagated shoots were allowedto grow in vivo, the resulting transgenic plantsexhibited a strongly attenuated change in pattern;in fact, although SAMDC activity was somewhatdecreased, no changes in free or conjugated PAlevels were observed. As far as the phenotype ofthese plants is concerned, no changes wereobserved. Genetic transformation, either sense orantisense, involving PA biosynthetic genes, in mostcases, leads to no morphological changes, with afew exceptions (Kumar et al. 1996; Rafart Pedroset al. 1999; Masgrau et al. 1997), or unless atruncated SAMDC construct is used (Hanfrey et al.2002). By contrast, Arabidopsis mutants deficientfor ADC activity displayed an anomalous rootapparatus (Watson et al. 1998), while a mutant inspermine synthase activity was found to be severelyaffected in growth and cell elongation (Hanzawaet al. 2000). These data suggest that altered plantphenotypes may be better achieved by PA biosynth-esis mutations.
Rhizogenesis is particularly sensitive to changesin PA levels (Couee et al. 2004). In root-formingtobacco explants both free, and soluble andinsoluble conjugated PAs accumulate dramatically(Torrigiani et al. 1989), and the perturbation of PAbiosynthesis interferes dramatically with organo-genesis (Altamura et al. 1991). In a comparativestudy on Solanum melongena explants induced toform either adventitious shoots or roots, a differ-ential response in terms of PA metabolism has beenreported (Scoccianti et al. 2000). In particular, rootformation, but not shoot formation, was associatedwith a dramatic increase in soluble conjugatedspermidine, prior to and during meristemoid andprimordium formation. Furthermore, the conju-gated-to-free spermidine, but not putrescine, ratiowas consistently higher in the organogenic bordersvs. the non-organogenic central zones of theexplants. Therefore, the diminished rhizogenicresponse in transgenic line V explants, while notcorrelating with the initial PA titre of leaf explants,may be associated with their reduced capacity to

ARTICLE IN PRESS
Expression of an antisense SAMDC in tobacco 569
accumulate conjugated spermidine under non-physiological conditions, as documented here forMJ-treated leaf discs. In fact, in root-formingtobacco thin layers inhibition of SAMDC activityled to a strong reduction of the rhizogenicresponse, and of soluble and insoluble conjugatedspermidine levels (Altamura et al. 1991).
An emerging aspect of the manipulation of PAbiosynthesis is that increased putrescine or spermi-dine levels are associated with increased stressresistance (Roy and Wu 2001, 2002; Kumria andRajan 2002; Waie and Rajam 2003). It has beenpreviously shown that exposure to MJ of tobaccoleaf discs leads to a dramatic accumulation ofconjugated PAs (spermidine more than putrescine)both in the PCA-soluble and -insoluble fraction, andto increased SAMDC mRNA levels and activity(Biondi et al. 2003). As observed in micropropa-gated shoots, the free putrescine-to-spermidineratio was higher in SAMDC antisense leaf discs thanin WT ones, mainly due to enhanced putrescinelevels, this being possibly indicative of depressedSAMDC activity. Moreover, under stress-mimickingconditions (MJ treatment), the relative accumula-tion of conjugated putrescine and spermidine in thetransgenic leaf discs was also different from WTcontrols. Although in absolute terms antisenseexplants exposed to MJ were able to accumulatemore conjugated PAs than WTones, the response tothe elicitor, quantified on the basis of fold increasesin soluble and insoluble conjugates in treated vs.untreated explants, was differentially expressed inthe two genotypes, with putrescine more stronglyinduced than spermidine in the transgenic discs.This again led to an altered putrescine-to spermi-dine ratio. Diversion of dcSAM from spermine tospermidine biosynthesis in order to justify thespermidine increase does not seem plausible inleaf discs since spermine levels are one order ofmagnitude lower than those of spermidine. Sinceconjugates have a role, both physiological andstructural, in defence (Martin-Tanguy 1985; Kelleret al. 1996), their enhanced and differentialaccumulation in transgenic leaf discs could berelevant in stress responses. Possibly, an increasein the putrescine-to-spermidine ratio renders theplants more resistant to stress, though less prone togrowth and differentiation (depressed rhizogen-esis). Besides the fact that the antisense mechan-ism needs to be clarified, these results open newperspectives for studies on response(s) to varioustypes of stress in transgenic plants with altered PAmetabolism.
In conclusion, the expression of an antisenseSAMDC in tobacco seems to be conditioned not onlyby plant developmental stage, but also by imposed
culture conditions. In particular, a stress-inducingevent, such as in vitro culture or MJ treatment, isneeded to obtain a response in terms of changes inPA levels.
Acknowledgements
This research was supported by funds from MIUR(ex-60%) to PTand SB. The authors wish to thank Dr.Anthony J. Michael (Institute of Food Research,Norwich, UK) for stimulating discussions and criticalreading of the manuscript, Tullia Costa, FrancescaPaglierani and Elena Galassi for their excellentcontribution to the experimental work, and Gio-vanni Bugamelli for technical assistance in growingthe plants.
References
Altamura MM, Torrigiani P, Capitani F, Scaramagli S, BagniN. De novo root formation in tobacco thin layers isaffected by inhibition of polyamine biosynthesis. J ExpBot 1991;42:1575–82.
Bagni N, Torrigiani P. Polyamines: a new class of growthsubstances. In: Karssen CM, Van Loon LC, VreugdenhilD, editors. Progress in plant growth regulation.Dordrecht: Kluwer Academic Publishers; 1992. p.264–75.
Bassie L, Noury M, Lepri O, Lahaye T, Christou P, Capell T.Promoter strength influences polyamine metabolismand morphogenetic capacity in transgenic rice tissueexpressing the oat adc cDNA constitutively. TransgenicRes 2000;9:33–42.
Bellincampi D, Cardarelli M, Zaghi D, Serino G, Salvi G,Gatz C, Cervone F, Altamura MM, Costantino P, DeLorenzo G. Oligogalacturonides prevent rhizogenesisin rolB-transformed tobacco explants by inhibitingauxin-induced expression of the rolB gene. Plant Cell1996;8:477–87.
Bennett EM, Ekstrom JL, Pegg AE, Ealick SE. MonomericS-adenosylmethionine decarboxylase from plantsprovides an alternative to putrescine stimulation.Biochemistry 2002;41:14509–17.
Bevan M. Binary Agrobacterium vectors for plant trans-formation. Nucl Acids Res 1984;12:8711–21.
Biondi S, Scaramagli S, Capitani F, Altamura MM,Torrigiani P. Methyl jasmonate upregulates biosyn-thetic gene expression, oxidation and conjugation ofpolyamines, and inhibits shoot formation in tobaccothin layers. J Exp Bot 2001;52:231–42.
Biondi S, Scaramagli S, Capitani F, Marino G, AltamuraMM, Torrigiani P. Ethylene involvement in vegetativebud formation in tobacco thin layers. Protoplasma1998;202:134–44.
Biondi S, Scoccianti V, Scaramagli S, Ziosi V, Torrigiani P.Auxin and cytokinin modify methyl jasmonate effects

ARTICLE IN PRESS
P. Torrigiani et al.570
on polyamine metabolism and ethylene biosynthesis intobacco leaf discs. Plant Sci 2003;165:95–101.
Bouchereau A, Aziz A, Larher F, Martin-Tanguy J.Polyamines and environmental challenges: recentdevelopments. Plant Sci 1999;140:103–25.
Bradford MM. A rapid and sensitive method for thequantitation of microgram quantities of proteinutilizing the principle of protein-dye binding. AnalBiochem 1976;72:248–54.
Capell T, Bassie L, Topsom L, Hitchin E, Christou P.Simultaneous reduction of the activity of two relatedenzymes, involved in early steps of the polyaminebiosynthetic pathway, by a single antisense cDNA intransgenic rice. Mol Gen Genet 2000;264:470–6.
Cohen SS. A guide to the polyamines. New York: OxfordUniversity Press; 1998.
Couee I, Hummel I, Sulmon C, Gouesbet G, El Amrani A.Involvement of polyamines in root development. PlantCell Tiss Org Cult 2004;76:1–10.
Creelman RA, Mullet JE. Biosynthesis and action ofjasmonates in plants. Annu Rev Plant Physiol PlantMol Biol 1997;48:355–81.
Franceschetti M, Hanfrey C, Scaramagli S, Torrigiani P,Bagni N, Burtin M, Michael AJ. Characterization ofmonocot and dicot plant S-adenosylmethionine dec-arboxylase gene families including identification in themRNA of a highly conserved pair of upstream over-lapping open reading frames. Biochem J 2001;353:403–9.
Franceschetti M, Fornale S, Tassoni A, Zuccherelli K,Mayer M, Bagni N. Effects of spermidine synthaseoverexpression on polyamine biosynthetic pathway intobacco plants. J Plant Physiol 2004;161:989–1001.
Guerineau F, Lucy A, Mullineaux P. Effect of twoconsensus sequences preceding the translation initia-tor codon on gene expression in plant protoplasts.Plant Mol Biol 1992;18:815–8.
Hanfrey C, Franceschetti M, Mayer MJ, Illingworth C,Michael AJ. Abrogation of upstream open readingframe-mediated translational control of a plant S-adenosylmethionine decarboxylase results in polya-mine disruption and growth perturbations. J BiolChem 2002;277:44131–9.
Hanzawa Y, Takahashi T, Michael AJ, Burtin D, Long D,Pineiro M, Coupland G, Komeda Y. ACAULIS5, anArabidopsis gene required for stem elongation, en-codes a spermine synthase. EMBO J 2000;19:4248–56.
Keller H, Hohlfeld H, Wray V, Hahlbrock K, Scheel D,Strack D. Changes in the accumulation of soluble andcell wall-bound phenolics in elicitor-treated cellsuspension cultures and fungus-infected leavesof Solanum tuberosum. Phytochemistry 1996;42:389–96.
Kumar A, Minocha SC. Transgenic manipulation of poly-amine metabolism. In: Lindsey K, editor. Transgenicresearch in plants. London: Harwood Academic Pub-lishing; 1998. p. 189–99.
Kumar A, Taylor MA, Mad Arif SA, Davies HV. Potato plantsexpressing antisense and sense S-adenosylmethioninedecarboxylase (SAMDC) transgenes show altered levels
of polyamines and ethylene: antisense plants displayabnormal phenotypes. Plant J 1996;9:147–58.
Kumria R, Rajam MV. Ornithine decarboxylase transgenein tobacco affects polyamines, in vitro-morphogenesisand response to stress. J Plant Physiol 2002;159:983–90.
Lewin B. Genes VII. New York: Oxford University Press;2000. pp. 312–8.
Malmberg RL, McIndoo J. Abnormal floral development ofthe tobacco mutant with elevated polyamine levels.Nature 1983;305:623–5.
Martin-Tanguy J. The occurrence and possible function ofhydroxycinnamoyl acid amides in plants. Plant GrowthRegul 1985;3:381–99.
Masgrau C, Altabella T, Farras R, Flores D, Thompson AJ,Besford RT, Tiburcio AF. Inducible overexpression ofoat arginine decarboxylase in transgenic tobaccoplants. Plant J 1997;11:465–73.
Mayer MJ, Michael AJ. Polyamine homeostasis in trans-genic plants overexpressing ornithine decarboxylaseincludes ornithine limitation. J Biochem 2003;134:765–72.
Mayer MJ, Narbad A, Parr AJ, Parker ML, Walton NJ,Mellon A, Michael AJ. Rerouting the plant phenylpro-panoid pathway by expression of a novel bacterialenoyl-coenzyme A hydratase/lyase enzyme function.Plant Cell 2001;13:1669–82.
Murashige T, Skoog F. Revised medium for rapid growthand bioassay with tobacco tissue cultures. PhysiolPlant 1962;15:473–97.
Noh EW, Minocha SC. Expression of a human S-adeno-sylmethionine decarboxylase cDNA in transgenic to-bacco and its effect on polyamine biosynthesis.Transgenic Res 1994;3:25–33.
Noury M, Bassie L, Lepri O, Kurek I, Christou P, Capell T. Atransgenic rice cell lineage expressing the oat argininedecarboxylase (ADC) cDNA constitutively accumulatesputrescine in callus and seeds but not in vegetativetissues. Plant Mol Biol 2000;43:537–44.
Quan Y, Minocha R, Minocha SC. Genetic manipulation ofpolyamine metabolism in poplar. II. Effects onethylene biosynthesis. Plant Physiol Biochem 2002;40:929–37.
Rafart Pedros A, MacLeod MR, Ross HA, McRae D, TiburcioAF, Davies HV, Taylor MA. Manipulation of S-adenosyl-methionine decarboxylase activity in potato tubers.Planta 1999;209:153–60.
Ravanel S, Gakiere B, Job D, Douce R. The specificfeatures of methionine biosynthesis and metabolism inplants. Proc Natl Acad Sci USA 1998;95:7805–12.
Roy M, Wu R. Arginine decarboxylase transgene expres-sion and analysis of environmental stress tolerance intransgenic rice. Plant Sci 2001;160:869–75.
Roy M, Wu R. Overexpression of S-adenosylmethioninedecarboxylase gene in rice increases polyamine leveland enhances sodium chloride-stress tolerance. PlantSci 2002;163:987–92.
Sambrook J, Fritsch EF, Maniatis T. Molecular cloning: Alaboratory manual. Cold Spring Harbor: Cold SpringHarbor Laboratory Press; 1989.

ARTICLE IN PRESS
Expression of an antisense SAMDC in tobacco 571
Scaramagli S, Biondi S, Torrigiani P. Methylglyoxal(bis-guanylhydrazone) inhibition of organogenesis is notdue to SAMDC inhibition/polyamine depletion intobacco thin layers. Physiol Plant 1999;107:353–60.
Scaramagli S, Bueno M, Torrigiani P, Altamura MM,Capitani F, Bagni N. Morphogenesis in cultured thinlayers and pith explants of tobacco. II Early hormone-modulated polyamine biosynthesis. J Plant Physiol1995;147:113–7.
Scoccianti V, Sgarbi E, Fraternale D, Biondi S. Organogen-esis from Solanum melongena L. (eggplant) cotyledonexplants is associated with hormone-modulated en-hancement of polyamine biosynthesis and conjuga-tion. Protoplasma 2000;211:51–63.
Serafini-Fracassini D, Del Duca S, Monti F, Poli F, SacchettiG, Bregoli AM, Biondi S, Della Mea M. Transglutaminaseactivity during senescence and programmed cell deathin the corolla of tobacco (Nicotiana tabacum) flowers.Cell Death Differ 2002;9:309–21.
Thu-Hang P, Bassie L, Safwat G, Trung-Nghia P, Christou P,Capell T. Expression of a heterologous S-adenosyl-methionine decarboxylase cDNA in plants demon-strates that changes in S-adenosylmethioninedecarboxylase activity determine levels of the higherpolyamines spermidine and spermine. Plant Physiol2002;129:1744–54.
Torrigiani P, Altamura MM, Capitani F, Falasca G, Bagni N,Serafini-Fracassini D. De novo root formation in thincell layers of tobacco: changes in free and boundpolyamines. Physiol Plant 1989;77:294–301.
Trung-Nghia P, Bassie L, Safwat G, Thu-Hang P, Lepri O,Rocha P, Christou P, Capell T. Reduction in theendogenous arginine decarboxylase transcript levelsin rice leads to depletion of the putrescine andspermidine pools with no concomitant changes in theexpression of downstream genes in the polyaminebiosynthetic pathway. Planta 2003;218:125–34.
Waie B, Rajam MW. Effect of increased polyaminebiosynthesis on stress responses in transgenic tobaccoby introduction of human S-adenosylmethionine gene.Plant Sci 2003;164:727–34.
Walters D. Resistance to plant pathogens: possible rolesfor free polyamines and polyamine catabolism. NewPhytol 2003;159:109–15.
Watson MB, Emory KK, Piatak RM, Malmberg RL. Argininedecarboxylase (polyamine synthesis) mutants of Ara-bidopsis thaliana exhibit altered root growth. Plant J1998;13:231–9.
Xiong H, Stanley BA, Tekwani BL, Pegg AE. Processing ofmammalian and plant S-adenosylmethionine decar-boxylase proenzymes. J Biol Chem 1997;272:28342–8.
Yoon SO, Lee YS, Cho YD. Polyamine synthesis in plants:isolation and characterisation of spermidine synthasefrom soybean (Glycine max) axes. Biochem BiophysActa 2000;1475:17–26.
Zhang Z, Honda C, Kita M, Hu C, Nakayama M, MoriguchiT. Structure and expression of spermidine synthasegenes in apple: two cDNAs are spatially and devel-opmentally regulated through alternative splicing. MolGen Genet 2003;268:799–807.














